gabaergic pump cells of solitary tract nucleus innervate retrotrapezoid nucleus chemoreceptors
TRANSCRIPT

GABAergic pump cells of solitary tract nucleus innervate retrotrapezoid nucleus chemoreceptors
Ana C. Takakura1,2, Thiago S. Moreira1,2, Gavin H. West1, Justin M. Gwilt1, Eduardo Colombari2, Ruth L. Stornetta1 and Patrice G. Guyenet1
1Department of Pharmacology, University of Virginia, Charlottesville, VA, 22908, USA;2Department of Physiology, UNIFESP-EPM, São Paulo, SP, 04023-060, Brazil.
Abbreviated title: pump cells and RTNKey words: lung stretch receptors, solitary tract nucleus, central chemoreceptors, medulla oblongata, breathing, respiration.
Number of text pages: 25Number of figures: 6
Address correspondence to:Patrice G. Guyenet, PhD,Department of PharmacologyUniversity of Virginia Health SystemP.O. Box 8007351300 Jefferson Park AvenueCharlottesville, VA 22908-0735.Tel: (434) 924-7433; Fax: (434) 982-3878; E-mail: [email protected]
Acknowledgement: This research was supported by grants from the National Institutes of Health to PGG (HL 74011 and HL 28785) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior to ACT (BEX 4402/05-7). A.C. Takakura and T.S. Moreiracontributed equally to the study.
Page 1 of 31 Articles in PresS. J Neurophysiol (April 25, 2007). doi:10.1152/jn.00322.2007
Copyright © 2007 by the American Physiological Society.

2
ABSTRACT
The retrotrapezoid nucleus (RTN) contains central respiratory chemoreceptors
that are inhibited by lung inflation via slowly-adapting pulmonary stretch receptors
(SARs). Here we examine whether RTN inhibition by lung inflation could be mediated
by a direct projection from SAR second-order neurons (pump cells).
Pump cells (N = 56 neurons, 13 rats) were recorded in the nucleus of solitary tract
(NTS) of halothane-anesthetized rats with intact vagus nerves. Pump cells had discharges
that coincided with lung inflation as monitored by the tracheal pressure. Their activity
increased when end-expiratory pressure was raised and stopped instantly when
ventilation was interrupted in expiration. Many pump cells could be antidromically
activated from RTN (12/36). Nine of those were labeled with biotinamide. Of these 9
cells, eight contained glutamic acid decarboxylase67- (GAD67-) mRNA and seven were
found to reside in the lower half of the interstitial subnucleus of NTS (iNTS). Using the
retrograde tracer cholera toxin-B, we confirmed that neurons located in or close to iNTS
innervate RTN (2 rats). Many such neurons contained GAD67-mRNA and a few
contained glycine transporter2 (GLYT2) mRNA. Anterograde tract-tracing with
biotinylated dextranamide (4 rats) applied to iNTS also confirmed that this region
innervates RTN via a predominantly GABAergic projection.
This work confirms that rat many NTS pump cells are located in and around the
interstitial subnucleus at area postrema level. We demonstrate that a GABAergic subset
of these pump cells innervates the RTN region. We conclude that these inhibitory
neurons probably contact RTN chemoreceptors and mediate their inhibition by lung
inflation.
Page 2 of 31

3
INTRODUCTION
The rat retrotrapezoid nucleus (RTN) is a cluster of acid-sensitive glutamatergic
neurons with properties consistent with a central respiratory chemoreceptor function
(Okada et al., 2002; Feldman et al., 2003; Mulkey et al., 2004; Putnam et al., 2004;
Takakura et al., 2006). These neurons selectively innervate the pontomedullary regions
that contain the respiratory pattern generator, CPG (Cream et al., 2002; Mulkey et al.,
2004; Feldman and Del Negro, 2006; Rosin et al., 2006). The excitatory input from RTN
neurons seems necessary for the CPG to be active under anesthesia and RTN neurons
also appear to contribute notably to respiration in the awake state and during sleep
(Akilesh et al., 1997; Nattie and Li, 2000; Feldman et al., 2003; Amiel et al., 2003;
Takakura et al., 2006; Stornetta et al., 2006).
The activity of RTN neurons is subject to several feedbacks consistent with their
postulated role as a pH-regulated source of excitatory drive to the CPG. For example,
their intrinsic acid sensitivity combined with their excitatory input from the carotid
bodies forms a long-loop feedback that seems appropriate to contribute to the
homeostatic regulation of pCO2 and pO2 (Nattie et al., 2001; Li and Nattie, 2002;
Feldman et al., 2003; Takakura et al., 2006). RTN neurons also receive inhibitory inputs
from the CPG (Guyenet et al., 2005) and are inhibited by the activation of slowly-
adapting lung stretch receptors (SARs) (Moreira et al., 2007). The latter two feedbacks
seem appropriate to reduce the intensity of the excitatory input from RTN
chemoreceptors to the CPG when the latter network is being adequately driven by other
inputs.
Page 3 of 31

4
According to our prior study, SARs may inhibit RTN neurons via the simplest
possible route namely via a direct projection from inhibitory second-order relay neurons
(pump cells) located in the solitary tract nucleus (NTS) (Moreira et al., 2007). The
present study is designed to test this possibility. Using a combination of
electrophysiological and anatomical experiments, we show here that GABAergic NTS
pump cells do indeed innervate the RTN region .
METHODS
Animals
The experiments were performed on 19 male Sprague-Dawley rats (Taconic;
Germantown, NY, USA) weighing 250-350 g. Procedures were in accordance with NIH
Animal Care and Use Guidelines and were approved by the University of Virginia’s
Animal Care and Use Committee.
In vivo recordings
General anesthesia was induced with 5% halothane in 100% oxygen. The rats
received a tracheostomy. Artificial ventilation with 1.4-1.5% halothane in 100% oxygen
was maintained throughout surgery. The surgical procedures (arterial cannulation,
phrenic nerve dissection, dorsal access to the medulla oblongata through the atlanto-
occipital membrane) have been described previously (Guyenet et al., 2005; Takakura et
al., 2006). A bilateral pneumothorax was performed for added stability and both vagus
nerves were left intact.
Page 4 of 31

5
Upon completion of surgical procedures, halothane concentration was reduced to
1% (still in 100% oxygen). This anesthetic level is sufficient to abolish the corneal reflex
and the following reactions to strong nociceptive stimulation of the hindpaw: retraction of
distal phalanges, increases in phrenic nerve discharge (PND) amplitude and rise of
arterial pressure (AP). Rectal temperature was maintained at 37°C and end-expiratory
CO2 was monitored throughout the experiment with a capnometer (Columbus
Instruments, Ohio, USA). The muscle relaxant pancuronium was administered at the
initial dose of 1 mg/kg i.v. and the adequacy of anesthesia was thereafter gauged by the
lack of increase in AP and PND amplitude to firm toe pinch. No adjustment of the
halothane concentration was needed.
Arterial pressure (AP), PND and tracheal CO2 were recorded as previously
described (Mulkey et al., 2004; Takakura et al., 2006; Stornetta et al., 2006). Lung
inflation was monitored by measuring tracheal pressure through a side port of the tracheal
cannula. Lung mechanoreceptors were activated by transiently elevating positive end-
expiratory pressure (PEEP) (5-20 sec) from a resting level of +1 cm H2O to +2, +4 or +6
cm H2O. Single-unit recording and juxtacellular labeling of the recorded units with
biotinamide were done as described previously (Mulkey et al., 2004; Takakura et al.,
2006; Stornetta et al., 2006). All recordings were done within the portion of the NTS that
extends from the rostral end to the caudal end (calamus scriptorius) of the area postrema.
Pump cells were encountered at a depth of 300 to 700 µm below the dorsal surface of the
medulla oblongata within a longitudinally oriented column of at most 300 µm in width
with its center 600 to 1000 µm from the midline depending on the rat. Most recordings
were made on the left side of the brain.
Page 5 of 31

6
All analog data (end-expiratory CO2, PND, unit activity, AP) were stored on a
microcomputer via a micro-1401 digitizer from Cambridge Electronics Design (CED,
Cambridge, UK) and were processed off-line using version 5 of the Spike 2 software
(CED). Processing included action potential discrimination and binning, neuronal
discharge rate measurement, and PND “integration” (iPND) consisting of rectification
and smoothing (τ: 0.015s) (Mulkey et al., 2004; Takakura et al., 2006; Stornetta et al.,
2006). The CED software was also used for acquisition of peri-event histograms of
neuronal activity and peri-event averages of iPND, tracheal CO2, or tracheal pressure.
The peri-event histograms of neuronal single-unit activity were triggered either on iPND
or on the tracheal pressure trace. Each histogram represents the summation of at least 100
consecutive central respiratory or ventilation cycles (350 to 800 action potentials per
histogram). At the end of the physiological experiment the rats were deeply anesthetized
with halothane (4% until AP reached 40 mmHg). Then they were perfused through the
left cardiac ventricle with phosphate-buffered saline (pH 7.4; 150 ml) followed by
paraformaldehyde (4% in 0.1 M phosphate buffer, pH 7.4, 500 ml). The brains were
postfixed overnight in the paraformaldehyde solution. The brains were then sectioned in
the coronal plane (30 µm) and the sections were kept in cryoprotectant awaiting
histological procedures.
Tracer injections
Survival surgery was done using aseptic procedures on rats anesthetized with a
mixture of ketamine, xylazine and acepromazine as described previously (Rosin et al.,
2006). After surgery and tracer injection, the rats received an antibiotic (ampicillin, 100
Page 6 of 31

7
mg/kg, i.m.; American Pharmaceutical Partners, Schaumberg, IL) and an analgesic
(Ketorolac tromethamine, 0.6 mg/kg, s.c.; Abbott Laboratories). One week later the rats
were deeply anesthetized with pentobarbital (60 mg/kg, i.p.; Abbott Laboratories) then
killed by intracardiac perfusion with 4% formaldehyde as detailed above. Tracer injection
was done exactly as described previously (Rosin et al., 2006). The procedure involved
iontophoretic delivery of the tracer through glass electrodes drawn from 2 mm glass with
an external tip diameter of no more than 5 microns. In 4 rats, the anterograde tracer
biotinylated dextranamide (BDA; lysine fixable, MW 10,000; 10% by weight in 10 mM
phosphate buffer, pH 7.4; Molecular Probe, Eugene, OR; 5 µA positive current 5s ON 5s
OFF) was injected into the region of the solitary tract nucleus that contains the pump
cells (500 µm rostral to the calamus scriptorius, 0.8 mm lateral to the midline and 0.5 mm
below the dorsal surface). Two other rats received an injection of cholera toxin B, CTB
(1% in 0.2 M phosphate buffer, pH 7.35; List Biological, Campbell, CA; 5 µA positive
current 5s ON 5s OFF) into the RTN. Injections into the RTN were guided by recording
the facial field potential and were placed 200-300 µm below the lower edge of the field,
1.6–1.9 mm lateral to the midline and 200–400 µm rostral to the caudal end of the field
(Rosin et al., 2006). This region contains the largest concentration of RTN
chemoreceptors (Stornetta et al., 2006).
Histology
All histochemical procedures were done using 30 µm-thick free-floating sections
according to previously described protocols (Takakura et al., 2006; Rosin et al., 2006;
Stornetta et al., 2006). Neurons labelled in vivo with biotinamide (juxtacellular
Page 7 of 31

8
recording) or terminal fields labeled anterogradely with BDA were identified by
incubating the sections with streptavidin conjuguated with Cy-3 according to previously
described methods (Mulkey et al., 2004; Rosin et al., 2006). Glutamic acid decarboxylase
67 (GAD67) mRNA and Glycine transporter2 (GLYT2) mRNA were detected by non-
radioactive in situ hybridization using digoxigenin-labeled cRNA probes as described
previously (Stornetta and Guyenet, 1999; Stornetta et al., 2003b). GAD
immunoreactivity was detected by immunofluorescence using a rabbit anti GAD65 and
67 antibody (AB 5907; Chemicon International, Temecula, CA, USA; dilution 1:5000
dilution). Guinea pig anti-GAD65/67 (Chemicon AB1511) was raised against a synthetic
peptide (Chemicon AG252) with amino acid sequence CDFLIEEIERLGQDL from the
C-terminal portion of rat glutamate decarboxylase (GAD65; Cys + C-terminal residues
572-585). The manufacturer has demonstrated by western blot of whole brain lysate that
the antiserum recognizes bands of the correct molecular weight corresponding to the two
isoforms of the enzyme. In our laboratory, preabsorption of the GAD65/67 antibody with
a 3-fold molar excess of the immunogen provided by Chemicon (AG252) eliminated
immunolabeling in brain sections.
Cell mapping and imaging
The computer-assisted mapping technique designed to map the location of tracer
injection sites, BDA-labelled terminal fields and biotinamide-labeled neurons has been
described in detail previously (Neurolucida; MicroBrightField, Williston, VT, USA; see
(Stornetta and Guyenet, 1999)). Section alignment between brains was done relative to a
reference section as follows. Specific segments of the ventral respiratory column were
Page 8 of 31

9
identified by their location caudal to the caudal end of the facial motor neurons in each
brain and brain levels were named according to the atlas of Paxinos and Watson (1998).
On the basis of our previous work in adult rats of the same age and strain, the pre-
Bötzinger region of the ventral respiratory column (for definition see (Alheid et al., 2002;
Feldman and Del Negro, 2006)) was judged to lie 900 ± 200 micron caudal to the facial
motor nucleus (Bregma -12.5 mm after Paxinos and Watson (Paxinos and Watson, 1998))
and the rostral ventral respiratory group to reside 1.3 mm or more caudal to facial nucleus
(caudal to -12.9 mm in the Paxinos and Watson atlas (Stornetta et al., 2003a)).
The Neurolucida files were exported to the Canvas 9 software drawing program
(ACD Systems of America, Miami, FLA USA) for final modifications. Photographs were
taken with a 12-bit color CCD camera (CoolSnap, Roper Scientific, Tuscon, AZ;
resolution 1392 X 1042 pixels). IPLab software (Scanalytics, Rockville, MD, USA) was
used for merging of color channels in photographs of dual labeling experiments.
RESULTS
Electrophysiological evidence that GABAergic NTS pump cells innervate RTN
NTS pump cells (N = 56) were identified as described by others previously (Ezure
et al., 2002; Ezure and Tanaka, 2004). These cells were phasically active and their
discharge coincided with lung inflation as monitored by the tracheal pressure (Fig. 1A1-
A3). Their activity increased when end-expiratory pressure was raised (Fig. 1A1, A2, A5)
and the cells became immediately silent when ventilation was interrupted in expiration
(Fig. 1A1). Such cells could only be found reliably within a narrow (< 300 µm)
rostrocaudally oriented cell column centered 0.83 ± 0.04 mm lateral to the midline (N =
Page 9 of 31

10
13 rats). All recordings were made in the NTS at area postrema level therefore the full
rostrocaudal extent of the cell column was not investigated. Although the pump cells that
were recorded had very homogeneous properties with respect to our test criteria, we
cannot state whether the sample is representative of the entire population of pump cells
present in the NTS.
The same region of the brain contained numerous other types of respiratory-
phasic neurons (not illustrated). These cells were inhibited by raising end-expiratory
pressure and/or remained phasically active after interrupting ventilation, indicating that
they were not pump cells. Recording was interrupted after this fact was established and
no detailed characterization was performed.
Thirty-six pump cells were tested for antidromic activation from the ipsilateral
RTN and twelve could be antidromically activated from this region (latency 6.3 ± 0.18
ms; range: 5.4 - 7.5 ms). The antidromic nature of the evoked spikes was demonstrated
with the collision test (Lipski, 1981) (Fig. 2A). There was no statistical difference
between the resting discharge rate of pump cells with or without axonal projection to
RTN level (Fig. 2B). All the stimulation sites were identified histologically (Fig. 2C).
These sites were centered ventromedial to the very caudal end of the facial motor nucleus
(1. 8 mm from the midline; -11.6 mm caudal to Bregma according to the atlas of Paxinos
and Watson (1998)), region that contains the highest density of chemoreceptors as we
have defined them previously (Fig. 2D) (Mulkey et al., 2004; Stornetta et al., 2006).
Nine of the 12 pump cells that were antidromically activated from RTN were
labeled with biotinamide. A representative example is shown in Fig. 3A. This
biotinamide-labeled pump cell (small red object photographed under fluorescence
Page 10 of 31

11
illumination) is shown superimposed on a dark-field image of the same field in which
myelinated fiber bundles appear white and neuron-rich regions are dark. The labeled
pump cell was located at mid-area postrema level between the bundles of myelinated
axons that define the solitary tract (sol). More precisely, this neuron was located just
below the distinctive circular portion of the interstitial subnucleus (Sol-I), a structure that
has been previously described as the main projection site of laryngeal afferents
(Patrickson et al., 1991; Furusawa et al., 1996). Seven out of the nine recovered pump
cells had virtually the same location (Fig. 3B). The labeled pump cell shown in Fig. 3A
contained GAD67mRNA (Fig. 3C1, C2). Two other examples of GAD67-positive NTS
pump cells that were backfired from RTN are shown in Figure 3 (panels D1-E2).
GAD67mRNA was detected in eight out of the nine cells recovered (Fig. 3B).
Histological evidence for a GABAergic projection from the pump cell region of the
NTS to RTN
The anterograde transport of biotinamide dextranamide (BDA) was used in an
effort to confirm the existence of a GABAergic projection to RTN from the region of the
NTS that contains the pump cells. BDA was injected by iontophoresis in 4 rats at the sites
indicated in Fig. 4A. A representative injection site is shown in Fig. 4B. An abundant
field of BDA-labeled varicosities was present throughout the ventrolateral medulla (Fig.
4C and D) extending into the RTN region (Fig. 4 D- F). Synaptic terminals were less
numerous in the RTN than in more caudal regions of the ventral respiratory column such
as the region of the preBötzinger complex and the rostral ventral respiratory group, rVRG
(Fig. 4G). Simultaneous histological detection of BDA and GAD revealed that a high
Page 11 of 31

12
proportion of the anterogradely labeled varicosities were GABAergic (Fig. 5A-C). A
random sampling of BDA-labeled varicosities located within RTN revealed that
approximately 63% of the BDA-labeled terminals were immunoreactive for GAD (total
of 55 varicosities sampled in 4 rats).
We used the retrograde transport of cholera-toxin B (CTB) as final confirmation
of the existence of a GABAergic projection from the region of the NTS that contains the
pump cells to RTN. A small iontophoretic deposit of CTB was made into RTN in two
rats (Fig. 6A) and the segment of the NTS located at area postrema level was examined
for the presence of CTB-labeled neurons. One series of sections was processed for
simultaneous detection of BDA and GAD67 mRNA and a second series was reacted for
detection of CTB and GLYT2 mRNA.
At the level of the area postrema, the CTB labeled neurons were clustered within
two regions. The first cluster (henceforth called lateral cluster) was located in and around
the solitary tract (sol) and included a high proportion of neurons that were positive for
GAD67 mRNA (Fig. 6B, C, D). GLYT2 mRNA could also be detected in a small number
of these neurons (Fig. 6B, D). The second cluster of CTB-labeled neurons (henceforth
called medial cluster) was located within the commissural nucleus and extended into the
dorsolateral edge of the median nucleus as defined by Paxinos and Watson (1998). This
larger cell group contained few GAD67 mRNA (Fig. 6C) and no detectable GLYT2
mRNA (not illustrated). To quantify the proportion of retrogradely labeled GAD67-
positive neurons present in the lateral and medial clusters, the NTS was divided in two
regions by a vertical line drawn 500 µm lateral to the midline. In the region of the NTS
situated 0-500 µm from the midline only 10% of the CTB-labeled neurons contained
Page 12 of 31

13
GAD67 mRNA (5 of 50 cells counted in 3 section/rat in 2 rats). This proportion was 40%
in the lateral portion of the NTS extending 500 – 1000 µm from the midline (10 of 24
CTB-labeled cells counted).
DISCUSSION
The present study confirms the presence of GABAergic pump cells within the
NTS in and around the tractus solitarius at area postrema level (Ezure and Tanaka, 2004).
We also show that a subset of these pump cells innervate the RTN region and we
conclude that this projection could be responsible for the previously described inhibitory
effect of lung inflation on RTN chemoreceptors (Moreira et al., 2007).
Inhibitory pump cells innervate RTN
The NTS neurons that we identified as pump cells displayed the same
characteristics as the cells previously recorded in both cats and rats and their location was
generally conform to prior descriptions (Bonham and McCrimmon, 1990; Ezure and
Tanaka, 1996; Ezure et al., 2002). In the present study, evidence that these cells receive
excitatory inputs from slowly-adapting pulmonary afferents (SARs) was based on the
following criteria: a discharge pattern that is strictly synchronized with lung inflation, a
slowly-adapting excitatory response to lung inflation and the absence of a central
respiratory activity. In addition, others have shown that these cells are driven at very
short latencies by low-intensity vagus nerve stimulation which suggests that their vagal
input is from myelinated afferents and is probably monosynaptic (Ezure and Tanaka,
1996; Miyazaki et al., 1999; Ezure et al., 2002). Finally, since our rats were paralyzed
Page 13 of 31

14
and artificially ventilated through a tracheal cannula, the pump cells that we recorded
could not have been driven by laryngeal or pharyngeal mechanoreceptor inputs sensitive
to air flow or to changes in upper airway diameter.
A minor debate persists as to the exact location of the NTS pump cells (Kubin et
al., 2006). Depending on the study, these neurons have been described as residing
predominantly lateral to the tractus solitarius (Ezure et al., 2002), within the interstitial
subnucleus (Ezure and Tanaka, 2004) or medial to the tractus solitarius (Bonham and
McCrimmon, 1990). These apparent discrepancies may be explained by the fact that the
tractus solitarius is not a compact fiber tract but a collection of bundles of differing size
and geometry with an imprecise ventrolateral border made of smaller fiber tracts. This
fact can be readily appreciated when non-delipidated brain tissue is observed under dark-
field illumination (see Fig. 3A). The interstitial subnucleus of the NTS is usually defined
as a sizable and roughly circular myelin-poor region located amidst the dorso-medial
fascicles of the tractus solitarius (Altschuler et al., 1989) (Fig. 3A). This zone and the
immediately overlying neuropil have been described as receiving a very dense input from
laryngeal and pharyngeal afferents in rats (Altschuler et al., 1989; Patrickson et al., 1991;
Furusawa et al., 1996). The pump cells that we labeled (8/9) were located at the periphery
of this circular region (in all directions except dorsal) but never in it. If one ignores the
smaller outer bundles of the tractus solitarius, one could easily conclude that some of the
pump cells reside ventrolateral to the solitary tract, as suggested by Ezure and colleagues
in their earlier work (Ezure and Tanaka, 1996; Miyazaki et al., 1998). Our
electrophysiological sampling of 56 pump cells suggests that these cells reside within a
longitudinal column that is about 300 microns-wide in the rat. This lateral spread clearly
Page 14 of 31

15
exceeds the width of the interstitial nucleus narrowly defined as the small myelin-poor
circular zone alluded to above. Based on this observation and the anatomical location of
the biotinamide labeled neurons, our results suggest that the cell bodies of the pump cells
reside at the immediate periphery of the interstitial nucleus. Some of these cells are
located medial to the tractus solitarius as suggested by Bonham and McCrimmon (1990).
In their latest work, Ezure and Tanaka’s (2004) describe the pump cells as residing in
“the interstitial subnucleus of the solitary tract nucleus and its immediate vicinity”. This
definition is in reasonable agreement with the present finding if one defines the
interstitial nucleus more broadly as the neuropil that is interspersed with the myelinated
bundles of the tractus solitarius.
Collectively and individually, NTS pump cells innervate large portions of the
ventral respiratory column as well as selected dorsal pontine regions that are also
involved in respiratory pattern generation (Ezure and Tanaka, 1996; Ezure et al., 2002).
The present study suggests that RTN is among the structures that receive a direct input
from these pump cells. This conclusion derives first and foremost from the fact that we
could antidromically activate about 30% of the pump cells by microstimulation within
RTN. The relatively long latency of the antidromic spikes (5.4 - 7.5 ms) suggests that this
projection probably consists of unmyelinated or lightly myelinated axons as is the case of
other pump cells according to Ezure and Tanaka (1996). The long antidromic latencies
that we observed may also denote the fact that we stimulated fine axonal collaterals rather
than the main axon of the pump cells. This interpretation is supported by the fact that the
minimum latency of activation of these cells from the dorsal pons is 1.9 to 4.5 ms (Ezure
et al., 2002), which is slightly less than from RTN. Nonetheless, we were unable to elicit
Page 15 of 31

16
antidromic spikes at multiple discrete latencies from the RTN stimulation sites, therefore
our electrophysiological approach alone did not provide definite evidence that the pump
cells that we recorded had a terminal field in RTN rather than just an axon of passage.
Accordingly, we sought additional anatomical evidence of pump cell projection to RTN.
The anterograde and retrograde tracing experiments provided supportive evidence by
showing that GABAergic neurons located in the region of the NTS that contains the
pump cells do innervate RTN.
The present study confirms and extends prior evidence that a large majority of
NTS pump cells in the rat are GABAergic and that a small fraction of these cells may
also release glycine (Ezure and Tanaka, 2004). In our case, 8 out of nine pump cells were
clearly positive for GAD67 (89%) vs. 10/14 (71%) in Ezure and Tanaka’s report (2004).
The difference could be due to slight variations in histological protocol or to the fact that
we only sampled pump cells with axonal projection to RTN whereas Ezure and Tanaka
labeled generic pump cells without regard to projection pattern.
If the NTS pump cells are inhibitory, how does the Breuer-Hering reflex work?
NTS pump cells mediate the Breuer-Hering reflex prolongation of expiration
(Bonham and McCrimmon, 1990; Bonham et al., 1993; Hayashi et al., 1996). The exact
neuronal targets of the NTS pump cells have been difficult to determine because most
ventral respiratory column (VRC) neurons are impacted by lung stretch afferent
stimulation due to the complex interactions that exist within the CPG network (Krolo et
al., 2005). Based on the assumption that the most likely targets of the pump cells are
neurons in which single-shock stimulation of the vagus nerve elicits the shortest latency
Page 16 of 31

17
PSCs, this reflex is currently thought to be mediated via the di-synaptic activation of
expiratory decremental (E-DEC a.k.a. post-inspiratory) medullary interneurons (Hayashi
et al., 1996; Kubin et al., 2006). The theory is compatible with the projection pattern of
NTS pump cells and consistent with the fact that E-DEC interneurons inhibit inspiratory
and other neurons (Richter et al., 1987) by releasing glycine (Ezure et al., 2003) and/or
GABA (Okazaki et al., 2001). Also in accordance with the theory, the latency of the
EPSCs recorded in E-DEC neurons is among the shortest of any VRC neuron and this
latency is shorter than that of the IPSCs recorded in other VRC neurons (Hayashi et al.,
1996; Kubin et al., 2006). The weakest link of this theory is that it postulates the
existence of excitatory pump cells for which no anatomical evidence exists yet. However,
this possibility is not ruled out by the present study nor by that of Ezure and Tanaka
(2004) because neither of these studies found that 100% of the pump cells are
GABAergic. Furthermore, the hypothetical excitatory pump cells may reside at some
distance from the region that was preferentially explored by Ezure and Tanaka (2004)
and by ourselves.
Physiological role of the GABAergic pump cell input to RTN
The existence of a direct projection from GABAergic NTS pump cells to RTN
chemoreceptors accounts for the fact that many of these cells are inhibited by lung
inflation (Moreira et al., 2007). Indeed, the inhibitory effect of lung inflation on RTN
neurons is eliminated by silencing neurons located in the NTS region that contains the
GABAergic pump cells (Moreira et al., 2007) and many RTN neurons are phasically
inhibited during each lung inflation as would be expected if they received a periodic
Page 17 of 31

18
inhibitory input from the type of pump cells identified in the present study (Moreira et al.,
2007). Finally, the inhibitory effect of lung inflation on RTN neurons is unaffected by
silencing neurons within the rVRG on both sides using the GABA-A agonist muscimol
(Moreira et al., 2007). This procedure is likely to silence the CPG altogether and, at the
very least, its ineffectiveness reduces the plausibility that the pathway between pump
cells and RTN neurons involves an interneuron located within the ventrolateral medulla.
As already mentioned, SAR activation reduces the duration of the inspiratory
phase and prolongs expiration by activating E-DEC inhibitory neurons (Hayashi et al.,
1996). In theory, the central apnea caused by higher levels of lung inflation could simply
result from a more intense activation of the E-DEC neurons by the hypothetical
excitatory pump cells located in the NTS. However, present and prior results suggest that
SAR activation may also reduce the frequency and intensity of the respiratory pattern
generator by suppressing the excitatory drive that originates from RTN chemoreceptors
(Moreira et al., 2007). This second mechanism is almost certainly mediated by NTS
pump cells that are inhibitory.
Chemoreceptor stimulation and lung inflation usually exert opposing effects on
the activity of the respiratory network (Hayashi et al., 1996; Coleridge and Coleridge,
2001; Vatner and Uemura, 2001; Kubin et al., 2006). The effect of chemoreceptors and
lung inflation are described as additive or as interacting in more complex ways depending
on the species, the anesthetic and the type of chemoreceptors (central, peripheral or both)
that were stimulated (Mitchell et al., 1982; Mitchell and Selby, 1988; Bajic et al., 1994;
Kubin et al., 2006). The present study suggests that the interaction between the chemical
drive of respiration and its control by lung stretch afferent occurs, at least in part, at RTN
Page 18 of 31

19
level. Since RTN neurons are central chemoreceptors that also receive strong excitatory
inputs from the carotid body (Takakura et al., 2006), RTN should also be viewed as a site
of interaction between lung stretch afferent input and both central and peripheral
chemoreception.
FIGURES LEGENDS
Figure 1: Identification of NTS pump cells.
A1: representative pump cell activated by lung inflation (+2, +4 and +6 cm H2O positive
end-expiratory pressure, PEEP) and silenced by interrupting ventilation in expiration. A2:
effect of lung inflation on the same cell (+6 cm H2O at bar) shown at higher resolution.
A3: peri-event histogram of the activity of the neuron shown in A1, A2 (PEEP: +1 cm
H2O). The cell discharges exclusively during lung inflation as monitored by the tracheal
pressure. B: Discharge rate of all recorded pump cells at normal level of PEEP (+1 cm
H2O) expressed as number of spikes per burst. C: effect of stepwise increments in PEEP
on the number of spikes per burst (ventilation cycle) in a group of 38 pump cells.
Abbreviations: AP, arterial pressure; iPND, integrated phrenic nerve discharge; Tp,
tracheal pressure.
Figure 2: Antidromic activation of NTS pump cells from RTN.
A: example of a pump cell that was antidromically activated from RTN. The asterisk
indicates sweeps when the antidromic spike (a) was absent due to collision with a
spontaneous spike (s) used to trigger the stimulus (stimulus artifact at arrow). B: spikes
Page 19 of 31

20
per burst during resting conditions of pump cells with or without projection to RTN. C:
representative stimulation site in RTN (white blotch at arrow) shown in a coronal
unstained section under darkfield illumination (scale bar: 400 microns). D: computer-
assisted plotting of 8 histologically verified stimulation sites (scale bar: 1 mm).
Abbreviation: P-cells, pump cells; AD-activated, antidromically activated.
Figure 3: Location and GABAergic nature of the pump cells with RTN projections
A: a single biotinamide-labelled pump cell antidromically activated from the RTN (Cy-3
red fluorescence, at arrow marked P cell) is shown superimposed with the darkfield
image of the same section where myelin appears white and neuron rich regions are dark.
The section is coronal and on the left side of the brain (scale bar: 200 µm). B: computer-
assisted plot describing the location of 9 biotinamide-labelled pump cells that were
antidromically activated from the RTN (scale bar: 400 µm). C: region represented by the
rectangle with a dashed line in panel A (C1, fluorescence illumination showing the
biotinamide labelled neuron; C2, image taken under brightfield illumination showing that
the biotinamide-labelled neuron contained GAD67 mRNA (scale bar: 20 µm). Panels
D1, D2 (scale bar: 20 µm in D1 applies to both panels) and E1, E2 (scale bar: 30 µm in
E1 applies to both panels) depict two additional biotinamide-labeled pump cells that were
antidromically activated from RTN and expressed high levels of GAD67 mRNA.
Abbreviations: AP, area postrema; cc, central canal; NTS, nucleus of the solitary tract;
NTS-I, interstitial subnucleus (definition after (Patrickson et al., 1991; Furusawa et al.,
1996)); sol, solitary tract; XII, hypoglossal nucleus.
Page 20 of 31

21
Figure 4: Pump cell region of NTS innervates RTN
A: computer-assisted plot of 4 BDA injection sites (scale bar: 0.5 mm). B: injection site
of biotindextranamide (BDA) centered on the tractus solitarius at area postrema level
(scale bar: 400 µm). C: BDA-labelled terminal fields in a coronal section at the level of
the preBötzinger complex. Each dot is a single varicosity. The 0.5 mm-square box
delineates the region in which varicosities were counted (scale bar: 1 mm). D: BDA-
labelled terminal fields in a coronal section at the level of the RTN. The rectangular box
delineates the region in which varicosities were counted (scale bar: 1 mm). E: low-power
photomicrograph of the BDA-labelled terminal field in the region of RTN (arrow) (scale
bar: 200 µm). F: higher-power photomicrograph of the BDA-labelled terminal field in the
region of RTN (scale bar: 25 µm). G: average number of BDA-labelled terminals per 30
µm section in the RTN region (defined by the rectangular box shown in D) and at two
levels of the ventral respiratory column (defined by the 0.5 x 0.5 mm box shown in C).
Figure 5: Pump cell region of NTS innervates RTN via GABAergic projection.
A-C: example of two GAD-immunoreactive terminals that were anterogradely labelled
with BDA following injection of the tracer in one of the sites shown in the previous
figure (A, BDA; B, GAD65/67 immunoreactivity; C, merged image). Scale bar in panel
A (20 µm) applies to all panels.
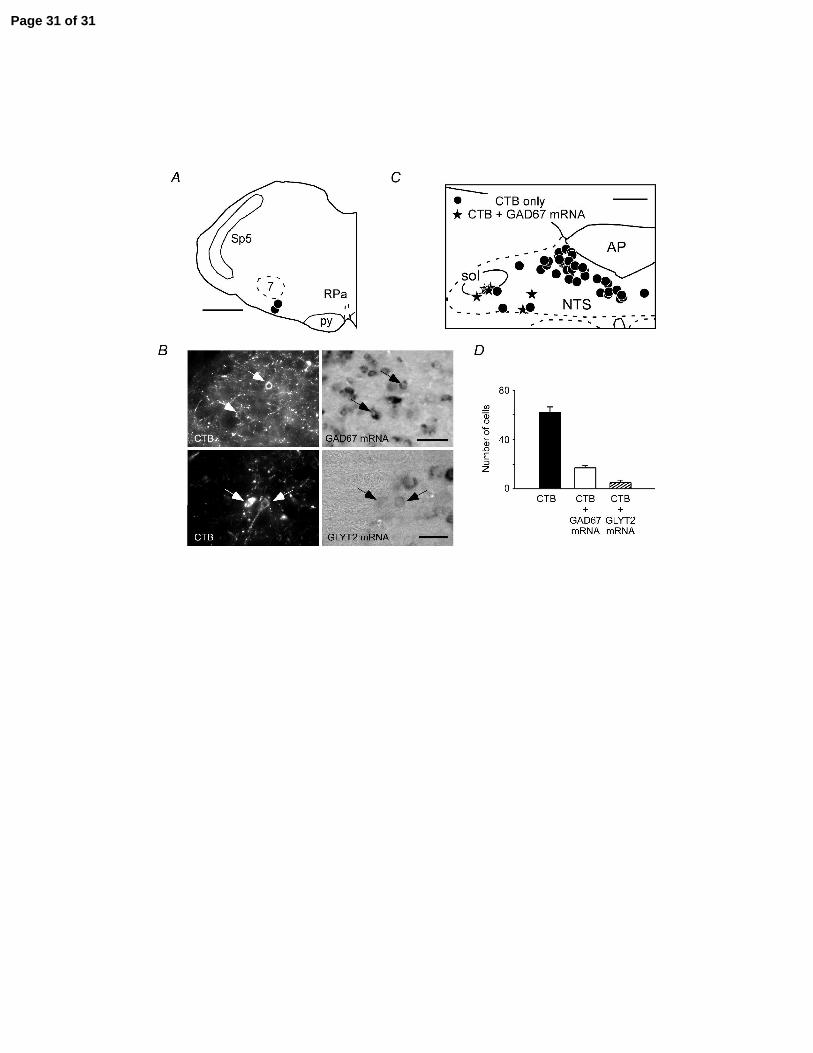
Figure 6: Location of GABAergic and glycinergic neurons with projection to RTN
A: location of the iontophoretic deposits of cholera toxin B (CTB) (scale bar: 0.5 mm). B,
top two panels: example of two CTB-labelled neurons (arrows, left panel, Cy-3
Page 21 of 31

22
fluorescence) located in the interstitial subnucleus of NTS that contained GAD67 mRNA
(ISH seen under tungsten light on right panel) (scale bar: 50 µm). B, lower two panels:
example of two CTB-labelled neurons (arrows, left panel, Cy-3 fluorescence) located in
the interstitial subnucleus of NTS that contained GLYT2 mRNA (ISH seen under
tungsten light on right panel) (scale bar: 30 µm). C: computer-assisted plotting of CTB-
labelled neurons in a representative coronal section located at mid-area postrema level
(scale bar at upper right corner: 200 µm). D: average number of CTB-labelled neurons
within the solitary tract nucleus (cells counted in 3 coronal sections per brain in 2 rats; all
sections were at area postrema level).
REFERENCES
Akilesh MR, Kamper M, Li A, Nattie EE. Effects of unilateral lesions of retrotrapezoid nucleus on breathing in awake rats. J Appl Physiol 82: 469-479, 1997.
Alheid GF, Gray PA, Jiang MC, Feldman JL, McCrimmon DR. Parvalbumin in respiratory neurons of the ventrolateral medulla of the adult rat. J Neurocytol 31: 693-717, 2002.
Altschuler SM, Bao X, Bieger D, Hopkins DA, Miselis RR. Viscerotopic representation of the upper alimentary tract in the rat: sensory ganglia and nuclei of the solitary and spinal trigeminal tracts. J Comp Neurol 283: 248-268, 1989.
Amiel J, Laudier B, Attie-Bitach T, Trang H, de PL, Gener B, Trochet D, Etchevers H, Ray P, Simonneau M, Vekemans M, Munnich A, Gaultier C, Lyonnet S. Polyalanine expansion and frameshift mutations of the paired-like homeobox gene PHOX2B in congenital central hypoventilation syndrome. Nat Genet 33: 459-461, 2003.
Bajic J, Zuperku EJ, Tonkovic-Capin M, Hopp FA. Interaction between chemoreceptor and stretch receptor inputs at medullary respiratory neurons. Am J Physiol 266: R1951-R1961, 1994.
Bonham AC, Coles SK, McCrimmon DR. Pulmonary stretch receptor afferents activate excitatory amino acid receptors in the nucleus-tractus-solitarii in rats. J Physiol (Lond)464: 725-746, 1993.
Bonham AC, McCrimmon DR. Neurones in a discrete region of the nucleus tractus solitarius are required for the Breuer-Hering reflex in rat. J Physiol (Lond) 427: 261-280, 1990.
Page 22 of 31

23
Coleridge HM, Coleridge JC. Afferent innervation of lungs, airways, and pulmonary artery. In: Reflex control of the circulation, edited by Zucker, I. H. and Gilmore, J. P.: CRC Press, 2001, p. 579-607.
Cream C, Li A, Nattie E. The retrotrapezoid nucleus (RTN): local cytoarchitecture and afferent connections. Respir Physiolo Neurobiol 130: 121-137, 2002.
Ezure K, Tanaka I. Pump neurons of the nucleus of the solitary tract project widely to the medulla. Neurosci Lett 215: 123-126, 1996.
Ezure K, Tanaka I. GABA, in some cases together with glycine, is used as the inhibitory transmitter by pump cells in the Hering-Breuer reflex pathway of the rat. Neurosci 127: 409-417, 2004.
Ezure K, Tanaka I, Kondo M. Glycine is used as a transmitter by decrementing expiratory neurons of the ventrolateral medulla in the rat. J Neurosci 23: 8941-8948, 2003.
Ezure K, Tanaka I, Saito Y, Otake K. Axonal projections of pulmonary slowly adapting receptor relay neurons in the rat. J Comp Neurol 446: 81-94, 2002.
Feldman JL, Del Negro CA. Looking for inspiration: new perspectives on respiratory rhythm. Nat Rev Neurosci 7: 232-242, 2006.
Feldman JL, Mitchell GS, Nattie EE. Breathing: Rhythmicity, Plasticity, Chemosensitivity. Annu Rev Neurosci 26: 239-266, 2003.
Furusawa K, Yasuda K, Okuda D, Tanaka M, Yamaoka M. Central distribution and peripheral functional properties of afferent and efferent components of the superior laryngeal nerve: Morphological and electrophysiological studies in the rat. J Comp Neurol 375: 147-156, 1996.
Guyenet PG, Mulkey DK, Stornetta RL, Bayliss DA. Regulation of ventral surface chemoreceptors by the central respiratory pattern generator. J Neurosci 25: 8938-8947, 2005.
Hayashi F, Coles SK, McCrimmon DR. Respiratory neurons mediating the Breuer-Hering reflex prolongation of expiration in rat. J Neurosci 16: 6526-6536, 1996.
Krolo M, Tonkovic-Capin V, Stucke AG, Stuth EA, Hopp FA, Dean C, Zuperku EJ. Subtype composition and responses of respiratory neurons in the pre-botzinger region to pulmonary afferent inputs in dogs. J Neurophysiol 93: 2674-2687, 2005.
Kubin L, Alheid GF, Zuperku EJ, McCrimmon DR. Central pathways of pulmonary and lower airway vagal afferents. Journal of Applied Physiology 101: 618-627, 2006.
Li A, Nattie E. CO2 dialysis in one chemoreceptor site, the RTN: stimulus intensity and sensitivity in the awake rat. Respir Physiol Neurobiol 133: 11-22, 2002.
Lipski J. Antidromic activation of neurones as an analytical tool in the study of the central nervous system. J Neurosci Methods 4: 1-32, 1981.
Mitchell GS, Cross BA, Hiramoto T, Scheid P. Interactions between lung stretch and PaCO2 in modulating ventilatory activity in dogs. J Appl Physiol 53: 185-191, 1982.
Mitchell GS, Selby BD. Ventilatory responses to lung inflation and arterial CO2 in halothane-anesthetized dogs. J Appl Physiol 64: 1433-1438, 1988.
Miyazaki M, Arata A, Tanaka I, Ezure K. Activity of rat pump neurons is modulated with central respiratory rhythm. Neurosci Lett 249: 61-64, 1998.
Miyazaki M, Tanaka I, Ezure K. Excitatory and inhibitory synaptic inputs shape the discharge pattern of pump neurons of the nucleus tractus solitarii in the rat. Exp Brain Res 129: 191-200, 1999.
Page 23 of 31

24
Moreira TS, Takakura AC, Colombari E, West GH, Guyenet PG. Inhibitory input from slowly-adapting lung stretch receptors to retrotrapezoid nucleus (RTN) chemoreceptors. J Physiol (London) 580: 285-300, 2007.
Mulkey DK, Stornetta RL, Weston MC, Simmons JR, Parker A, Bayliss DA, Guyenet PG. Respiratory control by ventral surface chemoreceptor neurons in rats. Nat Neurosci 7: 1360 -1369, 2004.
Nattie E, Li A. Muscimol dialysis in the retrotrapezoid nucleus region inhibits breathing in the awake rat. J Appl Physiol 89: 153-162, 2000.
Nattie E, Shi J, Li A. Bicuculline dialysis in the retrotrapezoid nucleus (RTN) region stimulates breathing in the awake rat. Respir Physiol 124: 179-193, 2001.
Okada Y, Chen Z, Jiang W, Kuwana S, Eldridge FL. Anatomical arrangement of hypercapnia-activated cells in the superficial ventral medulla of rats. J Appl Physiol93: 427-439, 2002.
Okazaki M, Takeda R, Haji A, Yamazaki H. Glutamic acid decarboxylase-immunoreactivity of bulbar respiratory neurons identified by intracellular recording and labeling in rats. Brain Res 914: 34-47, 2001.
Patrickson JW, Smith TE, Zhou SS. Afferent projections of the superior and recurrent laryngeal nerves. Brain Res 539(1): 169-174, 1991.
Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. Academic Press. San Diego, 1998.
Putnam RW, Filosa JA, Ritucci NA. Cellular mechanisms involved in CO(2) and acid signaling in chemosensitive neurons. Am J Physiol Cell Physiol 287: C1493-C1526, 2004.
Richter DW, Ballantyne D, Remmers JE. The differential organization of medullary post-inspiratory activities. Pflugers Arch 410: 420-427, 1987.
Rosin DL, Chang DA, Guyenet PG. Afferent and efferent connections of the rat retrotrapezoid nucleus. J Comp Neurol 499: 64-89, 2006.
Stornetta RL, Guyenet PG. Distribution of glutamic acid decarboxylase mRNA-containing neurons in rat medulla projecting to thoracic spinal cord in relation to monoaminergic brainstem neurons. J Comp Neurol 407: 367-380, 1999.
Stornetta RL, Moreira TS, Takakura AC, Kang BJ, Chang DA, West GH, Brunet JF, Mulkey DK, Bayliss DA, Guyenet PG. Selective expression of Phox2b by brainstem neurons involved in chemosensory integration in the adult rat. J Neurosci26: 10305-10314, 2006.
Stornetta RL, Rosin DL, Wang H, Sevigny CP, Weston MC, Guyenet PG. A group of glutamatergic interneurons expressing high levels of both neurokinin-1 receptors and somatostatin identifies the region of the pre-Botzinger complex. J Comp Neurol 455: 499-512, 2003a.
Stornetta RL, Sevigny CP, Guyenet PG. Inspiratory augmenting bulbospinal neurons express both Glutamatergic and enkephalinergic phenotypes. J Comp Neurol 455: 113-124, 2003b.
Takakura AC, Moreira TS, Colombari E, West GH, Stornetta RL, Guyenet PG. Peripheral chemoreceptor inputs to retrotrapezoid nucleus (RTN) CO2-sensitive neurons in rats. J Physiol (Lond) 572: 503-523, 2006.
Page 24 of 31

25
Vatner SF, Uemura N. Integrative cardiovascular control by pulmonary inflation reflexes. In: Reflex control of the circulation, edited by Zucker, I. H. and Gilmore, J. P.: CRC Press, 2001, p. 609-626.
Page 25 of 31

Page 26 of 31

Page 27 of 31

Page 28 of 31

Page 29 of 31

Page 30 of 31
Page 31 of 31