excitotoxic profiles of novel, low-affinity kainate receptor agonists in primary cultures of murine...
TRANSCRIPT

Neuropharmacology 41 (2001) 421–432www.elsevier.com/locate/neuropharm
Excitotoxic profiles of novel, low-affinity kainate receptor agonistsin primary cultures of murine cerebellar granule cells
S.F. Giardina, P.M. Beart*
Department of Pharmacology, Monash University, PO Box 13E, Clayton, Victoria, 3800, Australia
Received 12 February 2001; received in revised form 4 June 2001; accepted 11 June 2001
Abstract
The involvement of low-affinity kainate (KA) receptors in neuronal injury was investigated by employing a variety of agonistsactive at GluR5–7. Their excitotoxic profiles were determined in primary cultures of cerebellar granule cells, which abundantlyexpressed low-affinity KA receptors, and in the absence of any AMPA receptor-mediated neurotoxicity. Neurotoxicity induced bythese compounds was analysed by phase contrast microscopy, a cell viability assay, the TUNEL technique (apoptosis), and byemploying propidium iodide (PI; necrosis). All agonists induced concentration-dependent neurotoxicity, with rank order (EC50
values; µM): (S)-iodowillardiine (IW) 0.2�(2S,4R)-4-methylglutamate (4-MG) 36�(2S,4R,6E)-2-amino-4-carboxy-7-(2-naphthyl)hept-6-enoic acid (LY339434) 46�KA 74�(RS)-2-amino-3-(hydroxy-5-tert-butylisoxazol-4yl)propanoic acid (ATPA) 88.IW exposure resulted in apoptosis at lower concentrations (�30µM) and necrosis at higher concentrations, both of which wereattenuated by CNQX (50µM), but not MK-801 (10µM). ATPA-mediated neurotoxicity was purely apoptotic and was attenuatedby the non-NMDA receptor antagonists. Both IW and ATPA induced injury with the morphological characteristics of apoptosisshown by the presence of TUNEL-positive neurones. LY339434-mediated neuronal injury was only attenuated by MK-801 andwas necrotic in nature. Similarly, 4-MG (�30 µM) exposure caused necrosis that was partially attenuated by MK-801 (10µM) andCNQX (50µM). The patterns of neurotoxicity possessed a complex pharmacological profile, demonstrated an apoptotic–necroticcontinuum and were inconsistent with past findings, further outlining the importance of characterizing novel compounds at nativereceptors. ATPA and to a lesser extent IW appear to be suitable drugs for low-affinity KA receptors. Since toxicity-mediated bylow-affinity KA receptors seem likely to contribute to neurodegenerative conditions, our study importantly examines the excitotoxicprofile of these novel agonists. 2001 Published by Elsevier Science Ltd.
Keywords: Kainate agonists; Low-affinity kainate receptors; Apoptosis; Excitotoxicity; Cerebellar granule cells
1. Introduction
l-glutamate (Glu) receptor-mediated neurotoxicity,termed excitotoxicity, is well documented, and is likelyto be involved in numerous neurological diseases(Lipton and Rosenberg, 1994; Leist and Nicotera, 1998).Glu receptor-mediated excitotoxicity appears to involvetwo forms of neuronal injury, necrosis and apoptosis(Ankarcrona et al., 1995; Cheung et al., 1998b). Necrosisis a passive process characterized by cellular swelling(Clarke, 1990), whereas apoptosis involves complexmechanisms requiring energy and protein synthesis
* Corresponding author. Tel.:+61-3-9905-3817; fax:+61-3-9905-5851.
E-mail address: [email protected] (P.M. Beart).
0028-3908/01/$ - see front matter 2001 Published by Elsevier Science Ltd.PII: S0028-3908 (01)00086-7
(Dessi et al., 1994). Morphological changes induced byapoptosis are characterized by cell shrinkage, nuclearcondensation, oligonucleosomal fragmentation of DNAand neurite degeneration (Clarke, 1990), and aremediated by various pathways involving caspases andvarious families of kinases (Sastry and Rao, 2000).
Glu acts at two types of receptors the G-proteincoupled metabotropic receptors and the ion-channelgated ionotropic receptors. The ionotropic Glu receptors(iGluRs), which are named after their preferring agon-ists, N-methyl-d-aspartate (NMDA), α-amino-3-hyd-roxy-5-methyl-4-isoxazole propionate (AMPA) andkainate (KA) (Hollmann and Heinemann, 1994), areheavily implicated in numerous neurodegenerative andacute neurological conditions (Lipton and Rosenberg,1994; Leist and Nicotera, 1998). Cloning experimentshave played an essential role in the understanding of the

422 S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
various Glu receptor subunits and their ion per-meabilities, and have revealed a number of families ofsubunits consisting of GluR1-4 (AMPA), NR1 andNR2A-D (NMDA), KA1-2 (high affinity kainatereceptors) and GluR5-7 (low-affinity kainate receptors)(Hollmann and Heinemann, 1994). Low-affinity KAreceptors are widely distributed throughout the rodentand primate brain (Wisden and Seeburg, 1993; Carrollet al., 1998; Bernard et al., 1999) and have been associa-ted with various disease states including Alzheimer’sdisease (Aronica et al., 1998), schizophrenia (Breese etal., 1995), amyotropic lateral sclerosis, cerebral ischae-mia and epilepsy (Paschen et al., 1996; Bernard et al.,1999). Other evidence indicates that low-affinity KAreceptors are likely to play significant physiological rolesin synaptic transmission, with involvements in long termpotentiation (Clarke et al., 1997; Vignes and Colling-ridge, 1997; Bortolotto et al., 1999) and pain processing(Procter et al., 1998). Recently novel Glu analogues havebeen developed and studies employing recombinantreceptor model systems have demonstrated their selec-tivity for low-affinity KA receptors subunits (Jane et al.,1997; Zhou et al., 1997; Bleakman and Lodge, 1998).These compounds include ATPA, IW, LY339434 and 4-MG (also commonly termed SYM 2081) (Bleakman andLodge, 1998).
Recombinant receptor studies have revealed some ofthe physiological and binding characteristics of theseGlu analogues. Amino-3-(hydroxy-5-tert-butylisoxazol-4yl)propanoic acid (ATPA), an analogue of AMPA, isan effective AMPA agonist, with activity at GluR5 butnot GluR6 (Clarke et al., 1997; Wahl et al., 1998), whileIW, like other willardiines, has some AMPA activity(Patneau et al., 1992), but is also a potent GluR5 receptoragonist (Jane et al., 1997). Binding studies have revealedthat 4-MG has higher affinity for GluR6 than NMDA orAMPA receptors and displaces [3H]KA binding moreeffectively than KA itself (Zhou et al., 1997). Whileligand binding studies demonstrated that LY339434 is1000-fold more selective for GluR5 than GluR6 (Smallet al., 1998), functional studies have demonstratedactivity at AMPA and NMDA receptors (Small et al.,1998; Moldrich et al., 2000b). Limited studies have beenreported on the excitotoxic profiles of 4-MG, ATPA, IWand LY339434, with investigations conducted in primarycultures of cortical neurones demonstrating that thesenovel compounds exhibited complex excitotoxic pro-files, involving actions not only at KA receptors, but alsoat AMPA and NMDA receptors (Small et al., 1998; Mol-drich et al., 2000a,b).
The current study examines these putative compoundsin primary cultures of cerebellar granule cells (CGCs).CGCs are a particularly useful model system in manyrespects, as they are homogenous cell preparation con-taining �95% neurones when maintained under serum-free conditions (Cheung et al., 1998a) and at 8 d in vitro
possess no functional AMPA receptors (Hack et al.,1995). CGCs cultured under the present conditions arepresumed to be glutamatergic (Fonnum, 1984) since theyactively accumulate d[3H]aspartate and not d[3H]GABA(Carroll et al., 1997). In addition, the current model sys-tem is particularly sensitive to KA receptor-mediatedtoxicity (Cheung et al., 1998a; Giardina et al., 1998).This study demonstrates for the first time the excitotoxicprofiles of these novel compounds at native KA recep-tors, in an AMPA receptor-free environment, using pri-mary cultures of murine CGCs.
2. Materials and methods
2.1. Reagents
KA, 4-MG, ATPA, AMPA, NMDA and 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX) were purchasedfrom Tocris Cookson (Bristol, UK). Neurobasalmedium (NBM), B27 nutrients, N2 supplements andCa2+-free-Hank’s balanced salt solution (HBSS) werepurchased from GibcoBRL Life Technologies(Melbourne, Australia). 1-(4-aminophenyl)-4-methyl-7,8-methylendioxy-5H-2,3-benzodiazepine (GYKI52466) and (5R,10S)-(+)-5-methyl-10,11-dihydro-5H-dibenzo[a,d]cyclohepten-5,10-imine (MK-801) werepurchased from Sigma RBI (Sydney, Australia).LY339434 was a kind gift from Dr R. Baker (Eli Lilly;Windlesham, UK). KA2 and GluR6/7 antibodies werepurchased from Upstate Biotechnology (Lake Placid,USA). All other reagents were purchased from Sigma orBoehringer Mannheim (Sydney, Australia) and were ofcell culture or molecular biology grade.
Experiments were performed in accordance with theethical code of the National Health and MedicalResearch Council (Australia) with permission from thestanding Committee for Ethics in Animal Experimen-tation (Monash University).
2.2. Cell culture
CGCs were prepared from 7 d old Swiss White miceand cultured as previously described (Cheung et al.,1998a; Giardina et al., 1998). CGCs were grown inNBM containing B27 components (Cheung et al.,1998a), 25.4 mM K +, 500 µM Glu and 100 U/ml penicil-lin–streptomycin and exposed to 10% dialysed fetal calfserum for the first 24 h and left in serum-free conditionsfrom day in vitro (div) 1. Cells were seeded at a celldensity of 0.3×106 cells/cm2 in 24 well NUNC plates(Denmark) precoated with poly-d-lysine (50 µg/ml).Aphidicolin (1 µg/ml) was added to the medium 18–24 hafter plating to inhibit non-neuronal cell proliferation(Giardina et al., 1998). Immunocytochemistry previously

423S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
established that �95% of the cells were neurones(Cheung et al., 1998a).
2.3. Immunocytochemical identification of KA receptorsubunits
CGCs were fixed in 4% paraformaldehyde in 0.1 Mphosphate buffered saline (pH 7.4) at room temperaturefor 15 min. Cultures were subsequently washed in Trisbuffered saline (TBS; 50 mM Tris, 0.9% NaCl; pH 7.6),permeabilized in 2% Triton X-100 (TX-100) for 5 min,and blocked in 10% normal goat serum (NGS), 0.1%TX-100 in TBS for 1 h at 4°C. Cells were then incubatedwith the primary antibodies, rat polyclonal anti-KA2(1:1000 dilution) or rat polyclonal anti-GluR6/7 (1:1000)in solution with 2% NGS and 0.1%TX-100 in TBS,overnight at 4°C. CGCs were subsequently washed inTBS and incubated with anti-rabbit-IgG horse radishperoxidase conjugated secondary antibody (1:1000) for3 h at room temperature. After further washing, cultureswere incubated with 3�,3�-diaminobenzidine in stableperoxidase buffer and colour was allowed to develop for1 h at room temperature. Immunopositive labelling wasvisualized by bright field microscopy with an Olympusinverted microscope (Olympus, IMT-2) and photo-graphed using Kodak Gold 100 film.
2.4. Agonist exposure and cell viability assays
Cultures were exposed to the low-affinity KA agonists(10–1000 µM) alone or in the presence of the non-NMDA receptor antagonists CNQX (50 µM) and GYKI52466 (20 µM), or the NMDA receptor antagonist MK-801 (10 µM), for 24 h at div 8 in N2 supplemented NBMcontaining 100 U/ml penicillin–streptomycin, 0.25%BSA, 83 µM d(+) galactose, 16 µM ethanolamine, 6 µMl-carnitine, 0.4 µM biotin and 25.4 mM K+ (Giardina etal., 1998). Excitotoxicity mediated via the AMPA andNMDA receptors in the cultures was examined byexposure to these agonists (10–1000 µM) on div 8 underthe same conditions used for the low-affinity KA studies.Cellular viability was determined at 24 h by thereduction of 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) (Cheung et al., 1998a),which is a measure of mitochondrial function which iscomprised in necrotic and apoptotic cells (Kroemer etal., 1998). MTT was incubated with the cells for 30 minat 37°C and the reduced formazan product was lysedfrom the cells in 20% sodium dodecyl sulphate and 40%dimethylformamide, and absorbance was subsequentlymeasured at 590 nm (Ceres UV900c microplate reader;Biotek Instruments, USA). Cultures treated with 500 µMGlu for 24 h were taken as 100% cell death and theresults were expressed as percentage of vehicle control.
Morphological changes in neurones after exposure tothe iGluR agonists were observed under an inverted light
microscope (Olympus, IMT). Cellular swelling 1–4 hafter treatment, and a decrease in cell density with evi-dence of cellular debris 24 h after treatment was con-sidered necrosis (Clarke, 1990). Apoptosis was morpho-logically characterized by cellular shrinkage, thebreakdown of neurites (neurite blebbing) with no distinctloss of cell density 24 h after exposure to excitotoxins(Philpott et al., 1996).
2.5. Propidium iodide labelling of necrotic cells
Necrotic cell death was analysed by propidium iodide(PI) labelling of living cells with damaged cell mem-branes (Moldrich et al., 2000a). PI (1 µg/ml) was incu-bated at div 8 1–2 h after agonist exposure for 30 minat 37°C as previously described (Moldrich et al., 2000a).Cultures were subsequently viewed using fluorescentmicroscopy and random fields photographed with KodakEktachrome P1600 film.
2.6. In situ labelling of nuclear DNA fragments
Apoptosis was analysed by terminal deoxynucleotidyltransferase (TdT)-mediated dUTP digoxigenin nick endlabelling (TUNEL) as previously described (Cheung etal., 1998a). CGCs were fixed overnight in 4% paraform-aldehyde, and subsequently permeabilized with 2% Tri-ton X-100 (TX-100) in TBS. Cultures were subsequentlywashed in TBS and blocked for 1 h at room temperaturein solution with 10% normal goat serum and 0.1% TX-100 in TBS and then incubated with TdT reaction mix-ture for 3 h at 37°C as previously described (Cheung etal., 1998a). DIG labelled dUTP was detected by anti-DIG alkaline phosphatase (AP; 1:1000 dilution) in sol-ution with 2% normal goat serum and 0.1% TX-100 andTBS. TUNEL-positive cells were detected using AP sub-strate solution (0.4 mg/ml 5-bromo-4-chloro-3-indolyl-phosphate; 0.6 mg/ml nitroblue tetrazolium chloride,100 mM Tris–HCl, 0.5 mM MgCl2; pH 9). Control cul-tures included the above treatment with the omission ofTdT. Cells were visualized under bright field microscopyand random fields photographed.
2.7. Data analyses
Data are given as mean±S.E.M. from at least quad-ruplicate experiments across 4–6 independent culturesand concentration–response curves were generated bynon-linear regression using computer-assisted curve fit-ting (GraphPad Prism ). Statistical significance(p�0.05) of data was examined by two way ANOVA.Characteristics of cell death, determined by PI incorpor-ation and TUNEL labelling, were conducted over 2–3independent experiments and representative photomicro-graphs are presented here.
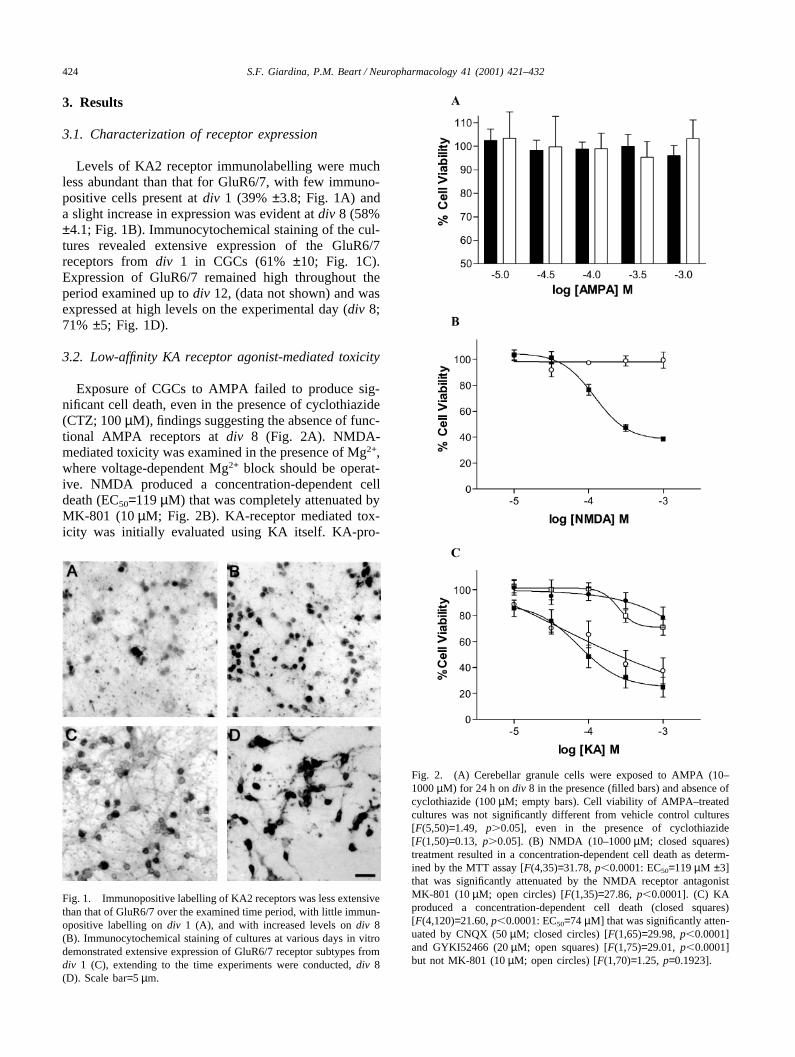
424 S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
3. Results
3.1. Characterization of receptor expression
Levels of KA2 receptor immunolabelling were muchless abundant than that for GluR6/7, with few immuno-positive cells present at div 1 (39% ±3.8; Fig. 1A) anda slight increase in expression was evident at div 8 (58%±4.1; Fig. 1B). Immunocytochemical staining of the cul-tures revealed extensive expression of the GluR6/7receptors from div 1 in CGCs (61% ±10; Fig. 1C).Expression of GluR6/7 remained high throughout theperiod examined up to div 12, (data not shown) and wasexpressed at high levels on the experimental day (div 8;71% ±5; Fig. 1D).
3.2. Low-affinity KA receptor agonist-mediated toxicity
Exposure of CGCs to AMPA failed to produce sig-nificant cell death, even in the presence of cyclothiazide(CTZ; 100 µM), findings suggesting the absence of func-tional AMPA receptors at div 8 (Fig. 2A). NMDA-mediated toxicity was examined in the presence of Mg2+,where voltage-dependent Mg2+ block should be operat-ive. NMDA produced a concentration-dependent celldeath (EC50=119 µM) that was completely attenuated byMK-801 (10 µM; Fig. 2B). KA-receptor mediated tox-icity was initially evaluated using KA itself. KA-pro-
Fig. 1. Immunopositive labelling of KA2 receptors was less extensivethan that of GluR6/7 over the examined time period, with little immun-opositive labelling on div 1 (A), and with increased levels on div 8(B). Immunocytochemical staining of cultures at various days in vitrodemonstrated extensive expression of GluR6/7 receptor subtypes fromdiv 1 (C), extending to the time experiments were conducted, div 8(D). Scale bar=5 µm.
Fig. 2. (A) Cerebellar granule cells were exposed to AMPA (10–1000 µM) for 24 h on div 8 in the presence (filled bars) and absence ofcyclothiazide (100 µM; empty bars). Cell viability of AMPA–treatedcultures was not significantly different from vehicle control cultures[F(5,50)=1.49, p�0.05], even in the presence of cyclothiazide[F(1,50)=0.13, p�0.05]. (B) NMDA (10–1000 µM; closed squares)treatment resulted in a concentration-dependent cell death as determ-ined by the MTT assay [F(4,35)=31.78, p�0.0001: EC50=119 µM ±3]that was significantly attenuated by the NMDA receptor antagonistMK-801 (10 µM; open circles) [F(1,35)=27.86, p�0.0001]. (C) KAproduced a concentration-dependent cell death (closed squares)[F(4,120)=21.60, p�0.0001: EC50=74 µM] that was significantly atten-uated by CNQX (50 µM; closed circles) [F(1,65)=29.98, p�0.0001]and GYKI52466 (20 µM; open squares) [F(1,75)=29.01, p�0.0001]but not MK-801 (10 µM; open circles) [F(1,70)=1.25, p=0.1923].

425S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
duced a concentration-dependent cell death(EC50=74 µM) that was significantly attenuated by thenon-NMDA receptor antagonists CNQX (50 µM) andGYKI 52466 (20 µM), but not by MK-801 (10 µM;Fig. 2C).
The present study found widespread, concentration-dependent cell death mediated by the putative low-affin-ity KA receptor agonists (Fig. 3). Rank order potencyand EC50 values (µM): IW (0.2)�4-MG(36)�LY339434 (46)�KA (74)�ATPA (88)[F(4,205)=32.24, p�0.0001]. These data coincide withthe rank order for excitotoxicity found in primary cul-tures of murine cortical neurones (Moldrich et al.,2000a).
ATPA-mediated toxicity was somewhat less potent
Fig. 3. Neurotoxicity mediated by putative low-affinity KA receptor agonists in cultured cerebellar granule cells was determined using an MTTcell viability assay. Concentration–response curves were generated for ATPA (A), IW (B), LY339434 (C), and 4-MG (D). Low-affinity KA agonistsalone are represented by a solid square, in the presence of CNQX (50 µM) by a solid circle, in the presence of GYKI 52466 (20 µM) by an opensquare and in the presence of MK-801 (10 µM) by an open circle for each representative graph. 4-MG in the presence of a combination of MK-801, GYKI52466 and CNQX is represented by a closed diamond (D). ATPA did not produce a concentration-dependent cell death [F(4,115)=2.29,p=0.46: EC50=88 µM] although the response was significantly different to concentration–response curves produced in the presence of CNQX andGYKI 52466 [F(2,95)=18.17, p�0.0001] (A). IW produced a concentration-dependent cell death [F(4,115)=15.29, p�0.0001: EC50=0.2 µM] thatwas significantly different to curves produced in the presence of CNQX and GYKI52466 [F(2,90)=21.22, p�0.0001], but not MK-801 [F(1,70)=0.02,p=0.875] (B). LY339434 produced a concentration-dependent cell death [F(4,145)=33.89, p=0.46: EC50=46 µM] that was attenuated by MK-801[F(1,80)=46.10, p�0.0001], but not CNQX and GYKI 52466 [F(2,104)=1.03, p=0.1135] (C). Cell death produced by 4-MG was concentration-dependent [F(4,140)=77.28, p�0.0001: EC50=36 µM] and was significantly attenuated, at lower concentrations of 4-MG, by all the iGluR antagonists[F(4,140)=5.33, p�0.0001] (E).
than that induced by the other compounds (Fig. 3A),with a maximal cell death of 25% (1000 µM) comparedto that of 100% cell death induced by 4-MG (1000 µM;Fig. 3C), 50% by IW (1000 µM; Fig. 3B) and 90% byLY339434 (1000 µM; Fig. 3D). Like KA, the neurotox-icity produced by ATPA was completely attenuated byboth the non-NMDA receptor antagonists, GYKI 52466(20 µM) and CNQX (50 µM), but not by MK-801(10 µM). The receptors involved in mediating IW tox-icity were of the non-NMDA subtype, where toxicitywas attenuated completely by CNQX and partially byGYKI 52466 (Fig. 3B). In contrast to previous findings(Moldrich et al., 2000a), MK-801 did not attenuate IW-mediated toxicity, [F(1,70)=0.02, p=0.88], implying thatIW acts through non-NMDA receptors in the current

426 S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
model system. Neurotoxicity mediated by high concen-trations of 4-MG was not attenuated by MK-801, CNQXor GYKI 52466 to any great extent implicating non-Glu-receptor mediated mechanisms by which toxicity occurs(Fig. 3C). However, injury induced by lower concen-trations of 4-MG (�30 µM) was significantly attenuatedby 10 µM MK-801 [F(1, 70)=5.01, p�0.0001], 50 µMCNQX [F(1,70)=1.34, p�0.05], but not 20 µM GYKI52466 [F(1,70)=0.45, p=0.77]. Combinations of theiGluR antagonists were employed to overcome the inef-fectiveness of the antagonists alone, and to evaluatewhether 4-MG induced neurotoxicity was mediated byboth NMDA and KA receptors. While the concen-tration–response curve to 4-MG is significantly shiftedto the right in the combined presence of CNQX (50 µM),GYKI 52466 (20 µM) and MK-801 (10 µM)[F(1,70)=9.77, p�0.0001], attenuation of neurotoxicitywas greatest at lower concentrations of 4-MG(�100 µM) and is only partial at higher concentrations(Fig. 3D). Neurotoxicity mediated by LY339434 wascompletely attenuated by MK801 (10 µM;[F(1,80)=46.10, p�0.0001]), and while the concen-tration–response curve was slightly shifted to the rightby the non-NMDA receptor antagonists GYKI andCNQX (Fig. 3D), this effect was not statistically signifi-cant (p�0.05).
3.3. Morphology of neurotoxicity induced by low-affinity KA receptor agonists
Morphological examination of the cultures by lightmicroscopy was undertaken to correlate cellular andpathological changes with the MTT injury data. AMPAexposure (300 µM) in the presence of CTZ (100 µM)demonstrated no significant neuronal injury at div 8 (Fig.4D), in agreement with MTT injury data. NMDA largelyproduced cellular swelling at higher concentrations(�100 µM; Fig. 4E), while little cellular damage wasevident at lower concentrations, implying NMDA recep-tors were functional in CGCs at this time in vitro. KA-mediated cell death induced negligible cellular swelling1–2 h after exposure (data not shown), but 24 h afterstimulation cells appeared shrunken and neurite net-works had broken down (Fig. 4B; 100 µM), and wascompletely attenuated by CNQX (50 µM; Fig. 4C). Theputative low-affinity KA receptor agonists producedconcentration-dependent neuronal damage, demonstrat-ing morphological changes consistent with apoptosis andnecrosis, dependent upon the intensity of the insult.Cellular swelling, indicative of necrosis, was evident incultures 1–2 h after exposure to 4-MG (10–1000 µM),LY339434 (100–1000 µM), IW (30–1000 µM), and KA(300–1000 µM) (data not shown). Cellular swelling wasnot evident in cells treated with lower concentrations ofIW, LY339434, KA or any concentrations examined forATPA (10–1000 µM). After exposure (24 h) of the cul-
tures to lower concentration of IW (10–30 µM),LY339434 (10–100 µM), KA (10–300 µM), and all con-centrations of ATPA (10–1000 µM; Fig. 4F), some neu-rones demonstrated neurite blebbing and cellular shrink-age, consistent with apoptosis. IW-mediated toxicity(100 µM; Fig. 4G) demonstrated morphological charac-teristics of both necrosis and apoptosis and was com-pletely attenuated by CNQX (Fig. 4H) and GYKI 52466(Fig. 4I), indicating a KA receptor response. Culturestreated with high concentrations of LY339434, IW, KA,and all concentrations of 4-MG (Fig. 4J) possessed highlevels of debris and a diminished cell density 24 h afterstimulation, representative of necrosis. High concen-trations of LY339434 (300 µM; Fig. 4K) produced rapidcellular swelling 1–2 h after exposure (necrosis) andafter 24 h remaining cells demonstrated shrunken cellbodies indicative of apoptosis. Neurotoxicity mediatedby LY339434 was completely attenuated in the presenceof MK-801 (Fig. 4L), but not the non-NMDA receptorantagonists (data not shown). These findings are summa-rized in Table 1.
3.4. Low-affinity KA receptor agonists: mode of celldeath
The DNA binding dye, PI, only penetrates cells withdamaged cellular membranes and when used after shortincubation times is a reliable index of rapid necrosis,especially when compared with findings from analysesusing the TUNEL technique for apoptosis. KA produceda slow cell death that was predominantly apoptotic innature as shown by the widespread presence of TUNEL-positive profiles (Fig. 5B) which were greatly decreasedby GYKI 52466 (Fig. 5C). Essentially no incorporationof PI was seen with KA (data not shown) in confirmation
Fig. 4. Photomicrographs of primary cultures of cerebellar granulecells treated with iGluR agonists 24 h after exposure revealed a com-plex mosaic of responses dependent upon the agonist itself and theconcentration. Treatment with 100 µM KA (B) showed typical charac-teristics of apoptosis including cell shrinkage and neurite blebbing,when compared to the control (A), that was attenuated by CNQX (C).AMPA (300 µM) in the presence of CTZ (100 µM; D) induced nomorphological change while NMDA (300 µM)-mediated toxicity dem-onstrated characteristics of necrosis and apoptosis, with a markedreduction in cell density and the presence of cellular debris (necrosis)and remaining cells appeared shrunken (E). ATPA (300 µM; F), whileless potent than KA, produced morphological changes that were indica-tive of apoptosis. Toxicity mediated by IW (100 µM; G) demonstratedmarked cellular loss (necrosis) and remaining cells appear shrunken(apoptosis). Toxicity induced by IW was completely attenuated by thenon-NMDA receptor antagonists CNQX (50 µM; H) and GYKI 52466(20 µM; I). 4-MG-induced toxicity was rapid and resulted in a markedreduction in cell density, indicative of necrosis (100 µM; J). Toxicitymediated by LY334394 (100 µM; K), was largely necrotic in natureand was not attenuated by the non-NMDA receptor antagonists andcompletely by the NMDA receptor antagonist MK-801 (10 µM; L).Photomicrographs presented here are representative of 4–6 inde-pendent experiments. Scale bar=5 µm.

427S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
of the microscopic observations that necrotic cells wererarely found. The modes of cell death mediated by theputative low-affinity KA receptor agonists were some-what different to that found for KA itself. Neurotoxicitymediated by ATPA was apoptotic rather than necrotic
in nature as determined by the increased presence ofTUNEL-positive cells (300 µM; Fig. 5D), when com-pared to the vehicle control (Fig. 5A), with few PI-posi-tive cells observed 1–2 h after stimulation (data notshown). Neuronal injury patterns found with IW and

428 S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
Table 1Neuronal injury patterns induced by the putative low-affinity kainate agonists are as indicated. Characteristics of apoptosis (neurite blebbing andcellular shrinkage) and necrosis (cellular swelling and loss of cellular density) were analysed by phase contrast microscopy, TUNEL (apoptosis)and PI incorporation (necrosis). The intensity of staining was graded as follows: +++ many of the cells are staining positive; ++ significant portionof cells are staining positive; + staining slightly more intense than that of the vehicle control; – represents no significant change when comparedto vehicle control. * indicates AMPA in the presence of cyclothiazide (100 µM)
Agonist concentration (µM) Morphological characteristics TUNEL labelling Propidium iodide labelling
KA (10–300 µM) Shrinkage, neurite blebbing +++ –KA (1000 µM) Shrinkage, neurite blebbing, some swelling and +++ +
reduction in densityNMDA (10–100 µM) Shrinkage, neurite blebbing +++ +NMDA (100–1000 µM) Swelling, reduction in density +++ +++AMPA* (10–1000 µM) No morphological changes – –ATPA (10–1000 µM) Shrinkage, neurite blebbing ++ –IW (10–30 µM) Shrinkage, neurite blebbing +++ –IW (30–1000 µM) Swelling, reduction in density + ++4-MG (10–30 µM) Shrinkage, neurite blebbing ++ +4-MG (30–1000 µM) Swelling, reduction in density + +++LY339434 (10–100 µM) Shrinkage, neurite blebbing +++ +LY339434 (100–1000 µM) Swelling, reduction in density + ++
LY339434 were more complex, demonstrating apoptotic(TUNEL-positive) nuclei in cells treated with lower con-centrations (Fig. 5E and J), and labelling with PI indica-tive of necrosis at higher concentrations (Fig. 5H and L). IW and LY339434 mediated TUNEL labelling wascompletely attenuated by CNQX (Fig. 5F) and MK-801(Fig. 5K), respectively. IW exposure, at high concen-trations, led to necrosis with the majority of cellsincorporating PI (Fig. 5G) 1–2 h after exposure. 4-MG-mediated toxicity was purely necrotic in nature as shownby wide-spread labelling of cells with PI (Fig. 5I) andfew cells staining TUNEL-positive for apoptosis 24 hafter stimulation (data not shown). These findings aresummarized in Table 1.
4. Discussion
The current study is the first to analyse the excitotoxicprofile of the putative low-affinity KA receptor agonistsATPA, IW, 4-MG and LY339434 in an essentiallyAMPA-receptor free environment. CGCs are a parti-cularly useful model system for pharmacological evalu-ation of compounds as they are an essentially homogen-ous cell preparation under serum-free conditions, with anegligible glial cell population, and naturally express KAreceptor subunits GluR5/6 and high affinity KA subunitsin agreement with earlier findings (Belcher and Howe,1997; Pemberton et al., 1998). While KA2 and GluR6expression occurs in equal abundance in the intact cere-bellum (Porter et al., 1997), the present study demon-strated differing abundance of the receptor subunitsdependent upon the time in culture. This difference ismost likely to be due to culture conditions not entirelymimicking the in vivo system, and directly affecting
receptor expression as previously described (Resink etal., 1994). The higher abundance of GluR6/7 expressionsuggests that low-affinity KA receptors are likely to bethe site of action of the various KA agonists. Previousstudies from our laboratory have found that KA exposurein CGCs results in apoptosis with TUNEL-positive neu-rones concomitant with increased expression of twoapoptotic indices (Kroemer et al., 1998; Sastry and Rao,2000), BAX (Giardina et al., 2000) and c-Jun (Cheunget al., 1998a) — moreover, for all three parameters theelevations were concentration- and CNQX-dependent.Together these findings indicate that the TUNEL datafrom the current study are consistent with the neurotox-icity mediated by the KA receptor is apoptotic in nature.
These cultures also do not express functional AMPAreceptors at the time in culture employed, as AMPAfailed to exhibit any loss of cell viability even in thepresence of CTZ, conditions that result in readily detect-able excitotoxicity in cortical cultures (Jensen et al.,1998; John et al., 1999), and data corresponding to pre-vious findings (Hack et al., 1995). Therefore the currentmodel system is particularly favourable to examine KAreceptor-mediated responses. Under these experimentalconditions the putative low-affinity KA agonists pro-duced neuronal cell death that was concentration-depen-dent with the rank order of potency being IW�4-MG�LY339434�KA�ATPA. The patterns of injuryproduced by the various agonists showed considerablevariation involving apoptosis and/or necrosis apparentlydependent on their different pharmacological activitiesat KA and/or NMDA receptors.
Under the present experimental conditions IW andATPA seemed likely to exert a considerable portion oftheir excitotoxicity via KA receptors in CGCs, given theneuroprotective actions of the receptor antagonists and

429S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
Fig. 5. The mode of cell death mediated by the putative low-affinity KA agonists was examined by TUNEL (apoptosis; after 24 h exposure) andPI incorporation (necrosis; 1–2 h after exposure), by bright field and fluorescent microscopy, respectively. Typical apoptotic neurones are indicatedby the black deposits (black arrows; B), while typical necrotic neurones by white filled cells (G–I, L). KA (100 µM, B), produced an increase inthe number of TUNEL positive profiles when compared to the control (A), that was attenuated by the non-NMDA receptor antagonist GYKI 52466(20 µM, C). ATPA (300 µM) produced an increase in TUNEL-positive cells (D), and cultures treated with IW (30 µM; E) demonstrated a markedincrease in the number of TUNEL-positive cells that was attenuated by (CNQX, F). Higher concentrations of IW (300 µM, H) induced a markedincrease in the incorporation of PI, indicative of necrosis relative to control cultures (G). 4MG (100 µM, I) mediated toxicity was largely necroticin nature with positive cells incorporating PI, when compared to the vehicle control (G). LY339434 produced an increase in TUNEL-positiveprofiles at lower concentrations (30 µM, J) that were attenuated by the presence of MK-801 (10 µM, K), whereas at a higher concentration (300 µM)a moderate density of PI-positive profiles were noted (L) indicating necrosis. Scale bar=5 µm.

430 S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
the abundant expression of these receptors as determinedby immunocytochemistry. ATPA was somewhat lesspotent than IW and has previously been shown to be aquite potent AMPA receptor agonist (Stensbol et al.,1999), whilst IW has recently been suggested to possessactivity at NMDA receptors in primary cortical cultures(Moldrich et al., 2000b). Since MK-801 was ineffectiveat preventing IW-mediated neurotoxicity, such a NMDArelated effect was not operative in CGCs. This discrep-ancy may not be due to the immediate action of IW, butrather a secondary response of necrotic cell death leadingto the release of Glu, which subsequently acts on neighb-oring NMDA receptors to elucidate further toxicity incortical neurones. While IW and ATPA were more andless potent than KA itself, respectively, neurotoxicitymediated by these compounds was not attenuated byMK-801. Indeed, NMDA under the present Mg2+-con-taining conditions was a weak excitotoxin only causingappreciable cell death at concentrations �100 µM. Thusthe experimental milieu would appear to mitigate againstany secondary mechanism subsequently dependent uponNMDA receptor activation. While KA producedapoptosis at lower concentrations and necrosis was a rareevent only evident at quite high concentrations, IW-mediated cell death was largely necrotic in nature exceptat low concentrations (�30 µM). ATPA did not produceany necrotic death at the concentrations examined andthe cell death that was evident was apoptotic, as determ-ined by TUNEL labelling and morphological examin-ation. The differences in the toxicity profiles of ATPAand IW, when compared to the toxicity mediated by KA,are likely to be explained by their differing affinitiesthese agents have for KA receptors, rather than differentsites of action. Since CNQX and GYKI 52466 protectedneurones against IW- and ATPA-mediated toxicity it canbe suggested that these agonists are acting on KA recep-tors. While GYKI 52466 has higher affinity for AMPAreceptors, but also acts on KA receptors (Donevan andRogawski, 1993), and these receptors are absent fromCGCs (Hack et al., 1995), it can be assumed that theprotection produced by GYKI 52466 is from activity atKA receptors.
4-MG has previously been shown to be a selective andpotent GluR5/6 receptor agonist in studies that expressedrecombinant receptors in HEK293 cells (Zhou et al.,1997). The present study demonstrated the toxicitymediated by 4-MG was not mediated to any great extent(�30% total injury) by any of the iGluRs, as non-NMDA and NMDA receptor antagonists failed to attenu-ated toxicity except at concentrations �30 µM. Oneexplanation for this discrepancy may be the blockingactivity of 4-MG at Glu transporters (Vandenberg et al.,1997). Blocking Glu transporter hinders uptake of Glu,resulting in an accumulation of extracellular Glu that inturn is free to act on various Glu receptors, causing exci-totoxic damage. While damage caused by this mech-
anism should be attenuated by the various Glu receptorantagonists, such an effect was not evident in the presentstudy, again indicating a non-iGluR mediated mech-anism of toxicity for 4-MG, perhaps involving cellularswelling and ultimately necrosis, that arises from the dis-ruption of associated ion transport (Kroemer et al.,1998).
Neurotoxicity mediated by LY339434, although con-sistent with studies conducted at native Glu receptors(Small et al., 1998; Moldrich et al., 2000a,b), contrastedwith studies conducted in recombinant receptor modelsystems (Small et al., 1998). Excitotoxicity mediated byLY339434 was only attenuated by MK-801, with no sig-nificant protection from the non-NMDA receptor antag-onists, and thus was likely to be only mediated byNMDA receptors. While NMDA-induced toxicity wassignificantly less than observed with LY339434, it wasalso mediated purely by the NMDA receptor. The pres-ence of Mg2+ in the medium during the experimentaltimes NMDA did not inhibit the ability of NMDA tocause neuronal injury, implying the NMDA receptor wasable to overcome the depolarization-dependent Mg2+
block to induce neuronal injury. How LY339434 andNMDA overcome the Mg2+ block to allow NMDAreceptor stimulation is unknown, but this effect mayhave occurred by an initial stimulation of the non-NMDA receptors, resulting in membrane depolarization,with release of the Mg2+ block and subsequentlyallowing the activation of the receptor. While previousstudies have shown LY339434 to be a selective GluR5agonist (Small et al., 1998) the current study did not findany evidence in support. Rather LY339434 was foundto be a potent NMDA receptor agonist with potent neur-otoxic properties.
While the pharmacological profiles of these agonistswere quite different, all produced an apoptotic–necroticcontinuum of cell death, except for ATPA, where thepattern of neuronal injury only exhibited apoptoticcharacteristics. Morphologically apoptotic cell bodieswere evident at low concentrations for all the agonists,with evidence of shrunken cell bodies and neurite bleb-bing (Clarke, 1990). Apoptosis is a complex mechanisminvolving cascades of events largely initiated by anincrease in intracellular Ca2+ (Lipton and Nicotera, 1998;Toescu, 1998) leading to the activation of various casp-ases and kinases which mediate the demise of the cell(Sastry and Rao, 2000). Hence apoptosis is a slowermechanism than necrotic cell death and cellular damageis not evident within the first hour of stimuli, unlikenecrosis, allowing the manipulation of labelling withvarious dyes that only penetrate cells with damagedmembranes at earlier time points for identification ofnecrotic cells. Necrotic cell death, as demonstrated athigher concentrations of IW, 4-MG and LY339434, wasevident 1–2 h after stimulation where the cells appearedlarge and swollen, and stained positively with PI. After

431S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
24 h of agonist exposure only debris and a fraction of thecells remained, characteristic of necrosis (Clarke, 1990),mediated, in large, by an influx of Na+ and Cl� ionsfollowed by a secondary passive influx of water (Choi etal., 1987). Glu receptor-mediated toxicity has previouslybeen shown to involve a complex continuum ofapoptosis–necrosis with a considerable NMDA receptor-mediated component of necrosis (Ankarcrona et al.,1995; Nicotera et al., 1997; Cheung et al., 1998b),whereas cellular damage that occurs from the oversti-mulation of the non-NMDA receptors generally occursby apoptosis (Jensen et al., 1998; Giardina et al., 1998;John et al., 1999), except when extremely severe insultsare used (van Lookeren Campagne et al., 1995). Thispattern of injury is consistent with the current findingsfor IW and KA, which produced extensive apoptotic celldeath at lower concentrations, but at higher concen-trations the agonists caused necrosis.
The current study highlights the importance of exam-ining pharmacological profiles of various novel com-pounds in native receptors, and the vast difference thatcan exist between artificially combined receptors andthose that are formed in the presence of other receptorsubunits. Recombinant receptors do not necessarilyallow the construction of heteromeric receptors from thevarious native receptor subunits and cells used in recom-binant technology do not always contain the enzymesresponsible for RNA editing, leading to a functional dif-ference in recombinant and native receptors (Puchalskiet al., 1994; Belcher and Howe, 1997). Therefore, whilerecombinant techniques are invaluable for the initialcharacterization of compounds it is essential to examinethe pharmacological characteristics in native receptors,where primary cell culture is invaluable due to the lackof influences from other neuronal inputs and cell typessuch as glia. Based on our evidence ATPA and low con-centrations of IW appear to be useful drugs for the analy-sis of low-affinity KA receptors. Using this strategy wehave identified new roles for low-affinity KA receptors,which appear additional to their involvement in synaptictransmission (Clarke et al., 1997), and whereby GluR5-7 via apoptosis and programmed cell death are likely tocontribute to neurodegeneration attributed to excitotox-icity (Ikonomidou and Turski, 1995; Leist and Nicot-era, 1998).
Acknowledgements
Supported by the National Health and MedicalResearch Council (Australia), of which P.M. Beart is aSenior Principal Research Fellow, and by grants fromthe Ramaciotti, Rebecca Cooper and William BucklandFoundations, and Perpetual Trustees.
References
Ankarcrona, M., Dypbukt, J.M., Bonfoco, E., Zhivotovsky, B., Orren-ius, S., Lipton, S.A., Nicotera, P., 1995. Glutamate-induced neu-ronal death: a succession of necrosis or apoptosis depending onmitochondrial function. Neuron 15, 961–973.
Aronica, E., Dickson, D.W., Kress, Y., Morrison, J.H., Zukin, R.S.,1998. Non-plaque dystrophic dendrites in Alzheimer hippocampus:a new pathological structure revealed by glutamate receptor immu-nocytochemistry. Neuroscience 82, 979–991.
Belcher, S.M., Howe, J.R., 1997. Characterization of RNA editing ofthe glutamate-receptor subunits GluR5 and GluR6 in granule cellsduring cerebellar development. Molecular Brain Research 52,130–138.
Bernard, A., Ferhat, L., Dessi, F., Charton, G., Represa, A., Ben-Ari,Y., Khrestchatisky, M., 1999. Q/R editing of the rat GluR5 andGluR6 kainate receptors in vivo and in vitro: evidence for inde-pendent developmental, pathological and cellular regulation. Euro-pean Journal of Neuroscience 11, 604–616.
Bleakman, D., Lodge, D., 1998. Neuropharmacology of AMPA andkainate receptors. Neuropharmacology 37, 1187–1204.
Bortolotto, Z.A., Clarke, V.R., Delany, C.M., Parry, M.C., Smolders,I., Vignes, M., Ho, K.H., Miu, P., Brinton, B.T., Fantaske, R.,Ogden, A., Gates, M., Ornstein, P.L., Lodge, D., Bleakman, D.,Collingridge, G.L., 1999. Kainate receptors are involved in synapticplasticity. Nature 402, 297–301.
Breese, G.R., Freedman, R., Leonard, S.S., 1995. Glutamate receptorsubtype expression in human postmortem brain tissue from schizo-phrenics and alcohol abusers. Brain Research 674, 82–90.
Carroll, F.Y., Cheung, N.S., Beart, P.M., 1997. Investigations of non-NMDA receptor-induced toxicity in serum-free antioxidant-richprimary cultures of murine cerebellar granule cells. NeurochemistryInternational 33, 23–28.
Carroll, F.Y., Finkelstein, D.I., Horne, M.K., Lawrence, A.J., Craw-ford, D., Paxinos, G., Beart, P.M., 1998. Regional distribution oflow affinity kainate receptors in brain of Macaca fascicularisdetermined by autoradiography using [3H](2S,4R)-4-methylgluta-mate. Neuroscience Letters 255, 71–74.
Cheung, N.S., Carroll, F.Y., Larm, J.A., Beart, P.M., Giardina, S.F.,1998a. Kainate-induced apoptosis correlates with c-Jun activationin cultured cerebellar granule cells. Journal of NeuroscienceResearch 52, 69–82.
Cheung, N.S., Pascoe, C.J., Giardina, S.F., John, C.A., Beart, P.M.,1998b. Micromolar l-glutamate induces extensive apoptosis in anapoptotic–necrotic continuum of insult-dependent, excitotoxicinjury in cultured cortical neurones. Neuropharmacology 37,1419–1429.
Choi, D.W., Maulucci-Gedde, M., Kriegstein, A.R., 1987. Glutamateneurotoxicity in cortical cell culture. Journal of Neuroscience 7,357–368.
Clarke, G.H., 1990. Developmental cell death: morphological diversityand multiple mechanisms. Anatomy and Embryology 181, 195–213.
Clarke, V.R., Ballyk, B.A., Hoo, K.H., Mandelzys, A., Pellizzari, A.,Bath, C.P., Thomas, J., Sharpe, E.F., Davies, C.H., Ornstein, P.L.,Schoepp, D.D., Kamboj, R.K., Collingridge, G.L., Lodge, D.,Bleakman, D., 1997. A hippocampal GluR5 kainate receptor reg-ulating inhibitory synaptic transmission. Nature 389, 599–603.
Dessi, F., Ben-Ari, Y., Charriaut-Marlangue, C., 1994. Increased syn-thesis of specific proteins during glutamate-induced neuronal deathin cerebellar culture. Brain Research 654, 27–33.
Donevan, S.D., Rogawski, M.A., 1993. GYKI 52466, a 2,3-benzodia-zepine, is a highly selective, noncompetitive antagonist ofAMPA/kainate receptor responses. Neuron 10, 51–59.
Fonnum, F., 1984. Glutamate: a neurotransmitter in the mammalianbrain. Journal of Neurochemistry 42, 1–11.
Giardina, S.F., Carroll, F.Y., Moldrich, R.X., Finklestein, D.I., Horne,

432 S.F. Giardina, P.M. Beart / Neuropharmacology 41 (2001) 421–432
M.K., Lawrence, A.J., Cheung, N.S., Beart, P.M., 2000. Kainatereceptors, cellular signalling and excitotoxic neuronal death: newpharmacological evidence for the functional involvement of low-affinity and metabotropic kainate receptors. In: Cavalheiro, E.A.,Schoepp, D., Turski, L. (Eds.), Excitatory Amino Acids: Ten YearsLater. IOS Press, Amsterdam, pp. 159–170.
Giardina, S.F., Cheung, N.S., Reid, M.T., Beart, P.M., 1998. Kainate-induced apoptosis in cultured murine cerebellar granule cells elev-ates expression of the cell cycle gene cyclin D1. Journal of Neuro-chemistry 71, 1325–1328.
Hack, N.J., Sluiter, A.A., Balazs, R., 1995. AMPA receptors in cer-ebellar granule cells during development in culture. DevelopmentalBrain Research 87, 55–61.
Hollmann, M., Heinemann, S., 1994. Cloned glutamate receptors.Annual Review of Neuroscience 17, 31–108.
Ikonomidou, C., Turski, L., 1995. Excitotoxicity and neurodegener-ative diseases. Current Opinion in Neurology 8, 487–497.
Jane, D.E., Hoo, K., Kamboj, R., Deverill, M., Bleakman, D., Mandel-zys, A., 1997. Synthesis of willardiine and 6-azawillardiine anal-ogs: pharmacological characterization on cloned homomeric humanAMPA and kainate receptor subtypes. Journal of Medical Chemis-try 40, 3645–3650.
Jensen, J.B., Schousboe, A., Pickering, D.S., 1998. AMPA receptormediated excitotoxicity in neocortical neurons is developmentallyregulated and dependent upon receptor desensitization. Neuroch-emistry International 32, 505–513.
John, C.A., Beart, P.M., Giardina, S.F., Pascoe, C.J., Cheung, N.S.,1999. Cyclothiazide and GYKI 52466 modulate AMPA receptor-mediated apoptosis in cortical neuronal cultures. Neuroscience Let-ters 268, 9–12.
Kroemer, G., Dallaporta, B., Resche-Rigon, M., 1998. The mitochon-drial death/life regulator in apoptosis and necrosis. Annual Reviewof Physiology 60, 619–642.
Leist, M., Nicotera, P., 1998. Apoptosis, excitotoxicity, and neuropath-ology. Experimental Cell Research 239, 183–201.
Lipton, S.A., Nicotera, P., 1998. Calcium, free radicals and excitotox-ins in neuronal apoptosis. Cell Calcium 23, 165–171.
Lipton, S.A., Rosenberg, P.A., 1994. Excitatory amino acids as a finalcommon pathway for neurologic disorders. New England Journalof Medecine 330, 613–622.
Moldrich, R.X., Beart, P.M., Pascoe, C.J., Cheung, N.S., 2000a. Low-affinity kainate receptor agonists induce insult-dependent apoptosisand necrosis in cultured murine cortical neurons. Journal of Neuro-science Research 59, 788–796.
Moldrich, R.X., Cheung, N.S., Pascoe, C.J., Baker, S.R., Pedregal, C.,Beart, P.M., 2000b. Excitotoxic profile of LY339434, a GluR5agonist, in cultured murine cortical neurons. Brain Research 862,270–275.
Nicotera, P., Ankarcrona, M., Bonfoco, E., Orrenius, S., Lipton, S.A.,1997. Neuronal necrosis and apoptosis: two distinct events inducedby exposure to glutamate or oxidative stress. Advances in Neurol-ogy 72, 95–101.
Paschen, W., Schmitt, J., Uto, A., 1996. RNA editing of glutamatereceptor subunits GluR2, GluR5 and GluR6 in transient cerebralischemia in the rat. Journal of Cerebral and Blood Flow Metab-olism 16, 548–556.
Patneau, D.K., Mayer, M.L., Jane, D.E., Watkins, J.C., 1992. Acti-vation and desensitization of AMPA/kainate receptors by novelderivatives of willardiine. Journal of Neuroscience 12, 595–606.
Pemberton, K.E., Belcher, S.M., Ripellino, J.A., Howe, J.R., 1998.High-affinity kainate-type ion channels in rat cerebellar granulecells. Journal of Physiology 510, 401–420.
Philpott, K.L., McCarthy, M.J., Becker, D., Gatchalian, C., Rubin,L.L., 1996. Morphological and biochemical changes in neurons:apoptosis versus mitosis. European Journal of Neuroscience 8,1906–1915.
Porter, R.H., Eastwood, S.L., Harrison, P.J., 1997. Distribution of kain-ate receptor subunit mRNAs in human hippocampus, neocortex andcerebellum, and bilateral reduction of hippocampal GluR6 and KA2transcripts in schizophrenia. Brain Research 751, 217–231.
Procter, M.J., Houghton, A.K., Faber, E.S., Chizh, B.A., Ornstein,P.L., Lodge, D., Headley, P.M., 1998. Actions of kainate andAMPA selective glutamate receptor ligands on nociceptive pro-cessing in the spinal cord. Neuropharmacology 37, 1287–1297.
Puchalski, R.B., Louis, J.C., Brose, N., Traynelis, S.F., Egebjerg, J.,Kukekov, V., Wenthold, R.J., Rogers, S.W., Lin, F., Moran, T.,Morrison, J.H., Heinemann, S., 1994. Selective RNA editing andsubunit assembly of native glutamate receptors. Neuron 13, 131–147.
Resink, A., Hack, N., Boer, G.J., Balazs, R., 1994. Growth conditionsdifferentially modulate the vulnerability of developing cerebellargranule cells to excitatory amino acids. Brain Research 655,222–232.
Sastry, P.S., Rao, K.S., 2000. Apoptosis and the nervous system. Jour-nal of Neurochemistry 74, 1–20.
Small, B., Thomas, J., Kemp, M., Hoo, K., Ballyk, B., Deverill, M.,Ogden, A.M., Rubio, A., Pedregal, C., Bleakman, D., 1998.LY339434, a GluR5 kainate receptor agonist. Neuropharmacology37, 1261–1267.
Stensbol, T.B., Slok, F.A., Trometer, J., Hurt, S., Ebert, B., Kjoller,C., Egebjerg, J., Madsen, U., Diemer, N.H., Krogsgaard-Larsen, P.,1999. Characterization of a new AMPA receptor radioligand,[3H]2-amino-3-(3-carboxy-5-methyl-4-isoxazolyl)propionic acid.European Journal of Pharmacology 373, 251–262.
Toescu, E.C., 1998. Apoptosis and cell death in neuronal cells: wheredoes Ca2+ fit in? Cell Calcium 24, 387–403.
van Lookeren Campagne, M., Lucassen, P.J., Vermeulen, J.P., Balazs,R., 1995. NMDA and kainate induce internucleosomal DNA cleav-age associated with both apoptotic and necrotic cell death in theneonatal rat brain. European Journal of Neuroscience 7, 1627–1640.
Vandenberg, R.J., Mitrovic, A.D., Chebib, M., Balcar, V.J., Johnston,G.A., 1997. Contrasting modes of action of methylglutamatederivatives on the excitatory amino acid transporters, EAAT1 andEAAT2. Molecular Pharmacology 51, 809–815.
Vignes, M., Collingridge, G.L., 1997. The synaptic activation of kain-ate receptors. Nature 388, 179–182.
Wahl, P., Frandsen, A.A., Madsen, U., Schousboe, A., Krogsgaard-Larsen, P., 1998. Pharmacology and toxicology of ATOA, anAMPA receptor antagonist and a partial agonist at GluR5 receptors.Neuropharmacology 37, 1205–1210.
Wisden, W., Seeburg, P.H., 1993. A complex mosaic of high-affinitykainate receptors in rat brain. Journal of Neuroscience 13, 3582–3598.
Zhou, L.M., Gu, Z.Q., Costa, A.M., Yamada, K.A., Mansson, P.E.,Giordano, T., Skolnick, P., Jones, K.A., 1997. (2S,4R)-4-methylglu-tamic acid (SYM 2081): a selective, high-affinity ligand for kainatereceptors. Journal of Pharmacology and Experimental Therapeutics280, 422–427.