estradiol alleviates acinar cell apoptosis and chronic pancreatitis in male wistar bonn/kobori rats
TRANSCRIPT

Estradiol Alleviates Acinar Cell Apoptosis and ChronicPancreatitis in Male Wistar Bonn/Kobori Rats
*Soichi Nakamura, *Tamaki Yamada, ‡Takashi Hashimoto, †Satoru Takahashi, §Mitsue Sogawa,*Hirotaka Ohara, *Takahiro Nakazawa, *Hitoshi Sano, *Atsushi Kuno, *Takashi Joh,
*Tomoyuki Nomura, §Tetsuo Arakawa, *Makoto Itoh
*Departments of Comprehensive Medicine, Internal Medicine, and Bioregulation and †Departments of Pathology andPathophysiology, Experimental Pathophysiology, and Tumor Biology, Nagoya City University Graduate School of Medical Sciences,
Nagoya; ‡Second Department of Internal Medicine, Aichi Medical University, Nagakute; and §Third Department of Gastroenterology,Graduate School of Medicine, Osaka City University Medical School, Osaka, Japan
Aims: To quantitatively determine the influence of estradiol onacinar cell apoptosis and chronic pancreatitis; assess its effects oninfiltration of CD4 and CD8 T cells in the pancreas; investigate therole of testosterone on chronic pancreatitis in 20-week-old maleWBN/Kob rats; and determine the impact of estradiol on prolif-eration of splenocytes derived from these animals in vitro. Meth-odology and Findings: Treatment with high (0.4 mg·kg·week)but not low (0.1 mg·kg·week) doses of estradiol for 10 weekssignificantly decreased the number of apoptotic acinar cells stainedwith an anti–single strand DNA antibody, histologic scores, andpancreatic myeloperoxidase activity in 20-week-old WBN/Kobrats, in comparison with control values. The high doses also sig-nificantly attenuated the increase in pancreatic hydroxyprolinecontent, an indicator of collagen deposition, at 20 weeks. They
caused significant decreases in the numbers of CD4 and CD8 Tcells infiltrating the pancreas. Both doses suppressed levels oftestosterone but without any influence on the serum corticosteroneconcentrations. Androgen receptors could not be immunohisto-chemically identified in the pancreas at 20 weeks, and dietarytreatment with flutamide, an androgen receptor antagonist, did notinfluence the chronic pancreatitis. Estradiol significantly reduced1% phytohemagglutinin-induced incorporation of bromodeoxyuri-dine into the splenocytes in vitro. Conclusions: We conclude thatestradiol dose-dependently attenuates acinar cell apoptosis and de-velopment of chronic pancreatitis, independent of any change inendogenous corticosterone and testosterone, by suppressing infil-tration and function of T cells. Key Words: Acinar cell—Apoptosis—Corticosterone—Sex hormones.
Male Wistar Bonn/Kobori (WBN/Kob) rats spontane-ously develop chronic pancreatitis (1–3). In the initialphase, edema of the interlobular and intralobular connectivetissue associated with inflammatory cells, predominantlyneutrophils, focally occurs in the pancreas. This alteration isfollowed by replacement with granulation tissue, featuringsevere infiltration of immature fibroblasts, neutrophils, andlymphocytes. Finally, chronic inflammation results in insuf-ficiencies of exocrine and endocrine functions (1–3). Hashi-moto et al., from our laboratory, earlier reported that apo-ptosis of acinar cells, closely related to decreased endog-enous corticosterone, is involved in chronic pancreatitis inmale WBN/Kob rats (4).
We have also demonstrated that predominantly CD8-positive and to a lesser extent CD4-positive T cells infiltrate
into the pancreatic parenchyma; the CD8 T cells closelyadhere to the apoptotic acinar cells, and the treatment withtacrolimus, an immunosuppressant, alleviates apoptosis ofacinar cells and chronic pancreatitis at 20 weeks of age inthis model (5). We concluded that CD8- and CD4-positiveT cells are responsible for the induction of acinar cell ap-optosis and the development and persistence of pancreati-tis (5).
Hormones are known to modulate the immune system(6), and castration or treatment with different sex steroidsresults in attenuation or exacerbation of autoimmune dis-eases in experimental animals (7–10). In WBN/Kob rats,chronic pancreatitis occurs only in males, and the diseasedevelops around the time of sexual maturity (1–3). Estradioltreatment or castration in males ameliorates the condition,whereas ovariectomy in females induces diabetic symptoms(11). Ohashi et al. reported that treatment with estradiol butnot luteinizing hormone–releasing hormone partially attenu-ates early pancreatic lesions observed at 16 weeks in maleWBN/Kob rats (12). Although they described protectiveeffects of estradiol against early pancreatic injury in thismodel, the dose was constant over time for each animal,with the result that the pharmacological effects decreased as
Manuscript received August 19, 2002; revised manuscript accepted No-vember 6, 2002.
Address correspondence and reprint requests to Dr. Tamaki Yamada,Department of Comprehensive Medicine, Internal Medicine, and Bioregu-lation, Nagoya City University Graduate School of Medical Sciences, 1Kawasumi, Mizuho-cho, Mizuho-ku Nagoya 467–8602, Japan. E-mail:[email protected]
PancreasVol. 26, No. 3, pp. e59–e66© 2003 Lippincott Williams & Wilkins, Inc., Philadelphia
e59

the body weight increased during the treatment period. Fur-thermore, their findings were presented in Japanese in anonquantitative fashion and were not accompanied by anyattempt at elucidation of mechanisms.
Therefore, we decided to conduct a quantitative assess-ment of the effects of estradiol doses based on body weightsof WBN/Kob rats and, moreover, to concentrate on under-lying processes. To our knowledge, there is no informationavailable about the effects of estradiol on apoptosis of aci-nar cells and infiltration of CD4 and CD8 T cells, and ourunderstanding of the mechanisms of the hormone protectionin WBN/Kob rats is incomplete.
Therefore, the objectives set for this study were to: quan-titatively determine the impact of estradiol on acinar cellapoptosis and chronic pancreatitis; assess the effects on in-filtration of CD4 and CD8 T cells into the pancreas; inves-tigate pancreatic androgen receptors immunohistochemical-ly and the influence of flutamide, an androgen receptorantagonist, on chronic pancreatitis in male WBN/Kob rats;and determine what effects estradiol has on proliferation ofsplenocytes derived from male WBN/Kob rats in vitro.
MATERIALS AND METHODS
Materials and animalsTen-week-old male Wistar Bonn/Kobori (WBN/Kob)
and 20-week-old male Wistar rats were obtained from SLC(Hamamatsu, JAPAN) and maintained in a temperature-controlled room under a constant light cycle, with free ac-cess to water and standard laboratory chow diet (containingprotein, 24.8%; fat, 4.4%; fiber, 3.9%; and carbon, 6.9%).The Animal Care Committee of Nagoya City Universityapproved the study protocol. Beta-estradiol was purchasedfrom Wako Co., Ltd. (Tokyo, Japan), and flutamide wasgenerously donated by Nippon Kayaku Co., Ltd. (Tokyo,Japan). All other chemicals were of the highest qualityavailable.
Experimental protocolsThe 10-week-old WBN/Kob rats were randomly divided
into three groups, receiving the vehicle (mineral oil; n � 5)and low and high doses of estradiol (0.1 and 0.4mg·kg·week; n � 6 and n � 5, respectively). In a separateexperiment, 10-week-old WBN/Kob rats were randomizedinto vehicle (n � 5) and high-dose estradiol (n � 7) groupsfor measurement of pancreatic hydroxyproline content andevaluation of immunohistochemical staining for CD4- andCD8-positive T cells in the pancreas.
Ten-week-old WBN/Kob rats were also randomly di-vided into three groups, as follows: no treatment (n � 4)and low- and high-dose estradiol (0.1 and 0.4 mg·kg·week,n � 3 for both). Serum estradiol levels were measuredwithout estradiol treatment in the no-treatment group andwithin 72 hours after estradiol treatment in the low-dose andhigh-dose groups, respectively. The 20-week-old maleWistar rats (n � 5) were used for comparison of hydroxy-proline content in the pancreas with that in WBN/Kob rats.
Ten-week-old WBN/Kob rats were also assigned to twogroups, given a normal powder diet (n � 5) and a powderdiet containing 0.1% flutamide (n � 8).
Administration of estradiolTen-week-old WBN/Kob rats were injected subcutane-
ously with either vehicle alone or with estradiol in the ve-hicle (0.05 and 0.2 mg/kg) twice a week for 10 weeks (intotal, 0.1 and 0.4 mg·kg·week, respectively). Weights of theanimals were recorded once a week.
Administration of flutamideTen-week-old WBN/Kob rats were fed either a normal
powder diet or a powder diet containing with 0.1% flu-tamide for 10 weeks. The amount of flutamide was deter-mined as reported previously (13). Approximately, the cal-culated dose from 0.1% powder–containing diet was 80mg·kg·day. Weights of the animals and food intake wererecorded once a week.
Tissue and blood samplingAt week 20, following the estradiol or flutamide treat-
ment, rats were weighed and anesthetized by intraperitonealinjection of pentobarbital sodium (50 mg per 100 g bodyweight; Abbott Laboratories, North Chicago, IL, USA). Af-ter blood was taken from the abdominal aorta, they werekilled with an overdose of the anesthetic. Then each pan-creas was removed and weighed.
Portions were stored at −40°C for subsequent measure-ment of pancreatic myeloperoxidase activity or fixed in10% formalin for morphologic assessment and stained withhematoxylin and eosin as well as immunohistochemicallyfor androgen receptors. Serum was also stored at −40°Cbefore measurement of corticosterone, testosterone, and flu-tamide (flutamide-treated group) levels.
For assessment of serum estradiol levels, blood was takenfrom the femoral vein at 3, 6, 24, and 72 hours after estra-diol administration in the low- and high-dose groups underpentobarbital sodium anesthesia. Blood was also taken inthe no-treatment group for determination of the serum levelsof estradiol (physiological levels).
Morphological examinationA pathologist who was blinded to the treatments carried
out the microscopic examination. Histological alterations ofthe pancreas, including inflammatory cell infiltration, inter-stitial edema, fibrosis, acinar cell necrosis, and hemorrhage,were graded on a scale of minimal to maximal (0–3), asdescribed previously (4).
Immunohistochemical staining forsingle-stranded DNA
Pancreas tissues from rats in the vehicle and low- andhigh-dose estradiol groups were fixed with formalin, cutinto 4–6-�m paraffin sections, and deparaffinized. Single-stranded DNA was immunohistochemically identified withuse of a commercially available kit (DAKO Japan, Kyoto,Japan) (14). Average counts of positive nuclei were deter-mined in 20 randomly selected areas at 200× magnification,as described previously (4).
Immunohistochemical staining for CD4- andCD8-positive T cells
For immunohistochemistry, pancreas tissue was fixedwith 4% paraformaldehyde in phosphate-buffered saline
e60 Pancreas, Vol. 26, No. 3, 2003

overnight at 4°C. After successive transfer through 8% to20% sucrose in 0.1 M phosphate buffer (pH, 7.4), portionswere frozen in liquid nitrogen and sections were cut in aCM3000 cryostat (Leika Instruments BmbH, Nussloch,Germany) and immediately air-dried. To block endogenousperoxidase activity, sections were incubated with 0.3%hydrogen peroxide in methanol for 20 minutes at roomtemperature and further incubated with diluted mouseanti-rat CD4 monoclonal antibody (mAb, 1:300; RM4–5;PharMingen, San Diego, CA, USA) and mouse anti-rat CD8mAb (1:300, YTS169.4; Cedalane, Ontario, Canada) over-night at 4°C.
After washing with phosphate-buffered saline, sectionswere treated with diluted biotinylated secondary rabbit anti-mouse IgG antibody (1:400; Dako, Copenhagen, Denmark)for 60 minutes at room temperature, followed by incubationwith avidin-biotin-peroxidase complex (Vectastain kit;Vector Laboratories, Peterborough, UK). Reaction productswere visualized by incubation with 0.025% diaminobenzi-dine and 0.003% hydrogen peroxide. Counterstaining wasachieved with methyl green. Average counts of CD4- andCD8-positive T cells were determined in 20 randomly se-lected areas at 200× magnification, as described previ-ously (4).
Immunohistochemical staining for androgen receptorsPancreas specimens were routinely fixed in 10% phos-
phate-buffered formalin for 48 hours and then processed forembedding in paraffin. Five-�m-thick sections were cut andimmunohistochemical analyses were performed with use ofrabbit polyclonal anti-rat androgen receptor antibody (Af-finity Bioreagents, Golden, CO, USA). Binding was visu-alized with a Vectastain Elite ABC kit (Vector Lab, Bur-lingame, CA, USA), and light hematoxylin counter-stainingwas conducted to facilitate microscopic examination.Wistar rat prostate specimen was used for positive staining,as reported previously (15).
Measurement of pancreatic myeloperoxidase(MPO) activity
Pancreatic MPO activity, an indirect quantitative index ofgranulocyte infiltration, was determined by means of a mi-nor modification of the method of Grisham et al. (16). Inbrief, pancreatic tissue was homogenized in 20 mM phos-phate buffer (pH, 7.4) and centrifuged at 6,000g for 20minutes at 4°C. The pellet was then homogenized and soni-cated with an equivalent volume of 50 mM acetic acid (pH,6.0) containing 0.5% (w/v) hexadecyltrimethylammoniumhydroxide, and MPO activity was determined by measuringthe H2O2-dependent oxidation of 3,3�5,5� tetramethylbenzi-dine. Results were expressed as units per gram of pancreas.
Measurement of pancreatic hydroxyproline contentHydroxyproline content, an indicator of collagen deposi-
tion, was determined by a modification of the method ofBlumenkrantz and Asobe-Hansen (17). In brief, approxi-mately 100 mg of pancreas tissue was homogenized in wa-ter and hydrolyzed in 10 N HCl at 110°C for 20 hours. HClwas evaporated under a nitrogen gas flush and the hydro-
lysate, dissolved in distilled water, was filtered. Then 0.5-mL aliquots were mixed with 3 mL of citrate-phosphatebuffer (0.15 M citric acid and 0.6 M dibasic sodium phos-phate) and 0.5 mL of 1 M periodic acid in 9 M H3PO4,added to 1.75 mL of extract buffer consisting of toluene (5parts), 2-methyl-1-propanol (5 parts), and 1-propanol (2parts), shaken for 30 minutes and centrifuged. The organicphase (0.6 mL) was mixed with Ehrlich’s reagent (0.15 mL)and left for 15 minutes.
Absorbance was read at 565 nm and hydroxyproline lev-els were calculated with reference to a standard curve for4-hydroxy-1-proline and expressed as micrograms per gramof tissue.
Measurements of serum levels of corticosterone,testosterone, and estradiol
Serum corticosterone levels in the vehicle and low- andhigh-dose estradiol groups were determined as reported pre-viously (18).
Serum testosterone and estradiol levels in the vehicle andlow- and high-dose estradiol groups were determined withuse of DPC total testosterone kits or DPC total estradiol kits,respectively (purchased from Diagnostic Products Co., LosAngeles, CA, USA).
Measurement of serum flutamideSerum flutamide levels were determined in the flutamide
group by high-speed liquid chromatographic analysis, asreported previously (19).
Proliferation activity of splenocytesProliferative activity of splenocytes was determined by
measuring incorporation of bromodeoxyuridine (BrdU) intothe cells after stimulation with phytohemagglutinin (PHA),which predominantly activates T cells, with use of a BrdUproliferation assay kit (Oncogene Research Products, Cam-bridge, MA, USA). Splenocytes were prepared from 20-week-old male WBN/Kob rats as reported previously (20)and stimulated with 1% PHA in the absence or presence ofestradiol at 37°C for 24 hours. Then cells were incubatedwith BrdU at 37°C for another 24 hours.
Amounts of incorporated label in each group were mea-sured according to the manufacturer’s protocol. The con-centrations of PHA and estradiol were in line with previousreports (21,22). The stimulation index was defined as theabsorbance ratio for each group against the negative controlvalue with cells incubated in the medium.
Splenocytes were divided into four groups as follows:negative control (cultured without PHA); positive control(cultured with 1% PHA); Es5 (cultured with 5 �M estradioland 1% PHA); and Es10 (cultured with 10 �M estradiol and1% PHA).
Statistical analysisEach data set is presented as the arithmetic mean ± stan-
dard error of the mean (SEM). Statistical differences amonggroups were identified by means of one-way analysis ofvariance, and multiple comparisons were performed by theleast-significant-difference method. Differences between
S. NAKAMURA ET AL. ESTRADIOL IN APOPTOSIS AND PANCREATITIS e61

two groups were analyzed by Student t test. Statistical sig-nificance was set at the 5% level.
RESULTS
Body and pancreatic weightsTreatment with estradiol for 10 weeks dose-dependently
and significantly decreased body weights and increasedpancreatic weights of WBN/Kob rats at the age of 20 weeksin comparison with the vehicle treatment (Table 1).
Morphological findings and inflammatory parametersHigh doses of estradiol completely prevented the macro-
scopic lesions of the pancreas, including atrophy, hemor-rhage, and pigmentation, which were generally observed inthe vehicle group, whereas low doses partially attenuatedtheir development. Estradiol also dose-dependently amelio-rated microscopic findings at the age of 20 weeks, reducinginflammatory cell infiltration, edema, fibrosis, acinar cellnecrosis, and hemorrhage (Fig. 1 A, B, C). These protectiveeffects were quantitatively confirmed by the significant in-crease in pancreatic weight and decreases in pancreaticMPO activity and histologic scores in the high-dose group,in comparison with measures in the vehicle and low-dosegroups (Table 1).
Apoptosis of acinar cellsHigh doses of estradiol significantly decreased the num-
bers of apoptotic acinar cells, assessed by counting nucleistained with the anti-single-stranded DNA antibody in 20-week-old WBN/Kob rats, in comparison with the vehicleand low-dose groups (Fig. 2 A and B and Table 1).
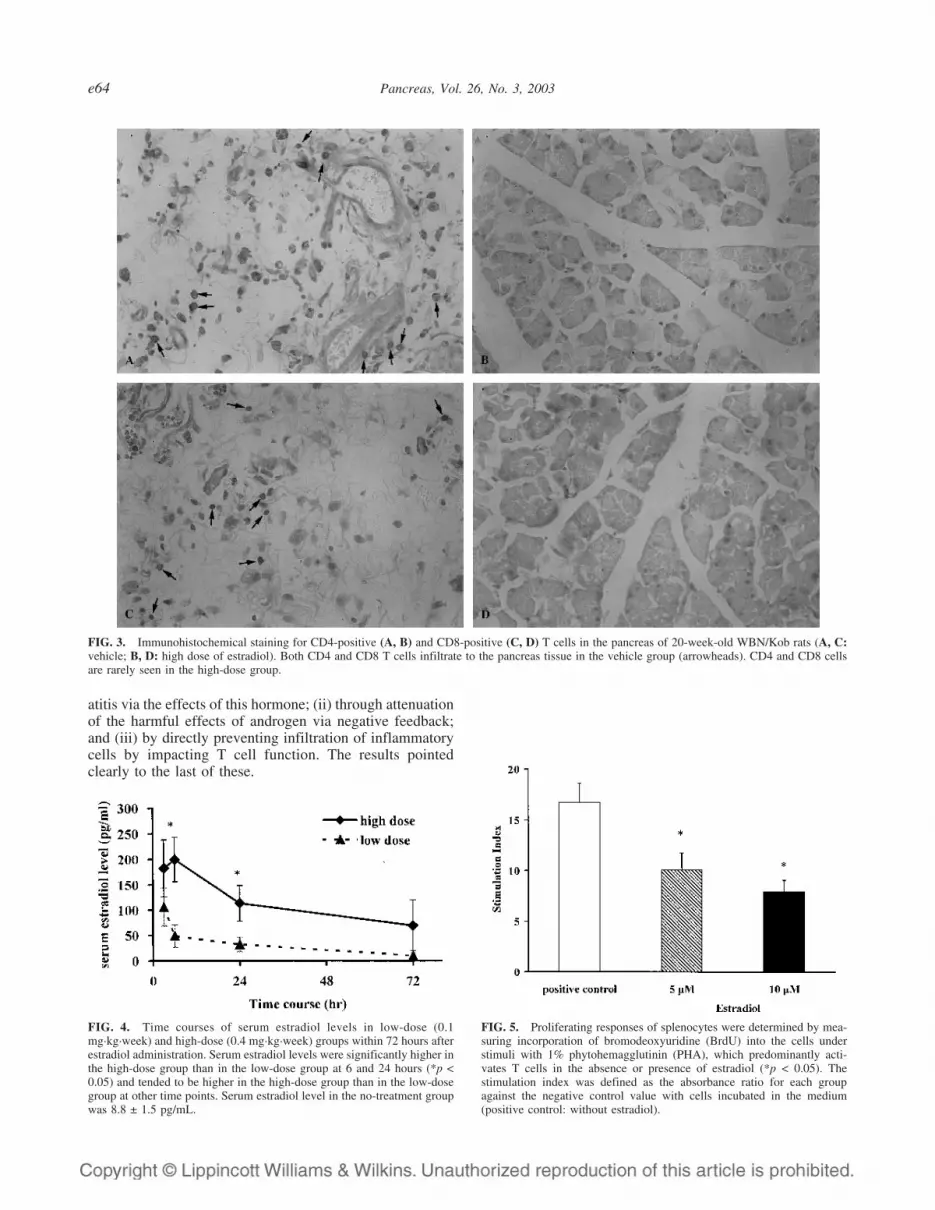
Infiltrating CD4- and CD8-positive T cells inthe pancreas
Immunohistochemical staining of T cells revealed bothCD4- and CD8-positive populations infiltrating the pan-
creas in the vehicle group (Fig. 3A, C). High doses of es-tradiol significantly decreased the numbers of both CD4-and CD8-positive T cells in the pancreas in 20-week-oldWBN/Kob rats, in comparison with the vehicle group (Fig.3B, D) (CD4: 0.4 ± 0.1 versus 3.4 ± 1.1 per field; CD8: 0.5± 0.1 versus 4.5 ± 1.0 per field; p < 0.05 for both).
FIG. 1. Pancreas tissue from 20-week-old WBN/Kob rats treated withvehicle (A), low-dose (0.1 mg·kg·week) estradiol (B), and high-dose (0.4mg·kg·week) estradiol (C) (hematoxylin and eosin staining; original mag-nification, ×25). A: Degeneration of acinar cells, formation of pseudotu-bules, and infiltration of inflammatory cells are evident in the inflamedlobules. Acinar cells are disappearing in the area with the most severeinflammation. B: Although lobular structure is partially maintained, thesame degree of inflammatory changes as the vehicle group is observed. C:Parenchyma of pancreas is almost intact. Areas of inflammatory cellsinfiltration or structural degeneration are rarely seen.
TABLE 1. The effects of estradiol treatment on body weight(BW), pancreas weight, pancreatic myeloperoxidase (MPO)
activity, histologic score, and number of apoptotic acinar cellsin 20-week-old male WBN/Kob rats
Vehicle(mineral oil)
(n � 5)
Estradiol
0.1 mg/kgBW/week(n � 6)
0.4 mg/kgBW/week(n � 5)
Body weight (g) 396.4 ± 11.5 301.5 ± 6.7a 274.0 ± 5.0a
Pancreas weight (g/kg) 2.4 ± 0.2 3.2 ± 0.1a 3.7 ± 0.2a
Pancreatic MPO activity(unit/g wet weight) 6.7 ± 0.7 3.9 ± 0.7a 3.3 ± 0.6a
Histologic scoreb 1.64 ± 0.46 0.63 ± 0.44 0.00 ± 0.00a
No. of apoptotic acinarcellsc (no./field) 2.00 ± 1.04 0.50 ± 0.36 0.02 ± 0.01a
a p < 0.05 compared with mineral oil.b Histologic alternations of the pancreas, including inflammatory cell
infiltration, interstitial edema, fibrosis, acinar cell necrosis, and hemor-rhage, were graded on a scale of minimal to maximal (0–3), and the valueswere averaged in each rat.
c Averaged counts of positive nuclei were determined in 20 randomlyselected areas at 200-fold magnification.
e62 Pancreas, Vol. 26, No. 3, 2003

Pancreatic hydroxyproline contentHydoxyproline content, an indicator of collagen deposi-
tion, was significantly increased in the pancreas inWBN/Kob versus Wistar rats at 20 weeks of age (589.0 ±122.6 versus 72.5 ± 7.6 �g/g, p < 0.05). High doses ofestradiol significantly suppressed this increase in hydroxy-proline content in the pancreas to 139.2 ± 27.6 �g/g (p <0.05).
Serum levels of corticosterone, testosterone,and estradiol
Serum corticosterone levels did not differ among the ve-hicle, low-dose, and high-dose groups (291.5 ± 15.3 versus168.4 ± 50.6 and 233.2 ± 5.7 ng/mL, respectively).
Serum testosterone levels were significantly lower in thelow-dose and high-dose groups than in the vehicle group(0.05 ± 0.03 and 0.00 ± 0.00 versus 0.95 ± 0.15 ng/mL,respectively; p < 0.05 for both). Although two rats in thelow-dose group had undetectable levels of endogenous tes-tosterone, no protection was observed in their pancreases.
Serum estradiol level was 8.8 ± 1.5 pg/mL in the no-treatment group. Serum estradiol levels were significantlyhigher in the high-dose group than in the low-dose group at
6 and 24 hours and tended to be elevated at other time pointsalso (Fig. 4). Serum estradiol levels in the high-dose groupat all points were much higher than in the no-treatmentgroup.
Immunohistochemical staining of androgen receptorsThere was no apparent immunohistochemical staining of
androgen receptors in the pancreas in 20-week-old WBN/Kobrats (figures not shown).
Effects of dietary treatment with 0.1% flutamide onchronic pancreatitis
The dose of flutamide calculated from food intake was49.3 ± 9.3 mg·kg·day. Serum flutamide level was 6.95 ±0.35 �g/mL. Dietary treatment of 10 weeks affected neitherincrease in pancreatic MPO activity nor histologic findingsat 20 weeks (data not shown).
Cell proliferative activity of splenocytesBrdU cell proliferation assays revealed both 5 and 10 �M
of estradiol to significantly reduce the proliferating re-sponses of splenocytes under stimulation with 1% PHA(Fig. 5).
DISCUSSION
We quantitatively demonstrated that subcutaneous treat-ment with high but not low doses of estradiol for 10 weeksprevents acinar cell apoptosis and development of chronicpancreatitis in male WBN/Kob rats. High doses of estradiolreduced the numbers of CD4 and CD8 T cells infiltratingthe pancreas. This protective effect appeared independent ofthe endogenous testosterone and corticosterone since bothdoses of estradiol significantly suppressed endogenous tes-tosterone levels without affecting endogenous corticoste-rone concentrations at 20 weeks.
Furthermore, androgen receptors could not be immuno-histochemically identified in the pancreas, and dietary treat-ment with flutamide, an androgen receptor antagonist, for10 weeks did not affect the level of pancreatic injury at 20weeks. A possible role in suppression of proliferation ofsplenocytes was also indicated by the in vitro results withPHA activation.
Many investigators have previously demonstrated that es-tradiol regulates the immune system and affects the hypo-thalamic-pituitary-adrenal axis (6,23). Although sex hor-mones are reported to play important roles in the pathogen-esis of pancreatic inflammation and diabetes in maleWBN/Kob rats (11,12), the mechanism of estradiol-mediated protection remains unclear. We recently demon-strated that T cells play a crucial role in acinar cell apoptosisand induction of pancreatitis (5).
Therefore, in the current study we focused on the effectsof estradiol on endogenous steroid hormones and T cells,testing the hypothesis that one of the following three theo-ries might be correct regarding the mechanism by whichestradiol ameliorates apoptosis of acinar cell and chronicpancreatitis in male WBN/Kob rats: (i) by increasing en-dogenous corticosterone and indirectly attenuating pancre-
FIG. 2. Immunohistochemical staining for single-stranded DNA in 20-week-old WBN/Kob rat pancreas (A, vehicle; B, high dose of estradiol;original magnification, ×25). A: The apoptotic cells are observed in thesurrounding area of the inflammation (arrowheads), and some of the in-flammatory cells are positively stained. B: Apoptotic cells are very rare inthe pancreatic tissue from the high-dose group.
S. NAKAMURA ET AL. ESTRADIOL IN APOPTOSIS AND PANCREATITIS e63
atitis via the effects of this hormone; (ii) through attenuationof the harmful effects of androgen via negative feedback;and (iii) by directly preventing infiltration of inflammatorycells by impacting T cell function. The results pointedclearly to the last of these.
FIG. 4. Time courses of serum estradiol levels in low-dose (0.1mg·kg·week) and high-dose (0.4 mg·kg·week) groups within 72 hours afterestradiol administration. Serum estradiol levels were significantly higher inthe high-dose group than in the low-dose group at 6 and 24 hours (*p <0.05) and tended to be higher in the high-dose group than in the low-dosegroup at other time points. Serum estradiol level in the no-treatment groupwas 8.8 ± 1.5 pg/mL.
FIG. 5. Proliferating responses of splenocytes were determined by mea-suring incorporation of bromodeoxyuridine (BrdU) into the cells understimuli with 1% phytohemagglutinin (PHA), which predominantly acti-vates T cells in the absence or presence of estradiol (*p < 0.05). Thestimulation index was defined as the absorbance ratio for each groupagainst the negative control value with cells incubated in the medium(positive control: without estradiol).
FIG. 3. Immunohistochemical staining for CD4-positive (A, B) and CD8-positive (C, D) T cells in the pancreas of 20-week-old WBN/Kob rats (A, C:vehicle; B, D: high dose of estradiol). Both CD4 and CD8 T cells infiltrate to the pancreas tissue in the vehicle group (arrowheads). CD4 and CD8 cellsare rarely seen in the high-dose group.
e64 Pancreas, Vol. 26, No. 3, 2003

Kimura et al. earlier reported that depletion of endog-enous glucocorticoids by adrenalectomy induced acinar cellapoptosis in rats, suggesting they might play an importantrole in maintaining the integrity of acinar cells (24). Werecently demonstrated that prednisolone completely pre-vented apoptosis of acinar cells and chronic pancreatitis inmale WBN/Kob rats (4). Estrogen generally elevates en-dogenous corticosterone levels by enhancing corticoste-rone-binding globulin synthesis in the liver, and femaleshave higher levels than males (25,26).
Indeed, Nakama et al. reported that treatment with estra-diol increased the endogenous levels of corticosterone at theage of 40 weeks and speculated that antiinflammatory ac-tion of corticosterone may be responsible for inhibition ofdiabetes by the hormone in male WBN/Kob rats (11). How-ever, the present data are not in agreement, as we observedno correlation between an increase of endogenous cortico-sterone and inhibition of acinar cell apoptosis and chronicinflammation. We speculate that an increase in endogenouscorticosterone may not be necessary for the preventive ef-fect of estradiol in this treatment.
It is well documented that the immune response is moreactive in females than males (8). In contrast to the findingsin WBN/Kob rats, nonobese diabetes in mice is more preva-lent in females than the males (10). In these latter animals,castration increases the incidence of diabetes in males, andcastration and androgen treatment decreases it in females(9,10). NZB/NZW1 mice, known as an experimental modelfor systemic lupus erythematosus, exhibit more rapid devel-opment of the disorder in females, and testosterone hastherapeutic potential (27).
Therefore, female hormones have been suspected to con-tribute to higher incidences of autoimmune diseases in fe-males. However, in the WBN/Kob rats, sex hormones seemto affect development of disease conversely. Castration inmales ameliorates and testosterone treatment in females in-duces diabetic symptoms, suggesting that androgens maybe important components of the development of diabetes(11).
In the current study, endogenous testosterone levels weredramatically depressed in both the low- and high-dose es-tradiol groups. Furthermore, among the rats with undetect-able and extremely low levels of endogenous testosterone,the pancreases in the low-dose group were injured, suggest-ing that the decrease in male hormone may not have beenresponsible.
Although it is reported that pancreas tissue from rats ex-presses transcripts for the androgen receptor gene and an-drogen receptors have been detected by binding assay in therat and human pancreas (28–30), they were not detected byimmunohistochemistry in the present model. We speculatethat levels may be lower than the detectable limit with thisapproach, as pancreatic transcription of the encoding gene isreported to be weaker than in other reproductive organs inrats (30). To further confirm a lack of major involvement ofandrogen receptors in the pathogenesis of pancreatic injury,we tested the effects of flutamide, an androgen receptorantagonist.
We previously estimated that rats fed 0.5% diosgenin ina powder diet consumed 432 mg·kg·day of diosgenin (13).In the current study, 49.3 mg·kg·day of flutamide, whichachieved an average serum level of 6.95 �g/mL, had nosuppressive effect on pancreatitis. Because the dose of flu-tamide applied, 15–25 mg·kg·day, was found to suppress thedevelopment of prostate cancer in rats (31) and the presentflutamide level is much higher than what is effective inhumans (32), it might have been expected to be effectivein blocking the action of androgen receptors if they werepresent in this model.
The available information thus supports our speculationthat the decrease in testosterone brought about by exog-enous estradiol was not responsible for the observed attenu-ation of chronic pancreatitis.
It is known that physiological doses of estradiol stimulatewhile pharmacological doses suppress T cell functions, in-cluding early T cell development in the thymus as well asproliferative responses (6,22,33). The serum estradiol levelswere much higher in the high-dose group than in the no-treatment group, suggesting that this dose was clearly phar-macological. Recently, we demonstrated that CD8 and CD4T cells infiltrate the pancreas and, in particular, CD8 T cellsclosely adhere to apoptotic acinar cells in male WBN/Kobrats (5). We also found that an immunosuppressant, tacro-limus, completely attenuated acinar cell apoptosis and de-velopment of chronic pancreatitis, a finding suggesting im-portant roles of these T cells in the pathogenesis (5).
Taking into account all of the findings, we can speculatethat estrogen prevents pancreatic injury through suppressionof T cell function in this model. To investigate the possibleinfluence of estradiol in extrathymic T cells, proliferation ofsplenocytes was assessed, and we found the response tostimulation by PHA was significantly reduced. Our resultsthus support the hypothesis that estrogen may prevent acinarcell apoptosis and pancreatic injury through suppression ofT cell activity in this model.
It is reported that estradiol inhibits myofibroblastic trans-formation of hepatic stellate cells in rats and suppressesdimethylnitrosamine-induced fibrosis in the liver by reduc-ing �–smooth muscle actin expression and collagen produc-tion (34,35). The increase in the content of pancreatic hy-droxyproline, an amino acid common to all collagens, ob-served in male 20-week-old WBN/Kob versus Wistar ratsand the reduction with estradiol are clear evidence of theeffects on pancreatic fibrosis in the present model. Althoughthe mechanisms remain unclear, we can speculate that es-tradiol either exerts effects on pancreatic “stellate” cells (ananalogy to hepatic stellate cells) or decreases the underlyinginflammation.
In conclusion, estradiol dose-dependently attenuated aci-nar cell apoptosis and development of chronic pancreatitis,independent of change in endogenous corticosterone andmale hormones, presumably by suppressing infiltration andfunctions of T cells. Further investigations are now neededto clarify the impact of estradiol on different models ofchronic pancreatitis and to determine whether suppressiveeffects are universal.
S. NAKAMURA ET AL. ESTRADIOL IN APOPTOSIS AND PANCREATITIS e65

REFERENCES1. Mori Y, Yokoyama J, Nishimura M, et al. A new diabetic strain of rat with
exocrine pancreatic insufficiency. In: Tiengo A, Del Prato S, Alberti KG, et al.,eds. Diabetes secondary to pancreatopathy. Amsterdam: Elsevier; 1988:107–112.
2. Ohashi K, Kim J-H, Hara H, et al. WBN/Kob rats: a new spontaneously occurringmodel of chronic pancreatitis. Int J Pancreatol. 1990;6:231–247.
3. Tsuchitani M, Saegusa T, Narama I, et al. A new diabetic strain of rat(WBN/Kob). Lab Anim. 1985;19:200–207.
4. Hashimoto T, Yamada T, Yokoi T, et al. Apoptosis of acinar cells is involved inchronic pancreatitis in WBN/Kob rats: role of glucocorticoids. Pancreas 2000;21:296–304.
5. Yamada T, Hashimoto T, Sogawa M, et al. Involvement of T cells in develop-ment of chronic pancreatitis in male Wistar Bonn/Kobori rats: protective effectsof tacrolimus. Am J Physiol. 2001;281:G1397–1404.
6. Cutolo M, Sulli A, Seriolo B, et al. Estrogens, the immune response and auto-immunity. Clin Exp Rheumatol. 1995;13:217–226.
7. Allen JB, Blatter D, Calandra GB, et al. Sex hormonal effects on the severity ofstreptococcal cell wall-induced polyarthritis in the rat. Arthritis Rheum. 1983;26:560–563.
8. Grossman C. Possible underlying mechanisms of sexual dimorphism in the im-mune responses, fact and hypothesis. J Steroid Biochem. 1989;34:241–251.
9. Hawkins T, Gala RR, Dunbar JC. The effect of neonatal sex hormone manipu-lation on the incidence of nonobese diabetes. Proc Soc Exp Biol. 1993;202:201–205.
10. Makino S, Kunimoto K, Muraoka Y, et al. Effect of castration on the appearanceof diabetes in NOD mouse. Exp Anim 1981;30:137–140.
11. Nakama K, Akimoto T, Shimizu M, et al. Inhibitory effect of estrogen on thedevelopment of diabetic symptoms in male WBN/Kob rats [in Japanese]. Dia-betic Animal, 2, IYAKU Journal, Osaka, 1988:130–136.
12. Ohashi K, Hara H, Aso R, et al. Effects of various drugs on chronic pancreatitis-like lesions in WBN/Kob rat [in Japanese]. Jikken Dobutsu. Exp Anim. 1989;3:169–177.
13. Yamada T, Hoshino M, Hayakawa T, et al. Dietary diosgenin attenuates subacuteintestinal inflammation associated with indomethacin in rats. Am J Physiol. 1997;273:G355–364.
14. Gavrieli Y, Sherman Y, Ben-Sasson SA. Identification of programmed cell deathin situ via specific labeling of nuclear DNA fragmentation. J Cell Biol. 1992;119:209–214.
15. Shirai T, Takahashi S, Mori T, et al. Immunohistochemically demonstrated an-drogen receptor expression in the rat prostate during carcinogenesis induced by3, 2�-dimethyl-4-aminobiphenyl with and without testosterone. Urol Oncol.1995;1:263–268.
16. Grisham MB, Benoit JN, Granger DN. Assessment of leukocyte involvementduring ischemia and perfusion of intestine. Method Enzymol. 1990;186:729–741.
17. Blumenkrantz N, Asobe-Hansen G. A quick and specific assay for hydroxypro-line. Anal Biochem. 1973;55:288–291.
18. Ueberberg H, Laque K, Trieb G. Comparative studies on the circadian rhythm ofcorticosterone lipid and cholesterol levels in adrenals and blood of rats. Chro-nobiol Int. 1984;1:41–49.
19. Asakawa N, Koyama M, Hashimoto Y, et al. Studies on the metabolic fate offlutamide (4); biliary secretion, enterohepatic circulation, absorption from gastro-intestinal tract, metabolism and drug-metabolizing enzymes induction in rats.Drug Metabolism (Yakubutsu Dotai). 1995;10:470–483.
20. Jung CG, Kamiyama T, Agui T. Elevated apoptosis of peripheral T lymphocytesin diabetic BB rats. Immunology. 1999;98:590–594.
21. Elder ME, Maclaren NK. Identification of profound peripheral T-lymphocyteimmunodeficiencies in the spontaneously diabetic BB rat. J Immunol. 1983;130:1723–1731.
22. Rijhsinghani AG, Thompson K, Bhatia SK, et al. Estrogen blocks early T celldevelopment in the thymus. Am J Reprod Immunol. 1996;36:269–277.
23. Bateman A, Singh A, Kral T, et al. The immune-hypothalamic-pituitary-adrenalaxis. Endocr Rev. 1989;10:92–112.
24. Kimura K, Shimosegawa T, Sasano H, et al. Endogenous glucocorticoids de-crease the acinar cell sensitivity to apoptosis during caerulein pancreatitis in rats.Gastroenterology. 1998;114:372–381.
25. Felig P, Baxter JD, Frohman LA. Endocrinology and metabolism. In: WilliamFG, ed. Review of medical physiology. New York: McGraw-Hill, 1981:218–270.
26. Richard RG, Ulrich W. Corticosteroid-binding globulin in the rat: studies on thesex difference. Endocrinology. 1965;77:841–851.
27. Roubinian JR, Talal N, Greenspan JS, et al. Effect of castration and sex hormonetreatment on survival, anti-nucleic acid antibodies, and glomerulonephritis inNZB/NZW1 mice. J Exp Med. 1978;147:1568–1583.
28. Corbishley TP, Iqbal MJ, Wilkinson ML, et al. Androgen receptor in human andmalignant pancreatic tissue cell lines. Cancer. 1986;57:1992–1995.
29. Diaz-Sanchez V, Morimoto S, Morales A, et al. Androgen receptor in the ratpancreas: genetic statement and steroid regulation. Pancreas. 1995;11:241–245.
30. Pousette Å. Demonstration of an androgen receptor in rat pancreas. Biochem J.1976;157:229–232.
31. Brooks JR, Berman C, Nguyen H, et al. Effect of castration, DES, flutamide, andthe 5�-reductase inhibitor, MK-906, on the growth of the Dunning rat prostaticcarcinoma, R-3327. Prostate. 1991;18:215–217.
32. Aso Y, Akaza H, Kumamoto Y, et al. Phase I study of flutamide, a nonsteroidalantiandrogen, in patients with prostatic cancer. Acta Urol Jpn. 1993;38:381–389.
33. Luster MI, Hayes HT, Korach K, et al. Estrogen immunosuppression is regulatedthrough estrogenic responses in the thymus. J Immunol. 1984;133:110–116.
34. Shimizu I, Mizobuchi Y, Yasuda M, et al. Inhibitory effect of estradiol onactivation of rat hepatic stellate cells in vivo and in vitro. Gut. 1999;44:127–136.
35. Yasuda M, Shimizu I, Shibata M, Ito S. Suppressive effect of estradiol on di-methylnitrosamine-induced fibrosis in rats. Hepatology. 1999;29:719–727.
e66 Pancreas, Vol. 26, No. 3, 2003