energy balance regulating neuropeptides are expressed through pregnancy and regulated by...
TRANSCRIPT

Research ArticleEnergy Balance Regulating Neuropeptides Are Expressedthrough Pregnancy and Regulated by Interleukin-6 Deficiencyin Mouse Placenta
Patricia Pazos123 Luis Lima12 Carlos Dieacuteguez123 and Mariacutea C Garciacutea123
1 Department of Physiology Research Center of Molecular Medicine and Chronic Diseases (CIMUS)University of Santiago de Compostela Avenida de Barcelona sn 15782 Santiago de Compostela Spain
2 Instituto de Investigacion Sanitaria de Santiago de Compostela (IDIS) 15706 Santiago de Compostela Spain3 CIBER Fisiopatologıa Obesidad y Nutricion (CB0603) Instituto de Salud Carlos III (ISCIII)Ministerio de Economıa y Competitividad (MINECO) 15706 Santiago de Compostela Spain
Correspondence should be addressed to Marıa C Garcıa mariagarciagarciausces
Received 21 November 2013 Revised 21 January 2014 Accepted 2 February 2014 Published 12 March 2014
Academic Editor Georgios Valsamakis
Copyright copy 2014 Patricia Pazos et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The placenta produces a number of signaling molecules including metabolic and reproductive hormones as well as severalinflammatory mediators Among them Interleukin-6 (IL-6) a well-known immune and metabolic regulator acts peripherallymodulating metabolic function and centrally increasing energy expenditure and reducing body fat IL-6 interacts with keyhypothalamic neuropeptidergic systems controlling energy homeostasis such as those producing the orexigenicanabolicneuropeptide Y (NPY) and agouti-related peptide (AgRP) and anorecticcatabolic neuropeptides proopiomelanocortin (POMC)and cocaine and amphetamine regulated transcript (CART) Human and rat placenta have been identified as source of theseneuropeptides but their expression and regulation in murine placental tissues remain unknownTherefore placental mRNA levelsof IL-6 NPY AgRP POMC andCART at different pregnancy stages (gestational days 13 15 and 18) were analyzed by real time PCRas were the effect of IL-6 deficiency (IL-6 knockout mice) on their placental expression Our results showed that placenta-derivedneuropeptides were regulated by gestational age and IL-6 throughout the second half of mouse pregnancy These data suggest thatIL-6 may participate in the fine tune control of energy balance during pregnancy by extending its action as a metabolic signal tothe main organ at the fetomaternal interface the placenta
1 Introduction
Energy homeostasis defined as the process whereby bodyweight and energy reserves are maintained stable over longperiods of time is tightly regulated through the complexinteractions of the brain and peripheral organs [1 2] Recip-rocal neural networks within the hypothalamus and the brainstem act in response to peripheral (metabolic endocrineand neural) signals of metabolic status matching energyintake with energy outputs such as basal metabolism activitythermogenesis and reproduction [3ndash5]
Afferent endocrine signals projecting to the brain toencode short- and long-term energy status comprise a widevariety of tissue-specific signalling molecules including
(i) adipokines (eg leptin adiponectin resistin andinterleukin-6 (IL-6)) secreted by the expanding adiposetissue (ii) pancreatic hormones such as insulin and amylin(iii) gastrointestinal peptides such as ghrelin glucagon-likepeptide I and peptide YY and (iv) gonadal and placentalhormones such as steroids and placental lactogens [2 6ndash8]Several hypothalamic neuronal populations express specificreceptors for these metabolic hormones and communicatewithin each other via chemical synapses and the releaseof neurotransmitters and neuropeptides characterizedby having either direct or indirect effects on body weightregulation and reproductive function [9]
Two opposing cell types in the arcuate nucleus (ARC)are a relevant example of the integrative nature of the
Hindawi Publishing CorporationInternational Journal of EndocrinologyVolume 2014 Article ID 537603 10 pageshttpdxdoiorg1011552014537603
2 International Journal of Endocrinology
hypothalamus in the regulation of energy metabolism andfertility [10] One type produces the orexigenic and anabolicneuropeptide Y (NPY) and agouti-related peptide (AgRP) aswell as gamma-aminobutyric acid (GABA) The other pro-duces cocaine and amphetamine regulated transcript (CART)and proopiomelanocortin- (POMC-) derived peptides suchas alpha melanocyte stimulating hormone (120572MSH) thatpromote anorexia by inhibiting food intake and increasescatabolic processes 120572MSHmodulates its downstream home-ostatic signalling via their action at melanocortin recep-tors MC3R and MC4R which are antagonized by AgRP[2] The coordinated regulation of these neurons andtheir efferent projections to key brain regions such as theparaventricular nucleus (PVN) and the medial preopticand lateral hypothalamic area (LH) contributes to regu-late energy metabolism gonadotropin-releasing hormone(GnRH) release and pituitary-gonadal axis activity [2 10 11]
The placenta a unique and autonomous transient organrepresents the primary immunological andnutrient transportbarrier between the mother and the fetus Throughout itsentire lifespan the placenta is also able to produce as wellas respond to a variety of signalling molecules requiredfor pregnancy establishment and maintenance but also formaternal adaptation to pregnancy as well as fetal growth anddevelopment Placental secretory factors include among oth-ers steroid hormones growth factors metabolic hormones(eg leptin [12 13] adiponectin [14 15] and resistin [16 17])and pro- and anti-inflammatory cytokines as interleukins-6 and -10 (IL-6 and IL-10) [18 19] Despite the lack ofinnervation in this organ production of energy balanceregulating neuropeptides has been reported in rat [20 21] andhuman [22ndash25] placenta where they may play a regulatoryrole in placental hormonal secretion that affectsmaternal andfetal metabolism However the physiological relevance andregulation of these placental neuropeptides remain unclear
IL-6 is a pluripotent cytokine not only involved in theimmune response but with multiple relevant effects onmanydifferent cell types including those of the nervous [26] andreproductive systems [27 28] Recent evidence from ourgroup [29] and others [30ndash32] points to the IL-6 system asa key pathway involved in the central regulation of foodintake and energy metabolism Hence centrally (astrocyte-restricted) [26] and total IL-6 deficiency in mice [33ndash35] areassociated with increased body weight and adiposity at youngand late ages while long-term central (intracerebroventric-ular) but not peripheral IL-6 treatment exerts a restorativeeffect on fat mass values [36] The antiobesity IL-6 actionlikely involves enhanced thermogenesis and lipid oxidationrather than reduced food intake [29 37ndash39] To control thisphysiological functions in health [29 38 40] and diseaseconditions [41] IL-6 engages its widely expressed specificreceptor IL-6 receptor alpha [42] to modulate the expressionof key anabolic and catabolic hypothalamic neuropeptides atthe level of the ARC PVN and LH [31 42 43]
Our group has recently demonstrated that pregnancyin the mouse is associated with a progressive increase incirculating IL-6 levels while hypothalamic IL-6 and IL-6Ra expression are depressed [29] These changes in thecentral and peripheral expression of the IL-6 system are
accompanied by specific adaptations in the damrsquos hypotha-lamic circuits governing energy balance notably at the levelof the ARC and PVN Major sources of circulating IL-6during this physiological state include the expanding adiposetissue as well as the placenta where it has been relatedto disarrangements in maternal-fetal nutrient (ie lipid)transfer in complicated human pregnancies [44] Howeverlittle is known about the possible influence of this cytokineon the placental production of energy balance regulatingneuropeptides Thus the present study was designed todetermine whether during the second half of pregnancyNPY AgRP POMC and CART mRNA are expressed andontogenetically modulated in the mouse placenta Usinginterleukin-6 knockout mice (IL-6 KO mice) as a modelthe effects of IL-6 deficiency on their gestational pattern ofplacental expression were also investigated
2 Materials and Methods21 Animals IL-6 deficient mice (B6129S2-IL-6 tm1 kopfJsourced from Jackson Laboratories) [45] that had undergoneeleven backcrosses to the C57BL6 background and theirwild-type congenic C57BL6J controls were purchased fromJackson laboratories (Charles River Barcelona Spain stocknumbers 002650 and 000664 resp) After weaning animalswere housed 5-6 per cage under controlled temperatureconditions (22∘C) and a 12-hour lightdark cycle with freeaccess to water and rodent chow (2019s Teklad GlobalHarlan Spain) Age-matched female WT and IL-6 KO mice(12 weeks old) were always used for experiments Ethicalapproval was obtained from the University of Santiago deCompostela Bio-ethics Committee and all the procedureswere conducted according to the regulations of the EuropeanCommunity for the care and use of experimental animals
22 Experimental Design To obtain pregnant animalstwelve-week-old female IL-6 KO or WT mice (119899 = 27 and38 resp) were mated for three days with stud mice of thesame genotype Successful matings as judged by the presenceof a vaginal plug in the early morning (day of occurrence =day 0 after coitum) were detected in a total of 22 IL-6 KOmice (81) and 28WT (74) On the corresponding day ofpregnancy (day 13 15 or 18) timed pregnant animals wereanesthetized and after collection of serum samples by cardiacpuncturewere sacrificed by decapitationAbout 82 and 61ofthe plugged IL-6KOandWTmicewere pregnant (119899 = 18 and17 resp) and only one dam of each genotype was excludedfor the analysis due to presence of less than 7 viable (normalappearing) fetuses (Table 1) Placental samples were removedweighed frozen on dry ice and stored in ndash80∘C until furtheranalysis At each gestational age analyzed the number offetuses per litter was similar in WT and IL-6 KO groups aswere the weights of conceptuses and placentas (Table 1) Weused 5-6 placentas per experimental group extracted fromdifferent mothers All the samples were analyzed individuallyand samples were not pooled
23 Serum Assays Circulating serum levels of interleukin6 were assayed by ELISA using a commercial kit (IL-6
International Journal of Endocrinology 3
Table 1 Litter size weights of dams conceptuses and placentas in WT and IL-6 KO dams
Gestational day Dam weight (g) Conceptus weight (mg) Placental weight (mg) Litter sizeWT IL-6 KO WT IL-6 KO WT IL-6 KO WT IL-6 KO
13 2987 plusmn 057 2815 plusmn 049lowast
410 plusmn 64 421 plusmn 41 77 plusmn 13 72 plusmn 9 88 plusmn 08 85 plusmn 08
(6) (6)
15 3461 plusmn 072 3167 plusmn 068lowast
756 plusmn 54 696 plusmn 37 101 plusmn 12 110 plusmn 11 98 plusmn 04 92 plusmn 06
(5) (5)
18 3864 plusmn 069 3627 plusmn 078lowast
1555 plusmn 70 1658 plusmn 74 96 plusmn 2 85 plusmn 12 85 plusmn 04 78 plusmn 03
(5) (6)Number of dams used in each group is shown in parentheseslowast
119875 lt 005 versus pregnant WT mice at the same gestational age two-tailed Studentrsquos 119905-test
Table 2 Primers and probes used for qPCR
Genes Primer sequences GenBank accesion number Product size (bp)
HPRTF 51015840-AGCCGACCGGTTCTGTCAT-31015840
NM 0135562 72R 51015840-GGTCATAACCTGGTTCATCATCAC-31015840
Pb 51015840-CGACCCTCAGTCCCAGCGTCGTGAT-31015840
IL-6F 51015840-CTATACCACTTCACAAGTCGGAGG-31015840
NM 0311681 77R 51015840-TGCACAACTCTTTTCTCATTTCC-3Pb 51015840-TTAATTACACATGTTCTCTGGGAAATCG-31015840
NPYF 51015840-ACAGAAAACGCCCCCAGAAC-31015840
NM 0234562 72R 51015840-CGGGAGAACAAGTTTCATTTCC-31015840
Pb 51015840-AGGCTTGAAGACCCTTCCATGTGG TGAT-31015840
AgRPF 51015840-ACAACTGCAGACCGAGCAGAA-31015840
NM 0074272 98R 51015840-CGACGCGGAGAACGAGACT-31015840
Pb 51015840-CAGAAGGCAGAAGCTTTGGC GG AGGT-31015840
CARTF 51015840-CGCAT TCCGATCT ACGAGAAGAA-31015840
NM 0010814932 84R 51015840-CCT GGCCCCTT TCCTCACT-31015840
Pb 51015840-CCAAGTCCCCATGTGTGACGCTGGAG-31015840
HPRT F 51015840-CAGTCCCAGCGTCGTATT-3 NM 0135562 139R 51015840-AGCAAGTCTTTCAGTCCTGTC-31015840
POMC F 51015840-TCCATAGACGTGTGAGCTG-31015840 NM 139326 174R 51015840-GACGTACTTCCGGGGATTTT-31015840
Abcam Cambridge UK) according to the manufacturerrsquosinstructions
24 Real Time qPCR Real-time quantitative RT-PCR(qPCR) analysis was carried out as previously described [46]Total RNA was obtained by homogenization in TRIZOLreagent according to the manufacturerrsquos instructions(Invitrogen Barcelona Spain) First-strand cDNA wassynthesized from 15 120583g of total RNA in a 30 120583L reaction using200U Maloney murine leukemia virus reverse transcriptaseand randomhexamer primers (Invitrogen Barcelona Spain)qPCR was performed using a Mastercycler EP Realplex real-time PCR system (Eppendorf) Probe detection (UniversalMaster mix Applied Biosystems) was used for IL-6 [47]and the neuropeptides NPY AgRP and CART expressionanalysis [48] (Table 2) SYBR Green was used for POMCmRNA detection [21] (Luminaris Color HiGreen qPCRMaster Mix Thermo Fisher Scientific) and was followed by amelting curve to assure the primers specificity (Table 2)
A standard curve was run in each assay with an arbitraryvalue assigned to the highest standard and correspondingvalues to the subsequent dilutions A nontemplate reactionwas included during each experiment to control for DNAcontamination Mouse hypoxanthine guanine phosphoribo-syl transferase (HPRT) was used as control house-keepinggene [49ndash51] The relative expression levels were estimatedusing the comparative threshold cycle method [52] and allsamples were normalized againstHPRTexpression valuesWehave previously usedHPRTas reference gene in the rat [14 21]but not in the mouse placenta Therefore we tested whetherthe HPRTmRNA levels were stable in placentas frommice ofthe same genotype at different gestational ages HPRTmRNAexpression was similar among gestational age groups hencevalidating its use as reference gene in the present study
To verify the identity of amplified cDNAs PCR productswere electrophoresed on a 15 agarose gel which yieldedDNA fragments of the expected length for all mRNAs andwere confirmed by sequencing (data not shown)
4 International Journal of Endocrinology
300
250
200
150
100
50
0
13 15 18
Seru
m IL
-6(p
gm
L)
Gestational day
lowast
WT
Figure 1 IL-6 levels in serum of WT (C57BL6) mice 12-week-oldWT mice were time-pregnant and serum IL-6 levels at differentpregnancy stages (13 15 and 18 days) were assessed by ELISA 119899 =5-6 Each value represents the mean plusmn SEM One-way ANOVAlowast
119875 lt 005 versus 13 days pregnant
25 Statistical Analysis Results are given as mean plusmn SEMAll data were analyzed using Graph Pad Prism version 600for Windows (GraphPad Software La Jolla California USA)Comparisons between two groups were performed withunpaired two-tailed Studentrsquos 119905-test and one-way analysis ofvariance (ANOVA) followed by the Bonferroni post hoctest when differences between more than two experimentalgroups were analyzed
3 Results
31 Increased Serum IL-6 Levels during Pregnancy in WTMice Firstly circulating levels of IL-6 were assessed duringpregnancy As expected [29 53] serum IL-6 levels increasedas pregnancy progressed although a significant stimulatoryeffect was only evident at gestational day 18 from values atgestational day 13 (119875 lt 005) (Figure 1)
32 IL-6 mRNA Expression Is Up-Regulated in the Placentaof Pregnant WT Mice Throughout Pregnancy Thereafter weanalyzed the transcriptional profile of placental IL-6 expres-sion during mouse pregnancy IL-6 mRNA was detected inall placentas tested from pregnantWT dams (C57BL6 femalemice) at gestational days 13 15 and 18 Our data showed thatIL-6 gene expression increased with gestational age in themouse placenta with significantly higher mRNA levels ondays 15 and 18 of pregnancy in comparison to values on day13 (119875 lt 001 and 119875 lt 0001 resp) (Figure 2)
33 NPYmRNAExpression Is Up-Regulated by IL-6Deficiencyin the Mouse Placenta Next we evaluated the placental NPYmRNA levels and whether absence of IL-6 might influenceits gestational profile during mouse pregnancy Althoughin the placenta of WT mice there was a trend towards an
0
5
4
3
2
1
lowastlowastlowast
lowastlowast
IL-6
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
Gestational day
WT
Figure 2 IL-6 mRNA expression in placenta of WT (C57BL6)mice 12-week-old WT mice were time-pregnant and placental IL-6 mRNA levels at different pregnancy stages (13 15 and 18 days)were assessed by real-time qPCR 119899 = 5-6 Relative mRNA levelswere normalized to 13 days pregnant mice values as 1 Each valuerepresents the mean plusmn SEM One-way ANOVA lowastlowast119875 lt 001 versus13 days pregnant mice values
increased NPY mRNA expression at gestational day 15 inrelation to the other stages of pregnancy studied no statisticaldifference was observed (Figure 3(a)) On the contrary inabsence of IL-6 placental NPY mRNA expression remainedunchanged between 13 and 15 days of pregnancy reaching thehighest values at the end of the gestational period (119875 lt 001)(Figure 3(b))
34 Lack of IL-6 Does Not Affect AgRP mRNA Expressionin the Mouse Placenta AgRP mRNA was expressed in themouse placenta at all gestational ages assessed but showeda slightly different profile during pregnancy from that ofNPYThus AgRP mRNA content in the placenta of pregnantWT mice did not change significantly from 13 to 18 days ofpregnancy (Figure 4(a)) being unaffected by IL-6 deficiency(Figure 4(b))
35 POMC mRNA Expression Is Down-Regulated by IL-6Deficiency in the Mouse Placenta Placental POMC mRNAexpression in WT mice was markedly reduced at the latestpregnancy stage (gestational day 18 119875 lt 005) whileit remained unchanged from 13 to 15 days of pregnancy(Figure 5(a)) However IL-6 KO mice showed a significantly(119875 lt 005) reducing trend for placental POMC mRNAcontent from gestational days 13 to 18 (Figure 5(b))
36 CART mRNA Expression Is Up-Regulated by IL-6 Defi-ciency in the Mouse Placenta Finally we studied the tran-scriptional profile of CART expression in the placenta ofWT and IL-6 KO mice during pregnancy Our data showedthat this catabolic neuropeptide had a constitutive and fairlyconstant expression in the placenta of WT mice throughout
International Journal of Endocrinology 5
0
Gestational day
WT
30
25
20
15
10
05
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
(a)
0
Gestational day
IL-6 KO
lowastlowast
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
6
5
4
3
2
1
13 15 18
(b)
Figure 3 NPYmRNA expression in placenta ofWT and IL-6 KOmice 12-week-oldWT and IL-6 KOmice were time-pregnant and placentalNPYmRNA levels in (a)WT and (b) IL-6 KOmice at different pregnancy stages (13 15 and 18 days) were assessed by real-time qPCR 119899 = 5-6Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents the mean plusmn SEMOne-way ANOVA lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(b)
Figure 4 AgRP mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental AgRP mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM
pregnancy (Figure 6(a)) A similar pattern was observed inthe placentas from IL-6 KO mice until gestational day 18when the highest levels of placental CARTmRNA expressionwere reached (119875 lt 001) (Figure 6(b))
4 Discussion
Pregnancy is now recognized as a state of adaptive low-grade-inflammation induced by a progressive increase ofcirculating levels inflammatory mediators such as cytokines
chemokines and acute-phase reactants [53 54] Cytokinescentral players of this inflammatory response are secreted bythe immune cells and adipocytes but also by placental cells[55] Placental cytokine production and action are crucialto control trophoblast implantation embryo developmentand feto-maternal tolerance [55] However once pregnancyis established the physiological relevance of most placentalcytokines remains unclear
Proinflammatory cytokines such as IL-6 TNF alphaand IL-1 are known to negatively influence peripheral lipid
6 International Journal of Endocrinology
Gestational day
WT
0
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)20
15
10
05lowast
13 15 18
(a)
0
Gestational day
IL-6 KO
20
15
10
05
lowast
lowastlowast
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)
13 15 18
(b)
Figure 5 POMC mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental POMC mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
CART
mRN
A le
vels
(rel
ativ
e val
ues)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
30
25
20
15
10
05
CART
mRN
A le
vels
(rel
ativ
e val
ues) lowast
13 15 18
(b)
Figure 6 CART mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental CART mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (1315 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 versus 13 days pregnant mice of respective genotype
and glucose metabolism Hence by secreting these immunefactors the placenta may enhance the contribution madeby adipose tissue to induce maternal insulin resistance andensure a proper glucose transfer to the growing fetus In thisstudy we found increased expression of IL-6 mRNA in themouse placenta during gestation These results are in agree-ment with previous reports of an enhanced IL-6 placentalproduction during the latest stages of rat [54] and humanpregnancies [56] To our knowledge the present work is thefirst one to determine the ontogenetic profile of IL-6 mRNA
levels in the mouse placenta In fact De and coworkers [57]showed high levels of placental IL-6 bioactivity but failed todemonstrate its constitutive transcriptional expression in thismouse reproductive organ The reason for this discrepancymay be the different methodologies employed since in thisprevious study IL-6mRNA levelsweremeasured byNorthernblot while we used a more sensitive technique RT-qPCRWe[29] and others [53 58] have previously found elevated IL-6 serum and adipose tissue mRNA levels during mouse latepregnancy Our data do not allow us to determine the relative
International Journal of Endocrinology 7
contribution of both tissues to the amount of this cytokinepresent in maternal circulation but the similar pattern of IL-6 mRNA overexpression in both tissues suggests that bothcould be a source of circulating IL-6 in pregnancy
Our group has recently shown that in addition toits peripheral metabolic effects IL-6 modulates the cen-tral mechanisms associated to maternal hyperphagia andincreased adiposity in the pregnantmouse [29] Although thetranscriptional expression of its hypothalamic neuropeptidetargets NPY AgRP POMC and CART in the human [2425] and rat placenta [20] is well established whether theyare also placentally produced and regulated by IL-6 duringmouse pregnancy remains unknown In order to clarify thisissue we determined the placental mRNA expression ofthese energy balance regulating neuropeptides throughoutpregnancy (from gestational days 13 to 18) in WT and IL-6deficient mice
The results of the present study revealed a similar tran-scriptional regulation of ARC neuropeptides in the mouseplacenta from that previously reported in the hypothalamus[29] Thus POMC mRNA levels were reduced from gesta-tional days 13 to 18 while the expression of the orexigenicand anabolic neuropeptides NPYAgRP remains unchangedA similar absence of relation between gestational age andplacental NPY mRNA content has been shown in otherspecies [20 25] On the contrary the placental ontogeneticexpression of the melanocortin system members POMC andAgRP seems to be a rather controversial issue Thereforesome studies reported a decrease in AgRP mRNA levels withgestational age and no change in POMC levels whilst othersshowed an upregulated mRNA expression of both neuropep-tides in human [24] and rat placentas [20 21] As far as we areaware the mRNA expression of CART has been previouslyexamined neither in human nor in murine placental tissuesIn this sense recent data from our group demonstrated thatthemRNA levels of this catabolic and anorectic neuropeptideare markedly increased in the rat placenta at term [20 21]However in the present study we failed to detect any changein CART gene expression throughout mouse pregnancy Thereason for this discrepancy is unclear but itmight reflect somebiological differences between species
Another novel finding of the present study is the fact thatIL-6 acting on an autocrine or paracrine manner modulatesthe placental expression of classical ARC energy balance reg-ulatory peptides as shown by IL-6 KOmice data Thus NPYand CART mRNA levels are upregulated by IL-6 deficiencyat late pregnancy while POMC gene expression is downregu-lated in placentas from 15-day pregnant IL-6 KOmice Theseresults indicate that during the third week of pregnancylack of IL-6 settles an anabolic milieu in the mouse placentaby reciprocally modulating the transcriptional expression ofneuropeptides known to either promote NPY or repressPOMC net energy gain As we have previously demonstratedin the hypothalamus of the mouse in late pregnancy a lackof IL-6 upregulates NPYAgRP mRNA levels in the ARCit seems that IL-6 deficiency during late pregnancy exertsa stimulatory effect on NPY gene transcription whatever isits source However the results obtained regarding placentalCARTPOMC gene expression were somewhat unexpected
Increased hypothalamic CART or POMC production hasbeen reported after endotoxemic challenge or central admin-istration of other well-known proinflammatory cytokinesas IL-1 [59 60] In addition exercise-induced hypothala-mic IL-6 and IL-10 in obese rats [61] and IL-6 astrocyte-targeted overexpression in female mice have been shownto increase POMC expression in the ARC [62] Converselytotal knockout of IL-6 in mice restored to virgin values thereduced POMC mRNA levels in the ARC of late pregnantmice probably reflecting a compensatorymechanism exertedby other factors [29] On the contrary the data reportedherein indicate that during the second half of pregnancy thiscompensatory effect on POMC gene expression is not settledin the mouse placenta Considering the above mentionedresults and the fact that CART and POMC are colocalized inthe ARC a similar pattern of placental expressionmight havebeen expected for both neuropeptides However we havenot performed in situ hybridization or immunohistochemicalstudies to characterize cellular distribution of CART andPOMC in placental tissue Therefore we cannot exclude thepossibility that different placental cell types could producethese neuropeptides and respond to absence of IL-6 in adifferent manner In fact with respect to their hypothalamicregulation it is worth to mention that CART mRNA hasbeen reported in a number of hypothalamic cell populationsbesides the ARC where its expression has been shown to bedifferentially modulated from that of POMC in response toendotoxin challenge [63]
Finally in relation to the previous point it is importantto take into account that the experimental approach used inthe current study was designed to assess during the secondhalf of pregnancy global changes in IL-6 and main energyregulating neuropeptides gene expression in the mouse pla-centa Additionally the IL-6 deficient mouse was chosen toaddress the influence of total absence of endogenous IL-6 onthis setting However the murine placenta consists of threemayor cell layers of maternal or placental origin the outermaternal decidua the middle junctional zone composedof fetoplacental trophoblast cells and the inner labyrinthzone [64] Therefore further in vitro studies involving eitherdecidual or trophoblast cells in culture would be helpful todetermine the ability of each cell population to produce andsecrete these neuropeptides and confirm the contribution ofIL-6 to their transcriptional modulation
Whatever is the case these results of the current studysuggest that IL-6 by exerting a direct or indirect regulatoryaction in the transcriptional control of NPY CART andPOMC expression may play a role in the homeostaticresponse to energy availability in mouse placenta Finallywhether placental neuropeptide protein levels correlate withthe ontogenetic neuropeptide mRNA pattern herein shownwill merit further investigation
5 Conclusions
In summary we demonstrate that as in other species theclassical hypothalamic neuropeptidergic systems involvedin energy homeostasis are present in the mouse placenta
8 International Journal of Endocrinology
These central signals display a specific ontogenetic expressionpattern in absence of IL-6 with major changes settled inplacentas at term Thus our results suggest that IL-6 a well-known immune and metabolic modulating factor mightmodulate their transcriptional expression by acting on anautocrine or paracrine manner Collectively these resultssuggest that during an energetic challenging condition suchas pregnancy IL-6 may also extend its action as a metabolicsignal to the main organ at the maternal-fetal interface theplacenta
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank Eva Vigo C Ruth Gonzalez and SiaraPerez for excellent technical assistance with the qPCRassays The HPRT gene primers for SYBR Green geneexpression analysis were kindly donated by professor ClaraV Alvarez Research Center of Molecular Medicine andChronicDiseases (CIMUS)This work has been supported byEuropean Community (FP72007n∘ 245009 ldquoNeuroFASTrdquo)Ministerio de Economıa y Competitividad (PP and MCGGBFU2007-62683BFI and PP MCGG and CD CIBERobn(CB0603)) and Xunta de Galicia Grants (MCGG and LLPGIDIT06PXIB208067PR) CIBER de Fisiopatologıa de laObesidad y Nutricion is an initiative of ISCIII
References
[1] J R Speakman D A Levitsky D B Allison et al ldquoSet pointssettling points and some alternative models theoretical optionsto understandhowgenes and environments combine to regulatebody adiposityrdquoDisease Models ampMechanisms vol 4 no 6 pp733ndash745 2011
[2] K W Williams and J K Elmquist ldquoFrom neuroanatomy tobehavior central integration of peripheral signals regulatingfeeding behaviorrdquoNature Neuroscience vol 15 no 10 pp 1350ndash1355 2012
[3] J E Schneider C M Klingerman and A Abdulhay ldquoSenseand nonsense in metabolic control of reproductionrdquo Frontiersin Endocrinology vol 3 article 26 2012
[4] D Richard B Monge-Roffarello K Chechi S M Labbe andE E Turcotte ldquoControl and physiological determinants ofsympatheticallymediated brown adipose tissue thermogenesisrdquoFrontiers in Endocrinology vol 3 article 36 2012
[5] MGMyers Jr andD POlson ldquoCentral nervous system controlof metabolismrdquo Nature vol 491 no 7424 pp 357ndash363 2012
[6] M Lopez S Tovar M J Vazquez L M Williams and CDieguez ldquoPeripheral tissue-brain interactions in the regulationof food intakerdquo Proceedings of the Nutrition Society vol 66 no1 pp 131ndash155 2007
[7] S R Ladyman R A Augustine and D R Grattan ldquoHormoneinteractions regulating energy balance during pregnancyrdquo Jour-nal of Neuroendocrinology vol 22 no 7 pp 805ndash817 2010
[8] K Sinchak and E J Wagner ldquoEstradiol signaling in theregulation of reproduction and energy balancerdquo Frontiers inEndocrinology vol 33 no 4 pp 342ndash363 2012
[9] A Crown D K Clifton and R A Steiner ldquoNeuropeptidesignaling in the integration of metabolism and reproductionrdquoNeuroendocrinology vol 86 no 3 pp 175ndash182 2007
[10] J W Hill J K Elmquist and C F Elias ldquoHypothalamicpathways linking energy balance and reproductionrdquo AmericanJournal of Physiology vol 294 no 5 pp E827ndashE832 2008
[11] S Kumar and G Kaur ldquoIntermittent fasting dietary restrictionregimen negatively influences reproduction in young rats astudy of hypothalamo-hypophysial-gonadal axisrdquo PLoS ONEvol 8 no 1 Article ID e52416 2013
[12] R Senaris T Garcia-Caballero X Casabiell et al ldquoSynthesis ofleptin in human placentardquo Endocrinology vol 138 no 10 pp4501ndash4504 1997
[13] N Hoogard L Hunter J S Duncan L M Williams PTrayhurn and J G Mercer ldquoLeptin and leptin receptor mRNAand protein expression in the murine fetus and placentardquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 20 pp 11073ndash11078 1997
[14] J E Caminos R Nogueiras R Gallego et al ldquoExpressionand regulation of adiponectin and receptor in human and ratplacentardquo The Journal of Clinical Endocrinology amp Metabolismvol 90 no 7 pp 4276ndash4286 2005
[15] H N Jones T Jansson and T L Powell ldquoFull-lengthadiponectin attenuates insulin signaling and inhibits insulin-stimulated amino acid transport in human primary trophoblastcellsrdquo Diabetes vol 59 no 5 pp 1161ndash1170 2010
[16] N di Simone F di Nicuolo D Marzioni et al ldquoResistinmodulates glucose uptake and glucose transporter-1 (GLUT-1) expression in trophoblast cellsrdquo Journal of Cellular andMolecular Medicine vol 13 no 2 pp 388ndash397 2009
[17] S Yura N Sagawa H Itoh et al ldquoResistin is expressed inthe human placentardquo The Journal of Clinical Endocrinology ampMetabolism vol 88 no 3 pp 1394ndash1397 2003
[18] N Noyola-Martıneza L Dıaz E Avila A Halhali F Larreaand D Barrera ldquoCalcitriol downregulates TNF-120572 and IL-6 expression in cultured placental cells from preeclampticwomenrdquo Cytokine vol 61 no 1 pp 245ndash250 2013
[19] S K Munro M D Mitchell and A P Ponnampalam ldquoHistonedeacetylase inhibition by trichostatin A mitigates LPS inducedTNF120572 and IL-10 production in human placental explantsrdquoPlacenta vol 34 no 7 pp 567ndash573 2013
[20] R Beloosesky D A Gayle F Amidi S N Ahanya M Desaiand M G Ross ldquoOntogenic expression of putative feedingpeptides in the rat fetal brain and placentardquo Nutritional Neu-roscience vol 9 no 1-2 pp 33ndash40 2006
[21] J E Caminos S B Bravo C R Gonzalez et al ldquoFood intakeregulating-neuropeptides are expressed and regulated throughpregnancy and following food restriction in rat placentardquoReproductive Biology and Endocrinology vol 6 article 14 2008
[22] S I Grigorakis E Anastasiou K Dai A Souvatzoglou andM Alevizaki ldquoThree mRNA transcripts of the proopiome-lanocortin gene in human placenta at termrdquo European Journalof Endocrinology vol 142 no 5 pp 533ndash536 2000
[23] J Robidoux L Simoneau A Masse and J Lafond ldquoActivationof L-type calcium channels induces corticotropin-releasing fac-tor secretion from human placental trophoblastsrdquo The Journalof Clinical Endocrinology ampMetabolism vol 85 no 9 pp 3356ndash3364 2000
International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003

2 International Journal of Endocrinology
hypothalamus in the regulation of energy metabolism andfertility [10] One type produces the orexigenic and anabolicneuropeptide Y (NPY) and agouti-related peptide (AgRP) aswell as gamma-aminobutyric acid (GABA) The other pro-duces cocaine and amphetamine regulated transcript (CART)and proopiomelanocortin- (POMC-) derived peptides suchas alpha melanocyte stimulating hormone (120572MSH) thatpromote anorexia by inhibiting food intake and increasescatabolic processes 120572MSHmodulates its downstream home-ostatic signalling via their action at melanocortin recep-tors MC3R and MC4R which are antagonized by AgRP[2] The coordinated regulation of these neurons andtheir efferent projections to key brain regions such as theparaventricular nucleus (PVN) and the medial preopticand lateral hypothalamic area (LH) contributes to regu-late energy metabolism gonadotropin-releasing hormone(GnRH) release and pituitary-gonadal axis activity [2 10 11]
The placenta a unique and autonomous transient organrepresents the primary immunological andnutrient transportbarrier between the mother and the fetus Throughout itsentire lifespan the placenta is also able to produce as wellas respond to a variety of signalling molecules requiredfor pregnancy establishment and maintenance but also formaternal adaptation to pregnancy as well as fetal growth anddevelopment Placental secretory factors include among oth-ers steroid hormones growth factors metabolic hormones(eg leptin [12 13] adiponectin [14 15] and resistin [16 17])and pro- and anti-inflammatory cytokines as interleukins-6 and -10 (IL-6 and IL-10) [18 19] Despite the lack ofinnervation in this organ production of energy balanceregulating neuropeptides has been reported in rat [20 21] andhuman [22ndash25] placenta where they may play a regulatoryrole in placental hormonal secretion that affectsmaternal andfetal metabolism However the physiological relevance andregulation of these placental neuropeptides remain unclear
IL-6 is a pluripotent cytokine not only involved in theimmune response but with multiple relevant effects onmanydifferent cell types including those of the nervous [26] andreproductive systems [27 28] Recent evidence from ourgroup [29] and others [30ndash32] points to the IL-6 system asa key pathway involved in the central regulation of foodintake and energy metabolism Hence centrally (astrocyte-restricted) [26] and total IL-6 deficiency in mice [33ndash35] areassociated with increased body weight and adiposity at youngand late ages while long-term central (intracerebroventric-ular) but not peripheral IL-6 treatment exerts a restorativeeffect on fat mass values [36] The antiobesity IL-6 actionlikely involves enhanced thermogenesis and lipid oxidationrather than reduced food intake [29 37ndash39] To control thisphysiological functions in health [29 38 40] and diseaseconditions [41] IL-6 engages its widely expressed specificreceptor IL-6 receptor alpha [42] to modulate the expressionof key anabolic and catabolic hypothalamic neuropeptides atthe level of the ARC PVN and LH [31 42 43]
Our group has recently demonstrated that pregnancyin the mouse is associated with a progressive increase incirculating IL-6 levels while hypothalamic IL-6 and IL-6Ra expression are depressed [29] These changes in thecentral and peripheral expression of the IL-6 system are
accompanied by specific adaptations in the damrsquos hypotha-lamic circuits governing energy balance notably at the levelof the ARC and PVN Major sources of circulating IL-6during this physiological state include the expanding adiposetissue as well as the placenta where it has been relatedto disarrangements in maternal-fetal nutrient (ie lipid)transfer in complicated human pregnancies [44] Howeverlittle is known about the possible influence of this cytokineon the placental production of energy balance regulatingneuropeptides Thus the present study was designed todetermine whether during the second half of pregnancyNPY AgRP POMC and CART mRNA are expressed andontogenetically modulated in the mouse placenta Usinginterleukin-6 knockout mice (IL-6 KO mice) as a modelthe effects of IL-6 deficiency on their gestational pattern ofplacental expression were also investigated
2 Materials and Methods21 Animals IL-6 deficient mice (B6129S2-IL-6 tm1 kopfJsourced from Jackson Laboratories) [45] that had undergoneeleven backcrosses to the C57BL6 background and theirwild-type congenic C57BL6J controls were purchased fromJackson laboratories (Charles River Barcelona Spain stocknumbers 002650 and 000664 resp) After weaning animalswere housed 5-6 per cage under controlled temperatureconditions (22∘C) and a 12-hour lightdark cycle with freeaccess to water and rodent chow (2019s Teklad GlobalHarlan Spain) Age-matched female WT and IL-6 KO mice(12 weeks old) were always used for experiments Ethicalapproval was obtained from the University of Santiago deCompostela Bio-ethics Committee and all the procedureswere conducted according to the regulations of the EuropeanCommunity for the care and use of experimental animals
22 Experimental Design To obtain pregnant animalstwelve-week-old female IL-6 KO or WT mice (119899 = 27 and38 resp) were mated for three days with stud mice of thesame genotype Successful matings as judged by the presenceof a vaginal plug in the early morning (day of occurrence =day 0 after coitum) were detected in a total of 22 IL-6 KOmice (81) and 28WT (74) On the corresponding day ofpregnancy (day 13 15 or 18) timed pregnant animals wereanesthetized and after collection of serum samples by cardiacpuncturewere sacrificed by decapitationAbout 82 and 61ofthe plugged IL-6KOandWTmicewere pregnant (119899 = 18 and17 resp) and only one dam of each genotype was excludedfor the analysis due to presence of less than 7 viable (normalappearing) fetuses (Table 1) Placental samples were removedweighed frozen on dry ice and stored in ndash80∘C until furtheranalysis At each gestational age analyzed the number offetuses per litter was similar in WT and IL-6 KO groups aswere the weights of conceptuses and placentas (Table 1) Weused 5-6 placentas per experimental group extracted fromdifferent mothers All the samples were analyzed individuallyand samples were not pooled
23 Serum Assays Circulating serum levels of interleukin6 were assayed by ELISA using a commercial kit (IL-6
International Journal of Endocrinology 3
Table 1 Litter size weights of dams conceptuses and placentas in WT and IL-6 KO dams
Gestational day Dam weight (g) Conceptus weight (mg) Placental weight (mg) Litter sizeWT IL-6 KO WT IL-6 KO WT IL-6 KO WT IL-6 KO
13 2987 plusmn 057 2815 plusmn 049lowast
410 plusmn 64 421 plusmn 41 77 plusmn 13 72 plusmn 9 88 plusmn 08 85 plusmn 08
(6) (6)
15 3461 plusmn 072 3167 plusmn 068lowast
756 plusmn 54 696 plusmn 37 101 plusmn 12 110 plusmn 11 98 plusmn 04 92 plusmn 06
(5) (5)
18 3864 plusmn 069 3627 plusmn 078lowast
1555 plusmn 70 1658 plusmn 74 96 plusmn 2 85 plusmn 12 85 plusmn 04 78 plusmn 03
(5) (6)Number of dams used in each group is shown in parentheseslowast
119875 lt 005 versus pregnant WT mice at the same gestational age two-tailed Studentrsquos 119905-test
Table 2 Primers and probes used for qPCR
Genes Primer sequences GenBank accesion number Product size (bp)
HPRTF 51015840-AGCCGACCGGTTCTGTCAT-31015840
NM 0135562 72R 51015840-GGTCATAACCTGGTTCATCATCAC-31015840
Pb 51015840-CGACCCTCAGTCCCAGCGTCGTGAT-31015840
IL-6F 51015840-CTATACCACTTCACAAGTCGGAGG-31015840
NM 0311681 77R 51015840-TGCACAACTCTTTTCTCATTTCC-3Pb 51015840-TTAATTACACATGTTCTCTGGGAAATCG-31015840
NPYF 51015840-ACAGAAAACGCCCCCAGAAC-31015840
NM 0234562 72R 51015840-CGGGAGAACAAGTTTCATTTCC-31015840
Pb 51015840-AGGCTTGAAGACCCTTCCATGTGG TGAT-31015840
AgRPF 51015840-ACAACTGCAGACCGAGCAGAA-31015840
NM 0074272 98R 51015840-CGACGCGGAGAACGAGACT-31015840
Pb 51015840-CAGAAGGCAGAAGCTTTGGC GG AGGT-31015840
CARTF 51015840-CGCAT TCCGATCT ACGAGAAGAA-31015840
NM 0010814932 84R 51015840-CCT GGCCCCTT TCCTCACT-31015840
Pb 51015840-CCAAGTCCCCATGTGTGACGCTGGAG-31015840
HPRT F 51015840-CAGTCCCAGCGTCGTATT-3 NM 0135562 139R 51015840-AGCAAGTCTTTCAGTCCTGTC-31015840
POMC F 51015840-TCCATAGACGTGTGAGCTG-31015840 NM 139326 174R 51015840-GACGTACTTCCGGGGATTTT-31015840
Abcam Cambridge UK) according to the manufacturerrsquosinstructions
24 Real Time qPCR Real-time quantitative RT-PCR(qPCR) analysis was carried out as previously described [46]Total RNA was obtained by homogenization in TRIZOLreagent according to the manufacturerrsquos instructions(Invitrogen Barcelona Spain) First-strand cDNA wassynthesized from 15 120583g of total RNA in a 30 120583L reaction using200U Maloney murine leukemia virus reverse transcriptaseand randomhexamer primers (Invitrogen Barcelona Spain)qPCR was performed using a Mastercycler EP Realplex real-time PCR system (Eppendorf) Probe detection (UniversalMaster mix Applied Biosystems) was used for IL-6 [47]and the neuropeptides NPY AgRP and CART expressionanalysis [48] (Table 2) SYBR Green was used for POMCmRNA detection [21] (Luminaris Color HiGreen qPCRMaster Mix Thermo Fisher Scientific) and was followed by amelting curve to assure the primers specificity (Table 2)
A standard curve was run in each assay with an arbitraryvalue assigned to the highest standard and correspondingvalues to the subsequent dilutions A nontemplate reactionwas included during each experiment to control for DNAcontamination Mouse hypoxanthine guanine phosphoribo-syl transferase (HPRT) was used as control house-keepinggene [49ndash51] The relative expression levels were estimatedusing the comparative threshold cycle method [52] and allsamples were normalized againstHPRTexpression valuesWehave previously usedHPRTas reference gene in the rat [14 21]but not in the mouse placenta Therefore we tested whetherthe HPRTmRNA levels were stable in placentas frommice ofthe same genotype at different gestational ages HPRTmRNAexpression was similar among gestational age groups hencevalidating its use as reference gene in the present study
To verify the identity of amplified cDNAs PCR productswere electrophoresed on a 15 agarose gel which yieldedDNA fragments of the expected length for all mRNAs andwere confirmed by sequencing (data not shown)
4 International Journal of Endocrinology
300
250
200
150
100
50
0
13 15 18
Seru
m IL
-6(p
gm
L)
Gestational day
lowast
WT
Figure 1 IL-6 levels in serum of WT (C57BL6) mice 12-week-oldWT mice were time-pregnant and serum IL-6 levels at differentpregnancy stages (13 15 and 18 days) were assessed by ELISA 119899 =5-6 Each value represents the mean plusmn SEM One-way ANOVAlowast
119875 lt 005 versus 13 days pregnant
25 Statistical Analysis Results are given as mean plusmn SEMAll data were analyzed using Graph Pad Prism version 600for Windows (GraphPad Software La Jolla California USA)Comparisons between two groups were performed withunpaired two-tailed Studentrsquos 119905-test and one-way analysis ofvariance (ANOVA) followed by the Bonferroni post hoctest when differences between more than two experimentalgroups were analyzed
3 Results
31 Increased Serum IL-6 Levels during Pregnancy in WTMice Firstly circulating levels of IL-6 were assessed duringpregnancy As expected [29 53] serum IL-6 levels increasedas pregnancy progressed although a significant stimulatoryeffect was only evident at gestational day 18 from values atgestational day 13 (119875 lt 005) (Figure 1)
32 IL-6 mRNA Expression Is Up-Regulated in the Placentaof Pregnant WT Mice Throughout Pregnancy Thereafter weanalyzed the transcriptional profile of placental IL-6 expres-sion during mouse pregnancy IL-6 mRNA was detected inall placentas tested from pregnantWT dams (C57BL6 femalemice) at gestational days 13 15 and 18 Our data showed thatIL-6 gene expression increased with gestational age in themouse placenta with significantly higher mRNA levels ondays 15 and 18 of pregnancy in comparison to values on day13 (119875 lt 001 and 119875 lt 0001 resp) (Figure 2)
33 NPYmRNAExpression Is Up-Regulated by IL-6Deficiencyin the Mouse Placenta Next we evaluated the placental NPYmRNA levels and whether absence of IL-6 might influenceits gestational profile during mouse pregnancy Althoughin the placenta of WT mice there was a trend towards an
0
5
4
3
2
1
lowastlowastlowast
lowastlowast
IL-6
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
Gestational day
WT
Figure 2 IL-6 mRNA expression in placenta of WT (C57BL6)mice 12-week-old WT mice were time-pregnant and placental IL-6 mRNA levels at different pregnancy stages (13 15 and 18 days)were assessed by real-time qPCR 119899 = 5-6 Relative mRNA levelswere normalized to 13 days pregnant mice values as 1 Each valuerepresents the mean plusmn SEM One-way ANOVA lowastlowast119875 lt 001 versus13 days pregnant mice values
increased NPY mRNA expression at gestational day 15 inrelation to the other stages of pregnancy studied no statisticaldifference was observed (Figure 3(a)) On the contrary inabsence of IL-6 placental NPY mRNA expression remainedunchanged between 13 and 15 days of pregnancy reaching thehighest values at the end of the gestational period (119875 lt 001)(Figure 3(b))
34 Lack of IL-6 Does Not Affect AgRP mRNA Expressionin the Mouse Placenta AgRP mRNA was expressed in themouse placenta at all gestational ages assessed but showeda slightly different profile during pregnancy from that ofNPYThus AgRP mRNA content in the placenta of pregnantWT mice did not change significantly from 13 to 18 days ofpregnancy (Figure 4(a)) being unaffected by IL-6 deficiency(Figure 4(b))
35 POMC mRNA Expression Is Down-Regulated by IL-6Deficiency in the Mouse Placenta Placental POMC mRNAexpression in WT mice was markedly reduced at the latestpregnancy stage (gestational day 18 119875 lt 005) whileit remained unchanged from 13 to 15 days of pregnancy(Figure 5(a)) However IL-6 KO mice showed a significantly(119875 lt 005) reducing trend for placental POMC mRNAcontent from gestational days 13 to 18 (Figure 5(b))
36 CART mRNA Expression Is Up-Regulated by IL-6 Defi-ciency in the Mouse Placenta Finally we studied the tran-scriptional profile of CART expression in the placenta ofWT and IL-6 KO mice during pregnancy Our data showedthat this catabolic neuropeptide had a constitutive and fairlyconstant expression in the placenta of WT mice throughout
International Journal of Endocrinology 5
0
Gestational day
WT
30
25
20
15
10
05
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
(a)
0
Gestational day
IL-6 KO
lowastlowast
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
6
5
4
3
2
1
13 15 18
(b)
Figure 3 NPYmRNA expression in placenta ofWT and IL-6 KOmice 12-week-oldWT and IL-6 KOmice were time-pregnant and placentalNPYmRNA levels in (a)WT and (b) IL-6 KOmice at different pregnancy stages (13 15 and 18 days) were assessed by real-time qPCR 119899 = 5-6Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents the mean plusmn SEMOne-way ANOVA lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(b)
Figure 4 AgRP mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental AgRP mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM
pregnancy (Figure 6(a)) A similar pattern was observed inthe placentas from IL-6 KO mice until gestational day 18when the highest levels of placental CARTmRNA expressionwere reached (119875 lt 001) (Figure 6(b))
4 Discussion
Pregnancy is now recognized as a state of adaptive low-grade-inflammation induced by a progressive increase ofcirculating levels inflammatory mediators such as cytokines
chemokines and acute-phase reactants [53 54] Cytokinescentral players of this inflammatory response are secreted bythe immune cells and adipocytes but also by placental cells[55] Placental cytokine production and action are crucialto control trophoblast implantation embryo developmentand feto-maternal tolerance [55] However once pregnancyis established the physiological relevance of most placentalcytokines remains unclear
Proinflammatory cytokines such as IL-6 TNF alphaand IL-1 are known to negatively influence peripheral lipid
6 International Journal of Endocrinology
Gestational day
WT
0
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)20
15
10
05lowast
13 15 18
(a)
0
Gestational day
IL-6 KO
20
15
10
05
lowast
lowastlowast
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)
13 15 18
(b)
Figure 5 POMC mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental POMC mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
CART
mRN
A le
vels
(rel
ativ
e val
ues)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
30
25
20
15
10
05
CART
mRN
A le
vels
(rel
ativ
e val
ues) lowast
13 15 18
(b)
Figure 6 CART mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental CART mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (1315 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 versus 13 days pregnant mice of respective genotype
and glucose metabolism Hence by secreting these immunefactors the placenta may enhance the contribution madeby adipose tissue to induce maternal insulin resistance andensure a proper glucose transfer to the growing fetus In thisstudy we found increased expression of IL-6 mRNA in themouse placenta during gestation These results are in agree-ment with previous reports of an enhanced IL-6 placentalproduction during the latest stages of rat [54] and humanpregnancies [56] To our knowledge the present work is thefirst one to determine the ontogenetic profile of IL-6 mRNA
levels in the mouse placenta In fact De and coworkers [57]showed high levels of placental IL-6 bioactivity but failed todemonstrate its constitutive transcriptional expression in thismouse reproductive organ The reason for this discrepancymay be the different methodologies employed since in thisprevious study IL-6mRNA levelsweremeasured byNorthernblot while we used a more sensitive technique RT-qPCRWe[29] and others [53 58] have previously found elevated IL-6 serum and adipose tissue mRNA levels during mouse latepregnancy Our data do not allow us to determine the relative
International Journal of Endocrinology 7
contribution of both tissues to the amount of this cytokinepresent in maternal circulation but the similar pattern of IL-6 mRNA overexpression in both tissues suggests that bothcould be a source of circulating IL-6 in pregnancy
Our group has recently shown that in addition toits peripheral metabolic effects IL-6 modulates the cen-tral mechanisms associated to maternal hyperphagia andincreased adiposity in the pregnantmouse [29] Although thetranscriptional expression of its hypothalamic neuropeptidetargets NPY AgRP POMC and CART in the human [2425] and rat placenta [20] is well established whether theyare also placentally produced and regulated by IL-6 duringmouse pregnancy remains unknown In order to clarify thisissue we determined the placental mRNA expression ofthese energy balance regulating neuropeptides throughoutpregnancy (from gestational days 13 to 18) in WT and IL-6deficient mice
The results of the present study revealed a similar tran-scriptional regulation of ARC neuropeptides in the mouseplacenta from that previously reported in the hypothalamus[29] Thus POMC mRNA levels were reduced from gesta-tional days 13 to 18 while the expression of the orexigenicand anabolic neuropeptides NPYAgRP remains unchangedA similar absence of relation between gestational age andplacental NPY mRNA content has been shown in otherspecies [20 25] On the contrary the placental ontogeneticexpression of the melanocortin system members POMC andAgRP seems to be a rather controversial issue Thereforesome studies reported a decrease in AgRP mRNA levels withgestational age and no change in POMC levels whilst othersshowed an upregulated mRNA expression of both neuropep-tides in human [24] and rat placentas [20 21] As far as we areaware the mRNA expression of CART has been previouslyexamined neither in human nor in murine placental tissuesIn this sense recent data from our group demonstrated thatthemRNA levels of this catabolic and anorectic neuropeptideare markedly increased in the rat placenta at term [20 21]However in the present study we failed to detect any changein CART gene expression throughout mouse pregnancy Thereason for this discrepancy is unclear but itmight reflect somebiological differences between species
Another novel finding of the present study is the fact thatIL-6 acting on an autocrine or paracrine manner modulatesthe placental expression of classical ARC energy balance reg-ulatory peptides as shown by IL-6 KOmice data Thus NPYand CART mRNA levels are upregulated by IL-6 deficiencyat late pregnancy while POMC gene expression is downregu-lated in placentas from 15-day pregnant IL-6 KOmice Theseresults indicate that during the third week of pregnancylack of IL-6 settles an anabolic milieu in the mouse placentaby reciprocally modulating the transcriptional expression ofneuropeptides known to either promote NPY or repressPOMC net energy gain As we have previously demonstratedin the hypothalamus of the mouse in late pregnancy a lackof IL-6 upregulates NPYAgRP mRNA levels in the ARCit seems that IL-6 deficiency during late pregnancy exertsa stimulatory effect on NPY gene transcription whatever isits source However the results obtained regarding placentalCARTPOMC gene expression were somewhat unexpected
Increased hypothalamic CART or POMC production hasbeen reported after endotoxemic challenge or central admin-istration of other well-known proinflammatory cytokinesas IL-1 [59 60] In addition exercise-induced hypothala-mic IL-6 and IL-10 in obese rats [61] and IL-6 astrocyte-targeted overexpression in female mice have been shownto increase POMC expression in the ARC [62] Converselytotal knockout of IL-6 in mice restored to virgin values thereduced POMC mRNA levels in the ARC of late pregnantmice probably reflecting a compensatorymechanism exertedby other factors [29] On the contrary the data reportedherein indicate that during the second half of pregnancy thiscompensatory effect on POMC gene expression is not settledin the mouse placenta Considering the above mentionedresults and the fact that CART and POMC are colocalized inthe ARC a similar pattern of placental expressionmight havebeen expected for both neuropeptides However we havenot performed in situ hybridization or immunohistochemicalstudies to characterize cellular distribution of CART andPOMC in placental tissue Therefore we cannot exclude thepossibility that different placental cell types could producethese neuropeptides and respond to absence of IL-6 in adifferent manner In fact with respect to their hypothalamicregulation it is worth to mention that CART mRNA hasbeen reported in a number of hypothalamic cell populationsbesides the ARC where its expression has been shown to bedifferentially modulated from that of POMC in response toendotoxin challenge [63]
Finally in relation to the previous point it is importantto take into account that the experimental approach used inthe current study was designed to assess during the secondhalf of pregnancy global changes in IL-6 and main energyregulating neuropeptides gene expression in the mouse pla-centa Additionally the IL-6 deficient mouse was chosen toaddress the influence of total absence of endogenous IL-6 onthis setting However the murine placenta consists of threemayor cell layers of maternal or placental origin the outermaternal decidua the middle junctional zone composedof fetoplacental trophoblast cells and the inner labyrinthzone [64] Therefore further in vitro studies involving eitherdecidual or trophoblast cells in culture would be helpful todetermine the ability of each cell population to produce andsecrete these neuropeptides and confirm the contribution ofIL-6 to their transcriptional modulation
Whatever is the case these results of the current studysuggest that IL-6 by exerting a direct or indirect regulatoryaction in the transcriptional control of NPY CART andPOMC expression may play a role in the homeostaticresponse to energy availability in mouse placenta Finallywhether placental neuropeptide protein levels correlate withthe ontogenetic neuropeptide mRNA pattern herein shownwill merit further investigation
5 Conclusions
In summary we demonstrate that as in other species theclassical hypothalamic neuropeptidergic systems involvedin energy homeostasis are present in the mouse placenta
8 International Journal of Endocrinology
These central signals display a specific ontogenetic expressionpattern in absence of IL-6 with major changes settled inplacentas at term Thus our results suggest that IL-6 a well-known immune and metabolic modulating factor mightmodulate their transcriptional expression by acting on anautocrine or paracrine manner Collectively these resultssuggest that during an energetic challenging condition suchas pregnancy IL-6 may also extend its action as a metabolicsignal to the main organ at the maternal-fetal interface theplacenta
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank Eva Vigo C Ruth Gonzalez and SiaraPerez for excellent technical assistance with the qPCRassays The HPRT gene primers for SYBR Green geneexpression analysis were kindly donated by professor ClaraV Alvarez Research Center of Molecular Medicine andChronicDiseases (CIMUS)This work has been supported byEuropean Community (FP72007n∘ 245009 ldquoNeuroFASTrdquo)Ministerio de Economıa y Competitividad (PP and MCGGBFU2007-62683BFI and PP MCGG and CD CIBERobn(CB0603)) and Xunta de Galicia Grants (MCGG and LLPGIDIT06PXIB208067PR) CIBER de Fisiopatologıa de laObesidad y Nutricion is an initiative of ISCIII
References
[1] J R Speakman D A Levitsky D B Allison et al ldquoSet pointssettling points and some alternative models theoretical optionsto understandhowgenes and environments combine to regulatebody adiposityrdquoDisease Models ampMechanisms vol 4 no 6 pp733ndash745 2011
[2] K W Williams and J K Elmquist ldquoFrom neuroanatomy tobehavior central integration of peripheral signals regulatingfeeding behaviorrdquoNature Neuroscience vol 15 no 10 pp 1350ndash1355 2012
[3] J E Schneider C M Klingerman and A Abdulhay ldquoSenseand nonsense in metabolic control of reproductionrdquo Frontiersin Endocrinology vol 3 article 26 2012
[4] D Richard B Monge-Roffarello K Chechi S M Labbe andE E Turcotte ldquoControl and physiological determinants ofsympatheticallymediated brown adipose tissue thermogenesisrdquoFrontiers in Endocrinology vol 3 article 36 2012
[5] MGMyers Jr andD POlson ldquoCentral nervous system controlof metabolismrdquo Nature vol 491 no 7424 pp 357ndash363 2012
[6] M Lopez S Tovar M J Vazquez L M Williams and CDieguez ldquoPeripheral tissue-brain interactions in the regulationof food intakerdquo Proceedings of the Nutrition Society vol 66 no1 pp 131ndash155 2007
[7] S R Ladyman R A Augustine and D R Grattan ldquoHormoneinteractions regulating energy balance during pregnancyrdquo Jour-nal of Neuroendocrinology vol 22 no 7 pp 805ndash817 2010
[8] K Sinchak and E J Wagner ldquoEstradiol signaling in theregulation of reproduction and energy balancerdquo Frontiers inEndocrinology vol 33 no 4 pp 342ndash363 2012
[9] A Crown D K Clifton and R A Steiner ldquoNeuropeptidesignaling in the integration of metabolism and reproductionrdquoNeuroendocrinology vol 86 no 3 pp 175ndash182 2007
[10] J W Hill J K Elmquist and C F Elias ldquoHypothalamicpathways linking energy balance and reproductionrdquo AmericanJournal of Physiology vol 294 no 5 pp E827ndashE832 2008
[11] S Kumar and G Kaur ldquoIntermittent fasting dietary restrictionregimen negatively influences reproduction in young rats astudy of hypothalamo-hypophysial-gonadal axisrdquo PLoS ONEvol 8 no 1 Article ID e52416 2013
[12] R Senaris T Garcia-Caballero X Casabiell et al ldquoSynthesis ofleptin in human placentardquo Endocrinology vol 138 no 10 pp4501ndash4504 1997
[13] N Hoogard L Hunter J S Duncan L M Williams PTrayhurn and J G Mercer ldquoLeptin and leptin receptor mRNAand protein expression in the murine fetus and placentardquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 20 pp 11073ndash11078 1997
[14] J E Caminos R Nogueiras R Gallego et al ldquoExpressionand regulation of adiponectin and receptor in human and ratplacentardquo The Journal of Clinical Endocrinology amp Metabolismvol 90 no 7 pp 4276ndash4286 2005
[15] H N Jones T Jansson and T L Powell ldquoFull-lengthadiponectin attenuates insulin signaling and inhibits insulin-stimulated amino acid transport in human primary trophoblastcellsrdquo Diabetes vol 59 no 5 pp 1161ndash1170 2010
[16] N di Simone F di Nicuolo D Marzioni et al ldquoResistinmodulates glucose uptake and glucose transporter-1 (GLUT-1) expression in trophoblast cellsrdquo Journal of Cellular andMolecular Medicine vol 13 no 2 pp 388ndash397 2009
[17] S Yura N Sagawa H Itoh et al ldquoResistin is expressed inthe human placentardquo The Journal of Clinical Endocrinology ampMetabolism vol 88 no 3 pp 1394ndash1397 2003
[18] N Noyola-Martıneza L Dıaz E Avila A Halhali F Larreaand D Barrera ldquoCalcitriol downregulates TNF-120572 and IL-6 expression in cultured placental cells from preeclampticwomenrdquo Cytokine vol 61 no 1 pp 245ndash250 2013
[19] S K Munro M D Mitchell and A P Ponnampalam ldquoHistonedeacetylase inhibition by trichostatin A mitigates LPS inducedTNF120572 and IL-10 production in human placental explantsrdquoPlacenta vol 34 no 7 pp 567ndash573 2013
[20] R Beloosesky D A Gayle F Amidi S N Ahanya M Desaiand M G Ross ldquoOntogenic expression of putative feedingpeptides in the rat fetal brain and placentardquo Nutritional Neu-roscience vol 9 no 1-2 pp 33ndash40 2006
[21] J E Caminos S B Bravo C R Gonzalez et al ldquoFood intakeregulating-neuropeptides are expressed and regulated throughpregnancy and following food restriction in rat placentardquoReproductive Biology and Endocrinology vol 6 article 14 2008
[22] S I Grigorakis E Anastasiou K Dai A Souvatzoglou andM Alevizaki ldquoThree mRNA transcripts of the proopiome-lanocortin gene in human placenta at termrdquo European Journalof Endocrinology vol 142 no 5 pp 533ndash536 2000
[23] J Robidoux L Simoneau A Masse and J Lafond ldquoActivationof L-type calcium channels induces corticotropin-releasing fac-tor secretion from human placental trophoblastsrdquo The Journalof Clinical Endocrinology ampMetabolism vol 85 no 9 pp 3356ndash3364 2000
International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003
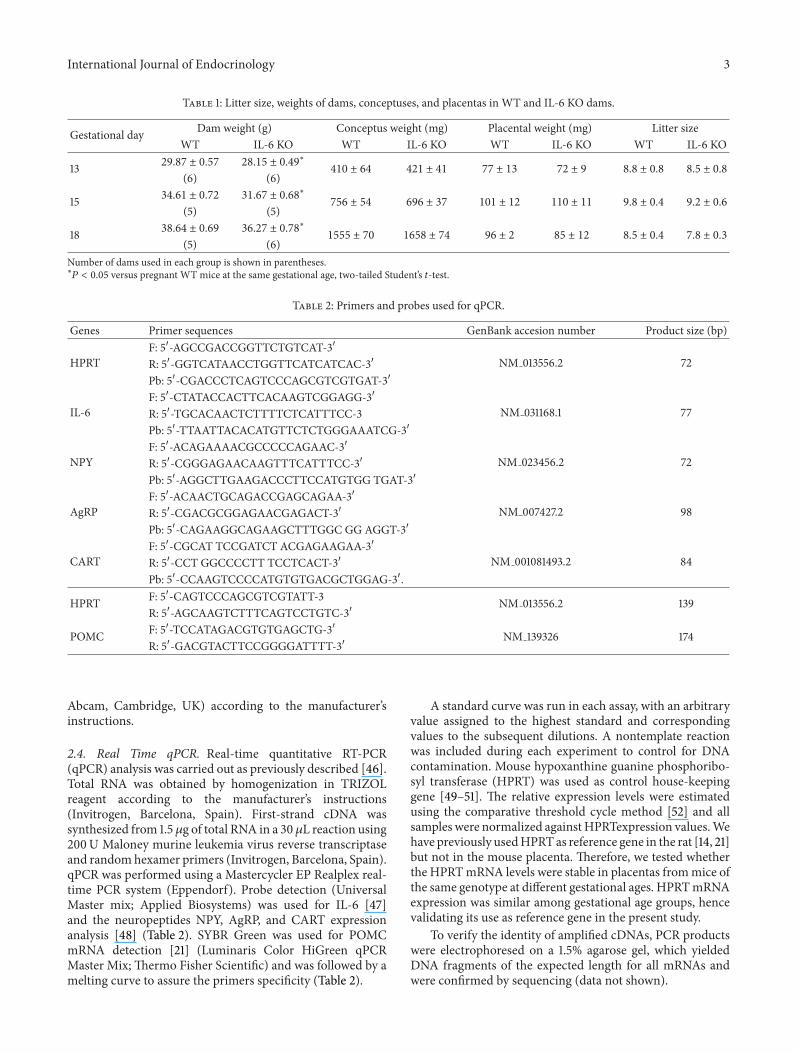
International Journal of Endocrinology 3
Table 1 Litter size weights of dams conceptuses and placentas in WT and IL-6 KO dams
Gestational day Dam weight (g) Conceptus weight (mg) Placental weight (mg) Litter sizeWT IL-6 KO WT IL-6 KO WT IL-6 KO WT IL-6 KO
13 2987 plusmn 057 2815 plusmn 049lowast
410 plusmn 64 421 plusmn 41 77 plusmn 13 72 plusmn 9 88 plusmn 08 85 plusmn 08
(6) (6)
15 3461 plusmn 072 3167 plusmn 068lowast
756 plusmn 54 696 plusmn 37 101 plusmn 12 110 plusmn 11 98 plusmn 04 92 plusmn 06
(5) (5)
18 3864 plusmn 069 3627 plusmn 078lowast
1555 plusmn 70 1658 plusmn 74 96 plusmn 2 85 plusmn 12 85 plusmn 04 78 plusmn 03
(5) (6)Number of dams used in each group is shown in parentheseslowast
119875 lt 005 versus pregnant WT mice at the same gestational age two-tailed Studentrsquos 119905-test
Table 2 Primers and probes used for qPCR
Genes Primer sequences GenBank accesion number Product size (bp)
HPRTF 51015840-AGCCGACCGGTTCTGTCAT-31015840
NM 0135562 72R 51015840-GGTCATAACCTGGTTCATCATCAC-31015840
Pb 51015840-CGACCCTCAGTCCCAGCGTCGTGAT-31015840
IL-6F 51015840-CTATACCACTTCACAAGTCGGAGG-31015840
NM 0311681 77R 51015840-TGCACAACTCTTTTCTCATTTCC-3Pb 51015840-TTAATTACACATGTTCTCTGGGAAATCG-31015840
NPYF 51015840-ACAGAAAACGCCCCCAGAAC-31015840
NM 0234562 72R 51015840-CGGGAGAACAAGTTTCATTTCC-31015840
Pb 51015840-AGGCTTGAAGACCCTTCCATGTGG TGAT-31015840
AgRPF 51015840-ACAACTGCAGACCGAGCAGAA-31015840
NM 0074272 98R 51015840-CGACGCGGAGAACGAGACT-31015840
Pb 51015840-CAGAAGGCAGAAGCTTTGGC GG AGGT-31015840
CARTF 51015840-CGCAT TCCGATCT ACGAGAAGAA-31015840
NM 0010814932 84R 51015840-CCT GGCCCCTT TCCTCACT-31015840
Pb 51015840-CCAAGTCCCCATGTGTGACGCTGGAG-31015840
HPRT F 51015840-CAGTCCCAGCGTCGTATT-3 NM 0135562 139R 51015840-AGCAAGTCTTTCAGTCCTGTC-31015840
POMC F 51015840-TCCATAGACGTGTGAGCTG-31015840 NM 139326 174R 51015840-GACGTACTTCCGGGGATTTT-31015840
Abcam Cambridge UK) according to the manufacturerrsquosinstructions
24 Real Time qPCR Real-time quantitative RT-PCR(qPCR) analysis was carried out as previously described [46]Total RNA was obtained by homogenization in TRIZOLreagent according to the manufacturerrsquos instructions(Invitrogen Barcelona Spain) First-strand cDNA wassynthesized from 15 120583g of total RNA in a 30 120583L reaction using200U Maloney murine leukemia virus reverse transcriptaseand randomhexamer primers (Invitrogen Barcelona Spain)qPCR was performed using a Mastercycler EP Realplex real-time PCR system (Eppendorf) Probe detection (UniversalMaster mix Applied Biosystems) was used for IL-6 [47]and the neuropeptides NPY AgRP and CART expressionanalysis [48] (Table 2) SYBR Green was used for POMCmRNA detection [21] (Luminaris Color HiGreen qPCRMaster Mix Thermo Fisher Scientific) and was followed by amelting curve to assure the primers specificity (Table 2)
A standard curve was run in each assay with an arbitraryvalue assigned to the highest standard and correspondingvalues to the subsequent dilutions A nontemplate reactionwas included during each experiment to control for DNAcontamination Mouse hypoxanthine guanine phosphoribo-syl transferase (HPRT) was used as control house-keepinggene [49ndash51] The relative expression levels were estimatedusing the comparative threshold cycle method [52] and allsamples were normalized againstHPRTexpression valuesWehave previously usedHPRTas reference gene in the rat [14 21]but not in the mouse placenta Therefore we tested whetherthe HPRTmRNA levels were stable in placentas frommice ofthe same genotype at different gestational ages HPRTmRNAexpression was similar among gestational age groups hencevalidating its use as reference gene in the present study
To verify the identity of amplified cDNAs PCR productswere electrophoresed on a 15 agarose gel which yieldedDNA fragments of the expected length for all mRNAs andwere confirmed by sequencing (data not shown)
4 International Journal of Endocrinology
300
250
200
150
100
50
0
13 15 18
Seru
m IL
-6(p
gm
L)
Gestational day
lowast
WT
Figure 1 IL-6 levels in serum of WT (C57BL6) mice 12-week-oldWT mice were time-pregnant and serum IL-6 levels at differentpregnancy stages (13 15 and 18 days) were assessed by ELISA 119899 =5-6 Each value represents the mean plusmn SEM One-way ANOVAlowast
119875 lt 005 versus 13 days pregnant
25 Statistical Analysis Results are given as mean plusmn SEMAll data were analyzed using Graph Pad Prism version 600for Windows (GraphPad Software La Jolla California USA)Comparisons between two groups were performed withunpaired two-tailed Studentrsquos 119905-test and one-way analysis ofvariance (ANOVA) followed by the Bonferroni post hoctest when differences between more than two experimentalgroups were analyzed
3 Results
31 Increased Serum IL-6 Levels during Pregnancy in WTMice Firstly circulating levels of IL-6 were assessed duringpregnancy As expected [29 53] serum IL-6 levels increasedas pregnancy progressed although a significant stimulatoryeffect was only evident at gestational day 18 from values atgestational day 13 (119875 lt 005) (Figure 1)
32 IL-6 mRNA Expression Is Up-Regulated in the Placentaof Pregnant WT Mice Throughout Pregnancy Thereafter weanalyzed the transcriptional profile of placental IL-6 expres-sion during mouse pregnancy IL-6 mRNA was detected inall placentas tested from pregnantWT dams (C57BL6 femalemice) at gestational days 13 15 and 18 Our data showed thatIL-6 gene expression increased with gestational age in themouse placenta with significantly higher mRNA levels ondays 15 and 18 of pregnancy in comparison to values on day13 (119875 lt 001 and 119875 lt 0001 resp) (Figure 2)
33 NPYmRNAExpression Is Up-Regulated by IL-6Deficiencyin the Mouse Placenta Next we evaluated the placental NPYmRNA levels and whether absence of IL-6 might influenceits gestational profile during mouse pregnancy Althoughin the placenta of WT mice there was a trend towards an
0
5
4
3
2
1
lowastlowastlowast
lowastlowast
IL-6
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
Gestational day
WT
Figure 2 IL-6 mRNA expression in placenta of WT (C57BL6)mice 12-week-old WT mice were time-pregnant and placental IL-6 mRNA levels at different pregnancy stages (13 15 and 18 days)were assessed by real-time qPCR 119899 = 5-6 Relative mRNA levelswere normalized to 13 days pregnant mice values as 1 Each valuerepresents the mean plusmn SEM One-way ANOVA lowastlowast119875 lt 001 versus13 days pregnant mice values
increased NPY mRNA expression at gestational day 15 inrelation to the other stages of pregnancy studied no statisticaldifference was observed (Figure 3(a)) On the contrary inabsence of IL-6 placental NPY mRNA expression remainedunchanged between 13 and 15 days of pregnancy reaching thehighest values at the end of the gestational period (119875 lt 001)(Figure 3(b))
34 Lack of IL-6 Does Not Affect AgRP mRNA Expressionin the Mouse Placenta AgRP mRNA was expressed in themouse placenta at all gestational ages assessed but showeda slightly different profile during pregnancy from that ofNPYThus AgRP mRNA content in the placenta of pregnantWT mice did not change significantly from 13 to 18 days ofpregnancy (Figure 4(a)) being unaffected by IL-6 deficiency(Figure 4(b))
35 POMC mRNA Expression Is Down-Regulated by IL-6Deficiency in the Mouse Placenta Placental POMC mRNAexpression in WT mice was markedly reduced at the latestpregnancy stage (gestational day 18 119875 lt 005) whileit remained unchanged from 13 to 15 days of pregnancy(Figure 5(a)) However IL-6 KO mice showed a significantly(119875 lt 005) reducing trend for placental POMC mRNAcontent from gestational days 13 to 18 (Figure 5(b))
36 CART mRNA Expression Is Up-Regulated by IL-6 Defi-ciency in the Mouse Placenta Finally we studied the tran-scriptional profile of CART expression in the placenta ofWT and IL-6 KO mice during pregnancy Our data showedthat this catabolic neuropeptide had a constitutive and fairlyconstant expression in the placenta of WT mice throughout
International Journal of Endocrinology 5
0
Gestational day
WT
30
25
20
15
10
05
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
(a)
0
Gestational day
IL-6 KO
lowastlowast
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
6
5
4
3
2
1
13 15 18
(b)
Figure 3 NPYmRNA expression in placenta ofWT and IL-6 KOmice 12-week-oldWT and IL-6 KOmice were time-pregnant and placentalNPYmRNA levels in (a)WT and (b) IL-6 KOmice at different pregnancy stages (13 15 and 18 days) were assessed by real-time qPCR 119899 = 5-6Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents the mean plusmn SEMOne-way ANOVA lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(b)
Figure 4 AgRP mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental AgRP mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM
pregnancy (Figure 6(a)) A similar pattern was observed inthe placentas from IL-6 KO mice until gestational day 18when the highest levels of placental CARTmRNA expressionwere reached (119875 lt 001) (Figure 6(b))
4 Discussion
Pregnancy is now recognized as a state of adaptive low-grade-inflammation induced by a progressive increase ofcirculating levels inflammatory mediators such as cytokines
chemokines and acute-phase reactants [53 54] Cytokinescentral players of this inflammatory response are secreted bythe immune cells and adipocytes but also by placental cells[55] Placental cytokine production and action are crucialto control trophoblast implantation embryo developmentand feto-maternal tolerance [55] However once pregnancyis established the physiological relevance of most placentalcytokines remains unclear
Proinflammatory cytokines such as IL-6 TNF alphaand IL-1 are known to negatively influence peripheral lipid
6 International Journal of Endocrinology
Gestational day
WT
0
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)20
15
10
05lowast
13 15 18
(a)
0
Gestational day
IL-6 KO
20
15
10
05
lowast
lowastlowast
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)
13 15 18
(b)
Figure 5 POMC mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental POMC mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
CART
mRN
A le
vels
(rel
ativ
e val
ues)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
30
25
20
15
10
05
CART
mRN
A le
vels
(rel
ativ
e val
ues) lowast
13 15 18
(b)
Figure 6 CART mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental CART mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (1315 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 versus 13 days pregnant mice of respective genotype
and glucose metabolism Hence by secreting these immunefactors the placenta may enhance the contribution madeby adipose tissue to induce maternal insulin resistance andensure a proper glucose transfer to the growing fetus In thisstudy we found increased expression of IL-6 mRNA in themouse placenta during gestation These results are in agree-ment with previous reports of an enhanced IL-6 placentalproduction during the latest stages of rat [54] and humanpregnancies [56] To our knowledge the present work is thefirst one to determine the ontogenetic profile of IL-6 mRNA
levels in the mouse placenta In fact De and coworkers [57]showed high levels of placental IL-6 bioactivity but failed todemonstrate its constitutive transcriptional expression in thismouse reproductive organ The reason for this discrepancymay be the different methodologies employed since in thisprevious study IL-6mRNA levelsweremeasured byNorthernblot while we used a more sensitive technique RT-qPCRWe[29] and others [53 58] have previously found elevated IL-6 serum and adipose tissue mRNA levels during mouse latepregnancy Our data do not allow us to determine the relative
International Journal of Endocrinology 7
contribution of both tissues to the amount of this cytokinepresent in maternal circulation but the similar pattern of IL-6 mRNA overexpression in both tissues suggests that bothcould be a source of circulating IL-6 in pregnancy
Our group has recently shown that in addition toits peripheral metabolic effects IL-6 modulates the cen-tral mechanisms associated to maternal hyperphagia andincreased adiposity in the pregnantmouse [29] Although thetranscriptional expression of its hypothalamic neuropeptidetargets NPY AgRP POMC and CART in the human [2425] and rat placenta [20] is well established whether theyare also placentally produced and regulated by IL-6 duringmouse pregnancy remains unknown In order to clarify thisissue we determined the placental mRNA expression ofthese energy balance regulating neuropeptides throughoutpregnancy (from gestational days 13 to 18) in WT and IL-6deficient mice
The results of the present study revealed a similar tran-scriptional regulation of ARC neuropeptides in the mouseplacenta from that previously reported in the hypothalamus[29] Thus POMC mRNA levels were reduced from gesta-tional days 13 to 18 while the expression of the orexigenicand anabolic neuropeptides NPYAgRP remains unchangedA similar absence of relation between gestational age andplacental NPY mRNA content has been shown in otherspecies [20 25] On the contrary the placental ontogeneticexpression of the melanocortin system members POMC andAgRP seems to be a rather controversial issue Thereforesome studies reported a decrease in AgRP mRNA levels withgestational age and no change in POMC levels whilst othersshowed an upregulated mRNA expression of both neuropep-tides in human [24] and rat placentas [20 21] As far as we areaware the mRNA expression of CART has been previouslyexamined neither in human nor in murine placental tissuesIn this sense recent data from our group demonstrated thatthemRNA levels of this catabolic and anorectic neuropeptideare markedly increased in the rat placenta at term [20 21]However in the present study we failed to detect any changein CART gene expression throughout mouse pregnancy Thereason for this discrepancy is unclear but itmight reflect somebiological differences between species
Another novel finding of the present study is the fact thatIL-6 acting on an autocrine or paracrine manner modulatesthe placental expression of classical ARC energy balance reg-ulatory peptides as shown by IL-6 KOmice data Thus NPYand CART mRNA levels are upregulated by IL-6 deficiencyat late pregnancy while POMC gene expression is downregu-lated in placentas from 15-day pregnant IL-6 KOmice Theseresults indicate that during the third week of pregnancylack of IL-6 settles an anabolic milieu in the mouse placentaby reciprocally modulating the transcriptional expression ofneuropeptides known to either promote NPY or repressPOMC net energy gain As we have previously demonstratedin the hypothalamus of the mouse in late pregnancy a lackof IL-6 upregulates NPYAgRP mRNA levels in the ARCit seems that IL-6 deficiency during late pregnancy exertsa stimulatory effect on NPY gene transcription whatever isits source However the results obtained regarding placentalCARTPOMC gene expression were somewhat unexpected
Increased hypothalamic CART or POMC production hasbeen reported after endotoxemic challenge or central admin-istration of other well-known proinflammatory cytokinesas IL-1 [59 60] In addition exercise-induced hypothala-mic IL-6 and IL-10 in obese rats [61] and IL-6 astrocyte-targeted overexpression in female mice have been shownto increase POMC expression in the ARC [62] Converselytotal knockout of IL-6 in mice restored to virgin values thereduced POMC mRNA levels in the ARC of late pregnantmice probably reflecting a compensatorymechanism exertedby other factors [29] On the contrary the data reportedherein indicate that during the second half of pregnancy thiscompensatory effect on POMC gene expression is not settledin the mouse placenta Considering the above mentionedresults and the fact that CART and POMC are colocalized inthe ARC a similar pattern of placental expressionmight havebeen expected for both neuropeptides However we havenot performed in situ hybridization or immunohistochemicalstudies to characterize cellular distribution of CART andPOMC in placental tissue Therefore we cannot exclude thepossibility that different placental cell types could producethese neuropeptides and respond to absence of IL-6 in adifferent manner In fact with respect to their hypothalamicregulation it is worth to mention that CART mRNA hasbeen reported in a number of hypothalamic cell populationsbesides the ARC where its expression has been shown to bedifferentially modulated from that of POMC in response toendotoxin challenge [63]
Finally in relation to the previous point it is importantto take into account that the experimental approach used inthe current study was designed to assess during the secondhalf of pregnancy global changes in IL-6 and main energyregulating neuropeptides gene expression in the mouse pla-centa Additionally the IL-6 deficient mouse was chosen toaddress the influence of total absence of endogenous IL-6 onthis setting However the murine placenta consists of threemayor cell layers of maternal or placental origin the outermaternal decidua the middle junctional zone composedof fetoplacental trophoblast cells and the inner labyrinthzone [64] Therefore further in vitro studies involving eitherdecidual or trophoblast cells in culture would be helpful todetermine the ability of each cell population to produce andsecrete these neuropeptides and confirm the contribution ofIL-6 to their transcriptional modulation
Whatever is the case these results of the current studysuggest that IL-6 by exerting a direct or indirect regulatoryaction in the transcriptional control of NPY CART andPOMC expression may play a role in the homeostaticresponse to energy availability in mouse placenta Finallywhether placental neuropeptide protein levels correlate withthe ontogenetic neuropeptide mRNA pattern herein shownwill merit further investigation
5 Conclusions
In summary we demonstrate that as in other species theclassical hypothalamic neuropeptidergic systems involvedin energy homeostasis are present in the mouse placenta
8 International Journal of Endocrinology
These central signals display a specific ontogenetic expressionpattern in absence of IL-6 with major changes settled inplacentas at term Thus our results suggest that IL-6 a well-known immune and metabolic modulating factor mightmodulate their transcriptional expression by acting on anautocrine or paracrine manner Collectively these resultssuggest that during an energetic challenging condition suchas pregnancy IL-6 may also extend its action as a metabolicsignal to the main organ at the maternal-fetal interface theplacenta
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank Eva Vigo C Ruth Gonzalez and SiaraPerez for excellent technical assistance with the qPCRassays The HPRT gene primers for SYBR Green geneexpression analysis were kindly donated by professor ClaraV Alvarez Research Center of Molecular Medicine andChronicDiseases (CIMUS)This work has been supported byEuropean Community (FP72007n∘ 245009 ldquoNeuroFASTrdquo)Ministerio de Economıa y Competitividad (PP and MCGGBFU2007-62683BFI and PP MCGG and CD CIBERobn(CB0603)) and Xunta de Galicia Grants (MCGG and LLPGIDIT06PXIB208067PR) CIBER de Fisiopatologıa de laObesidad y Nutricion is an initiative of ISCIII
References
[1] J R Speakman D A Levitsky D B Allison et al ldquoSet pointssettling points and some alternative models theoretical optionsto understandhowgenes and environments combine to regulatebody adiposityrdquoDisease Models ampMechanisms vol 4 no 6 pp733ndash745 2011
[2] K W Williams and J K Elmquist ldquoFrom neuroanatomy tobehavior central integration of peripheral signals regulatingfeeding behaviorrdquoNature Neuroscience vol 15 no 10 pp 1350ndash1355 2012
[3] J E Schneider C M Klingerman and A Abdulhay ldquoSenseand nonsense in metabolic control of reproductionrdquo Frontiersin Endocrinology vol 3 article 26 2012
[4] D Richard B Monge-Roffarello K Chechi S M Labbe andE E Turcotte ldquoControl and physiological determinants ofsympatheticallymediated brown adipose tissue thermogenesisrdquoFrontiers in Endocrinology vol 3 article 36 2012
[5] MGMyers Jr andD POlson ldquoCentral nervous system controlof metabolismrdquo Nature vol 491 no 7424 pp 357ndash363 2012
[6] M Lopez S Tovar M J Vazquez L M Williams and CDieguez ldquoPeripheral tissue-brain interactions in the regulationof food intakerdquo Proceedings of the Nutrition Society vol 66 no1 pp 131ndash155 2007
[7] S R Ladyman R A Augustine and D R Grattan ldquoHormoneinteractions regulating energy balance during pregnancyrdquo Jour-nal of Neuroendocrinology vol 22 no 7 pp 805ndash817 2010
[8] K Sinchak and E J Wagner ldquoEstradiol signaling in theregulation of reproduction and energy balancerdquo Frontiers inEndocrinology vol 33 no 4 pp 342ndash363 2012
[9] A Crown D K Clifton and R A Steiner ldquoNeuropeptidesignaling in the integration of metabolism and reproductionrdquoNeuroendocrinology vol 86 no 3 pp 175ndash182 2007
[10] J W Hill J K Elmquist and C F Elias ldquoHypothalamicpathways linking energy balance and reproductionrdquo AmericanJournal of Physiology vol 294 no 5 pp E827ndashE832 2008
[11] S Kumar and G Kaur ldquoIntermittent fasting dietary restrictionregimen negatively influences reproduction in young rats astudy of hypothalamo-hypophysial-gonadal axisrdquo PLoS ONEvol 8 no 1 Article ID e52416 2013
[12] R Senaris T Garcia-Caballero X Casabiell et al ldquoSynthesis ofleptin in human placentardquo Endocrinology vol 138 no 10 pp4501ndash4504 1997
[13] N Hoogard L Hunter J S Duncan L M Williams PTrayhurn and J G Mercer ldquoLeptin and leptin receptor mRNAand protein expression in the murine fetus and placentardquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 20 pp 11073ndash11078 1997
[14] J E Caminos R Nogueiras R Gallego et al ldquoExpressionand regulation of adiponectin and receptor in human and ratplacentardquo The Journal of Clinical Endocrinology amp Metabolismvol 90 no 7 pp 4276ndash4286 2005
[15] H N Jones T Jansson and T L Powell ldquoFull-lengthadiponectin attenuates insulin signaling and inhibits insulin-stimulated amino acid transport in human primary trophoblastcellsrdquo Diabetes vol 59 no 5 pp 1161ndash1170 2010
[16] N di Simone F di Nicuolo D Marzioni et al ldquoResistinmodulates glucose uptake and glucose transporter-1 (GLUT-1) expression in trophoblast cellsrdquo Journal of Cellular andMolecular Medicine vol 13 no 2 pp 388ndash397 2009
[17] S Yura N Sagawa H Itoh et al ldquoResistin is expressed inthe human placentardquo The Journal of Clinical Endocrinology ampMetabolism vol 88 no 3 pp 1394ndash1397 2003
[18] N Noyola-Martıneza L Dıaz E Avila A Halhali F Larreaand D Barrera ldquoCalcitriol downregulates TNF-120572 and IL-6 expression in cultured placental cells from preeclampticwomenrdquo Cytokine vol 61 no 1 pp 245ndash250 2013
[19] S K Munro M D Mitchell and A P Ponnampalam ldquoHistonedeacetylase inhibition by trichostatin A mitigates LPS inducedTNF120572 and IL-10 production in human placental explantsrdquoPlacenta vol 34 no 7 pp 567ndash573 2013
[20] R Beloosesky D A Gayle F Amidi S N Ahanya M Desaiand M G Ross ldquoOntogenic expression of putative feedingpeptides in the rat fetal brain and placentardquo Nutritional Neu-roscience vol 9 no 1-2 pp 33ndash40 2006
[21] J E Caminos S B Bravo C R Gonzalez et al ldquoFood intakeregulating-neuropeptides are expressed and regulated throughpregnancy and following food restriction in rat placentardquoReproductive Biology and Endocrinology vol 6 article 14 2008
[22] S I Grigorakis E Anastasiou K Dai A Souvatzoglou andM Alevizaki ldquoThree mRNA transcripts of the proopiome-lanocortin gene in human placenta at termrdquo European Journalof Endocrinology vol 142 no 5 pp 533ndash536 2000
[23] J Robidoux L Simoneau A Masse and J Lafond ldquoActivationof L-type calcium channels induces corticotropin-releasing fac-tor secretion from human placental trophoblastsrdquo The Journalof Clinical Endocrinology ampMetabolism vol 85 no 9 pp 3356ndash3364 2000
International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003
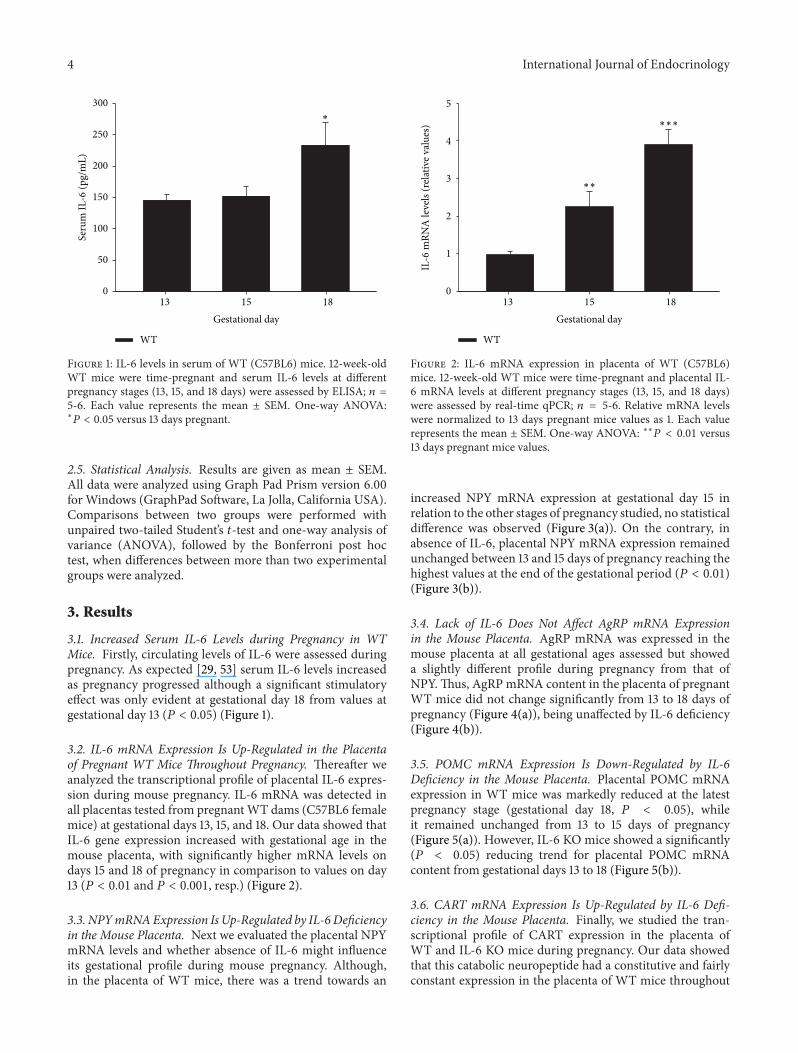
4 International Journal of Endocrinology
300
250
200
150
100
50
0
13 15 18
Seru
m IL
-6(p
gm
L)
Gestational day
lowast
WT
Figure 1 IL-6 levels in serum of WT (C57BL6) mice 12-week-oldWT mice were time-pregnant and serum IL-6 levels at differentpregnancy stages (13 15 and 18 days) were assessed by ELISA 119899 =5-6 Each value represents the mean plusmn SEM One-way ANOVAlowast
119875 lt 005 versus 13 days pregnant
25 Statistical Analysis Results are given as mean plusmn SEMAll data were analyzed using Graph Pad Prism version 600for Windows (GraphPad Software La Jolla California USA)Comparisons between two groups were performed withunpaired two-tailed Studentrsquos 119905-test and one-way analysis ofvariance (ANOVA) followed by the Bonferroni post hoctest when differences between more than two experimentalgroups were analyzed
3 Results
31 Increased Serum IL-6 Levels during Pregnancy in WTMice Firstly circulating levels of IL-6 were assessed duringpregnancy As expected [29 53] serum IL-6 levels increasedas pregnancy progressed although a significant stimulatoryeffect was only evident at gestational day 18 from values atgestational day 13 (119875 lt 005) (Figure 1)
32 IL-6 mRNA Expression Is Up-Regulated in the Placentaof Pregnant WT Mice Throughout Pregnancy Thereafter weanalyzed the transcriptional profile of placental IL-6 expres-sion during mouse pregnancy IL-6 mRNA was detected inall placentas tested from pregnantWT dams (C57BL6 femalemice) at gestational days 13 15 and 18 Our data showed thatIL-6 gene expression increased with gestational age in themouse placenta with significantly higher mRNA levels ondays 15 and 18 of pregnancy in comparison to values on day13 (119875 lt 001 and 119875 lt 0001 resp) (Figure 2)
33 NPYmRNAExpression Is Up-Regulated by IL-6Deficiencyin the Mouse Placenta Next we evaluated the placental NPYmRNA levels and whether absence of IL-6 might influenceits gestational profile during mouse pregnancy Althoughin the placenta of WT mice there was a trend towards an
0
5
4
3
2
1
lowastlowastlowast
lowastlowast
IL-6
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
Gestational day
WT
Figure 2 IL-6 mRNA expression in placenta of WT (C57BL6)mice 12-week-old WT mice were time-pregnant and placental IL-6 mRNA levels at different pregnancy stages (13 15 and 18 days)were assessed by real-time qPCR 119899 = 5-6 Relative mRNA levelswere normalized to 13 days pregnant mice values as 1 Each valuerepresents the mean plusmn SEM One-way ANOVA lowastlowast119875 lt 001 versus13 days pregnant mice values
increased NPY mRNA expression at gestational day 15 inrelation to the other stages of pregnancy studied no statisticaldifference was observed (Figure 3(a)) On the contrary inabsence of IL-6 placental NPY mRNA expression remainedunchanged between 13 and 15 days of pregnancy reaching thehighest values at the end of the gestational period (119875 lt 001)(Figure 3(b))
34 Lack of IL-6 Does Not Affect AgRP mRNA Expressionin the Mouse Placenta AgRP mRNA was expressed in themouse placenta at all gestational ages assessed but showeda slightly different profile during pregnancy from that ofNPYThus AgRP mRNA content in the placenta of pregnantWT mice did not change significantly from 13 to 18 days ofpregnancy (Figure 4(a)) being unaffected by IL-6 deficiency(Figure 4(b))
35 POMC mRNA Expression Is Down-Regulated by IL-6Deficiency in the Mouse Placenta Placental POMC mRNAexpression in WT mice was markedly reduced at the latestpregnancy stage (gestational day 18 119875 lt 005) whileit remained unchanged from 13 to 15 days of pregnancy(Figure 5(a)) However IL-6 KO mice showed a significantly(119875 lt 005) reducing trend for placental POMC mRNAcontent from gestational days 13 to 18 (Figure 5(b))
36 CART mRNA Expression Is Up-Regulated by IL-6 Defi-ciency in the Mouse Placenta Finally we studied the tran-scriptional profile of CART expression in the placenta ofWT and IL-6 KO mice during pregnancy Our data showedthat this catabolic neuropeptide had a constitutive and fairlyconstant expression in the placenta of WT mice throughout
International Journal of Endocrinology 5
0
Gestational day
WT
30
25
20
15
10
05
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
(a)
0
Gestational day
IL-6 KO
lowastlowast
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
6
5
4
3
2
1
13 15 18
(b)
Figure 3 NPYmRNA expression in placenta ofWT and IL-6 KOmice 12-week-oldWT and IL-6 KOmice were time-pregnant and placentalNPYmRNA levels in (a)WT and (b) IL-6 KOmice at different pregnancy stages (13 15 and 18 days) were assessed by real-time qPCR 119899 = 5-6Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents the mean plusmn SEMOne-way ANOVA lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(b)
Figure 4 AgRP mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental AgRP mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM
pregnancy (Figure 6(a)) A similar pattern was observed inthe placentas from IL-6 KO mice until gestational day 18when the highest levels of placental CARTmRNA expressionwere reached (119875 lt 001) (Figure 6(b))
4 Discussion
Pregnancy is now recognized as a state of adaptive low-grade-inflammation induced by a progressive increase ofcirculating levels inflammatory mediators such as cytokines
chemokines and acute-phase reactants [53 54] Cytokinescentral players of this inflammatory response are secreted bythe immune cells and adipocytes but also by placental cells[55] Placental cytokine production and action are crucialto control trophoblast implantation embryo developmentand feto-maternal tolerance [55] However once pregnancyis established the physiological relevance of most placentalcytokines remains unclear
Proinflammatory cytokines such as IL-6 TNF alphaand IL-1 are known to negatively influence peripheral lipid
6 International Journal of Endocrinology
Gestational day
WT
0
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)20
15
10
05lowast
13 15 18
(a)
0
Gestational day
IL-6 KO
20
15
10
05
lowast
lowastlowast
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)
13 15 18
(b)
Figure 5 POMC mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental POMC mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
CART
mRN
A le
vels
(rel
ativ
e val
ues)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
30
25
20
15
10
05
CART
mRN
A le
vels
(rel
ativ
e val
ues) lowast
13 15 18
(b)
Figure 6 CART mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental CART mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (1315 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 versus 13 days pregnant mice of respective genotype
and glucose metabolism Hence by secreting these immunefactors the placenta may enhance the contribution madeby adipose tissue to induce maternal insulin resistance andensure a proper glucose transfer to the growing fetus In thisstudy we found increased expression of IL-6 mRNA in themouse placenta during gestation These results are in agree-ment with previous reports of an enhanced IL-6 placentalproduction during the latest stages of rat [54] and humanpregnancies [56] To our knowledge the present work is thefirst one to determine the ontogenetic profile of IL-6 mRNA
levels in the mouse placenta In fact De and coworkers [57]showed high levels of placental IL-6 bioactivity but failed todemonstrate its constitutive transcriptional expression in thismouse reproductive organ The reason for this discrepancymay be the different methodologies employed since in thisprevious study IL-6mRNA levelsweremeasured byNorthernblot while we used a more sensitive technique RT-qPCRWe[29] and others [53 58] have previously found elevated IL-6 serum and adipose tissue mRNA levels during mouse latepregnancy Our data do not allow us to determine the relative
International Journal of Endocrinology 7
contribution of both tissues to the amount of this cytokinepresent in maternal circulation but the similar pattern of IL-6 mRNA overexpression in both tissues suggests that bothcould be a source of circulating IL-6 in pregnancy
Our group has recently shown that in addition toits peripheral metabolic effects IL-6 modulates the cen-tral mechanisms associated to maternal hyperphagia andincreased adiposity in the pregnantmouse [29] Although thetranscriptional expression of its hypothalamic neuropeptidetargets NPY AgRP POMC and CART in the human [2425] and rat placenta [20] is well established whether theyare also placentally produced and regulated by IL-6 duringmouse pregnancy remains unknown In order to clarify thisissue we determined the placental mRNA expression ofthese energy balance regulating neuropeptides throughoutpregnancy (from gestational days 13 to 18) in WT and IL-6deficient mice
The results of the present study revealed a similar tran-scriptional regulation of ARC neuropeptides in the mouseplacenta from that previously reported in the hypothalamus[29] Thus POMC mRNA levels were reduced from gesta-tional days 13 to 18 while the expression of the orexigenicand anabolic neuropeptides NPYAgRP remains unchangedA similar absence of relation between gestational age andplacental NPY mRNA content has been shown in otherspecies [20 25] On the contrary the placental ontogeneticexpression of the melanocortin system members POMC andAgRP seems to be a rather controversial issue Thereforesome studies reported a decrease in AgRP mRNA levels withgestational age and no change in POMC levels whilst othersshowed an upregulated mRNA expression of both neuropep-tides in human [24] and rat placentas [20 21] As far as we areaware the mRNA expression of CART has been previouslyexamined neither in human nor in murine placental tissuesIn this sense recent data from our group demonstrated thatthemRNA levels of this catabolic and anorectic neuropeptideare markedly increased in the rat placenta at term [20 21]However in the present study we failed to detect any changein CART gene expression throughout mouse pregnancy Thereason for this discrepancy is unclear but itmight reflect somebiological differences between species
Another novel finding of the present study is the fact thatIL-6 acting on an autocrine or paracrine manner modulatesthe placental expression of classical ARC energy balance reg-ulatory peptides as shown by IL-6 KOmice data Thus NPYand CART mRNA levels are upregulated by IL-6 deficiencyat late pregnancy while POMC gene expression is downregu-lated in placentas from 15-day pregnant IL-6 KOmice Theseresults indicate that during the third week of pregnancylack of IL-6 settles an anabolic milieu in the mouse placentaby reciprocally modulating the transcriptional expression ofneuropeptides known to either promote NPY or repressPOMC net energy gain As we have previously demonstratedin the hypothalamus of the mouse in late pregnancy a lackof IL-6 upregulates NPYAgRP mRNA levels in the ARCit seems that IL-6 deficiency during late pregnancy exertsa stimulatory effect on NPY gene transcription whatever isits source However the results obtained regarding placentalCARTPOMC gene expression were somewhat unexpected
Increased hypothalamic CART or POMC production hasbeen reported after endotoxemic challenge or central admin-istration of other well-known proinflammatory cytokinesas IL-1 [59 60] In addition exercise-induced hypothala-mic IL-6 and IL-10 in obese rats [61] and IL-6 astrocyte-targeted overexpression in female mice have been shownto increase POMC expression in the ARC [62] Converselytotal knockout of IL-6 in mice restored to virgin values thereduced POMC mRNA levels in the ARC of late pregnantmice probably reflecting a compensatorymechanism exertedby other factors [29] On the contrary the data reportedherein indicate that during the second half of pregnancy thiscompensatory effect on POMC gene expression is not settledin the mouse placenta Considering the above mentionedresults and the fact that CART and POMC are colocalized inthe ARC a similar pattern of placental expressionmight havebeen expected for both neuropeptides However we havenot performed in situ hybridization or immunohistochemicalstudies to characterize cellular distribution of CART andPOMC in placental tissue Therefore we cannot exclude thepossibility that different placental cell types could producethese neuropeptides and respond to absence of IL-6 in adifferent manner In fact with respect to their hypothalamicregulation it is worth to mention that CART mRNA hasbeen reported in a number of hypothalamic cell populationsbesides the ARC where its expression has been shown to bedifferentially modulated from that of POMC in response toendotoxin challenge [63]
Finally in relation to the previous point it is importantto take into account that the experimental approach used inthe current study was designed to assess during the secondhalf of pregnancy global changes in IL-6 and main energyregulating neuropeptides gene expression in the mouse pla-centa Additionally the IL-6 deficient mouse was chosen toaddress the influence of total absence of endogenous IL-6 onthis setting However the murine placenta consists of threemayor cell layers of maternal or placental origin the outermaternal decidua the middle junctional zone composedof fetoplacental trophoblast cells and the inner labyrinthzone [64] Therefore further in vitro studies involving eitherdecidual or trophoblast cells in culture would be helpful todetermine the ability of each cell population to produce andsecrete these neuropeptides and confirm the contribution ofIL-6 to their transcriptional modulation
Whatever is the case these results of the current studysuggest that IL-6 by exerting a direct or indirect regulatoryaction in the transcriptional control of NPY CART andPOMC expression may play a role in the homeostaticresponse to energy availability in mouse placenta Finallywhether placental neuropeptide protein levels correlate withthe ontogenetic neuropeptide mRNA pattern herein shownwill merit further investigation
5 Conclusions
In summary we demonstrate that as in other species theclassical hypothalamic neuropeptidergic systems involvedin energy homeostasis are present in the mouse placenta
8 International Journal of Endocrinology
These central signals display a specific ontogenetic expressionpattern in absence of IL-6 with major changes settled inplacentas at term Thus our results suggest that IL-6 a well-known immune and metabolic modulating factor mightmodulate their transcriptional expression by acting on anautocrine or paracrine manner Collectively these resultssuggest that during an energetic challenging condition suchas pregnancy IL-6 may also extend its action as a metabolicsignal to the main organ at the maternal-fetal interface theplacenta
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank Eva Vigo C Ruth Gonzalez and SiaraPerez for excellent technical assistance with the qPCRassays The HPRT gene primers for SYBR Green geneexpression analysis were kindly donated by professor ClaraV Alvarez Research Center of Molecular Medicine andChronicDiseases (CIMUS)This work has been supported byEuropean Community (FP72007n∘ 245009 ldquoNeuroFASTrdquo)Ministerio de Economıa y Competitividad (PP and MCGGBFU2007-62683BFI and PP MCGG and CD CIBERobn(CB0603)) and Xunta de Galicia Grants (MCGG and LLPGIDIT06PXIB208067PR) CIBER de Fisiopatologıa de laObesidad y Nutricion is an initiative of ISCIII
References
[1] J R Speakman D A Levitsky D B Allison et al ldquoSet pointssettling points and some alternative models theoretical optionsto understandhowgenes and environments combine to regulatebody adiposityrdquoDisease Models ampMechanisms vol 4 no 6 pp733ndash745 2011
[2] K W Williams and J K Elmquist ldquoFrom neuroanatomy tobehavior central integration of peripheral signals regulatingfeeding behaviorrdquoNature Neuroscience vol 15 no 10 pp 1350ndash1355 2012
[3] J E Schneider C M Klingerman and A Abdulhay ldquoSenseand nonsense in metabolic control of reproductionrdquo Frontiersin Endocrinology vol 3 article 26 2012
[4] D Richard B Monge-Roffarello K Chechi S M Labbe andE E Turcotte ldquoControl and physiological determinants ofsympatheticallymediated brown adipose tissue thermogenesisrdquoFrontiers in Endocrinology vol 3 article 36 2012
[5] MGMyers Jr andD POlson ldquoCentral nervous system controlof metabolismrdquo Nature vol 491 no 7424 pp 357ndash363 2012
[6] M Lopez S Tovar M J Vazquez L M Williams and CDieguez ldquoPeripheral tissue-brain interactions in the regulationof food intakerdquo Proceedings of the Nutrition Society vol 66 no1 pp 131ndash155 2007
[7] S R Ladyman R A Augustine and D R Grattan ldquoHormoneinteractions regulating energy balance during pregnancyrdquo Jour-nal of Neuroendocrinology vol 22 no 7 pp 805ndash817 2010
[8] K Sinchak and E J Wagner ldquoEstradiol signaling in theregulation of reproduction and energy balancerdquo Frontiers inEndocrinology vol 33 no 4 pp 342ndash363 2012
[9] A Crown D K Clifton and R A Steiner ldquoNeuropeptidesignaling in the integration of metabolism and reproductionrdquoNeuroendocrinology vol 86 no 3 pp 175ndash182 2007
[10] J W Hill J K Elmquist and C F Elias ldquoHypothalamicpathways linking energy balance and reproductionrdquo AmericanJournal of Physiology vol 294 no 5 pp E827ndashE832 2008
[11] S Kumar and G Kaur ldquoIntermittent fasting dietary restrictionregimen negatively influences reproduction in young rats astudy of hypothalamo-hypophysial-gonadal axisrdquo PLoS ONEvol 8 no 1 Article ID e52416 2013
[12] R Senaris T Garcia-Caballero X Casabiell et al ldquoSynthesis ofleptin in human placentardquo Endocrinology vol 138 no 10 pp4501ndash4504 1997
[13] N Hoogard L Hunter J S Duncan L M Williams PTrayhurn and J G Mercer ldquoLeptin and leptin receptor mRNAand protein expression in the murine fetus and placentardquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 20 pp 11073ndash11078 1997
[14] J E Caminos R Nogueiras R Gallego et al ldquoExpressionand regulation of adiponectin and receptor in human and ratplacentardquo The Journal of Clinical Endocrinology amp Metabolismvol 90 no 7 pp 4276ndash4286 2005
[15] H N Jones T Jansson and T L Powell ldquoFull-lengthadiponectin attenuates insulin signaling and inhibits insulin-stimulated amino acid transport in human primary trophoblastcellsrdquo Diabetes vol 59 no 5 pp 1161ndash1170 2010
[16] N di Simone F di Nicuolo D Marzioni et al ldquoResistinmodulates glucose uptake and glucose transporter-1 (GLUT-1) expression in trophoblast cellsrdquo Journal of Cellular andMolecular Medicine vol 13 no 2 pp 388ndash397 2009
[17] S Yura N Sagawa H Itoh et al ldquoResistin is expressed inthe human placentardquo The Journal of Clinical Endocrinology ampMetabolism vol 88 no 3 pp 1394ndash1397 2003
[18] N Noyola-Martıneza L Dıaz E Avila A Halhali F Larreaand D Barrera ldquoCalcitriol downregulates TNF-120572 and IL-6 expression in cultured placental cells from preeclampticwomenrdquo Cytokine vol 61 no 1 pp 245ndash250 2013
[19] S K Munro M D Mitchell and A P Ponnampalam ldquoHistonedeacetylase inhibition by trichostatin A mitigates LPS inducedTNF120572 and IL-10 production in human placental explantsrdquoPlacenta vol 34 no 7 pp 567ndash573 2013
[20] R Beloosesky D A Gayle F Amidi S N Ahanya M Desaiand M G Ross ldquoOntogenic expression of putative feedingpeptides in the rat fetal brain and placentardquo Nutritional Neu-roscience vol 9 no 1-2 pp 33ndash40 2006
[21] J E Caminos S B Bravo C R Gonzalez et al ldquoFood intakeregulating-neuropeptides are expressed and regulated throughpregnancy and following food restriction in rat placentardquoReproductive Biology and Endocrinology vol 6 article 14 2008
[22] S I Grigorakis E Anastasiou K Dai A Souvatzoglou andM Alevizaki ldquoThree mRNA transcripts of the proopiome-lanocortin gene in human placenta at termrdquo European Journalof Endocrinology vol 142 no 5 pp 533ndash536 2000
[23] J Robidoux L Simoneau A Masse and J Lafond ldquoActivationof L-type calcium channels induces corticotropin-releasing fac-tor secretion from human placental trophoblastsrdquo The Journalof Clinical Endocrinology ampMetabolism vol 85 no 9 pp 3356ndash3364 2000
International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003
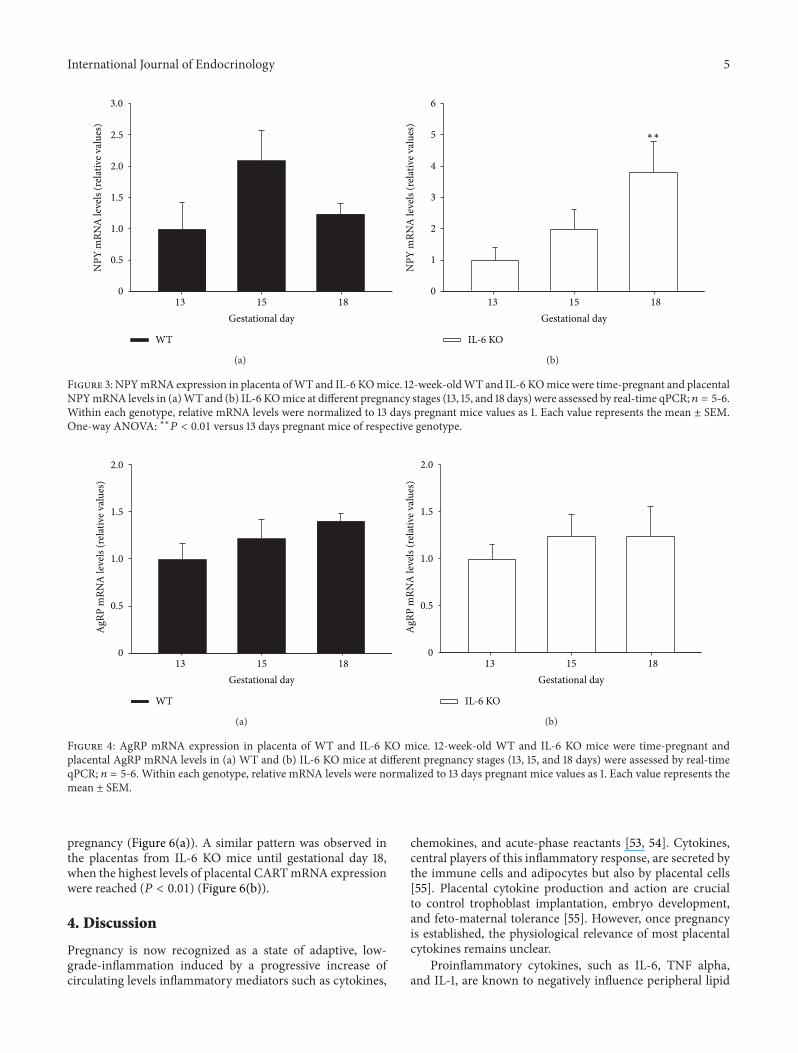
International Journal of Endocrinology 5
0
Gestational day
WT
30
25
20
15
10
05
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
13 15 18
(a)
0
Gestational day
IL-6 KO
lowastlowast
NPY
mRN
A le
vels
(rel
ativ
e val
ues)
6
5
4
3
2
1
13 15 18
(b)
Figure 3 NPYmRNA expression in placenta ofWT and IL-6 KOmice 12-week-oldWT and IL-6 KOmice were time-pregnant and placentalNPYmRNA levels in (a)WT and (b) IL-6 KOmice at different pregnancy stages (13 15 and 18 days) were assessed by real-time qPCR 119899 = 5-6Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents the mean plusmn SEMOne-way ANOVA lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
AgR
P m
RNA
leve
ls (r
elat
ive v
alue
s)
20
15
10
05
13 15 18
(b)
Figure 4 AgRP mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental AgRP mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM
pregnancy (Figure 6(a)) A similar pattern was observed inthe placentas from IL-6 KO mice until gestational day 18when the highest levels of placental CARTmRNA expressionwere reached (119875 lt 001) (Figure 6(b))
4 Discussion
Pregnancy is now recognized as a state of adaptive low-grade-inflammation induced by a progressive increase ofcirculating levels inflammatory mediators such as cytokines
chemokines and acute-phase reactants [53 54] Cytokinescentral players of this inflammatory response are secreted bythe immune cells and adipocytes but also by placental cells[55] Placental cytokine production and action are crucialto control trophoblast implantation embryo developmentand feto-maternal tolerance [55] However once pregnancyis established the physiological relevance of most placentalcytokines remains unclear
Proinflammatory cytokines such as IL-6 TNF alphaand IL-1 are known to negatively influence peripheral lipid
6 International Journal of Endocrinology
Gestational day
WT
0
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)20
15
10
05lowast
13 15 18
(a)
0
Gestational day
IL-6 KO
20
15
10
05
lowast
lowastlowast
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)
13 15 18
(b)
Figure 5 POMC mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental POMC mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
CART
mRN
A le
vels
(rel
ativ
e val
ues)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
30
25
20
15
10
05
CART
mRN
A le
vels
(rel
ativ
e val
ues) lowast
13 15 18
(b)
Figure 6 CART mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental CART mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (1315 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 versus 13 days pregnant mice of respective genotype
and glucose metabolism Hence by secreting these immunefactors the placenta may enhance the contribution madeby adipose tissue to induce maternal insulin resistance andensure a proper glucose transfer to the growing fetus In thisstudy we found increased expression of IL-6 mRNA in themouse placenta during gestation These results are in agree-ment with previous reports of an enhanced IL-6 placentalproduction during the latest stages of rat [54] and humanpregnancies [56] To our knowledge the present work is thefirst one to determine the ontogenetic profile of IL-6 mRNA
levels in the mouse placenta In fact De and coworkers [57]showed high levels of placental IL-6 bioactivity but failed todemonstrate its constitutive transcriptional expression in thismouse reproductive organ The reason for this discrepancymay be the different methodologies employed since in thisprevious study IL-6mRNA levelsweremeasured byNorthernblot while we used a more sensitive technique RT-qPCRWe[29] and others [53 58] have previously found elevated IL-6 serum and adipose tissue mRNA levels during mouse latepregnancy Our data do not allow us to determine the relative
International Journal of Endocrinology 7
contribution of both tissues to the amount of this cytokinepresent in maternal circulation but the similar pattern of IL-6 mRNA overexpression in both tissues suggests that bothcould be a source of circulating IL-6 in pregnancy
Our group has recently shown that in addition toits peripheral metabolic effects IL-6 modulates the cen-tral mechanisms associated to maternal hyperphagia andincreased adiposity in the pregnantmouse [29] Although thetranscriptional expression of its hypothalamic neuropeptidetargets NPY AgRP POMC and CART in the human [2425] and rat placenta [20] is well established whether theyare also placentally produced and regulated by IL-6 duringmouse pregnancy remains unknown In order to clarify thisissue we determined the placental mRNA expression ofthese energy balance regulating neuropeptides throughoutpregnancy (from gestational days 13 to 18) in WT and IL-6deficient mice
The results of the present study revealed a similar tran-scriptional regulation of ARC neuropeptides in the mouseplacenta from that previously reported in the hypothalamus[29] Thus POMC mRNA levels were reduced from gesta-tional days 13 to 18 while the expression of the orexigenicand anabolic neuropeptides NPYAgRP remains unchangedA similar absence of relation between gestational age andplacental NPY mRNA content has been shown in otherspecies [20 25] On the contrary the placental ontogeneticexpression of the melanocortin system members POMC andAgRP seems to be a rather controversial issue Thereforesome studies reported a decrease in AgRP mRNA levels withgestational age and no change in POMC levels whilst othersshowed an upregulated mRNA expression of both neuropep-tides in human [24] and rat placentas [20 21] As far as we areaware the mRNA expression of CART has been previouslyexamined neither in human nor in murine placental tissuesIn this sense recent data from our group demonstrated thatthemRNA levels of this catabolic and anorectic neuropeptideare markedly increased in the rat placenta at term [20 21]However in the present study we failed to detect any changein CART gene expression throughout mouse pregnancy Thereason for this discrepancy is unclear but itmight reflect somebiological differences between species
Another novel finding of the present study is the fact thatIL-6 acting on an autocrine or paracrine manner modulatesthe placental expression of classical ARC energy balance reg-ulatory peptides as shown by IL-6 KOmice data Thus NPYand CART mRNA levels are upregulated by IL-6 deficiencyat late pregnancy while POMC gene expression is downregu-lated in placentas from 15-day pregnant IL-6 KOmice Theseresults indicate that during the third week of pregnancylack of IL-6 settles an anabolic milieu in the mouse placentaby reciprocally modulating the transcriptional expression ofneuropeptides known to either promote NPY or repressPOMC net energy gain As we have previously demonstratedin the hypothalamus of the mouse in late pregnancy a lackof IL-6 upregulates NPYAgRP mRNA levels in the ARCit seems that IL-6 deficiency during late pregnancy exertsa stimulatory effect on NPY gene transcription whatever isits source However the results obtained regarding placentalCARTPOMC gene expression were somewhat unexpected
Increased hypothalamic CART or POMC production hasbeen reported after endotoxemic challenge or central admin-istration of other well-known proinflammatory cytokinesas IL-1 [59 60] In addition exercise-induced hypothala-mic IL-6 and IL-10 in obese rats [61] and IL-6 astrocyte-targeted overexpression in female mice have been shownto increase POMC expression in the ARC [62] Converselytotal knockout of IL-6 in mice restored to virgin values thereduced POMC mRNA levels in the ARC of late pregnantmice probably reflecting a compensatorymechanism exertedby other factors [29] On the contrary the data reportedherein indicate that during the second half of pregnancy thiscompensatory effect on POMC gene expression is not settledin the mouse placenta Considering the above mentionedresults and the fact that CART and POMC are colocalized inthe ARC a similar pattern of placental expressionmight havebeen expected for both neuropeptides However we havenot performed in situ hybridization or immunohistochemicalstudies to characterize cellular distribution of CART andPOMC in placental tissue Therefore we cannot exclude thepossibility that different placental cell types could producethese neuropeptides and respond to absence of IL-6 in adifferent manner In fact with respect to their hypothalamicregulation it is worth to mention that CART mRNA hasbeen reported in a number of hypothalamic cell populationsbesides the ARC where its expression has been shown to bedifferentially modulated from that of POMC in response toendotoxin challenge [63]
Finally in relation to the previous point it is importantto take into account that the experimental approach used inthe current study was designed to assess during the secondhalf of pregnancy global changes in IL-6 and main energyregulating neuropeptides gene expression in the mouse pla-centa Additionally the IL-6 deficient mouse was chosen toaddress the influence of total absence of endogenous IL-6 onthis setting However the murine placenta consists of threemayor cell layers of maternal or placental origin the outermaternal decidua the middle junctional zone composedof fetoplacental trophoblast cells and the inner labyrinthzone [64] Therefore further in vitro studies involving eitherdecidual or trophoblast cells in culture would be helpful todetermine the ability of each cell population to produce andsecrete these neuropeptides and confirm the contribution ofIL-6 to their transcriptional modulation
Whatever is the case these results of the current studysuggest that IL-6 by exerting a direct or indirect regulatoryaction in the transcriptional control of NPY CART andPOMC expression may play a role in the homeostaticresponse to energy availability in mouse placenta Finallywhether placental neuropeptide protein levels correlate withthe ontogenetic neuropeptide mRNA pattern herein shownwill merit further investigation
5 Conclusions
In summary we demonstrate that as in other species theclassical hypothalamic neuropeptidergic systems involvedin energy homeostasis are present in the mouse placenta
8 International Journal of Endocrinology
These central signals display a specific ontogenetic expressionpattern in absence of IL-6 with major changes settled inplacentas at term Thus our results suggest that IL-6 a well-known immune and metabolic modulating factor mightmodulate their transcriptional expression by acting on anautocrine or paracrine manner Collectively these resultssuggest that during an energetic challenging condition suchas pregnancy IL-6 may also extend its action as a metabolicsignal to the main organ at the maternal-fetal interface theplacenta
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank Eva Vigo C Ruth Gonzalez and SiaraPerez for excellent technical assistance with the qPCRassays The HPRT gene primers for SYBR Green geneexpression analysis were kindly donated by professor ClaraV Alvarez Research Center of Molecular Medicine andChronicDiseases (CIMUS)This work has been supported byEuropean Community (FP72007n∘ 245009 ldquoNeuroFASTrdquo)Ministerio de Economıa y Competitividad (PP and MCGGBFU2007-62683BFI and PP MCGG and CD CIBERobn(CB0603)) and Xunta de Galicia Grants (MCGG and LLPGIDIT06PXIB208067PR) CIBER de Fisiopatologıa de laObesidad y Nutricion is an initiative of ISCIII
References
[1] J R Speakman D A Levitsky D B Allison et al ldquoSet pointssettling points and some alternative models theoretical optionsto understandhowgenes and environments combine to regulatebody adiposityrdquoDisease Models ampMechanisms vol 4 no 6 pp733ndash745 2011
[2] K W Williams and J K Elmquist ldquoFrom neuroanatomy tobehavior central integration of peripheral signals regulatingfeeding behaviorrdquoNature Neuroscience vol 15 no 10 pp 1350ndash1355 2012
[3] J E Schneider C M Klingerman and A Abdulhay ldquoSenseand nonsense in metabolic control of reproductionrdquo Frontiersin Endocrinology vol 3 article 26 2012
[4] D Richard B Monge-Roffarello K Chechi S M Labbe andE E Turcotte ldquoControl and physiological determinants ofsympatheticallymediated brown adipose tissue thermogenesisrdquoFrontiers in Endocrinology vol 3 article 36 2012
[5] MGMyers Jr andD POlson ldquoCentral nervous system controlof metabolismrdquo Nature vol 491 no 7424 pp 357ndash363 2012
[6] M Lopez S Tovar M J Vazquez L M Williams and CDieguez ldquoPeripheral tissue-brain interactions in the regulationof food intakerdquo Proceedings of the Nutrition Society vol 66 no1 pp 131ndash155 2007
[7] S R Ladyman R A Augustine and D R Grattan ldquoHormoneinteractions regulating energy balance during pregnancyrdquo Jour-nal of Neuroendocrinology vol 22 no 7 pp 805ndash817 2010
[8] K Sinchak and E J Wagner ldquoEstradiol signaling in theregulation of reproduction and energy balancerdquo Frontiers inEndocrinology vol 33 no 4 pp 342ndash363 2012
[9] A Crown D K Clifton and R A Steiner ldquoNeuropeptidesignaling in the integration of metabolism and reproductionrdquoNeuroendocrinology vol 86 no 3 pp 175ndash182 2007
[10] J W Hill J K Elmquist and C F Elias ldquoHypothalamicpathways linking energy balance and reproductionrdquo AmericanJournal of Physiology vol 294 no 5 pp E827ndashE832 2008
[11] S Kumar and G Kaur ldquoIntermittent fasting dietary restrictionregimen negatively influences reproduction in young rats astudy of hypothalamo-hypophysial-gonadal axisrdquo PLoS ONEvol 8 no 1 Article ID e52416 2013
[12] R Senaris T Garcia-Caballero X Casabiell et al ldquoSynthesis ofleptin in human placentardquo Endocrinology vol 138 no 10 pp4501ndash4504 1997
[13] N Hoogard L Hunter J S Duncan L M Williams PTrayhurn and J G Mercer ldquoLeptin and leptin receptor mRNAand protein expression in the murine fetus and placentardquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 20 pp 11073ndash11078 1997
[14] J E Caminos R Nogueiras R Gallego et al ldquoExpressionand regulation of adiponectin and receptor in human and ratplacentardquo The Journal of Clinical Endocrinology amp Metabolismvol 90 no 7 pp 4276ndash4286 2005
[15] H N Jones T Jansson and T L Powell ldquoFull-lengthadiponectin attenuates insulin signaling and inhibits insulin-stimulated amino acid transport in human primary trophoblastcellsrdquo Diabetes vol 59 no 5 pp 1161ndash1170 2010
[16] N di Simone F di Nicuolo D Marzioni et al ldquoResistinmodulates glucose uptake and glucose transporter-1 (GLUT-1) expression in trophoblast cellsrdquo Journal of Cellular andMolecular Medicine vol 13 no 2 pp 388ndash397 2009
[17] S Yura N Sagawa H Itoh et al ldquoResistin is expressed inthe human placentardquo The Journal of Clinical Endocrinology ampMetabolism vol 88 no 3 pp 1394ndash1397 2003
[18] N Noyola-Martıneza L Dıaz E Avila A Halhali F Larreaand D Barrera ldquoCalcitriol downregulates TNF-120572 and IL-6 expression in cultured placental cells from preeclampticwomenrdquo Cytokine vol 61 no 1 pp 245ndash250 2013
[19] S K Munro M D Mitchell and A P Ponnampalam ldquoHistonedeacetylase inhibition by trichostatin A mitigates LPS inducedTNF120572 and IL-10 production in human placental explantsrdquoPlacenta vol 34 no 7 pp 567ndash573 2013
[20] R Beloosesky D A Gayle F Amidi S N Ahanya M Desaiand M G Ross ldquoOntogenic expression of putative feedingpeptides in the rat fetal brain and placentardquo Nutritional Neu-roscience vol 9 no 1-2 pp 33ndash40 2006
[21] J E Caminos S B Bravo C R Gonzalez et al ldquoFood intakeregulating-neuropeptides are expressed and regulated throughpregnancy and following food restriction in rat placentardquoReproductive Biology and Endocrinology vol 6 article 14 2008
[22] S I Grigorakis E Anastasiou K Dai A Souvatzoglou andM Alevizaki ldquoThree mRNA transcripts of the proopiome-lanocortin gene in human placenta at termrdquo European Journalof Endocrinology vol 142 no 5 pp 533ndash536 2000
[23] J Robidoux L Simoneau A Masse and J Lafond ldquoActivationof L-type calcium channels induces corticotropin-releasing fac-tor secretion from human placental trophoblastsrdquo The Journalof Clinical Endocrinology ampMetabolism vol 85 no 9 pp 3356ndash3364 2000
International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003
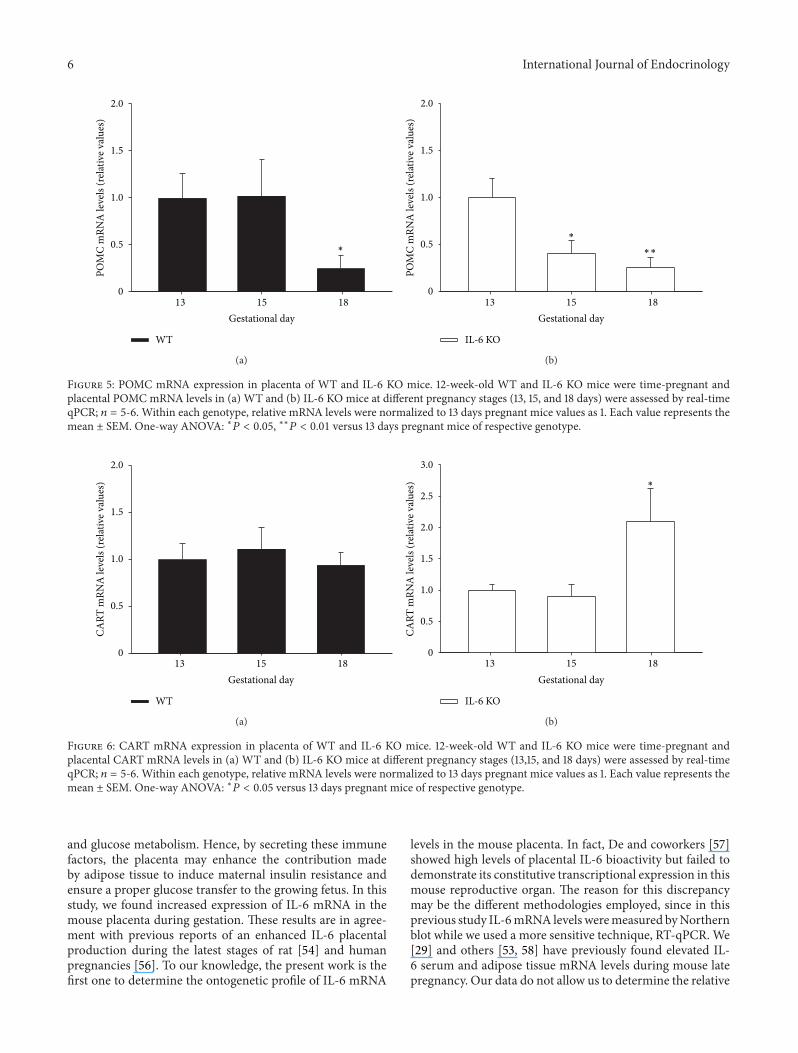
6 International Journal of Endocrinology
Gestational day
WT
0
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)20
15
10
05lowast
13 15 18
(a)
0
Gestational day
IL-6 KO
20
15
10
05
lowast
lowastlowast
POM
C m
RNA
leve
ls (r
elat
ive v
alue
s)
13 15 18
(b)
Figure 5 POMC mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental POMC mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (13 15 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 lowastlowast119875 lt 001 versus 13 days pregnant mice of respective genotype
Gestational day
WT
0
CART
mRN
A le
vels
(rel
ativ
e val
ues)
20
15
10
05
13 15 18
(a)
0
Gestational day
IL-6 KO
30
25
20
15
10
05
CART
mRN
A le
vels
(rel
ativ
e val
ues) lowast
13 15 18
(b)
Figure 6 CART mRNA expression in placenta of WT and IL-6 KO mice 12-week-old WT and IL-6 KO mice were time-pregnant andplacental CART mRNA levels in (a) WT and (b) IL-6 KO mice at different pregnancy stages (1315 and 18 days) were assessed by real-timeqPCR 119899 = 5-6 Within each genotype relative mRNA levels were normalized to 13 days pregnant mice values as 1 Each value represents themean plusmn SEM One-way ANOVA lowast119875 lt 005 versus 13 days pregnant mice of respective genotype
and glucose metabolism Hence by secreting these immunefactors the placenta may enhance the contribution madeby adipose tissue to induce maternal insulin resistance andensure a proper glucose transfer to the growing fetus In thisstudy we found increased expression of IL-6 mRNA in themouse placenta during gestation These results are in agree-ment with previous reports of an enhanced IL-6 placentalproduction during the latest stages of rat [54] and humanpregnancies [56] To our knowledge the present work is thefirst one to determine the ontogenetic profile of IL-6 mRNA
levels in the mouse placenta In fact De and coworkers [57]showed high levels of placental IL-6 bioactivity but failed todemonstrate its constitutive transcriptional expression in thismouse reproductive organ The reason for this discrepancymay be the different methodologies employed since in thisprevious study IL-6mRNA levelsweremeasured byNorthernblot while we used a more sensitive technique RT-qPCRWe[29] and others [53 58] have previously found elevated IL-6 serum and adipose tissue mRNA levels during mouse latepregnancy Our data do not allow us to determine the relative
International Journal of Endocrinology 7
contribution of both tissues to the amount of this cytokinepresent in maternal circulation but the similar pattern of IL-6 mRNA overexpression in both tissues suggests that bothcould be a source of circulating IL-6 in pregnancy
Our group has recently shown that in addition toits peripheral metabolic effects IL-6 modulates the cen-tral mechanisms associated to maternal hyperphagia andincreased adiposity in the pregnantmouse [29] Although thetranscriptional expression of its hypothalamic neuropeptidetargets NPY AgRP POMC and CART in the human [2425] and rat placenta [20] is well established whether theyare also placentally produced and regulated by IL-6 duringmouse pregnancy remains unknown In order to clarify thisissue we determined the placental mRNA expression ofthese energy balance regulating neuropeptides throughoutpregnancy (from gestational days 13 to 18) in WT and IL-6deficient mice
The results of the present study revealed a similar tran-scriptional regulation of ARC neuropeptides in the mouseplacenta from that previously reported in the hypothalamus[29] Thus POMC mRNA levels were reduced from gesta-tional days 13 to 18 while the expression of the orexigenicand anabolic neuropeptides NPYAgRP remains unchangedA similar absence of relation between gestational age andplacental NPY mRNA content has been shown in otherspecies [20 25] On the contrary the placental ontogeneticexpression of the melanocortin system members POMC andAgRP seems to be a rather controversial issue Thereforesome studies reported a decrease in AgRP mRNA levels withgestational age and no change in POMC levels whilst othersshowed an upregulated mRNA expression of both neuropep-tides in human [24] and rat placentas [20 21] As far as we areaware the mRNA expression of CART has been previouslyexamined neither in human nor in murine placental tissuesIn this sense recent data from our group demonstrated thatthemRNA levels of this catabolic and anorectic neuropeptideare markedly increased in the rat placenta at term [20 21]However in the present study we failed to detect any changein CART gene expression throughout mouse pregnancy Thereason for this discrepancy is unclear but itmight reflect somebiological differences between species
Another novel finding of the present study is the fact thatIL-6 acting on an autocrine or paracrine manner modulatesthe placental expression of classical ARC energy balance reg-ulatory peptides as shown by IL-6 KOmice data Thus NPYand CART mRNA levels are upregulated by IL-6 deficiencyat late pregnancy while POMC gene expression is downregu-lated in placentas from 15-day pregnant IL-6 KOmice Theseresults indicate that during the third week of pregnancylack of IL-6 settles an anabolic milieu in the mouse placentaby reciprocally modulating the transcriptional expression ofneuropeptides known to either promote NPY or repressPOMC net energy gain As we have previously demonstratedin the hypothalamus of the mouse in late pregnancy a lackof IL-6 upregulates NPYAgRP mRNA levels in the ARCit seems that IL-6 deficiency during late pregnancy exertsa stimulatory effect on NPY gene transcription whatever isits source However the results obtained regarding placentalCARTPOMC gene expression were somewhat unexpected
Increased hypothalamic CART or POMC production hasbeen reported after endotoxemic challenge or central admin-istration of other well-known proinflammatory cytokinesas IL-1 [59 60] In addition exercise-induced hypothala-mic IL-6 and IL-10 in obese rats [61] and IL-6 astrocyte-targeted overexpression in female mice have been shownto increase POMC expression in the ARC [62] Converselytotal knockout of IL-6 in mice restored to virgin values thereduced POMC mRNA levels in the ARC of late pregnantmice probably reflecting a compensatorymechanism exertedby other factors [29] On the contrary the data reportedherein indicate that during the second half of pregnancy thiscompensatory effect on POMC gene expression is not settledin the mouse placenta Considering the above mentionedresults and the fact that CART and POMC are colocalized inthe ARC a similar pattern of placental expressionmight havebeen expected for both neuropeptides However we havenot performed in situ hybridization or immunohistochemicalstudies to characterize cellular distribution of CART andPOMC in placental tissue Therefore we cannot exclude thepossibility that different placental cell types could producethese neuropeptides and respond to absence of IL-6 in adifferent manner In fact with respect to their hypothalamicregulation it is worth to mention that CART mRNA hasbeen reported in a number of hypothalamic cell populationsbesides the ARC where its expression has been shown to bedifferentially modulated from that of POMC in response toendotoxin challenge [63]
Finally in relation to the previous point it is importantto take into account that the experimental approach used inthe current study was designed to assess during the secondhalf of pregnancy global changes in IL-6 and main energyregulating neuropeptides gene expression in the mouse pla-centa Additionally the IL-6 deficient mouse was chosen toaddress the influence of total absence of endogenous IL-6 onthis setting However the murine placenta consists of threemayor cell layers of maternal or placental origin the outermaternal decidua the middle junctional zone composedof fetoplacental trophoblast cells and the inner labyrinthzone [64] Therefore further in vitro studies involving eitherdecidual or trophoblast cells in culture would be helpful todetermine the ability of each cell population to produce andsecrete these neuropeptides and confirm the contribution ofIL-6 to their transcriptional modulation
Whatever is the case these results of the current studysuggest that IL-6 by exerting a direct or indirect regulatoryaction in the transcriptional control of NPY CART andPOMC expression may play a role in the homeostaticresponse to energy availability in mouse placenta Finallywhether placental neuropeptide protein levels correlate withthe ontogenetic neuropeptide mRNA pattern herein shownwill merit further investigation
5 Conclusions
In summary we demonstrate that as in other species theclassical hypothalamic neuropeptidergic systems involvedin energy homeostasis are present in the mouse placenta
8 International Journal of Endocrinology
These central signals display a specific ontogenetic expressionpattern in absence of IL-6 with major changes settled inplacentas at term Thus our results suggest that IL-6 a well-known immune and metabolic modulating factor mightmodulate their transcriptional expression by acting on anautocrine or paracrine manner Collectively these resultssuggest that during an energetic challenging condition suchas pregnancy IL-6 may also extend its action as a metabolicsignal to the main organ at the maternal-fetal interface theplacenta
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank Eva Vigo C Ruth Gonzalez and SiaraPerez for excellent technical assistance with the qPCRassays The HPRT gene primers for SYBR Green geneexpression analysis were kindly donated by professor ClaraV Alvarez Research Center of Molecular Medicine andChronicDiseases (CIMUS)This work has been supported byEuropean Community (FP72007n∘ 245009 ldquoNeuroFASTrdquo)Ministerio de Economıa y Competitividad (PP and MCGGBFU2007-62683BFI and PP MCGG and CD CIBERobn(CB0603)) and Xunta de Galicia Grants (MCGG and LLPGIDIT06PXIB208067PR) CIBER de Fisiopatologıa de laObesidad y Nutricion is an initiative of ISCIII
References
[1] J R Speakman D A Levitsky D B Allison et al ldquoSet pointssettling points and some alternative models theoretical optionsto understandhowgenes and environments combine to regulatebody adiposityrdquoDisease Models ampMechanisms vol 4 no 6 pp733ndash745 2011
[2] K W Williams and J K Elmquist ldquoFrom neuroanatomy tobehavior central integration of peripheral signals regulatingfeeding behaviorrdquoNature Neuroscience vol 15 no 10 pp 1350ndash1355 2012
[3] J E Schneider C M Klingerman and A Abdulhay ldquoSenseand nonsense in metabolic control of reproductionrdquo Frontiersin Endocrinology vol 3 article 26 2012
[4] D Richard B Monge-Roffarello K Chechi S M Labbe andE E Turcotte ldquoControl and physiological determinants ofsympatheticallymediated brown adipose tissue thermogenesisrdquoFrontiers in Endocrinology vol 3 article 36 2012
[5] MGMyers Jr andD POlson ldquoCentral nervous system controlof metabolismrdquo Nature vol 491 no 7424 pp 357ndash363 2012
[6] M Lopez S Tovar M J Vazquez L M Williams and CDieguez ldquoPeripheral tissue-brain interactions in the regulationof food intakerdquo Proceedings of the Nutrition Society vol 66 no1 pp 131ndash155 2007
[7] S R Ladyman R A Augustine and D R Grattan ldquoHormoneinteractions regulating energy balance during pregnancyrdquo Jour-nal of Neuroendocrinology vol 22 no 7 pp 805ndash817 2010
[8] K Sinchak and E J Wagner ldquoEstradiol signaling in theregulation of reproduction and energy balancerdquo Frontiers inEndocrinology vol 33 no 4 pp 342ndash363 2012
[9] A Crown D K Clifton and R A Steiner ldquoNeuropeptidesignaling in the integration of metabolism and reproductionrdquoNeuroendocrinology vol 86 no 3 pp 175ndash182 2007
[10] J W Hill J K Elmquist and C F Elias ldquoHypothalamicpathways linking energy balance and reproductionrdquo AmericanJournal of Physiology vol 294 no 5 pp E827ndashE832 2008
[11] S Kumar and G Kaur ldquoIntermittent fasting dietary restrictionregimen negatively influences reproduction in young rats astudy of hypothalamo-hypophysial-gonadal axisrdquo PLoS ONEvol 8 no 1 Article ID e52416 2013
[12] R Senaris T Garcia-Caballero X Casabiell et al ldquoSynthesis ofleptin in human placentardquo Endocrinology vol 138 no 10 pp4501ndash4504 1997
[13] N Hoogard L Hunter J S Duncan L M Williams PTrayhurn and J G Mercer ldquoLeptin and leptin receptor mRNAand protein expression in the murine fetus and placentardquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 20 pp 11073ndash11078 1997
[14] J E Caminos R Nogueiras R Gallego et al ldquoExpressionand regulation of adiponectin and receptor in human and ratplacentardquo The Journal of Clinical Endocrinology amp Metabolismvol 90 no 7 pp 4276ndash4286 2005
[15] H N Jones T Jansson and T L Powell ldquoFull-lengthadiponectin attenuates insulin signaling and inhibits insulin-stimulated amino acid transport in human primary trophoblastcellsrdquo Diabetes vol 59 no 5 pp 1161ndash1170 2010
[16] N di Simone F di Nicuolo D Marzioni et al ldquoResistinmodulates glucose uptake and glucose transporter-1 (GLUT-1) expression in trophoblast cellsrdquo Journal of Cellular andMolecular Medicine vol 13 no 2 pp 388ndash397 2009
[17] S Yura N Sagawa H Itoh et al ldquoResistin is expressed inthe human placentardquo The Journal of Clinical Endocrinology ampMetabolism vol 88 no 3 pp 1394ndash1397 2003
[18] N Noyola-Martıneza L Dıaz E Avila A Halhali F Larreaand D Barrera ldquoCalcitriol downregulates TNF-120572 and IL-6 expression in cultured placental cells from preeclampticwomenrdquo Cytokine vol 61 no 1 pp 245ndash250 2013
[19] S K Munro M D Mitchell and A P Ponnampalam ldquoHistonedeacetylase inhibition by trichostatin A mitigates LPS inducedTNF120572 and IL-10 production in human placental explantsrdquoPlacenta vol 34 no 7 pp 567ndash573 2013
[20] R Beloosesky D A Gayle F Amidi S N Ahanya M Desaiand M G Ross ldquoOntogenic expression of putative feedingpeptides in the rat fetal brain and placentardquo Nutritional Neu-roscience vol 9 no 1-2 pp 33ndash40 2006
[21] J E Caminos S B Bravo C R Gonzalez et al ldquoFood intakeregulating-neuropeptides are expressed and regulated throughpregnancy and following food restriction in rat placentardquoReproductive Biology and Endocrinology vol 6 article 14 2008
[22] S I Grigorakis E Anastasiou K Dai A Souvatzoglou andM Alevizaki ldquoThree mRNA transcripts of the proopiome-lanocortin gene in human placenta at termrdquo European Journalof Endocrinology vol 142 no 5 pp 533ndash536 2000
[23] J Robidoux L Simoneau A Masse and J Lafond ldquoActivationof L-type calcium channels induces corticotropin-releasing fac-tor secretion from human placental trophoblastsrdquo The Journalof Clinical Endocrinology ampMetabolism vol 85 no 9 pp 3356ndash3364 2000
International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003

International Journal of Endocrinology 7
contribution of both tissues to the amount of this cytokinepresent in maternal circulation but the similar pattern of IL-6 mRNA overexpression in both tissues suggests that bothcould be a source of circulating IL-6 in pregnancy
Our group has recently shown that in addition toits peripheral metabolic effects IL-6 modulates the cen-tral mechanisms associated to maternal hyperphagia andincreased adiposity in the pregnantmouse [29] Although thetranscriptional expression of its hypothalamic neuropeptidetargets NPY AgRP POMC and CART in the human [2425] and rat placenta [20] is well established whether theyare also placentally produced and regulated by IL-6 duringmouse pregnancy remains unknown In order to clarify thisissue we determined the placental mRNA expression ofthese energy balance regulating neuropeptides throughoutpregnancy (from gestational days 13 to 18) in WT and IL-6deficient mice
The results of the present study revealed a similar tran-scriptional regulation of ARC neuropeptides in the mouseplacenta from that previously reported in the hypothalamus[29] Thus POMC mRNA levels were reduced from gesta-tional days 13 to 18 while the expression of the orexigenicand anabolic neuropeptides NPYAgRP remains unchangedA similar absence of relation between gestational age andplacental NPY mRNA content has been shown in otherspecies [20 25] On the contrary the placental ontogeneticexpression of the melanocortin system members POMC andAgRP seems to be a rather controversial issue Thereforesome studies reported a decrease in AgRP mRNA levels withgestational age and no change in POMC levels whilst othersshowed an upregulated mRNA expression of both neuropep-tides in human [24] and rat placentas [20 21] As far as we areaware the mRNA expression of CART has been previouslyexamined neither in human nor in murine placental tissuesIn this sense recent data from our group demonstrated thatthemRNA levels of this catabolic and anorectic neuropeptideare markedly increased in the rat placenta at term [20 21]However in the present study we failed to detect any changein CART gene expression throughout mouse pregnancy Thereason for this discrepancy is unclear but itmight reflect somebiological differences between species
Another novel finding of the present study is the fact thatIL-6 acting on an autocrine or paracrine manner modulatesthe placental expression of classical ARC energy balance reg-ulatory peptides as shown by IL-6 KOmice data Thus NPYand CART mRNA levels are upregulated by IL-6 deficiencyat late pregnancy while POMC gene expression is downregu-lated in placentas from 15-day pregnant IL-6 KOmice Theseresults indicate that during the third week of pregnancylack of IL-6 settles an anabolic milieu in the mouse placentaby reciprocally modulating the transcriptional expression ofneuropeptides known to either promote NPY or repressPOMC net energy gain As we have previously demonstratedin the hypothalamus of the mouse in late pregnancy a lackof IL-6 upregulates NPYAgRP mRNA levels in the ARCit seems that IL-6 deficiency during late pregnancy exertsa stimulatory effect on NPY gene transcription whatever isits source However the results obtained regarding placentalCARTPOMC gene expression were somewhat unexpected
Increased hypothalamic CART or POMC production hasbeen reported after endotoxemic challenge or central admin-istration of other well-known proinflammatory cytokinesas IL-1 [59 60] In addition exercise-induced hypothala-mic IL-6 and IL-10 in obese rats [61] and IL-6 astrocyte-targeted overexpression in female mice have been shownto increase POMC expression in the ARC [62] Converselytotal knockout of IL-6 in mice restored to virgin values thereduced POMC mRNA levels in the ARC of late pregnantmice probably reflecting a compensatorymechanism exertedby other factors [29] On the contrary the data reportedherein indicate that during the second half of pregnancy thiscompensatory effect on POMC gene expression is not settledin the mouse placenta Considering the above mentionedresults and the fact that CART and POMC are colocalized inthe ARC a similar pattern of placental expressionmight havebeen expected for both neuropeptides However we havenot performed in situ hybridization or immunohistochemicalstudies to characterize cellular distribution of CART andPOMC in placental tissue Therefore we cannot exclude thepossibility that different placental cell types could producethese neuropeptides and respond to absence of IL-6 in adifferent manner In fact with respect to their hypothalamicregulation it is worth to mention that CART mRNA hasbeen reported in a number of hypothalamic cell populationsbesides the ARC where its expression has been shown to bedifferentially modulated from that of POMC in response toendotoxin challenge [63]
Finally in relation to the previous point it is importantto take into account that the experimental approach used inthe current study was designed to assess during the secondhalf of pregnancy global changes in IL-6 and main energyregulating neuropeptides gene expression in the mouse pla-centa Additionally the IL-6 deficient mouse was chosen toaddress the influence of total absence of endogenous IL-6 onthis setting However the murine placenta consists of threemayor cell layers of maternal or placental origin the outermaternal decidua the middle junctional zone composedof fetoplacental trophoblast cells and the inner labyrinthzone [64] Therefore further in vitro studies involving eitherdecidual or trophoblast cells in culture would be helpful todetermine the ability of each cell population to produce andsecrete these neuropeptides and confirm the contribution ofIL-6 to their transcriptional modulation
Whatever is the case these results of the current studysuggest that IL-6 by exerting a direct or indirect regulatoryaction in the transcriptional control of NPY CART andPOMC expression may play a role in the homeostaticresponse to energy availability in mouse placenta Finallywhether placental neuropeptide protein levels correlate withthe ontogenetic neuropeptide mRNA pattern herein shownwill merit further investigation
5 Conclusions
In summary we demonstrate that as in other species theclassical hypothalamic neuropeptidergic systems involvedin energy homeostasis are present in the mouse placenta
8 International Journal of Endocrinology
These central signals display a specific ontogenetic expressionpattern in absence of IL-6 with major changes settled inplacentas at term Thus our results suggest that IL-6 a well-known immune and metabolic modulating factor mightmodulate their transcriptional expression by acting on anautocrine or paracrine manner Collectively these resultssuggest that during an energetic challenging condition suchas pregnancy IL-6 may also extend its action as a metabolicsignal to the main organ at the maternal-fetal interface theplacenta
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank Eva Vigo C Ruth Gonzalez and SiaraPerez for excellent technical assistance with the qPCRassays The HPRT gene primers for SYBR Green geneexpression analysis were kindly donated by professor ClaraV Alvarez Research Center of Molecular Medicine andChronicDiseases (CIMUS)This work has been supported byEuropean Community (FP72007n∘ 245009 ldquoNeuroFASTrdquo)Ministerio de Economıa y Competitividad (PP and MCGGBFU2007-62683BFI and PP MCGG and CD CIBERobn(CB0603)) and Xunta de Galicia Grants (MCGG and LLPGIDIT06PXIB208067PR) CIBER de Fisiopatologıa de laObesidad y Nutricion is an initiative of ISCIII
References
[1] J R Speakman D A Levitsky D B Allison et al ldquoSet pointssettling points and some alternative models theoretical optionsto understandhowgenes and environments combine to regulatebody adiposityrdquoDisease Models ampMechanisms vol 4 no 6 pp733ndash745 2011
[2] K W Williams and J K Elmquist ldquoFrom neuroanatomy tobehavior central integration of peripheral signals regulatingfeeding behaviorrdquoNature Neuroscience vol 15 no 10 pp 1350ndash1355 2012
[3] J E Schneider C M Klingerman and A Abdulhay ldquoSenseand nonsense in metabolic control of reproductionrdquo Frontiersin Endocrinology vol 3 article 26 2012
[4] D Richard B Monge-Roffarello K Chechi S M Labbe andE E Turcotte ldquoControl and physiological determinants ofsympatheticallymediated brown adipose tissue thermogenesisrdquoFrontiers in Endocrinology vol 3 article 36 2012
[5] MGMyers Jr andD POlson ldquoCentral nervous system controlof metabolismrdquo Nature vol 491 no 7424 pp 357ndash363 2012
[6] M Lopez S Tovar M J Vazquez L M Williams and CDieguez ldquoPeripheral tissue-brain interactions in the regulationof food intakerdquo Proceedings of the Nutrition Society vol 66 no1 pp 131ndash155 2007
[7] S R Ladyman R A Augustine and D R Grattan ldquoHormoneinteractions regulating energy balance during pregnancyrdquo Jour-nal of Neuroendocrinology vol 22 no 7 pp 805ndash817 2010
[8] K Sinchak and E J Wagner ldquoEstradiol signaling in theregulation of reproduction and energy balancerdquo Frontiers inEndocrinology vol 33 no 4 pp 342ndash363 2012
[9] A Crown D K Clifton and R A Steiner ldquoNeuropeptidesignaling in the integration of metabolism and reproductionrdquoNeuroendocrinology vol 86 no 3 pp 175ndash182 2007
[10] J W Hill J K Elmquist and C F Elias ldquoHypothalamicpathways linking energy balance and reproductionrdquo AmericanJournal of Physiology vol 294 no 5 pp E827ndashE832 2008
[11] S Kumar and G Kaur ldquoIntermittent fasting dietary restrictionregimen negatively influences reproduction in young rats astudy of hypothalamo-hypophysial-gonadal axisrdquo PLoS ONEvol 8 no 1 Article ID e52416 2013
[12] R Senaris T Garcia-Caballero X Casabiell et al ldquoSynthesis ofleptin in human placentardquo Endocrinology vol 138 no 10 pp4501ndash4504 1997
[13] N Hoogard L Hunter J S Duncan L M Williams PTrayhurn and J G Mercer ldquoLeptin and leptin receptor mRNAand protein expression in the murine fetus and placentardquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 20 pp 11073ndash11078 1997
[14] J E Caminos R Nogueiras R Gallego et al ldquoExpressionand regulation of adiponectin and receptor in human and ratplacentardquo The Journal of Clinical Endocrinology amp Metabolismvol 90 no 7 pp 4276ndash4286 2005
[15] H N Jones T Jansson and T L Powell ldquoFull-lengthadiponectin attenuates insulin signaling and inhibits insulin-stimulated amino acid transport in human primary trophoblastcellsrdquo Diabetes vol 59 no 5 pp 1161ndash1170 2010
[16] N di Simone F di Nicuolo D Marzioni et al ldquoResistinmodulates glucose uptake and glucose transporter-1 (GLUT-1) expression in trophoblast cellsrdquo Journal of Cellular andMolecular Medicine vol 13 no 2 pp 388ndash397 2009
[17] S Yura N Sagawa H Itoh et al ldquoResistin is expressed inthe human placentardquo The Journal of Clinical Endocrinology ampMetabolism vol 88 no 3 pp 1394ndash1397 2003
[18] N Noyola-Martıneza L Dıaz E Avila A Halhali F Larreaand D Barrera ldquoCalcitriol downregulates TNF-120572 and IL-6 expression in cultured placental cells from preeclampticwomenrdquo Cytokine vol 61 no 1 pp 245ndash250 2013
[19] S K Munro M D Mitchell and A P Ponnampalam ldquoHistonedeacetylase inhibition by trichostatin A mitigates LPS inducedTNF120572 and IL-10 production in human placental explantsrdquoPlacenta vol 34 no 7 pp 567ndash573 2013
[20] R Beloosesky D A Gayle F Amidi S N Ahanya M Desaiand M G Ross ldquoOntogenic expression of putative feedingpeptides in the rat fetal brain and placentardquo Nutritional Neu-roscience vol 9 no 1-2 pp 33ndash40 2006
[21] J E Caminos S B Bravo C R Gonzalez et al ldquoFood intakeregulating-neuropeptides are expressed and regulated throughpregnancy and following food restriction in rat placentardquoReproductive Biology and Endocrinology vol 6 article 14 2008
[22] S I Grigorakis E Anastasiou K Dai A Souvatzoglou andM Alevizaki ldquoThree mRNA transcripts of the proopiome-lanocortin gene in human placenta at termrdquo European Journalof Endocrinology vol 142 no 5 pp 533ndash536 2000
[23] J Robidoux L Simoneau A Masse and J Lafond ldquoActivationof L-type calcium channels induces corticotropin-releasing fac-tor secretion from human placental trophoblastsrdquo The Journalof Clinical Endocrinology ampMetabolism vol 85 no 9 pp 3356ndash3364 2000
International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003

8 International Journal of Endocrinology
These central signals display a specific ontogenetic expressionpattern in absence of IL-6 with major changes settled inplacentas at term Thus our results suggest that IL-6 a well-known immune and metabolic modulating factor mightmodulate their transcriptional expression by acting on anautocrine or paracrine manner Collectively these resultssuggest that during an energetic challenging condition suchas pregnancy IL-6 may also extend its action as a metabolicsignal to the main organ at the maternal-fetal interface theplacenta
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors thank Eva Vigo C Ruth Gonzalez and SiaraPerez for excellent technical assistance with the qPCRassays The HPRT gene primers for SYBR Green geneexpression analysis were kindly donated by professor ClaraV Alvarez Research Center of Molecular Medicine andChronicDiseases (CIMUS)This work has been supported byEuropean Community (FP72007n∘ 245009 ldquoNeuroFASTrdquo)Ministerio de Economıa y Competitividad (PP and MCGGBFU2007-62683BFI and PP MCGG and CD CIBERobn(CB0603)) and Xunta de Galicia Grants (MCGG and LLPGIDIT06PXIB208067PR) CIBER de Fisiopatologıa de laObesidad y Nutricion is an initiative of ISCIII
References
[1] J R Speakman D A Levitsky D B Allison et al ldquoSet pointssettling points and some alternative models theoretical optionsto understandhowgenes and environments combine to regulatebody adiposityrdquoDisease Models ampMechanisms vol 4 no 6 pp733ndash745 2011
[2] K W Williams and J K Elmquist ldquoFrom neuroanatomy tobehavior central integration of peripheral signals regulatingfeeding behaviorrdquoNature Neuroscience vol 15 no 10 pp 1350ndash1355 2012
[3] J E Schneider C M Klingerman and A Abdulhay ldquoSenseand nonsense in metabolic control of reproductionrdquo Frontiersin Endocrinology vol 3 article 26 2012
[4] D Richard B Monge-Roffarello K Chechi S M Labbe andE E Turcotte ldquoControl and physiological determinants ofsympatheticallymediated brown adipose tissue thermogenesisrdquoFrontiers in Endocrinology vol 3 article 36 2012
[5] MGMyers Jr andD POlson ldquoCentral nervous system controlof metabolismrdquo Nature vol 491 no 7424 pp 357ndash363 2012
[6] M Lopez S Tovar M J Vazquez L M Williams and CDieguez ldquoPeripheral tissue-brain interactions in the regulationof food intakerdquo Proceedings of the Nutrition Society vol 66 no1 pp 131ndash155 2007
[7] S R Ladyman R A Augustine and D R Grattan ldquoHormoneinteractions regulating energy balance during pregnancyrdquo Jour-nal of Neuroendocrinology vol 22 no 7 pp 805ndash817 2010
[8] K Sinchak and E J Wagner ldquoEstradiol signaling in theregulation of reproduction and energy balancerdquo Frontiers inEndocrinology vol 33 no 4 pp 342ndash363 2012
[9] A Crown D K Clifton and R A Steiner ldquoNeuropeptidesignaling in the integration of metabolism and reproductionrdquoNeuroendocrinology vol 86 no 3 pp 175ndash182 2007
[10] J W Hill J K Elmquist and C F Elias ldquoHypothalamicpathways linking energy balance and reproductionrdquo AmericanJournal of Physiology vol 294 no 5 pp E827ndashE832 2008
[11] S Kumar and G Kaur ldquoIntermittent fasting dietary restrictionregimen negatively influences reproduction in young rats astudy of hypothalamo-hypophysial-gonadal axisrdquo PLoS ONEvol 8 no 1 Article ID e52416 2013
[12] R Senaris T Garcia-Caballero X Casabiell et al ldquoSynthesis ofleptin in human placentardquo Endocrinology vol 138 no 10 pp4501ndash4504 1997
[13] N Hoogard L Hunter J S Duncan L M Williams PTrayhurn and J G Mercer ldquoLeptin and leptin receptor mRNAand protein expression in the murine fetus and placentardquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 94 no 20 pp 11073ndash11078 1997
[14] J E Caminos R Nogueiras R Gallego et al ldquoExpressionand regulation of adiponectin and receptor in human and ratplacentardquo The Journal of Clinical Endocrinology amp Metabolismvol 90 no 7 pp 4276ndash4286 2005
[15] H N Jones T Jansson and T L Powell ldquoFull-lengthadiponectin attenuates insulin signaling and inhibits insulin-stimulated amino acid transport in human primary trophoblastcellsrdquo Diabetes vol 59 no 5 pp 1161ndash1170 2010
[16] N di Simone F di Nicuolo D Marzioni et al ldquoResistinmodulates glucose uptake and glucose transporter-1 (GLUT-1) expression in trophoblast cellsrdquo Journal of Cellular andMolecular Medicine vol 13 no 2 pp 388ndash397 2009
[17] S Yura N Sagawa H Itoh et al ldquoResistin is expressed inthe human placentardquo The Journal of Clinical Endocrinology ampMetabolism vol 88 no 3 pp 1394ndash1397 2003
[18] N Noyola-Martıneza L Dıaz E Avila A Halhali F Larreaand D Barrera ldquoCalcitriol downregulates TNF-120572 and IL-6 expression in cultured placental cells from preeclampticwomenrdquo Cytokine vol 61 no 1 pp 245ndash250 2013
[19] S K Munro M D Mitchell and A P Ponnampalam ldquoHistonedeacetylase inhibition by trichostatin A mitigates LPS inducedTNF120572 and IL-10 production in human placental explantsrdquoPlacenta vol 34 no 7 pp 567ndash573 2013
[20] R Beloosesky D A Gayle F Amidi S N Ahanya M Desaiand M G Ross ldquoOntogenic expression of putative feedingpeptides in the rat fetal brain and placentardquo Nutritional Neu-roscience vol 9 no 1-2 pp 33ndash40 2006
[21] J E Caminos S B Bravo C R Gonzalez et al ldquoFood intakeregulating-neuropeptides are expressed and regulated throughpregnancy and following food restriction in rat placentardquoReproductive Biology and Endocrinology vol 6 article 14 2008
[22] S I Grigorakis E Anastasiou K Dai A Souvatzoglou andM Alevizaki ldquoThree mRNA transcripts of the proopiome-lanocortin gene in human placenta at termrdquo European Journalof Endocrinology vol 142 no 5 pp 533ndash536 2000
[23] J Robidoux L Simoneau A Masse and J Lafond ldquoActivationof L-type calcium channels induces corticotropin-releasing fac-tor secretion from human placental trophoblastsrdquo The Journalof Clinical Endocrinology ampMetabolism vol 85 no 9 pp 3356ndash3364 2000
International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003

International Journal of Endocrinology 9
[24] C L Chen C C Chang D T Krieger and C W BardinldquoExpression and regulation of proopiomelanocortin-like genein the ovary and placenta comparison with the testisrdquoEndocrinology vol 118 no 6 pp 2382ndash2389 1986
[25] J Dotsch K-D Nusken I Knerr M Kirschbaum R Reppand W Rascher ldquoLeptin and neuropeptide Y gene expressionin human placenta ontogeny and evidence for similarities tohypothalamic regulationrdquoThe Journal of Clinical Endocrinologyamp Metabolism vol 84 no 8 pp 2755ndash2758 1999
[26] M Erta A Quintana and J Hidalgo ldquoInterleukin-6 a majorcytokine in the central nervous systemrdquo International Journalof Biological Sciences vol 8 no 9 pp 1254ndash1266 2012
[27] S Lager N Jansson A L Olsson M Wennergren T Janssonand T L Powell ldquoEffect of IL-6 and TNF-120572 on fatty acid uptakein cultured human primary trophoblast cellsrdquo Placenta vol 32no 2 pp 121ndash127 2011
[28] R Shao M Nutu L Karlsson-Lindahl et al ldquoDownregulationof cilia-localized Il-6R120572 by 17120573-estradiol in mouse and humanfallopian tubesrdquo American Journal of Physiology vol 297 no 1pp C140ndashC151 2009
[29] P Pazos L Lima F F Casanueva C Dieguez andM C GarcialdquoInterleukin 6 deficiency modulates the hypothalamic expres-sion of energy balance regulating peptides during pregnancy inmicerdquo PLoS ONE vol 8 no 8 Article ID e72339 2013
[30] J-O Jansson K Wallenius I Wernstedt C Ohlsson S LDickson and V Wallenius ldquoOn the site and mechanism ofaction of the anti-obesity effects of interleukin-6rdquo GrowthHormone and IGF Research vol 13 supplement A pp S28ndashS322003
[31] E Schele A Benrick L Grahnemo et al ldquoInter-relationbetween interleukin (IL)-1 IL-6 and body fat regulating circuitsof the hypothalamic arcuate nucleusrdquo Journal of Neuroen-docrinology vol 25 no 6 pp 580ndash589 2013
[32] K Stenlof I Wernstedt T Fjallman V Wallenius K Wal-lenius and J-O Jansson ldquoInterleukin-6 levels in the centralnervous system are negatively correlated with fat mass in over-weightobese subjectsrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 88 no 9 pp 4379ndash4383 2003
[33] V Wallenius K Wallenius B Ahren et al ldquoInterleukin-6-deficient mice develop mature-onset obesityrdquo Nature Medicinevol 8 no 1 pp 75ndash79 2002
[34] C T de Souza E P Araujo S Bordin et al ldquoConsumption of afat-rich diet activates a proinflammatory response and inducesinsulin resistance in the hypothalamusrdquo Endocrinology vol 146no 10 pp 4192ndash4199 2005
[35] MMilanski GDegasperi A Coope et al ldquoSaturated fatty acidsproduce an inflammatory response predominantly through theactivation of TLR4 signaling in hypothalamus implications forthe pathogenesis of obesityrdquoThe Journal of Neuroscience vol 29no 2 pp 359ndash370 2009
[36] K Wallenius V Wallenius D Sunter S L Dickson and J-O Jansson ldquoIntracerebroventricular interleukin-6 treatmentdecreases body fat in ratsrdquoBiochemical and Biophysical ResearchCommunications vol 293 no 1 pp 560ndash565 2002
[37] I Wernstedt A Edgley A Berndtsson et al ldquoReducedstress- and cold-induced increase in energy expenditure ininterleukin-6-deficient micerdquo American Journal of Physiologyvol 291 no 3 pp R551ndashR557 2006
[38] J Faldt I Wernstedt S M Fitzgerald K Wallenius GBergstrom and J-O Jansson ldquoReduced exercise endurance ininterleukin-6-deficient micerdquo Endocrinology vol 145 no 6 pp2680ndash2686 2004
[39] J Hidalgo S Florit M Giralt B Ferrer C Keller and HPilegaard ldquoTransgenicmice with astrocyte-targeted productionof interleukin-6 are resistant to high-fat diet-induced increasesin body weight and body fatrdquo Brain Behavior and Immunityvol 24 no 1 pp 119ndash126 2010
[40] M B Flores M F Fernandes E R Ropelle et al ldquoExerciseimproves insulin and leptin sensitivity in hypothalamus ofWistar ratsrdquo Diabetes vol 55 no 9 pp 2554ndash2561 2006
[41] B C Borges R Rorato Y Avraham et al ldquoLeptin resistance anddesensitization of hypophagia during prolonged inflammatorychallengerdquo American Journal of Physiology vol 300 no 5 ppE858ndashE869 2011
[42] A Benrick E Schele S B Pinnock et al ldquoInterleukin-6 geneknockout influences energy balance regulating peptides in thehypothalamic paraventricular and supraoptic nucleirdquo Journal ofNeuroendocrinology vol 21 no 7 pp 620ndash628 2009
[43] E Schele C Fekete P Egri et al ldquoInterleukin-6 receptor 120572 is co-localised with melanin-concentrating hormone in human andmouse hypothalamusrdquo Journal of Neuroendocrinology vol 24no 6 pp 930ndash943 2012
[44] S Lager and T L Powell ldquoRegulation of nutrient transportacross the placentardquo Journal of Pregnancy vol 2012 Article ID179827 14 pages 2012
[45] M Kopf H Baumann G Freer et al ldquoImpaired immune andacute-phase responses in interleukin-6-deficient micerdquo Naturevol 368 no 6469 pp 339ndash342 1994
[46] M J Vazquez C R Gonzalez L Varela et al ldquoCentral resistinregulates hypothalamic and peripheral lipid metabolism in anutritional-dependent fashionrdquo Endocrinology vol 149 no 9pp 4534ndash4543 2008
[47] I Rioja K A Bush J B Buckton M C Dickson and PF Life ldquoJoint cytokine quantification in two rodent arthritismodels kinetics of expression correlation of mRNA andprotein levels and response to prednisolone treatmentrdquo Clinicalamp Experimental Immunology vol 137 no 1 pp 65ndash73 2004
[48] F Isken M O Weickert M H Tschop et al ldquoMetaboliceffects of diets differing in glycaemic index depend on age andendogenous glucose-dependent insulinotrophic polypeptide inmicerdquo Diabetologia vol 52 no 10 pp 2159ndash2168 2009
[49] S B Bravo J E Caminos C R Gonzalez et al ldquoLeptinand fasting regulate rat gastric glucose-regulated protein 58rdquoInternational Journal of Peptides vol 2011 Article ID 96981811 pages 2011
[50] M Garcia-Lavandeira V Quereda I Flores et al ldquoAGRFa2Prop1Stem (GPS) cell niche in the pituitaryrdquo PLoSONE vol 4 no 3 Article ID e4815 2009
[51] S B Bravo M E Garcia-Rendueles A R Garcia-Rendueleset al ldquoHumanized medium (h7H) allows long-term primaryfollicular thyroid cultures from human normal thyroid benignneoplasm and cancerrdquoThe Journal of Clinical Endocrinology ampMetabolism vol 98 no 6 pp 2431ndash2441 2013
[52] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the2-ΔΔ119862T methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[53] N M Orsi N Gopichandran U V Ekbote and J J WalkerldquoMurine serum cytokines throughout the estrous cycle preg-nancy and post partum periodrdquo Animal Reproduction Sciencevol 96 no 1-2 pp 54ndash65 2006
[54] P JMark J L LewisM L Jones J A Keelan and B JWaddellldquoThe inflammatory state of the rat placenta increases in lategestation and is further enhanced by glucocorticoids in thelabyrinth zonerdquo Placenta vol 34 no 7 pp 559ndash566 2013
10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003

10 International Journal of Endocrinology
[55] S Hauguel-de Mouzon and M Guerre-Millo ldquoThe placentacytokine network and inflammatory signalsrdquo Placenta vol 27no 8 pp 794ndash798 2006
[56] R Agarwal A Loganath A C Roy Y C Wong C Lindoffand S C Ng ldquoIncreased expression of interleukin 6 in termcompared to the first trimester human placental villirdquoHormoneand Metabolic Research vol 32 no 5 pp 164ndash168 2000
[57] M de T H Sanford and G W Wood ldquoDetection ofinterleukin-1 interleukin-6 and tumor necrosis factor-120572 in theuterus during the second half of pregnancy in the mouserdquoEndocrinology vol 131 no 1 pp 14ndash20 1992
[58] L Zhang T Sugiyama N Murabayashi et al ldquoThe inflamma-tory changes of adipose tissue in late pregnant micerdquo Journal ofMolecular Endocrinology vol 47 no 2 pp 157ndash165 2011
[59] T M Reyes and P E Sawchenko ldquoInvolvement of the arcu-ate nucleus of the hypothalamus in interleukin-1-inducedanorexiardquoThe Journal of Neuroscience vol 22 no 12 pp 5091ndash5099 2002
[60] B C Borges J Antunes-Rodrigues M Castro J C BittencourtC F Elias and L L K Elias ldquoExpression of hypothalamicneuropeptides and the desensitization of pituitary-adrenal axisand hypophagia in the endotoxin tolerancerdquo Hormones andBehavior vol 52 no 4 pp 508ndash519 2007
[61] E R Ropelle M B Flores D E Cintra et al ldquoIL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamicinsulin and leptin sensitivity through IKK120573 and ER stressinhibitionrdquoPLoSBiology vol 8 no 8Article ID e1000465 2010
[62] R M Senaris M L Trujillo B Navia et al ldquoInterleukin-6 regulates the expression of hypothalamic neuropeptidesinvolved in body weight in a gender-dependent wayrdquo Journalof Neuroendocrinology vol 23 no 8 pp 675ndash686 2011
[63] V Sergeyev C Broberger and T Hokfelt ldquoEffect of LPSadministration on the expression of POMC NPY galaninCART and MCH mRNAs in the rat hypothalamusrdquo MolecularBrain Research vol 90 no 2 pp 93ndash100 2001
[64] A Malassine J-L Frendo and D Evain-Brion ldquoA comparisonof placental development and endocrine functions between thehuman andmouse modelrdquoHuman Reproduction Update vol 9no 6 pp 531ndash539 2003