ultrastructure of the normal human placenta
TRANSCRIPT

Electron Microsc. Rev., Vol. 4, pp. 129 178, 1991. 0892-0354/91 $0.00 + 0.50 Printed in Great Britain. All rights reserved. © 1991 Pergamon Press plc.
U L T R A S T R U C T U R E O F T H E N O R M A L H U M A N P L A C E N T A
C A R O L Y N J. P . J O N E S a n d H A R O L D F O X
Depar tment o f Pathological Sciences, University o f Manchester, O x f o r d Road, Manches ter M 1 3 9PT, U.K.
Abstract--In this review, the ultrastructure of the normal h u m a n chorionic villus is examined and illustrated. The outer covering of trophoblast, comprising the generative cytotrophoblast and the multinucleated syncytiotro- phoblast which is derived from it, is described, including such features as the microvillous surface, vesicles and vacuoles, endoplasmic reticulum and secretory droplets. The structure, composition, development and inclusions of the trophoblastic basement membrane are discussed, and the ultrastructure of the various components of the stroma, including reticulum cells, fibroblasts, Hofbauer ceils, capillaries and the non-cellular matrix are illustrated and described, with special reference to their inter-relationships and function.
C O N T E N T S
I. In t roduc t ion ........................................................................................................................................................................... 129 II. Structure of the chorionic villus ............................................................................................................................................ 130
III. Cytotrophoblast ..................................................................................................................................................................... 130 A. First trimester ................................................................................................................................................................... 130 B. Second trimester ............................................................................................................................................................... 138 C. Third trimester ................................................................................................................................................................. 138 D. Funct ion ........................................................................................................................................................................... 138
IV. Syncytiotrophoblast ............................................................................................................................................................... 139 A. Microvillous surtace ......................................................................................................................................................... 139 B. Endoplasmic reticulum ..................................................................................................................................................... 145 C. Ribosomes ........................................................................................................................................................................ 147 D. Mitochondria .................................................................................................................................................................... 147 E. Vacuoles, multivesicular and dense bodies ...................................................................................................................... 147 F. Lipid droplets ................................................................................................................................................................... 148 G. Secretory droplets/Golgi appara tus /hormone biosynthesis ............................................................................................. 148 H. Nuclei ............................................................................................................................................................................... 151 I. Cytoskeleton ..................................................................................................................................................................... 151 J. Basal plasma membrane .................................................................................................................................................. 151
V. Trophoblastic basement membrane ....................................................................................................................................... 152 A. Structure ........................................................................................................................................................................... 152 B. Composi t ion ..................................................................................................................................................................... 152 C. Development ..................................................................................................................................................................... 152 D. Inclusions .......................................................................................................................................................................... 153
VI. The s t roma of the chorionic villus ........................................................................................................................................ 153 A. Reticulum cells ................................................................................................................................................................. 153 B. Fibroblasts ........................................................................................................................................................................ 155 C. Hofbauer cells .................................................................................................................................................................. 157 D. Capillaries ......................................................................................................................................................................... 165 E. Matrix ............................................................................................................................................................................... 170 Acknowledgements ................................................................................................................................................................. 171 References .............................................................................................................................................................................. 171
I . I N T R O D U C T I O N
T h e p l a c e n t a is a m u l t i f a c e t t e d f e t a l s t r u c t u r e ,
c o m b i n i n g i n o n e o r g a n m a n y f u n c t i o n a l a c t i v i t i e s
which are separate in the adult. Thus it is involved in the gaseous and nutritive transfer between mother and fetus, provides substrates for fetal metabolism and disposes of the fetal waste prod-
129

130 c.J .P. Jones and H. Fox
ucts. It is a major site of hormone synthesis and acts, in an as yet unknown manner, as an immuno- logical barrier between the mother and the fetal 'graft'. During the 280 days of a normal gestation it continuously adapts to the requirements of the fetus, a process entailing rapid modification of structure and function, and at birth becomes separ- ated from the mother to be expelled harmlessly, despite its extreme vascularity.
Wigglesworth (1973), Kaufmann et al. (1979) and Ramsey (1987). However, in order to orientate those unfamiliar with placental structure, the basic features of the terminal chorionic villus are illus- trated in Fig. 1, while the main ultrastructural characteristics are diagrammatically represented in Fig. 2.
III. THE C Y T O T R O P H O B L A S T
II. STRUCTURE OF THE CHORIONIC VILLUS
The terminal chorionic villi may be likened to the smallest 'twigs' of the placental 'tree' and are the basic functional units of the placenta. They vary in diameter from about 170/~m in the first trimester to approximately 40 #m at term and have a central mesenchymal core which is covered by a mantle of trophoblast. The trophoblastic covering consists of an outer multinucleated layer, the syn- cytiotrophoblast, and an inner layer of single cells, the cytotrophoblast. This inner cellular layer, which gives rise to the syncytium by a process of cell multiplication and subsequent fusion, is almost continuous in early pregnancy but, in the later stages of gestation, only occasional cells are found.
The mesenchymal villous core is initially avascu- lar but a capillary network eventually develops, this arising in situ from the chorionic mesenchyme. Fibroblasts, macrophages (Hofbauer cells) and a collagenous stroma are also present. During the course of pregnancy, the fetal capillaries dilate while the overlying syncytiotrophoblast becomes attenuated in some areas to form the so-called vasculo-syncytial membranes--these allow the maternal and fetal circulations to become closely apposed to each other, thus facilitating the materno-fetal exchange of oxygen. This barrier may become extremely thin, less than 2 # m at term, which is only slightly greater than the pulmonary alveolar blood/air barrier.
This review is intended to cover the ultra- structural features of the terminal chorionic villus; for a detailed analysis of the development and histology of the placenta the reader is referred to the publications by Boyd and Hamilton (1970),
A. First Trimester
The cytotrophoblast is the stem cell of the trophoblastic mantle and, in the first trimester of pregnancy, forms an almost complete layer be- neath the syncytium (Fig. 3). Three differing types can be identified (Jones, 1976): undifferentiated cells, those intermediate in character between the cyto- and syncytiotrophoblast and electron-dense cells which are often closely associated with the endothelium of the stromal capillaries.
1. Undifferentiated cells
The undifferentiated cytotrophoblastic cells (Fig. 3) are generally cuboidal with a large, ovoid nucleus containing evenly dispersed chromatin. A prominent nucleolus is often present and oc- casional pores can be seen in the nuclear mem- brane which is studded with ribosomes on its outer aspect. The cytoplasm is relatively electron-lucent with a paucity of organelles; rough endoplasmic reticulum generally forms narrow cisternae although sometimes an expanded form is seen which contains light, flocculent material (Fig. 4). These expanded cisternae tend to be associated with round mitochondria having an electron-dense matrix and pale, irregularly-shaped cristae, whereas the narrow cisternae are associated with much paler mitochondria having narrow cristae of a similar electron density. It is possible that the dark mitochondria are demonstrating the 'con- densed' (as opposed to the 'orthodox') configur- ation (Hackenbrock, 1966, 1968; Jasper, 1975) which is found in an actively phosphorylating state, and is thought to be indicative of intense biosynthesis. All types of mitochondria exhibit
Ultrastructure of the Human Placenta 131
C A P
c"
7 ,
• Q , , ~-~,
1~ A ,J ~
• i
/ ..:': Cy, j I : N i '
~ •
~ r
.r' ("
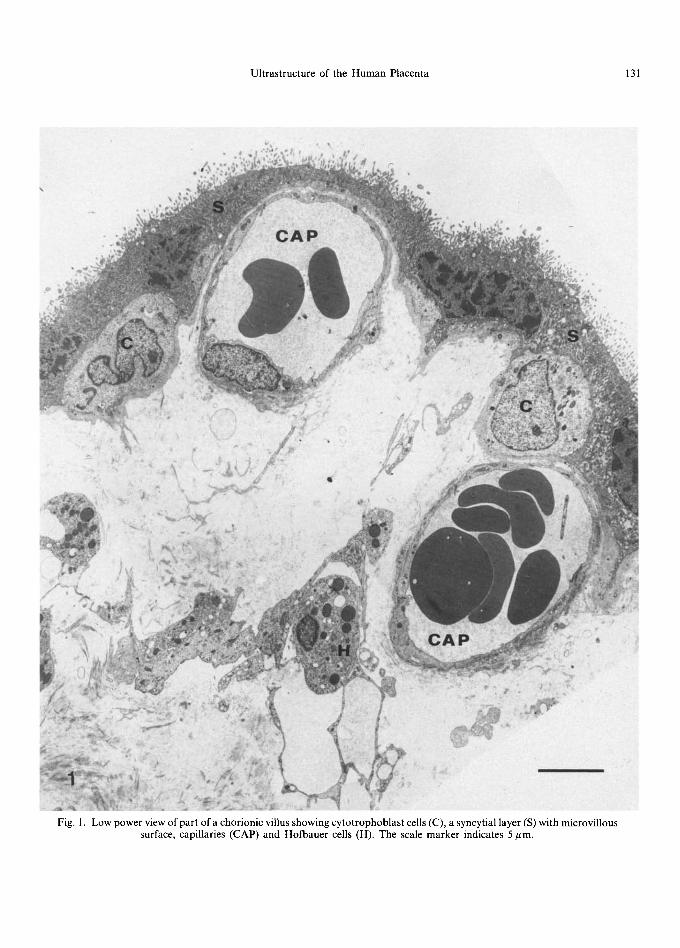
Fig. 1. Low power view of part ofa chorionic villus showing cytotrophoblast cells (C), a syncytial layer (S) with microvillous surface, capillaries (CAP) and Hofbauer cells (H). The scale marker indicates 5 pm.
132 C . J . P . Jones and H. Fox
NEMATOSOME--,--
DESMOSOME--
COATED VESICLE--'--"
MITOCHONDRION
MICROTUBU
CYTOTROPHO- BLASTIC CELl NUCLEUS
RIBOSOMES--
GOLGI BODY & DROPLETS "="
CYTOPLASMIC FILAMENTS""
R O U G H - - ENDOPLASMIC RETICULUM
BASEMENT MEMBRANE
BASAL YNTERDIGI TATIONS
2
O @ e ,
<__
f
DENSE MULTI- VESICULAR BODY
ROUGH ENDOPLASMIC RETICULUM
COATED
: ! / / : : : . . • o e o l l a ~ i o 4
4
i e o o • • •
• • u o i ~ e o e • •
~ a a o o m e o j
":.
.o
MACRO- ~ PINOCYTOTIC
~ _ ~ _ ~ CHANNEL
BODY
DENSE BODY
SECRETORY DROPLET
GOLGI BODY
APICAL '~kCUOLE
SYNCYTIAL
MICROVlIIUS
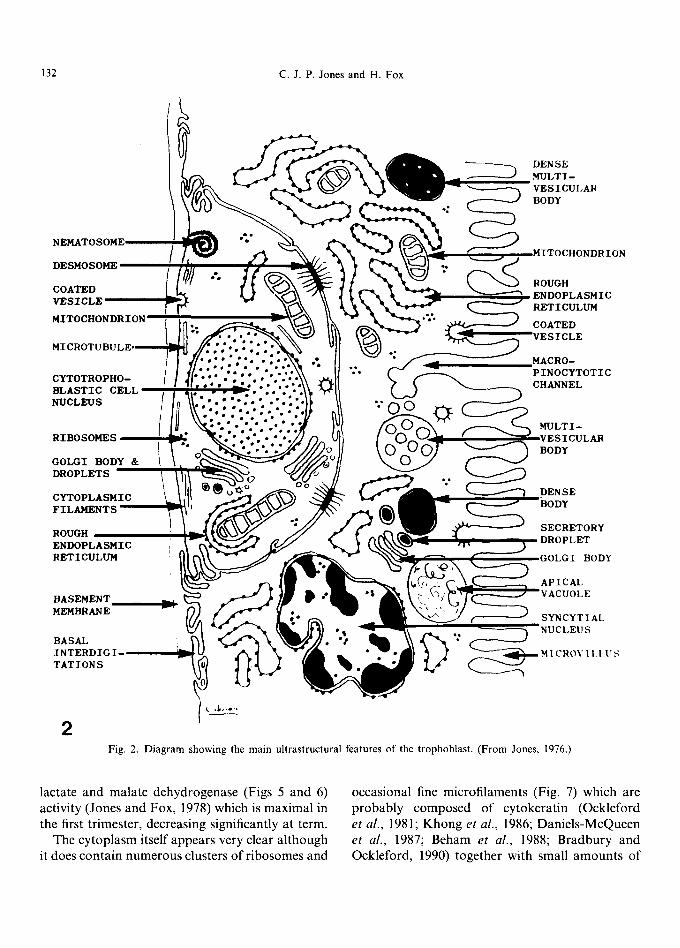
Fig. 2. Diagram showing the main ultrastructural features of the trophoblast. (From Jones, 1976.)
lactate and malate dehydrogenase (Figs 5 and 6) activity (Jones and Fox, 1978) which is maximal in the first trimester, decreasing significantly at term.
The cytoplasm itself appears very clear although it does contain numerous clusters of ribosomes and
occasional fine micro filaments (Fig. 7) which are probably composed of cytokeratin (Ockleford et al., 1981; Khong et al., 1986; Daniels-McQueen et al., 1987; Beham et al., 1988; Bradbury and Ockleford, 1990) together with small amounts of
Ultrastructure of the Human Placenta 133
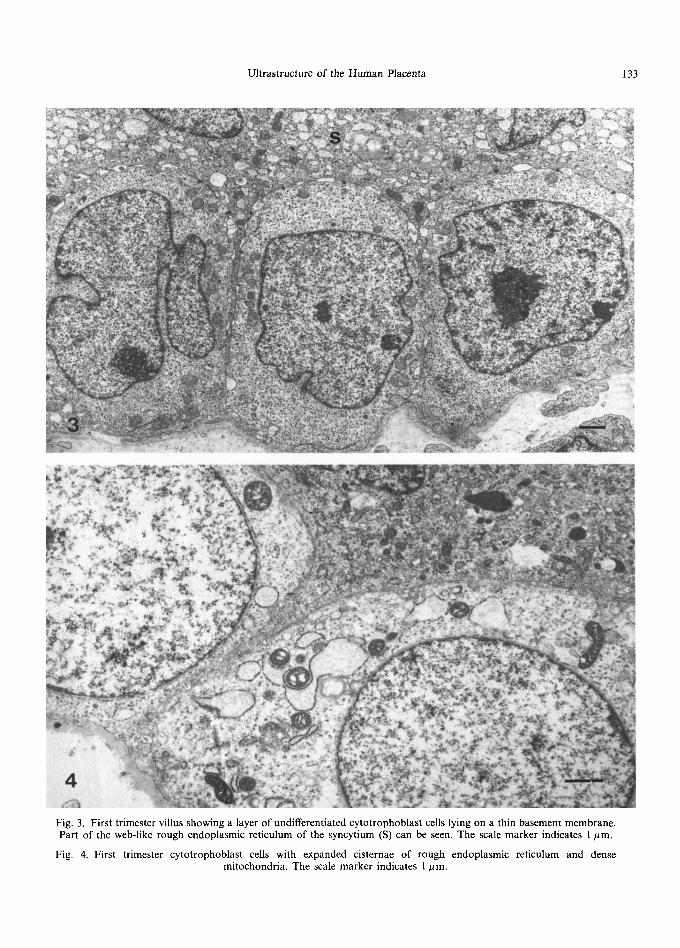
Fig. 3. First trimester villus showing a layer of undifferentiated cytotrophoblast cells lying on a thin basement membrane. Part of the web-like rough endoplasmic reticulum of the syncytium (S) can be seen. The scale marker indicates 1 #m.
Fig. 4. First trimester cytotrophoblast cells with expanded cisternae of rough endoplasmic reticulum and dense mitochondria. The scale marker indicates 1 ttm.
134 C . J . P . Jones and H. Fox
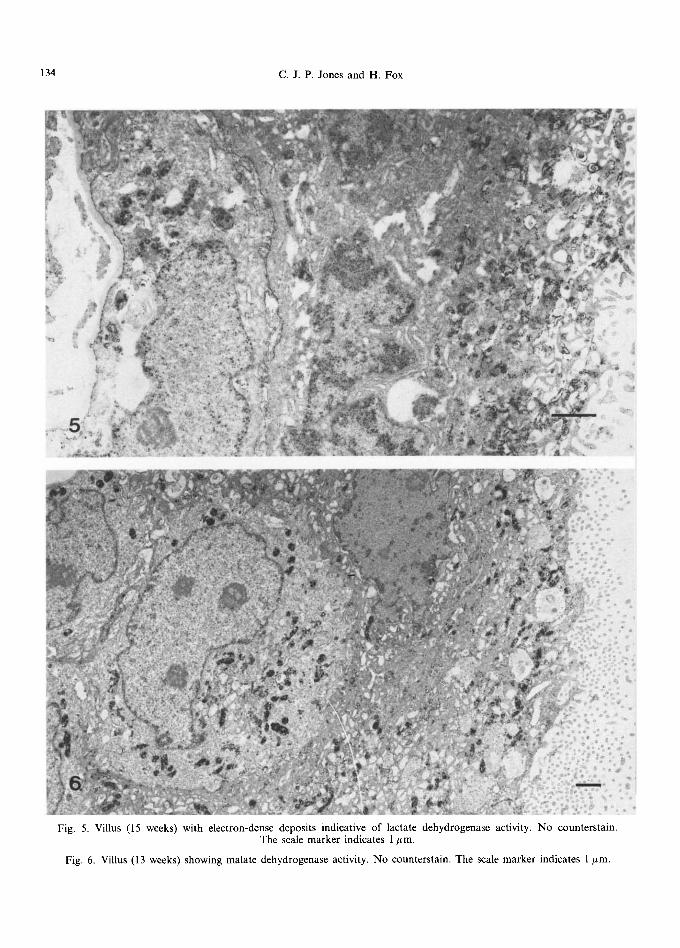
Fig. 5. Villus (15 weeks) with electron-dense deposits indicative of lactate dehydrogenase activity. No counterstain. The scale marker indicates 1 #m.
Fig. 6. Villus (13 weeks) showing malate dehydrogenase activity. No counterstain. The scale marker indicates 1 #m.
Ultrastructure of the Human Placenta 135
actin (King, 1983); microtubules with their associ- ated centrioles are also found. Glycogen deposits are present in some cells (Inoue, 1960; Terzakis, 1963; Anderson and McKay, 1966; Knoth, 1968; Boyd and Hamilton, 1970; Jones, 1976). Secretory droplets are infrequent, and Golgi bodies are variable in their distribution. Occasional micro- peroxisomes have been demonstrated in the cyto- trophoblast: these are about 200-300nm in diameter but are difficult to detect without special staining methods which reveal their catalase activity (King, 1982a).
The basal plasma membrane is generally smooth, apart f rom occasional caveolae and sur- face irregularities, and abuts on a thin basement membrane, although interdigitations are often pre- sent on the area adjoining the syncytium (Ashley, 1965). On the outer surface, where the cells lie against the syncytium, there are occasional local dilatations between the adjacent plasma mere-
branes which contain small villous protrusions of the cytotrophoblast . Receptors for epidermal growth factor (EGF) are present on this membrane (Kawagoe et al., 1990) as well as alkaline phospha- tase activity (Fig. 8) (Jones and Fox, 1976). Desmosomes attach the cytotrophoblast ic cells to the syncytiotrophoblast and, where their lateral walls are in contact, to each other (Metz and Weihe, 1980).
2. In termedia te cells
The ultrastructure of cells intermediate in char- acter between the undifferentiated cytotropho- blastic cell and the syncytium was first described by Boyd and Hughes (1954), and later by Terzakis (1963), Pierce and Midgley (1963) and Pierce et al. (1964). A typical intermediate cell is illustrated in Fig. 9. The nucleus is somewhat irregular in shape, with chromatin aggregated beneath the nuclear
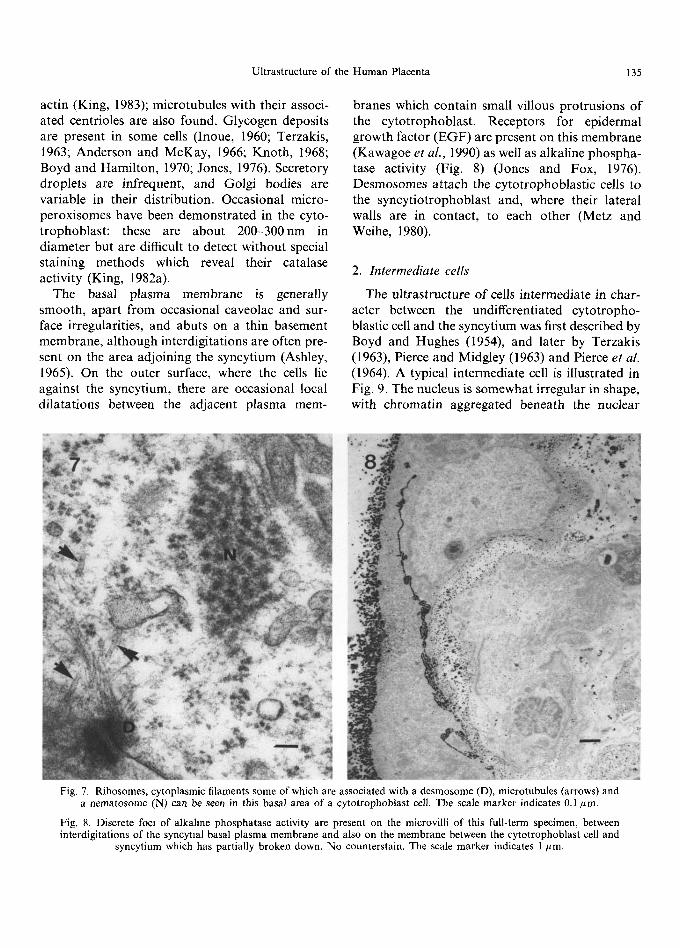
Fig. 7. Ribosomes, cytoplasmic filaments some of which are associated with a desmosome (D), microtubules (arrows) and a nematosome (N) can be seen in this basal area of a cytotropboblast cell. The scale marker indicates 0.1/~m.
Fig. 8. Discrete foci of alkaline phosphatase activity are present on the microvilli of this full-term specimen, between interdigitations of the syncytial basal plasma membrane and also on the membrane between the cytotrophoblast cell and
syncytium which has partially broken down. No counterstain. The scale marker indicates 1 kLm.
136 C.J.P. Jones and H. Fox
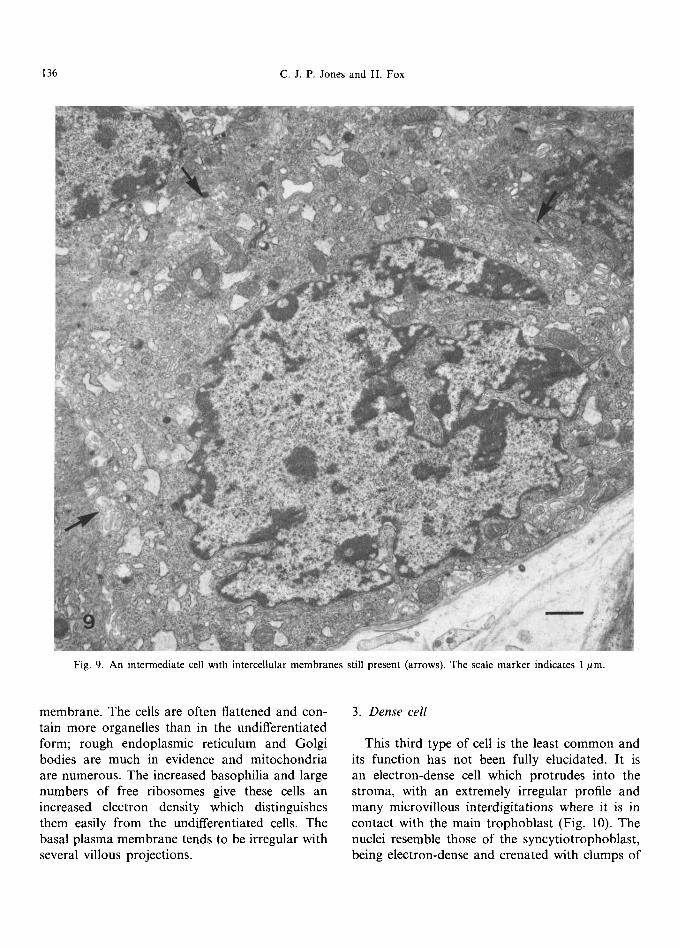
Fig. 9. An intermediate cell with intercellular membranes still present (arrows). The scale marker indicates 1 #m.
membrane. The cells are often flattened and con- tain more organelles than in the undifferentiated form; rough endoplasmic reticulum and Golgi bodies are much in evidence and mitochondria are numerous. The increased basophilia and large numbers of free ribosomes give these cells an increased electron density which distinguishes them easily from the undifferentiated cells. The basal plasma membrane tends to be irregular with several villous projections.
3. Dense cell
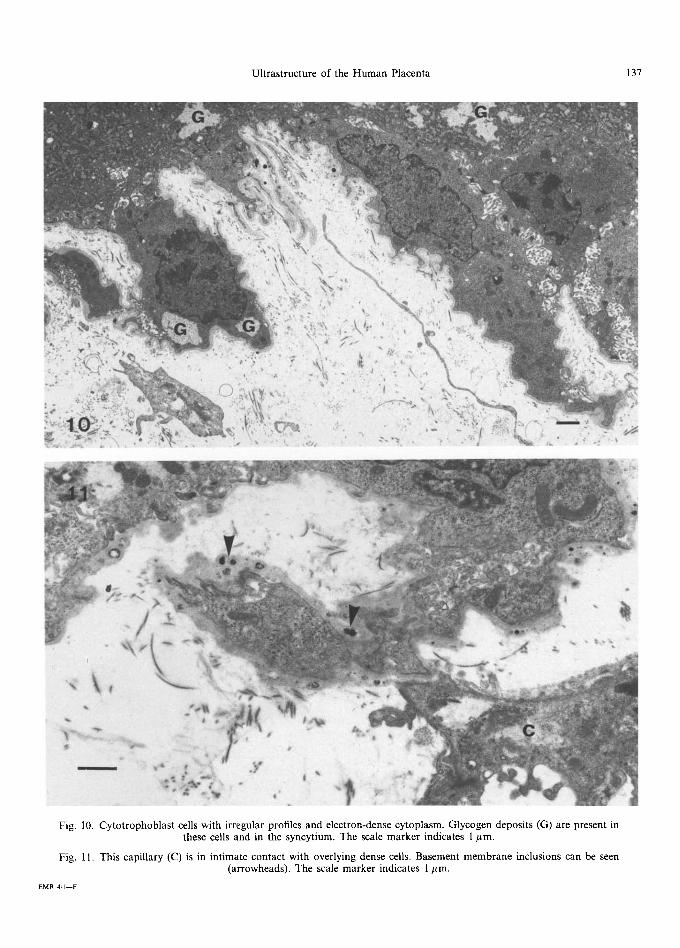
This third type of cell is the least common and its function has not been fully elucidated. It is an electron-dense cell which protrudes into the stroma, with an extremely irregular profile and many microvillous interdigitations where it is in contact with the main t rophoblast (Fig. 10). The nuclei resemble those of the syncytiotrophoblast, being electron-dense and crenated with clumps of
Ultrastructure of the Human Placenta 137
t
>
2 :~ . '
• )'--<.,,
t ' t,
.r ~ ,L
,, # f ~ i
ii
Fig. 10. Cytotrophoblast cells with irregular profiles and electron-dense cytoplasm. Glycogen deposits (G) are present in these cells and in the syncytium. The scale marker indicates 1 #m.
Fig. 11. This capillary (C) is in intimate contact with overlying dense cells. Basement membrane inclusions can be seen (arrowheads). The scale marker indicates 1 #m.
EMR 4/I--F

138 c . J .P . Jones and H. Fox
irregularly dispersed chromatin. The cytoplasm contains numerous electron-dense mitochondria and thin cisternae of rough endoplasmic reticulum with contents of moderate electron density. Free ribosomes are plentiful whilst several small vesicles and associated channels can be found. Clumps of cytoplasmic filaments are present whilst secretory droplets and occasional deposits of glycogen are also seen. Desmosomes are well developed between these cells and their neighbours and the intervening spaces are frequently occupied by the interdigita- tions of the cell membrane. Basement membrane material surrounds the projecting areas of the cells but does not extend between the interdigitations. These dense cells are often in intimate contact with the endothelial cells of the stromal capillaries (Fig. 11), being separated from them by a very thin strip of basement membrane; no evidence of true fusion between the two cell types has, however, been seen. Forms intermediate between all three cytotrophoblast cell types can be seen in first trimester material.
B. Second Trimester
By the second trimester there are fewer cytotro- phoblastic cells, the gaps between them are more prominent and many appear to be slightly flat- tened. Most are of the undifferentiated type but contain more organelles than in the first trimester, while the nuclei are more irregular in outline. At least one nucleolus is invariably present. Rough endoplasmic reticulum is extensively distributed throughout the cell and there appears to be an increase in the number of secretory droplets pre- sent compared with the first trimester. Golgi bodies are frequently seen, free ribosomes are numerous and nematosomes can occasionally be found. At this stage, no dense cells are found and inter- mediate cells are uncommon.
C. Third Tr imester
By term, cytotrophoblast cells (Fig. 12) are greatly reduced in number, although occasional mitotic figures can be seen indicating that tro- phoblastic growth, albeit slow, is still in progress. Nematosomes (Fig. 7) are more frequently encoun-
tered in the fully mature placenta. These organelles have been fully described by Jones and Ockleford (1985) and Ockleford et al. (1987) who detailed reports of their occurrence in rodent, baboon and human placentae, and noted that similar organelles can be found in neural and plant tissues. Serial sectioning shows that nematosomes have an aver- age profile of 0.45/~m, and may have various configurations including spirals, concentric rings, parallel arrays, random coils and paracrystalline structures. Ribonucleic acid appears to be a major constituent of this organelle, the exact function of which is, as yet, unknown.
In the term placenta, intermediate cells are rare and, when present, are at an advanced stage of differentiation so that they are indistinguishable from the syncytium except for the presence of a plasma membrane.
D. Funct ion
That the cytotrophoblast gives rise to the syn- cytiotrophoblast has been established by auto- radiography (Midgley et al., 1963; Tao and Hertig, 1965) and microspectrophotometry (Galton, 1962). Electron microscopy has, as noted above, confirmed the presence of differentiating cells having a cytoplasmic complexity intermediate between the undifferentiated cytotrophoblast cell, with its paucity of organelles, and the organelle- rich syncytiotrophoblast (Terzakis, 1963; Pierce and Midgley, 1963; Pierce et al., 1964; Jones, 1976), free membrane remnants bearing desmosomes from assimilated cytotrophoblast cells are found in the syncytium (Carter, 1964; Enders, 1965; Jones, 1976) and cytotrophoblastic cells with partial membrane breakdown are occasionally detected, as seen in Fig. 8 (Knoth, 1968; Jones, 1976). The presence of various types of cellular junctions on the maternal surface of the syncytium has also been taken as evidence of the multicellular cyto- trophoblastic origin of the syncytiotrophoblast (Cavicchia, 1971; Metz et al., 1979; Reale et al., 1980; Wang and Schneider, 1987).
The evidence that the cytotrophoblast is largely a germinative zone of undifferentiated stem cells is therefore overwhelming (Pierce et al., 1964; Boyd and Hamilton, 1970) but several workers have

Ultrastructure of the Human Placenta 139
postulated that these cells may also play a role in hormone synthesis, largely because the rise and subsequent decline in the number of cells parallels the level of chorionic gonadotropin (hCG) pro- duction (Wislocki and Bennett, 1943; Lister, 1963b). It has further been argued that the large mitochondria, well-developed Golgi bodies and potentially active endoplasmic reticulum in these cells indicate a capability for endocrine activity (Hashimoto et al., 1960b; Lister, 1963b; Nagy et aL, 1965; Garancis et aL, 1970; Dec and Bartel, 1976). However, the development of sensitive immunohistochemical techniques has shown that hCG is mainly localised in the syncytium (Morinaga et al., 1984; Morrish et al., 1987, 1988; Hamasaki et al., 1987; Suemizu et al., 1988). Nevertheless, Hoshina et al. (1982) demonstrated positive staining for human chorionic gonado- tropin (hCG) in the cisternae of rough endoplasmic reticulum and perinuclear spaces of differentiating intermediate cytotrophoblastic cells and in situ hybridisation studies (Hoshina et al., 1985) have shown the presence of hCG alpha and beta messen- ger RNAs in occasional cytotrophoblastic cells. In general, it appears that only cells that are progress- ing along the differentiation pathway towards a syncytial phenotype (i.e. intermediate cells) are able to synthesise these hormones. The genes for the different hormones are activated at different stages of placental differentiation, with hCG alpha and beta subunit expression dependent on the presence of proliferating cytotrophoblast while human placental lactogen (hPL) expression is sus- tained in differentiated syncytium (Hoshina, 1983; Kliman et al., 1987). The cytotrophoblast also synthesises, in part at least, laminin, one of the constituents of the trophoblastic basement mem- brane, this substance being detected, by immuno- cytochemical techniques, in the rough endoplasmic reticulum of these cells (Amenta et al., 1986; Leivo et al., 1989).
IV. THE SYNCYTIOTROPHOBLAST
A. Microv i l lous Sur face
In the first trimester, the syncytium, which is of a fairly uniform thickness, often has a charac-
teristic scalloped outline (Sawasaki et al., 1957; Boyd et al., 1968b; Hamilton and Boyd, 1970; Jones, 1976) created by the presence of syncytial projections (Fig. 13) which are generally covered by slender microvilli. This microvillous surface forms the interface between maternal blood and fetal tissues and, as such, is vitally concerned with absorption of nutrients, transfer of oxygen, immunological protection, excretion of waste products and secretion of hormones. A detailed discussion of the biochemistry of this unique mem- brane is beyond the scope of this review and the reader is referred to the writings of Whyte (1983), Wild (1983), Truman and Ford (1984) and Webb et al. (1987).
1. S t ruc ture
The outer surface of the syncytium is covered by microvilli which form a brush border; this was first described by Kastschenko (1885), rather charmingly, as a covering of "Wimpern", which translates literally as "eyelashes". These microvilli alter in height during the course of gestation reaching their maximum height at 10-11 weeks' gestation and then subsequently shortening (Ikawa, 1959; Tighe et al., 1967; Burton, 1987). They are mainly cylindrical but sometimes spatulate or clavate at their tips: branching mi- crovilli have been described by Bergstr6m (1971), Ferenczy and Richart (1972) and Fox and Agrafojo-Blanco (1974) in early pregnancy. Jones (1976) found the distribution of clavate as opposed to rod-shaped forms to be quite variable at all stages of pregnancy. Scanning electron microscopy has also shown the microvilli to be arranged in an irregular honeycomb pattern in early pregnancy (Burton, 1987), giving rise to the characteristic 'scalloping' profile seen in thin sections (Boyd et al., 1968b; Jones, 1976).
Microvilli normally form a continuous covering over the chorionic villi except where there is syncy- tial damage (Burton, 1987), although there is a depletion in numbers over vasculo-syncytial mem- branes (Fox and Agrafojo-Blanco, 1974; Jones, 1976), those areas where sinusoidally dilated capil- laries bulge into the trophoblast and the syncytial covering is anuclear and thin. There also appears
140 C. J, P. Jones and H. Fox
°s',l . ° p
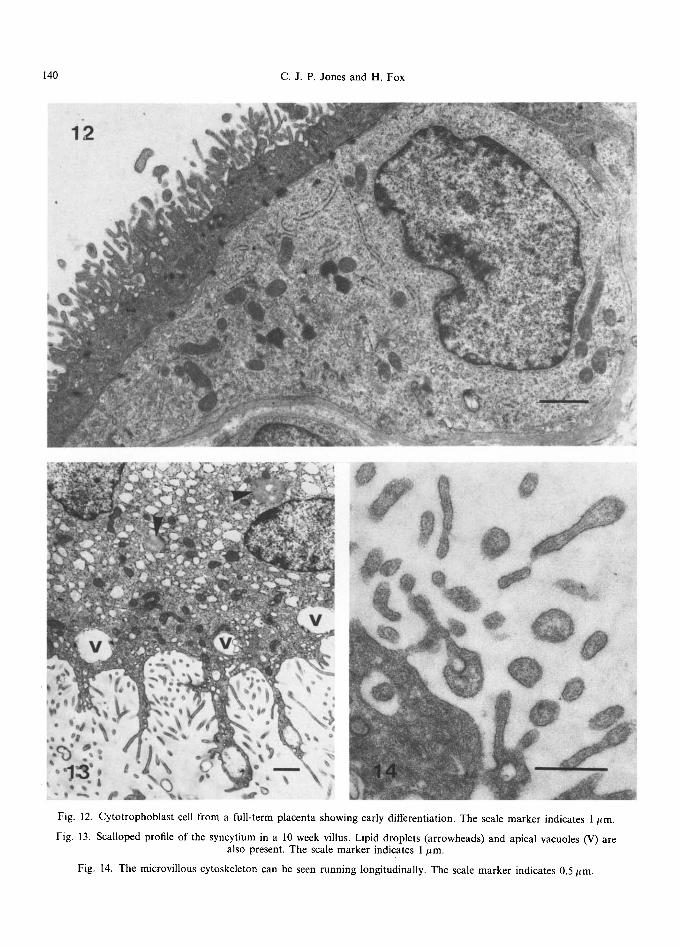
Fig. 12. Cytotrophoblast cell from a full-term placenta showing early differentiation. The scale marker indicates 1/~m.
Fig. 13. Scalloped profile of the syncytium in a 10 week villus. Lipid droplets (arrowheads) and apical vacuoles (V) are also present. The scale marker indicates 1/tm.
Fig. 14. The microvillous cytoskeleton can be seen running longitudinally. The scale marker indicates 0.5 #m.

Ultrastructure of the Human Placenta 141
to be a functional relationship between the pres- ence of microvilli and pinocytosis as the incidence of invaginations is markedly reduced in areas with depleted brush borders (Boyd and Hughes, 1954; Boyd et al., 1968b; Knoth, 1968; Jones, 1976; Jones and Fox, 1977).
Some degree of internal structure can be seen (Fig. 14) in favourably orientated thin sections of microvilli (Terzakis, 1963; Tighe et al., 1967; Hamilton and Boyd, 1970). Actin-like filaments, 6-9nm in diameter, have been described (Ockleford and Whyte, 1977; Ockleford et al., 1981; Ockleford and Wakely, 1982) and actin has been demonstrated by immunocytochemistry (Faulk and Johnson, 1977; Ockleford and Wakely, 1982; Beham et al., 1988), biochemically (Whitsett and Lessard, 1978; Truman and Ford, 1984, 1986a) and by myosin (SI) decoration (King, 1983). Calcium-dependent binding pro- teins, associated with a submembranous micro- villous cytoskeleton, are also present (Webb and Mahadevan, 1987) as well as alpha-actinin and ezrin (Edwards and Booth, 1987). The microvilli appear to be quite different from those found on intestinal or renal brush borders; they are more dynamic and are involved in receptor-mediated endocytosis (Ockleford and Whyte, 1977; Edwards and Booth, 1987).
No evidence was found by Jones (1976) of the production of excretory vacuoles by microvilli as described by Salazar and Gonzalez-Angulo (1967) although clavate forms were common, and a cer- tain degree of electron-lucidity was seen in the cores of the bulbous areas, within a circlet of supporting microfilaments. Experimental work on the choroid plexus, which bears clavate microvilli very similar to those seen in the placenta, indicates that excretion does not occur by microvillous blebbing but via small vesicles in the case of proteins, and intercellular clefts in the case of salts and water (Brightman, 1975; Tennyson, 1975).
2. Glycosarninoglycans and pro teog lycans
The microvilli bear a well-developed 'glycocalyx' which has been the subject of many histochemical (Bradbury et al., 1969, 1970; Rovasio and Monis, 1973; Liebhart, 1974a; Martin et al., 1974; Nelson
et al., 1975; Whyte et al., 1978; Whyte, 1980; King, 1981; Lee and Damjanov, 1984; Jones et al., 1990) and biochemical studies (Whyte and Loke, 1978; Whyte, 1983; Webb et al., 1987). According to Rovasio and Monis (1973), its staining character- istics suggest the presence of 'acid mucopoly- saccharides'. In modern terminology, this would be glycosaminoglycan, possibly with a glycoprotein containing sialic acid. If the glycosaminoglycan were keratan sulphate, the sialic acid could be part of a side-chain associated with the linkage sequence of the glycosaminoglycan and its core protein. Rovasio and Monis thought that the glycoproteins may be linked to the presence of human chorionic gonadotropin (hCG) which has also been found on microvilli (Amoroso and Perry, 1975; Dreskin et al., 1970; Hamanaka et al., 1971; De Ikonicoff, 1973). Other workers have found progesterone bound to microvilli (Smith and Brush, 1973; Smith et al., 1974), which they thought might mask the antigenicity of the placenta, a function also ascribed to h C G by Amoroso and Perry. Martin et al. (1974) concluded from their many histochemical experiments, that the glycocalyx was composed of hyaluronic acid as opposed to the highly sulphated 'acid muco- polysaccharides' that were proposed by Bradbury et al. (1969, 1970), and suggested that it provided an optimum microenvironment for transport mechanisms as well as preventing the adherence of immunologically active lymphocytes.
Martin et aL (1974) and Whyte (1980) also agreed that hCG might be secreted onto the maternal surface of the syncytium, as demon- strated by lectin-binding techniques. King (1981) described anionic sites that were neuraminidase- resistant and pointed out that both hCG and alkaline phosphatase, which are associated with the microvilli, contain sialic acid. Recent bio- chemical studies have shown that the microvillous surface contains highly glycosylated and highly sialylated glycosphingolipids of distinctive types (Cheng and Johnson, 1985), while lectin histo- chemistry has demonstrated the presence of several classes of N-linked oligosaccharides (Jones et al., 1990). The presence of abundant sialyl residues was confirmed by Jones et al. (1990) both on cell surfaces and within granules, and sub-terminal

142 c.J .P. Jones and H. Fox
galactosyl residues were also demonstrated. King and Loke (1988) have identified Le a antigen expression in syncytiotrophoblastic cells at the maternofetal interface, but found no evidence for sialyl Le a. In contrast, they found sialyl Le X antigen in extravillous trophoblastic cells in normal and molar pregnancies, and in choriocarcinoma. These results clearly indicate subtle and specific vari- ations in the patterns of expression of fucosylated oligosaccharides in these tissues.
3. E n z y m e s
The microvillous surface is richly endowed with enzymes. Alkaline phosphatase activity (Figs 8 and 15) is present in discrete foci along the surface of the membrane of term placentae (Lister, 1967; Hempel and Geyer, 1969; Martin et al., 1974; Jones and Fox, 1976; Jemmerson et al., 1985) although Hulstaert et al., (1973) were exceptional in finding the localisation to be continuous. The distribution of this enzyme in the immature placenta may be very variable (Jones, 1976; Jones and Fox, 1976) with only some specimens showing activity; this discrepancy i~ probably due to the fact that alka- line phosphatase activity tends to diminish towards the middle of the first trimester and only reappears again in the middle of the second trimester (McKay et al., 1958; Kedzia and Pilarski, 1971).
Various forms of alkaline phosphatase have been isolated from the placenta (Webb et al., 1986) and suggestions as to its function include mem- brane transport mechanisms (Jones, 1976; Truman and Ford, 1984), phosphoprotein phosphatase ac- tivity and a contribution to the regulation of cell growth by selective tyrosyl dephosphorylation (Risk and Johnson, 1985; Webb et al., 1987). Other enzymes found on the microvilli include 5'-nucle- otidase (Smith et al., 1977; Booth et al., 1980), neutral arylaminidase and galactosyl and sialyl transferases (Whyte, 1983; Khalfoun et al., 1986) together with steroid sulphatases (Guller et al., 1986).
4. Recep tors
The range of receptors found on the microvillous membrane has been reviewed by Wild (1983) and Truman and Ford (1984) and includes those for
epidermal growth factor (Richards et al., 1983; Magid et al., 1985; Rao et al., 1985; Kawagoe et al., 1990), insulin (Haour and Bertrand, 1974; Whitsett and Lessard, 1978), native and acetylated- low-density lipoprotein (Alsat et al., 1984, 1987; Malassin6 et al., 1987), Fc fragment of IgG (Matre et al., 1975; Wild, 1981; Brown and Johnson, 1981; Pearse, 1982) and transferrin (Wada et al., 1979; Loh et al., 1980; Truman and Ford, 1986b; Bierings et al., 1988, 1990; Turkewitz and Harrison, 1989). These receptors tend to cluster in specialised regions of the surface membrane called coated pits (Goldstein et al., 1979; Turkewitz and Harrison, 1989) that invaginate into the cell to form coated vesicles which have a characteristic structure and protein composition (Ockleford and Whyte, 1977; Pearse, 1982; Wild, 1980). It has been suggested (Ockleford and Whyte, 1977; Ockleford and Menon, 1977) that one of the main functions of syncytial microvilli is to expose a large surface area of membrane containing receptors which are then moved laterally towards the differentiated regions which constitute the coated pits; the pres- ence of subplasmalemmal actin filaments may be related to this phenomenon.
5. C o a t e d vesicles
Coated vesicles have, as their name suggests, a very characteristic appearance in the electron microscope (Fig. 16). Also known as acantho- somes, bristle-coated vesicles and rhopheosomes (Wild, 1980), their occurrence has been linked to the process of selective endocytosis whereby extra- cellular materials are taken up by cells. They can be distinguished from smooth-walled micropinocy- totic vesicles by the coating on the cytoplasmic aspect of the vesicle membrane, which appears as radially arranged projections (Fig. 16) each about 5 nm in width, that form the walls of a 'basket' of regular hexagons and pentagons (Kanaseki and Kadota, 1969; Ockleford, 1976, 1981). Isolation of coated vesicles has enabled the major constituent of the 'coat' to be characterised--a protein of molecular weight 180,000 which has been named 'clathrin' (Pearse, 1976) after the lattice-like structure that it forms. Coated vesicles in the placenta have diameters ranging from 70 to 120 nm
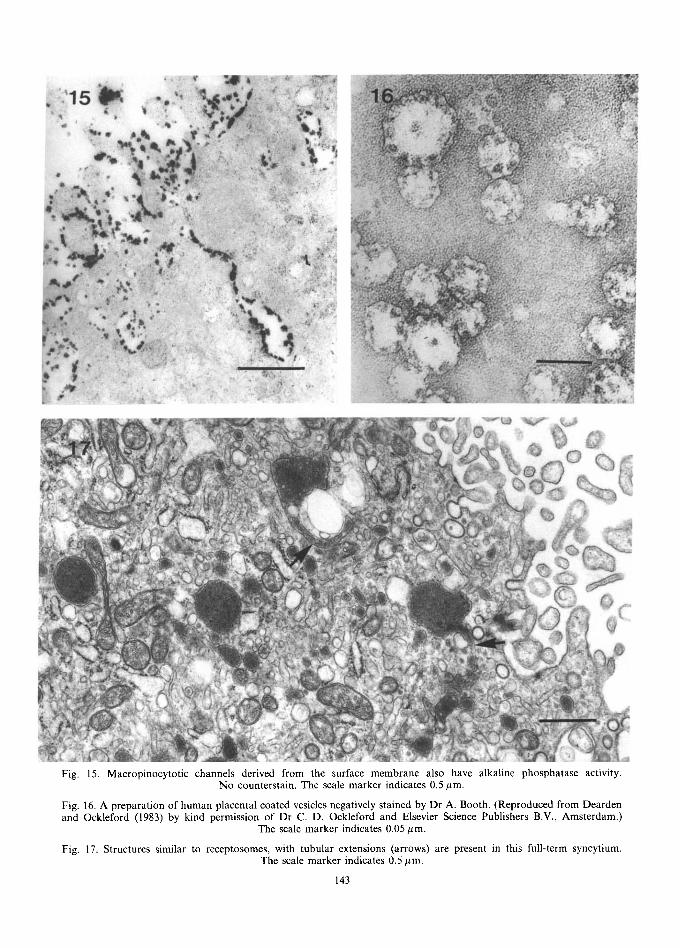
Fig. 15. Macropinocytotic channels derived from the surface membrane also have alkaline phosphatase activity. No counterstain. The scale marker indicates 0.5 #m.
Fig. 16. A preparation of human placental coated vesicles negatively stained by Dr A. Booth. (Reproduced from Dearden and Ockleford (1983) by kind permission of Dr C. D. Ockleford and Elsevier Science Publishers B.V., Amsterdam.)
The scale marker indicates 0.05/~m.
Fig. 17. Structures similar to receptosomes, with tubular extensions (arrows) are present in this full-term syncytium. The scale marker indicates 0.5 #m,
143

144 c.J.P. Jones and H. Fox
(Ockleford and Whyte, 1977) and their distribution is linked to that of the syncytial microvilli. They occur mainly at or near the cell surface; the fact that the majority are in direct contact with the surface membrane suggests that the process of vesicle formation is slow compared with the rate of their subsequent movement away from the cell surface (Ockleford and Whyte, 1977). The inner face of the coated vesicles is lined by an electron- dense layer which stains with Ruthenium red and Alcian blue, indicating the presence of an abun- dant glycocalyx related to its function as a site of selective protein uptake and reflecting the presence of receptors composed of molecules with promi- nent saccharide portions (Whyte, 1980; Ockleford, 1981). The vesicle actually forms by the gradual indentation of a specialised region of the plasma membrane, the curvature of which increases to form a flask-shaped structure which is eventually pinched off to form a free vesicle. The fate of coated vesicles in the human placenta is not clear. It has been shown that the clathrin lattice may be lost by vesicles (Kanaseki and Kadota, 1969) while Wild (1975) proposed that the clathrin coat pre- vented subsequent fusion with lysosomes, thus protecting the vesicle contents from proteolytic degradation. In many systems, the internalised protein is rapidly broken down (Goldstein et al., 1979) whereas in others the contents are directed to specific subcellular organelles. The mechanism for sorting the various proteins is not, at present, understood although it has been suggested that, in some systems, the macromolecular ligands internalised via coated pits are delivered to a common intracellular compartment--the endosome (Hopkins, 1986), receptosome (Willingham and Pastan, 1980) or CURL (compartment of un- coupling of receptor and ligand) (Geiaze et al., 1982). This compartment has a relatively low pH maintained by an ATP-dependent proton pump, and within this acid environment ligands become uncoupled from their receptors which may then be recycled, with the vesicle membrane being re- incorporated into the wall for subsequent reloca- tion. It is possible that a similar pump may operate within the coated vesicle itself as such a mechanism has been demonstrated in vesicles isolated from calf brain, to acidify the endocytosed material as
soon as it has been internalised (Forgac et al., 1983). In the case of transferrin, iron is dissociated from the transferri~receptor complex at low pH, and the complex is recycled to the cell surface where apo-transferrin is dissociated from the receptor at pH 7.0 (Dautry-Varsat et al., 1983). The endosome may have its syncytiotrophoblastic equivalent in the multivesicular body (Fig. 17) (Ockleford and Dearden, 1984) which has been shown to have an acid microenvironment (Martin et al., 1974; Jones and Fox, 1976).
6. Endocytic /macropinocytotic channels
Apart from the coated vesicles at the base of microvilli and in the apical cytoplasm, many endo- cytic or macropinocytotic channels can be seen, especially in the first half of gestation. These have been linked with non-selective uptake of materials (Wild, 1975). Their derivation from the maternal surface has been shown by their positive staining for alkaline phosphatase activity (Jones, 1976; Fig. 15); in some cases they can be seen to form tortuous channels leading from the surface of the syncytium down to the trophoblastic basement membrane, as shown by thorium labelling (Ashley, 1965) and alkaline phosphatase activity. These may be particularly prominent in damaged syn- cytium where cell turnover is high (Jones, 1976).
7. Surface junctional complexes
A feature of the maternal surface of the syn- cytium is the presence of cellular junctions (Fig. 18) such as desmosomes, desmosome-like junctions and tight junctions (Burgos and Rodriguez, 1966; Cavicchia, 1971; Metz et al., 1979; Reale et al., 1980; Wang and Schneider, 1987). These are gener- ally considered to be remnants of the desmosomes originally connecting the cytotrophoblastic cells to the syncytium although Metz and colleagues (1979) thought that they may represent remnants of intervillous contact areas. Similar membrane remnants may be found elsewhere in the syncytium as can also intermediate cells showing partial dissolution of their plasma membranes (Fig. 8) as they become incorporated into the main syn- cytium (Carter, 1964; Pierce et al., 1964; Enders, 1965).
Ultrastructure of the Human Placenta 145
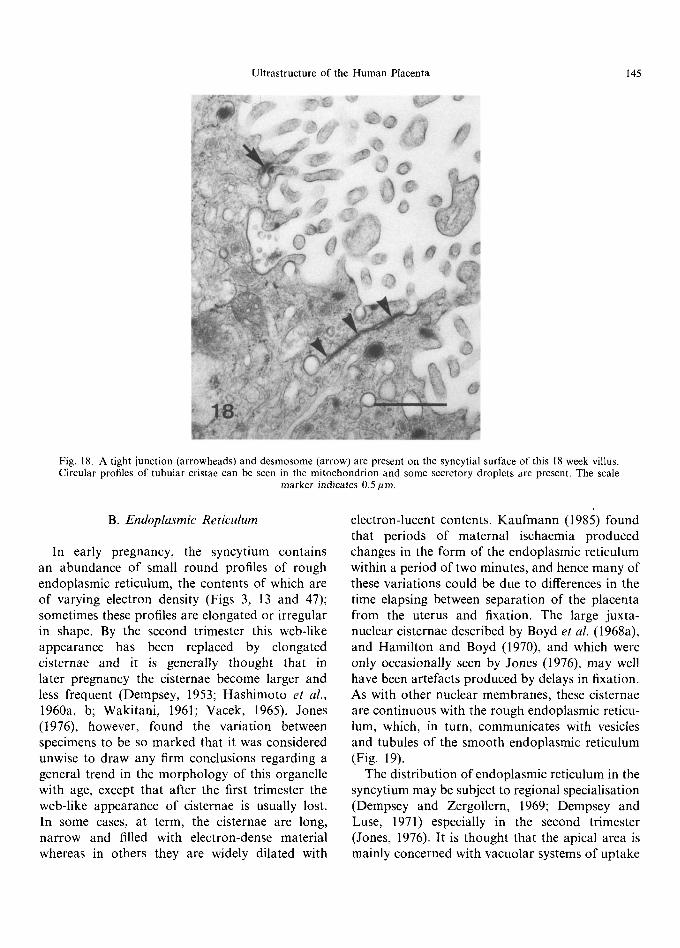
Fig. 18. A tight junction (arrowheads) and desmosome (arrow) are present on the syncytial surface of this 18 week villus. Circular profiles of tubular cristae can be seen in the mitochondrion and some secretory droplets are present. The scale
marker indicates 0.5/~m.
B. Endoplasmic Reticulum
In early pregnancy, the syncytium contains an abundance of small round profiles of rough endoplasmic reticulum, the contents of which are of varying electron density (Figs 3, 13 and 47); sometimes these profiles are elongated or irregular in shape. By the second trimester this web-like appearance has been replaced by elongated cisternae and it is generally thought that in later pregnancy the cisternae become larger and less frequent (Dempsey, 1953; Hashimoto et al., 1960a, b; Wakitani, 1961; Vacek, 1965). Jones (1976), however, found the variation between specimens to be so marked that it was considered unwise to draw any firm conclusions regarding a general trend in the morphology of this organelle with age, except that after the first trimester the web-like appearance of cisternae is usually lost. In some cases, at term, the cisternae are long, narrow and filled with electron-dense material whereas in others they are widely dilated with
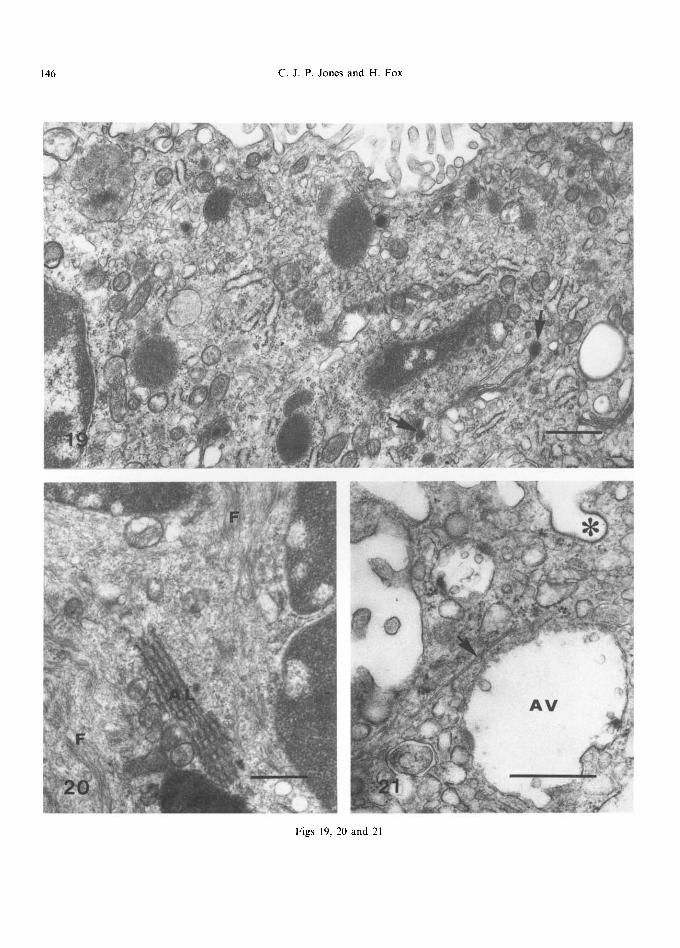
electron-lucent contents. Kaufmann (1985) found that periods of maternal ischaemia produced changes in the form of the endoplasmic reticulum within a period of two minutes, and hence many of these variations could be due to differences in the time elapsing between separation of the placenta from the uterus and fixation. The large juxta- nuclear cisternae described by Boyd et al. (1968a), and Hamilton and Boyd (1970), and which were only occasionally seen by Jones (1976), may well have been artefacts produced by delays in fixation. As with other nuclear membranes, these cisternae are continuous with the rough endoplasmic reticu- lum, which, in turn, communicates with vesicles and tubules of the smooth endoplasmic reticulum (Fig. 19).
The distribution of endoplasmic reticulum in the syncytium may be subject to regional specialisation (Dempsey and Zergollern, 1969; Dempsey and Luse, 1971) especially in the second trimester (Jones, 1976). It is thought that the apical area is mainly concerned with vacuolar systems of uptake
146 C . J . P . Jones and H. Fox
Figs 19, 20 and 21

Ultrastructure of the Human Placenta 147
and digestion and that the middle zone, which contains a higher proport ion of endoplasmic reticulum, is the main site of biosynthetic activity. A specialised form of endoplasmic re t icu lum-- annulate lamellae--is occasionally found in the syncytial cytoplasm, especially in areas rich in 10 nm intermediate filaments (Fig. 20). Jones and Fox (1977) considered them to be involved in the synthesis of these filaments as annulate lamellae appear to be associated with protein biosynthesis (Wischnitzer, 1970; Benzo, 1974).
C. Ribosomes
Free ribosomes are plentiful in the syncytium at all stages of pregnancy and are responsible, to a large extent, for the increased electron density of the syncytium over that of the cytotrophoblast . Also present, and of very similar size, are oc- casional masses of glycogen (Fig. 10), especially in the first trimester of pregnancy (Terzakis, 1963; Dempsey and Luse, 1971). These are less easy to identify later in pregnancy as their presence is obscured by the density of cytoplasmic organelles.
D. Mitochondria
Mitochondria are numerous, round, ovoid or branching, more electron-dense and smaller than those found in the cytotrophoblast (Hamilton and Boyd, 1970; Mart in and Spicer, 1973a). Tubular cristae, of the type characteristic of steroidogenic cells, may be present (Terzakis, 1963; Tighe et al., 1967) and are prevalent from the end of the second trimester to term (Fig. 18) (Wynn, 1972). Kaufmann and Stegner (1972), Wakitani (1961), Lister (1963a) and Demir (1980) thought that syncytial mitochondria are more numerous in the first than in the third trimester, while Panigel and
Anh (1964a) found that the cristae of syncytial mitochondria became blurred at term. Haigh et al. (1989), using a computerised image analysis sys- tem, made a morphometr ic study of syncytial mitochondria and found that their numerical den- sity increased throughout gestation. Their mean volume, however, declined, and hence there was no significant difference in the proport ion of the syn- cytial cytoplasm occupied by these organelles. Malate and lactate dehydrogenase activity is pre- sent in syncytial mitochondria (Jones and Fox, 1978), the highest level of activity being found in first trimester specimens with a significant decrease at term (Figs 5 and 6).
E. Vacuoles, Multivesicular and Dense Bodies
In the syncytium, a large number of different vacuoles and dense bodies are found (Terzakis, 1963; Martin and Spicer, 1973a). Near the apical surface are large, round vacuoles, about 70~1000 nm in diameter at term but sometimes as large as 1800 nm in the immature placenta (Figs ! 3 and 21); these have either diffuse or flocculent contents but sometimes contain a number of small vesicles, especially early in pregnancy. On occasion, fusion of these vacuoles with other small vesicles can be seen. Ovoid or round masses, often up to 1200 nm in length, with contents of medium electron density can also be present-- these may be analagous to the light multivesicular bodies of Martin and Spicer (1973a) although vesicles may not always be evident within them. Also noted are irregularly-shaped, electron-dense, osmiophilic masses, up to about 1200nm long, bound by a limiting membrane, and which occasionally appear to be multivesicular (Fig. 19); some of these may correspond to the dense multivesicular bodies de- scribed by Martin and Spicer (1973a), others may
Fig. 19. Various multivesicular and dense bodies are present in this term villus, and secretory droplets associated with Golgi bodies (arrows). The complexity of smooth and rough endoplasmic reticular channels and vesicles can be appreciated.
The scale marker indicates 0.5 #m.
Fig. 20. Masses of 10 nm cytoplasmic filaments (F) and annulate lamellae (AL) are associated with this syncytial knot. The scale marker indicates 0.5 #m.
Fig. 21. An apical vacuole and vesicles (AV), a coated vesicle at the base of a microvillus (*) and a microtubule (arrow) can be seen. The scale marker indicates 0.5 ~m.

148 C.J.P. Jones and H. Fox
form aggregates within the syncytium and be associated with the presence of hormones (vide infra). Dense bodies, smaller than multivesicular bodies (about 400 nm in diameter) have been described by Martin and Spicer (1973a) who proposed that they were residual multivesicular bodies; experiments with tracers have certainly indicated a pathway from coated pits and endo- cytic channels through to multivesicular bodies and finally to dense bodies (King and Enders, 1971; Lin, 1980; King, 1982b; Sideri et al., 1987). The fate of the dense bodies is not clear; Lin (1980) described discharge of the contents of residual bodies through exocytosis via the basal plasma membrane, but Sideri et al. (1987) found only a very rare example of tracer close to the syncytial basement membrane. King (1982b) also failed to find fusion of vesicles containing label with the basal surface. It is possible that some of the apical vacuoles discharge their contents into the inter- villous space and so function as a type of residual body; it seems unlikely that dense bodies are retained in the trophoblast as there is no accumu- lation of them in the term placenta and lipofuscin pigment is absent (Haigh et al., 1984).
That multivesicular bodies and dense bodies are associated with enzymatic breakdown of material is indicated by their acid phosphatase activity (Martin and Spicer, 1973a; Jones and Fox, 1976); such activity is also present in small round or elongated inclusions thought to be primary lyso- somes (Fig. 22) (Jones and Fox, 1976). These findings are paralleled by sedimentation velocity analysis (Corash and Gross, 1974) by which two lysosome fractions were separated out. Acid phos- phatase activity is generally most evident in the first half of gestation where it is thought to be concerned not only with degrading the contents of multivesicular bodies (Friend and Farquhar, 1967) but also with the considerable architectural
refashioning that occurs in the developing placenta (Corash and Gross, 1974; Jones and Fox, 1976). These lysosomal structures are also thought to play a part in the breakdown of the cytotro- phoblast cell membrane during the formation of syncytiotrophoblast (Contractor et al., 1977).
Aryl sulphatase activity has been demonstrated in the trophoblast (Jones, 1976; Contractor et al., 1977), although the number of positively-stained bodies is lower than that showing acid phosphatase activity; the pattern is, however, similar, with maximum activity in the first trimester (Fig. 23).
F. Lipid Droplets
Syncytial osmiophilic lipid droplets (Figs 13 and 47) are particularly prominent in the first trimester of pregnancy (Sawasaki et al., 1957; Lister, 1963a, Anderson and McKay, 1966; Tighe et al., 1967), generally have a smooth round profile and measure about 50(~1500 nm in diameter. They do not have a limiting membrane and their contents are usually homogeneous, though electron-lucent islands may be present where the fat has leached away (Fig. 47). Terzakis (1963) and Strauss et al. (1965) thought they represented nutritional lipid in the process of being transferred from mother to fetus whereas Wislocki and Dempsey (1955), Tighe et al. (1967) and Jones (1976) suggested that their presence was related to steroid biosynthesis, representing a lipid 'pool' of precursors.
G. Secretory Droplets /Golgi Apparatus/Hormone Biosynthesis
Small, electron-dense, osmiophilic, membrane- bound granules, 80-350 nm in diameter, are fre- quently in evidence at all stages of gestation and are probably secretory droplets (Figs 18 and 19). There is usually a well-defined space between the
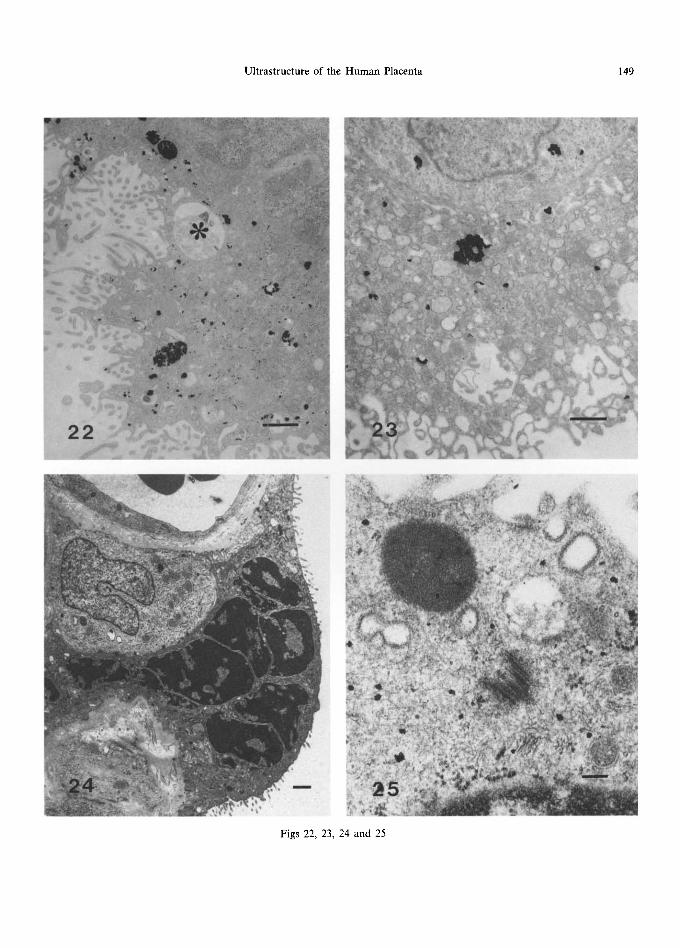
Fig. 22. Acid phosphatase activity in a 16 week villus. An apical vacuole is unreactive (*) while other inclusions show intense deposition of reaction product. No counterstain. The scale marker indicates 1 ktm.
Fig. 23. Aryl sulphatase activity in this 10 week villus is restricted to some primary lysosomes and occasional multivesicular and dense bodies. No counterstain. The scale marker indicates 1/~m.
Fig. 24. Aggregated nuclei forming a syncytial knot in a full-term villus. The scale marker indicates 1 ~m.
Fig. 25. A residual centriole in the apical part of the syncytium. The scale marker indicates 0.1 ~tm.
Ultrastructure of the Human Placenta 149
22.:"'~
Figs 22, 23, 24 and 25

150 c.J.P. Jones and H. Fox
limiting membrane and the granule itself (Rhodin and Terzakis, 1962; Anderson and McKay, 1966); they are often associated with Golgi bodies, which are generally small in the syncytium with short, plump cisternae. Golgi bodies may be difficult to discern amongst the plethora of smooth-surfaced vesicles and vacuoles of the syncytium, especially in term placentae (Wakitani, 1961; Ashley, 1965; Hamilton and Boyd, 1970). Granules with lamel- lar, periodic inclusions, as described by Terzakis (1963), may also occasionally be seen, especially in the first trimester.
Many of these granules have been linked with hormone production. Immunocytochemical techniques have indicated that small secretory granules, 100-300 nm in diameter, and large, irreg- ularly-shaped granule complexes, 400-1200nm in size, contain human chorionic gonadotropin (hCG) in first trimester syncytial tissue (Morinaga et al., 1984; Morrish et al., 1987, 1988; Hamasaki et al., 1987; Suemizu et al., 1988). Hamasaki et al. (1987) and Suemizu et al. (1988) considered the granule complexes as sites for the storage and/or digestion of free excess hormone, especially beta- subunits, in the early placenta. Fusion of secretory products with lysosomes has been described as a regulatory phenomenon (crinophagy) by Farquhar (1969) in the pituitary, and possibly these bodies correspond to the acid phosphatase positive dense multivesicular bodies of Martin and Spicer (1973a); certainly the electron micrographs of Morrish et al. (1987) imply a degree of variability in the electron density, or vesiculation, within the hCG-positive structures. Anti-hCG staining of granules has not been seen in term placenta (Morrish et al., 1987, 1988) but a population of small granules, about 80-250 nm in diameter, has been found in the apical portions of the syncytium at term, sometimes associated with Golgi bodies, which shows human placental lactogen (hPL) activity (Fujimoto et al., 1986; Morrish et al., 1988). These were absent from early placenta and their numbers were found to increase during the course of gestation. These find- ings--a preponderance of hCG-positive granules in early pregnancy, falling off towards term, and an increase in hPL-positive granules with increasing gestational age, parallels changes in maternal hor- mone levels (see Pattillo et al. (1983) for a review).
Apart from secretory granules, other sites have been implicated in syncytial hormone biosynthesis. Cis t e rnae of rough endoplasmic reticulum were shown to contain hCG activity by Dreskin et al. (1970), Hamanaka et al. (1971), de Ikonicoff (1974), Amoroso and Perry (1975), Morinaga et al. (1984), Johnson and Wooding (1988) and Suemizu et al. (1988), while reaction product in the peri- nuclear space was also described by many of the above workers. De Ikonicoff (1973, 1974) described hPL and hCG activity on ribosomes and membranes of the rough endoplasmic reticulum and also on the surface of microvilli, but eliminated localisation to the nuclear membrane, which she attributed to a non-specific reaction, by working at a pH below 7.4. hCG was also localised to the surface of microvilli by Dreskin et al. (1970), Hamanaka et al. (1971) and Amoroso and Perry (1975) and, to a much lesser extent, on the basal plasma membrane of the syncytium (Dreskin et al., 1970). This has led various workers to propose a non-classical model of secretion of the hormone i.e. packaging into granules by the Golgi apparatus is by-passed and the hormone is secreted by fusion of the rough endoplasmic reticulum directly with the apical plasma membrane (Dreskin et al., 1970; Yorde et al., 1979; Morinaga et al., 1984). Morrish et al. (1987), however, attributed these findings to low sensitivity of the methodology and poor fixation protocols, as they were able to demonstrate numerous secretory granules undergoing exo- cytosis in the manner of the conventional secretory pathway.
Recent molecular biology studies have con- firmed the immunocytochemical findings, with the genes for hCG alpha and beta subunits, and hPL, being activated at different stages of placental differentiation (Hoshina et al., 1982, 1983, 1985). The hCG alpha gene is activated after the for- mation of committed (as opposed to undifferenti- ated) cytotrophoblasts, while the beta subunit is expressed at the intermediate cell stage, which thus elaborates both subunits. During the course of gestation, the hCG alpha-mRNA decreases six- fold, while hCG beta decreases to barely detectable levels in term placenta, hPL mRNA is only ex- pressed in fully differentiated syncytial tissue and

Ultrastructure of the Human Placenta 151
continues to be transcribed at a fairly constant rate throughout gestation.
H. Nuclei
The nuclei within the syncytiotrophoblast have more condensed chromatin than do their counter- parts in the cytotrophoblast (Figs 1 and 24) and are also rather irregular in outline. Their chromatin tends to be aggregated beneath the nuclear mem- brane except immediately subjacent to the nuclear pores where electron-lucent areas are evident. Nucleoli are frequently present, and changes in their ultrastructure, and that of the nuclei, with maturation of the placenta, have been described by Martin and Spicer (1973b). In the first trimester, the nuclei are fairly evenly spaced; however, the formation of anuclear vasculo-syncytial mem- branes later in pregnancy tends to cause aggre- gation of nuclei into so-called syncytial knots (Fig. 24), the ultrastructure of which has been described in detail by Jones and Fox (1977). Within these knots, the profiles of nuclei tend to mould into each other so that only a narrow, even strip of cytoplasm is left between them; the nuclei themselves may become extremely pyknotic, with smooth regular outlines closely apposed to each other, to the extent that sometimes fusion of adjoining nuclear membranes may occur.
I. Cytoskeleton
Cytoskeletal elements within the syncytium have been demonstrated at the light microscopic level using immunofluorescence; cytokeratins including 8, 18 and 19 (Khong et al., 1986; Beham et al., 1988; Bradbury and Ockleford, 1990) and keratin polypeptides 40, 45, 52 and 54 K have been found in both cyto- and syncytiotrophoblast, while anti- serum to desmoplakin, which recognises desmo- somes, binds to areas between the cyto- and syncytiotrophoblast as well as to regions on the apical plasma membrane where cell junctions may be found (Burgos and Rodriguez, 1966; Cavicchia, 1971; Metz et al., 1979; Reale et al., 1980; Wang and Schneider, 1987). Bundles of intermediate filaments 10 nm in diameter have been described in the syncytium, especially around nuclei (Martin
and Spicer, 1973b) and in the apical cytoplasm of the syncytium (Lister, 1963a); their number may be greatly increased in the cytoplasm around syncytial knots where thick filament bundles lie between the aggregated nuclei (Fig. 20), often orientated in a plane parallel to the nuclear membrane. Jones and Fox (1977) suggested that such bundles provide mechanical support, especially where syncytial knots fuse to form intervillous bridges, thus forming an internal strut system as originally proposed by H6rmann (1953).
Tubulin can be identified in the apical regions of the syncytium using immunofluorescence (Ockleford et al., 1981; Ockleford and Wakely, 1982); the microtubules appear to form an open lattice and run in parallel bands along the long axis of proximal villi. Microtubules may also be seen in ultrathin sections as narrow structures 20-24 nm in diameter (Fig. 21) and, very occasionally, a syncy- tial centriole may be seen (Fig. 25). Microtubules are generally associated with the maintenance of cell shape, intracellular transport, and secretion (Stephens and Edds, 1976), but their exact function in the placenta has not been clearly elucidated. Actin is present in microvilli (vide supra) and also in the superficial region of the syncytium where it forms a mat-like arrangement in the apical cytoplasm; these filaments may be involved in the translocation of coated vesicles (King, 1983).
J. Basal Plasma Membrane
The basal surface of the syncytium may form short microvillous processes, where it abuts on the cytotrophoblast between desmosomal junctions (Lister, 1964a; Ashley, 1965) and, when there is direct contact with the trophoblastic basement membrane, these may become quite complex. Various workers have considered this to be import- ant in water transfer and other metabolic processes (Rhodin and Terzakis, 1962; Panigel and Anh, 1963; Vacek, 1965; Salazar and Gonzalez-Angulo, 1967; Tighe et al., 1967; Knoth, 1968; Dec and Bartel, 1976) and have noted that the degree of interdigitation increases during the course of gestation. Transferrin receptors have also been located in this site (Vanderpuye et al., 1986) as

152 c.J.P. Jones and H. Fox
well as variable amounts of alkaline phosphatase activity (Fig. 8) (Jones and Fox, 1976).
V. TROPHOBLASTIC BASEMENT M E M B R A N E
A. S t ruc ture
The trophoblastic basement membrane separ- ates the trophoblast, for which it forms a support- ing matrix, from the underlying connective tissue stroma (Duance and Bailey, 1983). It is composed of three separate layers--the lamina lucida or lamina rara is, as its name suggests, very electron lucent with little observable structure and separates the plasma membrane of the trophoblast from the underlying lamina densa which resembles a felt- work of fine filaments in an amorphous matrix. Under this is the reticular layer of narrow fibrils, each about 15nm in diameter, which merges imperceptibly with the extracellular matrix of the stroma.
B. Compos i t ion
Immunocytochemical techniques have identified many of the components of the placental basement membrane. At the light microscopic level, the presence of type IV collagen has been demon- strated at all stages of pregnancy by immuno- fluorescence (Isemura et al., 1985; Kurosawa et al., 1985; Rukosuev et al., 1989) and immuno- peroxidase techniques (Yamada et al., 1987). At the ultrastructural level, using a peroxidase label, Amenta and co-workers (1986) found the lamina densa, but not the lamina rara, to be strongly stained for type IV collagen: remnants of basement membrane material free in the interstitium also showed positive staining. Laminin is also present in the basement membrane (Charpin et al., 1985; Kurosawa et al., 1985; Yamada et al., 1987; Leivo et al., 1989; Earl et al., 1990) and is preferentially localised in the lamina rara, and around the surface interdigitations of the syncytium that project down onto the basement membrane (Amenta et al., 1986). Some laminin is also present in the cytotro- phoblastic rough endoplasmic reticulum (Amenta et al., 1986; Leivo et al., 1989) which may, in part,
be responsible for its biosynthesis. Fibronectin is present in the trophoblastic basement membrane early in pregnancy but, apart from occasional residual specks, disappears later (Kurosawa et aL, 1985; Isemura et al., 1985; Yamaguchi et al., 1985; Amenta et al., 1986; Pfister et al., 1988; Rukosuev et al., 1989) although Earl et al. (1990) failed to detect it in their series of first trimester specimens. Merosin has also been demonstrated (Leivo et al., 1989) and, occasionally, traces of type III and type VI collagen may be found at the interface of the basement membrane with the stroma (Amenta et al., 1986).
A rich variety of other glycoproteins and gly- cosaminoglycans is also present in the basement membrane: lectin histochemistry has demonstrated a wide range of N-linked oligosaccharides, distinct from each other in terms of their lectin-binding properties (Johnson and Faulk, 1976; Whyte et al., 1978; Johnson and White, 1979; Whyte, 1983; Lee and Damjanov, 1984) and showing differing con- centrations at different stages of pregnancy. The presence of chondroitin sulphate and sulphated glycosaminoglycans has also been demonstrated by ultrastructural histochemistry (Parmley et al., 1984).
C. D e v e l o p m e n t
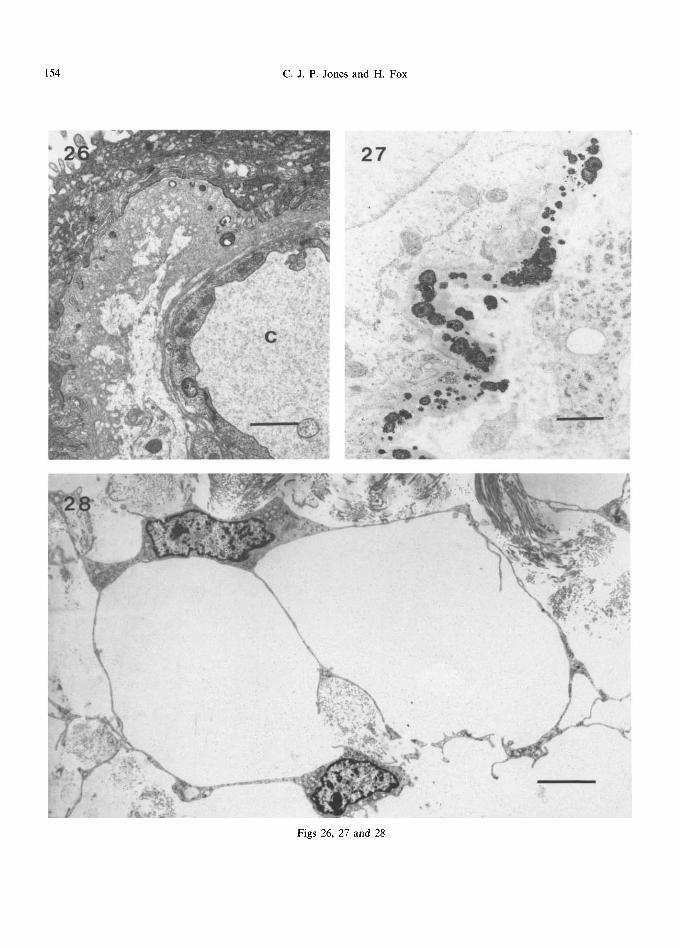
In the first trimester, the trophoblastic basement membrane is generally uniformly thin, with a faint suggestion of lamination (Fig. 3). The outermost layer appears to be slightly more electron dense than the others and is frequently in contact with collagen fibres in the stroma. Protrusion of the syncytium may be seen into the basement mem- brane, but this tissue never penetrates through to the stroma. At term, the basement membrane is more prominent but is extremely variable in thick- ness, ranging from 150 nm under vasculo-syncytial membranes to as much as 2600 nm in other areas (though part of this thickening may be due to tangential sectioning). In some areas a lacy or cribriform pattern, often with embedded small electron-dense inclusions and fibrils, can be seen (Fig. 26). In areas of vasculo-syncytial membranes, the basement membrane of the subjacent capillary is often confluent with that of the trophoblast.

Ultrastructure of the Human Placenta 153
This thickening of the basement membrane with increasing gestational age has been described by many workers (Sawasaki et al., 1957; Ikawa, 1959). Tighe et al. (1967) described a ten-fold, regular, increase in thickness from 10 to 100 nm, whereas Burgos and Rodriguez (1966) found unequal thickening between the alpha (thin, anuclear trans- port zone) and the beta (thicker, biosynthetic) zone, with thin basement membranes under the alpha zones. Jones (1976) also noted minimal basement membrane thickening under vasculo- syncytial membranes.
D. Inclusions
In the first and second trimesters, electron-dense inclusions may be present within, or just external to, the basement membrane. They are generally up to about 800nm in diameter with a concentric structure composed of layers of differing electron density (Figs 11 and 27) and show acid and alkaline phosphatase activity. The nature of these structures is controversial. Terzakis (1963), Lister (1963b, 1964a), Ashley (1965), Vacek (1965) and Anderson and McKay (1966) regarded them, almost certainly incorrectly, as lipid droplets, simi- lar to those seen in the syncytium, but it has more convincingly been suggested that they are deposits of iron (Salazar and Gonzalez-Angulo, 1967; McKay et al., 1958), calcium (Tighe et al., 1967) or both (Wislocki and Dempsey, 1946). Examination with an analytical electron microscope (Jones, 1976) has confirmed the presence of both calcium and iron, suggesting that the two elements are complexed together. These accumulations may be acting as a reserve pool prior to solubilisation and transfer to the fetus or, alternatively, could simply be excess material.
VI. THE STROMA OF THE CHORIONIC VILLUS
A. Re t i cu lum Cells
Our understanding of the internal structure of the connective tissue core of the chorionic villus has been greatly enhanced by new developments
in the field of scanning electron microscopy, including the technique of cryofracture, and by improved instrumentation and new fixation protocols (Burton, 1987). As a result of these advances, it has been possible to delineate the three-dimensional relationships that exist between 'fixed' stromal cells and the collagen fibrils they secrete (Castellucci et al., 1980, 1984; Castellucci and Kaufmann, 1982; Martinoli et al., 1984 and Burton, 1987), and to augment the descriptions of the internal architecture of various stromal cell types by Vacek (1970), Hoyes (1971), Liebhart (1974b) and Kaufmann et al. (1977) who used thin sections viewed by transmission electron microscopy.
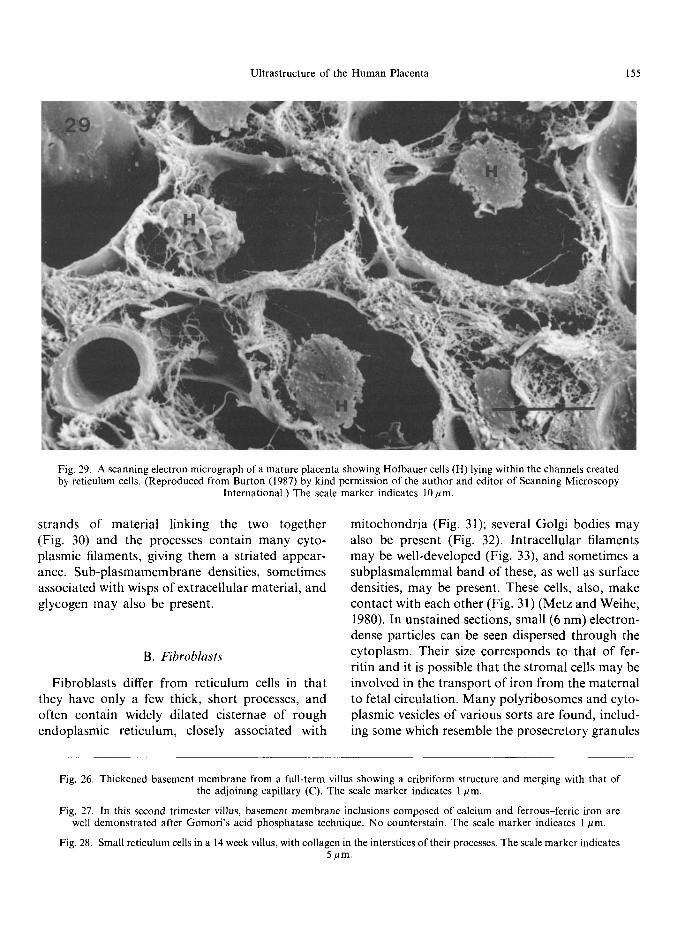
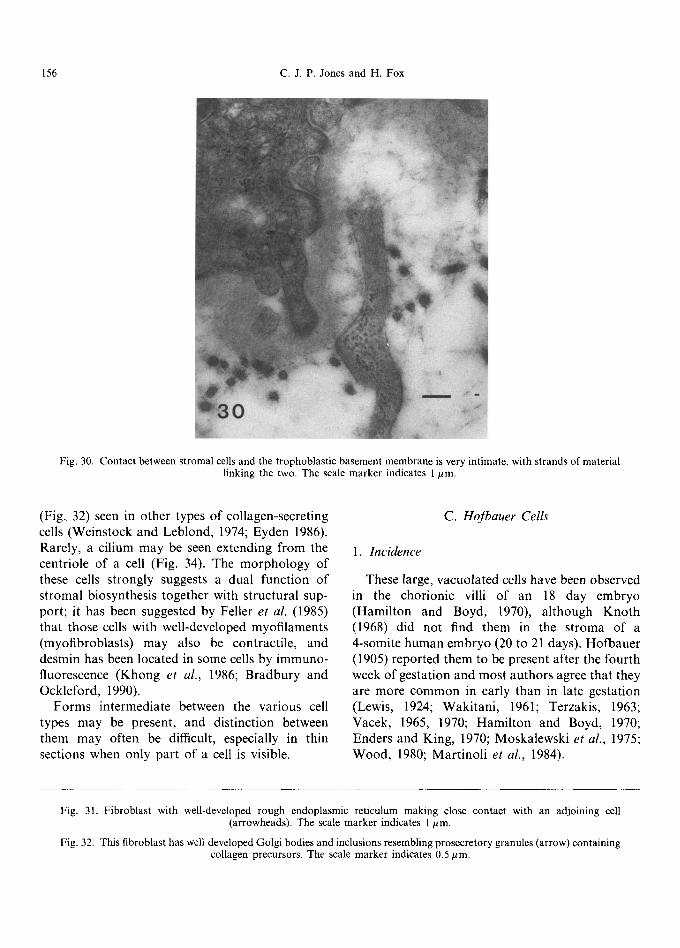
Early in gestation, undifferentiated mesenchy- mal cells form a loose network in the villous core, with collagen fibrils forming a delicate mesh- work around them (Martinoli et al., 1984; Burton, 1987). They are small, spindle-shaped cells with little cytoplasm and long, thin cytoplasmic pro- cesses (Kaufmann et al., 1977; Castellucci and Kaufmann, 1982; Vacek, 1970). These cells give rise to small reticulum cells (Kaufmann et al., 1977) characterised by a bizarre-shaped body and numerous processes (Fig. 28) which, under the scanning electron microscope, resemble 'sails' of cytoplasm that anastomose and make close contact with each other via tight junctions. Within the cell body and along parts of the processes are a variety of organelles, including narrow cisternae of rough endoplasmic reticulum, mitochondria and poly- ribosomes. The Golgi apparatus is frequently juxtanuclear in position (Kaufmann et al., 1977). Pinocytotic activity is usually concentrated at the dilated ends of the 'sails' (Kaufmann et al., 1977). Large reticulum cells have more cytoplasm and fewer processes, with a well-developed endo- plasmic reticulum. Reticulum cells form a network between the trophoblastic basement membrane, each other and the capillaries, and form the chan- nels through which Hofbauer cells move (Fig. 29). These channels are generally parallel to the long axis of the villus with intercommunicating fenestra- tions between them (Castellucci et al., 1984; Martinoli et al., 1984). The contact of the cyto- plasmic processes with the trophoblastic basement membrane is often a very intimate one, with
154 C . J . P . Jones and H. Fox
27
' / ' " 2" ..'a:.
m
• < . . %
,. . ; O
/ ~'e,. , : k /
t .
r ~ ~
k
Figs 26, 27 and 28
Ultrastructure of the Human Placenta 155
Fig. 29. A scanning electron micrograph of a mature placenta showing Hofbauer cells (H) lying within the channels created by reticulum cells. (Reproduced from Burton (1987) by kind permission of the author and editor of Scanning Microscopy
International.) The scale marker indicates 10/~m.
strands o f material linking the two together (Fig. 30) and the processes contain many cyto- plasmic filaments, giving them a striated appear- ance. Sub-p lasmamembrane densities, sometimes associated with wisps o f extracellular material, and glycogen may also be present.
B. Fibroblasts
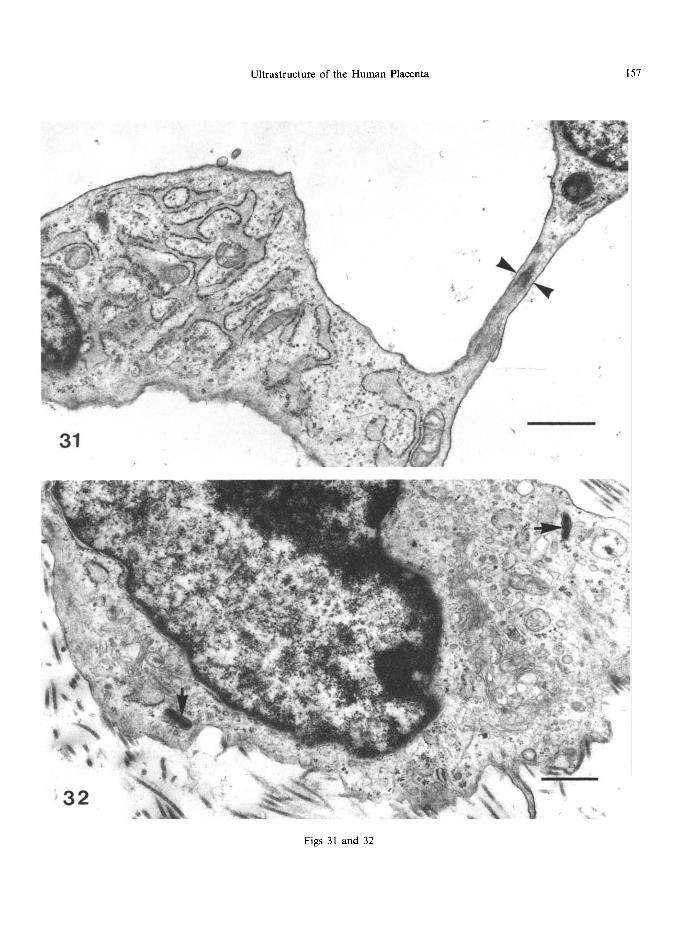
Fibroblasts differ f rom reticulum cells in that they have only a few thick, short processes, and often contain widely dilated cisternae o f rough endoplasmic reticulum, closely associated with
mi tochondr ia (Fig. 31); several Golgi bodies may also be present (Fig. 32). Intracellular filaments may be well-developed (Fig. 33), and sometimes a subplasmalemmal band of these, as well as surface densities, may be present. These cells, also, make contact with each other (Fig. 31) (Metz and Weihe, 1980). In unstained sections, small (6 nm) electron- dense particles can be seen dispersed th rough the cytoplasm. Their size corresponds to that o f fer- ritin and it is possible that the stromal cells may be involved in the t ranspor t o f iron f rom the maternal to fetal circulation. M a n y polyr ibosomes and cyto- plasmic vesicles o f various sorts are found, includ- ing some which resemble the prosecretory granules
Fig. 26. Thickened basement membrane from a full-term villus showing a cribriform structure and merging with that of the adjoining capillary (C). The scale marker indicates 1/~m.
Fig. 27. In this second trimester villus, basement membrane inclusions composed of calcium and ferrous-ferric iron are well demonstrated after Gomori's acid phosphatase technique. No counterstain. The scale marker indicates 1/~m.
Fig. 28. Small reticulum cells in a 14 week villus, with collagen in the interstices of their processes. The scale marker indicates 5#m.
156 C.J.P. Jones and H. Fox
Fig. 30. Contact between stromal cells and the trophoblastic basement membrane is very intimate, with strands of material linking the two. The scale marker indicates 1/~m.
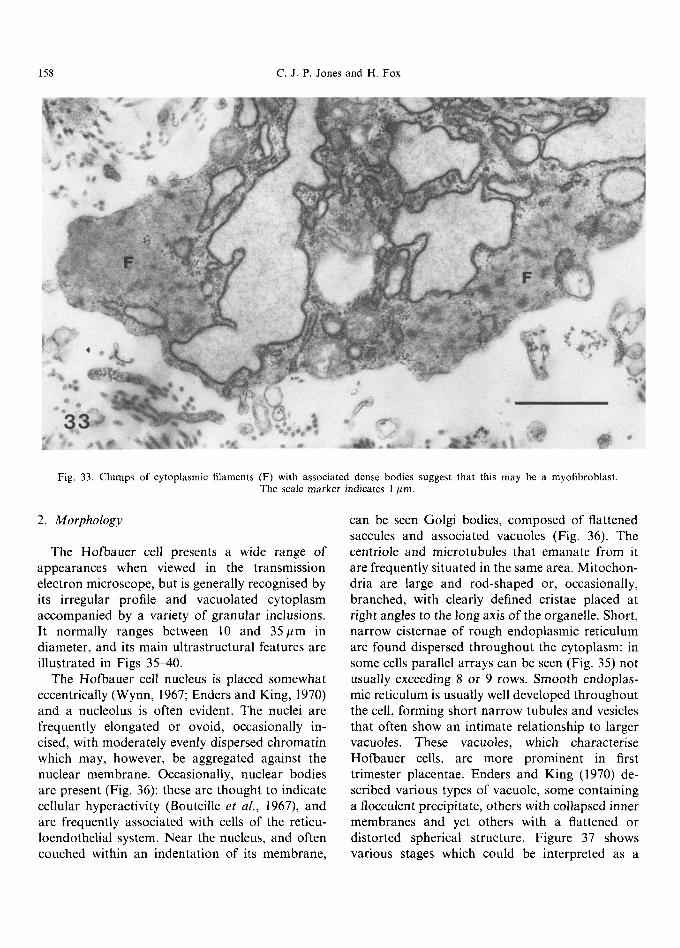
(Fig, 32) seen in other types of collagen-secreting cells (Weinstock and Leblond, 1974; Eyden 1986). Rarely, a cilium may be seen extending from the centriole of a cell (Fig. 34). The morphology of these cells strongly suggests a dual function of stromal biosynthesis together with structural sup- port; it has been suggested by Feller et al. (1985) that those cells with well-developed myofilaments (myofibroblasts) may also be contractile, and desmin has been located in some cells by immuno- fluorescence (Khong et al., 1986; Bradbury and Ockleford, 1990).
Forms intermediate between the various cell types may be present, and distinction between them may often be difficult, especially in thin sections when only part of a cell is visible.
C. Hofbauer Cells
1. Incidence
These large, vacuolated cells have been observed in the chorionic villi of an 18 day embryo (Hamilton and Boyd, 1970), although Knoth (1968) did not find them in the stroma of a 4-somite human embryo (20 to 21 days). Hofbauer (1905) reported them to be present after the fourth week of gestation and most authors agree that they are more common in early than in late gestation (Lewis, 1924; Wakitani, 1961; Terzakis, 1963; Vacek, 1965, 1970; Hamil ton and Boyd, 1970; Enders and King, 1970; Moskalewski et al., 1975; Wood, 1980; Martinoli et al., 1984).
Fig. 31. Fibroblast with well-developed rough endoplasmic reticulum making close contact with an adjoining cell (arrowheads). The scale marker indicates 1 #m.
Fig. 32. This fibroblast has well-developed Golgi bodies and inclusions resembling prosecretory granules (arrow) containing collagen precursors. The scale marker indicates 0.5 #m.
Ultrastructure of the Human Placenta 157
S
j
31 ~ ~
Figs 31 and 32
158 C.J.P. Jones and H. Fox
ao~
a;
m i '̧ ' ~
• ' , z , ,
• P . , 2
~,t ~.. , , o.~e D . /
Fig. 33. Clumps of cytoplasmic filaments (F) with associated dense bodies suggest that this may be a myofibroblast. The scale marker indicates 1 pm.
2. Morphology
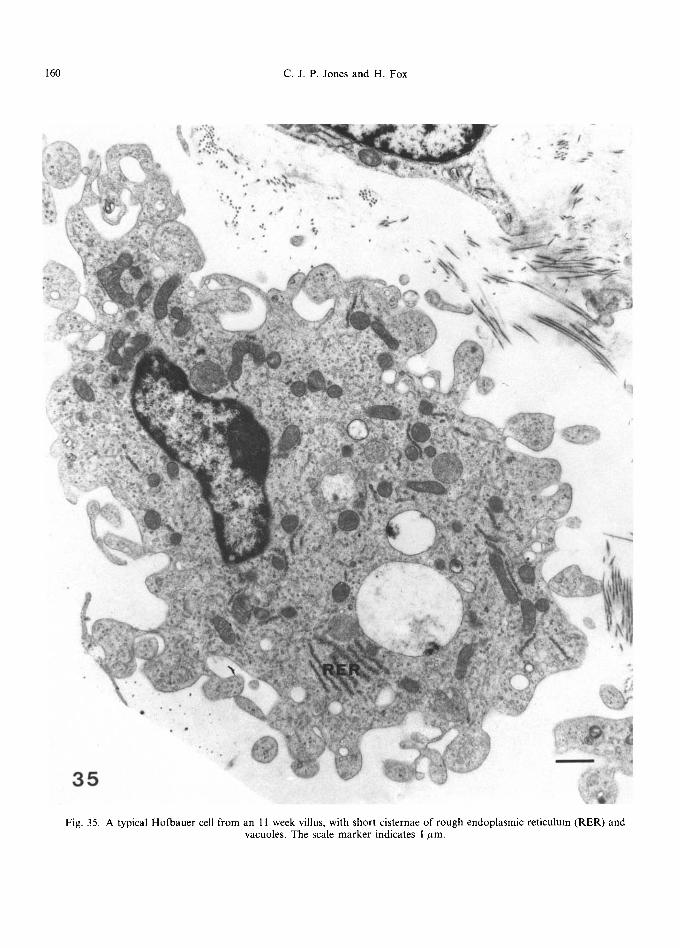
The Hofbauer cell presents a wide range of appearances when viewed in the transmission electron microscope, but is generally recognised by its irregular profile and vacuolated cytoplasm accompanied by a variety of granular inclusions. It normally ranges between 10 and 35/~m in diameter, and its main ultrastructural features are illustrated in Figs 3540 .
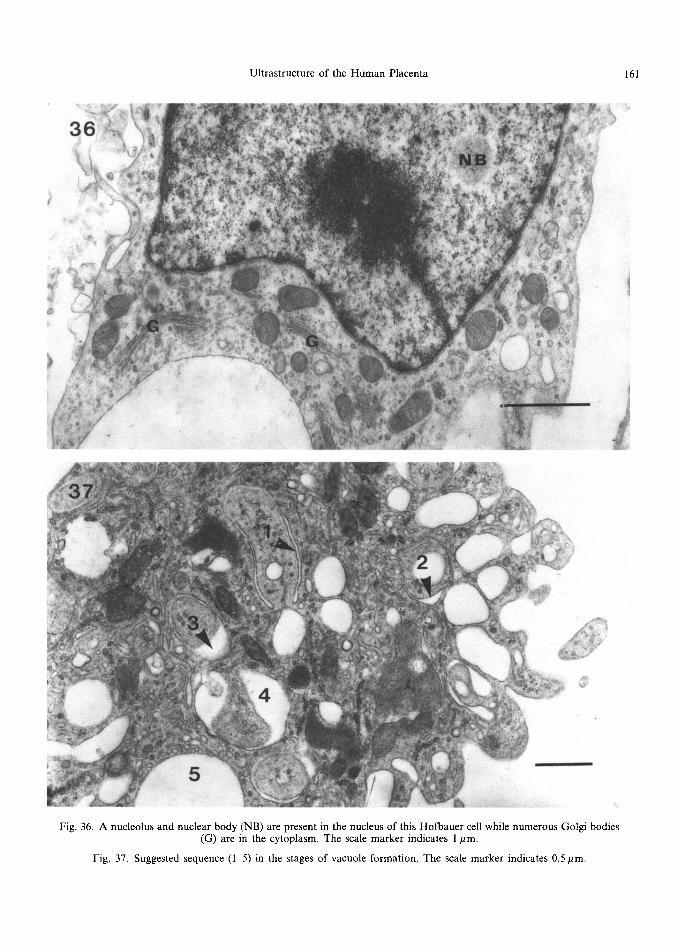
The Hofbauer cell nucleus is placed somewhat eccentrically (Wynn, 1967; Enders and King, 1970) and a nucleolus is often evident. The nuclei are frequently elongated or ovoid, occasionally in- cised, with moderately evenly dispersed chromatin which may, however, be aggregated against the nuclear membrane. Occasionally, nuclear bodies are present (Fig. 36): these are thought to indicate cellular hyperactivity (Bouteille et al., 1967), and are frequently associated with cells of the reticu- loendothelial system. Near the nucleus, and often couched within an indentation of its membrane,
can be seen Golgi bodies, composed of flattened saccules and associated vacuoles (Fig. 36). The centriole and microtubules that emanate from it are frequently situated in the same area. Mitochon- dria are large and rod-shaped or, occasionally, branched, with clearly defined cristae placed at right angles to the long axis of the organelle. Short, narrow cisternae of rough endoplasmic reticulum are found dispersed throughout the cytoplasm: in some cells parallel arrays can be seen (Fig. 35) not usually exceeding 8 or 9 rows. Smooth endoplas- mic reticulum is usually well developed throughout the cell, forming short narrow tubules and vesicles that often show an intimate relationship to larger vacuoles. These vacuoles, which characterise Hofbauer cells, are more prominent in first trimester placentae. Enders and King (1970) de- scribed various types of vacuole, some containing a flocculent precipitate, others with collapsed inner membranes and yet others with a flattened or distorted spherical structure. Figure 37 shows various stages which could be interpreted as a

Ultrastructure of the Human Placenta 159
Fig. 34. Part of a fibroblast containing Golgi bodies and a cilium extending into the extracellular milieu. The scale marker indicates 1 ~m.
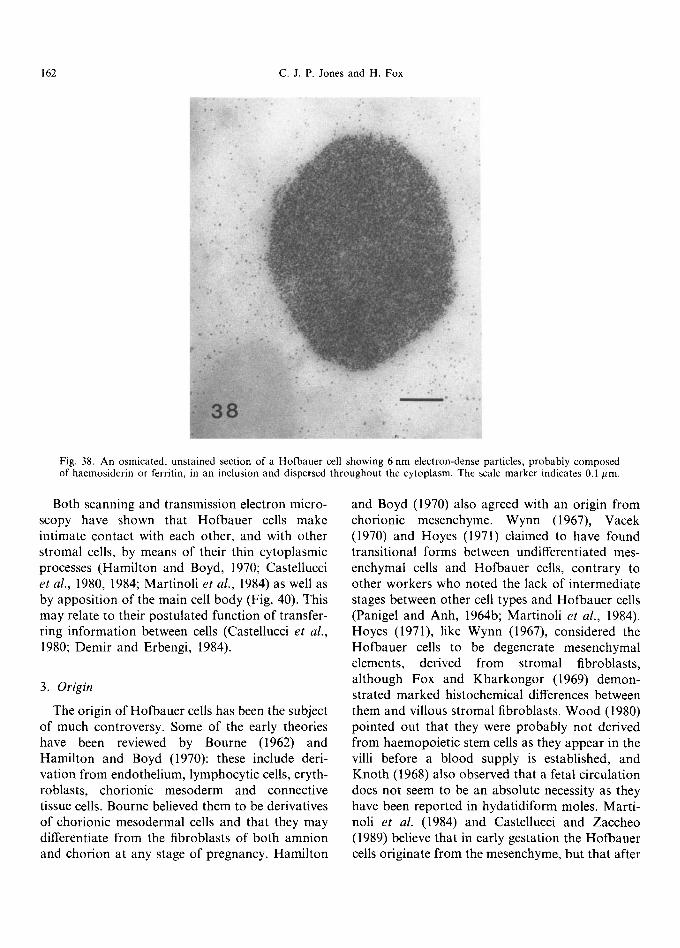
sequence in the formation of a vacuole, starting with a flask-shaped membranous intracytoplasmic invagination which then shows dilatation of its 'neck' until the inner tongue of cytoplasm is gradually displaced by the swelling contents. Many vacuoles have flocculent contents and sometimes one or more dense, osmiophilic bodies. An assort- ment of granules of varying electron density are also characteristic of these cells; examination of unstained material shows some of these bodies to contain masses of electron-dense particles, about 6 nm in diameter, probably composed of haemosiderin or ferritin (Fig. 38), which may also be dispersed throughout the cytoplasm. The pres- ence of intracellular iron has been confirmed by histochemical studies at the light microscope level (McKay et al., 1958; Bourne, 1962). The granules are also rich in acid phosphatase activity (Fig. 39), demonstrat ing their lysosomal origin. Coated and micropinocytotic vesicles are evident on the surface of the cells as well as dispersed through the cyto- plasm, and microtubules and cytoplasmic filaments
can also be found, some of which have been shown to be vimentin (Ocklef'ord et al., 1981).
Recent scanning and cryofracture electron microscopy studies have shed new light on to the three dimensional ultrastructure of these cells (Castellucci et al., 1980, 1984; Castellucci and Kaufmann, 1982; Martinoli et al., 1984; Burton, 1987) and have elucidated the nature of the narrow processes and folds seen in thin sections. It is now known that complex systems of lamellipodia extend out from the surface of the cell to form trumpet or funnel-like structures, often folding in around themselves, together with surface blebs and protrusions or microplicae. In early (6 week) placentae these cells are enmeshed in a network of collagen fibres, but as pregnancy progresses they are mainly found within a system of channels lined by resident or fixed cells. These reduce in size by term and the Hofbauer cells are then found within the collagenous stroma or in small compart - ments that are probably remnants of the channel system.
160 C . J . P . Jones and H. Fox
\ .
~t
3~" , , . r
3 5
Fig. 35. A typical Hofbauer cell from an 11 week villus, with short cisternae of rough endoplasmic reticulum (RER) and vacuoles, The scale marker indicates 1 #m.
Ultrastructure of the Human Placenta 161
Fig. 36. A nucleolus and nuclear body (NB) are present in the nucleus of this Hofbauer cell while numerous Golgi bodies (G) are in the cytoplasm. The scale marker indicates 1 pm.
Fig. 37. Suggested sequence (I 5) in the stages of vacuole formation. The scale marker indicates 0.5 pm.
162 C.J.P. Jones and H. Fox
~, .: ~ • i s • • ~ ~ /.
Fig. 38. An osmicated, unstained section of a Hofbauer cell showing 6 nm electron-dense particles, probably composed of haemosiderin or ferritin, in an inclusion and dispersed throughout the cytoplasm. The scale marker indicates 0.1 ~m.
Both scanning and transmission electron micro- scopy have shown that Hofbauer cells make intimate contact with each other, and with other stromal cells, by means of their thin cytoplasmic processes (Hamilton and Boyd, 1970; Castellucci et al., 1980, 1984; Martinoli et al., 1984) as well as by apposition of the main cell body (Fig. 40). This may relate to their postulated function of transfer- ring information between cells (Castellucci et al., 1980; Demir and Erbengi, 1984).
3. Origin
The origin of Hofbauer cells has been the subject of much controversy. Some of the early theories have been reviewed by Bourne (1962) and Hamilton and Boyd (1970): these include deri- vation from endothelium, lymphocytic cells, eryth- roblasts, chorionic mesoderm and connective tissue cells. Bourne believed them to be derivatives of chorionic mesodermal cells and that they may differentiate from the fibroblasts of both amnion and chorion at any stage of pregnancy. Hamilton
and Boyd (1970) also agreed with an origin from chorionic mesenchyme. Wynn (1967), Vacek (1970) and Hoyes (1971) claimed to have found transitional forms between undifferentiated mes- enchymal cells and Hofbauer cells, contrary to other workers who noted the lack of intermediate stages between other cell types and Hofbauer cells (Panigel and Anh, 1964b; Martinoli et al., 1984). Hoyes (1971), like Wynn (1967), considered the Hofbauer cells to be degenerate mesenchymal elements, derived from stromal fibroblasts, although Fox and Kharkongor (1969) demon- strated marked histochemical differences between them and villous stromal fibroblasts, Wood (1980) pointed out that they were probably not derived from haemopoietic stem cells as they appear in the villi before a blood supply is established, and Knoth (1968) also observed that a fetal circulation does not seem to be an absolute necessity as they have been reported in hydatidiform moles. Marti- noli et al. (1984) and Castellucci and Zaccheo (1989) believe that in early gestation the Hofbauer cells originate from the mesenchyme, but that after
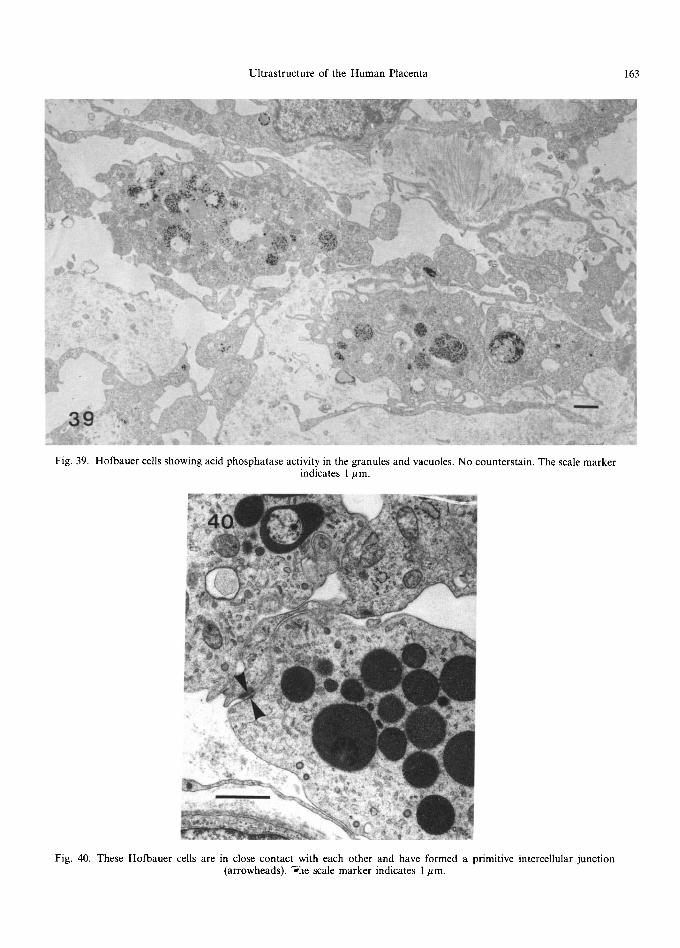
Ultrastructure of the Human Placenta 163
Fig. 39. Hofbauer cells showing acid phosphatase activity in the granules and vacuoles. No counterstain. The scale marker indicates 1/~m.
Fig. 40. These Hofbauer cells are in close contact with each other and have formed a primitive intercellular junction (arrowheads). 7"he scale marker indicates 1 #m.

164 c.J.P. Jones and H. Fox
a fetal villous circulation is established, they could be supplemented by a further population of cells derived from the fetal liver and from fetal bone marrow-derived monocytes (Castellucci et al., 1987), producing sub-populations of cells with different past histories and functions. Fox and Kharkongor (1969) noted a heterogeneity in en- zyme activity of Hofbauer cells, a feature that has been seen in other macrophage populations that exhibit functional and immunological variability (Harbison et al., 1984). These variations could be perpetuated by mitotic division for, although Wynn (1967) considered Hofbauer cells to be degenerate and only rarely capable of division, Castellucci et al. (1987) demonstrated Hofbauer cells undergoing mitosis in every placenta exam- ined between 5 and 20 weeks' gestation, thus showing them to be a self-replicating population. Castellucci et al. (1987) proposed that, in the first half of pregnancy, the large vacuolated Hofbauer cells were probably more mature than the poorly vacuolated or unvacuolated ones and suggested either that Hofbauer cells could divide at least twice during their development or that subpopu- lations exist with the capacity to divide at different times in their life cycle.
4. Function
Many functions have been proposed for Hofbauer cells, these including, with a varying degree of credibility, an endocrine (Katabuchi et al., 1989) or secretory role (Vacek, 1970; Dec et al., 1979), regulation of stromal water content (Enders and King, 1970; Vacek, 1970; Castellucci et al., 1980; Martinoli et al., 1984; Demir and Erbengi, 1984; Castellucci and Zaccheo, 1989; Katabuchi et al., 1989), transfer of information between cells (Castellucci et al., 1980; Demir and Erbengi, 1984), removal and remodelling of stro- mal components, including modulation of endo- thelial and fibroblast cell growth (Castellucci et al., 1980, 1983; Demir and Erbengi, 1984; Martinoli et al., 1984; Castellucci and Zaccheo, 1989), fetal nutrition (Hamilton and Boyd, 1970) and various immunological roles including antigen presen- tation, suppression of immune function and host defence (Wood, 1980; Bulmer and Johnson, 1984;
Castellucci et al., 1984; Mues et al., 1989). Most of the above authors also agreed that the large vacuoles and lysosomal activity were indicative of phagocytosis, a function confirmed histochemi- cally (McKay et al., 1958; Bourne, 1962; Vacek, 1965; Fox and Kharkongor, 1969) and by the ultrastructural features described above. That the cells are also motile has been proposed by Castel- lucci et al. (1980, 1984) and Martinoli et al. (1984) who showed scanning electron micrographs of Hofbauer cells crossing intercommunicating stro- mal channels. Their pleomorphism suggests a high capacity for shape change, though Bourne (1962) considered they were only capable of moving within tissues for a very limited distance.
5. I m m u n o l o g y
Immunological studies on Hofbauer cells have confirmed their similarity to tissue macrophages (Moskalewski et al., 1975; Wood, 1980; Loke et al., 1982; Bulmer and Johnson, 1984; Braunhut et al., 1984; Katabuchi et al., 1989; Castellucci and Zaccheo, 1989); they show positive rosette for- mation, immune and non-immune phagocytosis, adherence to plastic dishes and stain positively with a range of monoclonal antibodies that recog- nise tissue macrophages. However, Goldstein et al. (1988) remarked on some major antigenic differ- ences between these and other types of macro- phage, while Castellucci et al. (1983) and Bulmer and Johnson (1984) were, unlike Wood (1980) and Loke et al. (1982) unable to demonstrate the Fc and C3 receptors that are normally characteristic of this cell type. Castellucci et al. (1983) con- cluded that the Hofbauer cell was "a very peculiar macrophage-type cell".
D. Capillaries
1. Morpho logy
The main ultrastructural features of the fetal capillaries in normal placental villi are shown in Figs 41~,8. Early in gestation these vessels are generally small, with prominent endothelial cells that appear to be metabolically active (Fig. 41), having a content of many cisternae of rough endoplasmic reticulum, free ribosomes, mitochon-
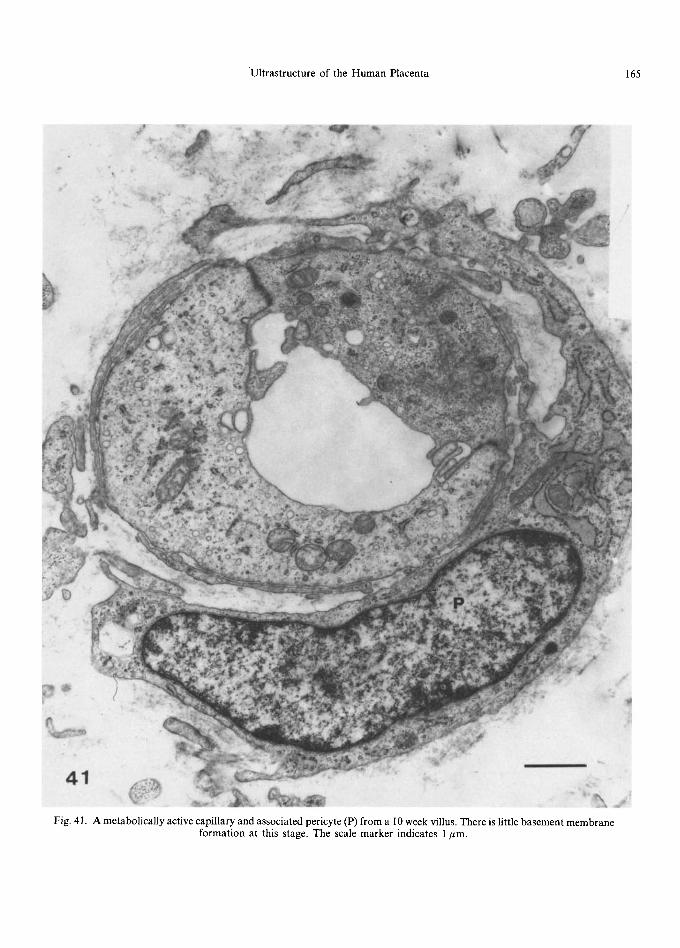
Ultrastructure of the Human Placenta 165
i i~• ~•
" ~ i - 5
41
Fig. 41. A metabolically active capillary and associated pericyte (P) from a 10 week villus. There is little basement membrane formation at this stage. The scale marker indicates 1/~m.

166 C.J.P. Jones and H. Fox
dria, small Golgi bodies, secretory droplets and, sometimes, centrioles. Weibel-Palade bodies may, occasionally, be seen (Fig. 42). Both luminal and external surfaces of the endothelial cells show cytoplasmic projections (Rhodin and Terzakis, 1962; Hoyes, 1971) and adjacent cells are united by a series of specialised junctions (Figs 43 and 44) resembling the zonulae occludentes of epithelial cells (Heinrich et al., 1976; Metz and Weihe, 1980). The adjoining limiting membranes are in contact but the cells maintain their individuality. Occasional deposits of glycogen are seen in the cells and their outgrowths (Lister, 1964b; Hamilton and Boyd, 1970; Heinrich et al., 1976; Jones, 1976; Bartel et al., 1978) in the early stages of pregnancy.
As gestation proceeds, the capillaries dilate and, in the second trimester, vasculo-syncytial mem- branes are beginning to be apparent in occasional villi (Lister, 1964b; Jones, 1976). The luminal plasma membrane becomes smoother and by term many vessels are sinusoidally dilated, with attenu- ated endothelial cells which are joined by tight junctions and contain few organelles (Fig. 45).
The fetal capillary plays a direct role in the passage of substances from mother to fetus and vice versa (Rigano and Sermann, 1963; Panigel and Anh, 1964a; Widmaier, 1969; Sideri et al., 1987) and therefore, not surprisingly, contains numerous pinocytotic vesicles (Fig. 43) which increase in number as gestation proceeds (Wakitani, 1961; Lister, 1964b; Hamilton and Boyd, 1970; Jones, 1976): Heinrich et al. (1976) did, however, point out that the overall number of such vesicles is small when compared to other types of capillary, such as those in muscle. The number of intra- cellular filaments in the endothelial cells also in- creases throughout pregnancy, these being present in small numbers at 8-10 weeks and numerous at term (Fig. 46) (Lister, 1964b; Strauss et al. 1965; Jones, 1976; Bartel et al., 1978). These filaments
have been thought to be responsible for the pur- ported contractile properties of the fetal capillaries (Terzakis, 1963; Hoyes, 1971; Heinrich et al., 1976; Bartel et al., 1978) but are predominantly vimentin (Ockleford et al., 1981; Clark and Damjanov, 1985; Beham et al., 1988; Bradbury and Ockelford, 1990) which has a mechanical or supportive role in these cells (Ockelford and Wakely, 1982; Lazarides, 1982): some actin filaments are, how- ever, present as has been shown by their decoration with myosin (S1) (King, 1983).
Jones (1976) found little evidence of an alter- ation in the size of the tight junctions between adjacent endothelial cells as gestation progresses: this was in contradiction to Lister's observation (1964b) of an increase in length of the junctions with time. Jones (1976) was also unable to confirm the increase in size of the luminal projections noted by Lister (1963b, 1964b) and, indeed, found the reverse, large intraluminal endothelial cell projections being frequently seen in the immature placenta but being uncommon at term. Jones (1976) also observed that protrusions through the capillary basement membrane often became vesicular later in pregnancy, with electron-lucent contents, similar to the 'blebs' described by Martinek et al. (1975).
2. Origin
Dempsey (1972) thought that pericytes are the principal progenitors of fetal blood vessels, claim- ing that endothelial cells do not undergo mitosis. Jones (1976) and Castellucci et al. (1987) did observe, however, clear evidence of dividing endo- thelial cells (Fig. 47) and therefore the endothelium is obviously not totally reliant on the pericytes for growth. Further, there is a possibility of a contri- bution of cells from the cytotrophoblast layer to the endothelial cell population. Hertig (1935) and
Fig. 42. A Weibel-Palade body (arrow) in a first trimester capillary. The scale marker indicates 0.5 #m.
Fig. 43. Part of a capillary from an 18 week villus showing specialised intercellular junctions (J) and micropinocytotic vesicles (arrows). The scale marker indicates 0.5/~m.
Fig. 44. In this capillary from an 8 week villus, the component membranes of the intercellular junction do not meet. There is electron-dense material between them. The scale marker indicates 0.1/~m.
Fig. 45. Full-term villus containing a widely dilated capillary with thin walls. The scale marker indicates 5 #m.
Ultrastructure of the Human Placenta 167
Figs 42, 43, 44 and 45
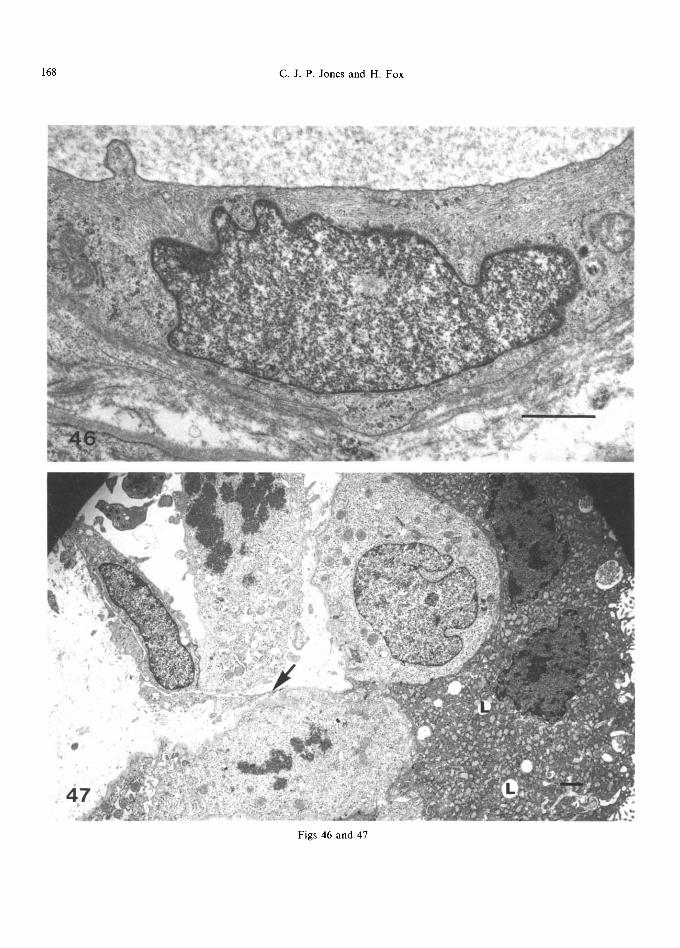
168 C . J . P . Jones and H. Fox
Figs 46 and 47

Ultrastructure of the Human Placenta 169
Cibils (1968) indeed maintained, from light micro- scopic evidence, that all the mesenchymal elements of the placental villi were derived from the cytotro- phoblast, a claim that was strongly denied by Dempsey (1972) and later by Luckett (1978). There may, however, be an element of truth in this theory as we have observed dense, elongated cells stream- ing out from the trophoblast into the stroma to make contact with existing endothelial cells (Fig. 11). Unfortunately, no direct evidence was found of an actual separation of these dense cells from the trophoblast although it appeared from the nature of the interdigitations between them that such a separation could easily be effected. The outer coating of basement membrane material would then be transferred to the capillary and there would be no question of the cells having to penetrate the trophoblastic basement membrane to effect their 'escape'. These cells have only been observed in the early stages of pregnancy, at the time when angiogenesis is prominent, and only in certain areas of the villi, which confirms the original observation of Hertig (1935) that this phenomenon is only seen whilst new primitive villi are being formed from solid trophoblast columns. The fact that endothelial cells have been seen in mitosis, and the descriptions of the involvement of pericytes in capillary development by Dempsey (1972) indicate that the contribution, if any, of the dense cytotro- phoblast cells to angiogenesis is only a partial one.
3. Per i cy t e s
Pericytes have been described in close associ- ation with the capillaries (Rhodin and Terzakis, 1962; Strauss et al., 1965; Lister, 1964b; Bartel et al., 1978) and Widmaier (1969) attributes to them participation in the transport function of the capillary. The pericyte itself, well-described by Bartel et al. (1978), is probably metabolically active as it contains a well-developed endoplasmic reticu- lum. These cells may be involved in capillary
basement membrane biosynthesis but not, how- ever, to the exclusion of endothelial cells, for Martinek et al. (1975) have shown that there is no correlation between capillary basement membrane thickness and the presence or absence of pericytes. Dempsey (1972) described desmosomal-like junc- tions between pericytes and endothelial cells but these are more likely to have been sub-plasmalem- mal densities as the basement membrane usually clearly separates the two cell types (Heinrich et al., 1976; Jones, 1976).
4. B a s e m e n t M e m b r a n e
The time at which a basement membrane can be found surrounding the fetal capillaries has been variably reported, although it is clearly not present in the first eight weeks of gestation (Knoth, 1968; Topilko and Pisarski, 1967). Lister (1964b) found the capillary basement membrane to be either absent or incomplete at 8-10 weeks of gestation, whilst Becker and Seifert (1965) failed to see endothelial basement membrane in their specimens of 4 and 5 months' gestation. Nakano (1963) observed, however, a basement membrane around fetal capillaries from the second month, as did Vacek (1965) at three months' gestation, while Jones (1976) noted a membrane in a 9 week old specimen.
In first trimester placentae, electron dense fibril- lar material resembling fibronectin (Chen, 1978; Hedman et al., 1978; Jones et al., 1980) can be seen around endothelial cells (Fig. 48). Fibronectin and laminin have been located around fetal blood vessels both in immature and term placentae using immunocytochemical techniques (Charpin et al., 1985; Isemura et al., 1985; Kurosawa et al., 1985; Yamaguchi et al., 1985; Amenta et al., 1986; Yamada et al., 1987) and also in the pinocytotic vesicles of endothelial cells (Amenta et al., 1986); fibronectin may function as the initial backbone for the fibrillogenesis of matrix proteins (Chen,
Fig. 46. There are numerous cytoplasmic filaments in this full-term capillary and the basement membrane is multilaminar. The scale marker indicates 1/~m.
Fig. 47. Cytotrophoblast and endothelium undergoing mitotic division in a first trimester villus. Note the close association of the capillary with the trophoblast (arrow). Lipid droplets (L) are present in the syncytium, which contains numerous
web-like cisternae of rough endoplasmic reticulum. The scale marker indicates I #m.
EMR 4 / 1 ~

170 C.J.P. Jones and H. Fox
Fig. 48. Material resembling fibronectin is present as short, electron-dense strands around this 14 week endothelial cell (E). The scale marker indicates 1 #m.
1978) such as type IV collagen, a normal constituent of basement membranes which has been demonstrated in this location by immuno- cytochemistry (Ockleford et al., 1981; Kurosawa et al., 1985; Amenta et al., 1986; Rukosuev et al., 1989), and type V collagen which may be associ- ated with, if not an integral part of, this structure.
E. M a t r i x
The amount of collagen present in chorionic villi progressively increases during the course of preg- nancy (Sawasaki et al., 1957; Vacek, 1965; Herbst et al., 1968; Liebhart, 1974a; Castellucci et al., 1982). The stromal cells of the early villous mesenchyme (up to 7 or 8 weeks' gestation) are enmeshed in a network of tiny bundles of collagen fibres with no preferred orientation (Martinoli et al., 1984). These villi gradually develop into imma- ture intermediate villi characterised by reticular cells, the cytoplasmic processes of which form the walls of the stromal channels described above;
collagen fibrils become condensed in the interstices of these cells (Fig. 28). By 14 weeks' gestation, the increase in collagen production compresses and fills the stromal channels of proximal intermediate villi to form the compact, fibrous stroma character- istic of the stem villus, while during the last trimester, peripheral immature intermediate villi develop into the mature form, with a tight fibrillar network filling the spaces between narrow and short stromal channels and fetal vessels. Terminal villi, which develop after about the 30th week, are mainly occupied by sinusoidally dilated capil- laries, with little room left for the collagenous stroma.
The various types of collagen secreted by the stromal cells have been identified at the ultrastruc- tural level using immunocytochemistry (Amenta et al., 1986). It was proposed that the large, cross- banded fibrils (30-35 nm) are composed of type I collagen, while thin, beaded fibrils (10-15nm) often forming a meshwork around type I collagen, are mainly type III collagen. However, recent

Ultrastructure of the Human Placenta 171
evidence suggests tha t c o p o l y m e r i s a t i o n o f col la- gens I and I I I occurs in m a n y tissues and can result in fibrils with a wide var ie ty o f d iamete r s (Keene et al., 1987) and so this might be an oversimplif i - ca t ion o f the t rue type d i s t r ibu t ion . W h e t h e r the thin b e a d e d type I I I fibrils are ana logous to the ' r e t i cu l in ' fibres o f l ight mic roscopy was discussed by A m e n t a et al. (1986), who conc luded tha t there is no cor re la t ion between ' re t icu l in ' and any specific ma t r ix componen t . Types V and VI col la- gen, with diameters o f 6-10 nm, have been localised on the surface o f col lagen fibres o f all d iameters , ma in ly in the pe r ivascu la r regions, type V tending to m a s k the b a n d i n g pa t t e rn o f type I col lagen while type VI enhanced it. Type VI col lagen has also been found assoc ia ted with 5-10 nm d iamete r b e a d e d microf i laments , of ten showing a per iod ic s ta in ing pa t t e rn a long their length, as well as being p resen t in a m o r p h o u s regions (Von der M a r k et
al., 1984; A m e n t a et al., 1986). Type IV col lagen was found by A m e n t a et al. (1986) in all basement m e m b r a n e s ( including r emnan t s o f lamel lae free in the in ters t i t ium), as is charac ter i s t ic o f this type o f co l lagen (Kefal ides , 1973), whereas type V was only occas iona l ly loca ted in vascu la r basement m e m b r a n e s ad jacen t to the vil lous in ters t i t ium, as descr ibed also by M a d r i et al. (1982) and R u k o s u e v et al. (1989) using immunof luorescence . F i b r o n e c t i n has been shown to be in t imate ly con- nected to the col lagen fibrils o f the s t roma, enhanc- ing thei r per iodic i ty , and also to be presen t as free f i laments or g lobu la r aggregates wi th in the inter- s t i t ium (Amen ta et al., 1986). S t roma l depos i t ion o f f ib ronec t in has been descr ibed at the l ight mic roscope level, where loca l i sa t ion was demon- s t ra ted within s t romal and endothe l i a l cells ( I s emura et al., 1985; Y a m a g u c h i et al., 1985; Y a m a d a et al., 1987; Pfister et al., 1988) and it was suggested tha t it was local ly synthesised and re ta ined within the tissue.
O the r g lycopro te ins and g lycosaminoglycans , d e m o n s t r a t e d by lec t in-b inding studies, are also presen t in the vi l lous s t roma ( Johnson and Fau lk , 1976; W h y t e et al., 1978; Johnson and Whi te , 1979; Why te , 1983; Lee and D a m j a n o v , 1984). The dis- t r i bu t ion o f these c o m p o n e n t s appea r s to al ter with increas ing ges ta t iona l age ( Johnson and White , 1979; Whyte , 1983).
Acknowledgements--We would like to thank Dr Colin Ockleford and Dr Graham Burton for their kind contribution of Figs 16 and 29 respectively, as well as Elsevier Science Publishers BV (Amsterdam) and the editor of Scanning Micro- scopy International (Chicago), Prof. Om Johari, for permission to reproduce the micrographs.
REFERENCES
Alsat, E., Bouali, Y., Goldstein, S., Malassin6, A., Berthelier, M., Mondon, F. and Cedard, L., 1984. Low-density lipo- protein binding sites in the microvillous membranes of human placenta at different stages of gestation. Molec. cell. Endocrinol., 38, 197 203.
Alsat, E., Mondon, F., Malassin6, A., Rebourcet, R., Goldstein, S. and Cedard, L., 1987. Identification of distinct receptors for native and acetylated-low-density lipoprotein in human placental microvilli. Trophobl. Res., 2, 17-27.
Amenta, P. S., Gay, S., Vaheri, A. and Martinez-Hernandez, A., 1986. The extracellular matrix is an integrated unit: ultrastructural localization of collagen types I, III, IV, V, VI, fibronectin and laminin in human term placenta. Coll. Rel. Res., 6, 125-152.
Amoroso, E. C. and Perry, J. S., 1975. The existence during gestation of an immunological buffer zone at the interface between maternal and foetal tissues. Phil. Trans. R. Soc. B, 271, 343-361.
Anderson, W. R. and McKay, D. G., 1966. Electron micro- scopic study of the trophoblast in normal and toxemic placentas. Am. J. Obstet. Gynec., 95, 1134-1148.
Ashley, C. A., 1965. Study of the human placenta with the electron microscope: functional implications of a canal system in the trophoblast. Archs Path., 80, 377-390.
Bartel, H., Dec, W., Cieciura, L. and Krajewski, J., 1978. ultrastructure of capillaries in the villi of human placenta. Folia Morph., 37, 315-320.
Becker, V. and Seifert, K., 1965. Die Ultrastructure den Kapillarwand in der menschlichen Placenta zur Zeit der Schwangerschaftsmitte. Z. Zellforsch. mikrosk. Anat., 65, 380-396.
Beham, A., Denk, H. and Desoye, G., 1988. The distribution of intermediate filament proteins, actin and desmoplakins in human placental tissue as revealed by polyclonal and mono- clonal antibodies. Placenta, 9, 479-492.
Benzo, C. A., 1974. Annulate lamellae in hepatic and pan- creatic beta cells of the chick embryo. Am. J. Anat., 140, 139-142.
Bergstr6m, S., 1971. Surface ultrastructure of human amnion and chorion in early pregnancy. Obstet. Gynec., 38, 513 524.
Bierings, M. B., Adriaansen, H. J. and van Dijk, J. P., 1988. The appearance of transferrin receptors on cultured human cytotrophoblast and in vitro-formed syncytiotrophoblast. Placenta, 9, 387-396.
Bierings, M., Jones, C., Adriaansen, H. and van Dijk, H., 1990. Transferrin receptors on cyto- and in vitro-formed syncytiotrophoblast. Trophobl. Res. (in press).
Booth, A. G., Olaniyan, R. O. and Vanderpuye, O. A., 1980. An improved method for the preparation of human placen- tal syncytiotrophoblast microvilli. Placenta, I, 327-336.
Bourne, G., 1962. The Human Amnion and Chorion, Year Book Medical Publisher, Inc., Chicago, Chapter VIII, 94-112.

172 C . J . P . Jones and H. Fox
Bouteille, M., Kalifat, S. R. and Delarue, J., 1967. Ultra- structural variations of nuclear bodies in human diseases. J. Ultrastruct. Res., 19, 474-486.
Boyd, J. D., Boyd, C. A. R. and Hamilton, W. J., 1968a. Observations on the vacuolar structure of the human syncy- tiotrophoblast. Z. Zellforsch. mikrosk. Anat., 88, 57-79.
Boyd, J. D. and Hamilton, W. J., 1970. The Human Placenta, W. Heifer and Sons, Ltd., Cambridge.
Boyd, J. D., Hamilton, W. J. and Boyd, C. A. R., 1968b. The surface of the syncytium of the human chorionic villus. J. Anat., 102, 553-563.
Boyd, J. D. and Hughes, A. F. W., 1954. Observations on human chorionic villi using electron microscope. J. Anat., 88, 356-362.
Bradbury, F. M. and Ockleford, C. D., 1990. A confocal and conventional epifluorescence microscope study of the inter- mediate filaments in chorionic villi. J. Anat., 169, 173-189.
Bradbury, S., Billington, W. D., Kirby, D. R. S. and Williams, E. A., 1969. Surface mucin of human trophoblast. Am. J. Obstet. Gynecol., 104, 416-418.
Bradbury, S., Billington, W. D., Kirby, D. R. S. and Williams, E. A., 1970. Histochemical characterization of the surface mucoprotein of normal and abnormal human trophoblast. Histochem. J., 2, 263-274.
Braunhut, S. J., Blanc, W. A., Ramanarayanan, M., Marboe, C. and Mesa-Tejada, R., 1984. Immunocytochemical local- ization of lysozyme and alpha-l-antichymotrypsin in the term placenta: an attempt to characterize the Hofbauer cell. J. Histochem. Cytochem., 32, 1204-1210.
Brightman, M. W., 1975. Ultrastructural characteristics of adult choroid plexus: relation to the blood--cerebrospinal fluid barrier to proteins. In: The Choroid Plexus in Health and Disease, Netsky, M. G. and Shuangshoti, S. (eds), John Wright and Sons, Bristol, 86-112.
Brown, P. J. and Johnson, P. M., 1981. Fc-receptor activity of isolated human placental syncytiotrophoblast plasma membrane. Immunology, 42, 313-319.
Bulmer, J. N. and Johnson, P. M., 1984. Macrophage popu- lations in the human placenta and amniochorion. Clin. exp. Immunol., 57, 393-403.
Burgos, M. H. and Rodriguez, E. M., 1966. Specialized zones in the trophoblast of the human term placenta. Am. J. Obstet. Gynec., 96, 342-356.
Burton, G. J., 1987. The fine structure of the human placental villus as revealed by scanning electron microscopy. Scanning Microsc., 1, 1811 1828.
Carter, J. E., 1964. Morphological evidence of syncytial for- mation from the cytotrophoblastic cells. Obstet. Gynec., 23, 647456.
Castellucci, M., Celona, A., Bartels, H., Steininger, B., Benedetto, V. and Kaufmann, P., 1987. Mitosis of the Hofbauer cell: Possible implications for a fetal macrophage. Placenta, 8, 65-76.
Castellucci, M. and Kaufmann, P., 1982. A three-dimensional study of the normal human placental villous core: II) Stromal architecture. Placenta, 3, 269-286.
Castellucci, M., Martinoli, C., Foidart, J. M., Zaccheo, D. and Kaufmann, P., 1983. The connective core of the human placental villi throughout pregnancy. An ultrastructural, immuno-histochemical and immunological study. J. Anat., 137, 788-789.
Castellucci, M., Schweikhart, G., Kaufmann, P. and Zaccheo, D., 1984. The stromal architecture of the immature intermediate villus of the human placenta. Functional
and clinical implications. Gynecol. Obstet. Invest., 18, 95-99.
Castellucci, M. and Zaccheo, D., 1989. The Hofbauer cells of the human placenta: Morphological and immunological aspects. Prog. din. biol. Res., 296, 443-451.
Castellucci, M., Zaccheo, D. and Pescetto, G., 1980. A three- dimensional study of the normal human placental villous core. I. The Hofbauer cells. Cell Tissue Res., 210, 235-247.
Cavicchia, J. C., 1971. Junctional complexes in the trophoblast of the human full term placenta. J. Anat., 108, 339 346.
Charpin, C., Kopp, F., Pourreau-Schneider, N., Lissitzky, J. C., Lavaut, M. N., Martin, P. M. and Toga, M., 1985. Laminin distribution in human decidua and immature placenta: An immunoelectron microscopic study (avidin-biotin- peroxidase complex method). Am. J. Obstet. Gynecol., 151, 882-826.
Chen, L. B., Burridge, K., Murray, A., Walsh, M. L., Copple, C. D., Bushnell, A., McDougall, J. K. and Gallimore, P. H., 1978. Modulation of cell surface glycocalyx: studies on large, external, transformation-sensitive protein. Ann. N.Y. Acad. Sci., 312, 366-381.
Cheng, H. M. and Johnson, P. M., 1985. A description of human placental syncytiotrophoblast membrane glycos- phingolipids. Placenta, 6, 229~38.
Cibils, L. A., 1968. Growth of placental villi in the first trimester. Lying-In: The Journal of Reproductive Medicine, 1, 377-387.
Clark, R. K. and Damjanov, I., 1985. Intermediate filaments of human trophoblast and choriocarcinoma cell lines. Virch. Arch. A Pathol. Anat., 407, 203-208.
Contractor, S. F., Banks, R. W., Jones, C. J. P. and Fox, H., 1977. A possible role for placental lysosomes in the for- mation of villous syncytiotrophoblast. Cell Tissue Res., 178, 411-419.
Corash, L. and Gross, E., 1974. Subcellular fractionation of human placenta. 2) Isolation and density distribution of lysosomes from first trimester tissue. Pedlar. Res., 8, 774--782.
Daniels-McQueen, S., Krichevsky, A. and Boime, I., 1987. Isolation and characterisation of human cytotrophoblast cells. Troph. Res., 2, 423-445.
Dautry-Varsat, A., Ciechanover, A. and Lodish, H. F., 1983. pH and the recycling of transferrin during receptor- mediated endocytosis. Proc. natn. Acad. Sci. U.S.A., 80, 2258-2262.
Dearden, L. and Ockleford, C. D., 1983. Structure of human trophoblast: correlation with function. In: Biology of Trophoblast, Loke, Y. W. and Whyte, A. (eds), Elsevier Science Publishers BV, Amsterdam, 69-110.
Dec, W. and Bartel, H. 1976. Ultrastructure of the chorionic villus of the human placenta of the second and third trimester of pregnancy. Folia Histochem. Cytochem., 14, 327-334.
Dec, W., Bartel, H., Cieciura, L. and Krajewski, J., 1979. Electron microscopic observation of the Hofbauer cells. Acta med. pol., Vars., 20, 375-376.
De Ikonicoff, L. K., 1973. Etude histochimique du placenta humain. Localisation ultrastructale de l'hormone chori- onique somatomammotrophique (HCS) par une technique immunocytochimique A la peroxydase. C. r. hebd. Seanc. Acad. Sci., Paris (D), 276, 3355-3358.
De Ikonicoff, L. K., 1974. Localisation immunohistochimique des hormones gonadotrope chorionique (HCG) et soma- tomammotrophique (HCS) dans le placenta humain. In:

Ultrastructure of the Human Placenta 173
Human Placental Biology, De Ikonicoff, L. K. and Cedard, L. (eds), INSERM, Paris, 31-47.
Demir, R., 1980. Ultrastructure of the epithelium of the chorionic villi of the human placenta. Acta Anat., 106, 18-29.
Demir, R. and Erbengi, T., 1984. Some new findings about Hofbauer cells in the chorionic villi of the human placenta. Acta Anat., 119, 18-26.
Dempsey, E. W., 1953. Electron microscopy of human placental villi. Anat. Rec., 117, 609.
Dempsey, E. W., 1972. The development of capillaries in the villi of early human placentas. Am. J. Anat., 134, 221~38.
Dempsey, E. W. and Luse, S. A., 1971. Regional specializations in the syncytial trophoblast of early human placentas. J. Anat., 108, 545-561.
Dempsey, E. W. and Zergollern, L., 1969. Zonal regions of the human placental barrier. Anat. Rec., 163, 177.
Dreskin, R. B., Spicer, S. S. and Greene, W. B., 1970. Ultra- structural localization of chorionic gonadotropin in human term placenta. J. Histochem. Cytochem., 18, 862-874.
Duance, V. C. and Bailey, A. J., 1983. Structure of the trophoblast basement membrane. In: Biology o f Tropho- blast, Loke, Y. W. and Whyte, A. (eds), Elsevier Science Publishers BV, Amsterdam, 597~25.
Earl, U., Estlin, C. and Bulmer, J. N., 1990. Fibronectin and laminin in the early human placenta. Placenta, I1,223-231.
Edwards, H. C. and Booth, A. G., 1987. Calcium-sensitive, lipid-binding cytoskeletal proteins of the human placental microvillar region. J. Cell Biol., 105, 303 311.
Enders, A. C., 1965. Formation of syncytium from cytotro- phoblast in the human placenta. Obstet. Gynecol., 25, 378-386.
Enders, A. C. and King, B. F., 1970. The cytology of the Hofbauer cell. Anat. Rec., 167, 231 251.
Eyden, B. P., Watson, R. J., Harris, M. and Howell, A., 1986. Intralobular stromal fibroblasts in the resting human mammary gland: ultrastructural properties and intercellular relationships. J. submicrosc. Cytol., 18, 397-408.
Farquhar, M. G., 1969. Lysosome function in regulating se- cretion: disposal of secretory granules in cells of the anterior pituitary gland. In: Lysosomes in Biology and Pathology, Vol. 2, Dingle, J. T. and Fell, H. B. (eds), North Holland Publishing Company, Amsterdam, 463-482.
Faulk, W. P. and Johnson, P. M., 1977. Immunological studies of human placentae: identification and distribution of pro- teins in mature chorionic villi. Clin. exp. Immunol., 27, 365-375.
Feller, A. C., Schneider, H., Schmidt, D. and Parwaresch, M. R., 1985. Myofibroblasts as a major cellular constituent of villous stroma in human placenta. Placenta, 6, 405-415.
Ferenczy, A. and Richart, R. M., 1972. Scanning electron microscopic study of normal and molar trophoblast. Gynecol. Oncol., 1, 95 110.
Forgac, M., Cantley, L., Wiedenmann, B., Altstiel, L. and Branton, D., 1983. Clathrin-coated vesicles contain an ATP-dependent proton pump. Proc. natn. Acad. Sci. U.S.A., 80, 1301~1303.
Fox, H. and Agrafojo-Blanco, A., 1974. Scanning electron microscopy of the human placenta in normal and abnormal pregnancies. Eur. J. Obstet. Gynecol. Reprod. Biol., 4, 45-50.
Fox, H. and Kharkongor, N. F., 1969. Enzyme histochemistry of the Hofbauer cells of the human placenta. J. Obstet. Gynaecol. Br. Commonw., 76, 918421.
Friend, D. A. and Farquhar, M. G., 1967. Functions of coated vesicles during protein absorption in the rat vas deferens. J. Cell Biol., 35, 357-376.
Fujimoto, S., Hamasaki, K., Ueda, H. and Kagawa, H., 1986. Immunoelectron microscope observations on secretion of human placental lactogen (hPL) in the human chorionic villus. Anat. Rec., 216, 68~2.
Galton, M., 1962. DNA content of placental nuclei. J. Cell Biol., 13, 183 191.
Garancis, J. C., Pattillo, R. A., Hussa, R. O., Schultz, J. and Mattingly, R. F., 1970. Electron microscopic and biochemi- cal patterns of the normal and malignant trophoblast. Am. J. Obstet. Gynecol., 108, 1257-1268.
Geuze, H. J., Slot, J. W., Strous, G. J. A. M., Lodish, H. F. and Schwartz, A. L., 1982. Intracellular site of asialoglyco- protein receptor-ligand uncoupling: double-label immuno- electron microscopy during receptor-mediated endocytosis. Cell, 32, 277-287.
Gey, G. O., Seegar, G. E. and Hellman, L. M., 1938. Pro- duction of gonadotropic substance (prolan) by placental cells in tissue culture. Science, 88, 306-307.
Goldstein, J. L., Anderson, R. G. W. and Brown, M. S., 1979. Coated pits, coated vesicles and receptor-mediated endocytosis. Nature, Lond., 279, 679~585.
Goldstein, J., Braverman, M., Salafia, C. and Buckley, P., 1988. The phenotype of human placental macrophages and its variation with gestational age. Am. J. Path., 133, 648~i59.
Guller, S., Gravanis, A. and Gurpide, E., 1986. Steroid metab- olizing enzymes associated with the microvillar membrane of human placenta. J. Steroid Biochem., 24, 935-944.
Hackenbrock, C. R., 1966. Ultrastructural bases for metaboli- cally linked mechanical activity in mitochondria. I) Revers- ible ultrastructural changes with change in metabolic steady state in isolated liver mitochondria. J. Cell Biol., 30, 269 297.
Hackenbrock, C. R., 1968. Ultrastructural bases for metaboli- cally linked mechanical activity in mitochondria. 2) Electron transport-linked ultrastructural transformations in mito- chondria. J. Cell Biol., 37, 345 369.
Haigh, M., Chawner, L. E. and Fox, H., 1984. The human placenta does not contain lipofuscin pigment. Placenta, 5, 459-464.
Haigh, M., Taylor, C. J. and Fox, H., 1989. A morphometric analysis of age-related changes in villous syncytiotropho- blastic subcellular organelles in the human placenta using a computerised image analysis system. The Fetus, 1, 27-42.
Hamanaka, N., Tanizawa, O., Hashimoto, T., Yoshinare, S. and Okudaire, Y., 1971. Electron microscopic study on the localization of human chorionic gonadotropin (HCG) in the chorionic tissue by enzyme-labeled antibody technique. J. Electron Microsc., 20, 128 130.
Hamasaki, K., Okamura, Y., Ueda, H., Kagawa, H. and Fujimoto, S., 1987. Immunocytochemistry of human chorionic gonadotropin in human chorionic villi. Am. J. Obstet. Gynecol., 156, 479-483.
Haour, F. and Bertrand, J., 1974. Insulin receptors in the plasma membranes of human placenta. J. clin. Endocr. Metab., 38, 334 337.
Harbison, M. L., Godleski, J. J., Mortara, M. and Brain, J. D., 1984. Correlation of lung macrophage age and surface antigen in the hamster. Lab. Invest., 50, 653~558.
Hashimoto, M., Kosaka, M., Mori, Y., Komori, A. and Akashi, K., 1960a. Electron microscopic studies on the

174 C . J . P . Jones and H. Fox
epithelium of the chorionic villi of the human placenta (Report 1). J. Jap. obstet, gynaecol. Soc., 7, 44-52.
Hashimoto, M., Kosaka, M., Shimoyama, T., Hirasawa, T., Komori, A., Kawasaki, T. and Akashi, K., 1960b. Electron microscopic studies on the epithelium of the chorionic villi of the human placenta (Report 2). J. Jap. obstet, gynaecol. Soc., 7, 122 129.
Hedman, K., Vaheri, A. and Wartiovaara, J., 1978. External fibronectin of cultured human fibroblasts is predominantly a matrix protein. J. Cell Biol., 76, 748-760.
Heinrich, D., Metz, J., Raviola, E. and Forssmann, W. G., 1976. Ultrastructure of perfusion-fixed fetal capillaries in the human placenta. Cell Tissue Res., 172, 157 169.
Hempel, von E. and Geyer, G., 1969. Submikroskopische Verteilung der alkalischen Phosphatase in der menschlichen Placenta. Acta Histochern., 34, 138 147.
Herbst, R., Multier, A-M. and Hormann, G., 1968. Die men- schlichen Plazentazotten des 2. Schwangerschaftstrimenon im elektronen-optischen Bild. Z. Geburtsch. Gyniik., 169, 1-16.
Hertig, A. T., 1935. Angiogenesis in the early human chorion and in the primary placenta of the macaque monkey. Contr. Embryol., 25, 37-81.
Hofbauer, J., 1905. Grundziige einer Biologic der menschlichen Plazenta mit besonderer Beriicksichtigung der Fragen der f6talen Erniihrung, Braumfiller, W. (ed.), Wien and Leipzig.
Hopkins, C. R., 1986. Membrane boundaries involved in the uptake and intracellular processing of cell surface receptors. Trends in Biochemical Sciences, 11, 473-477.
H6rmann, G., 1953. Ein Beitrag zur funktionellen Morphologie der menschlichen Placenta. Arch. Gynaekol., 184, 109-123.
Hoshina, M., Boothby, M. and Boime, I., 1982. Cytological localization of chorionic gonadotropin alpha and placental lactogen mRNAs during development of the human placenta. J. Cell Biol., 93, 190-198.
Hoshina, M., Hussa, R., Pattillo, R. and Boime, I. 1983. Cytological distribution of chorionic gonadotropin subunit and placental lactogen messenger RNA in neoplasms derived from the human placenta. J. Cell Biol., 97, 1200-1206.
Hoshina, M., Boothby, M., Hussa, R., Pattillo, R., Camel, H. M. and Boime, I., 1985. Linkage of human chorionic gonadotropin and placental lactogen biosynthesis to trophoblast differentiation and tumorigenesis. Placenta, 6, 163-172.
Hoyes, A. D., 1971. Ultrastructure of the mesenchymal layers of the human chorion laeve. J. Anat., 109, 17-30.
Hulstaert, C. E., Torringa, J. L., Koudstaal, J., Hardonk, M. J. and Molenaar, I., 1973. The characteristic distribution of alkaline phosphatase in the full-term human placenta. Gynecol. Invest., 4, 24-30.
Ikawa, A., 1959. Observations on the epithelium of human chorionic villi with the electron microscope. J. Jap. obstet. gynaecol. Soc., 6, 219-234.
Inoue, T., 1960. Observations on human placenta chorionic villi under the electron microscope. J. Jap. obstet, gynaecol. Soc., 7, 173-174.
Isemura, M., Yamaguchi, Y. and Munakata, H. 1985. Distri- bution of fibronectin and other connective tissue components in human placenta. Tohoku J. exp. Med., 145, 373 379.
Jasper, D. K., 1975. Mitochondrial morphological changes in mucosal epithelial cells. Experientia, 31, 35 36.
Jemmerson, R., Klier, F. G. and Fishman, W. H., 1985. Clustered distribution of human placental alkaline phos- phatase on the surface of both placental and cancer cells.
Electron microscopic observations using gold-labelled anti- bodies. J. Histochem. Cytochem., 33, 1227-1234.
Johnson, P. M. and Faulk, W. P., 1976. Immunological studies of human placentae. Lectin binding to villous stroma and to trophoblastic basement membrane. Clin. exp. Immunol., 24, 435~,40.
Johnson, P. M. and White, C. M., 1979. Distribution of lectin binding on human placental tissue sections. Protides biol. Fluid~, 27, 503-506.
Johnson, S. A. and Wooding, F. B. P., 1988. Synthesis and storage of chorionic gonadotrophin and placental lactogen in human syncytiotrophoblast. J. Physiol., 403, 29P.
Jones, C. J. P., 1976. An Ultrastructural and Ultrahistochemical Study o f the Human Placenta in Normal and Abnormal Pregnancy. PhD Thesis, University of Manchester.
Jones, C. J. P. and Fox, H., 1976. An ultrahistochemical study of the distribution of acid and alkaline phosphatases in placentae from normal and complicated pregnancies. J. Pathol., 118, 143-151.
Jones, C. J. P. and Fox, H., 1977. Syncytial knots and inter- villous bridges in the human placenta: an ultrastructural study. J. Anat., 124, 275-286.
Jones, C. J. P. and Fox, H., 1978. An ultrahistochemical study of the placental content of respiratory enzymes in normal and prolonged pregnancies. Invest. Cell Pathol., 1, 217 225.
Jones, C. J. P. and Ockleford, C. D., 1985. Nematosomes in the human placenta. Placenta, 6, 355-362.
Jones, C. J. P., Sear, C. H. J. and Grant, M. E., 1980. An ultrastructural study of fibroblasts derived from bovine ligamentum nuchae and their capacity for elastogenesis in culture. J. Pathol., 131, 35-53.
Jones, C. J. P., Stoddart, R. W., Bierings, M. B. and van Dijk, H. P., 1990. Expression of glycoproteins on and within placental cytotrophoblast cells in culture. Trophobl. Res. (in press).
Kanaseki, T., and Kadota, K., 1969. The 'vesicle in a basket'. A morphological study of the coated vesicle isolated from nerve endings of guinea-pig brain with special reference to mechanisms of membrane movement. J. Cell Biol., 42, 202 220.
Kastschenko, N., 1885. Das menschliche Chorionepithel und dessen Rolle bei der Histogenese der Placenta. Arch. Anat. Physiol. Jahr 1885, 451-480.
Katabuchi, H., Naito, M., Miyamura, S., Takahashi, K. and Okamura, H., 1989. Macrophages in human chorionic villi. Prog. clin. biol. Res., 296, 453-458.
Kaufmann, P., 1985. Influence of ischaemia and artificial perfusion on placental ultrastructure and morphometry. Contrib. Gynecol. Obstet., 13, 18-26.
Kaufmann, P., Sen, D. K. and Schweikhart, G., 1979. Classifi- cation of human placenta villi. 1) Histology, Cell Tissue Res., 200, 409423.
Kaufmann, P., Stark, J. and Stegner, H. E., 1977. The villous stroma of the human placenta. 1) The ultrastructure of fixed connective tissue cells. Cell Tissue Res., 177, 105-121.
Kaufmann, P. and Stegner, H. E., 1972. ~ber die funktionelle Differenzierung des Zottensyncytiums in der menslichen Placenta. Z. Zellforsch. mikrosk. Anat., 135, 361 382.
Kawagoe, K., Akiyama, J., Kawamoto, T., Morishita, Y. and Mori, S., 1990. Immunohistochemical demonstration of Epidermal Growth Factor (EGF) receptors in normal human placental villi. Placenta, 11, 7-15.

Ultrastructure of the Human Placenta 175
Kedzia, H. and Pilarski, T., 1971. Activity and distribution of hydrolytic enzymes in the developing human placenta. Pol. Endocr., 22, 374-385.
Keene, D. R., Sakai, L. Y., B/ichinger, H. P. and Burgeson, R. E., 1987. Type III collagen can be present on banded collagen fibrils regardless of fibril diameter. J. Cell Biol., 105, 2393 2402.
Kefalides, N. A., 1973. Structure and biosynthesis of basement membrane. Int. Rev. Connect. Tissue Res., 6, 63 104.
Khalfoun, B., Lacord-Bonneau, M., Degenne, D., Clement, L. and Bardos, P., 1986. Characterization of the human syncy- tiotrophoblast plasma membrane associated components. Int. J. Biochem., 18, 351-360.
Khong, T. Y., Lane, E. B. and Robertson, W. B., 1986. An immunocytochemical study of fetal cells at the maternal- placental interface using monoclonal antibodies to keratins, vimentin and desmin. Ceil Tissue Res., 246, 189-195.
King, B. F., 1981. The distribution and mobility of anionic sites on the surface of human placental syncytial trophoblast. Anat. Rec., 199, 15-22.
King, B. F., 1982a. Morphologic and cytochemical identifi- cation of microperoxisomes in human and non-human primate placental trophoblast. Histochemistry, 74, 115 121.
King, B. F., 1982b. Absorption of peroxidase-conjugated im- munoglobulin G by human placenta: an in vitro study. Placenta, 3, 395~,06.
King, B. F., 1983. The organization of actin filaments in human placental villi. J. Ultrastruct. Res., 85, 320 328.
King, B. F. and Enders, A. C., 1971. Protein absorption by the guinea pig chorioallantoic placenta. Am. J. Anat., 130, 409~,30.
King, A. and Loke, Y. W., 1988. Differential expression of blood-group-related carbohydrate antigens by trophoblast subpopulations. Placenta, 9, 513 521.
Kliman, H. J., Feinman, M. A. and Strauss, J. F., 1987. Differentiation of human cytotrophoblast into syncytiotro- phoblast in culture. Trophobl. Res., 2, 407~,21.
Knoth, M., 1968. Ultrastructure of chorionic villi from a four- somite human embryo. J. Ultrastruct. Res., 25, 423~,40.
Kurosawa, K., lsemura, M. and Yamaguchi, Y., 1985. Changes in distribution of connective tissue components of human placenta with maturation. Tohoku J. exp. Med., 147, 261-265.
Lazarides, E., 1982. Intermediate filaments: a chemically heterogeneous, developmentally regulated class of proteins. A. Rev. Biochem., 51, 219-250.
Lee, M-C. and Damjanov, I., 1984. Lectin histochemistry of human placenta. Differentiation, 28, 123 128.
Leivo, I., Laurila, P., Wahlstr6m, T. and Engvall, E., 1989. Expression of merosin, a tissue-specific basement membrane protein, in the intermediate trophoblast cells of chorio- carcinoma and placenta. Lab. Invest., 60, 783 790.
Lewis, W. H., 1924. Hofbauer cells (clasmatocytes) of the human chorionic villus. Bull. Johns Hopkins Hosp., 35, 183-185.
Liebhart, M., 1974a. Polysaccharide surface coat (glycocalix) of human placental villi. Pathol. Eur., 9, 3 5.
Liebhart, M., 1974b. Ultrastructure of the stromal connective tissue of normal placenta and of placenta in diabetes mellitus of mother. Pathol. Eur., 9, 177-184.
Lin, C-T., 1980. Immunoelectron microscopic localization of immunoglobulin G in human placenta. J. Histochem. Cytochern., 28, 339 347.
Lister, U. M., 1963a. Ultrastructure of the human mature
placenta. 1) The maternal surface. J. Obstet. Gynaecol. Br. Commonw., 70, 373 386.
Lister, U. M., 1963b. Ultrastructure of the human mature placenta. 2) The foetal surface. J. Obstet. Gynaecol. Br. Commonw., 70, 766-776.
Lister, U. M., 1964a. Ultrastructure of the early human placenta. J. Obstet. Gynaecol. Br. Commonw., 71, 21 32.
Lister, U. M., 1964b. Structural changes in the capillaries of human chorionic villi occurring with age. J. R. microsc. Soc., 83, 455458.
Lister, U. M., 1967. The localization of placental enzymes with the electron microscope. J. Obstet. Gynaecol. Br. Commonw., 74, 34-39.
Loh, T. T., Higuchi, D. A., van Bocksmeer, F. M., Smith, C. M. and Brown, E. B., 1980. Transferrin receptors on the human placental microvillous membrane. J. clin. Invest., 65, 1182 1191.
Loke, Y. W., Eremin, O., Ashby, J. and Day, S., 1982. Characterization of the phagocytic cells isolated from the human placenta. J. Reticuloendothel. Soc., 31, 317 324.
Luckett, W. P., 1978. Origin and differentiation of the yolk sac and extra-embryonic mesoderm in presomite human and rhesus monkey embryos. Am. J. Anat., 152, 59-98.
Madri, J. A., Foellmer, H. G. and Furthmayr, H., 1982. Type V collagens of the human placenta: trimer alpha- chain composition, ultrastructural morphology and peptide analysis. Coll. Rel. Res., 2, 19 29.
Magid, M., Nanny, L. B., Stoscheck, C. M. and King, L. E., 1985. Epidermal growth factor binding and receptor distri- bution in term human placenta. Placenta, 6, 519 526.
Malassin6, A., Besse, C., Roche, A., Alsat, E., Rebourcet, R., Mondon, E. and Cedard, L., 1987. Ultrastructural visualisa- tion of the internalization of low density lipoprotein by human placental cells. Histochemistry, 87, 457~,64.
Martin, B. J. and Spicer, S. S., 1973a. Multivesicular bodies and related structures of the syncytiotrophoblast of human term placenta. Anat. Rec., 175, 15-36.
Martin, B. J. and Spicer, S. S., 1973b. Ultrastructural features of cellular maturation and aging in human trophoblast. J. Ultrastruct. Res., 43, 133--149.
Martin, B. J., Spicer, S. S. and Smythe, N. M., 1974. Cytochem- ical studies of the maternal surface of the syncytiotro- phoblast of human early and term placenta. Anat. Rec., 178, 769-785.
Martinek, J. J., Gallagher, M. L. and Essig, G. F., 1975. An electron microscopic study of fetal capillary basal laminas of "normal" human term placentas. Am. J. Obstet. Gynecol., 121, 17-24.
Martinoli, C., Castellucci, M., Zaccheo, D. and Kaufmann, P., 1984. Scanning electron microscopy of stromal cells of human placental villi throughout pregnancy. Ceil Tissue Res., 235, 647~i55.
Matre, R., T6nder, O. and Endresen, C., 1975. Fc receptors in the human placenta. Scand. J. lmmunol., 4, 741 745.
McKay, D. G., Hertig, A. T., Adams, E. C. and Richardson, M. V., 1958. Histochemical observations on the human placenta. Obstet. Gynecol., 12, 1 36.
Metz, J. and Weihe, E., 1980. Intercellular junctions in the full-term human placenta. 2) Cytotrophoblast cells, intra- villous stroma ceils and blood vessels. Acta Embryol. (Berl), 158, 167-178.
Metz, J., Weihe, E. and Heinrich, D., 1979. Intercellular junctions in the human full-term placenta. 1) Syncytiotro- phoblastic layer. Acta Embryol. (Berl), 158, 41 50.

176 C . J . P . Jones and H. Fox
Midgley, A. R., Pierce, G. B., Deneau, G. A. and Gosling, J. R. G., 1963. Morphogenesis of syncytiotrophoblast in vivo: an autoradiographic demonstration. Science, 141, 349-350.
Morinaga, S., Haga, N. and Sasano, N., 1984. Ultra- structural localization of hCG in the placenta and in testicular germ cell tumours. Virch. Arch. B Cell Pathol., 45, 397-406.
Morrish, D. W., Marusyk, H. and Siy, O., 1987. Demon- stration of specific secretory granules for human chorionic gonadotropin in placenta. J. Histochem. Cytochem., 35, 93-101.
Morrish, D. W., Marusyk, H. and Bhardwaj, D., 1988. Ultrastructural localization of human placental lactogen in distinctive granules in human term placenta: comparison with granules containing human chorionic gonadotropin. J. Histochem. Cytochem., 36, 193 197.
Moskalewski, S., Ptak, W. and Czarnik, Z., 1975. Demon- stration of cells with IgG receptor in human placenta. Biol. Neonate, 26, 268~73.
Mues, B., Langer, D., Zwaldo, G. and Sorg, C., 1989. Phenotypic characterization of macrophages in human term placenta. Immunology, 67, 303-307.
Nagy, T., Boros, A. and Benko, K., 1965. Elektronen- mikroskopische Untersuchung junger und reifer men- schlicher Placenten. Arch. Gynaekol., 200, 428-440.
Nakano, M., 1963. Electron microscopic study of the transport mechanism of the human placental villi. Tohoku J. exp. Med., 78, 398-409.
Nelson, D. M., Smith, C. H., Enders, A. C. and Donohue, T. M., 1975. The non-uniform distribution of acidic com- ponents on the human placental syncytial trophoblast surface membrane. A cytochemical and analytical study. Anat. Rec., 184, 159 182.
Ockleford, C. D., 1976. A three-dimensional reconstruction of the polygonal pattern on placental coated vesicle membranes. J. Cell Sci., 21, 83-91.
Ockleford, C. D., 1981. Coated vesicles. In: Electron Micro- scopy of Proteins, Vol. 2, Harris, R. J. (ed.), Academic Press, London, 255~99.
Ockleford, C. D. and Dearden, L., 1984. The acquisition of immunity by the fetus. In: Immunological Aspects of Repro- duction in Mammals, Crichton, D. B. (ed.), Butterworth, London, 251-263.
Ockleford, C. D. and Menon, G., 1977. Differentiated regions of human placental cell surface associated with exchange of materials between maternal and foetal blood: a new organelle and the binding of iron. J. Cell Sci., 25, 279-291.
Ockleford, C. D., Nevard, C. H. F., Indans, I. and Jones, C. J. P., 1987. The structure and function of the nemato- some. J. Cell Sci., 87, 27-44.
Ockleford, C. D. and Wakely, J., 1982. The skeleton of the placenta. In: Progress in Anatomy, Vol. 2, Harrison, R. J. and Navaratnam, V. (eds), Cambridge University Press, Cambridge, 19-47.
Ockleford, C. D., Wakely, J. and Badley, R. A., 1981. Morpho- genesis of human placental chorionic villi: cytoskeletal, syncytioskeletal and extracellular matrix proteins. Proc. R. Soc. B, 212, 305-316.
Ockleford, C. D. and Whyte, A., 1977. Differentiated regions of human placental cell surface associated with exchange of material between maternal and foetal blood: coated vesicles. J. Cell Sci., 25, 293 312.
Panigel, M. M. and Anh, J. N. H., 1963. Precisions sur l'~volution de l'ultrastructure des elements trophoblastiques et sur leurs modifications superficielles dans le placenta humain. C. r. hebd. Seanc. Acad. Sci., Paris., 257, 3669-3671.
Panigel, M. M. and Anh, J. N. H., 1964a. Ultrastructure des villosit6s placentaires humaines. Pathol. Biol., Paris, 12, 927 949.
Panigel, M. M. and Anh, J. N. H., 1964b. Ultrastructure des cellules de Hofbauer dans le placenta humain. C. r. hebd. Seanc. Acad. Sci., Paris., 258, 3556-3558.
Parmley, R. T., Takagi, M. and Denys, F. R., 1984. Ultra- structural localization of glycosaminoglycans in human term placenta. Anat. Rec., 210, 477-484.
Pattillo, R. A., Hussa, R. O., Yorde, D. E. and Cole, L. A., 1983. Hormone synthesis by normal and neoplastic human trophoblast. In: Biology of Trophoblast, Loke, Y. W. and Whyte, A. (eds), Elsevier Science Publishers BV, Amsterdam, 283-316.
Pearse, B. M. F., 1976. Clathrin: a unique protein associated with intracellular transfer of membrane by coated vesicles. Proc. natn. Acad. Sci. U.S.A., 73, 1255 1259.
Pearse, B. M. F., 1982. Coated vesicles from human placenta carry ferritin, transferrin, and immunoglobulin G. Proc. natn. Acad. Sci. U.S.A., 79, 451-455.
Pfister, C., Scheuner, G., Bahn, H. and Stiller, D., 1988. Immunohistochemical demonstration of fibronectin in the human placenta. Acta Histochem., 84, 83 92.
Pierce, G. B. and Midgley, A. R., 1963. The origin and function of human syncytiotrophoblast giant cells. Am. J. Pathol., 43, 153-173.
Pierce, G. B., Midgley, A. R. and Beals, T. F., 1964. An ultrastructural study of differentiation and maturation of trophoblast of the monkey. Lab. Invest., 13, 451~,64.
Ramsey, E. M., 1987. Development and anatomy of the placenta. In: Haines and Taylor Obstetrical and Gynaecolog- ical Pathology, Fox, H. (ed.), Churchill, Livingstone, Edinburgh, 959471.
Rao, C. V., Ramani, N., Chegini, N., Stadig, B. K., Carman, F. R., Woost, P. G., Schultz, G. S. and Cook, C. L., 1985. Topography of human placental receptors for epidermal growth factor. J. biol. Chem., 260, 1705-1710.
Reale, E., Wang, T., Zaccheo, D., Maganza, C. and Pescetto, G., 1980. Junctions on the maternal blood surface of the human placental syncytium. Placenta, I, 245 258.
Rhodin, J. A. G. and Terzakis, J., 1962. The ultrastructure of the human full-term placenta. J. Ultrastruct. Res., 6, 88-106.
Richards, R. C., Beardmore, J. M., Brown, P. J., Molloy, C. M. and Johnson, P. M., 1983. Epidermal growth factor receptors on isolated human placental syncytiotrophoblast plasma membrane. Placenta, 4, 133 138.
Rigano, A. and Sermann, R., 1963. Ultrastruttura e funzione dei capillari sel villo coriale umano. Riv. ostet. Ginec., 18, 475-498.
Risk, J. M. and Johnson, P. M., 1985. Antigen expression by human trophoblast and tumour cells: models for gene regulation? Contrib. Gynecol. Obstet., 14, 74 82.
Rovasio, R. A. and Monis, B., 1973. Cytochemical changes of a glycocalyx of human placenta with maturation. Experientia, 29, 1115-1117.
Rukosuev, V. S., Fokin, E. I. and Milovanov, A. P., 1989. Immunofluorescence study of the extracellular matrix of the human placenta. Ontogenez, 20, 171 178.

Ultrastructure of the Human Placenta 177
Salazar, H. and Gonzalez-Angulo, A., 1967. The fine structure of human chorionic villi and placental transfer of iron in late pregnancy. Am. J. Obstet. Gynecol., 97, 851-865.
Sawasaki, C., Mori, T., lnoue, T. and Shinmi, K., 1957. Observations on the human placental membrane under the electron microscope. Endocr. jap., 4, I 11.
Sideri, M., Zannoni, E. and Challier, J. C., Transfer of horse- radish peroxidase across the human placental cotyledon perfused in vitro. Trophobl. Res., 2, 573-584.
Smith, C. H., Nelson, D. M., King, B. K., Donohue, T. M., Ruzycki, S. M. and Kelly, L. K., •977. Characterization of a microvillous membrane preparation from human placen- tal syncytiotrophoblast: a morphologic, biochemical and physiologic study. Am. J. Obstet. Gynecol., 128, 190 196.
Smith, N. C. and Brush, M. G., 1973. Progesterone-macro- molecular interactions in preparations of human placental villous surface membrane. J. Endocr., 59, xvii.
Smith, N. C., Brush, M. G. and Luckett, S., 1974. Preparation of human placental villous surface membrane. Nature, Lond., 252, 302-303.
Stephens, R. E. and Edds, K. T., 1976. Microtubules: structure, chemistry and function. PhysioL Rev., 56, 709 777.
Strauss, L., Goldenberg, N., Hirota, K. and Okudaira, Y., 1965. Structure of the human placenta with observations on ultrastructure of the terminal chorionic villus. Birth Defects' Original Article Series, 1, 13-26.
Suemizu, H., Osamura, Y. and Watanabe, K., 1988. Ultra- structural localization of HCG and subunits in the human placenta and choriocarcinoma cell lines. Morphological approach to secretory pathway. Acta Histochem. Cytochem., 21, 265 272.
Tao, T-W. and Hertig, A. T., 1965. Viability and differentiation of human trophoblast in organ culture. Am. J. Anat., ! 16, 315 327.
Tennyson, V., 1975. Ultrastructural characteristics of the telen- cephalic and myelencephalic choroid plexuses in fetus of man and rabbit, and a comparison with the adult choroid plexus in rabbit. In: The Choroid Plexus in Health and Disease, Netsky, M. G. and Shuangshoti, S. (eds), John Wright and Sons, Bristol, 36-71.
Terzakis, J. A., 1963, The ultrastructure of normal human first trimester placenta. J. Ultrastruct, Res., 9, 268-284.
Tighe, J. R., Garrod, P. R. and Curran, R. C., 1967. The trophoblast of the human chorionic villus. J. Pathol. Bact., 93, 559-567.
Topilko, A. and Pisarski, T., 1967. Structure of chorionic villi in early pregnancy under light and electron microscopes. Bull. Soc. Amis. Lett. Poznan. Ser. C, 16, 115--127.
Truman, P. and Ford, H. C., 1984. The brush border of the human term placenta. Bioch#n. biophys. Acta, 779, 139-160.
Truman, P. and Ford, H. C., 1986a. Proteins of human placental microvilli. 1) Cytoskeletal proteins. Placenta, 7, 95-110.
Truman, P. and Ford, H. C., 1986b. Proteins of human placental microvilli. 2) Identification and topology of the plasma membrane proteins. Placenta, 7, 111-120.
Turkewitz, A. P. and Harrison, S. C., 1989. Concentration of transferrin receptor in human placental coated vesicles. J. Cell Biol., 108, 2127-2135.
Vacek, Z., 1965. Submicroscopic structure of the human chorion from third month of gestation to term. Folia morph., 13, 104 115.
Vacek, Z., 1970. Derivation and ultrastructure of the stroma cells of the human chorionic villus. Folia morph., 18, 1-13.
Vanderpuye, O. A., Kelley, L. K. and Smith, C. H., 1986. Transferrin receptors in the basal plasma membrane of the human placental syncytiotrophoblast. Placenta, 7, 391-403.
Von der Mark, H., Aumailley, M., Wick, G., Fleischmajer, R. and Timpl, R., 1984. Immunochemistry, genuine size and tissue localization of collagen VI. Eur. J. Biochem., 142, 493-502.
Wada, H. G., Hass, P. E. and Sussman, H. H., 1979. Transfer- rin receptors in human placental brushborder membranes. J. biol. Chem., 254, 12629 12635.
Wakitani, T., 1961. Electron microscopic observation on the chorionic villi of the normal human placenta and chorio- epithelioma malignum. 1) Electronmicroscopic observation on the chorionic villi of the normal human placenta. J. Jap. obstet. Gynaecol. Sot., 8, 208-224.
Wang, T. and Schneider, J., 1987. Cellular junctions on the free surface of human placental syncytium. Arch. Gynaekol., 240, 211-216.
Webb, P. D., McLaughlin, P. J., Risk, J. M. and Johnson, P. M., 1986. Isolation of placental-type alkaline phospha- tase associated with human syncytiotrophoblast membranes using monoclonal antibodies. Placenta, 7, 405-415.
Webb, P. D., Hole, N., McLaughlin, P. J., Stern, P. L. and Johnson, P. M., 1987. Biochemical and immunological aspects of the human trophoblast cell surface. A review. Trophobl. Res., 2, 3-15.
Webb, P. D. and Mahadevan, L. C., 1987, Calcium-dependent binding proteins associated with human placental syncy- tiotrophoblast microvillous cytoskeleton. Biochem. biophys. Acta, 916, 288 297.
Weinstock, M. and Leblond, C. P., 1974. Formation of collagen. Fedn Proc., 33, 1205-1218.
Whitsett, J. A. and Lessard, J. L., 1978. Characteristics of the microvillous brush border of human placenta: insulin recep- tor localization in brush border membranes. Endocrinology, 103, 1458-1468.
Whyte, A., 1980. Lectin binding by microvillous membranes and coated-pit regions of human syncytial trophoblast. Histochem. J., 12, 599~)07.
Whyte, A., 1983. Biochemistry of the human syncytiotro- phoblast plasma membrane. In: Biology of Trophoblast, Loke, Y. W. and Whyte, A. (eds), Elsevier Science Publishers BV, Amsterdam, 513--533.
Whyte, A. and Loke, Y. W., 1978. Increased expression of surface glycopeptides of human trophoblast compared with fetal cells from the same conceptus. J. exp. Med., 148, 1087-1092.
Whyte, A., Loke, Y. W. and Stoddart, R. W., 1978. Saccharide distribution in human trophoblast demonstrated using fluorescein-labelled lectins. Histochem. J., 10, 417-423.
Widmaier, G., 1969. Zur Ultrastruktur der menschlichen Placentazotten. Arch. Gynaekol., 207, 513 527.
Wigglesworth, J. S., 1973. Development, anatomy and physiology of the placenta. In: Postgraduate Obstetrical and Gynaecological Pathology, Fox, H. and Langley, F. A. (eds), Pergamon, Oxford, 391-408.
Wild, A. E., 1975. Mechanism of protein transport across the rabbit yolk sac endoderm. In: Maternofoetal Transmission of Immunoglobulins, Hemmings, W. A. (ed.), Cambridge University Press, London, 155 167.
Wild, A. E., 1980. Coated vesicles: a morphologically distinct subclass of endocytic vesicles. In: Coated Vesicles, Ockleford, C. D.and Whyte, A. (eds), Cambridge University Press, Cambridge and London, 154.

178 C . J . P . Jones and H. Fox
Wild, A. E., 1981. Endocytic mechanisms in protein transfer across the placenta. Placenta (Supplement 1), 165-186.
Wild, A. E., 1983. Trophoblast cell surface receptors. In: Biology of Trophoblast, Loke, Y. W. and Whyte, A. (eds), Elsevier Science Publishers BV, Amsterdam, 471-512.
Willingham, M. C. and Pastan, I., 1980. The receptosome: an intermediate organelle of receptor-mediated endocytosis in cultured fibroblasts. Cell, 21, 67-77.
Wischnitzer, S., 1970. The annulate lamellae. Int. Rev. Cytol., 27, 65 100.
Wislocki, G. B. and Bennett, H. S., 1943. The histology and cytology of the human and monkey placenta, with special reference to the trophoblast. Am. J. Anat., 73, 335~449.
Wislocki, G. B. and Dempsey, E. W., 1946. Histochemical age-changes in normal and pathological placental villi (hydatidiform mole, eclampsia). Endocrinology, 38, 90 109.
Wislocki, G. B. and Dempsey, E. W., 1955. Electron micro- scopy of the human placenta. Anat. Rec., 123, 133-167.
Wood, G. W., 1980. Mononuclear phagocytes in the human placenta. Placenta, 1, 113 123.
Wynn, R. M., 1967. Derivation and ultrastructure of the so-called Hofbauer cell. Am. J. Obstet. Gynecol., 97, 235-248.
Wynn, R. M., 1972. Cytotrophoblastic specialisations: an ultra- structural study of the human placenta. Am. J. Obstet. Gynecol., 114, 339 355.
Yamada, T., Isemura, M., Yamaguchi, Y., Munakata, H., Hayashi, N. and Kyogoku, M., 1987. Immunochemical localization of fibronectin in the human placentas at their different stages of maturation. Histochemistry, 86, 579 584.
Yamaguchi, Y., Isemura, M., Yosizawa, Z., Kurosawa, K., Yoshinaga, K., Sato, A. and Suzuki, M., 1985. Changes in the distribution of fibronectin in the placenta during normal human pregnancy. Am. J. Obstet. Gynecol., 152, 715 718.
Yorde, D. E., Hussa, R. O., Garancis, J. C. and Pattillo, R. A., 1979. Immunocytochemical localization of human chorio- gonadotropin in human malignant trophoblast. Model for human choriogonadotropin secretion. Lab. Invest., 40, 391-398.