efficient identification of malassezia yeasts by matrix-assisted laser desorption ionization-time of...
TRANSCRIPT

These articles have been accepted for publication in the British Journal of Dermatology and are currently being edited and typeset. Readers should note that articles published below have been fully refereed, but have not been through the copy-editing and proof correction process. Wiley-Blackwell and the British Association of Dermatologists cannot be held responsible for errors or consequences arising from the use of information contained in these articles; nor do the views and opinions expressed necessarily reflect those of Wiley-Blackwell or the British Association of Dermatologists This article is protected by copyright. All rights reserved.
Article type : Original Article
Efficient identification of Malassezia yeasts by matrix-assisted laser desorption ionization-
time of flight mass spectrometry (MALDI-TOF MS)
A. Kolecka1, K. Khayhan1,2, M. Arabatzis3, A. Velegraki3, M. Kostrzewa4, A. Andersson5, A.
Scheynius5, C. Cafarchia6, R. Iatta6, M.T. Montagna7, S. Youngchim8, F.J. Cabañes9, P.
Hoopman1, B. Kraak1, M. Groenewald1, T. Boekhout1,10,11
Running title: MALDI-TOF/MS identification of Malassezia spp.
1CBS-KNAW, Fungal Biodiversity Centre, Utrecht, the Netherlands.
2Department of Microbiology and Parasitology, Faculty of Medical Sciences, University of
Phayao, Phayao, Thailand.

This article is protected by copyright. All rights reserved.
3Research Mycology Laboratory (K.A. 70/3/6915), Microbiology Department, Medical School
of University of Athens, Athens, Greece.
4Bioanalytical Development, Bruker Daltonics GmbH, Bremen, Germany.
5Karolinska Institutet, Translational Immunology Unit, Department of Medicine Solna,
Stockholm, Sweden.
6Department of Veterinary Medicine, Aldo Moro University of Bari, Valenzano, Bari, Italy.
7Department of Biomedical Science and Human Oncology, Section of Hygiene, Aldo Moro
University of Bari, Bari, Italy.
8Department of Microbiology, Faculty of Medicine, Chiang Mai University, Chiang Mai,
Thailand.
9Department of Animal Health and Anatomy, Universitat Autònoma de Barcelona, Bellaterra,
Barcelona, Spain.
10Department of Internal Medicine and Infectious Diseases, University Medical Center, Utrecht,
the Netherlands.
11Department of Dermatology, Shanghai Key Laboratory of Molecular Medical Mycology,
Second Military Medical University, Changzheng Hospital, Institute of Dermatology and
Medical Mycology, Shanghai, People's Republic of China.
Corresponding author: Teun Boekhout, Department of Yeast Research, CBS-KNAW Fungal
Biodiversity Centre, Uppsalalaan 8, 3584CT Utrecht, The Netherlands.

This article is protected by copyright. All rights reserved.
Tel. +31 30 2122 600, Fax. +31 30 2512 097
e-mail: [email protected]
Statement of all funding sources that supported the work:
This publication was made possible by a NPRP grant 5-298-3-086 from the Qatar National
Research Fund (a member of the Qatar Foundation) to Teun Boekhout. The statements herein are
solely the responsibility of the authors. Anna Kolecka received a travel grant from the Walter
Gams non-profit foundation Studienstiftung Mykologische Systematik und Ökologie to attend
the 5th FEBS Advanced Lecture Course Human Fungal Pathogens: Molecular Mechanisms of
Host-Pathogen Interactions and Virulence, May 25-31, 2013, La Colle sur Loup, France
(Kolecka et al., abstract C12) where this work was partially presented.
Any conflict of interest:
Markus Kostrzewa is an employee of Bruker Daltonics GmbH (Bremen, Germany), the
manufacturer of the MALDI Biotyper system, which was used for this study. Bruker Daltonics
GmbH did not influence the design of the study. All other authors do not report conflicts of
interest. All authors contributed to the content and the writing of this manuscript.

This article is protected by copyright. All rights reserved.
What’s already known about this topic?
• Matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-
TOF MS) has been developed for the rapid identification of microorganisms and proved
to be a robust tool for the recognition of bacterial species.
• Recently, it was shown that MALDI-TOF MS provided reliable identifications of yeasts
and filamentous fungi.
• Identification of Malassezia isolates by routine methods remains laborious and results in
a relative high number of errorneous results.
What does this study add?
• A database of reference mass spectra resulting in efficient and correct identification of all
Malassezia species by MALDI-TOF MS in a short turnaround time.
• MALDI-TOF MS results were consistent with those obtained by sequence analyses.
• The method may efficiently demonstrate the involvement of different Malassezia species
in skin diseases.
• Application of MALDI-TOF MS for the identification of Malassezia isolates is of clinical
importance as fast and reliable identification will result in early application of treatment.

This article is protected by copyright. All rights reserved.
Abstract
Background: Infections caused by Malassezia yeasts are most likely underdiagnosed, because
fatty acid supplementation is needed for growth. Rapid identification of Malassezia species is
essential for appropriate treatment of Malassezia related skin-infections, fungemia and
nosocomial outbreaks in neonates, children and adults and can be live saving for those patients.
Matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS)
was reported as a rapid and reliable diagnostic tool to identify clinically important yeasts, but so
far no data have been reported on identification of Malassezia isolates with this technique.
Objectives: To create an extensive database of main mass spectra (MSPs) that will allow quick
identification of Malassezia species by MALDI-TOF MS.
Methods: An in-house library of 113 MSPs was created from 48 reference strains from the CBS-
KNAW yeast collection. The in-house library was challenged with two test sets of Malassezia
strains, namely 165 reference strains from the CBS collection and 338 isolates collected in
Greece, Italy, Sweden, and Thailand.
Results: MALDI-TOF MS allowed correct identification of all 14 Malassezia spp. MALDI-TOF
MS results were concordant with those of sequence analyses of the internal transcribed spacers
(ITS1/ITS2) and the D1/D2 domains of the large subunit (LSU) of the ribosomal DNA (rDNA).
Conclusions: Implementation of the MALDI-TOF MS system as a routine identification tool
will contribute to correct identification of Malassezia yeasts with a minimal effort and in a short
turnaround time which is specifically important for the rapid identification of Malassezia in skin
diseases and nosocomial outbreaks.
Key words: lipophilic yeasts, mass spectrometry, identification, skin, nosocomial outbreak

This article is protected by copyright. All rights reserved.
Introduction
Malassezia yeasts are anamorphic basidiomycetes and 14 species of this monophyletic genus are
presently known (1). Findley et al. (2) revealed the presence of eleven Malassezia species as part
of the human skin microbiome. Five species are mainly known from animal skin (i.e. M. caprae,
M. cuniculi, M. equina, M. nana, M. pachydermatis), but can also occur on humans (1, 3).
Depending on the host immune condition, Malassezia yeasts can be a skin-related commensal or
an opportunistic fungal pathogen (3, 4). They occur on healthy and diseased skin of humans and
warm-blooded animals and may be involved in catheter related infections of neonates and adults.
The occurrence of Malassezia species on skin is due to their ability to adapt to dermal
physiological properties (2, 5, 6). Malassezia species are known as etiological agents of skin
diseases, such as pityriasis (tinea) versicolor, folliculitis, seborrheic dermatitis (including
dandruff), and have been associated with several dermal conditions, such as atopic dermatitis,
and psoriasis (1, 4, 7-11). Malassezia species can cause invasive and systemic infections in
neonatal and paediatric patients as well as in adult individuals whose immune system is
suppressed or compromised that usually receive lipid supplementation via central venous
catheters (CVC) (12-19). Malassezia furfur is most commonly found in conjunction with
nosocomial outbreaks in neonatal intensive care units (20, 21), but outbreaks due to M.
pachydermatis and M. sympodialis can occur as well (16, 20, 22, 23). Early onset of antifungal
treatment for these patients is hampered due to difficult diagnostics of Malassezia yeasts and,
consequently, these infections are most likely underdiagnosed.
Before the introduction of molecular methods the identification of Malassezia spp. was usually
based on biochemical characteristics that involve many days, are not always reliable, and have an
error rate of 14 % (1, 3, 24). Additionally, the routine laboratories most commonly use non-lipid

This article is protected by copyright. All rights reserved.
supplemented media, like Sabouraud glucose agar (SGA) to differentiate yeasts, but for the
isolation, enumeration and identification of Malassezia spp. complex and specialized media
should be used (e.g. modified Dixon’s agar, mDA or modified Leeming & Notman agar, MLNA)
(1).
The culture-based conventional identification methods usually provide ambiguous results of
differentiation between Malassezia spp., and thus application of molecular techniques has
resulted in continuous revision of this genus (11). PCR- and sequence-based methods have been
applied to detect and identify Malassezia spp. Restriction fragment length polymorphism (RFLP)
and amplified fragment length polymorphism (AFLP) analyses of rDNA have also been used
(25-29). Non-invasive methods to detect Malassezia species directly from the infected skin
included terminal fragment length polymorphism and real-time PCR (27, 30-33). Sequence
analyses of universal barcoding markers, such as ITS and the D1/D2 regions of the rDNA,
became also the “gold standard” for accurate identification of Malassezia spp. (24, 34, 35) and
allowed recognition of new species (24, 34, 36-38). Pyrosequencing identified 11 Malassezia
species (39). In contrast to the culture-based methods, molecular techniques that involve DNA
can confirm the presence of dead or at least metabolically inert cells of Malassezia on the skin
(11).
Recently, matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-
TOF MS) became the state-of-the-art procedure in medical microbiology diagnostics. This
method allows identification of closely related species and subspecies of yeasts with high
resolution (40-44). Many studies showed rapid and reliable identification of microorganisms,
including bacteria and fungi, in clinical microbiological laboratories by this method (45-57).
While the high discriminatory power of species identification by MALDI-TOF MS is certainly

This article is protected by copyright. All rights reserved.
true for pure cultures, the method cannot be applied directly to identify clinical skin samples.
However, it was recently reported, that MALDI-TOF MS was applied on positive blood cultures
without the need for prior culturing and this resulted in reliable identification of Candida spp.
causing bloodstream infection within 30 min (58). Here, we provide evidence that MALDI-TOF
MS is highly suitable to detect, identify and discriminate between all 14 described Malassezia
spp. regardless on the culture media used. Results obtained from the MALDI-TOF MS-based
identification system were compared and validated with isolates from human clinical and
veterinary origins, namely healthy and diseased skin as well as bloodstream isolates.
Material and methods
Strains and culture conditions
Two sets of Malassezia strains representing all 14 species were selected (Table 1-2). The first set
contained 165 type and reference strains from the Yeast Collection of the CBS Fungal
Biodiversity Centre (CBS-KNAW, www.cbs.knaw.nl/collection) (i.e. reference set, Table 1,
Table S1). The second set (i.e. the clinical set, Table 2) consisted of 338 clinical isolates obtained
from four countries, including isolates from infected skin and blood of neonates (Greece),
isolates from the skin of patients with seborrheic dermatitis, atopic dermatitis and from healthy
individuals of which 58 isolates were previously identified as M. sympodialis (5) (Sweden), and
isolates from a nosocomial outbreak among neonates, isolated from their skin, urine, and blood
as well as from CVC and incubators of the ward (Italy). All these isolates were identified at the
genus level by routine methods used in the respective laboratories. The isolates from Thailand
were from the skin of healthy individuals and two isolates from patients with pityriasis
versicolor. The clinical set was complemented by 17 veterinary isolates that were identified in

This article is protected by copyright. All rights reserved.
Italy; 3 came from Brazil and 14 from Italy (59, 60). Validation of these strains at the species
level was achieved by rDNA sequence analysis of ITS and/or D1/D2 for veterinary isolates from
Italy (60) and part of the clinical isolates from Sweden (5), and rDNA sequence analysis of ITS
(GenBank acc. No KC464507; KC464508) and PCR-RFLP for the isolates from Greece (27).
Italian clinical isolates were validated by rDNA sequencing of ITS and D1/D2 in this study. The
Thai sub-set was used without further molecular validation as the results of MALDI-TOF MS
and ITS/D1/D2 sequencing identifications of isolates from the other origins were concordant.
All cultures grew on MLNA plates at 30 oC (1). Prior to MALDI-TOF MS identification, all
Malassezia spp. were re-cultured on mDA plates and cultivated at 30 °C (1). After 24h sufficient
growth was observed for M. pachydermatis and M. furfur, 48-72h were necessary to get
sufficient biomass of M. globosa, M. equina, M. sympodialis, M. slooffiae, M. nana, M. obtusa,
M. caprae, M. yamatoensis, M. dermatis and M. japonica, whereas the slow growing strains of
M. restricta and M. cuniculi needed seven up to 14 days to grow. A set of nine strains of M.
pachydermatis (L2, L9, CBS 1879NT, CBS 1885, CBS 4165, CBS 6534, CBS 6535, CBS
6535_2008, CBS 6537) was sub-cultured on SGA, mDA and MLNA plates and incubated for
24h at 30 oC. Origin and isolation site of L2 and L9 M. pachydermatis strains were previously
described by Theelen et al. (61).
Sequence analyses
DNA extractions were performed from strains grown on mDA plates according to the protocol of
Bolano et al. (62) and ITS and D1/D2 regions of the rDNA were analysed as reported by Gupta
et al. (24). The sequence alignments and consensus sequences were processed as described
previously (44).

This article is protected by copyright. All rights reserved.
MALDI-TOF MS - sample preparation and identification runs
Sample preparation was carried out using the ethanol/formic acid (EtOH/FA) protein extraction
method, according to the Bruker Daltonics GmbH protocol (56). Once sufficient growth of the
tested strains was detected on mDA (additionally on MLNA and SGA for M. pachydermatis),
two loops of yeast biomass (1 µL volume, sterile inoculation loop, Greiner bio-one) were
resuspended in 300 µL of milliQ water followed by 900 µL of absolute ethanol. The volume of
FA used for optimising ranged between 5–30 µL. However, for most of the tested samples and
depending on their hydrophobicity a volume of 10-20 µL of FA was found to be optimal and an
equal volume of acetonitrile (ACN) was added later. The volume of FA used was dependent on
the pellet size and the incubation time was important due to the hydrophobic nature of the cells.
The crude protein extract was used for two purposes: i) to generate Main mass SPectra (MSPs) to
be stored as an entry in the CBS-KNAW in-house library, and ii) for identification runs operated
by flex control version 3.3.108.0 and measured by the MALDI Biotyper RTC software 3.0
(Bruker, Germany).
The strains from the reference and clinical sets were measured and identified by MALDI
Biotyper RTC software 3.0 (Bruker, Germany) by using a 96-spot polished steel target plate
(Bruker, Germany) and Bacterial Test Standard (BTS, Bruker, Germany) as a positive control, as
described by (44). MALDI-TOF MS identification results were achieved automatically as the
log-score values: secure species identification (>2.0), secure genus identification (1.7-2.0) and
not reliable identification (<1.7, NRI). Strains with low scores (<1.7, NRI) and no peaks found
(NPF) were repeated. The identification was considered correct if at least one spot from the
duplicates gave a reliable identification with score >1.7.

This article is protected by copyright. All rights reserved.
CBS-KNAW in-house library
The CBS-KNAW in-house database contained 113 reference MSPs (Fig. 1) from 48 strains,
including type and reference strains covering all 14 Malassezia species (Table S1). These MSPs
were generated using the MALDI Biotyper Automation Control software version 3.0 (Bruker,
Germany). Additionally, ethanol extracts of 39 strains of Malassezia spp. cultivated on MLNA
and mDA were sent to Bruker Daltonics (Bremen, Germany) for creation of MSPs according to
the manufacturer’s standard operating procedures (SOPs). For identification runs CBS in-house
database was used simultaneously with the Bruker Daltonics commercial database (BDAL).
Results
Effect of growth media
Nine strains of M. pachydermatis growing on SGA, mDA and MLNA plates were identified
using the BDAL database alone and in combination with CBS in-house database. Results showed
that when the BDAL database was applied, only five out of nine (5/9) strains on MLNA and
mDA were identified as M. pachydermatis with a log-score >2.0 and three with scores (2.0-1.7).
When growing on SGA, seven (7/9) strains had log-scores >2.0 and one a score (2.0-1.7). One
strain had NRI (<1.7) when grew on SGA, mDA, and NPF on MLNA. Using the CBS in-house
database, irrespective of the media used, eight strains were correctly identified with scores >2.0
and one strain had a score (2.0-1.7) on SGA and mDA, and the same strain had NPF on MLNA.
We recommend the use of mDA plates because oil residues may be transferred from the surface
of MLNA plates together with the yeast biomass, which makes it difficult to dry and measure on
the spots on the target plate.

This article is protected by copyright. All rights reserved.
MALDI-TOF MS based identification of reference CBS-KNAW strains
Identification by MALDI-TOF MS of 165 reference Malassezia strains from CBS resulted in
accurate recognition at the genus and species level for all strains with different log-score values
(Table 1) and was concordant with ITS and D1/D2 sequences of rDNA. One hundred and forty
strains (84.8%) had log-score values >2.0 and 25 strains (15.2%) had values (2.0-1.7) for at least
on one spot from duplicates (Table 1). Strains of M. caprae (n= 3), M. cuniculi (n=1), M. equina
(n=3), M. japonica (n=3), M. nana (n=6), M. sympodialis (n=10) and M. yamatoensis (n=2) were
correctly identified with log-score values >2.0. For other species, namely M. dermatis, M. furfur,
M. globosa, M. obtusa, M. pachydermatis M. restricta, M. slooffiae, 112 strains were correctly
identified with log-score values >2.0 and 25 strains with scores (2.0-1.7).
For 19/165 Malassezia strains identifications provided by MALDI-TOF MS (with log-scores
>2.0 either 2.0-1.7) did not match with the primary identifications given by CBS-KNAW
collection. These initially discrepant data underwent validation by sequencing analyses of the
ITS and D1/D2 regions of the rDNA. The final results confirmed that the MALDI-TOF MS
identifications were correct and consistent with the sequencing results. Thus, nine strains initially
thought to represent M. globosa turned out to be M. furfur, four M. restricta turned out to be M.
slooffiae, from two M. furfur strains one turned out to be M. japonica and one M. pachydermatis,
one M. slooffiae turned out to be M. furfur, and from three M. sympodialis strains one turned out
to be M. furfur, M. nana and M. pachydermatis, respectively.
Identification of clinical isolates by MALDI-TOF MS and ITS/D1-D2 validation
In total, 338 clinical isolates of Malassezia spp. were examined (Table 2). No major errors were
observed in identifications provided by MALDI-TOF MS, as the correct identification at the

This article is protected by copyright. All rights reserved.
genus level was accomplished for all tested isolates and these were concordant with ITS and
D1/D2 validation. Based on the MALDI-TOF MS results, the sub-set of validated clinical
isolates (n=206, 100%) comprised M. dermatis (n=1), M. furfur (n=94), M. globosa (n=20), M.
pachydermatis (n=15), M. slooffiae (n=2), and M. sympodialis (n=74). Identification at the
species level was accomplished for all isolates; 193 isolates (93.7%) with log-score value >2.0,
for 10 isolates (4.8%) with log-scores (2.0-1.7) and three isolates (1.5%) were correctly
identified with log-scores <1.7 (Table 3). These three isolates were veterinary M. pachydermatis
strains from Italy (CD9, CD14, CD69). Three veterinary isolates from Italy (CD864, CD865,
CD866) were identified as M. furfur with log-score values >2.0 which was in concordance to ITS
and D1/D2 rDNA sequence data (60) and clustering of MSPs (Fig. 1).
Because of the robust identification of the above set of reference and clinical isolates, a set of
132 clinical isolates from Thailand was identified by MALDI-TOF MS only. MALDI-TOF MS
identification resulted in correct genus recognition for all isolates (100%). Identification at the
species level was detected with log-scores >2.0 for 128 isolates (96.9%), with log-scores (2.0-
1.7) for three isolates (2.3%) and one isolate (0.8%) matched with M. globosa with NRI score
1.7>value>1.6 (Table 3). Based on the MALDI-TOF MS identifications the Thai set comprised
M. dermatis (n=15), M. furfur (n=39), M. globosa (n=1), M. japonica (n=2), M. slooffiae (n=6),
and M. sympodialis (n=69) (Table 2). This set was used to determine the number of runs needed
to accomplish identification at the species level. The cumulative percentage of correct
identification scores obtained during successive runs showed that during the first run a correct
species match with reliable identification results (log-scores >2.0 and log-scores 2.0-1.7) was
achieved for 92.4% of tested isolates (n=122), which after the second run increased to 94.7%

This article is protected by copyright. All rights reserved.
(n=125). Four runs were needed to achieve species identification for 98.5% of tested isolates,
and one isolate remained identified with log-scores <1.7 (1.5%) only.
Discussion
Isolation and identification of Malassezia spp. from clinical specimens remains a problem due to
the lipid-dependent nature of these yeasts and differences in growth rates between species (1, 4).
Phenotypic identification of Malassezia strains involves the use of morphological and
physiological characteristics. Although these methods are cheap and easy to use, they remain
laborious and results turned out to be not very reliable when interpreted by personnel with a
limited expertise in diagnostic mycology. Also, Malassezia spp. misidentification rates were
reported. Makimura and colleagues (63) confirmed that 27 out of 46 clinical isolates of
Malassezia were misidentified, i.e. an error rate of 59%. Other studies employing AFLP and
rDNA sequencing analyses revealed approximately 14% misidentified Malassezia isolates by
traditional methods (24, 64).
Next to the rDNA sequence-based species identification, successful applications of MALDI-TOF
MS to identify pathogenic yeasts have been reported (43, 44, 64-67). So far Malassezia spp.
were insufficiently represented in the commercially available reference database BDAL that
contained hitherto only two MSPs, namely one for M. furfur (DSM 6170) and one for M.
pachydermatis (VML, no strain number provided). Therefore, a CBS in-house database of 113
MSPs of all 14 Malassezia spp. was created and results showed that MALDI-TOF MS was
capable to reliably identify and discriminate between all known Malassezia spp. (Fig. 1).
Technical variation and reproducibility of measurements were tested by using the same strains

This article is protected by copyright. All rights reserved.
that were used to create the database (Table 1). No effect of different media was observed when
applied to nine strains of M. pachydermatis. Identification rates increased from as low as 55.6%
(mDA, MLNA) with the original BDAL database with two entries only to 89% when using the
new CBS in-house database. Different media did not influence the identification results as the
species remained separate from each other (Fig. 2). Cluster analysis of MSPs generated for type
and reference strains revealed species-specific clusters (Fig. 2). M. nana, however, showed two
clusters (Fig. S1). The top cluster links together MSPs of three M. nana strains isolated from ear
of cows in Brazil, viz. CBS 9558, CBS 9559, and CBS 9560 (37), and the bottom one contains
two strains from cat; namely CBS 9557T from Japan (37) and CBS 8334 from UK. Interestingly,
Cabañes and colleagues proposed that M. nana may represent a genetically and taxonomically
heterogenous complex (68), which is corroborated here by MALDI-TOF MS.
Performance of the CBS-KNAW in-house database was challenged by testing a set of clinical
isolates. Importantly, all tested strains were identified at the genus level and for most of the
tested strains (98.2%) designation at the species level was achieved with high accuracy as only 4
out of 338 (1.2%) tested clinical isolates were identified with not reliable log-scores <1.7 (Table
3). Importantly, not only the log-scores >2.0 but also those of value (2.0-1.7) gave identifications
that were concordant at the species level with ITS and/or D1/D2 sequencing data. Thus we
suggest that the cut-off value for correct species identification of Malassezia isolates may be
lowered to log-scores >1.7, as it was already suggested for other yeasts (44, 53). Major errors
defined as incorrect genus ID were never observed with log-scores >1.7 and minor errors, viz.
misidentification at the species level, were not found either.
To conclude, MALDI-TOF MS identification of Malassezia isolates facilitates accurate species
identification. MALDI-TOF MS is a robust identification system for Malassezia isolates if an

This article is protected by copyright. All rights reserved.
extended database is used. The mass spectra generated per strain are a unique fingerprint and
therefore MALDI-TOF MS is ideal for species discrimination but may also have potential for
strain typing. This, however, needs to be further explored. It remains to be seen whether skin
scrapings contain sufficient biomass to allow direct examination by MALDI-TOF MS. Hsieh et
al. (69) showed that 106 of bacterial cells were sufficient to gain correct identifications with log-
scores >2.0 while 102-104 always gave log-scores <1.7. Furthermore, mixed skin infections are
difficult to deal with as well.
Once a culture (or cultures) is available from a clinical skin sample or blood, the correct
management of neonates and other individuals suffering from Malassezia related infections will
be strongly improved with the MALDI-TOF MS based identification of Malassezia isolates, due
to its accuracy and speed. Also the epidemiological connection between the skin microbiota and
Malassezia-related sepsis remains to be investigated. Finally, MALDI-TOF MS can also be used
for quality control of Malassezia culture, as it generates results consistent with rDNA sequencing
data in a short time.
Acknowledgements
Technical support and useful insights related to ITS/D1-D2 sequencing from Bart Theelen (CBS-
KNAW) is highly acknowledged. We also appreciate the assistance of Luuk van Haren (CBS-
KNAW) and we would like to thank Dr. Gustav Wikberg (Karolinska University Hospital,
Stockholm, Sweden) for collecting clinical samples used in this study.

This article is protected by copyright. All rights reserved.
References
1. Boekhout T, Gueho, E., Mayser, P., Velegraki, A. (eds.). Malassezia and the skin. Berlin
Heidelberg: Springer-Verlag; 2010. 319 p.
2. Findley K, Oh J, Yang J, Conlan S, Deming C, Meyer JA, et al. Topographic diversity of
fungal and bacterial communities in human skin. Nature. 2013;498(7454):367-70. Epub
2013/05/24.
3. Gueho-Kellerman E, Batra, R., Boekhout, T. Malassezia Baillon (1889) In: Kurtzman
CP, Fell, J.W., Boekhout, T., editor. The Yeasts: a taxonomic study. San Diego: Elsevier B.V.;
2011. p. 1807-32.
4. Gaitanis G, Magiatis P, Hantschke M, Bassukas ID, Velegraki A. The Malassezia genus
in skin and systemic diseases. Clinical microbiology reviews. 2012;25(1):106-41. Epub
2012/01/11.
5. Gioti A, Nystedt B, Li W, Xu J, Andersson A, Averette AF, et al. Genomic insights into
the atopic eczema-associated skin commensal yeast Malassezia sympodialis. mBio.
2013;4(1):e00572-12. Epub 2013/01/24.
6. Xu J, Saunders CW, Hu P, Grant RA, Boekhout T, Kuramae EE, et al. Dandruff-
associated Malassezia genomes reveal convergent and divergent virulence traits shared with
plant and human fungal pathogens. Proceedings of the National Academy of Sciences of the
United States of America. 2007;104(47):18730-5. Epub 2007/11/15.
7. Ashbee HR. Recent developments in the immunology and biology of Malassezia species.
FEMS immunology and medical microbiology. 2006;47(1):14-23. Epub 2006/05/19.
8. Batra R, Boekhout T, Gueho E, Cabanes FJ, Dawson TL, Jr., Gupta AK. Malassezia
Baillon, emerging clinical yeasts. FEMS yeast research. 2005;5(12):1101-13. Epub 2005/08/09.

This article is protected by copyright. All rights reserved.
9. Tragiannidis A, Bisping G, Koehler G, Groll AH. Minireview: Malassezia infections in
immunocompromised patients. Mycoses. 2010;53(3):187-95. Epub 2009/12/24.
10. Hay RJ. Malassezia, dandruff and seborrhoeic dermatitis: an overview. The British
journal of dermatology. 2011;165 Suppl 2:2-8. Epub 2011/09/23.
11. Gaitanis G, Velegraki A, Mayser P, Bassukas ID. Skin diseases associated with
Malassezia yeasts: facts and controversies. Clinics in dermatology. 2013;31(4):455-63. Epub
2013/06/29.
12. Kaneko T, Murotani M, Ohkusu K, Sugita T, Makimura K. Genetic and biological
features of catheter-associated Malassezia furfur from hospitalized adults. Medical mycology :
official publication of the International Society for Human and Animal Mycology.
2012;50(1):74-80. Epub 2011/05/31.
13. Giusiano G, Mangiaterra M, Saito VG, Rojas F, Gomez V, Diaz MC. Etiology of
fungaemia and catheter colonisation in Argentinean paediatric patients. Mycoses. 2006;49(1):49-
54. Epub 2005/12/22.
14. Curvale-Fauchet N, Botterel F, Legrand P, Guillot J, Bretagne S. Frequency of
intravascular catheter colonization by Malassezia spp. in adult patients. Mycoses. 2004;47(11-
12):491-4. Epub 2004/12/17.
15. Kessler AT, Kourtis AP, Simon N. Peripheral thromboembolism associated with
Malassezia furfur sepsis. The Pediatric infectious disease journal. 2002;21(4):356-7. Epub
2002/06/22.
16. Chryssanthou E, Broberger U, Petrini B. Malassezia pachydermatis fungaemia in a
neonatal intensive care unit. Acta Paediatr. 2001;90(3):323-7. Epub 2001/05/03.

This article is protected by copyright. All rights reserved.
17. Nguyen ST, Lund CH, Durand DJ. Thrombolytic therapy for adhesion of percutaneous
central venous catheters to vein intima associated with Malassezia furfur Infection. Journal of
perinatology : official journal of the California Perinatal Association. 2001;21(5):331-3. Epub
2001/09/06.
18. Schleman KA, Tullis G, Blum R. Intracardiac mass complicating Malassezia furfur
fungemia. Chest. 2000;118(6):1828-9. Epub 2000/12/15.
19. Devlin RK. Invasive fungal infections caused by Candida and Malassezia species in the
neonatal intensive care unit. Advances in neonatal care : official journal of the National
Association of Neonatal Nurses. 2006;6(2):68-77; quiz 8-9. Epub 2006/04/19.
20. van Belkum A, Boekhout T, Bosboom R. Monitoring spread of Malassezia infections in a
neonatal intensive care unit by PCR-mediated genetic typing. Journal of clinical microbiology.
1994;32(10):2528-32. Epub 1994/10/01.
21. Oliveri S, Trovato L, Betta P, Romeo MG, Nicoletti G. Malassezia furfur fungaemia in a
neonatal patient detected by lysis-centrifugation blood culture method: first case reported in
Italy. Mycoses. 2011;54(5):e638-40. Epub 2011/05/04.
22. Chang HJ, Miller HL, Watkins N, Arduino MJ, Ashford DA, Midgley G, et al. An
epidemic of Malassezia pachydermatis in an intensive care nursery associated with colonization
of health care workers' pet dogs. The New England journal of medicine. 1998;338(11):706-11.
Epub 1998/03/12.
23. Welbel SF, McNeil MM, Pramanik A, Silberman R, Oberle AD, Midgley G, et al.
Nosocomial Malassezia pachydermatis bloodstream infections in a neonatal intensive care unit.
The Pediatric infectious disease journal. 1994;13(2):104-8. Epub 1994/02/01.

This article is protected by copyright. All rights reserved.
24. Gupta AK, Boekhout T, Theelen B, Summerbell R, Batra R. Identification and typing of
Malassezia species by amplified fragment length polymorphism and sequence analyses of the
internal transcribed spacer and large-subunit regions of ribosomal DNA. Journal of clinical
microbiology. 2004;42(9):4253-60. Epub 2004/09/15.
25. Mirhendi H, Makimura K, Zomorodian K, Yamada T, Sugita T, Yamaguchi H. A simple
PCR-RFLP method for identification and differentiation of 11 Malassezia species. Journal of
microbiological methods. 2005;61(2):281-4. Epub 2005/02/22.
26. Guillot J, Deville M, Berthelemy M, Provost F, Gueho E. A single PCR-restriction
endonuclease analysis for rapid identification of Malassezia species. Letters in applied
microbiology. 2000;31(5):400-3. Epub 2000/11/09.
27. Gaitanis G, Velegraki A, Frangoulis E, Mitroussia A, Tsigonia A, Tzimogianni A, et al.
Identification of Malassezia species from patient skin scales by PCR-RFLP. Clinical
microbiology and infection : the official publication of the European Society of Clinical
Microbiology and Infectious Diseases. 2002;8(3):162-73. Epub 2002/05/16.
28. Giusiano G, Bustillo S, Mangiaterra M, Deluca G. [Identification of Malassezia species
by PCR-REA]. Revista Argentina de microbiologia. 2003;35(3):162-6. Epub 2003/11/01.
Identificacion de especies de Malassezia por PCR-REA.
29. Giusiano G, Sosa Mde L, Rojas F, Vanacore ST, Mangiaterra M. Prevalence of
Malassezia species in pityriasis versicolor lesions in northeast Argentina. Revista iberoamericana
de micologia. 2010;27(2):71-4. Epub 2010/03/30.
30. Gemmer CM, DeAngelis YM, Theelen B, Boekhout T, Dawson Jr TL, Jr. Fast,
noninvasive method for molecular detection and differentiation of Malassezia yeast species on

This article is protected by copyright. All rights reserved.
human skin and application of the method to dandruff microbiology. Journal of clinical
microbiology. 2002;40(9):3350-7. Epub 2002/08/31.
31. Paulino LC, Tseng CH, Blaser MJ. Analysis of Malassezia microbiota in healthy
superficial human skin and in psoriatic lesions by multiplex real-time PCR. FEMS yeast
research. 2008;8(3):460-71. Epub 2008/02/26.
32. Sugita T, Suto H, Unno T, Tsuboi R, Ogawa H, Shinoda T, et al. Molecular analysis of
Malassezia microflora on the skin of atopic dermatitis patients and healthy subjects. Journal of
clinical microbiology. 2001;39(10):3486-90. Epub 2001/09/28.
33. Sugita T, Tajima M, Tsubuku H, Tsuboi R, Nishikawa A. Quantitative analysis of
cutaneous malassezia in atopic dermatitis patients using real-time PCR. Microbiology and
immunology. 2006;50(7):549-52. Epub 2006/07/22.
34. Cabanes FJ, Theelen B, Castella G, Boekhout T. Two new lipid-dependent Malassezia
species from domestic animals. FEMS yeast research. 2007;7(6):1064-76. Epub 2007/03/21.
35. Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA, et al. Nuclear
ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi.
Proceedings of the National Academy of Sciences of the United States of America.
2012;109(16):6241-6. Epub 2012/03/29.
36. Sugita T, Tajima M, Takashima M, Amaya M, Saito M, Tsuboi R, et al. A new yeast,
Malassezia yamatoensis, isolated from a patient with seborrheic dermatitis, and its distribution in
patients and healthy subjects. Microbiology and immunology. 2004;48(8):579-83. Epub
2004/08/24.

This article is protected by copyright. All rights reserved.
37. Hirai A, Kano R, Makimura K, Duarte ER, Hamdan JS, Lachance MA, et al. Malassezia
nana sp. nov., a novel lipid-dependent yeast species isolated from animals. International journal
of systematic and evolutionary microbiology. 2004;54(Pt 2):623-7. Epub 2004/03/17.
38. Cabanes FJ, Vega S, Castella G. Malassezia cuniculi sp. nov., a novel yeast species
isolated from rabbit skin. Medical mycology : official publication of the International Society for
Human and Animal Mycology. 2011;49(1):40-8. Epub 2010/06/22.
39. Kim JY, Hahn HJ, Choe YB, Lee YW, Ahn KJ, Moon KC. Molecular biological
identification of malassezia yeasts using pyrosequencing. Annals of dermatology. 2013;25(1):73-
9. Epub 2013/03/08.
40. Posteraro B, Vella A, Cogliati M, De Carolis E, Florio AR, Posteraro P, et al. Matrix-
assisted laser desorption ionization-time of flight mass spectrometry-based method for
discrimination between molecular types of Cryptococcus neoformans and Cryptococcus gattii.
Journal of clinical microbiology. 2012;50(7):2472-6. Epub 2012/05/11.
41. Firacative C, Trilles L, Meyer W. MALDI-TOF MS enables the rapid identification of
the major molecular types within the Cryptococcus neoformans/C. gattii species complex. PloS
one. 2012;7(5):e37566. Epub 2012/06/06.
42. Karger A, Stock R, Ziller M, Elschner MC, Bettin B, Melzer F, et al. Rapid identification
of Burkholderia mallei and Burkholderia pseudomallei by intact cell Matrix-assisted Laser
Desorption/Ionisation mass spectrometric typing. BMC microbiology. 2012;12:229. Epub
2012/10/11.
43. Cendejas-Bueno E, Kolecka A, Alastruey-Izquierdo A, Theelen B, Groenewald M,
Kostrzewa M, et al. Reclassification of the Candida haemulonii complex as Candida haemulonii
(C. haemulonii group I), C. duobushaemulonii sp. nov. (C. haemulonii group II), and C.

This article is protected by copyright. All rights reserved.
haemulonii var. vulnera var. nov.: three multiresistant human pathogenic yeasts. Journal of
clinical microbiology. 2012;50(11):3641-51. Epub 2012/09/07.
44. Kolecka A, Khayhan K, Groenewald M, Theelen B, Arabatzis M, Velegraki A, et al.
Identification of medically relevant species of arthroconidial yeasts by use of matrix-assisted
laser desorption ionization-time of flight mass spectrometry. Journal of clinical microbiology.
2013;51(8):2491-500. Epub 2013/05/17.
45. Fedorko DP, Drake SK, Stock F, Murray PR. Identification of clinical isolates of
anaerobic bacteria using matrix-assisted laser desorption ionization-time of flight mass
spectrometry. European journal of clinical microbiology & infectious diseases : official
publication of the European Society of Clinical Microbiology. 2012;31(9):2257-62. Epub
2012/03/01.
46. de Respinis S, Tonolla M, Pranghofer S, Petrini L, Petrini O, Bosshard PP. Identification
of dermatophytes by matrix-assisted laser desorption/ionization time-of-flight mass
spectrometry. Medical mycology : official publication of the International Society for Human
and Animal Mycology. 2012. Epub 2012/12/12.
47. Tekippe EM, Shuey S, Winkler DW, Butler MA, Burnham CA. Optimizing Identification
of Clinically Relevant Gram-positive Organisms Using the Bruker Biotyper MALDI-TOF MS
System. Journal of clinical microbiology. 2013. Epub 2013/02/22.
48. Ford BA, Burnham CA. Optimization of Routine Identification of Clinically Relevant
Gram-Negative Bacteria Using MALDI-TOF MS and the Bruker Biotyper. Journal of clinical
microbiology. 2013. Epub 2013/02/22.

This article is protected by copyright. All rights reserved.
49. Khot PD, Couturier MR, Wilson A, Croft A, Fisher MA. Optimization of matrix-assisted
laser desorption ionization-time of flight mass spectrometry analysis for bacterial identification.
Journal of clinical microbiology. 2012;50(12):3845-52. Epub 2012/09/21.
50. Tan KE, Ellis BC, Lee R, Stamper PD, Zhang SX, Carroll KC. Prospective evaluation of
a matrix-assisted laser desorption ionization-time of flight mass spectrometry system in a
hospital clinical microbiology laboratory for identification of bacteria and yeasts: a bench-by-
bench study for assessing the impact on time to identification and cost-effectiveness. Journal of
clinical microbiology. 2012;50(10):3301-8. Epub 2012/08/03.
51. Sendid B, Ducoroy P, Francois N, Lucchi G, Spinali S, Vagner O, et al. Evaluation of
MALDI-TOF mass spectrometry for the identification of medically-important yeasts in the
clinical laboratories of Dijon and Lille hospitals. Medical mycology : official publication of the
International Society for Human and Animal Mycology. 2013;51(1):25-32. Epub 2012/06/19.
52. Nenoff P, Erhard M, Simon JC, Muylowa GK, Herrmann J, Rataj W, et al. MALDI-TOF
mass spectrometry - a rapid method for the identification of dermatophyte species. Medical
mycology : official publication of the International Society for Human and Animal Mycology.
2013;51(1):17-24. Epub 2012/05/12.
53. Van Herendael BH, Bruynseels P, Bensaid M, Boekhout T, De Baere T, Surmont I, et al.
Validation of a modified algorithm for the identification of yeast isolates using matrix-assisted
laser desorption/ionisation time-of-flight mass spectrometry (MALDI-TOF MS). European
journal of clinical microbiology & infectious diseases : official publication of the European
Society of Clinical Microbiology. 2012;31(5):841-8. Epub 2011/08/24.

This article is protected by copyright. All rights reserved.
54. Del Chierico F, Masotti A, Onori M, Fiscarelli E, Mancinelli L, Ricciotti G, et al.
MALDI-TOF MS proteomic phenotyping of filamentous and other fungi from clinical origin.
Journal of proteomics. 2012;75(11):3314-30. Epub 2012/04/17.
55. Cassagne C, Ranque S, Normand AC, Fourquet P, Thiebault S, Planard C, et al. Mould
routine identification in the clinical laboratory by matrix-assisted laser desorption ionization
time-of-flight mass spectrometry. PloS one. 2011;6(12):e28425. Epub 2011/12/24.
56. Marklein G, Josten M, Klanke U, Muller E, Horre R, Maier T, et al. Matrix-assisted laser
desorption ionization-time of flight mass spectrometry for fast and reliable identification of
clinical yeast isolates. Journal of clinical microbiology. 2009;47(9):2912-7. Epub 2009/07/03.
57. Dieckmann R, Helmuth R, Erhard M, Malorny B. Rapid classification and identification
of salmonellae at the species and subspecies levels by whole-cell matrix-assisted laser desorption
ionization-time of flight mass spectrometry. Applied and environmental microbiology.
2008;74(24):7767-78. Epub 2008/10/28.
58. Spanu T, Posteraro B, Fiori B, D'Inzeo T, Campoli S, Ruggeri A, et al. Direct maldi-tof
mass spectrometry assay of blood culture broths for rapid identification of Candida species
causing bloodstream infections: an observational study in two large microbiology laboratories.
Journal of clinical microbiology. 2012;50(1):176-9. Epub 2011/11/18.
59. Cafarchia C, Gasser RB, Latrofa MS, Parisi A, Campbell BE, Otranto D. Genetic variants
of Malassezia pachydermatis from canine skin: body distribution and phospholipase activity.
FEMS yeast research. 2008;8(3):451-9. Epub 2008/02/26.
60. Cafarchia C, Latrofa MS, Figueredo LA, da Silva Machado ML, Ferreiro L, Guillot J, et
al. Physiological and molecular characterization of atypical lipid-dependent Malassezia yeasts
from a dog with skin lesions: adaptation to a new host? Medical mycology : official publication

This article is protected by copyright. All rights reserved.
of the International Society for Human and Animal Mycology. 2011;49(4):365-74. Epub
2010/11/13.
61. Theelen B, Silvestri M, Gueho E, van Belkum A, Boekhout T. Identification and typing
of Malassezia yeasts using amplified fragment length polymorphism (AFLP), random amplified
polymorphic DNA (RAPD) and denaturing gradient gel electrophoresis (DGGE). FEMS yeast
research. 2001;1(2):79-86. Epub 2003/04/19.
62. Bolano A, Stinchi S, Preziosi R, Bistoni F, Allegrucci M, Baldelli F, et al. Rapid methods
to extract DNA and RNA from Cryptococcus neoformans. FEMS yeast research. 2001;1(3):221-
4. Epub 2003/04/19.
63. Makimura K, Tamura Y, Kudo M, Uchida K, Saito H, Yamaguchi H. Species
identification and strain typing of Malassezia species stock strains and clinical isolates based on
the DNA sequences of nuclear ribosomal internal transcribed spacer 1 regions. Journal of
medical microbiology. 2000;49(1):29-35. Epub 2000/01/11.
64. Bader O, Weig M, Taverne-Ghadwal L, Lugert R, Gross U, Kuhns M. Improved clinical
laboratory identification of human pathogenic yeasts by matrix-assisted laser desorption
ionization time-of-flight mass spectrometry. Clinical microbiology and infection : the official
publication of the European Society of Clinical Microbiology and Infectious Diseases.
2011;17(9):1359-65. Epub 2010/10/16.
65. Castanheira M, Woosley LN, Diekema DJ, Jones RN, Pfaller MA. Candida
guilliermondii and other species of candida misidentified as Candida famata: assessment by vitek
2, DNA sequencing analysis, and matrix-assisted laser desorption ionization-time of flight mass
spectrometry in two global antifungal surveillance programs. Journal of clinical microbiology.
2013;51(1):117-24. Epub 2012/10/27.

This article is protected by copyright. All rights reserved.
66. Martinez-Lamas L, Perez del Molino ML, Pardo F, Varela E, Regueiro BJ. [Matrix-
assisted laser desorption ionization time-of-flight (MALDI-TOF) mass spectrometry vs
conventional methods in the identification of Candida non-albicans]. Enfermedades infecciosas y
microbiologia clinica. 2011;29(8):568-72. Epub 2011/07/26. Espectrometria de masas matrix-
assisted laser desorption ionization time-of-flight vs. metodologia convencional en la
identificacion de Candida no-albicans.
67. Santos C, Lima N, Sampaio P, Pais C. Matrix-assisted laser desorption/ionization time-
of-flight intact cell mass spectrometry to detect emerging pathogenic Candida species.
Diagnostic microbiology and infectious disease. 2011;71(3):304-8. Epub 2011/08/23.
68. Cabanes FJ, Hernandez JJ, Castella G. Molecular analysis of Malassezia sympodialis-
related strains from domestic animals. Journal of clinical microbiology. 2005;43(1):277-83.
Epub 2005/01/07.
69. Hsieh SY, Tseng CL, Lee YS, Kuo AJ, Sun CF, Lin YH, et al. Highly efficient
classification and identification of human pathogenic bacteria by MALDI-TOF MS. Molecular &
cellular proteomics : MCP. 2008;7(2):448-56. Epub 2007/11/30.
Figures
Figure 1. Cluster analysis of 113 MALDI-TOF Main mass Spectra’s (MSPs) of strains
belonging to 14 Malassezia species that were used to create the CBS-KNAW in-house database.
Detailed information on CBS strains selected for MSPs creations can be found in Supplementary
Table S1. The MSPs were obtained from strains growing on mDa and MLNA and were created
by CBS-KNAW (as manual entries) and Bruker laboratories. Additionally, the MSPs of three
atypical M. furfur isolates of veterinary origin from Italy (CD864, CD865, CD866) were

This article is protected by copyright. All rights reserved.
included. Distance level of subdivision is displayed in relative units. n-number of MSPs created
per species
Figure 2. Cluster analysis of 32 MALDI-TOF Main mass Spectra’s (MSPs) of CBS type
strains of 14 Malassezia species. These strains were validated by sequencing of the
D1/D2 and ITS of regions. Distance level of subdivision is displayed in relative units. The
media used (mDA or MLNA) did not influence species separation. MSPs were created by
Bruker Daltonics and those made in-house by CBS-KNAW (as manual entries) are
marked with superscript (*).
Tables
Table 1. The MALDI-TOF MS identification score results of 165 reference strains belonging to
14 Malassezia species from the CBS-KNAW yeast collection.
M.
yam
atoe
nsis
2 0 0 0 0 0 0 0 0 0 2
M.
sym
podi
alis
7 2 0 1 0 0 0 0 0 0 10
M.
sloo
ffia
e
6 0 0 1 4 0 1 0 0 0 12
M.
rest
rict
a
1 0 0 0 1 0 1 0 0 0 3

This article is protected by copyright. All rights reserved.
M.
pach
yder
mat
is
17
1 0 7 4 4 1 0 0 0 34
M.
obtu
sa
1 0 0 0 1 0 0 0 0 0 2
M.
nana
3 1 0 2 0 0 0 0 0 0 6
M.
japo
nica
3 0 0 0 0 0 0 0 0 0 3
M.
glob
osa
8 0 0 1 0 0 2 0 0 0 11
M.
furf
ur
59
2 0 6 4 0 1 0 0 0 72
M.
equi
na
2 1 0 0 0 0 0 0 0 0 3
M.
derm
atis
2 0 0 0 0 0 1 0 0 0 3
M.
cuni
culi
1 0 0 0 0 0 0 0 0 0 1
M.
capr
ae
3 0 0 0 0 0 0 0 0 0 3
CB
S-K
NA
W
refe
renc
e se
t n(
%)
115
(69.
7)
7 (4
.2)
0 18
(10.
9)
14
(8.5
)
4 (2
.4)
7 (4
.2)
0 0 0 165
(100
)
Spot
2
(sco
re)
>2.0
2.0-
1.7
NR
I (<
1.7)
NP
F
2.0-
1.7
NR
I (<
1.7)
NP
F
NR
I (<
1.7)
NP
F
NP
F
tota
l no
. of
stra
ins
Spot
1
(sco
re)
>2.0
>2.0
>2.0
>2.0
2.0-
1.7
2.0-
1.7
2.0-
1.7
NR
I (<
1.7)
NR
I (<
1.7)
NP
F
This article is protected by copyright. All rights reserved.
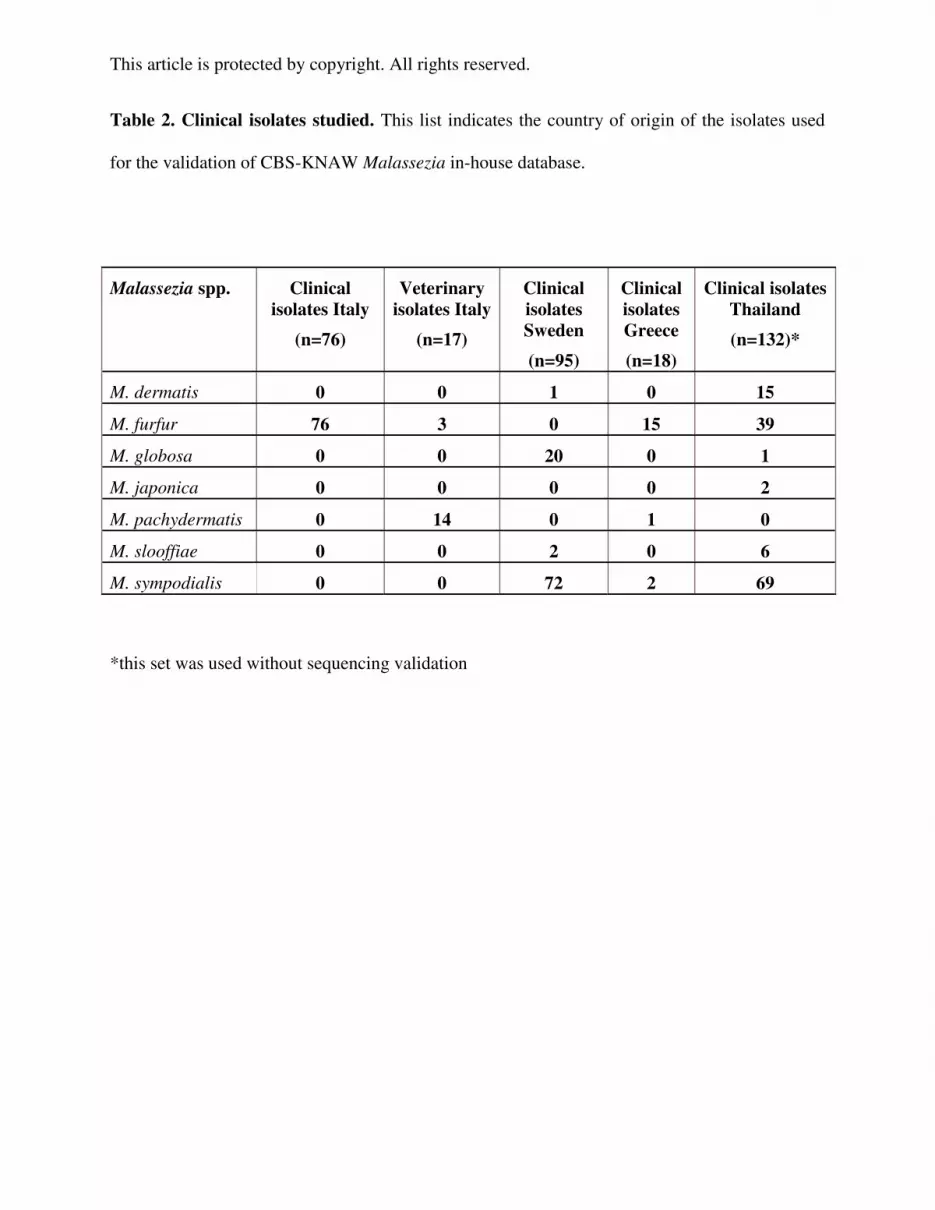
Table 2. Clinical isolates studied. This list indicates the country of origin of the isolates used
for the validation of CBS-KNAW Malassezia in-house database.
Malassezia spp. Clinical isolates Italy
(n=76)
Veterinary isolates Italy
(n=17)
Clinical isolates Sweden
(n=95)
Clinical isolates Greece
(n=18)
Clinical isolates Thailand
(n=132)*
M. dermatis 0 0 1 0 15
M. furfur 76 3 0 15 39
M. globosa 0 0 20 0 1
M. japonica 0 0 0 0 2
M. pachydermatis 0 14 0 1 0
M. slooffiae 0 0 2 0 6
M. sympodialis 0 0 72 2 69
*this set was used without sequencing validation

This article is protected by copyright. All rights reserved.
Table 3. MALDI-TOF MS identification results of Malassezia species from previously validated
and non-validated subsets of clinical isolates.
Score (log values) Validated clinical isolates from Greece,
Italy and Sweden (n = 206)
Non-validated clinical isolates from Thailand
(n = 132)
Spot 1 Spot 2 Number of correct ID (%)
Number of correct ID (%)
>2.0 >2.0 141 (68.4) 115 (87.1) >2.0 2.0-1.7 8 (3.9) 2 (1.5) >2.0 NRI (<1.7) 1 (0.5) 0
>2.0 NPF 43 (20.9) 11 (8.3) 2.0-1.7 2.0-1.7 6 (2.9) 0
2.0-1.7 NRI (<1.7) 1 (0.5) 1 (0.8)
2.0-1.7 NPF 3 (1.5) 2 (1.5) NRI (<1.7) NRI (<1.7) 3 (1.5) 1 (0.8)
NRI (<1.7) NPF 0 0 NPF NPF 0 0
total 206 (100) 132 (100)
ID – identification, NRI – not reliable ID, NPF – no peaks found

This article is protected by copyright. All rights reserved.

This article is protected by copyright. All rights reserved.