effects of the 5-ht2c receptor agonist cp809101 in the amygdala on reinstatement of cocaine-seeking...
TRANSCRIPT

Effects of the 5-HT2C receptor agonist CP809101in the amygdala on reinstatement of cocaine-seeking behavior and anxiety-like behavior
Lara A. Pockros-Burgess1, Nathan S. Pentkowski1,2, Taleen Der-Ghazarian2 andJanet L. Neisewander1,21Department of Psychology, Arizona State University, 950 S. McAllister, Tempe, AZ 85287, USA2 School of Life Sciences, Arizona State University, 427 East Tyler Mall, Tempe, AZ 85287, USA
Abstract
Serotonin 2C receptor (5-HT2CR) agonists attenuate reinstatement of cocaine-seeking behavior. These receptorsare found throughout the limbic system, including the basolateral amygdala (BlA), which is involved in formingassociations between emotional stimuli and environmental cues, and the central amygdala (CeA), which is impli-cated in the expression of conditioned responding to emotional stimuli. This study investigated whether5-HT2CRs in the amygdala are involved in cue and cocaine-primed reinstatement of cocaine-seeking behavior.Rats were trained to self-administer cocaine (0.75mg/kg, i.v.) which that was paired with light and tone cues,and then subsequently they underwent daily extinction training. Rats then received bilateral microinfusionsof the 5-HT2CR agonist CP809101 (0.01–1.0 μg/0.2 μl/side) into either the BlA or CeA prior to tests for cue orcocaine-primed (10mg/kg, i.p.) reinstatement. Rats were also tested for CP809101 effects on anxiety-like behavioron the elevated plus-maze (EPM). Surprisingly, intra-BlA CP809101 had no effect on cue reinstatement, thoughit did increase anxiety-like behavior on the EPM. Intra-CeA infusions of CP809101 attenuated cocaine-primedreinstatement, an effect that was prevented with concurrent administration of the 5-HT2CR antagonistSB242084 (0.1 μg/0.2 μl/side). CP809101 had no effect on cue reinstatement or anxiety-like behavior on theEPM. These findings suggest that 5-HT2CRs in the BlA modulate anxiety, whereas those in the CeA modulateincentive motivational effects induced by cocaine priming injections.
Received 20 March 2014; Reviewed 10 April 2014; Revised 19 April 2014; Accepted 27 April 2014
Key words: Addiction, elevated plus-maze, incentive motivation, relapse, serotonin.
NIH R01DA11064 and F32DA025413
Introduction
Serotonin 2C receptors (5-HT2CRs) modulate cocaine ef-fects. For instance, 5-HT2CR agonists attenuate cue andcocaine-primed reinstatement of cocaine-seeking beha-vior (Neisewander and Acosta, 2007; Fletcher et al.,2008; Pentkowski et al., 2010), while 5-HT2CR antagonistsenhance cocaine hyperlocomotion, cocaine-primed re-instatement, and cocaine self-administration (McMahonet al., 2001; Fletcher et al., 2002). The effects of 5-HT2CRagonists and indirect serotonin agonists on cue reinstate-ment of cocaine seeking are blocked by 5-HT2CR antago-nists, demonstrating 5-HT2CR mediation (Burmeisteret al., 2004; Pentkowski et al., 2010). 5-HT2CRs also modu-late anxiety as anxiety-like behavior on the elevatedplus-maze (EPM) is enhanced in transgenic mice
overexpressing 5-HT2CRs (Kimura et al., 2009a) butreduced in 5-HT2CR knockout mice (Heisler et al., 2007).
5-HT2CRs are found throughout the mesolimbic dopa-mine (DA) system (Pompeiano et al., 1994; Doherty andPickel, 2000), including the basolateral (BlA) and central(CeA) nuclei of the amygdala, key regions in the neurocir-cuitry of cocaine addiction (O’Dell et al., 1999; See, 2005;Alleweireldt et al., 2006) and anxiety disorders (LeDoux,2003). Indeed, the amygdala is involved in emotionallearning (Bechara et al., 1995) and memory (Cahill,2000), including fear conditioning (Blanchard andBlanchard, 1972; Lieblich et al., 1976; Pribram et al.,1979; Davis, 2000), avoidance learning (Weiskrantz,1956), and appetitive conditioning (Parkinson et al.,2000; Everitt et al., 2003). The ability of cocaine andcocaine-conditioned cues to prime motivation for cocainerelies on memory circuits involving the amygdala (Grantet al., 1996; Everitt et al., 2000). Specifically, the BlA hasbeen implicated in processing and modifying the incen-tive motivational value of drug-associated contextualand discrete cues (Everitt et al., 1999; Fuchs et al., 2002;McLaughlin and See, 2003). Although the role of
Address for correspondence: Dr J. L. Neisewander, School of Life Sciences,Arizona State University, 427 East Tyler Mall, Tempe, AZ 85287, USA.Tel.: +1 480 965 0803 Fax: +1 480 965 6899Email: [email protected]
International Journal of Neuropsychopharmacology, Page 1 of 12. © CINP 2014doi:10.1017/S1461145714000856
ARTICLE

amygdala 5-HT2CRs in processing the significance ofcocaine-associated cues has not been investigated, thereis evidence that they mediate anxiety-like behavior(Campbell and Merchant, 2003; de Mello Cruz et al.,2005; Heisler et al., 2007; Kimura et al., 2009b). Forinstance, intra-BlA 5-HT2CR agonists potentiate anxiety-like behavior in open-field and social exploration tests(Campbell and Merchant, 2003; Christianson et al.,2010). Further, the anxiogenic effects of a 5-HT2CR agonistcan be reversed by intra-BlA infusion of a 5-HT2CR antag-onist (de Mello Cruz et al., 2005).
The CeA is implicated in the unconditioned effects ofdrugs of abuse, including reward (O’Dell et al., 1999), re-inforcement (Caine et al., 1995), stress, and drug with-drawal (Koob and Nestler, 1997; Koob and Le Moal,2005). Inactivation of the CeA eliminates stress-primed re-instatement in both conditioned place preference (CPP)(Wang et al., 2002; Ma et al., 2008) and self-administrationmodels (McFarland et al., 2004). Studies also suggest arole for the CeA in responding for cocaine-paired cues(Kruzich and See, 2001; Thiel et al., 2010), though selec-tive inactivation of the CeA does not affect acquisitionof (Kruzich and See, 2001), or responding with (Burnset al., 1996; Robledo et al., 1996), conditioned reinforce-ment. However, the CeA appears to amplify conditionedresponses and is likely involved in the ‘incubation effect’in which cue reinstatement intensifies during abstinence(Robledo et al., 1996; Tran-Nguyen et al., 1998; Lu et al.,2005; Li et al., 2008; Thiel et al., 2012). Although the dis-tribution of 5-HT2CRs in the CeA is not well character-ized, it has been shown that 5-HT2CR knockout miceexhibit lower levels of Fos activation in the CeA followingsocial-defeat stress (Heisler et al., 2007) and 5-HT2CR ago-nists increase Fos expression in the CeA (Singewald et al.,2003; Somerville et al., 2007). Furthermore, there is a posi-tive correlation between 5-HT2CR levels in the CeA andanxiety-like behavior on the EPM (Li et al., 2012); how-ever, other studies failed to find effects of intra-CeA5-HT2CR agonists on anxiety-like behavior (Campbelland Merchant, 2003; Christianson et al., 2010).
In the present study, we hypothesized that BlA5-HT2CRs contribute to the inhibitory effects of systemic5-HT2CR agonists on cue reinstatement of extinguishedcocaine-seeking behavior based on the known involve-ment of the BlA in the incentive motivational effects ofcocaine-paired cues and the presence of 5-HT2CRs in theBlA. We further hypothesized that CeA 5-HT2CRs me-diate the inhibitory effects of systemic 5-HT2CR agonistson cocaine-primed reinstatement based on the known in-volvement of the CeA in unconditioned effects of cocaine.To examine these hypotheses, we used the selective5-HT2CR agonist CP890101, which has >500-fold selec-tivity for 5-HT2C over other 5-HT2Rs with EC50 valuesof 0.11, 153, and 65.3 nM for 5-HT2C, 5-HT2A, and5-HT2BRs, respectively (Siuciak et al., 2007; Fletcheret al., 2009). We predicted that intra-BlA CP809101would attenuate cue reinstatement of cocaine seeking
and increase anxiety-like behavior on the EPM, whilehaving no effect on cocaine-primed reinstatement.Conversely, we predicted that intra-CeA CP809101would attenuate cocaine-primed reinstatement andanxiety-like behavior on the EPM.
Method
Animals
Adult male Sprague–Dawley rats weighing 275–325 gat the start of the experiments were housed in aclimate-controlled colony room with a 14 h reversedlight/dark cycle (lights off at 07:00 hours) and cared forin accordance with the ‘Guide for the Care and Use ofLaboratory Animals’ (Institute of Laboratory AnimalResources on Life Sciences, National Research Council,2011).
Surgery
Animals were handled daily for at least 6 d beforeimplanting catheters into the right jugular vein as de-scribed by Pockros et al. (2011). During surgery, stainlesssteel guide cannulae were also implanted to a depth2.5 mm above the BlA and 2.7 mm above the CeA.The coordinates, selected based on previous research(Fuchs and See, 2002; Christianson et al., 2010; Thielet al., 2010), were: AP=−2.5, ML=5.0mm to the leftand 4.9mm to the right relative to bregma, and DV=−8.2 mm from the skull surface for BlA; AP=−2.5,ML=4.2 mm to the left and 4.1 mm to the right relativeto bregma, and DV=−8.2 mm from the skull surfacefor CeA (Paxinos, 2007). The guide cannulae and themetal end of the catheter were secured to the skulland anchor screws using dental acrylic cement. Metalstylets were inserted into the cannulae to maintainpatency. Catheters were flushed daily with a solutionof 0.1 ml saline containing heparin sodium (70 U/ml;APP Pharmaceuticals, USA) and Timentin (66.7mg/ml;GlaxoSmithKline, USA). Animals were given at least7 d of recovery from surgery before beginning self-administration training. Catheter patency was testedperiodically by administering 0.05ml Brevital (16.6 mg/ml, Jones Pharma Inc., USA), which briefly anesthetizesthe animal only if delivered i.v.
Intracranial drug infusions
CP809101 (Tocris, USA) was dissolved in artificialcerebral spinal fluid. Microinjections were deliveredover a 1min period using a 30 gauge injector (PlasticsOne) connected via polyethylene 50 tubing (BectonDickinson, USA) to a 25 μl syringe (Hamilton Co., USA)housed in an infusion pump (CMA Microdialysis,USA). Movement of an air bubble the proper distancethrough the drug infusion line confirmed successfuldrug infusion. After the infusion, the injectors remained
2 L. A. Pockros-Burgess et al.

in place for 1min to ensure thorough diffusion. After re-moving the injectors, metal stylets and caps were replacedbefore the animal was placed into the conditioning cham-ber for the test sessions.
Self-administration
Cocaine self-administration training sessions tookplace for 2 h, 6 d/wk in operant conditioning chambers(28×10×20 cm; Med Associates, USA), each containingan active lever, a cue light 4 cm above the active lever,an inactive lever, a tone generator (500Hz, 10 dB aboveground noise), and a house light on the wall oppositethe levers. Upon pressing the active lever to completea reinforcement schedule, the light and tone cues weresimultaneously activated, followed 1 s later by 0.1ml co-caine (0.75mg/kg, i.v.) infused over 6 s. The infusionpump, light and tone were then inactivated with theonset of the house light which signaled a 20 s time-outperiod during which active lever presses were recordedbut had no effects. Responses on the inactive lever wererecorded but had no effects.
For the first 5 d of training, all rats began on a fixedratio (FR) 1 schedule of reinforcement with the capabilityto progress to a variable ratio (VR) 3, and finally VR5schedule within a session. After ending the session on aVR5 schedule for five consecutive days, rats began thenext sessions on a VR3 schedule. Once rats began on aVR3 schedule and ended on a VR5 schedule, thereafterthey began on a VR5 schedule. All rats started on aVR5 schedule by day 14 and were on a VR5 scheduleexclusively for at least the last 5 d of self-administration.All rats were initially restricted to 16 g of food to facilitateacquisition of self-administration (Carroll et al., 1981) andremained food-restricted until they ended on a VR5schedule for three consecutive sessions. After meetingthis criterion, food rations were gradually increased toad libitum access which remained in effect throughoutthe rest of the experiment.
Extinction
Extinction training began once rats completed at least15 self-administration sessions and had received foodad libitum for at least the last five sessions. Extinctionoccurred daily for 1 h/d. Rats were placed into theself-administration chambers and lever presses wererecorded, but produced no consequences (i.e. no infusionsor cues were presented). Extinction sessions continued for10–14 d based on the criteria of either an 80% reduction inactive lever pressing from the rats’ highest response rateduring extinction or less than 20 active lever presses forthree consecutive days.
Experiments
Upon meeting the extinction criterion, rats with BlA andCeA cannulae (experiments 1 and 2, respectively)
underwent CP809101-primed reinstatement, cue re-instatement, cocaine-primed reinstatement, and EPM test-ing. Rats received their assigned dose of CP809101(0, 0.01, 0.1, or 1.0 μg/0.2 μg/side; final n/group=5–8)prior to each type of reinstatement test. In experiment 3,a new cohort of rats with CeA cannulae underwentCP809101-primed reinstatement testing for the effectsof CP809101 (0.01 μg/0.2 μl/side; n=6) or SB242082(0.1 μg/0.2 μl/side; n=5), cocaine-primed reinstatementtesting for the effects of CP809101 (0.01 μg/0.2 μl/side,n=6) or CP809101+SB242084 (0.01 and 0.1 μg/0.2 μl/side,respectively, n=5), and a subset of rats underwent EPMtesting for the effects of CP809101 (0 and 0.01 μg/0.2 μl/side, n=6–10). Group assignments were counterbalancedfor cocaine intake during self-administration.
CP809101-primed reinstatement
Following extinction, rats were assigned to CP809101dose groups, counterbalanced for cocaine intake duringself-administration as this has been shown to affect re-instatement (Deroche et al., 1999; Baker et al., 2001).Rats underwent two tests for CP809101-primed reinstate-ment of extinguished cocaine-seeking behavior, receivingvehicle prior to one test and their assigned dose ofCP809101 prior to the other test, with the order of thesepretreatments counterbalanced. Immediately after receiv-ing their assigned microinjection, rats were tested for1 h during which responding on either lever had noconsequences.
Cue reinstatement
Next, rats were given a minimum of three extinctionsessions to restabilize extinction response rates. Thenthey underwent two tests for cue reinstatement, receivinga vehicle microinjection prior to one test and theirassigned dose of CP809101 prior to the other test,with the order of these pretreatments counterbalanced.Immediately after receiving their assigned microinjection,rats were tested for cue reinstatement for 1 h. During thetest the same stimulus complex as that paired with co-caine during self-administration was presented response-contingently on an FR1 schedule; no cocaine was deliv-ered during cue tests. The FR1 schedule was used insteadof the VR5 training schedule because we have found thatthe FR1 schedule yields greater sensitivity for detectingthe predicted decrease in responding (Acosta et al.,2008). A noncontingent cue presentation was deliveredif the rat did not receive a response-contingent cue withinthe first 5 min of the test session.
Cocaine-primed reinstatement
After the two cue reinstatement tests, rats received at leastthree extinction sessions to restabilize baseline extinctionresponse rate. They were then given two tests for cocaine-primed reinstatement. Prior to one test, they received the
5-HT2C receptors in the amygdala and cocaine-seeking 3

same dose of CP809101 as they had received during cuereinstatement testing, and vehicle prior to the other test.The order of the two pretreatments was counterbalanced.Immediately after the microinjection, rats received apriming injection of cocaine (10mg/kg, i.p.) and wereimmediately placed into the conditioning chamber.Lever presses were recorded, but produced no conse-quences. To control for injection stress, rats were givenmock i.p. injections immediately before the extinctionsession preceding each of their cocaine-reinstatementtests, and the average response rates during these sessionswas used as the extinction baseline. Rats were given aminimum of three extinction sessions between tests torestabilize baseline response rates.
Elevated plus-maze
Following reinstatement testing, rats were tested for theeffects of CP809101 on anxiety-like behavior on theEPM at least 5 d after their last CP809101 treatment.Rats received one test on the EPM and were assigned toreceive either the same dose of CP809101 from reinstate-ment testing or vehicle. EPM testing was performed asdetailed previously (Pentkowski et al., 2009). Briefly,rats received their assigned microinjection and 5minlater they were individually placed in the center of theapparatus facing one of the two closed arms (File et al.,2004). Behavior was video recorded for 10min underdim lighting, and later scored using Observer 5.0 software(Noldus Information Technology BV, The Netherlands)by a trained observer blind to group assignment. Thebehaviors scored were entries and time spent in theopen arms, closed arms, and the middle of the maze.Locomotor activity was also analyzed by computer-automated video tracking (Clever Systems, USA). Theapparatus was cleaned with 5% ethanol between eachtest.
Histology
Rats were anesthetized with 3% isoflurane, given intra-cranial infusions (0.2 ml/side) of 1% methylene blue,and then decapitated. Harvested brains were stored at−80 °C and later sliced coronally (40 μm). Sections werestained with cresyl violet and cannula placementswere determined under a microscope by observers un-aware of group assignment (Fig. 1). Rats with incorrectplacements were excluded from the analyses.
Statistical analyses
For self-administration/reinstatement, data were analyzedusing mixed-factor analyses of variance (ANOVA) withsession (e.g. extinction baseline, vehicle test, andCP809101 test) as a within-subjects factor and dosagegroup (0.01, 0.1 and 1.0 μg/0.2 μl/side) as a between-subject factor. A Greenhouse–Geisser correction wasused to correct for heterogeneity of variance. Subsequent
post-hoc comparisons were made using tests of simplemain effects. In addition, planned t-tests were used totest the predictions that cocaine-seeking behavior wouldincrease after cocaine priming or cue presentation relativeto baseline. Baseline values were calculated as the averageof the sessions that occurred before each test day (i.e. thedays before agonist vs. vehicle testing). For cue andcocaine-primed reinstatement, if rats failed to meet thereinstatement criteria of doubling the extinction base-line response rate and at least twenty responses on the ac-tive lever on either of the two test days, they wereconsidered ‘non-reinstaters’ and were excluded from theanalysis (see results for details). Outliers were identifiedand excluded from analyses if they were more than2 S.D. from the mean.
Percent of time in the open arms of the EPM wasanalyzed using ANOVAs with dosage group as abetween-subjects factor. In addition, planned t-testswere used to test the prediction that anxiety-like behaviorwould increase after CP809101 relative to vehicle pretreat-ment. We also calculated an anxiety index score (Huynhet al., 2011) as follows: Anxiety Index=1−[(open armtime/10min)+(open arm entry/total entries)]/2.
(a)
(b)
Fig. 1. Histological reconstructions (left) with shaded regionsshowing where cannulae tips were considered correctly placedfor the basolateral amygdala (BlA) (a) and central amygdala(CeA) (b). Pictographs (right) of cresyl violet stained tissuesections to demonstrate methylene blue microinfusions into theBlA (a) and CeA (b). Tissue sections were taken at a levelapproximately 2.7mm from Bregma.
4 L. A. Pockros-Burgess et al.

Results
Cocaine self-administration
Table 1 shows the average number of cocaine infusions,and active and inactive lever presses/2 h (mean±S.E.M)during the last five cocaine self-administration trainingsessions for each group. While counterbalancing wasslightly affected by exclusion at the end of the studiesdue to misplaced cannulae, there were no group differ-ences in cocaine intake during self-administration.
Inactive lever presses
There were no differences in inactive lever presses be-tween groups for the first or last days of extinction, norfor any reinstatement test in any experiment (Table 2).
Extinction
The ANOVAs of the number of active lever presses/h onthe first day of extinction vs. the last day of extinctionbefore testing showed main effects of day (F1,19=87.18,p<0.001 and F1,20=90.92, p<0.001) for rats with BlA andCeA cannulae, respectively, but no dose effect or interac-tion with dose. The main effects indicated a significantdrop in responding across training sessions (Table 2).
Effects of CP809101 and SB242084 primes
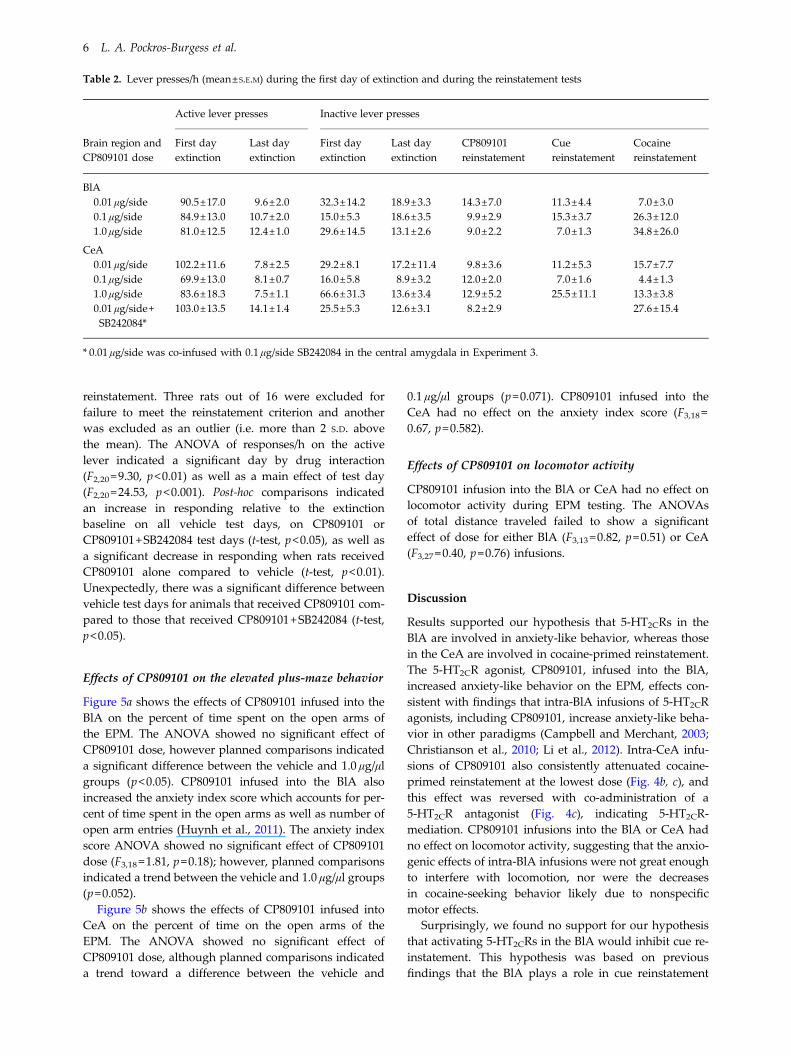
Figure 2 shows the effects of CP809101 priming injectionsinto the BlA (2a) or CeA (2b), and SB242084 priming injec-tions into the CeA (2c). None of these drug primes alteredresponding relative to extinction baseline.
Effects of CP809101 on cue reinstatement
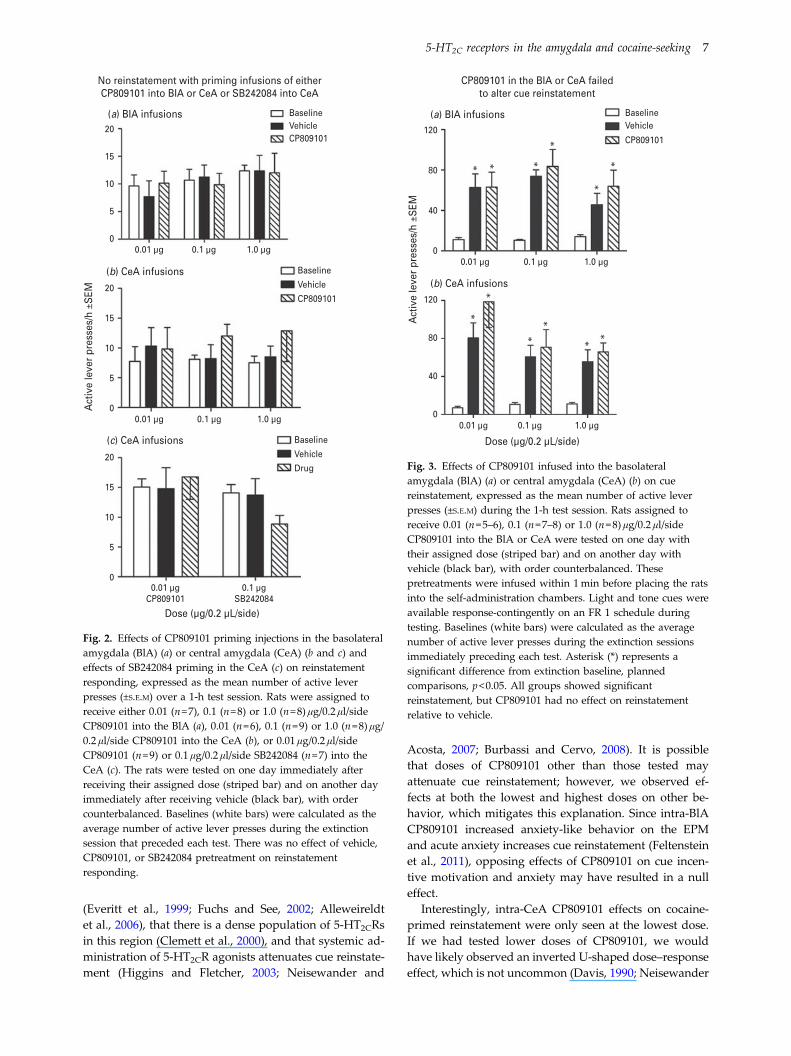
Figure 3 shows the effects of CP809101 infusions into theBlA (3a) or CeA (3b) on cue reinstatement. One rat out of27 tested for BlA infusion effects was excluded for failure
to meet the reinstatement criterion. The ANOVAs ofresponses/h on the active lever showed main effects oftest day (F1.30,24.77=20.51 for the BlA and F1.33,26.60=23.78 for the CeA, p<0.001) but no interactions withdose or main effects of dose. There was a trend towarda main effect of dose of intra-CeA CP809101 infusions(F2,20=3.22, p=0.061), however there were no differencesbetween CP809101 and vehicle test days at any specificdose. Tests of simple main effects indicated that when col-lapsed across dose, rats in both experiments showed cuereinstatement evident as an increase in responding onboth vehicle and CP809101 pretreatment test days relativeto the extinction baseline (p<0.05). The increase inresponding relative to extinction baseline was confirmedfor individual dosage groups on both vehicle and drugpretreatment tests using planned comparisons (acrossboth experiments, t values ranged from 2.87 to 10.54,p<0.05).
Effects of CP809101 with or without SB242084 oncocaine-primed reinstatement
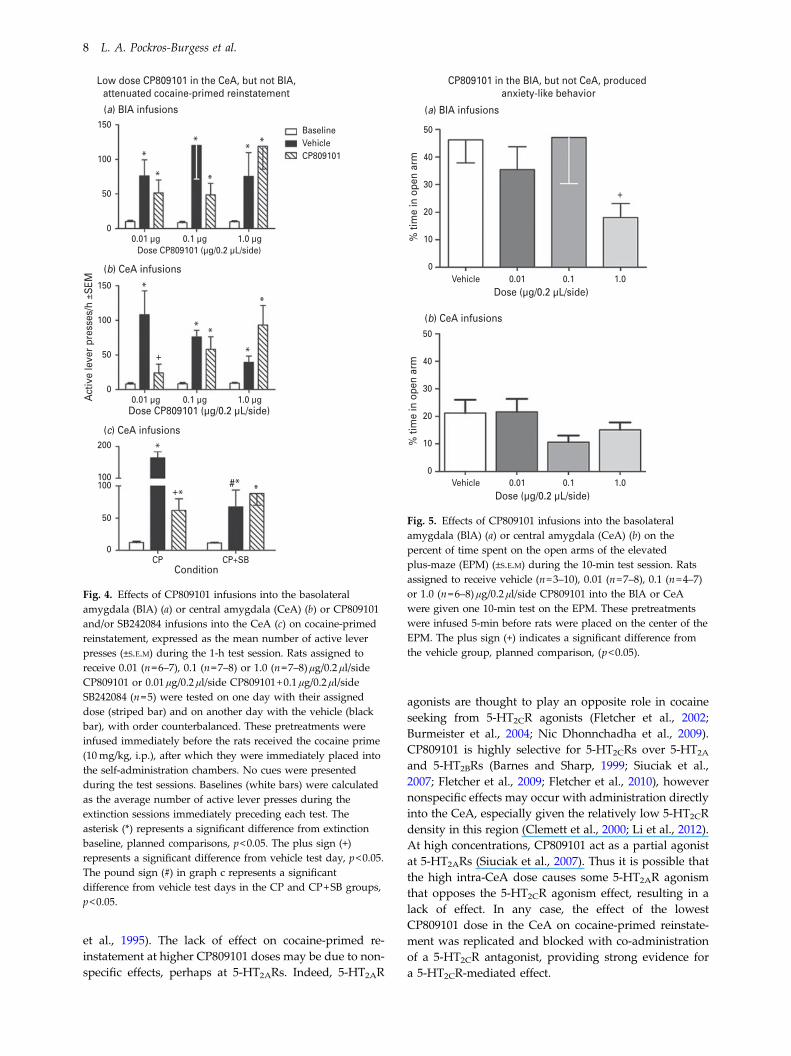
Figure 4a illustrates the effects of intra-BlA infusions ofCP809101 on cocaine-primed reinstatement. Four out of27 animals were excluded for failure to meet the reinstate-ment criterion. The ANOVA of responses/h on the activelever indicated a significant main effect of test day(F1.45,24.71=6.84, p<0.01) but no interaction with dose ormain effect of dose. Tests of simple main effects indicatedthat when collapsed across dose, all animals showedcocaine-primed reinstatement evident as an increase inresponding on the vehicle and CP809101 pretreatmenttest days relative to the extinction baseline (p<0.05). Theincrease in responding relative to extinction baselinewas confirmed for individual dosage groups on both ve-hicle and drug pretreatment tests using planned compar-isons (t values ranged from 2.14 to 5.80, p<0.05).
Figure 4b illustrates the effects of intra-CeA infusions ofCP809101 on cocaine-primed reinstatement. Three out of23 animals were excluded for failure to meet the reinstate-ment criterion. The ANOVA of responses/h on the activelever indicated a significant day by dose interaction(F4,34=4.06, p<0.01) as well as a main effect of test dayF2,34=12.33, p<0.001. Post-hoc comparisons indicated anincrease in responding relative to the extinction baselineon all vehicle test days and CP809101 test days at the0.1 and 1.0 μg/μl doses (t-test, p<0.05) and a significantdecrease in lever pressing at the 0.01 μg/μl dose ofCP809101 compared to vehicle (t-test, p<0.05). Therewere no differences on the vehicle test day across dosagegroups; however, there was a high amount of variance inthese measures. Thus, it is noteworthy that there were nodifferences between rats that received their vehicle testday first compared to those that received their vehicletest day second.
Figure 4c illustrates the effects of intra-CeA infusionsof CP809101 or CP809101+SB242084 on cocaine-primed
Table 1. The average number of cocaine infusions, and active andinactive lever presses/2 h (mean±S.E.M) during the last five cocaineself-administration training sessions
Brain region andCP809101 dose
Cocaineinfusions
Active leverpresses
Inactive leverpresses
BlA0.01 μg/side 27.2±2.8 163.2±29.1 17.9±11.80.1 μg/side 27.7±4.0 162.4±38.4 14.2±6.21.0 μg/side 28.7±4.5 158.1±32.2 3.7±1.8
CeA0.01 μg/side 23.3±2.5 113.8±11.9 2.2±1.20.1 μg/side 27.3±2.5 154.2±24.8 1.9±0.91.0 μg/side 26.6±1.4 130.4±12.1 428.6±425.60.01 μg/side+SB242084*
26.0±4.2 180.0±78.9 75.7±73.0
* 0.01 μg/side was co-infused with 0.1 μg/side SB242084 in thecentral amygdala in Experiment 3.
5-HT2C receptors in the amygdala and cocaine-seeking 5
reinstatement. Three rats out of 16 were excluded forfailure to meet the reinstatement criterion and anotherwas excluded as an outlier (i.e. more than 2 S.D. abovethe mean). The ANOVA of responses/h on the activelever indicated a significant day by drug interaction(F2,20=9.30, p<0.01) as well as a main effect of test day(F2,20=24.53, p<0.001). Post-hoc comparisons indicatedan increase in responding relative to the extinctionbaseline on all vehicle test days, on CP809101 orCP809101+SB242084 test days (t-test, p<0.05), as well asa significant decrease in responding when rats receivedCP809101 alone compared to vehicle (t-test, p<0.01).Unexpectedly, there was a significant difference betweenvehicle test days for animals that received CP809101 com-pared to those that received CP809101+SB242084 (t-test,p<0.05).
Effects of CP809101 on the elevated plus-maze behavior
Figure 5a shows the effects of CP809101 infused into theBlA on the percent of time spent on the open arms ofthe EPM. The ANOVA showed no significant effect ofCP809101 dose, however planned comparisons indicateda significant difference between the vehicle and 1.0 μg/μlgroups (p<0.05). CP809101 infused into the BlA alsoincreased the anxiety index score which accounts for per-cent of time spent in the open arms as well as number ofopen arm entries (Huynh et al., 2011). The anxiety indexscore ANOVA showed no significant effect of CP809101dose (F3,18=1.81, p=0.18); however, planned comparisonsindicated a trend between the vehicle and 1.0 μg/μl groups(p=0.052).
Figure 5b shows the effects of CP809101 infused intoCeA on the percent of time on the open arms of theEPM. The ANOVA showed no significant effect ofCP809101 dose, although planned comparisons indicateda trend toward a difference between the vehicle and
0.1 μg/μl groups (p=0.071). CP809101 infused into theCeA had no effect on the anxiety index score (F3,18 =0.67, p=0.582).
Effects of CP809101 on locomotor activity
CP809101 infusion into the BlA or CeA had no effect onlocomotor activity during EPM testing. The ANOVAsof total distance traveled failed to show a significanteffect of dose for either BlA (F3,13 =0.82, p=0.51) or CeA(F3,27=0.40, p=0.76) infusions.
Discussion
Results supported our hypothesis that 5-HT2CRs in theBlA are involved in anxiety-like behavior, whereas thosein the CeA are involved in cocaine-primed reinstatement.The 5-HT2CR agonist, CP809101, infused into the BlA,increased anxiety-like behavior on the EPM, effects con-sistent with findings that intra-BlA infusions of 5-HT2CRagonists, including CP809101, increase anxiety-like beha-vior in other paradigms (Campbell and Merchant, 2003;Christianson et al., 2010; Li et al., 2012). Intra-CeA infu-sions of CP809101 also consistently attenuated cocaine-primed reinstatement at the lowest dose (Fig. 4b, c), andthis effect was reversed with co-administration of a5-HT2CR antagonist (Fig. 4c), indicating 5-HT2CR-mediation. CP809101 infusions into the BlA or CeA hadno effect on locomotor activity, suggesting that the anxio-genic effects of intra-BlA infusions were not great enoughto interfere with locomotion, nor were the decreasesin cocaine-seeking behavior likely due to nonspecificmotor effects.
Surprisingly, we found no support for our hypothesisthat activating 5-HT2CRs in the BlA would inhibit cue re-instatement. This hypothesis was based on previousfindings that the BlA plays a role in cue reinstatement
Table 2. Lever presses/h (mean±S.E.M) during the first day of extinction and during the reinstatement tests
Brain region andCP809101 dose
Active lever presses Inactive lever presses
First dayextinction
Last dayextinction
First dayextinction
Last dayextinction
CP809101reinstatement
Cuereinstatement
Cocainereinstatement
BlA0.01 μg/side 90.5±17.0 9.6±2.0 32.3±14.2 18.9±3.3 14.3±7.0 11.3±4.4 7.0±3.00.1 μg/side 84.9±13.0 10.7±2.0 15.0±5.3 18.6±3.5 9.9±2.9 15.3±3.7 26.3±12.01.0 μg/side 81.0±12.5 12.4±1.0 29.6±14.5 13.1±2.6 9.0±2.2 7.0±1.3 34.8±26.0
CeA0.01 μg/side 102.2±11.6 7.8±2.5 29.2±8.1 17.2±11.4 9.8±3.6 11.2±5.3 15.7±7.70.1 μg/side 69.9±13.0 8.1±0.7 16.0±5.8 8.9±3.2 12.0±2.0 7.0±1.6 4.4±1.31.0 μg/side 83.6±18.3 7.5±1.1 66.6±31.3 13.6±3.4 12.9±5.2 25.5±11.1 13.3±3.80.01 μg/side+SB242084*
103.0±13.5 14.1±1.4 25.5±5.3 12.6±3.1 8.2±2.9 27.6±15.4
* 0.01 μg/side was co-infused with 0.1 μg/side SB242084 in the central amygdala in Experiment 3.
6 L. A. Pockros-Burgess et al.
(Everitt et al., 1999; Fuchs and See, 2002; Alleweireldtet al., 2006), that there is a dense population of 5-HT2CRsin this region (Clemett et al., 2000), and that systemic ad-ministration of 5-HT2CR agonists attenuates cue reinstate-ment (Higgins and Fletcher, 2003; Neisewander and
Acosta, 2007; Burbassi and Cervo, 2008). It is possiblethat doses of CP809101 other than those tested mayattenuate cue reinstatement; however, we observed ef-fects at both the lowest and highest doses on other be-havior, which mitigates this explanation. Since intra-BlACP809101 increased anxiety-like behavior on the EPMand acute anxiety increases cue reinstatement (Feltensteinet al., 2011), opposing effects of CP809101 on cue incen-tive motivation and anxiety may have resulted in a nulleffect.
Interestingly, intra-CeA CP809101 effects on cocaine-primed reinstatement were only seen at the lowest dose.If we had tested lower doses of CP809101, we wouldhave likely observed an inverted U-shaped dose–responseeffect, which is not uncommon (Davis, 1990; Neisewander
No reinstatement with priming infusions of eitherCP809101 into BIA or CeA or SB242084 into CeA
(a) BIA infusions
(b) CeA infusions
Act
ive
leve
r pr
esse
s/h
±SEM
(c) CeA infusions
Baseline
20
15
10
5
0
20
15
10
5
0
20
15
10
5
0
0.01 µg 0.1 µg 1.0 µg
0.01 µg
0.01 µgCP809101
0.1 µgSB242084
Dose (µg/0.2 µL/side)
0.1 µg 1.0 µg
VehicleCP809101
Baseline
Vehicle
CP809101
Baseline
Vehicle
Drug
Fig. 2. Effects of CP809101 priming injections in the basolateralamygdala (BlA) (a) or central amygdala (CeA) (b and c) andeffects of SB242084 priming in the CeA (c) on reinstatementresponding, expressed as the mean number of active leverpresses (±S.E.M) over a 1-h test session. Rats were assigned toreceive either 0.01 (n=7), 0.1 (n=8) or 1.0 (n=8) μg/0.2 μl/sideCP809101 into the BlA (a), 0.01 (n=6), 0.1 (n=9) or 1.0 (n=8) μg/0.2 μl/side CP809101 into the CeA (b), or 0.01 μg/0.2 μl/sideCP809101 (n=9) or 0.1 μg/0.2 μl/side SB242084 (n=7) into theCeA (c). The rats were tested on one day immediately afterreceiving their assigned dose (striped bar) and on another dayimmediately after receiving vehicle (black bar), with ordercounterbalanced. Baselines (white bars) were calculated as theaverage number of active lever presses during the extinctionsession that preceded each test. There was no effect of vehicle,CP809101, or SB242084 pretreatment on reinstatementresponding.
CP809101 in the BIA or CeA failedto alter cue reinstatement
(a) BIA infusions
(b) CeA infusions
* *
*
**
***
*
*
*
*
120
80
40
0
120
80
40
0
0.01 µg 0.1 µg 1.0 µg
0.01 µg 0.1 µg 1.0 µg
BaselineVehicle
CP809101
Dose (µg/0.2 µL/side)
Act
ive
leve
r pr
esse
s/h
±SEM
Fig. 3. Effects of CP809101 infused into the basolateralamygdala (BlA) (a) or central amygdala (CeA) (b) on cuereinstatement, expressed as the mean number of active leverpresses (±S.E.M) during the 1-h test session. Rats assigned toreceive 0.01 (n=5–6), 0.1 (n=7–8) or 1.0 (n=8) μg/0.2 μl/sideCP809101 into the BlA or CeA were tested on one day withtheir assigned dose (striped bar) and on another day withvehicle (black bar), with order counterbalanced. Thesepretreatments were infused within 1min before placing the ratsinto the self-administration chambers. Light and tone cues wereavailable response-contingently on an FR 1 schedule duringtesting. Baselines (white bars) were calculated as the averagenumber of active lever presses during the extinction sessionsimmediately preceding each test. Asterisk (*) represents asignificant difference from extinction baseline, plannedcomparisons, p<0.05. All groups showed significantreinstatement, but CP809101 had no effect on reinstatementrelative to vehicle.
5-HT2C receptors in the amygdala and cocaine-seeking 7
et al., 1995). The lack of effect on cocaine-primed re-instatement at higher CP809101 doses may be due to non-specific effects, perhaps at 5-HT2ARs. Indeed, 5-HT2AR
agonists are thought to play an opposite role in cocaineseeking from 5-HT2CR agonists (Fletcher et al., 2002;Burmeister et al., 2004; Nic Dhonnchadha et al., 2009).CP809101 is highly selective for 5-HT2CRs over 5-HT2A
and 5-HT2BRs (Barnes and Sharp, 1999; Siuciak et al.,2007; Fletcher et al., 2009; Fletcher et al., 2010), howevernonspecific effects may occur with administration directlyinto the CeA, especially given the relatively low 5-HT2CRdensity in this region (Clemett et al., 2000; Li et al., 2012).At high concentrations, CP809101 act as a partial agonistat 5-HT2ARs (Siuciak et al., 2007). Thus it is possible thatthe high intra-CeA dose causes some 5-HT2AR agonismthat opposes the 5-HT2CR agonism effect, resulting in alack of effect. In any case, the effect of the lowestCP809101 dose in the CeA on cocaine-primed reinstate-ment was replicated and blocked with co-administrationof a 5-HT2CR antagonist, providing strong evidence fora 5-HT2CR-mediated effect.
Low dose CP809101 in the CeA, but not BIA,attenuated cocaine-primed reinstatement
(a) BIA infusions
*
*
*
*
**
150
100
50
0
150
200
100100
50
0CP CP+SB
Condition
100
50
0
(b) CeA infusions
(c) CeA infusions
Dose CP809101 (µg/0.2 µL/side)0.01 µg 0.1 µg 1.0 µg
Dose CP809101 (µg/0.2 µL/side)0.01 µg 0.1 µg 1.0 µgA
ctiv
e le
ver
pres
ses/
h ±S
EM *
*
+
*
*
*
*
+*#* *
BaselineVehicleCP809101
Fig. 4. Effects of CP809101 infusions into the basolateralamygdala (BlA) (a) or central amygdala (CeA) (b) or CP809101and/or SB242084 infusions into the CeA (c) on cocaine-primedreinstatement, expressed as the mean number of active leverpresses (±S.E.M) during the 1-h test session. Rats assigned toreceive 0.01 (n=6–7), 0.1 (n=7–8) or 1.0 (n=7–8) μg/0.2 μl/sideCP809101 or 0.01 μg/0.2 μl/side CP809101+0.1 μg/0.2 μl/sideSB242084 (n=5) were tested on one day with their assigneddose (striped bar) and on another day with the vehicle (blackbar), with order counterbalanced. These pretreatments wereinfused immediately before the rats received the cocaine prime(10mg/kg, i.p.), after which they were immediately placed intothe self-administration chambers. No cues were presentedduring the test sessions. Baselines (white bars) were calculatedas the average number of active lever presses during theextinction sessions immediately preceding each test. Theasterisk (*) represents a significant difference from extinctionbaseline, planned comparisons, p<0.05. The plus sign (+)represents a significant difference from vehicle test day, p<0.05.The pound sign (#) in graph c represents a significantdifference from vehicle test days in the CP and CP+SB groups,p<0.05.
CP809101 in the BIA, but not CeA, produced anxiety-like behavior
(a) BIA infusions
(b) CeA infusions
+
50
40
% ti
me
in o
pen
arm
30
20
10
Vehicle 0.01Dose (µg/0.2 µL/side)
0.1 1.0
Vehicle 0.01Dose (µg/0.2 µL/side)
0.1 1.0
0
50
40
% ti
me
in o
pen
arm
30
20
10
0
Fig. 5. Effects of CP809101 infusions into the basolateralamygdala (BlA) (a) or central amygdala (CeA) (b) on thepercent of time spent on the open arms of the elevatedplus-maze (EPM) (±S.E.M) during the 10-min test session. Ratsassigned to receive vehicle (n=3–10), 0.01 (n=7–8), 0.1 (n=4–7)or 1.0 (n=6–8) μg/0.2 μl/side CP809101 into the BlA or CeAwere given one 10-min test on the EPM. These pretreatmentswere infused 5-min before rats were placed on the center of theEPM. The plus sign (+) indicates a significant difference fromthe vehicle group, planned comparison, (p<0.05).
8 L. A. Pockros-Burgess et al.

There was an unusual amount of variability inresponses during intra-CeA vehicle tests for cocaine-primed reinstatement. When CP809101 was injected intothe CeA in experiment 3, the lowest dose attenuatedcocaine-primed reinstatement consistent with the findingsin experiment 2; however, we observed unusually highresponding on the vehicle test day in experiment 3 com-pared to experiment 2. Statistically, there were no signifi-cant differences in vehicle tests across the groups inexperiment 2. We also did not observe an order effect be-tween rats that had their vehicle test first compared to se-cond. In experiment 3, we did find a significant differencebetween vehicle groups, and although there was again nosignificant order effect detected, likely due to low n/con-dition, we suspect that order of drug vs. vehicle contribu-ted to the high variance in response rates. Specifically,when the low dose of CP809101 was tested before vehicle,the suppression of cocaine-primed reinstatement mayhave resulted in a higher amount of lever pressing onthe subsequent vehicle reinstatement test. By contrast,when the highest dose of CP809101 was tested first in ex-periment 2, there was no suppression of responding, soextinction learning during that test likely resulted inlower responding on the subsequent vehicle test.
While the between-group variance in responding onvehicle test days in experiments 2 and 3 is curious, itdoes not detract from the reliability of our within-subjectscomparisons showing that 0.01 μg/0.2 μl CP809101 infu-sions into the CeA attenuated cocaine-primed reinstate-ment relative to vehicle, and this effect was not seenwith the addition of a 5-HT2CR antagonist. There wasan inconsistency in that the between group analysesrevealed no effect of CP809101, however the within-subjects comparison showed that the lowest dose ofCP809101 significantly decreased cocaine-seeking beha-vior relative to vehicle, while CP809101+SB242084 hadno effect. The lack of consistency of an antagonist reversaleffect across these two types of comparisons is likely dueto the high amount of variance in vehicle response rates.Given the variance and n/group in this experiment, itstands to reason that only the within-subjects comparisonindicates an antagonist effect because it is a more power-ful test than the between-group comparison.
Anatomical specificity and potential damage from re-peated drug infusions are important to consider whenusing intracranial drug administration. The regionspecific effects of the BlA and CeA infusions mitigates ef-fects of drug spread since these regions border each other.Furthermore, rats underwent cocaine-primed reinstate-ment and EPM testing as their last tests, yet we observedan effect of intra-BlA CP809101 on this test, suggestingthat tissue damage was not likely. Moreover, histologicalverification of cannula placements showed no evidence ofexcessive tissue damage from repeated infusions.
We targeted the rostral part of the BlA. The rostral(rBlA) and caudal (cBlA) subsections of the BlA have di-vergent projections. The rBLA projects to the nucleus
accumbens (NAc) core and dorsal agranular insular cor-tex, whereas the cBLA projects to the NAc shell and pre-limbic cortex (Groenewegen et al., 1990; Kita and Kitai,1990; Shinonaga et al., 1994). Inactivation of the rBlAusing excitotoxic lesions, tetrodotoxin (TTX) or lidocaineconsistently disrupts cue reinstatement (Meil and See,1997; Grimm and See, 2000; Kantak et al., 2002), whilecBlA lidocaine inactivation reduces drug-seeking be-havior during cocaine self-administration on a secondorder schedule (Kantak et al., 2002). Other studies havefound effects of pharmacological manipulations in thecBlA on both cue and cocaine-primed reinstatement(Alleweireldt et al., 2006; Berglind et al., 2006), and inthe rBlA on cue reinstatement (Alleweireldt et al., 2006;Mashhoon et al., 2009). Thus, if 5-HT2CRs in the BlA areinvolved in cue reinstatement, it seems likely that wewould have observed an effect with intra-rBlA infusionof CP809101.
Based on previous literature, we expected CP809101 inthe BlA, but not the CeA, to affect anxiety-like behavior.Our results support this prediction, as the slight increasein anxiety-like behavior from intra-CeA CP809101 wasnot statistically significant. Together with the inconsist-encies among previous studies examining 5-HT2CRsin the CeA on anxiety-like behavior (Campbell andMerchant, 2003; Heisler et al., 2007; Christianson et al.,2010; Li et al., 2012), our results suggest that 5-HT2CRsin this region may play a less prominent role in anxietythan those in the BlA. Accordingly, Campbell andMerchant (2003) found that a 5-HT2CR agonist, mCPP,in the CeA did not affect ultrasonic vocalizations or ex-ploratory behavior on a novel-object task, though in theBlA mCPP increased anxiety-like behaviors during thistask. Similarly, Christianson et al. (2010) found that a5-HT2CR antagonist, SB242084, in the CeA did not affectanxiety-like behavior on a juvenile social explorationtask, though in the BlA the antagonist decreased anxiety-like behavior.
We hypothesize that the mechanism underlying the ef-fects of CP809101 in the BlA on anxiety-like behavior andin the CeA on cocaine-primed reinstatement involvesmodulation of GABA release. In the BlA, 5-HT2CR ago-nists activate GABA interneurons at low doses (Steinet al., 2000), however it is hypothesized that high dosesof 5-HT2CR agonists activate excitatory pyramidal projec-tion neurons (Rainnie, 1999), which overshadows GABAactivation leading to the anxiogenic effects of 5-HT2CRagonists (Campbell and Merchant, 2003). 5-HT2CR-induced GABA activation may inhibit GABA interneur-ons that inhibit projection neurons, resulting in netdisinhibition and anxiety-like behavior. To our knowl-edge, the cellular localization of 5-HT2CRs in the CeA isunknown, however they may be located on GABA inter-neurons and thus 5-HT2CR agonists in this region wouldincrease GABA inhibition. Consistent with this idea,GABA receptor agonists in the CeA attenuate footshock-primed reinstatement of cocaine seeking (McFarland
5-HT2C receptors in the amygdala and cocaine-seeking 9

et al., 2004). While we did not assess the effects ofCP809101 on footshock-primed reinstatement, the CeAis involved in both stress and cocaine reinforcement(O’Dell et al., 1999; McFarland et al., 2004; Cain et al.,2008). However, since we did not observe an anxiogeniceffect of CP809101 infusions into the CeA, involvementof 5-HT2CRs in the CeA in stress-primed reinstatementmay be due to the motivational impact of the stressorrather than an anxiogenic effect.
In conclusion, our results suggest that 5-HT2CR acti-vation in the CeA reduces cocaine-primed reinstatement,whereas 5-HT2CR activation in the BlA increases anxiety,but does not affect cue reinstatement. The null effect ofCP809101 in the BlA on cue reinstatement was surprisingand warrants additional investigation, perhaps withapproaches that alter 5-HT2CR expression. These findingsare the first to show that 5-HT2CRs in the CeA attenuatecocaine-seeking behavior and support the idea that5-HT2CR agonists have potential as treatments for cocaineaddiction.
Acknowledgments
The authors thank Drs M. Foster Olive, Cheryl Conrad,and Federico Sanabria for their input as members of thefirst author’s dissertation committee. We also thankNatalie Peartree, Ryan Bastle, Timothy Cheung, SamuelBrunwasser, Kimberly Zwick, Theresa Ullman, SineadhConway, Jeffrey Lynn, Julianna Goenaga, and BryanHarder for their expert technical assistance. This researchwas supported by NIH grants RO1DA110644 andF32DA025413.
Statement of Interest
None.
References
Acosta JI, Thiel KJ, Sanabria F, Browning JR, Neisewander JL(2008) Effect of schedule of reinforcement on cue-elicitedreinstatement of cocaine-seeking behavior. Behav Pharmacol19:129–136.
Alleweireldt AT, Hobbs RJ, Taylor AR, Neisewander JL (2006)Effects of SCH-23390 infused into the amygdala or adjacentcortex and basal ganglia on cocaine seeking andself-administration in rats. Neuropsychopharmacol31:363–374.
Baker DA, Tran-Nguyen TL, Fuchs RA, Neisewander JL (2001)Influence of individual differences and chronic fluoxetinetreatment on cocaine-seeking behavior in rats.Psychopharmacol (Berl) 155:18–26.
Barnes NM, Sharp T (1999) A review of central 5-HT receptorsand their function. Neuropharmacol 38:1083–1152.
Bechara A, Tranel D, Damasio H, Adolphs R, Rockland C,Damasio AR (1995) Double dissociation of conditioning anddeclarative knowledge relative to the amygdala andhippocampus in humans. Science (New York) 269:1115–1118.
Berglind WJ, Case JM, Parker MP, Fuchs RA, See RE (2006)Dopamine D1 or D2 receptor antagonism within thebasolateral amygdala differentially alters the acquisition ofcocaine-cue associations necessary for cue-inducedreinstatement of cocaine-seeking. Neuroscience 137:699–706.
Blanchard DC, Blanchard RJ (1972) Innate and conditionedreactions to threat in rats with amygdaloid lesions. J CompPhysiol Psychol 81:281–290.
Burbassi S, Cervo L (2008) Stimulation of serotonin2C receptorsinfluences cocaine-seeking behavior in response todrug-associated stimuli in rats. Psychopharmacol (Berl)196:15–27.
Burmeister JJ, Lungren EM, Kirschner KF, Neisewander JL (2004)Differential roles of 5-HT receptor subtypes in cue and cocainereinstatement of cocaine-seeking behavior in rats.Neuropsychopharmacol 29:660–668.
Burns LH, Annett L, Kelley AE, Everitt BJ, Robbins TW (1996)Effects of lesions to amygdala, ventral subiculum, medialprefrontal cortex, and nucleus accumbens on the reaction tonovelty: implication for limbic-striatal interactions. BehavNeurosci 110:60–73.
Cahill L (2000) Neurobiological mechanisms of emotionallyinfluenced, long-term memory. Prog Brain Res 126:29–37.
Cain ME, Denehy ED, Bardo MT (2008) Individual differencesin amphetamine self-administration: the role of the centralnucleus of the amygdala. Neuropsychopharmacol33:1149–1161.
Caine SB, Heinrichs SC, Coffin VL, Koob GF (1995) Effects of thedopamine D-1 antagonist SCH 23390 microinjected into theaccumbens, amygdala or striatum on cocaineself-administration in the rat. Brain Res 692:47–56.
Campbell BM, Merchant KM (2003) Serotonin 2C receptorswithin the basolateral amygdala induce acute fear-likeresponses in an open-field environment. Brain Res 993:1–9.
Carroll ME, France CP, Meisch RA (1981) Intravenousself-administration of etonitazene, cocaine and phencyclidinein rats during food deprivation and satiation. J Pharmacol ExpTher 217:241–247.
Christianson JP, Ragole T, Amat J, Greenwood BN, Strong PV,Paul ED, Fleshner M, Watkins LR, Maier SF (2010)5-hydroxytryptamine 2C receptors in the basolateral amygdalaare involved in the expression of anxiety after uncontrollabletraumatic stress. Biol Psychiatry 67:339–345.
Clemett DA, Punhani T, Duxon MS, Blackburn TP, Fone KC(2000) Immunohistochemical localisation of the 5-HT2Creceptor protein in the rat CNS. Neuropharmacol 39:123–132.
Davis DM, Svensgaard DJ (1990) U-shaped dose-responsecurves: their occurrence and implications for risk assessment.J Toxic Environ Health 30:71–83.
Davis M (2000) The role of the amygdala in conditioned andunconditioned fear and anxiety. In: The Amygdala: afunctional analysis (Aggleton JP, ed), pp213–288. Oxford:Oxford University Press.
de Mello Cruz AP, Pinheiro G, Alves SH, Ferreira G, Mendes M,Faria L, Macedo CE, Motta V, Landeira-Fernandez J (2005)Behavioral effects of systemically administered MK-212 areprevented by ritanserin microinfusion into the basolateralamygdala of rats exposed to the elevated plus-maze.Psychopharmacol 182:345–354.
Deroche V, Le Moal M, Piazza PV (1999) Cocaineself-administration increases the incentive motivationalproperties of the drug in rats. Eur J Neurosci 11:2731–2736.
10 L. A. Pockros-Burgess et al.

Doherty MD, Pickel VM (2000) Ultrastructural localization of theserotonin 2A receptor in dopaminergic neurons in the ventraltegmental area. Brain Res 864:176–185.
Everitt BJ, Parkinson JA, Olmstead MC, Arroyo M, Robledo P,Robbins TW (1999) Associative processes in addiction andreward. The role of amygdala-ventral striatal subsystems. AnnN Y Acad Sci 877:412–438.
Everitt BJ, Cardinal RN, Hall J, Parkinson JA, Robbins TW(2000) Differential involvement of amygdala subsystemsin appetitive conditioning and drug addiction. In: TheAmygdala: A functional analysis (Aggleton JP, ed), pp353–390.Oxford: Oxford University Press.
Everitt BJ, Cardinal RN, Parkinson JA, Robbins TW (2003)Appetitive behavior: impact of amygdala-dependentmechanisms of emotional learning. Ann N Y Acad Sci985:233–250.
Feltenstein MW, Henderson AR, See RE (2011) Enhancement ofcue-induced reinstatement of cocaine-seeking in rats byyohimbine: sex differences and the role of the estrous cycle.Psychopharmacol 216:53–62.
File SE, Lippa AS, Beer B, Lippa MT (2004) Animal tests ofanxiety. Curr Protoc Neurosci Chapter 8:Unit 8.3.
Fletcher PJ, Grottick AJ, Higgins GA (2002) Differential effects ofthe 5-HT(2A) receptor antagonist M100907 and the 5-HT(2C)receptor antagonist SB242084 on cocaine-induced locomotoractivity, cocaine self-administration and cocaine-inducedreinstatement of responding. Neuropsychopharmacol27:576–586.
Fletcher PJ, Rizos Z, Sinyard J, Tampakeras M, Higgins GA(2008) The 5-HT2C receptor agonist Ro60-0175 reduces cocaineself-administration and reinstatement induced by the stressoryohimbine, and contextual cues. Neuropsychopharmacol33:1402–1412.
Fletcher PJ, Tampakeras M, Sinyard J, Slassi A, Isaac M,Higgins GA (2009) Characterizing the effects of 5-HT(2C)receptor ligands on motor activity and feeding behaviour in5-HT(2C) receptor knockoutmice.Neuropharmacol 57:259–267.
Fletcher PJ, Sinyard J, Higgins GA (2010) Genetic andpharmacological evidence that 5-HT2C receptor activation, butnot inhibition, affects motivation to feed under a progressiveratio schedule of reinforcement. Pharmacol Biochem Behav97:170–178.
Fuchs RA, See RE (2002) Basolateral amygdala inactivationabolishes conditioned stimulus- and heroin-inducedreinstatement of extinguished heroin-seeking behavior in rats.Psychopharmacol 160:425–433.
Fuchs RA, Weber SM, Rice HJ, Neisewander JL (2002) Effects ofexcitotoxic lesions of the basolateral amygdala oncocaine-seeking behavior and cocaine conditioned placepreference in rats. Brain Res 929:15–25.
Grant S, London ED, Newlin DB, Villemagne VL, Liu X,Contoreggi C, Phillips RL, Kimes AS, Margolin A (1996)Activation of memory circuits during cue-elicited cocainecraving. Proc Natl Acad Sci USA 93:12040–12045.
Grimm JW, See RE (2000) Dissociation of primary and secondaryreward-relevant limbic nuclei in an animal model of relapse.Neuropsychopharmacol 22:473–479.
Groenewegen HJ, Berendse HW, Wolters JG, Lohman AH(1990) The anatomical relationship of the prefrontal cortexwith the striatopallidal system, the thalamus and theamygdala: evidence for a parallel organization. Prog Brain Res85:95–116; discussion 116–118.
Heisler LK, Zhou L, Bajwa P, Hsu J, Tecott LH (2007) Serotonin5-HT(2C) receptors regulate anxiety-like behavior. Genes BrainBehav 6:491–496.
Higgins GA, Fletcher PJ (2003) Serotonin and drug reward: focuson 5-HT2C receptors. Eur J Pharmacol 480:151–162.
Huynh TN, Krigbaum AM, Hanna JJ, Conrad CD (2011) Sexdifferences and phase of light cycle modify chronic stresseffects on anxiety and depressive-like behavior. Behav BrainRes 222:212–222.
Kantak KM, Black Y, Valencia E, Green-Jordan K,Eichenbaum HB (2002) Dissociable effects of lidocaineinactivation of the rostral and caudal basolateral amygdala onthe maintenance and reinstatement of cocaine-seekingbehavior in rats. J Neurosci 22:1126–1136.
Kimura A, Stevenson PL, Carter RN, Maccoll G, French KL, PaulSimons J, Al-Shawi R, Kelly V, Chapman KE, Holmes MC(2009a) Overexpression of 5-HT2C receptors in forebrainleads to elevated anxiety and hypoactivity. Eur J Neurosci30:299–306.
Kimura A, Stevenson PL, Carter RN, Maccoll G, French KL,Simons JP, Al-Shawi R, Kelly V, Chapman KE, Holmes MC(2009b) Overexpression of 5-HT2C receptors in forebrain leadsto elevated anxiety and hypoactivity. Eur J Neurosci30:299–306.
Kita H, Kitai ST (1990) Amygdaloid projections to the frontalcortex and the striatum in the rat. J Comp Neurol 298:40–49.
Koob GF, Le Moal M (2005) Plasticity of reward neurocircuitryand the ‘dark side’ of drug addiction. Nat Neurosci8:1442–1444.
Koob GF, Nestler EJ (1997) The neurobiology of drug addiction.J Neuropsychiatry Clin Neurosci 9:482–497.
Kruzich PJ, See RE (2001) Differential contributions of thebasolateral and central amygdala in the acquisition andexpression of conditioned relapse to cocaine-seeking behavior.J Neurosci 21:RC155.
LeDoux J (2003) The emotional brain, fear, and the amygdala.Cell Mol Neurobiol 23:727–738.
Li Q, Luo T, Jiang X, Wang J (2012) Anxiolytic effects of 5-HT(1)Areceptors and anxiogenic effects of 5-HT(2)C receptors in theamygdala of mice. Neuropharmacol 62:474–484.
Li YQ, Li FQ, Wang XY, Wu P, Zhao M, Xu CM, Shaham Y, Lu L(2008) Central amygdala extracellular signal-regulated kinasesignaling pathway is critical to incubation of opiate craving.J Neurosci 28:13248–13257.
Lieblich I, Yitzhaky J, Cohen E (1976) Effects of septal lesions onbehavior elicited by stimulation of the amygdaloid complex.Behav Biol 17:1–16.
Lu L, Hope BT, Dempsey J, Liu SY, Bossert JM, Shaham Y (2005)Central amygdala ERK signaling pathway is critical toincubation of cocaine craving. Nat Neurosci 8:212–219.
Ma DY, Xu MY, Yang HC, Yang LZ (2008) Effect of inhibition ofthe central nucleus of the amygdala and drug experience onthe regions underlying footshock-induced reinstatement ofmorphine seeking. J Int Med Res 36:992–1000.
Mashhoon Y, Tsikitas LA, Kantak KM (2009) Dissociable effectsof cocaine-seeking behavior following D1 receptor activationand blockade within the caudal and rostral basolateralamygdala in rats. Eur J Neurosci 29:1641–1653.
McFarland K, Davidge SB, Lapish CC, Kalivas PW (2004) Limbicand motor circuitry underlying footshock-inducedreinstatement of cocaine-seeking behavior. J Neurosci24:1551–1560.
5-HT2C receptors in the amygdala and cocaine-seeking 11

McLaughlin J, See RE (2003) Selective inactivation of thedorsomedial prefrontal cortex and the basolateral amygdalaattenuates conditioned-cued reinstatement of extinguishedcocaine-seeking behavior in rats. Psychopharmacol 168:57–65.
McMahon LR, Filip M, Cunningham KA (2001) Differentialregulation of the mesoaccumbens circuit by serotonin5-hydroxytryptamine (5-HT)2A and 5-HT2C receptors.J Neurosci 21:7781–7787.
Meil WM, See RE (1997) Lesions of the basolateral amygdalaabolish the ability of drug associated cues to reinstateresponding during withdrawal from self-administered cocaine.Behav Brain Res 87:139–148.
National Research Council (2011) Guide for the care and use oflaboratory animals: eighth edition. Washington, DC: TheNational Academies Press.
Neisewander JL, Acosta JI (2007) Stimulation of 5-HT2Creceptors attenuates cue and cocaine-primed reinstatementof cocaine-seeking behavior in rats. Behav Pharmacol18:791–800.
Neisewander JL, O’Dell LE, Redmond JC (1995) Localization ofdopamine receptor subtypes occupied by intra-accumbensantagonists that reverse cocaine-induced locomotion. Brain Res671:201–212.
Nic Dhonnchadha BA, Fox RG, Stutz SJ, Rice KC,Cunningham KA (2009) Blockade of the serotonin 5-ht2areceptor suppresses cue-evoked reinstatement ofcocaine-seeking behavior in a rat self-administration model.Behav Neurosci 123:382–396.
O’Dell LE, Sussman AN, Meyer KL, Neisewander JL (1999)Behavioral effects of psychomotor stimulant infusions intoamygdaloid nuclei. Neuropsychopharmacology 20:591–602.
Parkinson JA, Robbins TW, Everitt BJ (2000) Dissociable roles ofthe central and basolateral amygdala in appetitive emotionallearning. Eur J Neurosci 12:405–413.
Paxinos GW, Watson C (2007) The rat brain in stereotaxiccoordinates. Amsterdam; Boston: Academic Press/Elsevier.
Pentkowski NS, Acosta JI, Browning JR, Hamilton EC,Neisewander JL (2009) Stimulation of 5-HT(1B) receptorsenhances cocaine reinforcement yet reduces cocaine-seekingbehavior. Addict Biol 14:419–430.
Pentkowski NS, Duke FD, Weber SM, Pockros LA, Teer AP,Hamilton EC, Thiel KJ, Neisewander JL (2010) Stimulation ofmedial prefrontal cortex serotonin 2C (5-HT(2C)) receptorsattenuates cocaine-seeking behavior. Neuropsychopharmacol35:2037–2048.
Pockros LA, Pentkowski NS, Swinford SE, Neisewander JL(2011) Blockade of 5-HT2A receptors in the medial prefrontalcortex attenuates reinstatement of cue-elicited cocaine-seekingbehavior in rats. Psychopharmacol 213(2–3):307–320.
Pompeiano M, Palacios JM, Mengod G (1994) Distribution of theserotonin 5-HT2 receptor family mRNAs: comparison between5-HT2A and 5-HT2C receptors. Brain Res Mol Brain Res23:163–178.
Pribram KH, Reitz S, McNeil M, Spevack AA (1979) The effect ofamygdalectomy on orienting and classical conditioning inmonkeys. Pavlov J Biol Sci 14:203–217.
Rainnie DG (1999) Serotonergic modulation ofneurotransmission in the rat basolateral amygdala.J Neurophysiol 82:69–85.
Robledo P, Robbins TW, Everitt BJ (1996) Effects of excitotoxiclesions of the central amygdaloid nucleus on the potentiationof reward-related stimuli by intra-accumbens amphetamine.Behav Neurosci 110:981–990.
See RE (2005) Neural substrates of cocaine-cue associations thattrigger relapse. Eur J Pharmacol 526:140–146.
Shinonaga Y, Takada M, Mizuno N (1994) Topographicorganization of collateral projections from the basolateralamygdaloid nucleus to both the prefrontal cortex and nucleusaccumbens in the rat. Neuroscience 58:389–397.
Singewald N, Salchner P, Sharp T (2003) Induction of c-Fosexpression in specific areas of the fear circuitry in rat forebrainby anxiogenic drugs. Biol Psychiatry 53:275–283.
Siuciak JA, Chapin DS, McCarthy SA, Guanowsky V, Brown J,Chiang P, Marala R, Patterson T, Seymour PA, Swick A,Iredale PA (2007) CP-809,101, a selective 5-HT2C agonist,shows activity in animal models of antipsychotic activity.Neuropharmacol 52:279–290.
Somerville EM, Horwood JM, Lee MD, Kennett GA, Clifton PG(2007) 5-HT(2C) receptor activation inhibits appetitive andconsummatory components of feeding and increases brainc-fos immunoreactivity in mice. Eur J Neurosci 25:3115–3124.
Stein C, Davidowa H, Albrecht D (2000) 5-HT(1A)receptor-mediated inhibition and 5-HT(2) as well as 5-HT(3)receptor-mediated excitation in different subdivisions of the ratamygdala. Synapse 38:328–337.
Thiel KJ, Wenzel JM, Pentkowski NS, Hobbs RJ, Alleweireldt AT,Neisewander JL (2010) Stimulation of dopamine D2/D3 but notD1 receptors in the central amygdala decreasescocaine-seeking behavior. Behav Brain Res 214:386–394.
Thiel KJ, Painter MR, Pentkowski NS, Mitroi D, Crawford CA,Neisewander JL (2012) Environmental enrichment counterscocaine abstinence-induced stress and brain reactivity tococaine cues but fails to prevent the incubation effect. AddictBiol 17:365–377.
Tran-Nguyen LT, Fuchs RA, Coffey GP, Baker DA, O’Dell LE,Neisewander JL (1998) Time-dependent changes incocaine-seeking behavior and extracellular dopamine levels inthe amygdala during cocaine withdrawal.Neuropsychopharmacol 19:48–59.
Wang B, Luo F, Ge XC, Fu AH, Han JS (2002) Effects of lesions ofvarious brain areas on drug priming or footshock-inducedreactivation of extinguished conditioned place preference.Brain Res 950:1–9.
Weiskrantz L (1956) Behavioral changes associated with ablationof the amygdaloid complex in monkeys. J Comp PhysiolPsychol 49:381–391.
12 L. A. Pockros-Burgess et al.