effects of salt depletion on the kidney: changes in medullary oxygenation and thick ascending limb...
TRANSCRIPT

1538 Volume 4 ‘ Number 8 ‘ 1994
Effects of Salt Depletion on the Kidney: Changes inMedullary Oxygenation and Thick Ascending Limb Size1
Isaac E. Stillman,2 Mayer Brezis, Samuel N. Heyman, Franklin H. Epstein, Kate Spokes, and
Seymour Rosen
I.E. Stillman, S. Rosen, Department of Pathology,Charles A. Dana Research Institute, Harvard Medical
School and Beth Israel Hospital, Boston, MA
SN. Heyman, F.H. Epstein, K. Spokes, Department ofMedicine, Charles A. Dana Research Institute, Harvard
Medical School and Beth Israel Hospital, Boston, MA
M. Brezis, Department of Medicine, Hadassah University
Hospital, Mt. Scopus, Jerusalem, Israel
(J. Am. Soc. Nephrol. 1994; 4:1538-1545)
tension was markedly increased from 29 ± 2 (incontrols) to 49 ± 2 mm Hg in salt-depleted rats (P <
0.0001), whereas cortical Po2 was reduced from 50± 2 to 27 ± 2 mm Hg, consistent with reduced trans-port activity In the inner stripe and/or the cortlco-meduilary redistribution of blood flow. Thus, chronic
salt depletion both diminishes mTAL size and reversesthe normal cortIcomeduilary 02 gradient, perhapsreflecting a reduced mTAL workload in conjunctionwith the reallocation of blood flow.
Key Words: Concentration, Inner stripe, cortical �2’ loop of
Henle, transport
ABSTRACTPrevious studies have shown that salt depletionenhances the susceptibility of the kidney to nephro-toxins (amphotericin, cyclosporine, and contrast). Tostudy the renal response to salt depletion, Sprague-
Dawiey rats were fed a sodium-deficient diet (N =
12)with pairfed controls (N= 13)for 4 wk. In addition,rats from each group underwent 24-h water depri-vation studies (N= 9; four salt deprived, five normal).Plastic 1-sm horizontal sections of mid-inner stripe
were examined, and cross-sectional areas of themedullary thick ascending limb (mTAL) were ana-lyzed. The mTAL ofthe salt-deprived rats were smaller(P = 0.04) and showed greater variance in size (P =
0.02) than control (618 ± 106 versus 693 ± 50 �m2).However, mean giomerular and collecting ductcross-sectional areas were unaffected by salt intake.Cross-sectional areas of long- and short-loop mTAL
were significantly different, regardless of group (518± 78 versus 732 ± 92 �m2). Maximal urinary concen-trating ability was found to correlate with mTAL cross-
sectional area (r = 0.85; P = 0.004) and with long-
loop mTAL size (r = 0.77; P = 0.016). However, it didnot significantly correlate with short loop mTAL size (1’
= 0.53; P = 0. 14). Thus, mTAL size varied with dietary
salt availability and correlated with concentratingability. In a parallel study of intrarenal oxygenation
with Clark-type 02 microelectrodes, meduilary 02
I Rc#{149}ivd April 30, 1993. Accptd September 6, 1993.2 Correspondence to Dr. I. E. 5tlllman, Department of Pathology, B&h lsrol
Hospital, 330 BrooklIne Avnue. Boston, MA 02215.
1046.6673/0408-1538$03.00/0Journal 01 th#{149}Amvlcon SocIety of NphrologyCopyrIght C 1994 by tt�. American socIety of Nephrology
T he medullary thick ascending limb (mTAL) playsan important robe in the countercurrent concen-
trating mechanism. Active chloride reabsorptionalong this water-impermeable part of the nephron,with the sodium ion following passively along theelectrical gradient, enables the mammalian kidney
to concentrate and dilute urine (1). This sodium-potassium ATPase-dependent ion transport systemis heavily dependent on aerobic metabolism (1). Be-cause of relatively low blood flow and the counter-
current exchange. a bow oxygen tension normallyexists within the renal medulla (2-5), rendering themTAL susceptible to hypoxic injury (6). IncreasedmTAL metabolic activity might increase medullary
oxygen insufficiency. This may explain the enhance-ment of mTAL hypoxic damage in hypertrophic kid-
neys, noted in vtvo (7,8) and in vitro (9), the injury tothe outer stripe associated with pobyene antibiotics(10), and the prevention of mTAL damage producedby the reduction of energy requirements (1 1,12).
Salt depletion is a well-established risk factor forthe syndrome of ATN and is known to potentiatenephrotoxins, both under experimental conditions
(7, 1 3, 1 4) and in clinical practice (1 5- 1 9). These stud-ies were designed to evaluate alterations in medullary
morphology and oxygen balance in chronically salt-depleted rats.
METHODS
Male Sprague-Dawley rats weighing 329 ± 8 g wereused for all experiments and were randomized to twoexperimental groups: control and salt depletion. Allanimal experimentation was conducted in accordwith the NIH Guide for the Care and Use of Laboratory

Stillman et al
Journal of the American Society of Nephrology 1539
Animals. Chronic salt depletion was induced over 4wk with a sodium-deficient diet (#902902; ICN Nu-tritional Biochemicals, Cleveland, OH). Rats in thesalt-depleted group were given tap water ad tibitum.
Control rats, pairfed with the same chow, were given0.25% NaC1 distilled water as their ad ttbitum drink-
ing water. The rat size was chosen to avoid the effect
of salt deprivation on the developing kidney (20).
Functional Studies
At the conclusion of the experiment, all animalswere subjected to a 24-h urine collection period, andurine and plasma samples were analyzed for thedetermination of creatinine, sodium, potassium, andurine osmolality. Standard laboratory techniqueswere used for all of these measurements. Creatinine
clearance, fractional tubular reabsorption of sodium,and fractional potassium excretion were calculatedby the use of standard formulas. In addition, in thefourth week of the experiment, randomly selectedrats (N = 4 salt deprived and five control) from eachgroup were placed in metabolic cages (Nalge Co.,
Rochester, NY) for 24 h of water deprivation andurine collection for the determination of sodium andpotassium excretion, creatinine, and maximal os-molality. These rats were given a 24-h recoveryperiod, before the initiation of terminal functionalstudies.
Measurements of Renal Parenchymal Po2
In a parallel study, intrarenal oxygen tension wasmeasured, as previously described (21 ,22), in a dif-ferent group of 1 5 salt-depleted rats, weighing 376 ±
48 g, after 1 4 ± 3 days of a saltfree diet; these resultswere compared with those obtained in 52 controlrats, as previously published (2 1). In brief, sensitive02 glass microelectrodes of the Clark type (handmadeby M.E. Traube for Omega Technologies, Haifa, Is-rael) with fine- (5- to 1 5-tim) tip diameter were used,
polarized at -0.75 V, connected to a picoampermeterand a recorder, and calibrated at 37#{176}C(with N2
and 12% 02) at the start and at the end of eachexperiment.
The rats were anesthetized with Inactin (BYKGulden, Konstanz, Germany; 100 mg/kg body wt).Tracheostomy was performed, and the femoral veinand artery were cannulated with PESO catheters(Clay-Adams, Parsippany, NJ) for the infusion of nor-mal saline with BSA (4 g/dL) (at a rate of 0.09 mL/mm) during the administration of indomethacin andfor the monitoring of the blood pressure by a pressuretransducer. The left kidney was exposed by a mid-abdominal incision and mechanically fixed. The tem-
perature of the kidney was monitored by a needlecopper probe connected to a type T thermocouple(Omega Engineering, Stamford, CT) and was kept at
37#{176}Cwith a heating lamp and by dripping warm
saline and mineral oil. Two or more electrodes,
mounted on micromanipubators, were inserted at dif-ferent depths of the kidney parenchyma.
The experiment was carried out in two phases,after an equilibration period of over 30 mm, for thestabilization of parenchymal oxygen tensions. Atfirst, repeated measurements of Po2 (averaging 13
per animal) were carried out with the electrodesplaced at various depths for the acquisition of Po2profiles in salt-depleted and control rats (total num-ber of measurements, 193 and 205, respectively). Inthe second phase of the experiment, cortical andouter medullary electrodes were positioned at depthsof 1 .5 ± 0.2 and 4.3 ± 0.4 mm, respectively. Bloodpressure and cortical and outer medullary oxygentension were monitored continuously at baseline and
over 30 mm after the iv injection of indomethacin(10 mg/kg body wt; Sigma Chemical Co., St. Louis,MO).
Morphometric Studies
At the end of the functional studies, the rats were
anesthetized with Inactin. Renal perfusion fixationin vivo was performed through the aorta at a pressure
of 140 mm Hg with 1 .25% glutaraldehyde (EastmanKodak Co., Rochester, NY) in 0. 1 M phosphate buffer(pH 7.4). The kidney that appeared more completelyfixed was selected. weighed, measured in three di-mensions, and cut in horizontal (i.e., parallel to thecorticomedullary axis) cross-sections. Selected 4 x 4-mm tissue slices were postfixed in buffered 2% 0504,
dehydrated, and embedded in an araldite-Epon 812mixture; sections were cut at 1 �m and stained with
methylene blue.Morphometric studies were performed with a Leitz
Aristoplan microscope (Wild Leitz USA, Inc., Rock-leigh, NJ), equipped with a color video camera andconnected to a color video monitor. The BioquantSystem N (versions 1 1/6/89 and 7/30/91) was usedfor morphometric analysis (R & M Biometrics, Nash-ville, TN).
Horizontal sections of the inner stripe of the outermedulla were studied, equivalent to the mid-innerstripe, or level 2, as defined by Bouby et at. (23). To
standardize the level of the examined section, weevaluated the percentage of inner stripe cross-sec-tional area that was taken up by vascular bundles,
for each slide studied. That value was not signifi-cantly different between salt-depleted and controlrats (means of 1 7 and 1 8%, respectively).
Sections of the inner stripe were examined at 50x(field area 9.9 x 1 O� � and the cross-sectionalarea of each mTAL and collecting duct (excluding thelumen) was quantified. Three full fields were exam-med per kidney, containing together 1 25 mTAL and24 collecting ducts on average.
Long- and short-looped mTAL were measured at a
Effects of Salt Depletion on the Kidney
1540 Volume 4 . Number 8 #{149}1994
separate session, with the same sections. They weredefined, respectively, by direct proximity to a vascu-bar bundle and juxtaposition to a collecting duct.
Thus, an mTAL was considered to be long looped ifit was both in contact with a vascular bundle and didnot touch a collecting duct. Conversely, a short-looped mTAL was defined as one that touched acollecting duct but did not lie adjacent to a vascular
bundle. These strict criteria resulted in many of themTAL not being counted as either a short- or long-looped mTAL.
Superficial and deep cortical sections were exam-
med as well, and all gbomeruli with clearly definedvascular poles were measured. Of these, the 20 larg-est measurements were retained for analysis.
Preliminary Studies
The morphometric study presented here followedtwo previous observations suggesting a smaller sizedmTAL in salt-depleted rats. The first observation wasmade in a study designed to investigate the synergis-tic effects of chronic amphotericin administrationand salt depletion (24). Morphometric analysis dis-
closed that the average mTAL size was 1 , 1 95 ± 48and 922 ± 45 Mm2 in control and salt-depleted ani-mals, respectively (P < 0.01 by analysis of variance
IANOVAJ). To validate this observation prospectively,a second pilot study was performed where two groupsof animals were evaluated-control and salt-dc-pleted-after 4 wk of treatment. Morphometricanalysis again revealed that the average mTAL sizewas smaller after salt depletion (887 ± 1 20 versus1 ,079 ± 279 �m2, salt-depleted rats versus controls).However, the study did not attain statistical signifi-cance because of variability among the animals ofboth groups. In addition, the animals were notpairfed. Thus, we were concerned that the findingswere, perhaps, related to protein and/or caloric mab-
nutrition. This study was therefore designed to verifythe effect of salt depletion on mTAL size in pairfedgroups of animals.
Statistics
Results are presented as the mean ± SD, in the
tables and text, and as the mean ± SE in the figures.Comparisons between the control and salt-depletedgroups were done with the independent group t testwith two-tailed P values. ANOVA with the Newman-Keuls test was applied for comparisons of repeatedP02 measurements. A P value of less than 0.05 wasconsidered to be statistically significant. Computa-tions were performed with the CRUNCH program(CRUNCH Software Corp., Oakland, CA).
RESULTS
Functional Studies
Despite initially identical body weights and rigor-
ous pairfeeding, by the end of the experiment, salt-depleted rats weighed significantly less than controls(413 ± 15 versus 443 ± 21 g; P < 0.001). Neverthe-
less, kidney weight and volume were virtually iden-tical. Salt deprivation in salt-depleted rats was con-firmed by 24-h sodium excretion (25) (Table 1). Both
groups differed by their terminal plasma concentra-tions of sodium and potassium but not by plasmaurea nitrogen, creatinine, or creatinine clearance.
Maximal urinary osmolality tended to be reduced insalt-depleted rats, but the difference between thegroups fell short of statistical significance (probablybecause of the small number of observations).
Po2 Measurements
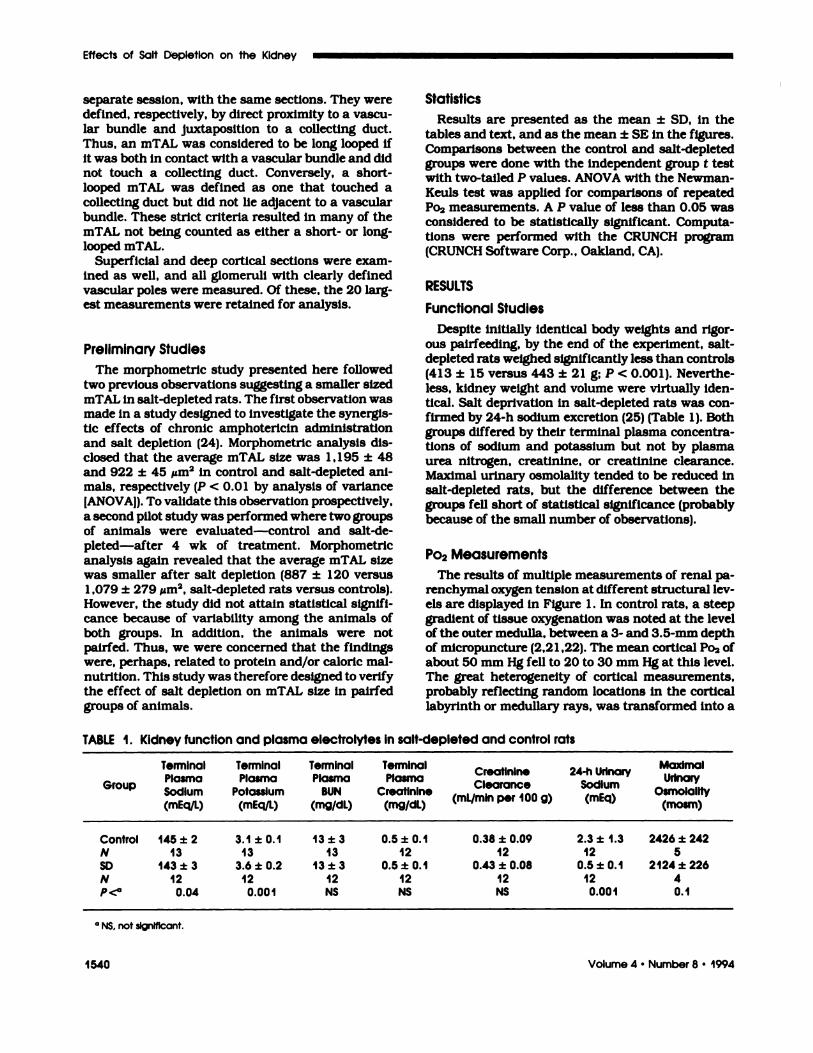
The results of multiple measurements of renal pa-renchymal oxygen tension at different structural 1ev-cbs are displayed in Figure 1 . In control rats, a steepgradient of tissue oxygenation was noted at the levelof the outer medulla, between a 3- and 3.5-mm depthof micropuncture (2,2 1 ,22). The mean cortical Po� ofabout 50 mm Hg fell to 20 to 30 mm Hg at this level.The great heterogeneity of cortical measurements,probably reflecting random locations in the cortical
labyrinth or medullary rays, was transformed into a
TABLE I . Kidney function and plasma electrolytes in salt-depleted and control rats
Group
TerminalPlasmaSodium(mEq/L)
TerminalPlasma
Potassium(mEq/L)
TerminalPlasma
BUN(mg/dL)
TerminalPlasma
Creatinine(mg/dL)
CreatinineClearance
(mi/mm per 100 g)
24-h UrinarySodium(mEg)
MaximalUrinary
0smoIali�(mosm)
Control 145±2 3.1±0.1 13±3 0.5±0.1 0.38±0.09 2.3±1.3 2426±242N 13 13 13 12 12 12 5
SD 143±3 3.6±0.2 13±3 0.5±0.1 0.43±0.08 0.5±0.1 2124±226N 12 12 12 12 12 12 4P.(o 0.04 0.001 NS NS NS 0.001 0.1
a NS. not significant.
Depth of micropuncture (mm)
Stiliman et al
Journal of the American Society of Nephrology 1541
Renal
tissue
P#{176}2
(mmHg)
CONTROL RATS SALT-DEPLETED RATS
Figure 1 . Multiple measurements of renal parenchymal P02 at various levels in control and salt-depleted rats. A sharp declineIn tissue oxygen tension occurs in controls between a 3- and 3.5-mm depth of electrode insertion, a level representing theinner stripe of the outer medulla. Substantial hypoxia normally exists within this region. In salt-depleted rats, this pattern oftissue oxygen profile Is completely inverted: low Po2 measurements are noted in the cortex, whereas medullary Po2 increases.
uniform tracing of low tissue oxygen tension withinthe deeper structures. In general, Po2 decreased withthe depth of the micropuncture.
This pattern of renal parenchymal oxygenationwas remarkably reversed in chronically salt-depletedrats. Mean cortical oxygen tension measurements
were as low as 30 mm Hg, whereas tissue Po2 grad-ually rose at the corticomedullary junction, reachingmean values of about SO mm Hg. The mean oxygentension measurements were significantly different (P
< 0.000 1) at every millimeter depth, except for thethird, where there was no significance. ANOVA (two-way) for salt and depth of Po2 gave an F value of 31
(P < 0.000 1 for salt plus depth [P < 0.02 for salt only;
P < 0.001 for depth onlyl). Thus, in salt-depletedrats, as opposed to controls, Po2 directly increasedwith depth of micropuncture.
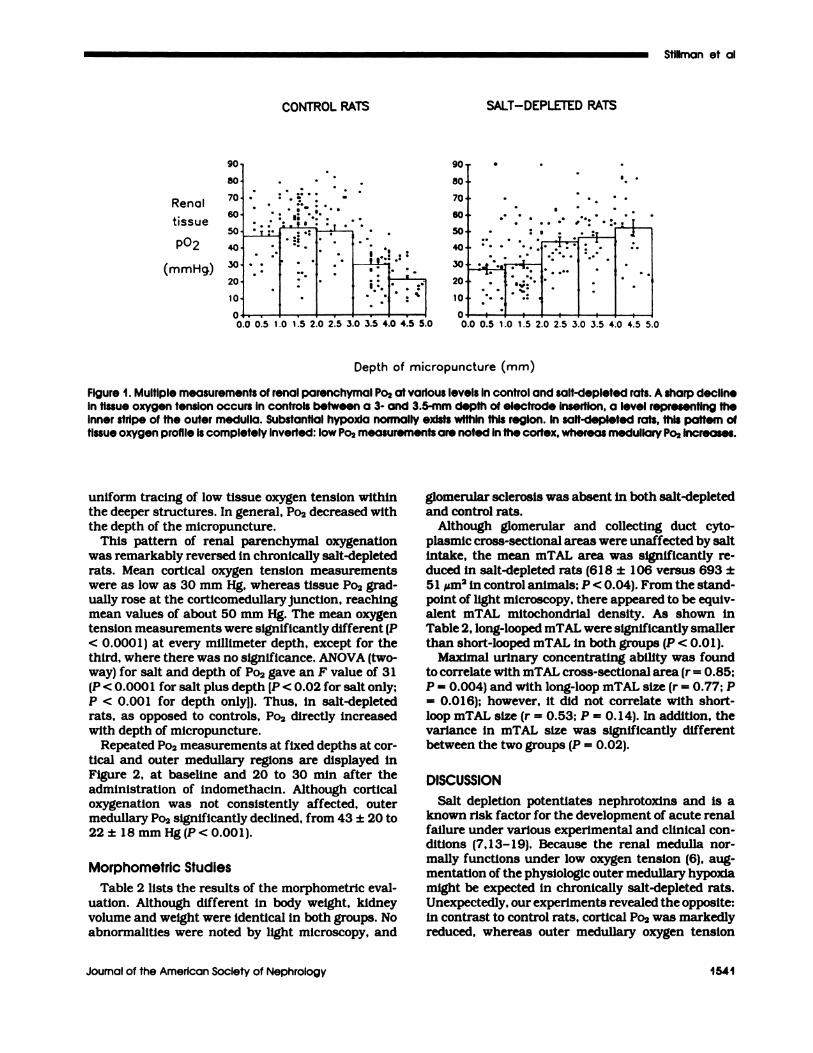
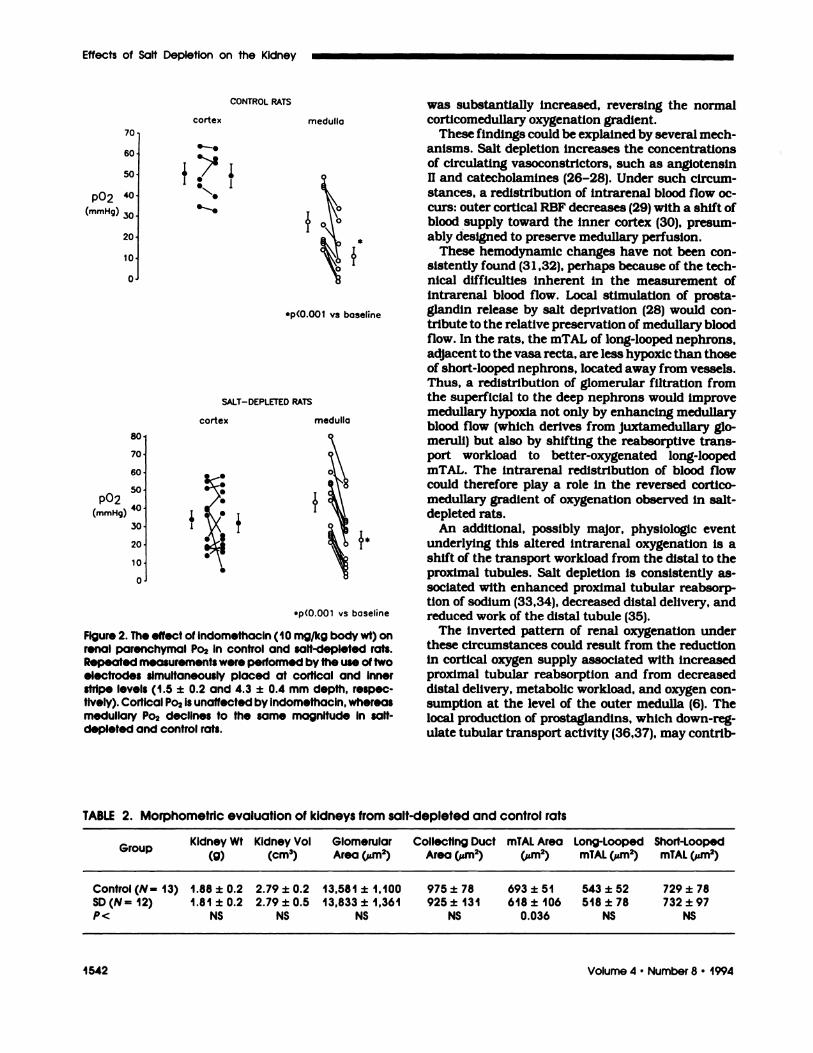
Repeated Po2 measurements at fixed depths at cor-tical and outer medullary regions are displayed inFigure 2, at baseline and 20 to 30 mm after theadministration of indomethacin. Although corticaloxygenation was not consistently affected, outermedullary Po2 significantly declined, from 43 ± 20 to22± 18 mm Hg(P<O.OO1).
Morphometric Studies
Table 2 lists the results of the morphometric evab-
uation. Although different in body weight, kidneyvolume and weight were identical in both groups. Noabnormalities were noted by light microscopy, and
gbomerubar sclerosis was absent in both salt-depleted
and control rats.Although gbomerular and collecting duct cyto-
plasmic cross-sectional areas were unaffected by saltintake, the mean mTAL area was significantly re-duced in salt-depleted rats (6 1 8 ± 1 06 versus 693 ±
51 �m2 in control animals; P < 0.04). From the stand-point of light microscopy, there appeared to be equiv-alent mTAL mitochondrial density. As shown in
Table 2, bong-looped mTAL were significantly smallerthan short-looped mTAL in both groups (P < 0.01).
Maximal urinary concentrating ability was foundto correlate with mTAL cross-sectional area (r = 0.85;P = 0.004) and with long-loop mTAL size (r = 0.77; P= 0.016); however, it did not correlate with short-loop mTAL size (r = 0.53; P = 0. 14). In addition, thevariance in mTAL size was significantly different
between the two groups (P = 0.02).
DISCUSSION
Salt depletion potentiates nephrotoxins and is aknown risk factor for the development of acute renalfailure under various experimental and clinical con-ditions (7, 1 3- 1 9). Because the renal medulla nor-
mally functions under low oxygen tension (6), aug-mentation of the physiologic outer medulbary hypoxiamight be expected in chronically salt-depleted rats.
Unexpectedly, our experiments revealed the opposite:in contrast to control rats, cortical P02 was markedlyreduced, whereas outer medullary oxygen tension
CONTROL RATS
cortex medulla
.�.
4
ep(O.OO1 vs baseline
SALT-DEPLETED RATS
cortex medulla
70
60
50
P#{176}240
(mmHg) .�o.
20
10�
0�
80�
70
60#{149}
50#{149}PO2
40#{149}(mmHg)
30�
20
10#{149}
0�
. .
Effects of Salt Depletion on the Kidney
1542 Volume 4 #{149}Number 8 #{149}1994
#{149}p(O.OOl vs baseline
Figure 2. The effect of indomethacin (10 mg/kg body wt) onrenal parenchymal Po2 in control and salt-depleted rats.Repeated measurements were performed by the use of twoelectrodes simultaneously placed at cortical and Innerstripe levels (1 .5 ± 0.2 and 4.3 ± 0.4 mm depth, respec-tively). Cortical Po2 Is unaffected by indomethacin, whereasmedullary Po2 declines to the same magnitude in salt-depleted and control rats.
was substantially increased, reversing the normalcorticomedullary oxygenation gradient.
These findings could be explained by several mech-
anisms. Salt depletion increases the concentrationsof circulating vasoconstrictors, such as angiotensinII and catecholamines (26-28). Under such circum-
stances, a redistribution of intrarenal blood flow oc-curs: outer cortical RBF decreases (29) with a shift ofblood supply toward the inner cortex (30), presum-ably designed to preserve medullary perfusion.
These hemodynamic changes have not been con-sistently found (31 ,32), perhaps because of the tech-nical difficulties inherent in the measurement ofintrarenal blood flow. Local stimulation of prosta-glandin release by salt deprivation (28) would con-
tribute to the relative preservation of medullary bloodflow. In the rats, the mTAL of long-looped nephrons,
adjacent to the vasa recta, are less hypoxic than thoseof short-looped nephrons, located away from vessels.Thus, a redistribution of glomerular filtration fromthe superficial to the deep nephrons would improvemedullary hypoxia not only by enhancing medublaryblood flow (which derives from juxtamedullary gb-meruli) but also by shifting the reabsorptive trans-port workload to better-oxygenated long-loopedmTAL. The intrarenal redistribution of blood flowcould therefore play a role in the reversed cortico-medullary gradient of oxygenation observed in salt-depleted rats.
An additional, possibly major, physiologic event
underlying this altered intrarenal oxygenation is ashift of the transport workload from the distal to theproximal tubules. Salt depletion is consistently as-sociated with enhanced proximal tubular reabsorp-tion of sodium (33,34), decreased distal delivery, andreduced work of the distal tubule (35).
The inverted pattern of renal oxygenation underthese circumstances could result from the reductionin cortical oxygen supply associated with increasedproximal tubular reabsorption and from decreased
distal delivery, metabolic workload, and oxygen con-sumption at the level of the outer medulla (6). Thelocal production of prostaglandins, which down-reg-
ulate tubular transport activity (36,37), may contrib-
TABLE 2. Morphometric evaluation of kidneys from salt-depleted and control rats
Group
Kidney WI�
Kidney Vol(cm3)
GiomerularArea (�m2)
Collecting DuctArea (�m2)
mTAL Area(�m2)
Long-LoopedmTAL (�m2)
Short-LoopedmTAL (�m2)
Controi(N= 13) 1.88±0.2 2.79±0.2 13,581±1,100 975±78 693±51 543±52 729±78
SD(N=12) 1.81±0.2 2.79±0.5 13,833±1,361 925±131 618±106 518±78 732±97
P< NS NS NS NS 0.036 NS NS

Stiliman et al
Journal of the American Society of Nephroiogy 1543
ute to this improvement in medullary oxygen bal-
ance. As illustrated in Figure 2, the inhibition ofprostaglandin synthesis by indomethacin reduced
outer medullary Po2, suggesting that increased pros-taglandin release may participate in the reversedprofile of intrarenal oxygenation noted in chronically
salt-depleted rats. However, it is to be noted that
prostaglandin secretion is important in the normalstate because the medulbary P02 in the controls was
reduced as well.We have recently observed a similar reversal of the
renal parenchymal oxygenation profile during acutehypotension (38). Paradoxically, acute reductions inrenal perfusion (produced by aortic ligation, hemor-rhage, or nitroprusside infusion) increased medullary
Po2, while reducing cortical Po2. In the presence offurosemide (which inhibits solute reabsorption in themTAL and increases medullary P02), acute hypoten-sion did not further increase Po2 in the medulla.These data suggest that the hypotension-induced re-versal of the corticomedullary oxygen gradient de-rives at least in part from a reduction in transportworkload in the outer medulla.
Our morphometric findings suggest an additionalmechanism for decreased oxygen consumption in the
outer medulla. The mean cross-sectional area ofmTAL cells at the level of the inner stripe of the outermedulla was smaller in salt-deprived rats, as corn-
pared with that in controls. Although both groups didnot differ by mitochondrial density, as assessed bylight microscopy, it is conceivable that oxygen con-
sumption may correlate with the reduction in mTALcellular mass. Supporting this hypothesis is the cor-relation found between mTAL cellular mass and
maximal urinary concentration. Because the latter isthe product of energy-dependent ion transport that
takes place in mTAL, a lower oxygen requirement isanticipated when cellular mass is reduced.
The association of chronic salt depletion with
trophic changes in the inner stripe is an additionalexample of the remarkable plasticity of the mTAL inresponse to various pathophysiobogic changes (39).
Vasopressin or water restriction (23), thyroid hor-mones (40), and a high-protein diet (41) have beenobserved to increase rnTAL cell size, often dispropor-
tionately to trophic changes in the remainder of thenephron. In addition, the activity of the Na-K-ATPasein mTAL Is increased by antidiuretic hormone (42),
mineralocorticoid (43), and high protein intake(44,45). Variation of salt intake may also havetrophic effects on the rnTAL. Increased dietary salt
load has been shown to stimulate the Na-K-ATPaseof mTAL segments In the rabbit (46). Sodium depri-vation in the mouse did not cause a clear change inrnTAL enzyme activity, probably reflecting the resultof opposing influences of decreased sodium deliveryand increased endogenous aldosterone (43). This ef-
fect contrasted with the stimulation of enzyme activ-
ity in the cortical collecting tubule (known to respondto mineralocorticoids).
Several studies have indicated a trophic robe of salton the kidney. A high-salt diet increased renal sizeand protein content, indicating stimulation of renalgrowth due to hyperplasia in the rat (47,48). Saltintake may be particularly important in modulating
the hypertrophic response of remnant kidneys. Re-cent studies have shown that dietary salt restrictionlessens renal damage by inhibiting compensatorykidney growth after severe renal ablation (49-S 1).The mechanism of protection, apparently independ-ent of changes in intragbomerular hypertension, mayrelate to some trophic effect of sodium chloride on
the renal parenchyma. In this study, the reduction
in epithelial volume after salt depletion was morereadily apparent in mTAL than in other nephronsegments, perhaps because of the greater plasticityof the mTAL in response to various stimuli (22,39-41). Recent insights into the regulation of geneexpression by extraceblular tonicity in the renal me-
dulla (52) may help in the future to clarify the mech-anism(s) of this trophic modulation of mTAL by so-dium chloride.
The trophic effect of salt upon the kidney may bemediated through growth factors. Augmenting distal
tubular workload with furosemide induced hypertro-phy (53) and increased immunoreactivity for insulin-like growth factor (IGF- 1) and its binding proteins inthe distal nephron (54). An altered IGF-1 signal,largely confined to the outer medulla (55), wouldsecondarily affect receptors locally and throughoutthe renal cortex (55) in an autocrine or paracrine
fashion to induce hemodynamic and growth signals.Looking at the stirnulatory effect of sodium on kidney
growth after injury (56), Finn and Yang recently sug-gested the presence of a circulating factor modifiedby salt intake and abbe to influence tubular epitheliabcell growth. Studies of the effects of dietary salt uponrenal tissue and circulating IGF and/or other growthfactors would be of great interest.
As previously reported (39), regardless of salt in-take, mTAL cell volume in short-looped nephronswas consistently larger than in long-looped neph-rons. Bankir has suggested (personal communica-tion) that epithelial cell size gradually decreases asthe thick limb ascends from its origin. Short-loopedmTAL would, therefore, be larger at any level simplybecause the cells are closer to the start of thesegment.
In conclusion, chronic salt depletion inverts theprofile of renal tissue oxygenation. Cortical Po2 falls,while medullary hypoxia is ameliorated. This processmaintains inner stripe oxygenation under conditionsthat would otherwise have resulted in substantialhypoxic injury. The increase in medulbary P02 may

Effects of Salt Depletion on the Kidney
1544 Volume 4 ‘ Number 8 . 1994
result from the redistribution of RBF and the reduc-
tion of oxygen requirements, in part due to reduced
rnTAL mass.
ACKNOWLEDGMENTS
This work was supported by Grants DK 1 8078 and DK39249 from
the NIH and grants from the BIH Pathology Foundation, Inc., and
from the USA-Israel Binatlonal Science Foundation. We thank Lena
Ellezian for her excellent technical assistance.
REFERENCES
1 . Dc Rouffignac C: The urinary concentratingmechanism. In: Kinne RKH, Ed. Urinary Con-centrating Mechanisms. Vol. 2. Base!: Karger;1990:31-102.
2. Leichtweiss HP, Lubbers DW, Weiss C, Baum-garth H, Reschke W: The oxygen supply of therat kidney: Measurements of intrarenal PO2.Pflugers Arch 1969;309:328-349.
3. BaUmgartl H, Leichtweiss HP, Lubbers DW,Weiss C, Huland H: The oxygen supply of thedog kidney: Measurements oT intrarenal p02.Microvasc Res 1972;4:247-257.
4. Leonhardt KO, Landes RR: Oxygen tension ofthe urine and renal structures. N Engl J Med1963;269:1 15-121.
S. Epstein FH, Balaban RS, Ross BD: Redox stateof cytochrome a,a3 in isolated perfused kidney.Am J Physiol 1 98 1 ;243:3S6-363.
6. Brezis M, Rosen 5, Silva P, Epstein FH: Renalischemia: A new perspective. Kidney mt 1984;26:375-383.
7. Heyman SN, Brezis M, Reubmoff CA, et at.:Acute renal failure with selective medullary in-jury in the rat. J Clin Invest 1988;82:401-412.
8. Heyman SN, Brezis M, Epstein FH, Spokes K,Rosen S: Effect of glycine and hypertrophy onrenal outer medullaryhypoxic injury in ischemiareflow and contrast nephropathy. Am J KidneyDis 1992;19:S78-S86.
9. Epstein FH, Silva P, Spokes K, Brezis M, Rosen5: Renal medullary Na-K-ATPase and hypoxicinjury in perfused rat kidneys. Kidney mt1989;36:768-772.
1 0. Brezis M, Rosen S, Silva P, Spokes K, EpsteinFH: Polyene toxicity in renal medulla. Injury me-diated by transport activity. Science 1984;224:66-68.
1 1 . Brezis M, Rosen S, Silva P, Epstein FH: Trans-port activity modifies thick ascending limb dam-a�e in the isolated perfused kidney. Kidney Int1�84;2S:6S-72.
12. Heyman SN, Brezis M, Greenfeld Z, Rosen 5:Protective role of furosemide and saline in radi-ocontrast-induced acute renal failure in the rat.Am J Kidney Dis 1989; 14:377-385.
1 3. Rosen 5, Greenfeld Z, Brezis M: Chronic cycbo-sporine-induced nephropathy in the rat: A mcd-ullary ray and inner stripe injury. Transplanta-tion 1990;49:445-452.
14. Ohnishl A, Ohnishi T, Stevenhead W, et at.:Sodium status influences amphotericin nephro-toxicity in rats. Antimicrob Agents Chemother1989;�3: 1222-1227.
1 S. Branch RA, Jackson EK, Jacqz E, et at.: Am-photericin B nephrotoxicity in humans de-
creased by sodium supplements with coadmin-istration of ticarcillin or intravenous saline. KlinWochenschr 1 987;65:500-506.
16. Lianos A, Cieza J, Bernardo J, et at.: Effect ofsalt supplementation on amphotericin B neph-rotoxicity. Kidney Int 1991 ;40:302-308.
17. Heidemann HTH, Gerkens JF, Spickard WA,Jackson EK, Branch PA: Amphotericin B neph-rotoxicity in humans decreased by salt repletion.Am J Med 1983:75:476-481.
18. Rasmussen HH, Ibels IS: Acute renal failure.Mubtivariate analysis of causes and risk factors.Am J Med 1982;’73:21 1-2 18.
1 9. Shusterman N, Storm BL, Murray TG, MorrisonG, West SL, Maislin G: Risk factors and outcomeof hospital-acquired acute renal failure. Am JMed 1987;83:6S-71.
20. Solomon 5, Romero C, Moore L: The effect ofage and salt intake on growth and renal devel-opment of rats. Arch Int Physiol Biochim1972;80:87 1-882.
2 1 . Brezis M, Heyman SN, Dinour D, Epstein FH,Rosen 5: Role of nitric oxide in renal medullaryoxygenation. Studies in isolated and intact ratkidneys. J Clin Invest 1991 ;88:390-395.
22. Heyman SN, Brezis M, Epstein FH, Spokes K,Silva P Rosen 5: Early renal medullary hypoxicinjury from radiocontrast and indomethacin.Kidney mt 1991;40:632-642.
23. Bouby N, Bankir L, Trinh-Trang-Tan MM, Mm-uth WW, Krltz W: Selective ADH-induced hyper-trophy of the medullary thick ascending limb inBrattleboro rats. Kidney Int 1 98S;28:456-466.
24. Heyman SN, Stillman IE, Brezis M, Epstein FH,Spokes K, Rosen 5: Chronic amphotericin neph-ropathy: Morphometric, electron microscopic,and functional studies. J Am Soc Nephrol 1993;4:69-80.
25. Brensilver JM, Daniels FH, Lefavour GS, et at.:Effect of variations in dietary sodium intake onsodium excretion in mature rats. Kidney mt1 98S;27:497-S02.
26. Hall JE, Guyton AC, Smith MJ Jr, Coleman TG:Chronic blockade of angiotensin H formationduring sodium deprivation. Am J Physiob1 979;237:F424-F432.
27. Friberg P, Meredith I, Jennings G, Lambert G,Fazio V, Esler M: Evidence for increased renalnorepinephrine overflow during sodium restric-tion in humans. Hypertension [99O;6: 1 2 1 -130.
28. Oliver JA, Pinto J, Sciacca RR, Cannon PJ:Increased renal secretion of norepinephrine andprostaglandin E2 during sodium depletion in thedog. J Clin Invest 1980;66:718-756.
29. Blaufox MD, Fromowltz A, Lee HB, Meng CH,Elkin M: Renal blood flow and renin activity inrenal venous blood in essential hypertension.Clrc Res 1970;27:913-920.
30. Mercer PF, Zusko EA: Whole kidney and corticalblood flow in rats on high and low sodium diets.J Physiol 1974;237:49�-5O4.
3 1 . Chou SY, Spitalewitz 5, Faubert PF, Park IY,Porush JG: Inner medullary hemodynamics inchronic salt-depleted dogs. Am J Physiob1984;246:F146-F 154.
32. Cambridge D, Chapman BJ, Haworth P, Mun-day KA, Vas Dias FW, Wilson RA: Intrarenaldistribution of blood flow in the rat measured by86Rb uptake and the effects of high- and low-sodium diets. J Physiol 1976;258:36P-37P.

Stillman et ol
Journal of the American Society of Nephrology I 545
33. Weiner MW, Weinman EJ, Kashgarian M, Hays-lett JP: Accelerated reabsorption in the proximaltubule produced by volume depletion. J Clin In-vest 1971:50:1379-1385.
34. Mohammad G, Di Scala V, Stein RM: Effects ofchronic sodium depletion on tubular sodium andwater reabsorption in the dog. Am J Physiol1 974;227:469-476.
35. Stein RM, Levitt BH, Goldstein MH, Porush JG,Eisner GM, Levitt MF: The effects of salt restric-tion on the renal concentrating operation in nor-ma!, hydropenic man. J Clin Invest 1962;41:2101-2111.
36. Lear S, Silva P. Kelley VE, Epstein FH: Prosta-glandin E2 inhibits oxygen consumption in rab-bit medulbary thick ascending limb. Am J Phys-iol 1990;258:F1372-F1378.
37. Wald H, Scherzer P, Rubinger D, PopovtzerMM: Effects of indomethacin in vivo and PGE2in vitro on mTAL Na-K-ATPase of the rat kidney.Pf!#{252}�ers Arch 1990;415:648-650.
38. Brezis M, Heyman SN, Fuhro R, Epstein FH:Determinants of intrarenal oxygenation gra-dients. Clin Res 1991;32:389A.
39. Bankir L, Bouby N, Trinh-Trang-Tan MM: Het-erogeneity of nephron anatomy. Kidney Int1987;31(Suppl 201:S25-S39.
40. Davis RG, Madsen KM. Fregly MJ, Tisher CC:Kidney structure in hypothyroidism. Am J Pa-thol 1983:113:41-49.
4 1 . Bouby N, Trinh-Trang-Tan MM, Laouari D, etat. : Role of urinary concentrating process in therenal effect of high protein intake. Kidney mt1 988;34:4- 12.
42. Trinh-Trang-Tan MM, Bankir L, Doucet A, etat. : Influence of chronic ADH treatment on aden-ylate cyclase and ATPase activity in distal neph-ron segments of diabetes insipidus Brattlebororats. Pflugers Arch 1985:405:216-222.
43. Grossman EB, Herbert SC: Modulation of Na-K-ATPase activity in the mouse medullary thickascending limb of Henle. Effects of mineralocor-ticoids and sodium. J Clin Invest 1988;81:885-892.
44. Bouby N, Chayen J, Trinh-Trang-Tan MM, Ban-kir L: Effect of a high protein intake on the Na-K-ATPase activity in the thick ascending limb ofHenle’s loop [Abstractj. Kidney Int 1986:30:638.
45. Seney FD Jr. Marver D: Effect of dietary proteinon medullary thick ascending limb Na-K-ATPase
activity lAbstract]. Clin Res 1986;34:609A.46. Scherzer P, Wald H, Czaczkes JW: Na-K-ATP-
ase in isolated rabbit tubules after unilateralnephrectomy and Na loading. Am J Physiol1985;248:F565-F73 1.
47. Binet L, Dejours P, Lacaisse A: Action du chbo-rure de sodium alimentaire sur le d#{233}veloppementpond#{233}ral du rein chez le rat. Comptes Renal SocBiol 1950:144:84-87.
48. McCormick CP, Rauch AL, Buckalew VM Jr:Differential effect of dietary salt on renal growthin Dahl salt-sensitive and salt-resistant rats. Hy-pertension 1 989; 13:122-127.
49. Benstein JA, Feiner HD, Parker M, DworkinLD: Superiority of salt restriction over diureticsin reducing renal hypertrophy and injury inuninephrectomized SHR. Am J Physiob 1990;258:F167S-F1681.
50. Lax DS, Benstein JA, Tolbert E, Dworkin LD:Effects of salt restriction on renal growth andglomerular injury in rats with remnant kidneys.Kidney Int 1992;1:1527-1534.
5 1 . Terzi F, Beaufils H, Laouari D, Burtin M,Kleinknecht C: Renal effects of anti-hyperten-sive drugs depend on sodium diet in the excisionremnant kidney model. Kidney mt 1992;42:354-363.
52. Burg MB, Garcia-Perez A: How tonicity regu-bates gene expression lEditorlall. J Am Soc Ne-phrol 1992:3:121-127.
53. Kobayashi 5, Venkatachalam MA, Roy AK,Stein JH: Work load induced by furosemide in-creases IGF- 1 and IGF-BP- 1 in rat distal nephron[Abstract]. Second International Symposium onInsulin-Like Growth Factors/Somatomedins;Growth Regul 199 1 ; 1:141.
54. Kaissling B, Stanton BA: Adaptation of distaltubule and collecting duct to increased sodiumdelivery. I. Ultrastructure. Am J Physiol 1988;25S:F1256-F1268.
55. Mulroney SE, Haramati A, Werner H, Bondy C,Roberts CT Jr, Leroith D: Altered expression ofinsulin-like growth factor-I (IGF-I) and IGF recep-tor genes after unilateral nephrectomy in im-mature rats. Endocrinology 1 992; 130:249.
56. Finn WF, Yang JJ: Influence of dietary sodiumon thymidine uptake following renal ischemia:
.In vivo and in vitro studies [Abstract]. J Am SocNephrol 1992;3:70S.