effect of dietary supplementation with vitamin e for lactating dairy cows fed tall fescue hay...
TRANSCRIPT

0021
doi:1
Corre
Effect of Dietary Supplementation with Vitamin Eand Stocking Density on Macrophage Recruitment
and Giant Cell Formation in the Teleost Fish,Piaractus mesopotamicus
M. A. A. Belo*, S. H. C. Schalch†, F. R. Moraes*†, V. E. Soares*,A. M. M. B. Otoboni‡ and J. E. R. Moraes*
*Department of Veterinary Pathology, †Aquaculture Centre and ‡Department of Technology, Sao Paulo State University
(Unesp), Jaboticabal, Brazil
Summary
-99
0
s
The effect of dietary supplementation with 0, 100 and 450 mg of vitamin E (DL-a tocopherylacetate)/kg of a dry diet on the kinetics of macrophage recruitment and giant cell formation in thepacu, maintained at different stocking densities (5 kg/m3 and 20 kg/m3), was investigated by insertionof round glass coverslips into the subcutaneous connective tissue. After a feeding period of 18 weeks,the coverslips were implanted and later removed for examination at 2, 7 and 15 days post-implantation. Fish fed diets supplemented with 450 mg of vitamin E showed an increase (P!0.05) inthe accumulation of macrophages, foreign body giant cells and Langhans type cells. The kinetics ofmacrophage recruitment and giant cell formation on the glass coverslips appeared to be stronglyinfluenced by vitamin E supplementation, since fish fed a basal diet and held at high stockingdensities showed low numbers of adhering cells on the coverslips, and high concentrations of plasmacorticosteroids. On the other hand, fish given a diet supplemented with 450 mg of vitamin E did notshow a similar difference in plasma cortisol concentrations related to stocking density. The effect ofcortisol concentrations on carbohydrate metabolism, analysed by assessment of plasma glycaemia, wasnot clear. Blood glucose concentrations did not vary substantially with the different treatmentsexamined. These results suggest that vitamin E may contribute to the efficiency of the fish’sinflammatory response by increasing macrophage recruitment and giant cell formation in the foreignbody granulomatous reaction. Vitamin E appeared to act on the stress response of pacus by preventinga stress-related immunosuppression.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: alpha-tocopherol; cortisol; fish; Piaractus mesopotamicus; polykaryon macrophage; vitamin E
Introduction
Immunostimulants are valuable for controllingdisease and may be useful in fish production(Sakai, 1999). Vitamin E is an important nutrientwhich acts as a lipid-soluble antioxidant, protecting
75/$ - see front matter
.1016/j.jcpa.2005.04.004
pondence to: F.R. Moraes.
biological membranes and lipoproteins againstoxidation, as well as being an essential dietarynutrient for fish (Bai and Lee, 1998; Montero et al.,1999). The ingestion of large amounts of dietaryvitamin E and its effect on the immune system offish has been investigated. Wise et al. (1993 a,b)showed that channel catfish fed high doses ofvitamin E had increased leucocyte phagocytic
J. Comp. Path. 2005, Vol. 133, 146–154
www.elsevier.com/locate/jcpa
q 2005 Elsevier Ltd. All rights reserved.

Table 1Ingredients of the basal diet
Ingredients %
Corn gluten meal 21.3Wheat bran 16.1Soybean meal 43.5Rice bran 10Yeast, brewers, dehydrated 8Methionine 0.27Mineral Premix* 0.25Vitamin Premix (no C or E)† 0.45Vitamin C 0.047
* The mineral premix (NUTREMIX Racoes Ltd, Monte Alto, Brazil)supplied the following to each kg of the basal diet: iron, 15 000 mg;copper, 5000 mg; iodine, 500 mg; manganese, 17 000 mg; zinc, 12000 mg; selenium, 70 mg.† The vitamin premix (NUTREMIX Racoes Ltd.) supplied thefollowing to each kg of the basal diet: vitamin A, 12 000 IU; vitaminD3, 1500 UI; vitamin K, 4 mg; vitamin B12, 7 mg; pantothenic acid,60 mg; ascorbil 2-poliphosphate, 200 mg; nicotinic acid, 120 mg;choline chloride, 600 mg.
Macrophages and Giant Cells in Fish 147
activity and superoxide anion production, andOrtuno et al. (2001) reported that gilthead sea-bream, Sparus aurata, fed a vitamin E-sup-plemented diet exhibited increased complementand phagocytic activities.
Inflammatory giant cells are formed in vivo bycell membrane fusion of blood-borne macro-phages, derived from circulating monocytes(Gillman and Wright, 1966; Mariano andSpector, 1974). Macrophage polykaryons in fishoccur in a variety of pathological processes, suchas infectious diseases or foreign body granulomas(Goodwin and Grizzle, 1991). The presence ofthese cells has been described in infections suchas Edwardsiella tarda in Nile tilapia (Oerochromisniloticus) (Miyazaki and Kaige, 1985), Ichthyopho-nus hoferi in plaice (Pleuronectes platessa L.)(Timur and Timur, 1984), Microsporidium sp. inNorway pout (Trisopterus esmarkii) (Pulsford andMatthews, 1991) and Mycobacterium marinum insea bass (Dicentrarchus labrax) (Colorni et al.,1998).
In several species of fish, crowding has beenreported to be an aquaculture-related chronicstress factor which reduces growth and affects theinflammatory and immune responses (Monteroet al., 1999). Cortisol is the main corticosteroid infish and its plasma concentrations are widely usedas both general and endocrine-related indicators ofstress (Pickering and Pottinger, 1989). Cortisol isan anti-inflammatory hormone that limits theincrease of permeability in blood vessels and,consequently, cell accumulation at the site ofinflammation (Garcia Leme, 1981).
Few studies have analysed the effect of vitamin Eon the inflammatory response of fish or thepossible effect produced by the interaction ofdietary supplementation and stocking density.Although the intake level of specific nutrientscan determine better organic responses andcrowding stress can increase disease outbreaks,few concomitant changes in the immunologicalparameters have been observed on these occasions(Hardie et al., 1990, 1991). With this in mind, anin-vivo study was performed to investigate theeffect of high doses of vitamin E or elevatedstocking density, or both, on the kinetics ofmacrophage recruitment and giant cell formation,as observed on round glass coverslips inserted intothe subcutaneous tissue of fish. The species chosenwas the pacu (Piaractus mesopotamicus,Osteichthyes:Characidae), a native teleost fish ofthe Parana-Paraguay Basin, of importance in Brazilin relation to human consumption, angling andaquaculture.
Materials and Methods
Experimental Procedure
The experiment was carried out at Sao Paulo StateUniversity. Pacus (nZ810), of average weight96.42 g (SDG25.23) and all originating from thesame spawn, were obtained from Aguas Claras Farm(Mococa, Brazil). The fish were initially distributedbetween three earth tanks of 51 m3 (270 fish pertank, at a flow rate of 1 litre/sec), where they werekept for 16 weeks on either a basal diet or on one oftwo experimental diets. After this period, fish fromeach tank were randomly redistributed between sixaquaria (i.e., 18 aquaria in all) each containing200 litres of water (flow rate 1 litre/min) at 29.7G1.8 8C. The aquaria, which were exposed to naturalperiods of light, contained fish allocated to obtain“control” and “crowded” stocking densities(5 kg/m3 and 20 kg/m3, respectively). Fish wereallowed to acclimatize to the new environment for15 days before implantation of coverslips. Sup-plementary aeration was provided to maintaindissolved oxygen near saturation point. On each ofthree occasions (2, 7 and 15 days post-implan-tation), seven fish from six aquaria were sampled(i.e., for 42 necropsies). During the experimentalperiod, the water was examined twice weekly forammonia and nitrite concentrations, and twice daily(at 7 am and 6 pm) for temperature, pH, electricalconductivity and dissolved oxygen.
Diets
The ingredients and nutritional values of thebasal diet are shown in Tables 1 and 2, respectively.

Table 2Nutritional values of the basal diet (dry matter basis)
Components %
Crude protein 27.16Crude fibre 2.97Ether extract 2.29Ash 5.35Calcium 0.21Phosphorus 0.51Nitrogen-free extract 41.86Dry matter 79.64
Composition calculated according to A.O.A.C. (1975). Thedigestible energy was 3650 kcal per kg of dry diet.
M.A.A. Belo et al.148
Three diets were prepared, to contain DL-a-tocopheryl acetate supplement at the followingconcentrations (mg/kg dry weight): 0 (basal diet),100 and 450. The basal diet was formulated withoutoil for provision of low vitamin E rates in an attemptto induce nutritional vitamin deficiency, since thedietary vitamin E requirement for young pacus hasnot yet been established.
All fish were fed twice a day (at 8 am and 5 pm)for 20 weeks, at a feeding rate of 2% of body weightper day. At the end of the feeding trial, the dietswere analysed for a-tocopherol by high perform-ance liquid chromatography (HPLC). The feedpellets were stored at K208 C before use.
Cell Evaluation on the Coverslips
Seven fish from each of the 18 groups wereanaesthetized with a water containing benzocaine(1 in 10 000) before coverslip implantation asdescribed by Petric (2000). Granulomatous inflam-matory responses were generated by insertinground glass coverslips (13 mm in diameter) intothe subcutaneous connective tissue of each fish(Mariano and Spector, 1974). At 2, 7 and 15 dayspost-implantation, fish were killed by an overdoseof a benzocaine aqueous solution (1 in 500), beforethe coverslips were removed, fixed in Bouin’s fluid,stained with haematoxylin and eosin (HE) andmounted for examination by light microscopy.
The number of cells adherent to the glasscoverslips was estimated microscopically with a!16 objective integrated to a Video Plan computersystem (Model KS-100, Version 3.0, Carl Zeiss). Foreach coverslip, five fields were randomly chosenand all cells counted; there were thus 35 counts pergroup. The number of nuclei present in multi-nucleated cells was also determined and cellmorphology was used to identify them as being aforeign body cell or a Langhans giant cell.
Blood Collection and Analysis
For the collection of blood samples at 2, 7 and 15days post-implantation, fish were briefly removedfrom the aquaria and blood was withdrawn into asyringe from a caudal vessel by needle puncture.The blood was immediately dispensed into two setsof heparinized tubes for separate analysis of cortisoland glucose concentrations in the plasma. Theblood was centrifuged at 3000 rpm (1700 g, at 48C)for 10 min and the plasma was either stored atK20 8C for cortisol analysis or processed immedi-ately for glucose determination. Cortisol concen-trations were determined by radioimmunoassaywith “DPC-Kits” (Gazola et al., 1996). Glucoseconcentrations were assessed colorimetrically, asdescribed by King and Garner (1947).
Diet Analysis
The experimental diets were analysed fora-tocopherol by HPLC as described by Manz andPhilipp (1981). This assay included saponificationof the samples and extraction of the unsaponifiablecomponents with a methanol:water mixture (96:4)as the mobile phase, containing butylhydroxyto-luene (BHT) as an antioxidant and potassiumhydroxide solution, followed by reversed-phasechromatography with fluorescence detection of a-tocopherol. The flow rate was 1.1 ml/min, and thedetection carried out at 295 to 345 nm. Calcu-lations were based on a comparison of the peakareas of a-tocopherol from the samples and thoserecovered from standard solution.
Growth
After 112 days, specific growth rates (SGRs: % bodyweight/day) for each group of fish were deter-mined by the equation SGRZ100ðlogWf
n K logWon Þ=t,
where Wo and Wf were the initial and final weightof each experimental group, respectively, after tdays (Ricker, 1979 cited by Ortuno et al., 2001).
Statistical Analysis
All data were examined statistically with a factorialscheme “split plot in time” (three levels of vitaminE [0, 100 and 450 mg/kg dry diet]!two levels ofdensity [5 and 20 kg/m3] ! three occasions [2, 7and 15 days post-implantation]). Comparison ofthe different experimental groups was carried outby applying a GLM procedure (General LinearModels). Significant differences (P!0.05) wereestimated on the basis of Tukey’s test according toSnedecor and Cochran (1974).
Macrophages and Giant Cells in Fish 149
Results
No mortality or abnormal behaviour attributable tothe treatments was observed throughout the exper-iment. The specific growth rates calculated after112 days (i.e., after the initial 16-week feedingperiod) were 0.30% body weight/day for fish fedthe basal diet, without dietary vitamin E supplemen-tation, and 0.32 and 0.33% body weight/day for fishfed the 100 and 450 mg vitamin E-supplementeddiets, respectively. No statistically significant differ-ences in growth rates were observed between fishgroups. Alpha-tocopherol concentrations (mg/kgdry weight) of 12.6, 58.2 and 310.4 mg wererecovered from the three diets (0, 100 and450 mg/kg), respectively, by HPLC analysis.
During the experimental period, water qualityparameters showed the following value ranges:temperature, 28.2–31.4 8C; pH, 6.9–7.6; dissolvedoxygen, 3.1–5.6 mg/litre; electric conductivity72.1–80.8 mS/cm; ammonia 60–123.4 mg/litre;nitrite, 18.7–23.5 mg/litre.
Glass coverslips were removed and examined 2, 7and 15 days after subcutaneous implantation into
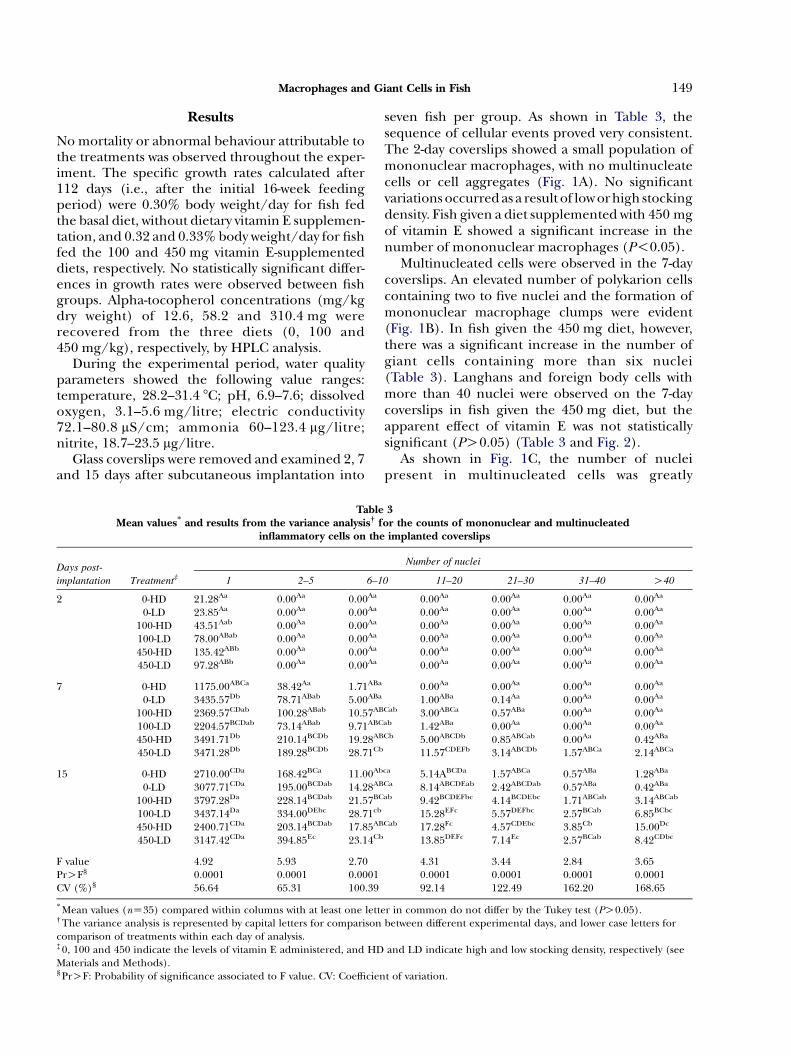
TableMean values* and results from the variance analysis† f
inflammatory cells on the
Days post-
implantation Treatment‡ 1 2–5 6–1
2 0-HD 21.28Aa 0.00Aa 0.00Aa
0-LD 23.85Aa 0.00Aa 0.00Aa
100-HD 43.51Aab 0.00Aa 0.00Aa
100-LD 78.00ABab 0.00Aa 0.00Aa
450-HD 135.42ABb 0.00Aa 0.00Aa
450-LD 97.28ABb 0.00Aa 0.00Aa
7 0-HD 1175.00ABCa 38.42Aa 1.71ABa
0-LD 3435.57Db 78.71ABab 5.00ABa
100-HD 2369.57CDab 100.28ABab 10.57AB
100-LD 2204.57BCDab 73.14ABab 9.71ABC
450-HD 3491.71Db 210.14BCDb 19.28AB
450-LD 3471.28Db 189.28BCDb 28.71Cb
15 0-HD 2710.00CDa 168.42BCa 11.00Ab
0-LD 3077.71CDa 195.00BCDab 14.28AB
100-HD 3797.28Da 228.14BCDab 21.57BC
100-LD 3437.14Da 334.00DEbc 28.71cb
450-HD 2400.71CDa 203.14BCDab 17.85AB
450-LD 3147.42CDa 394.85Ec 23.14Cb
F value 4.92 5.93 2.70PrOF§ 0.0001 0.0001 0.0001CV (%)§ 56.64 65.31 100.39
* Mean values (nZ35) compared within columns with at least one lette† The variance analysis is represented by capital letters for comparisoncomparison of treatments within each day of analysis.‡ 0, 100 and 450 indicate the levels of vitamin E administered, and HDMaterials and Methods).§ PrOF: Probability of significance associated to F value. CV: Coefficien
seven fish per group. As shown in Table 3, thesequence of cellular events proved very consistent.The 2-day coverslips showed a small population ofmononuclear macrophages, with no multinucleatecells or cell aggregates (Fig. 1A). No significantvariations occurred as a result of low orhigh stockingdensity. Fish given a diet supplemented with 450 mgof vitamin E showed a significant increase in thenumber of mononuclear macrophages (P!0.05).
Multinucleated cells were observed in the 7-daycoverslips. An elevated number of polykarion cellscontaining two to five nuclei and the formation ofmononuclear macrophage clumps were evident(Fig. 1B). In fish given the 450 mg diet, however,there was a significant increase in the number ofgiant cells containing more than six nuclei(Table 3). Langhans and foreign body cells withmore than 40 nuclei were observed on the 7-daycoverslips in fish given the 450 mg diet, but theapparent effect of vitamin E was not statisticallysignificant (PO0.05) (Table 3 and Fig. 2).
As shown in Fig. 1C, the number of nucleipresent in multinucleated cells was greatly
3or the counts of mononuclear and multinucleatedimplanted coverslips
Number of nuclei
0 11–20 21–30 31–40 O40
0.00Aa 0.00Aa 0.00Aa 0.00Aa
0.00Aa 0.00Aa 0.00Aa 0.00Aa
0.00Aa 0.00Aa 0.00Aa 0.00Aa
0.00Aa 0.00Aa 0.00Aa 0.00Aa
0.00Aa 0.00Aa 0.00Aa 0.00Aa
0.00Aa 0.00Aa 0.00Aa 0.00Aa
0.00Aa 0.00Aa 0.00Aa 0.00Aa
1.00ABa 0.14Aa 0.00Aa 0.00Aa
Cab 3.00ABCa 0.57ABa 0.00Aa 0.00Aa
ab 1.42ABa 0.00Aa 0.00Aa 0.00Aa
Cb 5.00ABCDb 0.85ABCab 0.00Aa 0.42ABa
11.57CDEFb 3.14ABCDb 1.57ABCa 2.14ABCa
ca 5.14ABCDa 1.57ABCa 0.57ABa 1.28ABa
Ca 8.14ABCDEab 2.42ABCDab 0.57ABa 0.42ABa
ab 9.42BCDEFbc 4.14BCDEbc 1.71ABCab 3.14ABCab
15.28EFc 5.57DEFbc 2.57BCab 6.85BCbc
Cab 17.28Fc 4.57CDEbc 3.85Cb 15.00Dc
13.85DEFc 7.14Ec 2.57BCab 8.42CDbc
4.31 3.44 2.84 3.650.0001 0.0001 0.0001 0.000192.14 122.49 162.20 168.65
r in common do not differ by the Tukey test (PO0.05).between different experimental days, and lower case letters for
and LD indicate high and low stocking density, respectively (see
t of variation.

Fig. 1 A–C. The kinetics of macrophage accumulation and giantcell formation on the glass coverslips. (1A) Two days afterimplantation, a few mononuclear cells (M) are present.(1B) Seven days after implantation, small polykaryonmacrophages are present (P). (1C) Fifteen days afterimplantation, foreign body (F) and Langhans (L) giantcells (inset) are present. HE. !200.
M.A.A. Belo et al.150
increased on the 15-day coverslips. This wasfollowed by a decrease in the total number ofmononuclear macrophages, since the relative areacovered by these cell types varied according to
the age of the lesion. These results show statisticallysignificant differences (P!0.05) in the inflamma-tory response between fish given the basal andvitamin E-supplemented diets (Table 3 and Fig. 2).The incidence of Langhans and foreign body cellswas highest in fish given the 450 mg diet, but thecellular counts between fish given 100 or 450 mgdiets did not differ significantly (PO0.05) (Fig. 2).
The first and second coverslip samples (2- and 7-day) showed a significant increase in plasmacortisol due to high stocking density (Fig. 3A).However, pacus fed the 450 mg diet showed nosuch effect when the second and third samples(7- and 15-day) were compared (Fig. 3A). Cortisolvalues decreased during the experiment, the day-2and day-15 values differing significantly (P!0.05).
Fish fed the basal diet were more susceptiblethan those of the other two groups to stockingdensity, as was shown by their increased plasmacortisol concentration (Fig. 3A). By examiningthese results in relation to the number of cellsfound on coverslips (Table 3 and Fig. 3A), theinfluence of plasma cortisol concentration on celladhesion is apparent from the 7- and 15-daycoverslips from fish fed low concentrations ofvitamin E.
Plasma glucose concentrations (Fig. 3B) wereunaffected by any of the treatments to which thefish were subjected (PO0.05).
Discussion
Results obtained showed a significant increase inthe number of macrophages and the formation ofpolykaryon cells on the implanted glass cover slipsas a result of vitamin E-supplementation, these cellsbeing particularly numerous in the pacus given the450 mg diet. Some authors have studied the effectof vitamin E ingestion on the defence mechanismof fish. Thus, gilthead seabream fed a vitamin E-supplemented diet exhibited increased haemolyticactivity of complement and an increased phagocy-tic leucocyte function (Ortuno et al., 2001) andenhancement of acidophilic granulocyte activityand migration of leucocytes from the head kidneys(Mulero et al., 1998). In channel catfish (Ictaluruspunctatus) high doses of vitamin E increased thephagocytic activity and superoxide anion pro-duction of leucocytes (Wise et al., 1993a, b). Blazerand Wolke (1984) reported that specific and cell-mediated immunity and macrophage phagocytosiswere all adversely affected in rainbow trout (Salmogairdneri) fed vitamin E-depleted diets. Accordingto these authors, the effect of vitamin E onphagocytosis seems to be related to the protection
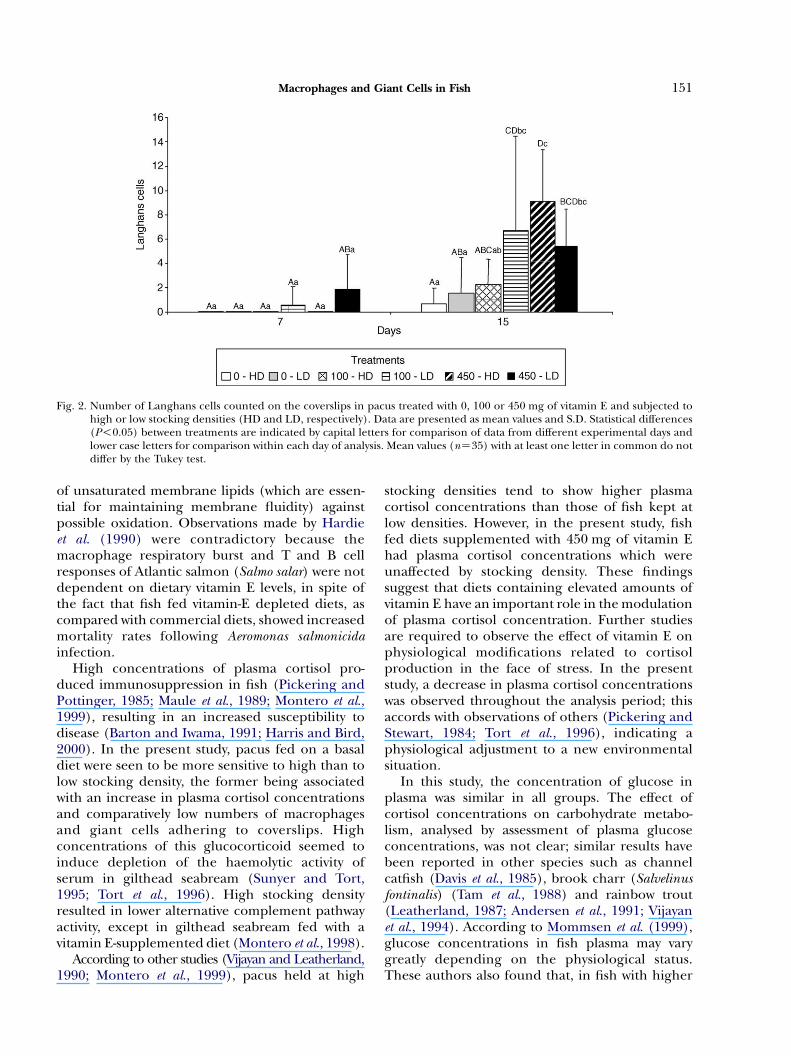
Fig. 2. Number of Langhans cells counted on the coverslips in pacus treated with 0, 100 or 450 mg of vitamin E and subjected tohigh or low stocking densities (HD and LD, respectively). Data are presented as mean values and S.D. Statistical differences(P!0.05) between treatments are indicated by capital letters for comparison of data from different experimental days andlower case letters for comparison within each day of analysis. Mean values (nZ35) with at least one letter in common do notdiffer by the Tukey test.
Macrophages and Giant Cells in Fish 151
of unsaturated membrane lipids (which are essen-tial for maintaining membrane fluidity) againstpossible oxidation. Observations made by Hardieet al. (1990) were contradictory because themacrophage respiratory burst and T and B cellresponses of Atlantic salmon (Salmo salar) were notdependent on dietary vitamin E levels, in spite ofthe fact that fish fed vitamin-E depleted diets, ascompared with commercial diets, showed increasedmortality rates following Aeromonas salmonicidainfection.
High concentrations of plasma cortisol pro-duced immunosuppression in fish (Pickering andPottinger, 1985; Maule et al., 1989; Montero et al.,1999), resulting in an increased susceptibility todisease (Barton and Iwama, 1991; Harris and Bird,2000). In the present study, pacus fed on a basaldiet were seen to be more sensitive to high than tolow stocking density, the former being associatedwith an increase in plasma cortisol concentrationsand comparatively low numbers of macrophagesand giant cells adhering to coverslips. Highconcentrations of this glucocorticoid seemed toinduce depletion of the haemolytic activity ofserum in gilthead seabream (Sunyer and Tort,1995; Tort et al., 1996). High stocking densityresulted in lower alternative complement pathwayactivity, except in gilthead seabream fed with avitamin E-supplemented diet (Montero et al., 1998).
According to other studies (Vijayan and Leatherland,1990; Montero et al., 1999), pacus held at high
stocking densities tend to show higher plasmacortisol concentrations than those of fish kept atlow densities. However, in the present study, fishfed diets supplemented with 450 mg of vitamin Ehad plasma cortisol concentrations which wereunaffected by stocking density. These findingssuggest that diets containing elevated amounts ofvitamin E have an important role in the modulationof plasma cortisol concentration. Further studiesare required to observe the effect of vitamin E onphysiological modifications related to cortisolproduction in the face of stress. In the presentstudy, a decrease in plasma cortisol concentrationswas observed throughout the analysis period; thisaccords with observations of others (Pickering andStewart, 1984; Tort et al., 1996), indicating aphysiological adjustment to a new environmentalsituation.
In this study, the concentration of glucose inplasma was similar in all groups. The effect ofcortisol concentrations on carbohydrate metabo-lism, analysed by assessment of plasma glucoseconcentrations, was not clear; similar results havebeen reported in other species such as channelcatfish (Davis et al., 1985), brook charr (Salvelinusfontinalis) (Tam et al., 1988) and rainbow trout(Leatherland, 1987; Andersen et al., 1991; Vijayanet al., 1994). According to Mommsen et al. (1999),glucose concentrations in fish plasma may varygreatly depending on the physiological status.These authors also found that, in fish with higher
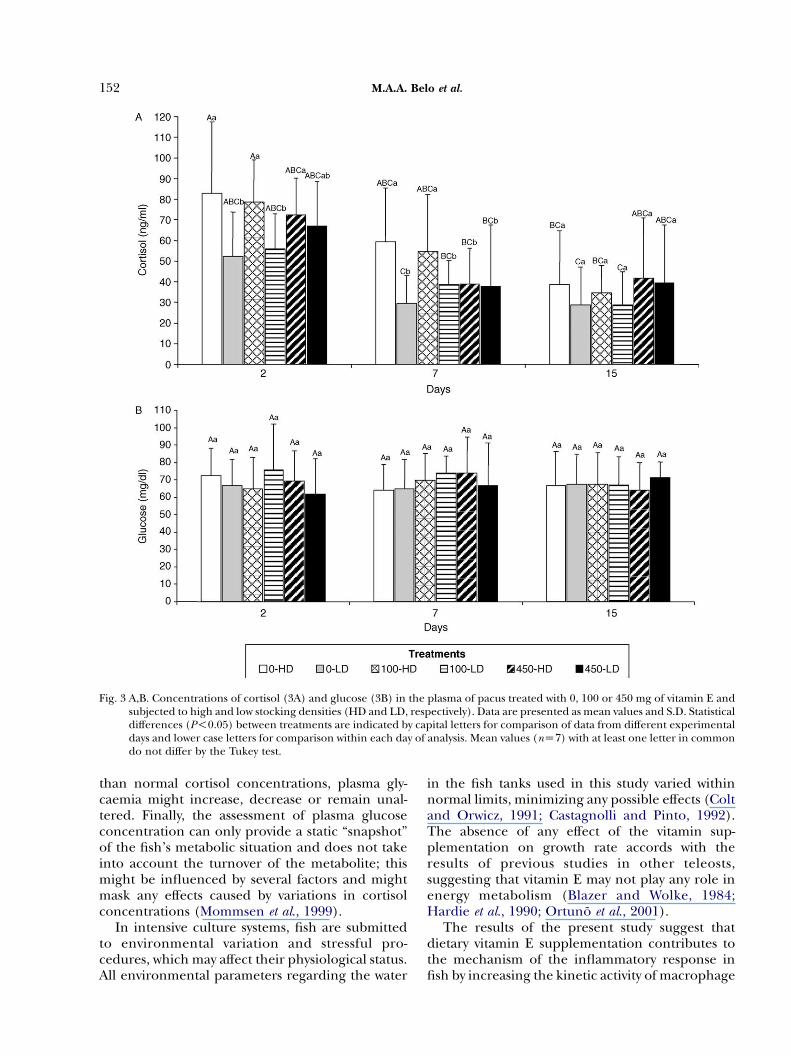
Fig. 3 A,B. Concentrations of cortisol (3A) and glucose (3B) in the plasma of pacus treated with 0, 100 or 450 mg of vitamin E andsubjected to high and low stocking densities (HD and LD, respectively). Data are presented as mean values and S.D. Statisticaldifferences (P!0.05) between treatments are indicated by capital letters for comparison of data from different experimentaldays and lower case letters for comparison within each day of analysis. Mean values (nZ7) with at least one letter in commondo not differ by the Tukey test.
M.A.A. Belo et al.152
than normal cortisol concentrations, plasma gly-caemia might increase, decrease or remain unal-tered. Finally, the assessment of plasma glucoseconcentration can only provide a static “snapshot”of the fish’s metabolic situation and does not takeinto account the turnover of the metabolite; thismight be influenced by several factors and mightmask any effects caused by variations in cortisolconcentrations (Mommsen et al., 1999).
In intensive culture systems, fish are submittedto environmental variation and stressful pro-cedures, which may affect their physiological status.All environmental parameters regarding the water
in the fish tanks used in this study varied withinnormal limits, minimizing any possible effects (Coltand Orwicz, 1991; Castagnolli and Pinto, 1992).The absence of any effect of the vitamin sup-plementation on growth rate accords with theresults of previous studies in other teleosts,suggesting that vitamin E may not play any role inenergy metabolism (Blazer and Wolke, 1984;Hardie et al., 1990; Ortuno et al., 2001).
The results of the present study suggest thatdietary vitamin E supplementation contributes tothe mechanism of the inflammatory response infish by increasing the kinetic activity of macrophage

Macrophages and Giant Cells in Fish 153
recruitment and giant cell formation in foreignbody reactions. Vitamin E would seem to act on thestress response of pacus by preventing a stress-related immunosuppression.
Acknowledgments
The authors thank the administrative directors andtechnicians of the Caunesp (Aquaculture Centre ofUnesp) for use of the aquarium facilities and fortechnical help. Thanks are due to Maria Ines Y. deCampos and Francisca de Assis Ardison forassistance in histological assessment. This researchwas financed by a Fapesp project (ResearchFoundation of Sao Paulo State, process number:00/04986-8).
References
Andersen, D. E., Reid, S. D., Moon, T. W. and Perry, S. F.(1991). Metabolic effects associated with chronicallyelevated cortisol in rainbow trout (Oncorhynchusmykiss). Canadian Journal of Fisheries and Aquatic Science,48, 1811–1817.
A.O.A.C. (Association of Official Agricultural Chemists)(1975). Official Methods of Analysis, 3rd Edit.,Washington.
Bai, S. C. and Lee, K. J. (1998). Different levels of dietaryDL-a-tocopherol acetate affect the vitamin E status ofjuvenile Korean rockfish, Sebastes schlegeli. Aquaculture,161, 405–414.
Barton, B. A. and Iwama, G. W. (1991). Physiologicalchanges in fish from stress in aquaculture withemphasis on the response and effects of corticoster-oids. Annual Review of Fish Disease, 1, 3–26.
Blazer, V. S. and Wolke, R. E. (1984). The effects of (a-tocopherol on the immune response and non-specificresistance factors of rainbow trout (Salmo gairdneriRichardson). Aquaculture, 37, 1–9.
Castagnolli, N. and Pinto, M. L. G. (1992). Piscicultura,3rd Edit, Funep Press, Jaboticabal.
Colorni, A., Avtalion, R., Knibb, W., Berger, E., Colorni,B. and Timan, B. (1998). Histopathology of sea bass(Dicentrarchus labrax) experimentally infected withMycobacterium marinum and treated with streptomycinand garlic (Allim sativum) extract. Aquaculture, 160, 1–17.
Colt, J. and Orwicz, K. (1991). Modeling productioncapacity in aquatic culture systems under freshwaterconditions. Aquacultural Engineering, 10, 1–29.
Davis, K. B., Torrance, P., Parker, N. C. and Suttle, M. A.(1985). Growth body composition and hepatictyrosine aminotransferase activity in cortisol-fedchannel catfish, Ictalurus punctatus Rafinesque. Journalof Fish Biology, 27, 177–184.
Garcia Leme, J. (1981). Regulatory mechanisms ininflammation: new aspects of autopharmacology.General Pharmacology, 12, 15–24.
Gazola, R., Bonella, M. I., Donaldson, E. M., Val-Sella,M. V., Moraes, F. F. and Bernardino, G. (1996). Plasmasteroids and corticosteroid levels in female pacuPiaractus mesopotamicus teleostei-characidae. BrazilianJournal of Medical and Biological Research, 29, 659–664.
Gillman, T. and Wright, L. J. (1966). Probable in vivoorigin of multinucleate giant cells from circulatingmononuclears. Nature, 209, 263–265.
Goodwin, A. E. and Grizzle, J. M. (1991). Granulomatousinflammation and monstrous giant cells in responseto intraperitoneal hormone implants in channelcatfish (Ictalurus punctatus). Journal of ComparativePathology, 104, 147–160.
Hardie, L. J., Fletcher, T. C. and Secombes, C. J. (1990).The effect of vitamin E on the immune response ofthe Atlantic salmon (Salmo salar L.). Aquaculture, 87,1–13.
Hardie, L. J., Fletcher, T. C. and Secombes, C. J. (1991).The effect of vitamin C on the immune response ofthe Atlantic salmon (Salmo salar L.). Aquaculture, 95,201–214.
Harris, J. and Bird, D. J. (2000). Modulation of the fishimmune system by hormones. Veterinary Immunologyand Immunopathology, 77, 163–176.
King, E. J. and Garner, R. J. (1947). Colorimetricdetermination of glucose. Journal of Clinical Pathology,1, 30–44.
Leatherland, J. F. (1987). Thyroid response to ovinethyrotropin challenge in cortisol-treated and dexa-methasone-treated rainbow-trout, Salmo gairdneri.Comparative Biochemistry and Physiology, 86B, 383–388.
Manz, U. and Philipp, K. (1981). A method for theroutine determination of tocopherols in animal feedand human foodstuffs with the aid of high perform-ance liquid chromatography. International Journal ofVitamin Nutrition Research, 51, 342–348.
Mariano, M. and Spector, W. G. (1974). The formationand properties of macrophage polykaryons (inflam-matory giant cells). Journal of Pathology, 113, 1–19.
Maule, A. G., Tripp, R. A., Kaattari, S. L. and Schreck,C. B. (1989). Stress alters the immune function anddisease resistance in Chinook salmon Oncorhynchustshawytscha. Journal of Endocrinology, 120, 135–142.
Miyazaki, T. and Kaige, N. (1985). Comparative histo-pathology of edwardsiellosis in fishes. Fish Pathology,20, 219–227.
Mommsen, T. P., Vijayan, M. M. and Moon, T. W. (1999).Cortisol in teleosts: dynamics, mechanisms of action,and metabolic regulation. Reviews in Fish Biology andFisheries, 9, 211–268.
Montero, D., Marrero, M., Izquierdo, M. S., Robaina, L.,Vergara, J. M. and Tort, L. (1999). Effect of vitamin Eand C dietary supplementation on some immuneparameters of gilthead seabream (Sparus aurata)juveniles subjected to crowding stress. Aquaculture,171, 269–278.

M.A.A. Belo et al.154
Montero, D., Tort, L., Izquierdo, M. S., Robaina, L. andVergara, J. M. (1998). Depletion of serum alternativecomplement pathway activity in gilthead seabreamcaused by a-tocopherol and n-3 HUFA dietarydeficiencies. Fish Physiology and Biochemistry, 18, 399–407.
Mulero, V., Esteban, M. A. and Meseguer, J. (1998).Effects of in vitro addition of exogenous vitamin Cand E on gilthead seabream (Sparus aurata L.)phagocytes. Veterinary Immunology and Immunopathol-ogy, 66, 185–199.
Ortuno, J., Cuesta, A., Esteban, M. A. and Meseguer, J.(2001). Effect of oral administration of high vitamin Cand E dosages on the gilthead seabream (Sparusaurata L.) innate immune system. Veterinary Immu-nology and Immunopathology, 79, 167–180.
Petric, M.C. (2000). Effect of vitamin C supplementationon giant cell formation on glass coverslips, implantedinto subcutaneous tissues of Piaractus mesopotamicus.MSc Thesis, University of Sao Paulo State, Jabotica-bal/SP, Brazil.
Pickering, A. D. and Pottinger, T. G. (1985). Cortisol canincrease the susceptibility of brown trout, Salmo truttaL., to disease without reducing the white blood cellcount. Journal of Fish Biology, 27, 611–619.
Pickering, A. D. and Pottinger, T. G. (1989). Stressresponses and disease resistance in salmonid fish:effects of chronic elevation of plasma cortisol. FishPhysiology and Biochemistry, 7, 253–258.
Pickering, A. D. and Stewart, A. (1984). Acclimation ofthe interregnal tissue of the brown trout, Salmo truttaL., to chronic crowding stress. Journal of Fish Biology,24, 731–740.
Pulsford, A. and Matthews, R. A. (1991). Macrophagesand giant cells associated with a microsporidianparasite causing liquefaction of the skeletal muscleof the Norway pout, Trisopterus esmarkii (Nilsson).Journal of Fish Diseases, 14, 67–78.
Sakai, M. (1999). Current research status of fishimmunostimulants. Aquaculture, 172, 63–92.
Snedecor, G. W. and Cochran, G. (1974). StatisticalMethods, Iowa State University Press, Ames.
Sunyer, J. O. and Tort, L. (1995). Natural hemolytic andbactericidal activities of sea bream Sparus aurataserum effected by the alternative complement path-way. Veterinary Immunology and Immunopathology, 45,333–345.
Tam, W. H., Fryer, J. N., Ali, L., Dallaire, M. R. andValentine, B. (1988). Growth inhibition, gluconeo-genesis, and morphometric studies of the pituitaryand interregnal cells of acid-stressed brook trout(Salvelinus fontinalis). Canadian Journal of Fisheries andAquatic Science, 45, 1197–1211.
Timur, G. and Timur, M. (1984). Giant-cell reactionassociated with Ichthyophonus hoferi infection in wildplaice, Pleuronectes platessa L. Journal of Fish Diseases, 7,513–514.
Tort, L., Sunyer, J. O., Gomez, E. and Molinero, A.(1996). Crowding stress induces changes in serumhaemolytic and agglutinating activity in the giltheadsea bream Sparus aurata. Veterinary Immunology andImmunopathololgy, 51, 179–188.
Vijayan, M. M. and Leatherland, J. F. (1990). Highstocking density affects cortisol secretion and tissuedistribution in brook charr, Salvelinus fontinalis.Journal of Endocrinology, 124, 311–318.
Vijayan, M. M., Pereira, C. and Moon, T. W. (1994).Hormonal stimulation of hepatocyte metabolism inrainbow trout following an acute handling stress.Comparative Biochemistry and Physiology, 108C, 321–329.
Wise, D. J., Tomasso, J. R., Gatlin, D. M., Bai, S. C. andBlazer, V. S. (1993a). Effects of dietary selenium andvitamin E on red blood cell peroxidation, glutathioneperoxidase activity, and macrophage superoxideanion production in channel catfish. Journal of AquaticAnimal Health, 5, 177–182.
Wise, D. J., Tomasso, J. R., Schewedler, T. E., Blazer, V. S.and Gatlin, D. M. (1993b). Effect of vitamin E on theimmune response of channel catfish to Edwardisiellaictaluri. Journal of Aquatic Animal Health, 5, 183–188.
Received; July 1st; 2004
Accepted ;March 2nd ; 2005
� �