early events during c1300 neuroblastoma cell interaction with syngeneic lymphocytes
TRANSCRIPT

Europ. J. Cancer Vol. 14. pp. 217-227. Pergamon Press 1978. Printed in Great Britain
Early Events During Neuroblastoma Cell
C1300 Interaction
with Syngeneic Lymphocytes
M. FIORANI, R. BUTLER, L. BERTOLINI and R. REVOLTELLA
Laborato O c.j Cell Biol~oj., C.N.R., v. Romagr, osi 18A, Rome, Italy
Abstract--Roset te formation leading to specific target cell lysis has been shown between spleen lymphocytes and syngeneic cloned C1300 neuroblastoma cells. These two phases of cell interaction could be separated by low temperature, E D T A or cytochalasin-B, which permitted the formation of rosettes but inhibited lysis. Signifcant differences were also seen between the various clones of neuroblastoma tested. Additon of ED TA or cytochalasin-B shortly after rosetteJormation prevented target cell lysis, but addition later than 5-10 min from rosette formation was ineffective, indicating a triggering of lysis and/or critical damage shortly after cell adherence.
INTRODUCTION
EVIDENCE has been recently provided that auto- genous immunity to C 1300 neuroblastoma (NB). cells exists in syngeneic A/J mice [ 1 ]. Spleen cells of unsensitized mice or mice with a growing NB tumor recognize antigenic differences on the membrane surface ofsyngeneic NB cells in a one way mixed lymphocyte-tumor blastogenic assay system [2] or by chromium release from pre- labeled NB target cells [3]. Morphologically, the first event of such cell recognition reactions consisted of adherence of lymphocytes to NB cells [4]. The purpose of this study was the characterization and analysis of the early phases. which follow first cell contact and lead to target cell lysis.
Cell adherence was documented by an in vitro rosette formation between attacking lym- phocytes and NB cells. After binding lym- phocytes were observed for a potential cytotoxic effect on target cells. This was measured by dye- exclusion or chromium release assays.
MATERJAL AND METHODS
1. Reagents The following' drugs were used: cytochalasin-
B (I.C.I. Research Lab., Cheshire, England),
Accepted 1 July 1977. For correspondence: Roberto Revohella, M.D., Ph.D., I.aboratory ot Cell Biology, C.N.R., Via Romagnosi 18A, Rome 00196, Italy.
colchicine, (Sigma Chemical Co., St. Louis, Mo), vinbtastine (Velbe, Eli Lilly and Co., Indianapolis, Ind.). All solutions of drugs were freshly prepared and used immediately. Ethylen-di-amino-tetracetic acid (EDTA) was from Sigma.
217
2. C1300 neuroblastoma ( N B ) cells Cloned neuroblastoma cells (Clone NB1R,
NB6R, NB9R and NB10R) were isolated in our laboratory from a C1300 NB line o}'iginally supplied 5yr ago by Dr. F. Jacob (Institut Pasteur, Paris). These cells are maintained in suspension culture as anaplastic cells in 20% Fetal Bovine Serum (FBS) in Dulbecco's mo- dified Eagle's medium in 12% CO2 and air atmosphere. When the cells are allowed to adhere to tissue culture vessels in the absence or in low concentration of FBS, they transform, rapidly emit cytoplasmic processes and acquire several biochemical and morphological charac- teristics of mature neurons [5]. Evidence has been recently provided that different clones derived from the same C 1300 NB cell line share a common set of tumor-associated antigenic de- terminants which are mainly responsible for tumor cell recognition by syngeneic lymphocytes in vitro. A different quantitative expression of such antigenic determinants was seen, however, among the clones and in respect to the original line. Clone NB6R appeared to be the most stimulatory for syngeneic spleen cells and exhi-

218 M. Fiorani, R. Butler, L. Bertolini and R. Revoltella
bited high expression of such common NB- associated antigen(s) [2].
3. Mice Normal 2 month male A/J mice (The Jackson
Laboratories, Bar Harbor, Maine) were in- oculated sub-cutaneously with 2 x 106 NB6R per mouse and spleens were dissected 7 days thereafter, at a time when the growing tumor mass at the site of cell injection was 2 x 2 mm in size.
4. Lymphocyte suspension Lymphocytes were obtained from spleens by
the Metrizoate-Ficoll method (Lymphoprep, Nyegaard and Co, AS, Oslo) [6]. Defibrinated non adherent-spleen cell preparations were cen- triluged at room temperature (300 x O tor 30 min) and the white cells at the interface between medium and Lymphoprep consisted mainly of lymphocytes (over 90%), monocytes and few granulocytes (1-2 % ).
5. Rosette technique NB cells in suspension culture were washed
twice and resuspended before the test in serum- free Earle's balanced salt solution (EBSS) to a concentration of 105 viable cells m l - 1 . Five hundred microliters of spleen lymphoid cells were mixed with 0.1 ml ofneuroblastoma cells at various cell ratios in round-bottomed plastic test tubes, centrituged (300 0) tot 5 rain at room temperature and gently resuspended. The cells were examined for rosette formation in a haemocytometer and Giemsa stained cytocen- trifuge preparations, using a Shandon Elliott Cytospin cytocentrifuge. In some time course kinetic studies of rosette formation, cells in pellet were resuspended in their own medium and immediately fixed in equal vol of 0.1% parafor- maldehyde in the same diluent to "freeze" the reaction at each desired time. NB cells with four or more lymphocytes attached were counted as rosettes. Rosettes were kepl at desired tempera- ture and examined for cell viability. Aldehyde (2.0% paraformaldehyde, 2.0% glutaral- dehyde, v/v 1:1) fixed rosettes were also pre- pared for electron microscopy [7].
6. Assay for cell cytotoxicity Labeled neuroblastoma cells [8] (5x 104
51Cr) were pipetted into round-bottom glass tubes (100 x 7.5). Varying concentrations of spleen lymphocytes were added at a lymphocyte to target (L/T) ratios ranging from 2: 1 to 200: 1 in a final vol of 0.5ml RPMI-1640 medium containing 2% heat decomplemented and fil- tered FBS. The cells were centrifuged (300 g) for
5 min. The pellets were then gently resuspended before incubation at ° " o 37 C m 5 / o C O 2 i n 9 5 % air. After 1 hr the tubes were centrifuged and 0.5 ml of medium were aspirated for gamma counting. Per cent specific cytotoxicity was expressed as:
s 1Cr release in exper imenta l - 5XCr release in controls (NB cells alone) maximum 51Cr release (+5°//o S D S ) -
51Cr release in controls (NB cells alone) x 100
Target cell killing in vitro was also demonstrated by the erythrosin-B method after resuspending the cells in their own medium.
7. Inhibitors Tumor membrane antigens. A soluble, crude cell
extract was prepared by stirring 2-5 x 108 NB6R cells for 16 hr at 4°C in 3 M KC1, pH 7.4 Tris- buffered saline followed by dialysis and high speed centrifugation (105,000 g for 2 hr at 4°C). This cell product, named crude soluble antigen (CSA), was shown to be immunogenic when added in vitro to syngeneic spleen cells from unprimed A/J mice; it also induced a signi- ficantly increased incorporation of 3H-Th by spleen lymphoid cells from neuroblastoma bear- ing mice inoculated 7 days before with 2 x 106 viable tumor cells [2]. A second tumor cell extract was obtained by a mild papain digestion of crude membrane preparations of NB6R cells followed by Sephadex G-150 fractionation. A soluble product, named crude membrane anti- gen (CMA, tool. wt below 30,000) was recovered in peak III as elsewhere described [2]. CMA was a very effective and specific inhibitor of blastogenesis of syngeneic A/J spleen lymphoid cells against NB cells in a unidirectional mixed cell culture system. On the contrary, it proved to be poor stimulatory when challenged in vitro with lymphocytes from normal A/J mice or from mice bearing neuroblastoma tumours. Soluble cell products from spleen, liver and kidney cells of normal A/J mice were prepared in a similar manner and were used as a source of negative controls.
Antibodies. The 7S fractions from "2 rabbits injected subcutaneously 15 days previously with 2x 107 aldehyde fixed (0.07% formal- dehyde and 0.02% glutaraldehyde) NB6R cells were obtained by recycling Sephadex-G-200 and DE-52 (Whatrnan) column chromato- graphy (0.005 M ionic strength pH 7.8 phosphate buffer) and was free of active complement or 19S antibody. This globulin fraction was then absorbed on spleen, liver and kidney of normal
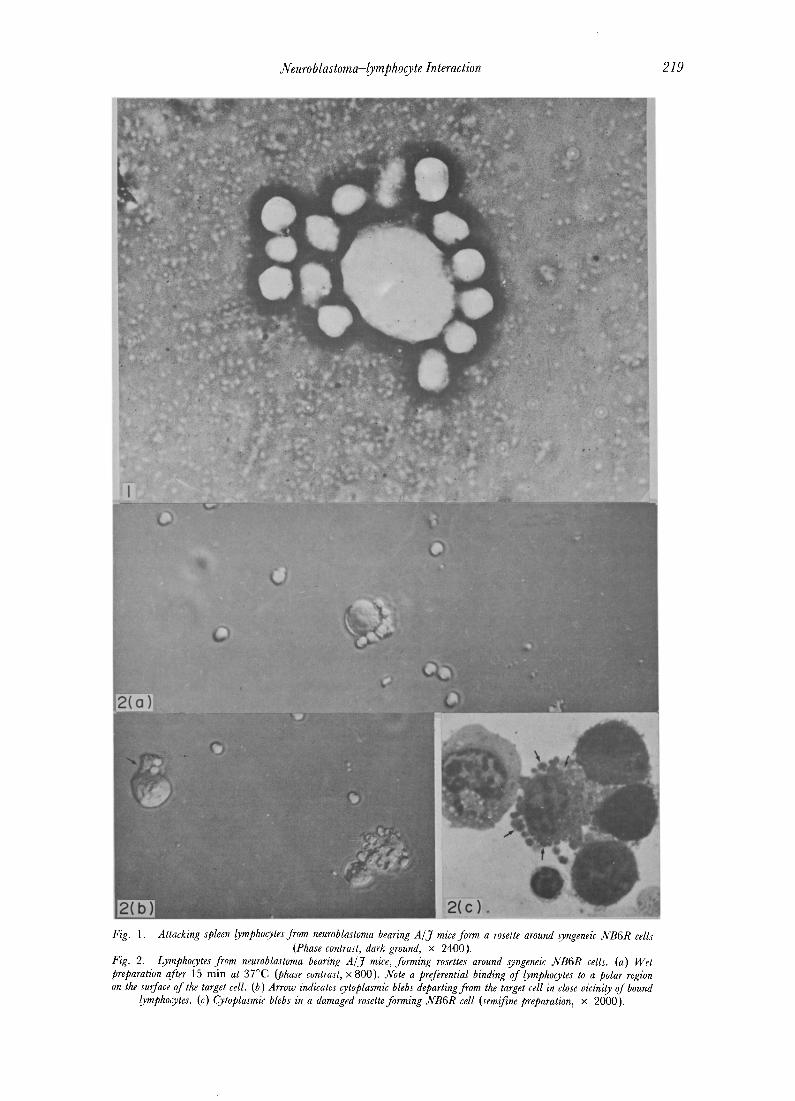
Neuroblastoma-lymphocyte Interaction 219
Fig. 1. Attacking spleen lymphocytes from neuroblastoma bearing A / j mice form a rosette around syngeneie NB6R cells (Phase contrast, dark ground, x 2400).
Fig. 2. Lymphocytes from neuroblastoma bearing A/J mice, forming rosettes around syngeneic NB6R cells. (a) Wet preparation alter 15 min at 37°C (phase contrast, x 800). Note a preferential binding of lymphocytes to a polar region on the surface of the target cell. (b ) Arrow indicates cytoplasmic blebs departing from the target cell in close vicinity of bound
lymphocytes. (c) Cytoplasmic blebs in a damaged rosette forming NB6R cell (semi~ne preparation, x 2000).

Neuroblastoma-lymphocyte Interaction 221
A/J mice. In the presence of guinea pig comple- ment (1:10) this 7S fraction was effective in killing in vitro 10()% NB6R cells at 1:30 dilution after 60 min at 37°C. In the absence of added complement no cell killing was obtained. The 7S immunoglobulin fraction from a rabbit hyper- immune serum against human serum albumin absorbed on normal A/J cells was used as a source of negative control. Fab preparations of 7S immunoglobulins were obtained by papain digestion [9]. Anti-theta serum was prepared by giving C3H mice: a course of seven injections of A/J thymus cells, at weekly intervals. All the sera were pooled and absorbed on normal liver and kidney A/J cells. The cytotoxic effect of anti- theta serum was tested on A/J mouse spleen cells by a chromium release assay or the erythrosin-B dye method: more than 70% of normal spleen lymphocytes were killed by this antiserum at a dilution of 1:10 in the presence of complement. Sepharose 4B beads (Bio-Rad, Richmond. Cal.) were covalently coated with rabbit anti mouse gamma globulin 7S immunoglobulins (MGG) as described by Wigzell et al. [10]; by sensitive radioimmunoassay, using 125 I-labeled 7S-MGG [11], it was calculated that l gg of beads (dry weight) bound at saturation about 10 v molecules ot 125I_MGG.
RESULTS
1. Rosette technique Firm adherence of syngeneic spleen lym-
phocytes to C1300 neuroblastoma cells in sus- pension was demonstrated by the rosette tech- nique. Lymphocytes bound to NB target cells at sites distributed at random on their surface (Fig. 1 ). When, however, the cells in the pellet were left to incubate for longer than 5-10 min at 24°C or higher temperature, there was a tendency for lymphocytes (usually less than 10 per target cell) to bind at a restricted polar area on the target cell and blebs from the cytoplasm of NB cells rapidly formed firstly in the proximity of the areas of contact of adhering lymphocytes and then over the whole cell surface (Fig. 2).
Increasing numbers of dead NB cells were then counted; dead cells did not form rosettes. When different NB clones were compared for rosette-forming ,capacity with lymphocytes, all clones were capable of forming rosettes. However, significant differences were shown among the clones. For example, cells of clone NB10R, even when they were incubated with a large number ot lymphocytes, lormed tew rosettes (but many cells with 1-3 lymphocytes attached). In contrast, the majority of cells from clone NB1R and NB6R formed rosettes with
usually 10-20 lymphocytes bound per cell, even if they were exposed to few lymphocytes in the test (Fig. 3). Using T-lymphocytes enriched spleen cell population (after filtration through Sepharose-beads covalently coated with anti- mouse immunoglobulins antibody) almost 100 °/o NB6R cells formed rosettes with 10-20 lym- phocytes firmly bound per cell (lymphocyte to target cell ratio 3'2" 1 ). Under the same experi- mental conditions, using a B-lymphocyte en- riched cell population (after treatment with anti-theta antibody and 1:10 fresh guinea pig serum as a source of complement, at 37°C for 60 min) few (5-6%) rosettes were obtained with the majority of NB6R cells binding a maximum of 1- 2 lymphocytes per cell. Preincubation of NB cells with anti-NB6R Fab-antibody consistently in- hibited rosette formation. Cell adherence was prevented by pre-exposing A/J lymphocytes to soluble NB cell extracts (CSA) or to a solubilized crude membrane antigen (CMA) from NB6R cells before exposure to NB cells. If similarly made preparations from normal A/J mouse cells or cell membrane were tested, no inhibition of rosette formation was obtained. These results are reported in Table 1.
2. Target cell lysis After binding, the lymphocytes were observed
to kill rosette forming NB target cells in vitro in the absence of added serum complement, as demonstrated by the erythrosin-B dye method. The kinetic of target cell lysis in experiment performed at 37°C followed an S-shaped curve. Using NB6R cells, an initial lag phase of less than 10 min was seen until 50% cell lysis took place within 15 min. In the chromium release assay, the cytotoxic effect of lymphocytes on these target cells was detected after an initial lag time of less than 15 min at room temperature (less than 5 min at 37°C), increasing rapidly and reaching 50% cell lysis in less than 20 min at room temperature (in about 15 min at 37°C). Using 30 min of incubation at room temperature as a standard procedure, it was shown that maximal target cell l'ysis was observed at an optimal lymphocyte to target cell ratio 16-32:1. The results of a typical experiment are reported in Fig. 4. When in these experimental conditions different NB clones were compared as target cells in the chromium release assay, following rosette formation with syngeneic normal lymphocytes, significantly different ratios of lymphocytes were required to induce a maximum release of chromium by a fixed number ofpre-labeled cells, with the different clones. As a general obser- vation, clones which more easily formed rosettes
2 2 2 M . Fiorani, R . Butler, L. Bertolini and R. Revoltella
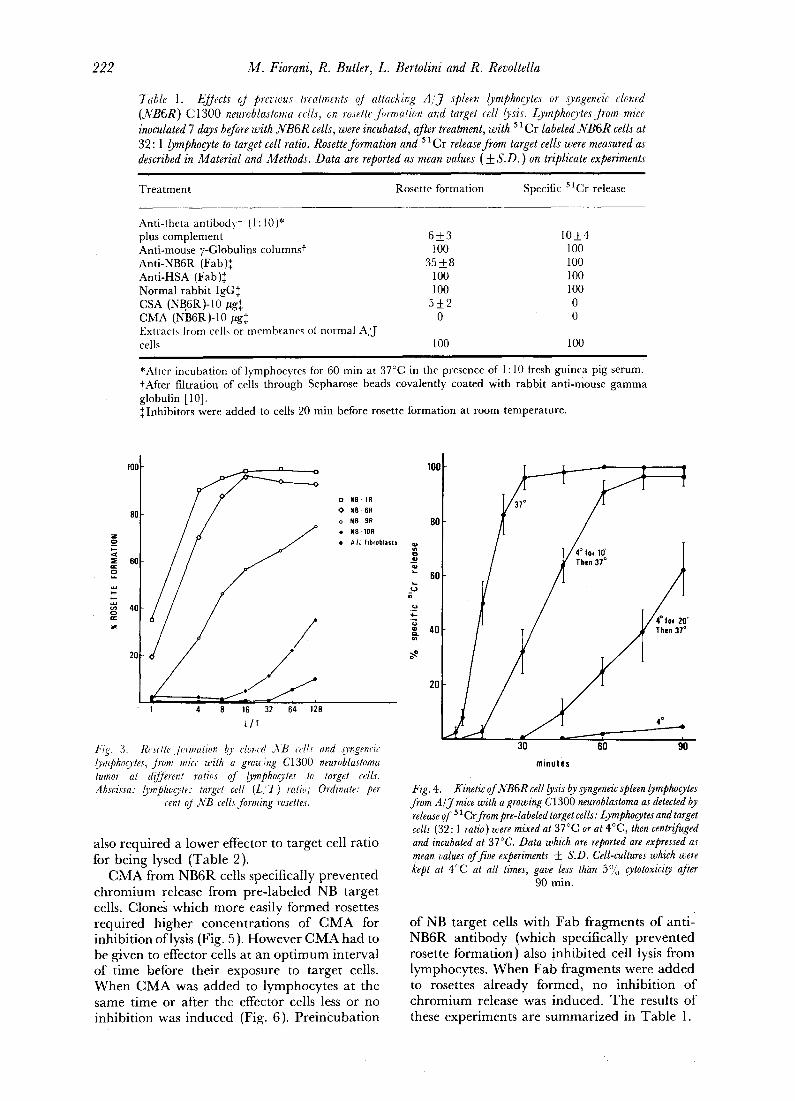
7able l. EJJects oJ pr~cious treatments oJ attacking A / j spleen lymphoo, tes or syt;geneic clc, t;ed (NB6R) C I300 neuroblastoma cells, on roselteJ~rmation and targel cell/,_)'sis. Lymphoo,tesJrom mice inoculated 7 days before with N B 6 R cells, were incubated, after treatment, with 51Cr labeled N B 6 R cells at 32:1 lymphocyte to target cell ratio. Rosette formation and 51Cr release from target cells were measured as described in Material and Methods. Data are reported as mean values (+_ S.D. ) on triplicate experiments
Treatment Rosette formation Specific 51Cr release
Anti-theta antibodyr (1: t0)* plus complement 6 + 3 Anti-mouse y-Globulins columns~ 100 Anti-NB6R (Fab)+ + 35 __.8 Anti-HSA (Fab)~ 100 Normal rabbit IgG+ + 100 CSA (NB6R)-10 #g++ 5 + 2 CMA (NB6R)-10 #g++ 0 Extracts trom cells or membranes ol normal A/J cells 100
10_+4 100 100 100 100 0 0
100
*Alter incubation of lymphocytes t0r 60 rain at 37°C in the presence ot 1 : 10 ti~esh guinea Pig serum. ~After filtration of cells through Sepharose beads covalently coated with rabbit anti-mouse gamma globulin [10]. +Inhibitors were added to cells 20 min before rosette formation at room temperature.
/ 1001 ' sa -o 100
/ D NB IR
80 0 N6- 6R o NB 9R 80 • NB -1OR
i • A]J ftbrob~/sts ~ 60' 60
40 .o
~ 40
20
20
- 4 8 16 64 128 L/T
4 ° fer 10' Then 37 °
Then 37 °
Fig. 3. Re~tte jormation b)" clvm'd ~]3 cdl,~ and s,),ngemic lymphocyes, jrom mice with a grcu'ing C1300 neuroblastoma tumor at diJfferent ratios of lymphocytes to target celL~. Abscissa: lymphocyte: target cell (L/T) ratio; Ordinate: per
cent oJ NB cells forming rosettes.
also required a lower effector to target cell ra t io for being lysed (Table 2).
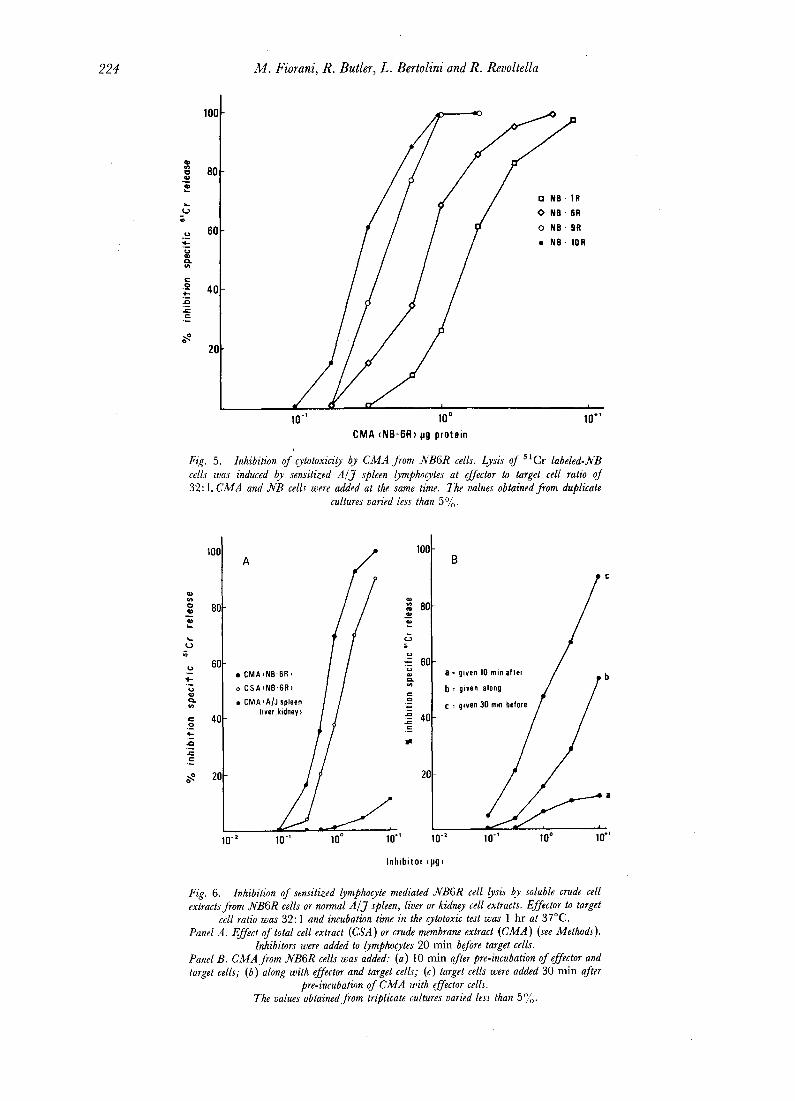
C M A from NB6R cells specifically p reven ted c h r o m i u m release f rom pre- labeled NB target cells. Clone; which more easily formed rosettes required higher concentra t ions of C M A for inhibi t ion oflysis (Fig. 5). H o w e v e r C M A had to be given to effector cells at an o p t i m u m interval of t ime before their exposure to target cells. W h e n C M A was added to lymphocytes at the same t ime or after the effector cells less or no inhibit ion was induced (Fig. 6). P re incuba t ion
30
minutes
60 90
Fig. 4. Kinetic of NB6R cell lysis by syngeneic spleen lymphocytes from A/J mice with a growing C1300 neuroblastoma as detected by release of 51Crfrom pre-labeled target cells: Lymphocytes and target cells (32:1 ratio) were mixed at 37°C or at 4°C, then centrifuged and incubated at 37°C. Data which are reported are expressed as mean values of five experiments ± S.D. Cell-cultures which were kept at 4°C at all times, gave less than 5% cytotoxicity aJter
90 rain.
of NB target cells with Fab f ragments of ant i - NB6R an t ibody (which specifically p reven ted rosette fo rmat ion) also inhibi ted cell lysis f rom lymphocytes . W h e n F a b f ragments were added to rosettes a l ready formed, no inhibi t ion of c h r o m i u m release was induced. T h e results of these exper iments are summar i zed in Tab l e 1.

Neuroblastoma-lymphocyte Interaction 223
Table 2. Effect of different lymphocyte to target cell (L/T) ratio i/ 51Cr release by pre-labeled NB6R cells Jorming rosettes with sensitized syngeneic spleen lymphocytes. Rosette formation and s 1Cr release were measured at 37 ° C
as described in Material and Methods
Cells Specific SlCr release (%)
(L/T ratio) 2:1 4:1 8:1 16:1 32:1 64:1 128:1 200:1
N B N B 1 R N B 6 R N B 7 R N B 9 R N B 1 0 R
10 21 25 25 36 52 70 70 12 25 41 65 80 92 85 90 20 38 56 86 92 95 92 96 5 15 12 31 35 45 65 62
- - 15 15 35 25 35 45 NT~ - - - - 10 15 25 40 45 N T t
J'NT: not tested.
3. Dissociation of tumor-lymphocyte adherence and cytotoxic phases
(a) Effect of temperature. Temperature signi- ficantly affected NB cell cytotoxicity by attack- ing lymphocytes. Lysis of target cells did not occur at 4°C but rapidly occurred at higher temperature reaching a maximum at 37-40°C (Fig. 7). When cells were pre-cooled for 10 rain at 4°C, spun to a cell pellet (350 0) 5 min at 4°C), resuspended and then transferred into a water bath at 37°C, the initial lag phase was prolonged as compared to cells always maintained at high temperature and the maximal rate of lysis took place within 2-3 hr (Fig. 4). Cells pre-cooled for 20-30 min before centrifugation, formed rosettes which could be easily detached simply by pipet- ting the cell suspension (2-3 times). Dead NB6R cells were shown only after 1-2 hr after the temperature was raised to 37°C.
(b) Effects of E D T A . The adhesion of lym- phocytes to rosette-forming NB cells could be reversed with EDTA (0.01 M) (Fig. 8). EDTA added shortly after cell centrifugation (tess than 5 min at room temperature) separated lym- phocytes from NB6R cells and target cells re- leased significantly less chromium than did EDTA-untreated controls. If EDTA was given after l0 min frora rosette formation, detachment of NB6R cells ti'om lymphocytes was still ob- tained but target cell lysis was not prevented. At room temperature, separation of cells forming a rosette by EDTA took about 5-10 rain to be completed: at 37°C the reaction was signi- ficantly shorter t12-3 rain). In absence of EDTA, mechanical shaking or pipetting and/or low temperature were usually ineffective in separat- ing the attached cells.
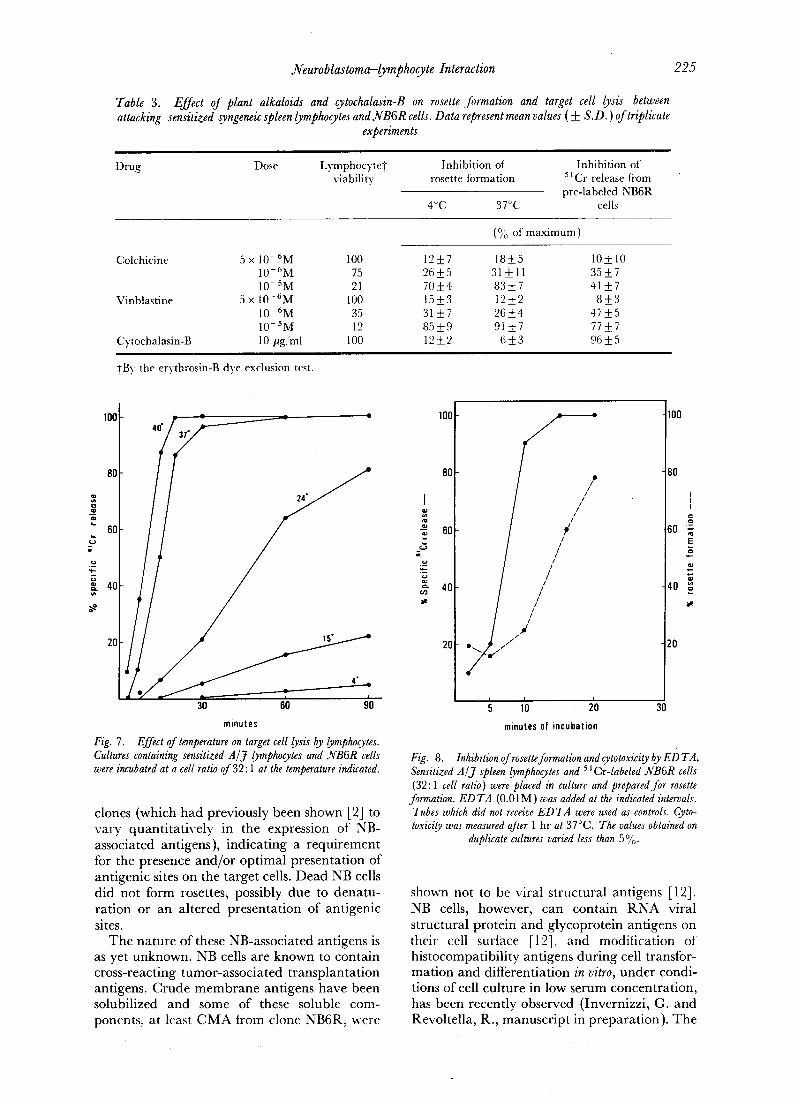
(c) Effect of plant alkaloids" and cytochalasin-B. Colchicine and vinblastine, two drugs which
induce irreversible disaggregation of microtu- bular proteins, had no effect in preventing rosette formation and chromium release from rosette-forming NB cells at any concentration tested which permitted cell viability (Table 3). Cytochalasin-B, a drug which has been shown to affect a variety of cellular functions involving microfilament protein organization, did not af- fect rosette formation but strongly inhibited NB cell lysis by attacking lymphocytes (Table 3). In the presence of cytochalasin-B, unifbrm binding by lymphocytes onto the target cell surface was seen, even at 37°C, without tendency of lymphocytes to gather on a polar region of the target cell.
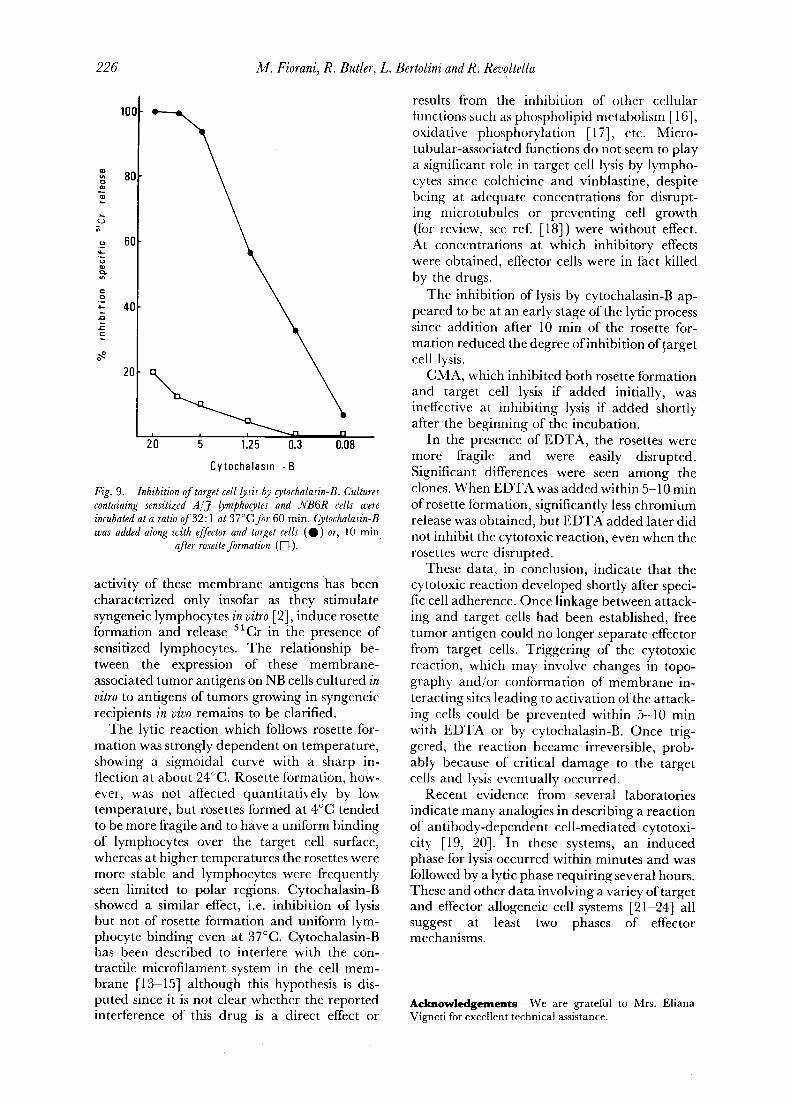
The effect of the drug was dose dependent with an optimum at 10 #g/ml (Fig. 9). The effect was reversible since washing and replacing of medium immediately induced target cell lysis. Cytochalasin-B inhibited cell lysis if given before or immediately after (less than 5 rain) cell adhesion. If given 10 min after rosette formation cytochalasin-B did not prevent target cell lysis (Fig. 9).
D I S C U S S I O N
Our findings clearly indicate that spleen lym- phocytes from A/J mice with a growing C 1300 neuroblastoma tumor bind to and induce lysis of syngeneic C1300 NB cells in vitro. The effector cells have been identified mainly as T (thymus derived) lymphocytes and the specificity of the reaction was shown by the ability of Fab fi~ag- ments of anti-NB antibodies and of solubilized NB membrane antigens to inhibit the rosette formation and cytotoxic tests. Significant differ- ences in their ability to form rosettes and in the effector/target cell ratios necessary for optimal lysis were observed among the different clones of the same NB line. The quantity of rosette formation also varied significantly among the
224 M. Fiorani, R. Butler, L. Bertolini and R. Revoltella
100
m
.o
,,a
80
6l]
4C
2(
0 N B ' I R 0 N6" 6R
o NB- 9R • NB- 1OR
f T~ Dr I i
10" I0 ° 10"
CMA (NB-6R) pg protein
Fig. 5. Inhibition of cytotoxicity by CMA Jrom NB6R cells. Lysis of 51Cr labeled-NB cells was induced by sensitized A / J spleen lymphocytes at effector to target cell ratio of 32: 1. CMA and NB cells were added at the same time. The values obtained from duplicate
cultures varied less than 50/0 .
==
C,
'4-
OZ. tn
o = -
4- .4= ,,¢: C
o<
4(
• CMA~NB-6R)
o CSA(NB-6R)
• CMAcA/J spleen liver kidney>
100
8C ~z
oa
6C
E g
.~_
10 "2 10" 10 ° 10" 10"
Inhibitor (pg)
C
a= given 10 min after / p b
b : given along / / C = given 30 min before
a
10" 100 10"
Fig. 6. Inhibition of sensitized lymphocyte mediated NB6R cell lysis by soluble crude cell extracts from NB6R cells or normal A / J spleen, liver or kidney cell extracts. Effector to target
cell ratio was 32:1 and incubation time in the cytotoxic test was 1 hr at 37°C. Panel A. Effect of total cell extract (CSA ) or crude membrane extract (CMA ) (see Methods).
Inhibitors were added to lymphocytes 20 rain before target cells. Panel B. CMA from NB6R cells was added: (a) 10 rain after pre-incubation of effector and target cells; (b ) along with effector and target cells; (c) target cells were added 30 min after
pre-incubation of C~IA with effector cells. The values obtained from triplicate cultures varied less than 5%.
Neuroblastoma-lymphocyte Interaction 2 2 5
Table 3. Effect oJ plant alkaloids and cytochalasin-B on rosette formation and target cell lysis between attacking sensitized syngeneic spleen lymphocytes and N B 6 R cells. Data represent mean values (+_ S.D. ) of triplicate
experiments
Drug Dose LymphocyteT Inhibition of Inhibition of viability rosette formation 51Cr release fiom
pre-labeled NB6R 4°(21 3T~C cells
(% of maximum)
Colchicine 5 x 10- 6M 100 12_7 18+5 10-+ 10 10-6M 75 26 _+5 31 _+ l l 35 -+ 7 10-SM 21 70 +4 83_+7 41_7
Vinblastine 5 x 10- 6 M 100 15 _ 3 12 -+ 2 8 -+_ 3 10-6M 35 31 -+7 26_+4 47+5 10-SM 12 85_+9 91 _+7 77_+7
• Cytochalasin-B 10/~g/ml 100 12 _+ 2 6 _+ 3 96 _ 5
+By the erythrosin-B dye exclusion test.
'°°I80 //:'-------- 24"
e " 60 [
g 4O Q. us
2 0 ; / / . , ~ 4"
30 60 90 minutes
Fig. 7. Effect of temperature on target cell lysis by lymphocytes. Cultures containing sensitized A /J lymphocytes and NB6R cells were incubated at a ceE ratio of 32:1 at the temperature indicated.
clones (which had previously been shown [2] to vary quant i ta t ive ly in the expression of NB- associated antigens), indicat ing a r equ i remen t for the presence and /o r op t imal presenta t ion of ant igenic sites on the target cells. Dead NB cells did not form rosettes, possibly due to dena tu - ra t ion or an al tered presenta t ion of ant igenic sites.
T h e na ture of these NB-associated antigens is as yet unknown. NB cells are known to contain cross-reacting tumor-associa ted t ransp lan ta t ion antigens. Crude m e m b r a n e antigens have been
' solubilized and some of these soluble com- ponents, at least C M A from clone NB6R, were
I
oa
ca. ctJ #
100
80
60
40
2G
A -'2
// ///
~////
illlllllllllll/)lillll IIIIIIIIII ~i 110 20 30
minutes of incubation
100
80
I
== 60 =
E
40
20
Fig. 8. Inhibition of rosette formation and cytotoxicity by ED TA. Sensitized A/J spleen lymphocytes and 51Cr-labeled NB6R cells (32:1 cell ratio) were placed in culture and prepared for rosette
formation. ED TA (0.01M) was added at the indicated intervals. Tubes which did not receive ED TA were used as controls. Cyto- toxicity was measured after 1 hr at 37 °C. The values obtained on
duplicate cultures varied less than 5%.
shown not to be viral s t ructural antigens [12]. NB cells, however , can contain R N A viral s t ructural prote in and g lycoprote in antigens on their cell surface [12], and modi t icat ion ot h is tocompat ibi l i ty antigens dur ing cell transfor- ma t ion and differentiat ion in vitro, under condi- tions of cell cul ture in low serum concentra t ion , has been recently observed (Invernizzi , G. and Revoltel la , R., manusc r ip t in p repara t ion) . T h e
100
20
== 8O
,3
._o 60 '4.-
O .
P, 7- 40 a ; i
a ~ c
20 5 1.25 0.3 0.08
2 2 6 M . Fiorani, R. Butler, L . Bertolini and R . Revoltella
Cytochalasin - B
Fig. 9. Inhibition of target cell lysis by cytochalasin-B. Cultures containing sensitized A /J lymphocytes and NB6R cells were incubated at a ratio 0f32:1 at 37°C for 60 min. Cytochalasin-B was added along with effector and target cells ( 0 ) or, 10 min
after rosette formation ([~ ).
activity of these membrane antigens has been characterized only insofar as they stimulate syngeneic lymphocytes in vitro [2], induce rosette formation and release 51Cr in the presence of sensitized lymphocytes. The relationship be- tween the expression of these membrane- associated tumor antigens on NB cells cultured in vitro to antigens of tumors growing in syngeneic recipients in vivo remains to be clarified.
The lytic reaction which follows rosette for- mation was strongly dependent on temperature, showing a sigmoidal curve with a sharp in- tlection at about 24°C. Rosette formation, how- ever, was not affected quantitatively by low temperature, but rosettes formed at 4°C tended to be more fragile and to have a uniform binding of lymphocytes over the target cell surface, whereas at higher temperatures the rosettes were more stable and lymphocytes were frequently seen limited to polar regions. Cytochalasin-B showed a similar effect, i.e. inhibition of lysis but not of rosette formation and uniform lym- phocyte binding even at 37°C. Cytochalasin-B has been described to interfere with the con- tractile microfilament system in the cell mem- brane [13-15] although this hypothesis is dis- puted since it is not clear whether the reported interference of this drug is a direct effect or
results from the inhibition of other cellular Iunctions such as phospholipid metabolism [ 16], oxidative phosphorylation [17], etc. Micro- tubular-associated functions do not seem to play a significant role in target cell lysis by lympho- cytes since colchicine and vinblastine, despite being at adequate concentrations for disrupt- ing microtubules or preventing cell growth (for review, see ref. [18]) were without effect. At concentrations at Which inhibitory effects were obtained, effector cells were in fact killed by the drugs.
The inhibition of lysis by cytochalasin-B ap- peared to be at an early stage of the lyric process since addition after 10 min of the rosette for- mation reduced the degree of inhibition of~arget cell lysis.
CMA, which inhibited both rosette formation and target cell lysis if added initially, was inettective at inhibiting lysis if added shortly after the beginning of the incubation.
In the presence of EDTA, the rosettes were more fragile and were easily disrupted. Significant differences were seen among the clones. When EDTA was added within 5-10 min of rosette formation, significantly less chromium release was obtained, but EDTA added later did not inhibit the cytotoxic reaction, even when the rosettes were disrupted.
These data, in conclusion, indicate that the cytotoxic reaction developed shortly after speci- fic cell adherence. Once linkage between attack- ing and target cells had been established, free tumor antigen could no longer separate effector from target cells. Triggering of the cytotoxic reaction, which may involve changes in topo- graphy and/or cont0rmation of membrane in- teracting sites leading to activation of the attack- ing cells could be prevented within 5--10 min with EDTA or by cytochalasin-B. Once trig- gered, the reaction became irreversible, prob- ably because of critical damage to the target cells and lysis eventually occurred.
Recent evidence from several laboratories indicate many analogies in describing a reaction ot antibody-dependent cell-mediated cytotoxi- city [19, 20]. In these systems, an induced phase for lysis occurred within minutes and was followed by a lytic phase requiring several hours. These and other data involving a variey of target and effector allogeneic cell systems [21-24] all suggest at least two phases of effector mechanisms.
Acknowledgements--We are grateful to Mrs. Eliana Vigneti for excellent technical assistance.

Neuroblastoma-lymphocyte interaction
REFERENCES 1. M.B.A. OLDSTONE, Immune complex in cancer: demonstration of complexes in
mice bearing neuroblastomas. J. nat. Cancer Inst. 54, 223 (1975). 2. R. REVOLTELLA, L. BERTOLINI and L. DIAMOND, In vitro stimulation of mouse
lymphoid cells by C 1300 neuroblastoma cells or tumor membrane extracts. Int. J. Cancer 18, 521 (1976).
3. R. REVOLTELLA, L. BERTOLINI, L. DIAMOND and R. Bt:TLER, Serum-mediated raodulation of murine lymphoid cells immune response to syngeneic C1300 neuroblastoma cells. (Submitted for publication.)
4. G. PROCICCHIANI, M. FIORANI and R. REVOLTELLA, Ultrastructural studies of C1300 neuroblastoma cell interaction with syngeneic spleen lymphocytes. (Submitted for publication.)
5. L). SCHUBERT, S. HUMPHREYS, F. S. DE VITRY and F. JACOB, Induced diiIerentiation of a neuroblastoma. Devel@. Biol. 25, 514 (1971).
6. E. THORSBY and A. BRATTIE, A rapid method for preparation of pure lymphocyte suspensions. Histocompatibility testing. (Edited by L. P. Terasaki) p. 655. Munskgaard, Copenhagen (1970).
7. (2. GUDAT and W. S. WILLINGER, A scanning and transmission electron microscope study of antigen binding sites on rosette-forming cells J . exp. Med. 137, 4,83 (1973).
8. .]i. C. CEROTTINI and T. B. BRUNN'ER, Cell mediated cytotoxicity, allografi rejection and tumor immunity. Advan. Imrnunol. 18, 67 (1974).
9. R .R. PORTER, The hydrolysis of rabbit ~'-globulin and antibodies with crystalline papain. Biochem. J. 73~ 119 (1959).
10. H. WIGZELL, P. GOLSTEIN, E. A. SVEDMYR and M. JONDHAL, Impact of fi'actionation procedures on lymphocyte activities in vitro and in vivo. Separation of cells with high concentrations of surface immunoglobulin. Transplant. Proc. 4, 311 (1973).
11. R. REVOLTELLA, L. BERTOLINI, L. DIAMOND, E. VmNETI and A. GRASSO, A radioimmunoassay for measuring nanogram levels of 14-3-2 protein in fluids or cell extracts. J. Neurochem. 26~ 831 (1976).
12. R. REVOLTELLA and R. BUTLER, Antigens solubilized from cloned murine C 1300 neuroblastoma cells. (Submitted for publication.)
13. R. WAGNER, M. ROSENBERG and R. D. ESTENSEN, Endocytosis in Chang liver cells. J. CellBiol. 50, 804 (1971).
14. N.K. WESSELS, B. S. SPOONER, J. FASH, M. O. BRADLEY, MA. A. LUDUENA, E. L. TAYLOR, J. T. WRENN and K. M. YAMADA, Microfilaments in cellular and developmental processes. Science 171~ 135 (1971 ).
15. E.L. BECKER , A. T. DAVIS, R. DESTENSEN and P. G. RrIE, Cytochalasin-B. 1V. Inhibition and stimulation of chemotaxis of rabbit and human polymorphonuc- lear leukocytes. Immunology. 108, 396 (1972).
16. K. RESCH, E. W. GELFAND, K. HANSEN and E. FERBER, Lymphocyte activation: rapid changes in the phospholipid metabolism of plasma membranes during stimulation. Europ. 07. Immunol. 9, 598 (1972).
17. ]~. D. ESTENSEN and P. G. W. PLAGEMAN, Cytochalasin-B; inhibition of glucose and glucosamine transport. Proc. nat. Acad. Sci. (Wash.)69, 1430 (1972).
18. J.B. OLMSTED and G. G. BORISY, Microtubules. Ann. Rev. Biochem. 42, 507 (1973). 19. J. C. SCORNICK, Antibody-dependent cell mediated cytotoxicity. II. Early
interactions between effector and target cells. 07. Immunol. 113~ 1519 (1974). 20. E.W. GELFAND, S. A. MORRIS and K. RESCH, Antibody-dependent cytotoxicity:
modulation bY the cytochalasin and microtubule disrupting agents. 07. Immunol. 114, 919 (1975).
21. H. WAGNER and M. ROLLINGHOFF, T-cell mediated cytotoxicity: discrimination between antigen recognition, lethal hit and cytolysis phase. Europ. 07. Immunol. 4, 1745 (1974).
22. i~. MARTZ, Early steps in specific tumor cell lysis by sensitized mouse T- ]ymphocytes. I. Resolution and characterization. 07. Immunol. 115, 261 (1975).
23. i~. MARTZ and B. BENACERRAF, T-lymphocyte mediated cytolysis: temperature dependance of killer cell dependent and independent phases and lack of recovery ti'om the lethal hit at low temperature. Cell. Immunol. 20, 81 (1975).
24. D. ZAOURY,J. BERNARD, N. THIERNESS, M. FELDMAN and G. BERKE, Isolation and characterization of individually functionally reactive cytotoxic T-lymphocytes: conjugation, killing and recycling at the single cell level. Europ. J. Immunol. 5, 818 t'1975).
227