dysregulated cc receptor/ligand in monocytes/macrophages from tongue squamous cell carcinoma...
TRANSCRIPT

SA
MMT
tp[a
rn
(
S
Human Immunology 73 (2012) 38-47
Contents lists available at SciVerse ScienceDirect
Dysregulated CC receptor/ligand in monocytes/macrophages from tonguesquamous cell carcinoma patients is partially rectified by interferon �-2bKrishnendu Chakraborty a,,†, Anamika Bose a,,‡, Kuntal K. Goswami a, Kalyan K. Mukherjee a,b,hyamal Goswami a, Diptendu Ghosh a, Tathagata Chakraborty a, Koustav Sarkar a,§, Smarajit Pal c,nup Bhowmick d, Jaydip Biswas b,e, Rathindranath Baral a,*
a Department of Immunoregulation and Immunodiagnostics, Chittaranjan National Cancer Institute, Kolkata 700026, Indiab Department of Medical Oncology, Chittaranjan National Cancer Institute, Kolkata 700026, Indiac Unit of Clinical Biochemistry, Chittaranjan National Cancer Institute, Kolkata 700026, Indiad Department of ENT & Head–Neck Oncology, Chittaranjan National Cancer Institute, Kolkata 700026, Indiae Department of Surgical Oncology, Chittaranjan National Cancer Institute, Kolkata 700026, India
A R T I C L E I N F O
Article history:Received 4 May 2011Accepted 3 October 2011Available online 29 October 2011
Keywords:ChemokinesCCR5IFN�2bonocytesacrophagesongue squamous cell carcinoma
A B S T R A C T
In an aim to rectify dysregulated CC chemokine receptor (CCR5)/ligand (RANTES, MIP-1�, MIP-1�) status ofmonocytes/macrophages in tongue squamous cell carcinoma (TSCC; n � 12) patients, we have testedinterferon �2b (IFN�2b), a novel immunomodulator with wide use in the management of several forms ofcancer. IFN�2b can upregulate reduced CCR5 expression and increases the suppressed secretory status of itsligands, as evidenced from in vitro studies on monocytes/macrophages from the peripheral blood of TSCCpatients as well as healthy individuals. Isolated monocytes of TSCC patients (n � 5) undergoing chemother-apeutic treatment along with IFN�2b immunotherapy demonstrated significant upregulation in CCR5 ex-pression and secretion of corresponding ligands. These rectifications in receptor/ligand levels are reflected inimproved CCR5-dependent migration of monocytes/macrophages after IFN�2b treatment. The rectifiedchemokine profile and cellular migration translate into better tumoricidal and antigen-presenting functionsof these cells. Accordingly, enhanced T-cell-mediated tumor cell killing is demonstrated upon IFN�2btreatment. Translating dual benefits on monocyte/macrophage functions, IFN�2b may emerge as a potentialform of immunotherapy for TSCC patients that may be combined with standard chemotherapy for betterclinical outcome.
� 2012 American Society for Histocompatibility and Immunogenetics. Published by Elsevier Inc. All rights
reserved.1. Introduction
Head and neck squamous cell carcinoma (HNSCC), includingtongue squamous cell carcinoma (TSCC), is a serious health prob-lem worldwide [1,2] and is also increasing in India [3]. Currentherapeutic approaches with new-generation chemotherapy showromise, but overall increases in 5-year survival are insignificant4]. Alternate approaches to combat this particular form of cancerre under evaluation [5,6]. Interferon �-2b (IFN�2b) is a pleotropic
cytokine with wide applications in the treatment of several formsof cancer [7,8], including immunocompetent cancers [9,10] such asenal cell carcinoma andmelanoma. TSCC, being a cancer of immu-osuppressed type [11], is not generally considered for IFN�2b
* Corresponding author.E-mail address: [email protected]; [email protected]
R. Baral).† Current address: Louisiana State University, Baton Rouge, LA 70803, USA.‡ Current address: Department of Molecular Medicine, Bose Institute, C.I.T.
cheme, Kolkata, India.§ Current address: Children’s Hospital of UPMC, Pittsburgh, PA 15224, USA.0198-8859/$36.00 � 2012 American Society for Histocompatibility and Immunogeneticsdoi:10.1016/j.humimm.2011.10.015
immunotherapeutic protocols. However, our in vitro [12,13] stud-ies clearly suggested that this cytokine is effective to increase thetumoricidal efficacy of blood T cells, natural killer cells, andmacro-phages (M�) from HNSCC patients. Such in vitro results were ex-trapolated in a pilot in vivo study with immunosuppressed TSCCpatients and significant immunopotentiation was noted in correla-tion with clinical outcome [14].
Monocytes (MO) and M� are highly versatile, multifunctionalcells characterized by their ability to engulf invading microbes orcell debris from injured sites, secrete a wide array of immuno-modulatory cytokines, and act as accessory cells in lymphocyteactivation [15]. For T-cell-mediated killing, optimization ofMO/M�
functions is important because they present antigens to T cells forinitiation of antigen-specific tumor killing [16]. To actively assistantitumor T effector functions against tumor,MO/M� should reachthe tumor site with primary help from chemokines [17,18]. Wepreviously reported dysregulated CXC chemokine signaling on Tlymphocytes and their rectification by IFN�2b was reported [19].To ascertain the status of MO/M� in chemotactic paths, we have
selected the CC chemokine receptor CCR5 [20]. Studies have con-. Published by Elsevier Inc. All rights reserved.
orap
filn
iwi
2
2
ttmO
ffSHcf
2
filiecht
wti
2
vd
ot
2
hdT
TCc
K. Chakraborty et al. / Human Immunology 73 (2012) 38-47 39
firmed that CCR5 and its ligands (RANTES, MIP-1�, andMIP-1�) arene of the main receptors and chemoattractants involved in theecruitment of type 1 polarized MO/M� at tumor sites [15,21–23]nd positive CCR5 signaling inMO/M� may result in better antigenresentation to T cells.Significant dysregulation in CCR5 expression on MO/M�, puri-
ed from TSCC patients, has been demonstrated. Similar dysregu-ation in secretion of corresponding ligands has also been alsooted. With regard to our previous observation reporting IFN�2b-
mediated augmentation of suppressed immune functions in tobacco-related HNSCC patients [12–14], it is of interest to assess the mod-ulatory role of IFN�2b on dysregulated CCR5 chemokine signalingn TSCC MO/M� to induce successful migration of these cells to-ard the tumor for ultimate enhancement of tumor cell cytotoxic-
ty, either directly by MO/M� or by presenting antigens to T cells.
. Subjects and methods
.1. Antibodies and reagents
Recombinant human IFN�2b was gifted by the manufacturer(Shanferon, Santha-Biotech, Hyderabad, India). Fluorescence-conjugated antihuman antibodies (CCR5, RANTES, MIP-1�, andMIP-1�) were procured from BD Pharmingen (San Diego, CA). Neu-ralizing antibodies such as antihuman CCR5 and CXCR3 were ob-ained from e-Biosciences (San Diego, CA). Lymphocyte separationedium was procured from MP Biomedicals (Irvine, CA). TheptEIA kit (to measure IFN-�) and the Ki-67 kit (for cell prolifera-
tion)were procured fromBDPharmingen. Cellular cytotoxicitywasdetermined by lactate dehydrogenase (LDH) release assay using acommercially available cytotoxicity detection kit (Roche Diagnos-tics, Mannheim, Germany). Reverse transcription polymerasechain reaction (RT-PCR) primers were designed and procured fromMWG-Biotech AG (Bangalore, India).
2.2. Tumor cell lines and tumor antigen
Oral cancer (HEp2) andmacrophage-specific lymphoma (U937)cell lines originally obtained from the National Center for CellSciences (Pune, India) were maintained in minimum essential me-
Table 1Characteristics of tongue squamous cell carcinoma (TSCC) patients and healthyindividuals in in vitro studies
Sr No. Age Sex Tobacco habits Histology Disease status
Pt 1 43 F Chewer Mod T3N1M0
Pt 2 65 M Chewer Mod T4N1M0
Pt 3 52 M No habits Well T3N0M0
Pt 4 37 M Smoker and chewer Poor T4N1M0
Pt 5 68 F Chewer Poor T3N1M0
Pt 6 49 M Smoker Mod T3N1M0
Pt 7 55 F Chewer Mod T4N1M0
Pt 8 56 M Smoker and chewer Poor T4N1M0
Pt 9 34 M Chewer Well T3N1M0
Pt 10 25 M No habits Mod T2N1M0
Pt 11 62 M Smoker Mod T3N1M0
Pt 12 57 F Chewer Mod T3N0M0
H 1 38 M No habits — —H 2 48 M Smoker — —H 3 27 M Smoker and chewer — —H 4 52 M No habits — —H 5 48 F Chewer — —H 6 67 M Smoker and chewer — —H 7 53 M Smoker — —H 8 29 F No habits — —H 9 42 F Chewer — —H 10 58 M Smoker and chewer — —H 11 70 M Chewer — —H 12 53 F Chewer — —
Pt � TSCC patient; H � healthy individual; M � male; F � female; well � well
Sifferentiated; mod � moderately differentiated; poor � poorly differentiated;NM � tumor node metastasis.
dium and RPMI 1640 (Life Technologies, Grand Island, NY), respec-tively, with 10% fetal bovine serum, penicillin (50 U/mL), and strep-tomycin (50 �g/mL) at 37�C with a supply of 5% CO2.
HEp2 cells harvested form confluent culturewere subjected to 3reeze–thaw cycles, stirred in cold phosphate-buffered saline (PBS)or 2 hours, dialyzed, and then centrifuged at 12,000g for 1 hour.upernatant containing tumor-associated antigen extracted fromEp2 cells (HEp2Ag)was collected and stored at�20�C. Theproteinoncentration of the extract was determined by Bradford assayollowing the manufacturer’s instructions.
.3. TSCC patients and healthy individuals
Patients (8 males, 4 females) with histopathologically con-rmed TSCC were included and their blood specimens were col-ected before initiation of any treatment after obtaining writtennformed consent from each donor (Table 1). Healthy individuals ofither sex in the sameage groupwere also included as age-matchedontrols (8 males, 4 females). In addition, patients (n � 5) withistopathologically confirmed inoperable TSCC (tumor node me-astasis stage IV; Table 2) treatedwith cisplatin (70mg/m2 iv onday1) � 5-fluorouracil (5-FU; 500 mg/m2 iv on days 1–3) in 2 cycles ata 21-day interval were included in this study. IFN�2b (3 mIU twiceeekly for 3 weeks) was administered between 2 cycles of chemo-herapy. The treatment protocol wasmodified in some cases accord-ng to clinical demand. For example, the IFN�2bcyclewasdivided into2 because of leukopenia. All of these patients had 1 or more form oftobacco habits. Bloodwas collected at 3 time points from the patientswho received IFN�2b therapy (i.e., before treatment, after the firstchemotherapy cycle, and after completion of IFN�2b therapy). Theinstitutional ethical committee approved the study design.
2.4. Preparation and antigen pulsation of MO/M�
Venous blood was collected from study subjects in heparinizedtubes and separated on lymphocyte separationmedium at 500g for30minutes. Leukocyteswere recovered from the interface,washed,counted, and adhered to a plastic surface for 2 hours. Adherentfractions were checked for CD14, CD68 positivity and �90%CD14�CD68� MO/M¦ were used in different in vitro assays. Forantigen pulsation, MO/M¦ from 3 TSCC patients (with or withoutIFN�2b priming for 24 hours) were incubated with HEp2Ag (1�g/mL) for another 24 hours.
.5. Preparation of T cells
Peripheral blood mononuclear cells (PBMC) from healthy indi-iduals (n � 3) were exposed to interleukin-2 (IL-2; 10 ng/mL) andivided into 2 sets. One set was cultured with HEp2Ag (1 �g/mL),
whereas the other set remained treatment free. Both sets of culturewere continued for 7dayswith feeding of IL-2 aloneor IL-2�HEp2Agn alternate days. The status of T-cell enrichmentwas checked beforehe experiment by assessing CD3� T cells using flow cytometry.
.6. Mixed lymphocyte reactions
HEp2Ag0pulsed TSCC MO/M¦ and IL-2/HEp2Ag-generatedealthy T cellsweremixed in 1:5 ratios and cocultured for 72hours.
able 2linical information of stage IV tongue squamous cell carcinoma patients havinghemotherapy and interferon �2b therapy
Sr No. Age Sex Tobacco habit Disease status
Pt 1 53 M Chewer T4N1M0
Pt 2 55 M Smoker and chewer T4N1M0
Pt 3 51 F No habits T4N1M0
Pt 4 34 F Chewer T4N0M0
Pt 5 60 M Chewer T4N1M0
upernatants from these cultures were stored at �80�C for IFN-�

ih
r
I
K. Chakraborty et al. / Human Immunology 73 (2012) 38-4740
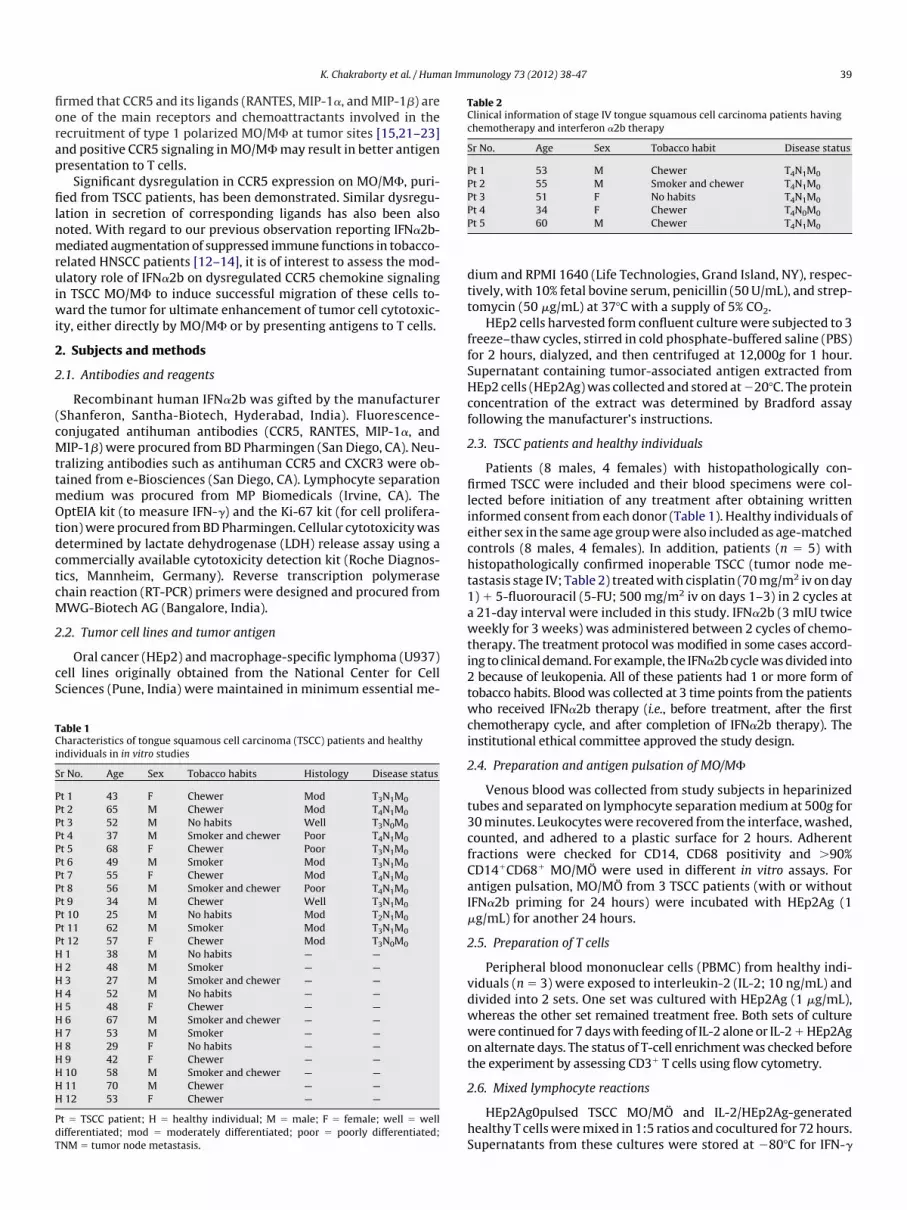
Fig. 1. Interferon (IFN)-�2b upregulates CCR5 expression in tongue squamous cell carcinoma (TSCC)monocytes/macrophages (MO/M�). Peripheral bloodmononuclear cells weresolated from the venous blood of TSCC patients (n� 12) and healthy individuals (n� 12). Adherent fractionswere taken and incubatedwith IFN�2b (0.09MIU/mL of culture) for 48ours. Cellswere surface labeledwith anti-CCR5 antibody for flow cytometric analysis. Themean� SD of the percentage positive cells is presented in the bar diagram (A) (*p� 0.05
in comparison with NT TSCC) and a representative figure from TSCC MO/M� on CCR5 expression is also presented. Values indicate the percentage positive cells, along with mean
fluorescence intensity inparentheses (B). Reverse transcriptionpolymerasechain reactionanalysis indicates that IFN�2bupregulatesmRNAexpressionofCCR5 inTSCCMO/M�withespect to GAPDH. A representative image (C) and relative expression of CCR5mRNA in relation to GAPDH (D) are presented. **p � 0.001. NT � no treatment.Fig. 2. Interferon-�2b (IFN�2b) upregulates expression of CCR5 ligands in tongue squamous cell carcinoma (TSCC)monocytes/macrophages (MO/M�). MO/M�were isolatedfrom peripheral blood mononuclear cells of TSCC patients (n � 12) and healthy individuals (n � 12) and incubated with IFN�2b (0.09 MIU/mL of culture) for 48 hours.ntracellular expression of RANTES, MIP-1�, and MIP-1� was assessed by flow cytometry. Values are presented in the bar diagrams as means � SD of the percentage positive
cells (A). A representative image in each case generated from TSCCMO/M� is presented. Values indicate the percentage positive cells, alongwithmean fluorescence intensityin parentheses (B). *p � 0.01, #p � 0.001, in comparison with TSCC NT; **p � 0.001, ***p � 0.001, in comparison with TSCC NT.
(R
K. Chakraborty et al. / Human Immunology 73 (2012) 38-47 41
analysis. Nonadherent cells from these cultures were subjected toproliferation assays (byMTT colorimetric assay andKi-67 staining).Adherent MO/M¦ were stained for CCR5 expression.
2.7. Proliferation assays
An aliquot (20 �L) of MTT solution (5 mg/mL) was added tononadherent cells from the coculture and incubated for 4 hours at37�C. The medium was removed by aspiration, the purple-coloredformazan precipitate was dissolved in dimethylsulfoxide (DMSO;100 �L), and absorbance was measured at 550 nm using a micro-plate reader (Bio-Tek,Winooski, VT). The absorbance obtained wasdirectly proportional to the number of viable cells.
Another aliquot of a similar cell population was fixed in chilledethanol and proliferating cells were stained with Ki-67–FITC withappropriate isotype control according to the manufacturer’s rec-ommendation.
2.8. Flow cytometry for CCR5, CD80, and CD86 expressionon MO/M�
Expression of CCR5, CD80, and CD86 on theMO/M� surfacewasstudied by flow cytometry. Purified MO/M� before and afterIFN�2b treatment (0.09 MIU/mL of culture) were washed with
Fig. 3. Interferon-�2b (IFN�2b) upregulates CC receptor/ligand expression inmonocchemotherapy. TSCC patients (n � 5) were treated with chemotherapy followed by I
i.e., before therapy, after chemotherapy, and after chemoimmunotherapy). Flow cytomANTES, MIP-1�, and MIP-1� (B), was assessed.FACS buffer and labeled with 20 �L of fluorescence-labeled antihu-man antibodies for 30 minutes at 4�C in the dark. After beinglabeled, cells were washed in FACS buffer and fixed in 1% parafor-maldehyde in PBS and cytometry was performed using Cell Questsoftware on a FACScan flow cytometer (Becton Dickinson, Moun-tain View, CA). Suitable negative isotype controls were used to ruleout the background fluorescence. The datawere generated by cyto-fluorometric analyses of 10,000 events. The percentage of eachpositive population and mean fluorescence intensity were deter-mined using quadrant statistics.
2.9. Intracellular secretion of chemokines
The intracellular secretion of RANTES, MIP-1�, and MIP-1� wasanalyzed by flow cytometry. In brief, purified MO/M¦were stimu-lated in vitro with IFN�2b in the presence of GolgiStop (2.0 mMmonensin; BD Pharmingen). After being fixed and permeabilizedusing cytofix/cytoperm solution (BD Pharmingen), cells werestained for intracellular secretion of RANTES, MIP-1�, and MIP-1�
using fluorescence-conjugated antibodies and incubated for 30 min-utes in thedarkat4�C.Afterbeingwashedby1Perm/Washsolution,cells were fixed in 1% paraformaldehyde solution and analyzed on aFACSCalibur using CellQuest software. Suitable negative isotype con-
acrophages (MO/M�) from tongue squamous cell carcinoma (TSCC) patients underb immunotherapy. Blood samples were collected from each patient at 3 time points
ytes/mFN�2
etric analysis for CCR5 expression (A) and intracellular secretion for its ligands,

r(tmm
2
5
qr
M
ab
K. Chakraborty et al. / Human Immunology 73 (2012) 38-4742
trols were used to rule out the background fluorescence. The datawere generated by cytofluorometric analyses of 10,000 events. Thepercentages of the positive population andmean fluorescence inten-sity were determined using quadrant statistics.
2.10. mRNA isolation and RT-PCR analysis
mRNA was isolated from TSCC MO/M¦ and cDNA was synthe-sized by RT. Amplification was performed using primer-specificpolymerization. Finally, the expression was detected by runningthe PCR product in agarose gel electrophoresis.
The oligonucleotide primers of human CCR5 and GAPDH are asfollows: CCR5-forward, 5=- CTG GCC ATC TCT GAC CTG TTT TTC -3=;CCR5-reverse, 5=- CAG CCC TGT GCC TCT TCT TCT CAT -3=; GAPDH-forward, 5=- TGTTGCCATCAATGACCCCTT -3=; GAPDH-reverse, 5=-CTC CAC GAC GTA CTC AGC G -3=. The thermal profile for the PCRreaction was as follows: 95�C for 30 seconds, 58�C for 45 seconds,and 72�C for 60 seconds.
PCR products were identified by image analysis software for geldocumentation (LabWorks Software; UVP, LLC, Upland, CA) follow-ing electrophoresis on 2% agarose gels and staining with ethidiumbromide.
2.11. Neutralization of CCR5
For neutralization of CCR5, the MO/M� preparation was incu-batedwith purified antihumanCCR5 antibody (1�g/mL culture) for30 minutes at 4�C before the experiment. After incubation, cellswerewashedwith PBS and used in a chemotaxis assay. Anti-CXCR3antibody was used in this experiment as a negative control.
2.12. Chemotaxis assay
The chemotaxis assay was performed in a Boyden chamber.Briefly, medium and culture supernatant were placed in the
Fig. 4. Suppressed CCR5-dependent chemotaxis of tongue squamous cell carcinomO/M� were isolated from venous blood of TSCC patients (n � 10) and healthy indiv
(A). MO/M� from TSCC patients (n � 6) were allowed to migrate against IFN�2b-indsupernatant (IFN�2b-Sup) and the same fromuntreated PBMC (NT-Sup) (B). TSCCMO
nd untreated cells was checked against recombinantMIP-1� with orwithout CCR5 neutraar diagram, *p � 0.01, **p � 0.001, in comparison with other groups (A.1, B.1, and C.1) anlower wells (25 �L/well). Cell suspension (50 �L) at a concentra-tion of 1 106 cells/mL was added to each well of the upperchamber, which was separated from the lower well by a 5-�mpore size polycarbonate polyvinyl pyrrolidone–free membrane.The chamber was incubated for 90 minutes at 37�C in an atmo-sphere containing 5% CO2. The membrane was then carefullyemoved, washed, fixed in methanol, stained in Giemsa stain1:10 dilution), and mounted on a slide. Migrated cells adheredo the lower surface of the membrane were counted using a lighticroscope. Five different fields at 40 were observed and theean cell count value was taken.
.13. Enzyme-linked immunosorbent assay (ELISA) for IFN-� release
IFN-� wasmeasured in the T-cell culture supernatant after theircoculture with TSCC MO/M¦ by ELISA using a commercially avail-able kit. In brief, 96-wellmicrotiter plateswere coatedwith captureantibody (anti-IFN-�), incubated overnight at 4�C, and blockedwith% bovine serum albumin for 1 hour. After being washed, 100 �L of
cell-free supernatant was added to each well to incubate for 2hours. Bound cytokines were detected using biotinylated mouseantihuman IFN-� and avidin–horseradish peroxidase subse-uently. Color was developed with TMB substrate solution. Theeaction was stopped with 2 N H2SO4 and absorbance was mea-sured at 450 nm using a microplate reader (Tecan Spectra, Grodig,Austria).
2.14. Cytotoxicity assay
The cytotoxicity of MO/M¦ and nonadherent fractions of PBMCwas tested against macrophage-specific U937 and HEp2 cells, re-spectively, by LDH release assay using a commercially availablecytotoxicity detection kit. In brief, cancer cells (1 104 of each)were plated overnight as target in 96-well, flat-bottom cell culture
C) monocytes/macrophages (MO/M�) can be rectified by interferon-�2b (IFN�2b).s (n � 10). Adherent fractionswere allowed tomigrate against recombinantMIP-1�
peripheral blood mononuclear cell (PBMC; from a heterologous individual) culture(n�6)were incubatedwith orwithout IFN�2b for 48hours andmigration of treated
a (TSCidualuced/M�
lization (C). Themean � SD ofmigrated cells from TSCC patients is presented in thed representative images of migrated cells are shown in each case (A.2, B.2, and C.2).
s
(H
T
i
c
T
Mt
Cu
m
1s
vdfia
(
m
tt
rc(m*
K. Chakraborty et al. / Human Immunology 73 (2012) 38-47 43
plates. The next day, adherent MO/M¦ or the nonadherent fractionof PBMCwas added as an effector in each well and cocultured for 4hours. After incubation the plate was centrifuged for 5 minutes at400g, and cell-free culture supernatant (100 �L) was used to mea-ure the level of released LDH.
Nonadherent fractions of PBMC obtained from TSCC patientsn � 5) at 3 different stages of treatment were also incubated withEp2 cells to assess their cytotoxic ability by LDH release assay.% Cytotoxicity � (lysis from effector–targetmixture – lysis from
effector only) – spontaneous lysis/maximum lysis – spontaneouslysis 100.
2.15. Statistical analysis
All results represent the average of separate in vitro experi-ments. Thenumber of experiments is indicated in theResults and inthe figure legends. In each experiment a value represents themeanof 3 individual observations presented as means � standard devi-ation (SD). Statistical significance was established using Tukey’stest based on oneway analysis of variance using INSTAT 3 software(INSTAT3, Albania).
3. Results
3.1. IFN�2b upregulates suppressed CC receptor/ligand expression inSCC patients in vitro
In view of the unique immunomodulatory property of IFN�2breported by us [12–14] and other laboratories [7,8], here we stud-ed the role of IFN�2b inmodulation of the dysregulated CC chemo-kine signaling in cancer. Purified MO/M� from TSCC patients (n �12) were treated with IFN�2b for 48 hours and assessed for CCR5expression. Downregulated CCR5 expression onMO/M� fromTSCCpatientswas significantly increased after IFN�2b treatment inmostcases (Fig. 1A and 1B). No such upregulation was noted in cases ofMO/M� from healthy individuals (Fig. 1A). mRNA analysis repre-senting the CCR5 gene also demonstrated upregulated expressionafter IFN�2b treatment (Fig. 1C and 1D).
Intracellular release of chemokines (RANTES, MIP-1�, and MIP-1�) fromMO/M� of TSCC (n � 12) patients was induced by IFN�2b(Fig. 2A and 2B). The results indicate that IFN�2b upregulates intra-ellular expression of RANTES, MIP-1�, and MIP-1� in MO/M�obtained from TSCC patients but not in healthy MO/M�.
3.2. IFN�2b upregulates suppressed CC receptor/ligand expression inSCC patients in vivo
We observed that in vitro treatment of MO/M� from TSCC pa-tients with IFN�2b upregulates suppressed CCR5 receptor/ligandexpression. Next, we analyzed whether the in vitro influence ofIFN�2b is also operable in an in vivo system. For these experiments,O/M� were isolated from TSCC patients who received chemo-
herapy followed by IFN�2b treatment.MO/M� fromTSCC patientsexhibited low CC receptor/ligand expression and cisplatin � 5-FUchemotherapy again intensified receptor/ligand downregulation.Interestingly, IFN�2b treatment upregulates the expression ofCR5 onMO/M� from 5 of 5 patients (Fig. 3A). Such treatment alsopregulates the expression of RANTES, MIP-1�, and MIP-1� in
MO/M� (Fig. 3B).
3.3. IFN�2b efficiently enhances the migratory ability of TSCConocytes
Trafficking of type 1 skewed MO/M� in tumor microenviron-ment is downregulated, possibly because of the downregulation ofCCR5 and corresponding ligands. Freshly isolated MO/M� fromTSCC (n � 10) and healthy individuals (n � 10) were allowed tomigrate against a standard chemoattractant (recombinant MIP-1�,0 ng/mL). The number of migrated cells from TSCC patients was
ignificantly less in comparison with the same from healthy indi-wfl
iduals (Fig. 4A). Neutralization of CCR5, but not CXCR3, furtherecreased the migration of monocytic cells (data not shown). Puri-ed MO/M� from TSCC patients (n � 6) were allowed to migrategainst the supernatant of IFN�2b-stimulated PBMC culture
(IFN�2b-Sup). Supernatant from the culturewithout any treatmentNT-Sup) was used as a control. Migration of MO/M� againstIFN�2b-Sup was greater than that of NT-Sup (Fig. 4B). In anotherexperiment,MO/M� fromTSCCpatientswere cultured for 48hourswith IFN�2b and chemotaxis of these cells against a standard che-oattractant (recombinant MIP-1�, 10 ng/mL) was tested (n � 6).
MO/M� from TSCC patients after IFN�2b treatment have demon-strated significantly greater migratory activity than those fromuntreated controls. Such enhanced migration was inhibited afterCCR5 neutralization following IFN�2b treatment (Fig. 4C) andreaches the same level of untreated MO/M� from TSCC with orwithout CCR5 neutralization. Such inhibition in migration was notnoted using CXCR3 in the neutralization study.
3.4. IFN�2b enhances tumoricidal functions of MO/M�
It is of interest to understand the functions of migratedMO/M�hatmay be either tumoricidal or antigen presenting. To determinehe tumoricidal functions of these cells,MO/M�were isolated from
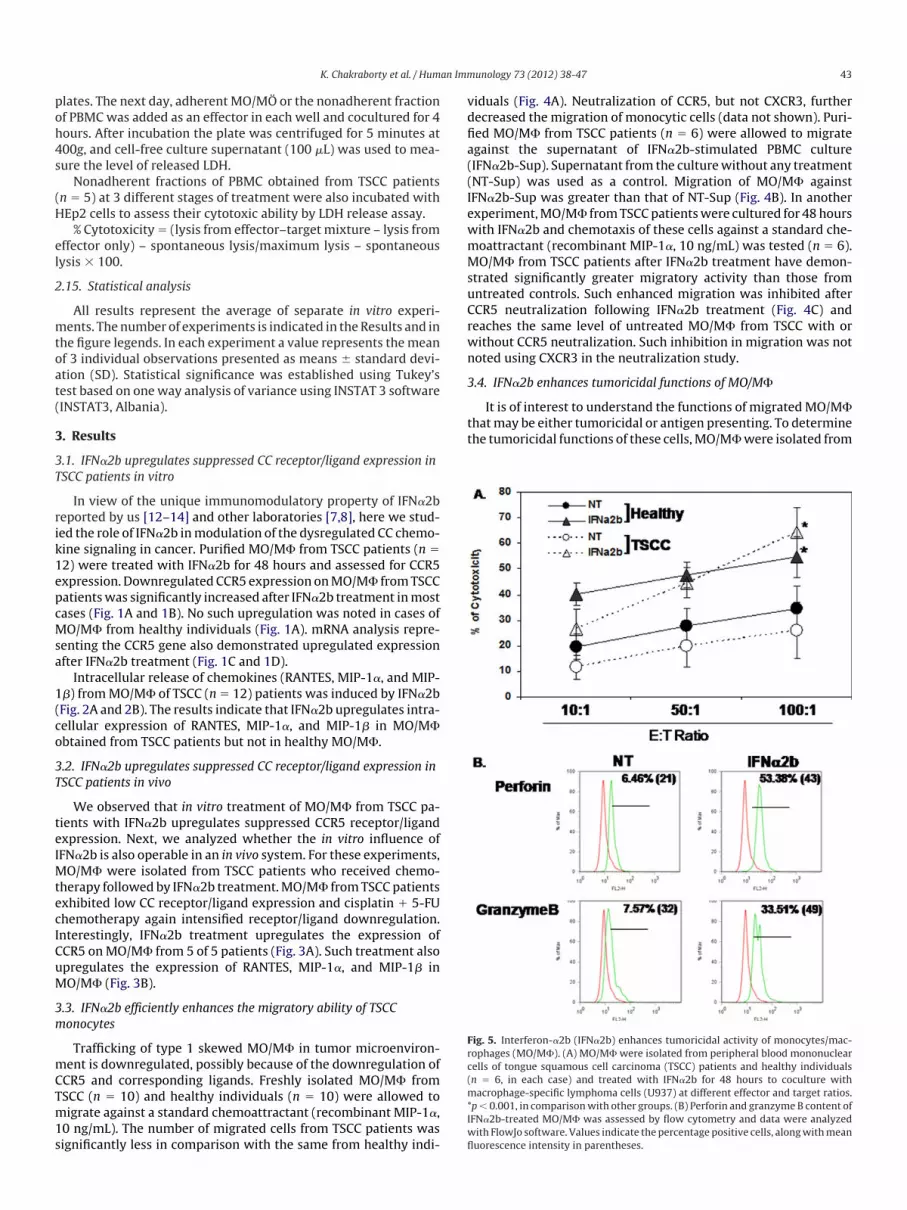
Fig. 5. Interferon-�2b (IFN�2b) enhances tumoricidal activity of monocytes/mac-ophages (MO/M�). (A) MO/M� were isolated from peripheral blood mononuclearells of tongue squamous cell carcinoma (TSCC) patients and healthy individualsn � 6, in each case) and treated with IFN�2b for 48 hours to coculture withacrophage-specific lymphoma cells (U937) at different effector and target ratios.
p� 0.001, in comparisonwith other groups. (B) Perforin and granzymeB content ofIFN�2b-treated MO/M� was assessed by flow cytometry and data were analyzed
ith FlowJo software. Values indicate thepercentagepositive cells, alongwithmeanuorescence intensity in parentheses.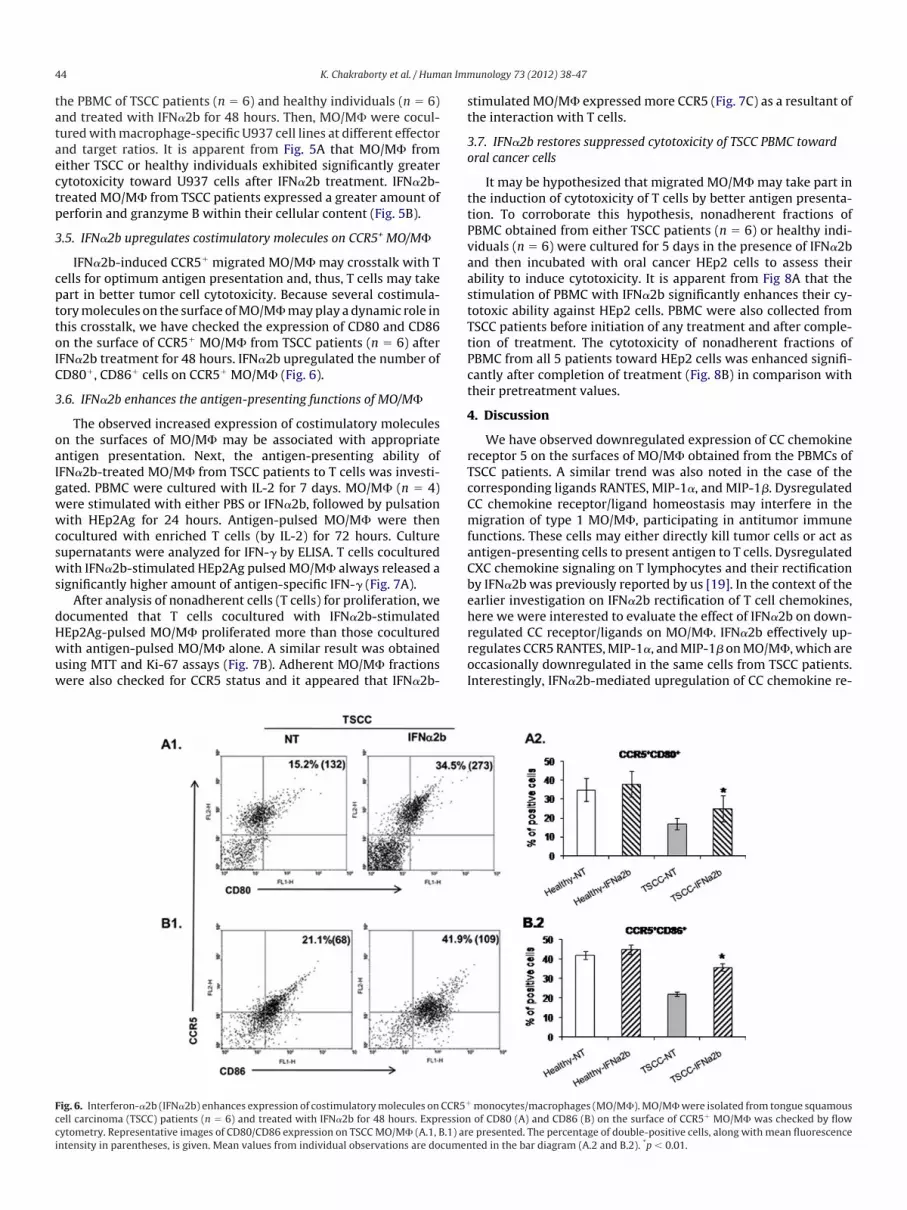
tp
3
cptto
oaIg
w
d
uw
st
3o
tTtPct
4
rTc
c
K. Chakraborty et al. / Human Immunology 73 (2012) 38-4744
the PBMC of TSCC patients (n � 6) and healthy individuals (n � 6)and treated with IFN�2b for 48 hours. Then, MO/M� were cocul-turedwithmacrophage-specific U937 cell lines at different effectorand target ratios. It is apparent from Fig. 5A that MO/M� fromeither TSCC or healthy individuals exhibited significantly greatercytotoxicity toward U937 cells after IFN�2b treatment. IFN�2b-reated MO/M� from TSCC patients expressed a greater amount oferforin and granzyme B within their cellular content (Fig. 5B).
.5. IFN�2b upregulates costimulatory molecules on CCR5+ MO/M�
IFN�2b-induced CCR5� migrated MO/M� may crosstalk with Tells for optimum antigen presentation and, thus, T cells may takeart in better tumor cell cytotoxicity. Because several costimula-orymolecules on the surface ofMO/M�mayplay a dynamic role inhis crosstalk, we have checked the expression of CD80 and CD86n the surface of CCR5� MO/M� from TSCC patients (n � 6) after
IFN�2b treatment for 48 hours. IFN�2b upregulated the number ofCD80�, CD86� cells on CCR5� MO/M� (Fig. 6).
3.6. IFN�2b enhances the antigen-presenting functions of MO/M�
The observed increased expression of costimulatory moleculesn the surfaces of MO/M� may be associated with appropriatentigen presentation. Next, the antigen-presenting ability ofFN�2b-treated MO/M� from TSCC patients to T cells was investi-ated. PBMC were cultured with IL-2 for 7 days. MO/M� (n � 4)
were stimulated with either PBS or IFN�2b, followed by pulsationwith HEp2Ag for 24 hours. Antigen-pulsed MO/M� were thencocultured with enriched T cells (by IL-2) for 72 hours. Culturesupernatants were analyzed for IFN-� by ELISA. T cells coculturedith IFN�2b-stimulated HEp2Ag pulsedMO/M� always released a
significantly higher amount of antigen-specific IFN-� (Fig. 7A).After analysis of nonadherent cells (T cells) for proliferation, we
ocumented that T cells cocultured with IFN�2b-stimulatedHEp2Ag-pulsed MO/M� proliferated more than those coculturedwith antigen-pulsed MO/M� alone. A similar result was obtainedsing MTT and Ki-67 assays (Fig. 7B). Adherent MO/M� fractionsere also checked for CCR5 status and it appeared that IFN�2b-
Fig. 6. Interferon-�2b (IFN�2b) enhances expression of costimulatorymolecules onell carcinoma (TSCC) patients (n � 6) and treated with IFN�2b for 48 hours. Expr
cytometry. Representative images of CD80/CD86 expression on TSCCMO/M� (A.1, B.1) arintensity in parentheses, is given. Mean values from individual observations are documentimulatedMO/M� expressedmore CCR5 (Fig. 7C) as a resultant ofhe interaction with T cells.
.7. IFN�2b restores suppressed cytotoxicity of TSCC PBMC towardral cancer cells
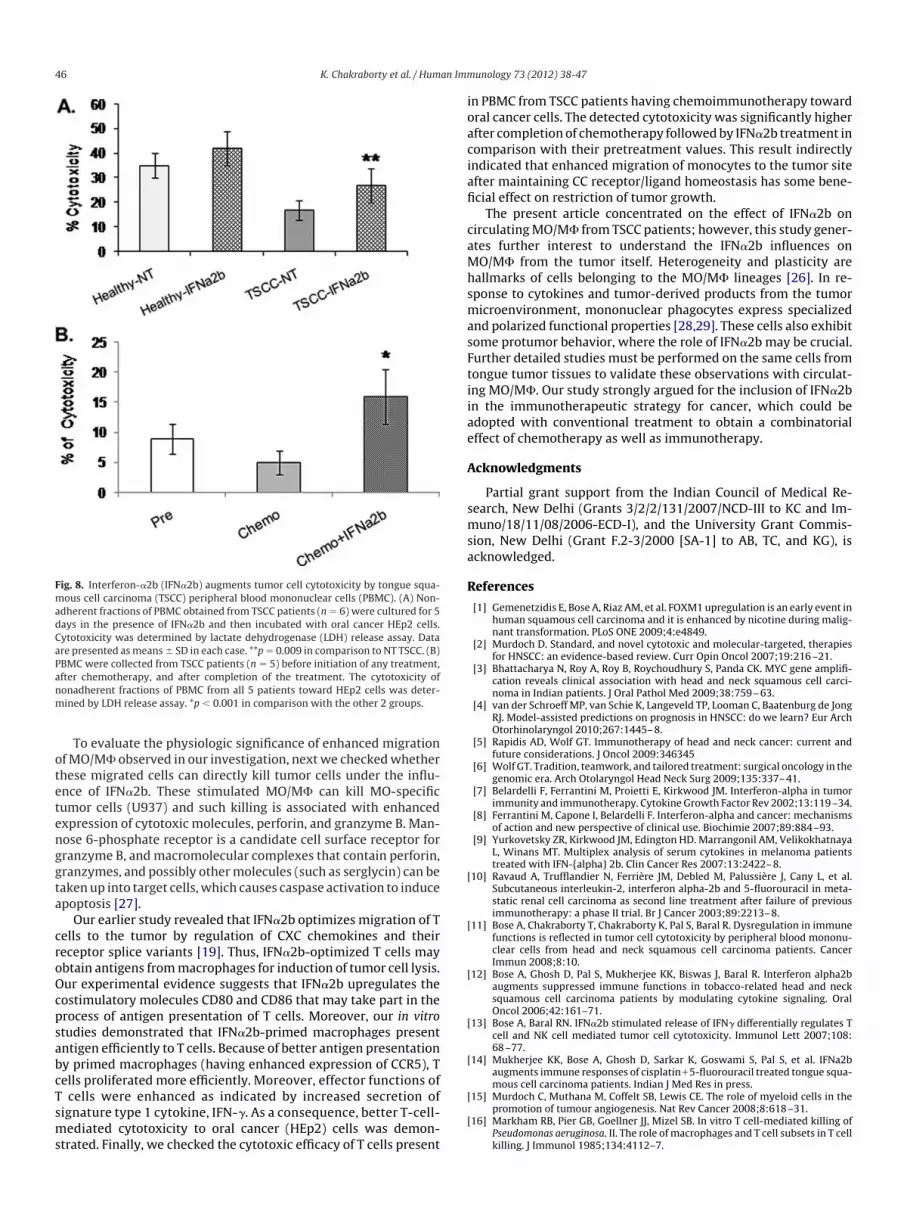
It may be hypothesized that migrated MO/M� may take part inthe induction of cytotoxicity of T cells by better antigen presenta-tion. To corroborate this hypothesis, nonadherent fractions ofPBMC obtained from either TSCC patients (n � 6) or healthy indi-viduals (n � 6) were cultured for 5 days in the presence of IFN�2band then incubated with oral cancer HEp2 cells to assess theirability to induce cytotoxicity. It is apparent from Fig 8A that thestimulation of PBMC with IFN�2b significantly enhances their cy-otoxic ability against HEp2 cells. PBMC were also collected fromSCC patients before initiation of any treatment and after comple-ion of treatment. The cytotoxicity of nonadherent fractions ofBMC from all 5 patients toward HEp2 cells was enhanced signifi-antly after completion of treatment (Fig. 8B) in comparison withheir pretreatment values.
. Discussion
We have observed downregulated expression of CC chemokineeceptor 5 on the surfaces of MO/M� obtained from the PBMCs ofSCC patients. A similar trend was also noted in the case of theorresponding ligands RANTES, MIP-1�, and MIP-1�. DysregulatedCC chemokine receptor/ligand homeostasis may interfere in themigration of type 1 MO/M�, participating in antitumor immunefunctions. These cells may either directly kill tumor cells or act asantigen-presenting cells to present antigen to T cells. DysregulatedCXC chemokine signaling on T lymphocytes and their rectificationby IFN�2b was previously reported by us [19]. In the context of theearlier investigation on IFN�2b rectification of T cell chemokines,here we were interested to evaluate the effect of IFN�2b on down-regulated CC receptor/ligands on MO/M�. IFN�2b effectively up-regulates CCR5 RANTES,MIP-1�, andMIP-1� onMO/M�, which areoccasionally downregulated in the same cells from TSCC patients.Interestingly, IFN�2b-mediated upregulation of CC chemokine re-
monocytes/macrophages (MO/M�).MO/M�were isolated from tongue squamousof CD80 (A) and CD86 (B) on the surface of CCR5� MO/M� was checked by flow
CCR5�
ession
e presented. The percentage of double-positive cells, along with mean fluorescenceted in the bar diagram (A.2 and B.2). *p � 0.01.
Mlgill
tamlnpad
h
lI1cctst
kT
Tnstpoat
fg
K. Chakraborty et al. / Human Immunology 73 (2012) 38-47 45
ceptor/ligands is restricted in TSCC MO/M�, but not in cells fromhealthy individuals. These results suggest that IFN�2b is active onO/M� onlywhen CCR5 expression is decreased below the normal
evel and their upregulation is required to maintain optimum mi-ration of mononuclear cells to the desired site. Apart from in vitronfluence of IFN�2b upon maintenance of CC chemokine receptor/igand homeostasis, we also checked the status of these receptor/igands after IFN�2b immunotherapy in TSCC patients along withcisplatin � 5-FU chemotherapy. We have experienced that theumor load itself decreases CC chemokines at receptor/ligand levelsnd was further downregulated following chemotherapy. This cu-ulative negative impact significantly hindered the CCR5 receptor/
igand dependent trafficking of MO/M�, thereby limiting theatural immune-mediated tumor eradication. The decreased ex-ression of CCR5 in monocytes from HNSCC patients was reportednd its association with upregulated Cox2 and PG expression wasescribed [24]. Evidences obtained from our in vitro and in vivo
studies suggested that IFN�2b increases CCR5 in isolated MO/M�from TSCC patients both in vitro and in vivo. Increment in thereceptor is correlatedwith the IFN�2b-mediated release of ligands.It may be hypothesized from our earlier report that IFN�2b induces
Fig. 7. Interferon-�2b (IFN�2b) enhances antigen presentation of monocytes and mPeripheral blood mononuclear cells were cultured in presence of interleukin (IL)-2buffered saline or IFN�2b, followed by pulsation with HEp2Ag for 24 hours. Antigenindividuals for 72 hours (A). Culture supernatant was analyzed for IFN-� by enzymeor proliferation by MTT (B.1) and Ki-67 (B.2) assays. (C) Adherent MO/M� fractionroups.
igher expression of STAT1 and STAT4 on monocytes, thereby re- a
easing a greater amount of IL-12, which ultimately propagates theFN-� secretion to polarize the system toward type 1 [12]. This typeimmune microenvironment may help in the upregulation of CChemokine receptor/ligand expression on MO/M� under canceronditions [25]. Upregulated CCR5 expression may impose a posi-ive impact on migration of monocytes to the tumor site. Ourtudies on monocytic migration in the Boyden chamber revealedhat direct stimulation of TSCC MO/M� with IFN�2b enhances itsmigratory ability. By contrast, IFN�2b-induced secreted chemo-ines also drive chemotaxis of an increased number of cells fromSCC patients. Both observations suggest that IFN�2b could en-
hance chemotaxis either by functional activation of CCR5� TSCCMO or by enhancing the release of RANTES, MIP-1�, and MIP-1�.he involvement of CCR5 in this process was confirmed by CCR5eutralization, resulting in inhibition ofmonocyte chemotaxis. Thistudy suggests that CCR5 is involved in classicalMO/M� migration athe tumor site and that for tumor eradication classical mononuclearhagocytes express IL-12high, IL-10low phenotypes, efficient producersf effector molecules (reactive oxygen and nitrogen intermediates)nd inflammatory cytokines (TNF) to participate as inducer and effec-or cells in polarized type 1 responses, thereby mediating resistance
phages (MO/M�) from tongue squamous cell carcinoma (TSCC) patients to T cells.days. MO/M� (n � 4) from TSCC patients were stimulated with either phosphate-dMO/M� were then cocultured with IL-2-treated or untreated T cells from healthyd immunosorbent assay. (B) Nonadherent fractions containing T cells were checkede checked for CCR5 status by flow cytometry. *p � 0.01 in comparison with other
acrofor 7-pulse-linkes wer
gainst intracellular parasites and tumors [26].
ote
cr
cpsabcTsms
ioa
MhsmasFti
[
[
[
[
madCa
K. Chakraborty et al. / Human Immunology 73 (2012) 38-4746
To evaluate the physiologic significance of enhanced migrationfMO/M� observed in our investigation, nextwe checkedwhetherhese migrated cells can directly kill tumor cells under the influ-nce of IFN�2b. These stimulated MO/M� can kill MO-specific
tumor cells (U937) and such killing is associated with enhancedexpression of cytotoxic molecules, perforin, and granzyme B. Man-nose 6-phosphate receptor is a candidate cell surface receptor forgranzyme B, andmacromolecular complexes that contain perforin,granzymes, and possibly othermolecules (such as serglycin) can betakenup into target cells,which causes caspase activation to induceapoptosis [27].
Our earlier study revealed that IFN�2b optimizes migration of Tells to the tumor by regulation of CXC chemokines and theireceptor splice variants [19]. Thus, IFN�2b-optimized T cells mayobtain antigens frommacrophages for induction of tumor cell lysis.Our experimental evidence suggests that IFN�2b upregulates theostimulatory molecules CD80 and CD86 that may take part in therocess of antigen presentation of T cells. Moreover, our in vitrotudies demonstrated that IFN�2b-primed macrophages presentntigen efficiently to T cells. Because of better antigen presentationy primed macrophages (having enhanced expression of CCR5), Tells proliferated more efficiently. Moreover, effector functions ofcells were enhanced as indicated by increased secretion of
ignature type 1 cytokine, IFN-�. As a consequence, better T-cell-ediated cytotoxicity to oral cancer (HEp2) cells was demon-
Fig. 8. Interferon-�2b (IFN�2b) augments tumor cell cytotoxicity by tongue squa-ous cell carcinoma (TSCC) peripheral blood mononuclear cells (PBMC). (A) Non-dherent fractions of PBMC obtained from TSCC patients (n � 6) were cultured for 5ays in the presence of IFN�2b and then incubated with oral cancer HEp2 cells.ytotoxicity was determined by lactate dehydrogenase (LDH) release assay. Datare presented asmeans � SD in each case. **p � 0.009 in comparison to NT TSCC. (B)
PBMC were collected from TSCC patients (n � 5) before initiation of any treatment,after chemotherapy, and after completion of the treatment. The cytotoxicity ofnonadherent fractions of PBMC from all 5 patients toward HEp2 cells was deter-mined by LDH release assay. *p � 0.001 in comparison with the other 2 groups.
trated. Finally, we checked the cytotoxic efficacy of T cells present
n PBMC from TSCC patients having chemoimmunotherapy towardral cancer cells. The detected cytotoxicity was significantly higherfter completion of chemotherapy followedby IFN�2b treatment in
comparison with their pretreatment values. This result indirectlyindicated that enhanced migration of monocytes to the tumor siteafter maintaining CC receptor/ligand homeostasis has some bene-ficial effect on restriction of tumor growth.
The present article concentrated on the effect of IFN�2b oncirculatingMO/M� from TSCC patients; however, this study gener-ates further interest to understand the IFN�2b influences onO/M� from the tumor itself. Heterogeneity and plasticity areallmarks of cells belonging to the MO/M� lineages [26]. In re-ponse to cytokines and tumor-derived products from the tumoricroenvironment, mononuclear phagocytes express specializednd polarized functional properties [28,29]. These cells also exhibitome protumor behavior, where the role of IFN�2b may be crucial.urther detailed studies must be performed on the same cells fromongue tumor tissues to validate these observations with circulat-ng MO/M�. Our study strongly argued for the inclusion of IFN�2bin the immunotherapeutic strategy for cancer, which could beadopted with conventional treatment to obtain a combinatorialeffect of chemotherapy as well as immunotherapy.
Acknowledgments
Partial grant support from the Indian Council of Medical Re-search, New Delhi (Grants 3/2/2/131/2007/NCD-III to KC and Im-muno/18/11/08/2006-ECD-I), and the University Grant Commis-sion, New Delhi (Grant F.2-3/2000 [SA-1] to AB, TC, and KG), isacknowledged.
References
[1] Gemenetzidis E, Bose A, Riaz AM, et al. FOXM1upregulation is an early event inhuman squamous cell carcinoma and it is enhanced by nicotine during malig-nant transformation. PLoS ONE 2009;4:e4849.
[2] Murdoch D. Standard, and novel cytotoxic and molecular-targeted, therapiesfor HNSCC: an evidence-based review. Curr Opin Oncol 2007;19:216–21.
[3] Bhattacharya N, Roy A, Roy B, Roychoudhury S, Panda CK. MYC gene amplifi-cation reveals clinical association with head and neck squamous cell carci-noma in Indian patients. J Oral Pathol Med 2009;38:759–63.
[4] van der Schroeff MP, van Schie K, Langeveld TP, Looman C, Baatenburg de JongRJ. Model-assisted predictions on prognosis in HNSCC: do we learn? Eur ArchOtorhinolaryngol 2010;267:1445–8.
[5] Rapidis AD, Wolf GT. Immunotherapy of head and neck cancer: current andfuture considerations. J Oncol 2009:346345
[6] Wolf GT. Tradition, teamwork, and tailored treatment: surgical oncology in thegenomic era. Arch Otolaryngol Head Neck Surg 2009;135:337–41.
[7] Belardelli F, Ferrantini M, Proietti E, Kirkwood JM. Interferon-alpha in tumorimmunity and immunotherapy. CytokineGrowth Factor Rev 2002;13:119–34.
[8] Ferrantini M, Capone I, Belardelli F. Interferon-alpha and cancer: mechanismsof action and new perspective of clinical use. Biochimie 2007;89:884–93.
[9] Yurkovetsky ZR, Kirkwood JM, Edington HD. Marrangonil AM, VelikokhatnayaL, Winans MT. Multiplex analysis of serum cytokines in melanoma patientstreated with IFN-{alpha} 2b. Clin Cancer Res 2007:13:2422–8.
10] Ravaud A, Trufflandier N, FerriÉre JM, Debled M, PalussiÉre J, Cany L, et al.Subcutaneous interleukin-2, interferon alpha-2b and 5-fluorouracil in meta-static renal cell carcinoma as second line treatment after failure of previousimmunotherapy: a phase II trial. Br J Cancer 2003;89:2213–8.
11] Bose A, Chakraborty T, Chakraborty K, Pal S, Baral R. Dysregulation in immunefunctions is reflected in tumor cell cytotoxicity by peripheral blood mononu-clear cells from head and neck squamous cell carcinoma patients. CancerImmun 2008;8:10.
12] Bose A, Ghosh D, Pal S, Mukherjee KK, Biswas J, Baral R. Interferon alpha2baugments suppressed immune functions in tobacco-related head and necksquamous cell carcinoma patients by modulating cytokine signaling. OralOncol 2006;42:161–71.
13] Bose A, Baral RN. IFN�2b stimulated release of IFN� differentially regulates Tcell and NK cell mediated tumor cell cytotoxicity. Immunol Lett 2007;108:68–77.
[14] Mukherjee KK, Bose A, Ghosh D, Sarkar K, Goswami S, Pal S, et al. IFNa2baugments immune responses of cisplatin�5-fluorouracil treated tongue squa-mous cell carcinoma patients. Indian J Med Res in press.
[15] Murdoch C, Muthana M, Coffelt SB, Lewis CE. The role of myeloid cells in thepromotion of tumour angiogenesis. Nat Rev Cancer 2008;8:618–31.
[16] Markham RB, Pier GB, Goellner JJ, Mizel SB. In vitro T cell-mediated killing of
Pseudomonas aeruginosa. II. The role ofmacrophages and T cell subsets in T cellkilling. J Immunol 1985;134:4112–7.
K. Chakraborty et al. / Human Immunology 73 (2012) 38-47 47
[17] Strieter RM, Belperio JA, Phillips RJ, Keane MP. CXC chemokines in angiogene-sis of cancer. Semin Cancer Biol 2004;14:195–200.
[18] Sallusto F, Lanzavecchia A, Mackay CR. Chemokines and chemokines receptorsin T-cell priming and Th1/Th2-mediated responses. Immunol Today 1998;19:568–74.
[19] Chakraborty K, Bose A, Pal S, Chattopadhyay U, Baral R. Interferon alpha2brestores the impaired chemotactic activity of peripheral blood mononuclearcells fromhead andneck squamous cell carcinomapatients bymodulating CXCreceptor ligand interaction. J Interferon Cytokine Res 2008;28:487–99.
[20] Giri RK, Rajagopal V, Shahi S, Zlokovic BV, Kalra VK. Mechanism of amyloidpeptide induced CCR5 expression inmonocytes and its inhibition by siRNA forEgr-1. Am J Physiol Cell Physiol 2005;289:C264–76.
[21] Samson M, Labbe O, Mollereau C, Vassart G, Parmentier M. Molecular cloningand functional expression of a new human CC-chemokine receptor gene.Biochemistry 1996;35:3362–7.
[22] Qin S, Rottman JB, Myers P, Kassam N, Weinblatt M, Loetscher M, et al. The
chemokine receptors CXCR3 and CCR5 mark subsets of T cells associated withcertain inflammatory reactions. J Clin Invest 1998;101:746–54.[23] Schall TJ, Bacon K, Toy KJ, Goeddel DV. Selective attraction of monocytes and Tlymphocytes of the memory phenotype by cytokine RANTES. Nature 1990;347:669–71.
[24] Lang S, Lauffer L, Clausen C, L×hr I, Schmitt B, H×lzel D, et al. Impaired mono-cyte function in cancer patients: restoration with a cyclooxygenase-2 inhibi-tor. FASEB J 2003;17:286–8.
[25] Dehmel S, Wang S, Schmidt C, Kiss E, Loewe RP, Chilla S, et al. Chemokinereceptor Ccr5 deficiency induces alternative macrophage activation and im-proves long-term renal allograft outcome. Eur J Immunol 2010;40:267–78.
[26] Mantovani A, Sica A, Locati M. Macrophage polarization comes of age. Immu-nity 2005;23:344–6.
[27] Trapani JA, Smyth MJ. Functional significance of the perforin/granzyme celldeath pathway. Nat Rev Immunol 2002;2:735–47.
[28] Gordon S. Alternative activation of macrophages. Nat Rev Immunol 2003;3:23–35.
[29] Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine
system in diverse forms of macrophage activation and polarization. TrendsImmunol 2004;25:677–86.