dysregulated physiological stress systems and accelerated cellular aging
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Dysregulated physiological stress systems and accelerated cellularaging
Dóra Révész a,*, Josine E. Verhoeven a, Yuri Milaneschi a, Eco J.C.N. de Geus b,Owen M. Wolkowitz c, Brenda W.J.H. Penninx a
aDepartment of Psychiatry, EMGO Institute for Health and Care Institute, VU University Medical Center, Amsterdam, the NetherlandsbDepartment of Biological Psychology, EMGO Institute for Health and Care Institute, VU University Medical Center, Amsterdam, the NetherlandscDepartment of Psychiatry, University of California San Francisco School of Medicine, San Francisco, CA, USA
a r t i c l e i n f o
Article history:Received 17 May 2013Received in revised form 20 December 2013Accepted 23 December 2013Available online 27 December 2013
Keywords:Cellular agingTelomeresStressAgingInflammationAutonomic nervous systemCortisol
a b s t r a c t
Exposure to chronic stressors is associated with accelerated biological aging as indicated by reducedleukocyte telomere length (LTL). This impact could be because of chronic overactivation of the body’sphysiological stress systems. This study examined the associations between LTL and the immune system,hypothalamic-pituitary-adrenal axis and autonomic nervous system. LTL was assessed in 2936 adultsfrom the Netherlands Study of Depression and Anxiety. Inflammation markers (interleukin-6, c-reactiveprotein, tumor necrosis factor-alpha), hypothalamic-pituitary-adrenal-axis indicators (salivary cortisolawakening curve [area under the curve indicators, with respect to the ground and increase], eveninglevels, 0.5 mg dexamethasone cortisol suppression ratio), and autonomic nervous system measures(heart rate, respiratory sinus arrhythmia, pre-ejection period) were determined. Linear regression ana-lyses were performed and adjusted for sociodemographic, lifestyle and clinical factors. Shorter LTL wassignificantly associated with higher c-reactive protein, interleukin-6, area under the curve with respect toincrease, and heart rate. A cumulative index score was calculated based on the number of highest tertilesof these 4 stress markers. LTL demonstrated a significant gradient within subjects ranging from havingzero (5528 base pairs) to having 4 elevated stress markers (5371 base pairs, p for trend ¼ 0.002), cor-responding to a difference of 10 years of accelerated biological aging. Contrary to the expectations,shorter LTL was also associated with longer pre-ejection period, indicating lower sympathetic tone. Thislarge-scale study showed that inflammation, high awakening cortisol response, and increased heart rateare associated with shorter LTL, especially when they are dysregulated cumulatively.
� 2014 Elsevier Inc. All rights reserved.
1. Introduction
Aging is accompanied by the onset of various somatic dis-eases, but variation in the age of onset of these diseases suggestsa high degree of variability in biological aging across humans.Lately, telomere length has emerged as a novel marker of bio-logical aging (Blackburn, 2001; Olovnikov, 1996). Telomeres aretandem repeat DNA sequences that form protective caps atchromosome ends (Blackburn, 2001). During each somatic celldivision, the DNA loses telomeric repeats with an estimatedshortening rate of 14e20 or more base pairs per year, eventuallycausing replicative cell senescence (Blackburn, 1991; Cawthonet al., 2003; Hartmann et al., 2010; Wikgren et al., 2012).
Successful maintenance of telomeres is crucial to human healthbefore they reach a critically short length and become dysfunc-tional at a threshold of 77 base pairs (Ledford, 2007). Normaltelomere maintenance requires the cellular enzyme telomerasethat adds telomeric DNA, thus preserving telomere length andhealthy cell function (Blackburn, 2001). Several recent studieshave demonstrated an inverse relationship between leukocytetelomere length (LTL) and the risk of subsequent somatic ill-nesses (e.g., cardiovascular diseases, dementia) and prematuredeath (Epel et al., 2006, 2008; Martin-Ruiz et al., 2006; Ruffet al., 2012; Weischer et al., 2012). LTL is a heritable trait, withgenetics contributing to approximately 70% of the variability(Broer et al., 2013), leaving a large part of the variability becauseof external factors such as environmental and lifestyle factors(Aubert and Lansdorp, 2008).
Shorter LTL has often been observed in individuals exposed topsychological stressors, such as childhood and caregiving stress(Epel et al., 2004; Kiecolt-Glaser et al., 2011), as well as in
* Corresponding author at: A.J. Ernststraat 1187, Room M1.06, PO Box 74077, 1070BB, 1081 HL Amsterdam, the Netherlands. Tel.: þ31 20 788 4596; fax: þ31 20 7885664.
E-mail address: [email protected] (D. Révész).
Contents lists available at ScienceDirect
Neurobiology of Aging
journal homepage: www.elsevier .com/locate/neuaging
0197-4580/$ e see front matter � 2014 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.neurobiolaging.2013.12.027
Neurobiology of Aging 35 (2014) 1422e1430

Author's personal copy
individuals with stress-related diseases such as depressive disor-ders (Hoen et al., 2011; Verhoeven et al., 2013). As individualsdiffer in their psychological and biological responses to stress, LTLmay represent a biomarker for assessing an individual’s cumula-tive exposure to stressful conditions (O’Donovan et al., 2012).Psychological stress might influence the length of telomericDNA and overall health, possibly through the alteration ofvarious physiological stress systems. The body’s major physio-logical stress systems are the immuno-inflammatory system, thehypothalamus-pituitary-adrenal (HPA)-axis, and the autonomicnervous system (ANS). Chronic stress can lead to systemic dys-regulations of these systems, or increased “allostatic load”,resulting in systemic inflammation, hyperactivity of the HPA-axis,sympathetic activation and parasympathetic withdrawal (Kiecolt-Glaser et al., 2003; McEwen, 2008). Such dysregulations have anegative impact on overall health (Carrero et al., 2008; Licht et al.,2010; van Reedt Dortland et al., 2012), possibly by affecting thetelomere maintenance system and contributing to the shorteningof telomeres. Indeed, in vitro studies in human T-lymphocytesreported that cortisol exposure may result in down-regulation oftelomerase activity, and that senescent immune cells showedmore inflammatory activity than cells with longer TL (Choi et al.,2008; Merino et al., 2011). Several in vivo studies have also re-ported that shorter telomeres are associated with dysregulationsin inflammatory markers (Carrero et al., 2008; Fitzpatrick et al.,2007; Kiecolt-Glaser et al., 2011, 2013; Masi et al., 2012; O’Do-novan et al., 2011), HPA-axis (Epel et al., 2006; Kroenke et al.,2011; Tomiyama et al., 2012; Wikgren et al., 2012), and auto-nomic nervous system (ANS) function (Epel et al., 2006; Kroenkeet al., 2011).
All these studies raise the suggestion that dysregulation ofcentral bodily stress systems is related to shorter LTL, but findingsare still inconsistent (Parks et al., 2009; Wikgren et al., 2012;Wolkowitz et al., 2011). Several of these previous studies werelimited by a small sample (n < 100; Epel et al., 2006; Kroenkeet al., 2011; O’Donovan et al., 2009; Ornish et al., 2008;Tomiyama et al., 2012; Wikgren et al., 2012) or a restrictedpopulation (one gender [Carrero et al., 2008; Epel et al., 2006;O’Donovan et al., 2009; Tomiyama et al., 2012]; elderly in-dividuals [Kiecolt-Glaser et al., 2011; O’Donovan et al., 2009,2011; Wikgren et al., 2012]; children [Kroenke et al., 2011];population with somatic diseases [Carrero et al., 2008; Ornishet al., 2008]); or the lack of adjustment for lifestyle, education,or somatic health indicators (Carrero et al., 2008; Epel et al.,2006; Fitzpatrick et al., 2007; Kiecolt-Glaser et al., 2011;Kroenke et al., 2011; Tomiyama et al., 2012; Wikgren et al., 2012;Wolkowitz et al., 2011). Above all, some studies have indicatedthat cumulative (or combinations of) dysregulations of multiplephysiological stress systems may have the strongest impact onbiological aging (Epel et al., 2006; Kroenke et al., 2011;O’Donovan et al., 2011), but this has not been examined yet forthe 3 major physiological stress systems together. As inflamma-tion, HPA and ANS markers are separately related to various so-matic diseases, the accumulation of these dysregulations mightlead to even worse health outcomes.
The present study examined to what extent dysregulations ofthe 3 major physiological stress systems (HPA-axis, inflammation,and ANS) are associated with LTL in a large-scale cohort studyincluding a sample of adult participants and adjusting for multiplelifestyle and somatic health indicators. In addition, the effect ofcumulative stress system dysregulations on LTL was investigated.We hypothesized that subjects with (over)activated stress systems,characterized by a pro-inflammatory state, hyperactivity of theHPA-axis, and high sympathetic activation combined with para-sympathetic withdrawal will have shorter LTL.
2. Methods
2.1. Study sample
Data were from the Netherlands Study of Depression and Anx-iety, a large ongoing longitudinal cohort study among 2981 adults(18e65 years), which is described elsewhere (Penninx et al., 2008).Briefly, respondents were recruited between September 2004 andFebruary 2007 from the community, primary care, and in special-ized mental health care. Baseline data collection consisted of amedical examination, blood draw, self-report questionnaires and adetailed interview including the lifetime version of the CompositeInterview Diagnostic Instrument (CIDI version 2.1) according to the4th Edition of the Diagnostic and Statistical Manual of MentalDisorders (DSM-IV) criteria, administered by specially trainedresearch staff. The research protocol was approved by the ethicalcommittee of participating universities and all respondents pro-vided written informed consent. Of all participants, 1737 personswere included with a current (i.e., within the past 6 months) DSM-IV diagnosis of depressive (major depressive disorders or dysthy-mia) and/or anxiety disorders (social phobia, generalized anxietydisorder, panic disorder with/without agoraphobia, and agora-phobia only), 592 persons with a remitted (i.e., lifetime, but notcurrent) depressive and/or anxiety disorder, and 652 healthy sub-jects with no lifetime psychiatric disorders. We then excluded 45subjects because of missing LTL data. These subjects were slightlyolder (45.5 vs. 41.8 years, p ¼ 0.06), but did not differ in years ofeducation or gender from the included subjects. The 2936remaining participants (95% from European origin) constitute thebasic sample for the present study, and had at least information onone of the physiological stress systems available. For specificphysiological stress markers, however, we had to exclude subjectswith missing data for that assessment, ranging from 27 to 47missing data points for inflammatory markers, 116e140 missingdata for ANS, and 1031e1183 missing data for HPA-axis measures(see below for further details). For HPA-axis analyses, we addi-tionally excluded 159 subjects who used corticosteroids (n ¼ 134)or were pregnant and/or breastfeeding (n ¼ 25).
2.2. Measurements
2.2.1. LTLFasting blood was drawn from participants in the morning be-
tween 8:30 and 9:30 AM. Peripheral blood mononuclear cells fromall samples were isolated from whole blood using density-gradientcentrifugation (with Ficoll-Paque PLUS) and stored at �80 �Cfreezers. Subsequently, in early 2012, LTL was determined at thelaboratory of Telomere Diagnostics, Inc (TDx, Menlo Park, CA, USA),using quantitative polymerase chain reaction as described earlier(Cawthon, 2002). All qPCRs were carried out on a Roche Lightcycler480 realtime PCR machine with 384-tube capacity (Roche Di-agnostics Corporation, Indianapolis, IN, USA). Telomere sequencecopy number in each patient’s sample (T) was compared with asingle-copy gene copy number (S), relative to a reference sample.The resulting T to S ratio is proportional to mean LTL (Aviv et al.,2011; Cawthon, 2002). To control for inter-assay variability, 8 con-trol DNA samples were included in each run. In each batch, the T toS ratio of each control DNAwas divided by the average T to S for thesame DNA from 10 runs to obtain a normalizing factor. This wasdone for all 8-control samples and the average normalizing factorfor these samples was used to correct the participant DNA samplesto obtain the final T to S ratio. The T to S ratio for each sample wasmeasured twice. If the duplicate T to S value and the initial valuevaried by more than 7%, the sample was run a third time and theaverage of the 2 closest values was reported. The reliability of the
D. Révész et al. / Neurobiology of Aging 35 (2014) 1422e1430 1423

Author's personal copy
assay was adequate: the included quality control DNA samples oneach PCR run illustrated a small intra-assay coefficient of variation(CV ¼ 5.1%), and the inter-assay CV was also sufficiently low (CV ¼4.6%), as well as for the telomere (CV ¼ 2.04%) and the single-geneassays (CV¼ 1.58%), separately. To compare T to S ratios to telomererestriction fragments reported by other studies using Southern blotanalysis, we used the following steps to derive a conversion for-mula. Published work from the Blackburn lab at University of Cal-ifornia, San Francisco (UCSF) used a formula of base pairs ¼ 3274 þ2413*T/S based on comparison of T to S ratios and telomere re-striction fragments analysis of a series of genomic DNA samplesfrom the human fibroblast cell line IMR90 (Lin et al., 2010). Com-parison of the T to S ratios of the 8 control DNA samples derivedfrom Blackburn lab and TDx lab generated the following formula: T/S (UCSF) ¼ (T/S[TDx] � 0.0545)/1.16. Therefore the final formula weused to convert T to S ratios to base pairs is: base pairs ¼ 3274 þ2413 � ([T/S � 0.0545]/1.16).
2.2.2. InflammationAfter fasting blood samples were kept frozen at �80 �C, circu-
lating plasma levels of interleukin-6 (IL-6, N ¼ 2908), c-reactiveprotein (CRP, N ¼ 2909), and tumor necrosis factor-alpha (TNF-a,N ¼ 2889) were assessed in duplicate (Vogelzangs et al., 2012).High-sensitivity plasma levels of CRP were measured in duplicateby an in-house enzyme-linked immunosorbent assay (ELISA) basedon purified protein and polyclonal anti-CRP antibodies (Dako,Glostrup, Denmark). Plasma IL-6 levels were measured in duplicateby a high sensitivity ELISA (PeliKine Compact ELISA, Sanquin,Amsterdam, The Netherlands). Plasma TNF-a levels were assayedusing a high-sensitivity solid phase ELISA (Quantikine HS HumanTNF-a Immunoassay, R&D systems, Minneapolis, MN, USA). Intra-and inter-assay coefficients of variation for inflammatory markerswere between 5%e10% and 10%e15%, respectively.
2.2.3. HPA-axis functionRespondentswere instructed to collect saliva samples at homeon
a regular (preferably working) day (Vreeburg et al., 2009b). This hasshown a reliable and minimally intrusive method to assess theactive, unbound form of cortisol (Kirschbaum and Hellhammer,1994). Instructions concerning saliva sampling prohibited eating,smoking, drinking tea or coffee, or brushing teethwithin 15minutesbefore sampling. Furthermore, no dental work 24 hours beforesampling was allowed. In total, 2204 subjects returned at least1-saliva sample and themedian timebetweenbloodwithdrawal andsaliva sampling was 8.0 days (25th to 75th percentile: 4e21). Salivasamples were obtained using Salivettes (Sarstedt, Nümbrecht, Ger-many) at 7 time points on a regular (work) day: at awakening (T1)and 30 (T2), 45 (T3), and 60 (T4)minutes later, and at 10 PM (T5) and11 PM (T6). In addition, dexamethasone suppression was measuredby cortisol sampling the next morning at awakening (T7) afteringestionof 0.5mgdexamethasonedirectlyafter the saliva sample at23:00 hours (T6). Respondents were instructed to write down theexact sampling times. The values of morning cortisol samples wereassigned missing when collected outside of a margin of 5 minutesbefore or after the time protocol. Samples were stored in re-frigerators and returned by regular mail. After receipt, Salivetteswere centrifuged at 2000� g for 10 minutes, aliquoted and storedat �80 �C. Cortisol analysis was performed by competitive electro-chemiluminescence immunoassay (Roche, Basel, Switzerland) (vanAken et al., 2003). The functional detection limit was 2.0 nmol/literand the intra- and inter-assay variability coefficients were <10%.
Various cortisol measures were used to characterize differentHPA-axis aspects. First, to indicate the 1-hour cortisol awakeningresponse we calculated the area under the curvewith respect to theincrease (AUCi) and ground (AUCg, N ¼ 1753) (Pruessner et al.,
2003). The AUCg is an estimate of the total cortisol secretion overthe first hour after awakening, whereas the AUCi is a measure of thedynamic of the cortisol awakening response, more indicative of thesensitivity of the system, emphasizing changes over time(Pruessner et al., 2003). Second, evening cortisol levels (N ¼ 1905)are indicative of basal HPA-axis activity, because cortisol levels aregenerally low at the end of the day. Since the 2 evening values werehighly correlated (r¼ 0.75, p < 0.001), the meanwas used to reflectevening cortisol (N ¼ 1905). At last, the cortisol suppression ratioafter dexamethasone intake is calculated by dividing the cortisolvalue at T1 by cortisol value at T7 (N ¼ 1806). This ratio examinesthe adequacy of the negative feedback of the HPA-axis (Carroll et al.,1981). A higher ratio indicates suppression by dexamethasone,which occurs when the feedback loop functions adequately, a lowerratio indicates non-suppression of the HPA-axis.
2.2.4. ANSDuring the baseline interview, subjects were wearing the VU
Universityambulatorymonitoring system(Lichtet al., 2012). This is alight-weight, unobtrusive device that records the electrocardiogram(ECG) and changes in thorax impedance (dZ) from 6 surface elec-trodesplaced at the chest andon the backof the subjects (Willemsenet al., 1996). The inter-beat interval time series were extracted fromthe ECG signal to obtain heart rate (HR, N ¼ 2819), an indicator ofcombined sympathetic and parasympathetic nervous system activ-ity. To separately index the cardiac effects of bothANSbranches, pre-ejection period (PEP, N ¼ 2796) and respiratory sinus arrhythmia(RSA, N ¼ 2820) were extracted from the combined dZ and ECGsignals (Licht et al., 2010). We refer to PEP as a measure of cardiacsympathetic control, as it can reliably index b-adrenergic inotropicdrive to the left ventricle (long PEP reflecting low cardiac sympa-thetic control) andwas defined as the interval from the beginning ofthe left ventricular electrical activity (ECG Q-wave onset), to thebeginning of left ventricular ejection (B point in the dZ/dt signal)(Berntson et al., 1994; Cacioppo et al., 1994). On the other hand, RSAreflects cardiac parasympathetic (vagal) control (high RSA reflectinghigh cardiac vagal control), and was obtained by subtracting theshortest inter-beat interval during heart rate acceleration in theinspirational phase from the longest inter-beat interval duringdeceleration in the expirational phase for all breaths as describedelsewhere (Task Force of the European Society of Cardiology theNorth American Society of Pacing Electrophysiology, 1996). Move-ment registration through vertical accelerometrywas used to exciseperiodswhere subjects were nonstationary (w15minutes), becausepostural changes have an effect on RSA and PEP that is partly unre-lated to changes in autonomic activity (Houtveen et al., 2005).Automated scoringof RSA and PEPwas checkedby visual inspection,and valid datawere averaged over 98.0� 24minutes time to create asingle PEP, RSA, and HR value.
2.3. Covariates
Sociodemographic factors included sex, age, andyears of attainededucation. Lifestyle variables included alcohol consumption (nodrinker, mild-moderate drinker 1e14 (women)/1e21 drinks perweek (men), heavy drinker >14 (women)/>21 (men) drinks perweek (Stuurgroep Multidisciplinaire Richtlijnontwikkeling, 2009)),smoking (never, former, current), andphysical activity (InternationalPhysicalActivityQuestionnaire [Craiget al., 2003], expressed in1000metabolic equivalent minutes in the past week). Health indicatorsincluded bodymass index (BMI: underweight<18.5, normalweight18.5e24.9, overweight 25e29.9, andobesity�30) and thenumberofself-reported chronic diseases for which medical treatment wasreceived (including cardiovascular diseases, diabetes, lung disease,osteoarthritis, rheumatic disease, cancer, ulcer, intestinal problem,
D. Révész et al. / Neurobiology of Aging 35 (2014) 1422e14301424

Author's personal copy
liver disease, epilepsy, and thyroid gland disease). Frequent medi-cation use (>50% of the time) was registered from the participant’smedication containers and classified using the World Health Orga-nization anatomic therapeutic chemical classification: cardiacmedication including beta-blocking agents (C07) and others such asantihypertensives (C01eC05, C08, C09), anti-inflammatory medi-cation (M01A, M01B, A07EB, A07EC), and antidepressants (N06AA,N06AB, N06AF, N06AG, N06AX).
Apart from the standard covariates mentioned previously, forthe HPA-axis analyses, sampling factors were additionally included,as suggested in a previous study (Vreeburg et al., 2009b): subjectsreported awakening time, working status on the sampling day, andseason was categorized into dark months (OctobereFebruary) andmonths with more daylight (MarcheSeptember). Menstrual cyclephase, menopausal status, the use of oral contraceptives (Vreeburget al., 2009b), or estrogen replacement therapy (data not shown)were not associated with salivary cortisol within this sample andwere therefore not included as covariates. For ANS analyses addi-tional adjustments weremade for respiratory rate (for RSA), and formean arterial pressure (systolic blood pressure þ 2 � diastolicblood pressure/3) to account for potential between-subject differ-ences in afterload in PEP analyses (Houtveen et al., 2005). There-fore, blood pressurewas recorded in a supine position by 2 repeatedmeasurements using the OMRON M4 IntelliSense (HEM-752A,Omron Healthcare, Bannockburn, IL, USA).
2.4. Statistical analyses
Baseline characteristics were compared across LTL (short, mid-dle, and long) tertiles using c2 and analysis of variance statistics todescribe the frequencies, means, and standard deviations. Thelevels of CRP, IL-6, TNF-a, mean evening cortisol, cortisol suppres-sion ratio, and RSA were not normally distributed. Therefore, nat-ural logarithm-transformations were used in analyses and thesevalues were presented back-transformed in tables. Pearson corre-lations were used to determine correlations between LTL and allstress system components. The associations between LTL and eachof the inflammatory, HPA-axis, and ANS markers (used both as acontinuous measure and categorized as the high-risk tertiles vs. thecombined middle- and low-risk tertiles) were tested in separatelinear regression analyses. All analyses were adjusted for socio-demographic, lifestyle, health indicators, and medication. Analyseson HPA-axis and ANS were additionally adjusted for a specific set ofcovariates as described in section 2.3.
In subsequent sensitivity analyses we examined whether cur-rent depression and/or anxiety diagnosis (current, remitted, or no),or antidepressant use (yes or no) modified the associations be-tween LTL and these stress systems, by checking whether theinteraction terms were significantly associated with LTL, and byadding these factors as covariates.
At last, to examine the joint impact of all significant physiolog-ical stress system mechanisms, a cumulative score was calculatedfrom all high-risk tertiles of the stress markers that showed sig-nificant associations with LTL in the a priori hypothesized di-rections. Fully corrected analyses of covariance were performed tocompare themean LTL of personswith gradually increasing numberof stress system elevations. All analyses were conducted using SPSSversion 20.0 (IBM Corp, Armonk, NY, USA).
3. Results
Table 1 shows the sample characteristics across LTL tertiles. Onaverage, subjects were 41.8 years (standard deviation; SD ¼ 13.1),66.4% were women, and had 12.2 years of education (SD ¼ 3.3). Theaverage T to S ratio was 1.11 (SD ¼ 0.30), corresponding to 5468
base pairs (bp; SD ¼ 617). Unadjusted linear regression showed LTLto be 14.5 bp shorter for each increasing year of age (standarderror ¼ 0.8; p < 0.001). Subjects in the shortest LTL tertile wereolder, more often male, higher in BMI, more often smokers andheavy drinkers, had more chronic diseases, used more cardiacand anti-inflammatory medication, and more often had their salivasampling on nonworking days in the months with more daylight.In subjects with the shortest LTL, higher levels of CRP, IL-6, and TNF-a were found, together with lower RSA and longer PEP. No differ-ences were found for the HPA-axis measures between the 3 LTLtertiles.
Unadjusted Pearson correlations between LTL and the individualstress system markers are shown in Table 2. Within the 3 stresssystems, stress markers were highly inter-correlated. LTL wassignificantly correlated with all 3 inflammatory markers, eveningcortisol levels, PEP, and RSA.
All inflammation markers were significantly associated with LTLafter adjustment for sociodemographics (Table 3). After fulladjustment for additional health and lifestyle factors, high levels ofCRP, and IL-6 remained significantly associated with LTL, but TNF-awas only borderline significant. Consistent with this, subjects withthe highest tertiles of CRP (>2.18 mg/L) and IL-6 (>1.04 pg/mL)showed shorter LTL, which was not confirmed for TNF-a.
Of the HPA-axis measures, no significant linear associationswere found with LTL. When high-risk tertiles were defined, how-ever, the highest AUCi tertile (>4.1 nmol/L/hr) was significantlyassociated with short LTL after sociodemographics adjustment andafter full adjustment, suggesting a nonlinear association. AUCg,evening cortisol levels and the cortisol suppression ratios were notassociated with LTL.
Regarding ANS indicators, contrary to our prior expectation,long PEP remained significantly associated with short LTL after fulladjustment, whereas only trends remained present for the associ-ations between RSA and LTL and between HR and LTL. However,shorter LTL was significantly associated to the highest tertiles of HR(>75.4 b/min) and PEP (>129.3 ms) compared with those in thelower tertiles.
Previously, it has been described in our sample that depressiveand anxiety disorders were associated with increased CRP and IL-6levels (Vogelzangs et al., 2012, 2013), increased AUCg and AUCi(Vreeburg et al., 2009a), but not to other stress system indicators(Licht et al., 2012). However, we found no significant interactionsbetween stress systemmarkers and current or remitted depressionand/or anxiety diagnosis (1707 current vs. 603 remitted vs. 626healthy controls) or antidepressant use (721 users vs. 2202 non-users) in the association with LTL (all p-values for interaction termswere >0.10, data not shown), and additional adjustment for psy-chopathology and antidepressant use resulted in similar associa-tions. Furthermore, we did not find significant sex interactionsbetween LTL and any of the stress system markers (all p-values forLTL-by-sex interaction terms were >0.10, data not shown).
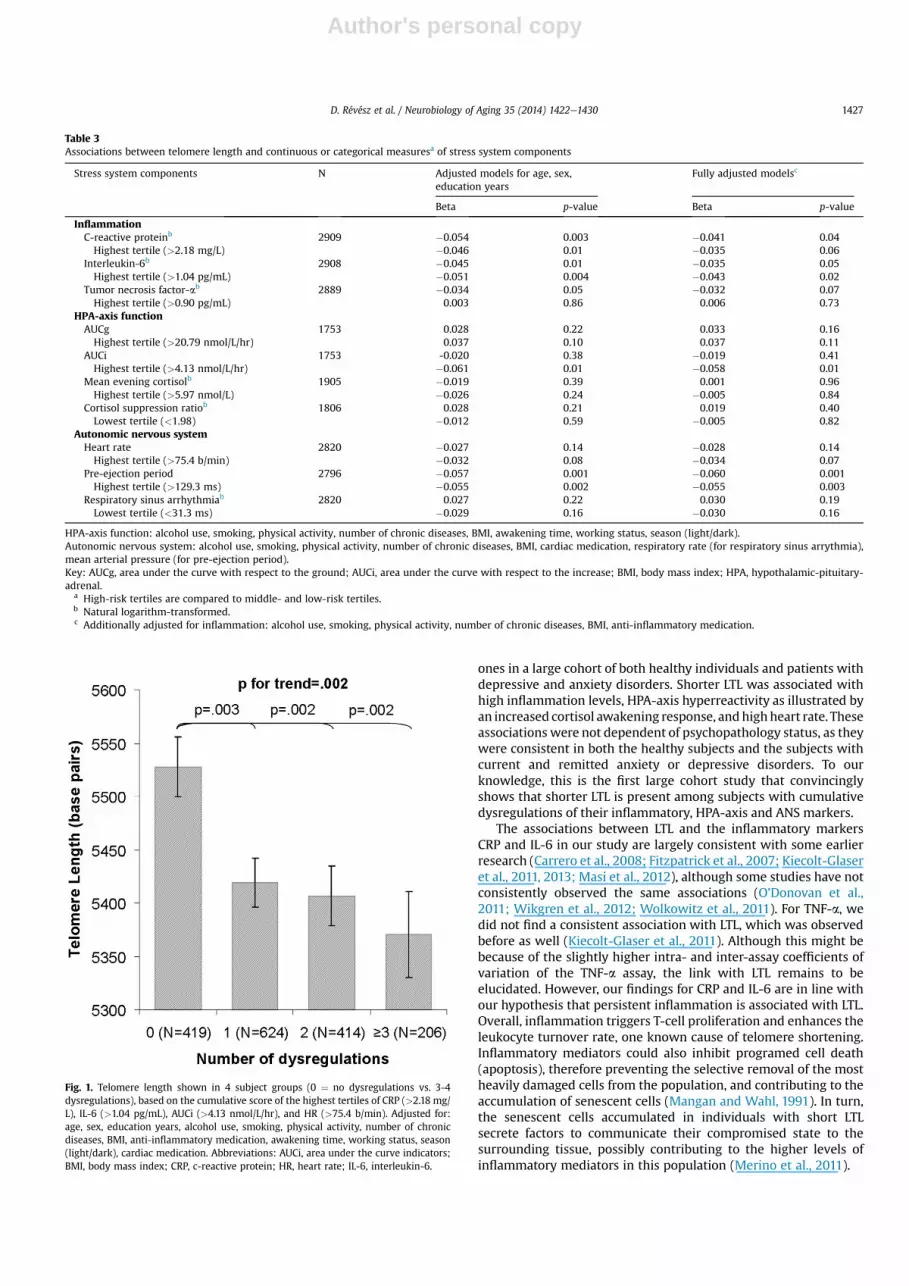
Subsequently, for the 4 stress system indicators that showed theexpected relation with LTL, and were significant at least at p < 0.10,we examined the cumulative index score. This score was defined asthe number of highest tertiles on CRP, IL-6, AUCi, and HR, for sub-jects with complete data on all significant stress indicators (N ¼1663). PEP was not included in the cumulative index score becauseof the unexpected direction of the associationwith LTL. Fig. 1 showsthe LTL of subject groups with various physiological risk profiles.Compared with having no dysregulations (LTL ¼ 5528 bp), eachadditional high-risk stress marker was significantly associated withshorter LTL after full adjustment for relevant confounders (1 dys-regulation: 5419 bp; p ¼ 0.003; 2 dysregulations: 5407 bp; p ¼0.002; 3 or 4 dysregulations: 5371 bp; p¼ 0.002). As the shorteningrate found in other studies was between 14e20 bp per year
D. Révész et al. / Neurobiology of Aging 35 (2014) 1422e1430 1425

Author's personal copy
(Cawthon et al., 2003; Hartmann et al., 2010; Wikgren et al., 2012),in line with our found shortening rate of 14.5 bp per year, thesedifferences correspond to an accelerated “biological aging” of5e8 years, 6e9 years, and 8e11 years, respectively.
4. Discussion
This study found that subjectswithmore activated stress systemshad shorter leukocyte telomere length than thosewith less activated
Table 1Sample characteristics shown per telomere length tertile
Sample characteristics Telomere length (T/S ratio) tertiles (N ¼ 2936) p-value
Short (N ¼ 978) Middle (N ¼ 979) Long (N ¼ 979)
T/S ratio, mean (range) 0.83 (0.33e0.95) 1.06 (0.95e1.17) 1.45 (1.18e2.03) d
TL (bp), mean (range) 4877 (3851e5135) 5357 (5135e5604) 6168 (5605e7392) d
DemographicsAge (y), mean (SD) 46.5 (11.9) 41.9 (12.8) 37.0 (12.8) <0.001Sex (female), n (%) 582 (19.8) 672 (22.9) 696 (23.7) <0.001Years of education, mean (SD) 12.1 (3.4) 12.2 (3.3) 12.2 (3.2) 0.74
Lifestyle and health factorsBMI (kg/m2), n (%)
Underweight 9 (0.3) 23 (0.8) 34 (1.2) <0.001Normal weight 429 (14.6) 517 (17.6) 540 (18.4)Overweight 347 (11.8) 287 (9.8) 258 (8.8)Obese 193 (6.6) 152 (5.2) 147 (5.0)
Smoking status, n (%)Never 227 (7.7) 276 (9.4) 323 (11.0) <0.001Former 355 (12.1) 315 (10.7) 304 (10.4)Current 396 (13.5) 388 (13.2) 352 (12.0)
Drinking behavior, n (%)Non-drinker 163 (5.6) 175 (6.0) 162 (5.5) 0.004Mild-moderate drinker 663 (22.6) 680 (23.2) 720 (24.5)Heavy drinker 152 (5.2) 124 (4.2) 97 (3.3)
Physical activity (*1000 MET-min/wk), median (IQR) 2.8 (3.3) 2.8 (3.4) 2.8 (3.4) 0.11Number of chronic diseases, mean (SD) 0.7 (1.0) 0.6 (0.9) 0.5 (0.8) <0.001Cardiac medication, n (%) 101 (3.4) 80 (2.7) 47 (1.6) <0.001Anti-inflammatory medication, n (%) 58 (2.0) 39 (1.3) 28 (1.0) 0.003Mean awakening time, mean (SD) 7:26 (1:00) 7:29 (1:00) 7:28 (1:00) 0.69Season light versus dark, n (%) 733 (25.0) 704 (24.0) 686 (23.4) 0.05Working day on saliva collection, n (%) 666 (22.7) 705 (24.0) 750 (25.5) <0.001
Physiological stress systems N
Inflammation, mean (SD)C-reactive protein (mg/L)a 2909 1.39 (3.3) 1.26 (3.5) 1.19 (3.6) 0.02Interleukin-6 (pg/mL)a 2908 0.82 (2.5) 0.79 (2.7) 0.67 (2.7) <0.001Tumor necrosis factor-alpha (pg/mL)a 2889 0.84 (1.8) 0.86 (1.9) 0.80 (1.9) 0.04
HPA-axis function, mean (SD)AUCg (nmol/L/hr) 1753 19.0 (7.0) 18.8 (6.6) 18.8 (7.4) 0.80AUCi (nmol/L/hr) 1753 2.4 (6.3) 2.1 (6.1) 2.0 (6.4) 0.45Mean evening cortisol (nmol/L)a 1905 4.78 (1.7) 4.74 (1.8) 4.53 (1.7) 0.16Cortisol suppression ratioa 1806 2.31 (1.7) 2.39 (1.6) 2.47 (1.7) 0.09
Autonomic nervous system, mean (SD)Heart rate (bpm) 2819 71.6 (9.7) 71.9 (10.0) 72.3 (9.1) 0.27Pre-ejection period (ms) 2796 121.9 (18.5) 119.9 (17.5) 118.8 (17.4) 0.001Respiratory sinus arrhythmia (ms)a 2820 33.5 (1.7) 38.8 (1.7) 43.5 (1.7) <0.001
Key: AUCg, area under the curve with respect to the ground; AUCi, area under the curve with respect to the increase; BMI, body mass index; M, mean; MET, metabolicequivalent; SD, standard deviation; TL, telomere length in base pairs; T/S, telomere-to-single copy gene ratio.
a Natural logarithm-transformed factors presented back-transformed.
Table 2Unadjusted Pearson correlations between telomere length and the stress system components of inflammation, HPA-axis and ANS
Markers TL CRP IL-6 TNF-a AUCg AUCi Evening Cortisol suppression HR PEP RSA
TL 1 �0.072b �0.094b �0.057b �0.007 �0.005 �0.058b 0.045 0.026 �0.055b 0.201b
CRPa d 1 0.312b 0.135b �0.020 �0.033 0.033 0.013 0.204b �0.118b 0.151b
IL-6a d d 1 0.125b �0.016 0.004 0.028 �0.042 0.108b �0.067b �0.175b
TNF-aa d d d 1 �0.019 0.020 0.005 �0.023 0.012 0.005 �0.074b
AUCg d d d d 1 0.473b 0.365b 0.187b 0.024 �0.008 �0.091b
AUCi d d d d d 1 0.085b �0.263b 0.042 �0.014 0.015Evening cortisola d d d d d d 1 �0.234b �0.001 0.022 �0.102b
Cortisol suppression ratioa d d d d d d d 1 0.041 �0.040 0.030HR d d d d d d d d 1 �0.249b �0.342b
PEP d d d d d d d d d 1 0.125b
RSAa d d d d d d d d d d 1
Key: ANS, autonomic nervous system; AUCg, area under the curve with respect to the ground; AUCi, area under the curve with respect to the increase; CRP, c-reactive protein;HPA, hypothalamic-pituitary-adrenal; HR, heart rate; IL-6, interleukine-6; PEP, pre-ejection period; RSA, respiratory sinus arrhythmia; TL, telomere length; TNF-a, tumornecrosis factor-a.
a Natural logarithm-transformed factors presented back-transformed.b p < 0.05, 2-tailed.
D. Révész et al. / Neurobiology of Aging 35 (2014) 1422e14301426
Author's personal copy
ones in a large cohort of both healthy individuals and patients withdepressive and anxiety disorders. Shorter LTL was associated withhigh inflammation levels, HPA-axis hyperreactivity as illustrated byan increased cortisol awakening response, andhighheart rate. Theseassociations were not dependent of psychopathology status, as theywere consistent in both the healthy subjects and the subjects withcurrent and remitted anxiety or depressive disorders. To ourknowledge, this is the first large cohort study that convincinglyshows that shorter LTL is present among subjects with cumulativedysregulations of their inflammatory, HPA-axis and ANS markers.
The associations between LTL and the inflammatory markersCRP and IL-6 in our study are largely consistent with some earlierresearch (Carrero et al., 2008; Fitzpatrick et al., 2007; Kiecolt-Glaseret al., 2011, 2013; Masi et al., 2012), although some studies have notconsistently observed the same associations (O’Donovan et al.,2011; Wikgren et al., 2012; Wolkowitz et al., 2011). For TNF-a, wedid not find a consistent association with LTL, which was observedbefore as well (Kiecolt-Glaser et al., 2011). Although this might bebecause of the slightly higher intra- and inter-assay coefficients ofvariation of the TNF-a assay, the link with LTL remains to beelucidated. However, our findings for CRP and IL-6 are in line withour hypothesis that persistent inflammation is associated with LTL.Overall, inflammation triggers T-cell proliferation and enhances theleukocyte turnover rate, one known cause of telomere shortening.Inflammatory mediators could also inhibit programed cell death(apoptosis), therefore preventing the selective removal of the mostheavily damaged cells from the population, and contributing to theaccumulation of senescent cells (Mangan and Wahl, 1991). In turn,the senescent cells accumulated in individuals with short LTLsecrete factors to communicate their compromised state to thesurrounding tissue, possibly contributing to the higher levels ofinflammatory mediators in this population (Merino et al., 2011).
Table 3Associations between telomere length and continuous or categorical measuresa of stress system components
Stress system components N Adjusted models for age, sex,education years
Fully adjusted modelsc
Beta p-value Beta p-value
InflammationC-reactive proteinb 2909 �0.054 0.003 �0.041 0.04Highest tertile (>2.18 mg/L) �0.046 0.01 �0.035 0.06
Interleukin-6b 2908 �0.045 0.01 �0.035 0.05Highest tertile (>1.04 pg/mL) �0.051 0.004 �0.043 0.02
Tumor necrosis factor-ab 2889 �0.034 0.05 �0.032 0.07Highest tertile (>0.90 pg/mL) 0.003 0.86 0.006 0.73
HPA-axis functionAUCg 1753 0.028 0.22 0.033 0.16Highest tertile (>20.79 nmol/L/hr) 0.037 0.10 0.037 0.11
AUCi 1753 -0.020 0.38 �0.019 0.41Highest tertile (>4.13 nmol/L/hr) �0.061 0.01 �0.058 0.01
Mean evening cortisolb 1905 �0.019 0.39 0.001 0.96Highest tertile (>5.97 nmol/L) �0.026 0.24 �0.005 0.84
Cortisol suppression ratiob 1806 0.028 0.21 0.019 0.40Lowest tertile (<1.98) �0.012 0.59 �0.005 0.82
Autonomic nervous systemHeart rate 2820 �0.027 0.14 �0.028 0.14Highest tertile (>75.4 b/min) �0.032 0.08 �0.034 0.07
Pre-ejection period 2796 �0.057 0.001 �0.060 0.001Highest tertile (>129.3 ms) �0.055 0.002 �0.055 0.003
Respiratory sinus arrhythmiab 2820 0.027 0.22 0.030 0.19Lowest tertile (<31.3 ms) �0.029 0.16 �0.030 0.16
HPA-axis function: alcohol use, smoking, physical activity, number of chronic diseases, BMI, awakening time, working status, season (light/dark).Autonomic nervous system: alcohol use, smoking, physical activity, number of chronic diseases, BMI, cardiac medication, respiratory rate (for respiratory sinus arrythmia),mean arterial pressure (for pre-ejection period).Key: AUCg, area under the curve with respect to the ground; AUCi, area under the curve with respect to the increase; BMI, body mass index; HPA, hypothalamic-pituitary-adrenal.
a High-risk tertiles are compared to middle- and low-risk tertiles.b Natural logarithm-transformed.c Additionally adjusted for inflammation: alcohol use, smoking, physical activity, number of chronic diseases, BMI, anti-inflammatory medication.
Fig. 1. Telomere length shown in 4 subject groups (0 ¼ no dysregulations vs. 3-4dysregulations), based on the cumulative score of the highest tertiles of CRP (>2.18 mg/L), IL-6 (>1.04 pg/mL), AUCi (>4.13 nmol/L/hr), and HR (>75.4 b/min). Adjusted for:age, sex, education years, alcohol use, smoking, physical activity, number of chronicdiseases, BMI, anti-inflammatory medication, awakening time, working status, season(light/dark), cardiac medication. Abbreviations: AUCi, area under the curve indicators;BMI, body mass index; CRP, c-reactive protein; HR, heart rate; IL-6, interleukin-6.
D. Révész et al. / Neurobiology of Aging 35 (2014) 1422e1430 1427

Author's personal copy
Wehypothesized that LTL would be shorter in subjects with highsympathetic and low parasympathetic activity, with long PEPreflecting low sympathetic activation, and high RSA reflecting highparasympathetic activation. As the combination of low para-sympathetic and high sympathetic activation would lead to ahigher HR, we further hypothesized that HR would be associatedwith LTL. The latter association was borne out by the results. Thissupports earlier observations that higher resting HR is stronglycorrelated to shorter lifespan, possibly because of higher basalmetabolic rate, increased oxidative stress, and accelerated deteri-oration of the cardiovascular system (Zhang and Zhang, 2009). LTLmay also have an impact on cardiovascular function, as functionaltelomeres are required for viable cardiovascular cells in vitro andfor cardiomyocyte renewal (Fuster and Andres, 2006; Kajstura et al.,2006). The extent to which LTL reflects telomeres in heart tissueremains largely unclear, although a high correlation betweenleukocyte and aortic wall tissue telomere length has been reported(Wilson et al., 2008). Furthermore, low RSA was associated withshorter LTL, as expected, although adjustment for sociodemo-graphics and lifestyle factors rendered the association nonsignifi-cant. However, we unexpectedly found that long PEP wassignificantly associated with shorter LTL. Two earlier relativelysmall-sampled studies investigated the link between ANS and TL,but also did not find any associations between TL and basal ANSmeasures (Epel et al., 2006; Kroenke et al., 2011). Rather they foundan association between lower parasympathetic activation duringstress testing and low telomerase activity (Epel et al., 2006), andbetween sympathetic activation during stress testing and shortbuccal cell telomere length (Kroenke et al., 2011). What could havedriven the anomalous finding of long PEP being associated withshort LTL? It is unlikely that this finding is because of an error in PEPassessment, since we haveein line with hypotheseseconfirmedthat short PEP in our sample is related to health factors such as BMI,antidepressant use (Licht et al., 2012), and with the number ofbaseline metabolic syndrome components (Licht et al., 2010), aswell as the increase of metabolic abnormalities over time (Lichtet al., 2013). An intriguing possibility is that chronic stress withconcurrent high levels of plasma (nor)epinephrine may have led toa down regulation of the cardiac beta-receptors. This would lead toa prolongation of the PEP, even in the presence of high cardiacsympathetic nervous system activity. Desensitization of beta-adrenergic receptors has earlier been associated with anxiousmood (Yu et al., 2008) and caregiving status (Mills et al., 2004).Studies that measure both PEP and cardiac beta-receptor status areneeded to test this speculative idea.
In HPA-axis analyses, we found an association between LTL andAUCi, a measure of HPA-axis sensitivity, but not for AUCg, cortisolevening levels, or cortisol suppression ratio. Most of the earlierstudies have shown significant associations between cortisol levelsand LTL (Epel et al., 2006; Kroenke et al., 2011; Parks et al., 2009;Tomiyama et al., 2012; Wikgren et al., 2012). However, comparingfindings directly is limited since these studies have either measuredHPA-axis function in 12-hours nocturnal urine (Epel et al., 2006;Tomiyama et al., 2012), first morning urine (Parks et al., 2009), orpre-post test salivary samples (Wikgren et al., 2012). Tomiyamaet al. (2012), have measured salivary cortisol awakening responsein a small post-menopausal sample, and have not shown significantassociations between AUCi andor AUCg and LTL, but thesemeasureswere calculated by subtracting the wakeup cortisol value from thewakeup þ30-minutes value. Our findings are not in line withWikgren et al. (2012) who linked short LTL with low responsivenessto a dexamethasone challenge, for which the dexamethasoneintake was lower, compared with our study (3.5 mg per kg of bodyweight vs. 500 mg in our study). The found association between thehighest tertile of AUCi with shorter LTL in our sample suggests that
cellular aging is especially linked to hyperactivity of the HPA-axis,and less with the more basal activity of the HPA-axis (as assessedwith AUCg or evening levels). In our prior work, we have alsoshown that especially the cortisol awakening response, but not theevening levels or dexamethasone stress test levels were associatedwith stress-related psychiatric disorders (Vreeburg et al., 2009a,2010). Such hyperactivity of the HPA-axis has shown to result inreduced telomerase activity in vitro and higher levels of oxidativestress, with subsequent shortening of LTL (Aschbacher et al., 2013;Choi et al., 2008). Shorter LTL might also lead to higher cortisollevels, possibly through inflammation and oxidative stress, but toour knowledge, this direction is not explored extensively yet.
Within our study, associations between shorter LTL and separatestress systemdysregulations were independent of psychopathologystatus or antidepressant medication use. To our knowledge, onlyfew studies have examined these associations within a psychiatricpopulation (Kiecolt-Glaser et al., 2011; Wikgren et al., 2012;Wolkowitz et al., 2011). Two of these studies have shown a differ-ential pattern of associations between LTL and IL-6 (Wolkowitzet al., 2011) and HPA-axis indicators (Wikgren et al., 2012) indepressed patients and healthy controls. The reason for these dif-ferences is unclear. Overall, our findings support the notion thatshorter LTL is associated with physiological stress markers, inde-pendently of psychopathology status. In recent analyses performedwithin our Netherlands Study of Depression and Anxiety sample,depressed patients were found to have shorter LTL compared withhealthy control subjects (Verhoeven et al., 2013). The lack of asignificant interaction within the present study suggests that thereis not merely a link between stress and shortened telomeres viadepression, but that other mechanistic pathways are likely to beinvolved as well.
Lastly, we analyzed whether the combination of high-impactphysiological stress system dysregulations added up to the short-ening of telomeres. Until now, the 3 main stress systems have beenmainly investigated in separate analysis, although 2 studiesexamined 2 combinations of these systems (Epel et al., 2006;Kroenke et al., 2011). We confirm that the simultaneous dysre-gulation of various inflammatory, HPA-axis and cardiac ANS factorsform a cumulative impact on cellular aging. Although these resultsneed further replication, they imply that multiple stress systemdysregulations may interact in creating an “allostatic load”. As theterm allostasis refers to the process whereby physiological stabilityis maintained, allostatic load represents the “wear and tear” in thebody during stressful events (McEwen, 2008). It still remains to beelucidated whether these (cumulative) dysregulations actuallylead to overall health decline, although 2 recent review paperssummarized the studies that indicate the importance of physio-logical stress systems dysregulations for numerous somatic healthoutcomes, such as mortality, cognitive decline, physical func-tioning, and cardiovascular health (Juster et al., 2010; Penninxet al., 2013).
Some limitations of this large cohort study should be taken intoaccount. Firstly, this is a cross-sectional study, therefore notcapturing the high variability of physiological stress markers, andnot fully clarifying the causal paths. Short LTL might be a directconsequence of stress physiology, but stress system dysregulationsmay also be the result of other underlying processes indexed byeorpassing througheLTL shortening. Future studies should investigatethese associations longitudinally to explain the causality. Secondly,no informationwas available on telomerase activity to shed light onthe maintenance mechanism of telomeres. Thirdly, the compliancewith saliva sampling might have been inaccurate, and althoughhardly feasible in large cohorts, multiple sampling days would haveenhanced the HPA function assays. On the other hand, the strengthsof this study were its large sample size, enabling us to adjust for
D. Révész et al. / Neurobiology of Aging 35 (2014) 1422e14301428

Author's personal copy
various lifestyle and health variables in a large adult age range.Furthermore, although circulating leukocytes might not directlyreflect the cellular aging process in all parts of the body, high cor-relations between LTL and telomere length in other body systems(Daniali et al., 2013) and in specific lymphocyte subsets have beenshown (Lin et al., 2010). This is, to our knowledge, the first study tolink LTL, measured with a frequently used well-validated method,with themajor physiological stress systems, and to shed light on thecumulative effect of multiple stress markers on LTL.
In summary, we found that subjects with shorter telomeres arecharacterized by a pro-inflammatory state, a hyperactive HPA-axis,and high heart rate. In addition, cumulative stress system dysre-gulations resulted in overall shorter LTL. These data clearly indicatethat dysregulation of physiological stress systems and cellular agingprocesses are intertwined, and may influence each other bi-directionally. This could directly explain why the exposure tochronic stressors and stress-related conditions has shown to impacton a multitude of aging-related conditions ranging from heartdisease, diabetes, obesity, cognitive and physical impairment toeven cancer (Penninx et al., 2013), and establishes a general un-derlying biological basis for the important psyche-soma interaction.
Disclosure statement
All authors declare that they have no conflicts of interest.
Acknowledgements
The infrastructure for the Netherlands Study of Depression andAnxiety study (www.nesda.nl) is funded through the Geestkrachtprogram of the Netherlands Organization for Health Research andDevelopment (ZonMW, grant number: 10-000e1002) and is sup-ported by participating universities and mental health care orga-nizations (VU University Medical Center, GGZ inGeest, Arkin, LeidenUniversity Medical Center, GGZ Rivierduinen, University MedicalCenter Groningen, Lentis, GGZ Friesland, GGZ Drenthe, IQ Health-care, Netherlands Institute for Health Services Research andNetherlands Institute of Mental Health and Addiction (Trimbos).Brenda Penninx, Dóra Révésza, Josine E. Verhoeven, and telomerelength assaying were supported through an NWO-VICI grantnumber: 91811602. Owen M. Wolkowitzc is on the ScientificAdvisory Board of Telomere Diagnostics, Inc. (TDx).
References
Aschbacher, K., O’Donovan, A., Wolkowitz, O., Dhabhar, F., Su, y., Epel, E., 2013. Goodstress, bad stress and oxidative stress: insights from anticipatory cortisolreactivity. Psychoneuroendocrinology 38, 1698e1708.
Aubert, G., Lansdorp, P.M., 2008. Telomeres and aging. Physiol. Rev. 88, 557e579.Aviv, A., Hunt, S.C., Lin, J., Cao, X., Kimura, M., Blackburn, E., 2011. Impartial
comparative analysis of measurement of leukocyte telomere length/DNA con-tent by Southern blots and qPCR. Nucleic Acids Res. 39, e134.
Berntson, G.G., Cacioppo, J.T., Binkley, P.F., Uchino, B.N., Quigley, K.S., Fieldstone, A.,1994. Autonomic cardiac control III. Psychological stress and cardiac response inautonomic space as revealed by pharmacological blockades. Psychophysiology31, 599e608.
Blackburn, E.H., 1991. Structure and function of telomeres. Nature 350, 569e573.Blackburn, E.H., 2001. Switching and signaling at the telomere. Cell 106, 661e673.Broer, L., Codd, V., Nyholt, D., Deelen, J., Mangino, M., Willemsen, G., Albrecht, E.,
Amin, N., Beekman, M., de Geus, E., Henders, A., Nelson, C., Steves, C.,Wright, M., de Craen, A., Isaacs, A., Matthews, M., Moayyeri, A., Montgomery, G.,Oostra, B., Vink, J., Spector, T., Slagboom, P., Martin, N., Samani, N., van Duijn, C.,Boomsma, D., 2013. Meta-analysis of telomere length in 19 713 subjects revealshigh heritability, stronger maternal inheritance and a paternal age effect. Eur. J.Hum. Genet. 21, 1163e1168.
Cacioppo, J.T., Berntson, G.G., Binkley, P.F., Quigley, K.S., Uchino, B.N., Fieldstone, A.,1994. Autonomic cardiac control II. Noninvasive indices and basal response asrevealed by autonomic blockades. Psychophysiology 31, 586e598.
Carrero, J.J., Stenvinkel, P., Fellstrom, B., Qureshi, A.R., Lamb, K., Heimburger, O.,Barany, P., Radhakrishnan, K., Lindholm, B., Soveri, I., Nordfors, L., Shiels, P.G.,2008. Telomere attrition is associated with inflammation, low fetuin-A levels
and high mortality in prevalent haemodialysis patients. J. Intern. Med. 263,302e312.
Carroll, B.J., Feinberg, M., Greden, J.F., Tarika, J., Albala, A.A., Haskett, R.F.,James, N.M., Kronfol, Z., Lohr, N., Steiner, M., de Vigne, J.P., Young, E., 1981.A specific laboratory test for the diagnosis of melancholia. Standardization,validation, and clinical utility. Arch. Gen. Psychiatry 38, 15e22.
Cawthon, R.M., 2002. Telomere measurement by quantitative PCR. Nucleic AcidsRes. 30, e47.
Cawthon, R.M., Smith, K.R., O’Brien, E., Sivatchenko, A., Kerber, R.A., 2003. Associ-ation between telomere length in blood and mortality in people aged 60 yearsor older. Lancet 361, 393e395.
Choi, J., Fauce, S.R., Effros, R.B., 2008. Reduced telomerase activity in human Tlymphocytes exposed to cortisol. Brain Behav. Immun. 22, 600e605.
Craig, C.L., Marshall, A.L., Sjostrom, M., Bauman, A.E., Booth, M.L., Ainsworth, B.E.,Pratt, M., Ekelund, U., Yngve, A., Sallis, J.F., Oja, P., 2003. International physicalactivity questionnaire: 12-country reliability and validity. Med. Sci. Sports Exerc.35, 1381e1395.
Daniali, L., Benetos, A., Susser, E., Kark, J.D., Labat, C., Kimura, M., Desai, K.,Granick, M., Aviv, A., 2013. Telomeres shorten at equivalent rates in somatictissues of adults. Nat. Commun. 4, 1597.
Epel, E.S., Merkin, S.S., Cawthon, R., Blackburn, E.H., Adler, N.E., Pletcher, M.J.,Seeman, T.E., 2009. The rate of leukocyte telomere shortening predicts mortalityfrom cardiovascular disease in elderly men. Aging (Albany.NY) 1, 81e88.
Epel, E.S., Blackburn, E.H., Lin, J., Dhabhar, F.S., Adler, N.E.,Morrow, J.D., Cawthon, R.M.,2004. Accelerated telomere shortening in response to life stress. Proc. Natl. Acad.Sci. U.S.A 101, 17312e17315.
Epel, E.S., Lin, J., Wilhelm, F.H., Wolkowitz, O.M., Cawthon, R., Adler, N.E., Dolbier, C.,Mendes, W.B., Blackburn, E.H., 2006. Cell aging in relation to stress arousal andcardiovascular disease risk factors. Psychoneuroendocrinology 31, 277e287.
Fitzpatrick, A.L., Kronmal, R.A., Gardner, J.P., Psaty, B.M., Jenny, N.S., Tracy, R.P.,Walston, J., Kimura, M., Aviv, A., 2007. Leukocyte telomere length and cardio-vascular disease in the cardiovascular health study. Am. J. Epidemiol. 165, 14e21.
Fuster, J.J., Andres, V., 2006. Telomere biology and cardiovascular disease. Circ. Res.99, 1167e1180.
Hartmann, N., Boehner, M., Groenen, F., Kalb, R., 2010. Telomere length of patientswith major depression is shortened but independent from therapy and severityof the disease. Depress. Anxiety 27, 1111e1116.
Hoen, P.W., de Jonge, P., Na, B.Y., Farzaneh-Far, R., Epel, E., Lin, J., Blackburn, E.,Whooley, M.A., 2011. Depression and leukocyte telomere length in patients withcoronary heart disease: data from the Heart and Soul Study. Psychosom. Med.73, 541e547.
Houtveen, J.H., Groot, P.F.C., Geus, E.J.C., 2005. Effects of variation in posture andrespiration on RSA and pre-ejection period. Psychophysiology 42, 713e719.
Juster, R.P., McEwen, B.S., Lupien, S.J., 2010. Allostatic load biomarkers of chronicstress and impact on health and cognition. Neurosci. Biobehav. Rev. 35, 2e16.
Kajstura, J., Rota, M., Urbanek, K., Hosoda, T., Bearzi, C., Anversa, P., Bolli, R., Leri, A.,2006. The telomere-telomerase axis and the heart. Antioxid. Redox. Signal. 8,2125e2141.
Kiecolt-Glaser, J.K., Epel, E.S., Belury, M.A., Andridge, R., Lin, J., Glaser, R.,Malarkey, W.B., Hwang, B.S., Blackburn, E., 2013. Omega-3 fatty acids, oxidativestress, and leukocyte telomere length: a randomized controlled trial. BrainBehav. Immun. 28, 16e24.
Kiecolt-Glaser, J.K., Gouin, J.P., Weng, N.P., Malarkey, W.B., Beversdorf, D.Q.,Glaser, R., 2011. Childhood adversity heightens the impact of later-life care-giving stress on telomere length and inflammation. Psychosom. Med. 73, 16e22.
Kiecolt-Glaser, J.K., Preacher, K.J., MacCallum, R.C., Atkinson, C., Malarkey, W.B.,Glaser, R., 2003. Chronic stress and age-related increases in the proin-flammatory cytokine IL-6. Proc. Natl. Acad. Sci. U.S.A 100, 9090e9095.
Kirschbaum, C., Hellhammer, D.H., 1994. Salivary cortisol in psychoneuroendocrineresearch: recent developments and applications. Psychoneuroendocrinology 19,313e333.
Kroenke, C.H., Epel, E., Adler, N., Bush, N.R., Obradovic, J., Lin, J., Blackburn, E.,Stamperdahl, J.L., Boyce, W.T., 2011. Autonomic and adrenocortical reactivityand buccal cell telomere length in kindergarten children. Psychosom. Med. 73,533e540.
Ledford, H., 2007. Minimum telomere length defined for healthy cells. Nature 449,515.
Licht, C.M., Vreeburg, S.A., van Reedt Dortland, A.K., Giltay, E.J., Hoogendijk, W.J.,Derijk, R.H., Vogelzangs, N., Zitman, F.G., de Geus, E.J., Penninx, B.W., 2010.Increased sympathetic and decreased parasympathetic activity rather thanchanges in hypothalamic-pituitary-adrenal axis activity is associated withmetabolic abnormalities. J. Clin. Endocrinol. Metab. 95, 2458e2466.
Licht, C.M.M., Penninx, B.W.J.H., de Geus, E.J.C., 2012. Effects of antidepressants, butnot psychopathology, on cardiac sympathetic control: a longitudinal study.Neuropsychopharmacology 37, 2487e2495.
Licht, C., de Geus, E., Penninx, B., 2013. Dysregulation of the autonomic nervoussystem predicts the development of the metabolic syndrome. J. Clin. Endocrinol.Metab. 98, 2484e2493.
Lin, J., Epel, E., Cheon, J., Kroenke, C., Sinclair, E., Bigos, M., Wolkowitz, O., Mellon, S.,Blackburn, E., 2010. Analyses and comparisons of telomerase activity andtelomere length in human T and B cells: insights for epidemiology of telomeremaintenance. J. Immunol. Methods 352, 71e80.
Mangan, D.F., Wahl, S.M., 1991. Differential regulation of human monocyte pro-grammed cell death (apoptosis) by chemotactic factors and pro-inflammatorycytokines. J. Immunol. 147, 3408e3412.
D. Révész et al. / Neurobiology of Aging 35 (2014) 1422e1430 1429

Author's personal copy
Martin-Ruiz, C., Dickinson, H.O., Keys, B., Rowan, E., Kenny, R.A., von Zglinicki, T.,2006. Telomere length predicts poststroke mortality, dementia, and cognitivedecline. Ann. Neurol. 60, 174e180.
Masi, S., Nightingale, C.M., Day, I.N.M., Guthrie, P., Rumley, A., Lowe, G.D.O., VonZglinicki, T., D’Aiuto, F., Taddei, S., Klein, N., Salpea, K., Cook, D.G.,Humphries, S.E., Whincup, P.H., Deanfield, J.E., 2012. Inflammation and notcardiovascular risk factors is associated with short leukocyte telomere length in13- to 16-year-old adolescents. Arterioscler. Thromb. Vasc. Biol. 32, 2029e2034.
McEwen, B.S., 2008. Central effects of stress hormones in health and disease: un-derstanding the protective and damaging effects of stress and stress mediators.Eur. J. Pharmacol. 583, 174e185.
Merino, A., Buendia, P., Martin-Malo, A., Aljama, P., Ramirez, R., Carracedo, J., 2011.Senescent CD14þCD16þ monocytes exhibit proinflammatory and proathero-sclerotic activity. J. Immunol. 186, 1809e1815.
Mills, P.J., Adler, K.A., Dimsdale, J.E., Perez, C.J., Ziegler, M.G., ncoli-Israel, S.,Patterson, T.L., Grant, I., 2004. Vulnerable caregivers of Alzheimer disease pa-tients have a deficit in beta 2-adrenergic receptor sensitivity and density. Am. J.Geriatr. Psychiatry 12, 281e286.
O’Donovan, A., Lin, J., Dhabhar, F.S., Wolkowitz, O., Tillie, J.M., Blackburn, E., Epel, E.,2009. Pessimism correlates with leukocyte telomere shortness and elevatedinterleukin-6 in post-menopausal women. Brain Behav. Immun. 23, 446e449.
O’Donovan, A., Pantell, M.S., Puterman, E., Dhabhar, F.S., Blackburn, E.H., Yaffe, K.,Cawthon, R.M., Opresko, P.L., Hsueh, W.C., Satterfield, S., Newman, A.B.,Ayonayon, H.N., Rubin, S.M., Harris, T.B., Epel, E.S., 2011. Cumulative inflam-matory load is associated with short leukocyte telomere length in the Health,Aging and Body Composition Study. PLoS One 6, e19687.
O’Donovan, A., Tomiyama, A.J., Lin, J., Puterman, E., Adler, N.E., Kemeny, M.,Wolkowitz, O.M., Blackburn, E.H., Epel, E.S., 2012. Stress appraisals and cellularaging: a key role for anticipatory threat in the relationship between psycho-logical stress and telomere length. Brain Behav. Immun. 26, 573e579.
Olovnikov, A.M., 1996. Telomeres, telomerase, and aging: origin of the theory. Exp.Gerontol. 31, 443e448.
Ornish, D., Lin, J., Daubenmier, J., Weidner, G., Epel, E., Kemp, C., Magbanua, M.J.,Marlin, R., Yglecias, L., Carroll, P.R., Blackburn, E.H., 2008. Increased telomeraseactivity and comprehensive lifestyle changes: a pilot study. Lancet Oncol. 9,1048e1057.
Parks, C.G., Miller, D.B., McCanlies, E.C., Cawthon, R.M., Andrew, M.E., DeRoo, L.A.,Sandler, D.P., 2009. Telomere length, current perceived stress, and urinary stresshormones in women. Cancer Epidemiol. Biomarkers Prev. 18, 551e560.
Penninx, B.W.J.H., Beekman, A.T.F., Smit, J.H., Zitman, F.G., Nolen, W.A., Spinhoven, P.,Cuijpers, P., de Jong, P.J., Van Marwijk, H.W.J., Assendelft, W.J.J., van der Meer, K.,Verhaak, P., Wensing, M., de Graaf, R., Hoogendijk, W.J., Ormel, J., van Dyck, R.,2008. The Netherlands Study of Depression and Anxiety (NESDA): rationale,objectives and methods. Int. J. Methods Psychiatr. Res. 17, 121e140.
Penninx, B.W.J.H., Milaneschi, Y., Lamers, F., Vogelzangs, N., 2013. Understanding thesomatic consequences of depression: biological mechanisms and the role ofdepression symptom profile. BMC Med. 11, 129.
Pruessner, J.C., Kirschbaum, C., Meinlschmid, G., Hellhammer, D.H., 2003. Two for-mulas for computation of the area under the curve represent measures of totalhormone concentration versus time-dependent change. Psychoneur-oendocrinology 28, 916e931.
Ruff, C.T., Mega, J., Morrow, D., Scirica, B., Frackelton, E.C., Hakonarson, H.,Crowley, K., Contant, C., Cannon, C., Braunwald, E., Sabatine, M., 2012. Telomerelength and cardiovascular outcomes. J. Am. Coll. Cardiol. 59, 356.
Stuurgroep Multidisciplinaire Richtlijnontwikkeling, 2009. Stoornissen in hetgebruik van alcohol: Richtlijn voor de diagnostiek en behandeling van patiëntenmet een stoornis in het gebruik van alcohol.
Task Force of the European Society of Cardiology the North American Societyof Pacing Electrophysiology, 1996. Heart rate variability: standards of mea-surement, physiological interpretation and clinical use. Circulation 93,1043e1065.
Tomiyama, A.J., O’Donovan, A., Lin, J., Puterman, E., Lazaro, A., Chan, J., Dhabhar, F.S.,Wolkowitz, O., Kirschbaum, C., Blackburn, E., Epel, E., 2012. Does cellular agingrelate to patterns of allostasis? An examination of basal and stress reactive HPAaxis activity and telomere length. Physiol. Behav. 106, 40e45.
van Aken, M.O., Romijn, J.A., Miltenburg, J.A., Lentjes, E.G.W.M., 2003. Automatedmeasurement of salivary cortisol. Clin. Chem 49, 1408e1409.
van Reedt Dortland, A., Vreeburg, S., Giltay, E., Licht, C., Vogelzangs, N., van Veen, T.,de Geus, E., Penninx, B., Zitman, F., 2012. The impact of stress systems andlifestyle on dyslipidemia and obesity in anxiety and depression. Psychoneur-oendocrinology 38, 209e218.
Verhoeven, J., Révész, D., Epel, E., Lin, J., Wolkowitz, O., Penninx, B., 2013. Majordepressive disorder and accelerated cellular aging: results from a large psy-chiatric cohort study. Mol. Psychiatry. Epub doi: 10.1038/mp.2013.151.
Vogelzangs, N., Beekman, A.T.F., de Jonge, P., Penninx, B.W.J.H., 2013. Anxiety dis-orders and inflammation in a large adult cohort. Transl. Psychiatry 3, e249.
Vogelzangs, N., Duivis, H.E., Beekman, A.T.F., Kluft, C., Neuteboom, J., Hoogendijk, W.,Smit, J.H., de Jonge, P., Penninx, B.W.J.H., 2012. Association of depressive dis-orders, depression characteristics and antidepressant medication with inflam-mation. Transl. Psychiatry 2, e79.
Vreeburg, S.A., Hoogendijk, W.J., van, P.J., Derijk, R.H., Verhagen, J.C., van, D.R.,Smit, J.H., Zitman, F.G., Penninx, B.W., 2009a. Major depressive disorder andhypothalamic-pituitary-adrenal axis activity: results from a large cohort study.Arch. Gen. Psychiatry 66, 617e626.
Vreeburg, S.A., Kruijtzer, B.P., van, P.J., van,D.R., Derijk, R.H.,Hoogendijk,W.J., Smit, J.H.,Zitman, F.G., Penninx, B.W., 2009b. Associations between sociodemographic,sampling and health factors and various salivary cortisol indicators in a largesample without psychopathology. Psychoneuroendocrinology 34, 1109e1120.
Vreeburg, S.A., Zitman, F.G., van, P.J., Derijk, R.H., Verhagen, J.C., van, D.R.,Hoogendijk,W.J., Smit, J.H., Penninx, B.W., 2010. Salivary cortisol levels in personswith and without different anxiety disorders. Psychosom. Med. 72, 340e347.
Weischer, M., Bojesen, S.E., Cawthon, R.M., Freiberg, J.J., Tybjaerg-Hansen, A.,Nordestgaard, B.G., 2012. Short telomere length, myocardial infarction, ischemicheart disease, and early death. Arterioscler. Thromb. Vasc. Biol. 32, 822e829.
Wikgren, M., Maripuu, M., Karlsson, T., Nordfjall, K., Bergdahl, J., Hultdin, J.,Del-Favero, J., Roos, G., Nilsson, L.G., Adolfsson, R., Norrback, K.F., 2012. Shorttelomeres in depression and the general population are associated with ahypocortisolemic state. Biol. Psychiatry 71, 294e300.
Willemsen, G.H., de Geus, E.J., Klaver, C.H., van Doornen, L.J., Carroll, D., 1996.Ambulatory monitoring of the impedance cardiogram. Psychophysiology 33,184e193.
Wilson, W.R., Herbert, K.E., Mistry, Y., Stevens, S.E., Patel, H.R., Hastings, R.A.,Thompson, M.M., Williams, B., 2008. Blood leucocyte telomere DNA contentpredicts vascular telomere DNA content in humans with and without vasculardisease. Eur. Heart J. 29, 2689e2694.
Wolkowitz,O.M.,Mellon, S.H., Epel, E.S., Lin, J., Dhabhar, F.S., Su, Y., Reus, V.I., Rosser, R.,Burke, H.M., Kupferman, E., Compagnone, M., Nelson, J.C., Blackburn, E.H., 2011.Leukocyte telomere length in major depression: correlations with chronicity,inflammation and oxidative stressepreliminary findings. PLoS One 6, e17837.
Yu, B.H., Kang, E.H., Ziegler, M.G., Mills, P.J., Dimsdale, J.E., 2008. Mood states,sympathetic activity, and in vivo beta-adrenergic receptor function in a normalpopulation. Depress. Anxiety 25, 559e564.
Zhang, G.Q., Zhang, W., 2009. Heart rate, lifespan, and mortality risk. Ageing Res.Rev. 8, 52e60.
D. Révész et al. / Neurobiology of Aging 35 (2014) 1422e14301430