crosstalk between the two sides of the thalamus through the reticular nucleus: a retrograde and...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 332:145-154 (1993)
Crosstalk Between the Two Sides of the Thalamus Through the Reticular Nucleus:
A Retrograde and Anterograde Tracing Study in the Rat
VASSILIS RAOS AND MARINA BENTIVOGLIO Institute of Anatomy, University of Verona, Italy
ABSTRACT In order to investigate the possible routes linking the thalamus in the two sides of the
brain, the connections of the reticular nucleus (RT), the major component of the ventral thalamus, with contralateral dorsal thalamic nuclei were systematically investigated in the adult rat. This study was performed with several tract-tracing techniques: single and double retrograde labeling with fluorescent tracers, and anterograde tracing with biocytin. Retrograde tracing was also combined with immunocytochemistry to provide additional criteria for the identification of labeled RT neurons.
The data obtained with the retrograde transport of one fluorescent tracer showed that RT neurons project to contralateral dorsal thalamic domains. In particular, retrograde labeling findings indicated that the anterior intralaminar nuclei, as well as the ventromedial (VM) nucleus, are preferential targets of the contralateral RT projections. Commissural neurons were concentrated in two portions of RT: its rostral part, including the rostral pole, which projects to the contralateral central lateral (CL) and paracentral (Pc) nuclei, and the ventro- medial sector of the middle third of RT, which projects to the contralateral VM and posterior part of CL and Pc. The double retrograde labeling study of the bilateral RT-intralaminar connection indicated that at least part of the commissural RT cells bifurcate bilaterally to symmetrical portions of the anterior intralaminar nuclei,
The targets of the RT commissural system inferred from the retrograde labeling data were largely confirmed by anterograde tracing. Moreover, it was shown that RT fibers cross the midline in the intrathalamic commissure.
The present data demonstrate that bilateral RT connections with the dorsal thalamus provide a channel for interthalamic crosstalk. Through these bilateral connections with thalamic VM and intralaminar neurons, RT could influence the activity of wide territories of the cerebral cortex and basal ganglia of both hemispheres.
Key words: commissural connections, thalamocortical system, ventral thalamus, fluorescent tracers,
e 1993 Wiley-Liss, Inc.
biocytin
The route of communication among different nuclei within each side of the thalamus is mediated by the connections of the reticular nucleus (RT), the major compo- nent of the ventral thalamus (Jones, '851, with dorsal thalamic domains (see Steriade et al., '90, for review). Much less is known about the relationship between the two sides of the thalamus, which have so far received very little attention. Glees and Wall ('48) provided evidence in the monkey with the Marchi technique of an interthalamic bundle of degenerating fibers following lesions of the lateral nuclear group of the thalamus. The occurrence of connec- tions linking dorsal thalamic structures of the two sides has not been subsequently confirmed. However, there is some
evidence of an interthalamic communication. For example, the involvement of thalamic nuclei in the transfer of information from one substantia nigra to contralateral basal ganglia components via an interthalamic pathway has been reported in both the rat and the cat (Romo et al., '84; Savaki et al., '84). Anatomical findings indicated the occur- rence in the cat of bilateral RT projections to the ventrome-
Accepted November 9,1992. V. Raos's current address is Laboratory of Physiology, Dept. of Basic
Address correspondence to Dr. M. Bentivoglio, Institute of' Anatomy Sciences, School of Health Sciences, University of Crete, Iraklion, Greece.
Medical Faculty, Strada Le Grazie, 37134 Verona, Italy.
o 1993 WILEY-LISS, INC.

146 V. RAOS AND M. BENTIVOGLIO
dial sector of the dorsal thalamus (Rinvik, '84). Connec- tions of the ventral lateral geniculate nucleus with the contralateral homologous structure and intralaminar nu- clei (Nakamura and Kawamura, '88), and a minor input to the mediodorsal nucleus (MD) from the contralateral RT (Velayos and Reinoso-Suarez, '851, have been incidentally reported in the cat. These data, as well as personal observa- tions of retrograde labeling in the contralateral RT after large tracer injections involving RT and dorsal thalamic nuclei in the rat (Chen et al., '921, suggest that ventral thalamic structures could provide channels for intertha- lamic crosstalk.
The present study was aimed at a systematic investiga- tion of the connections that could allow communication between the two sides of the dorsal thalamus in the rat, and was pursued with tract-tracing techniques in three steps. The retrograde transport of one tracer first revealed that only RT cells project bilaterally to the dorsal thalamus. In this part of the study, RT neurons were identified not only on the basis of cytoarchitectonic criteria but also with immunocytochemistry by using antibodies antiparvalbu- min, a calcium binding protein intensely expressed only by RT cells in the rat thalamus (Celio, '90; Frassoni et al., '91), which helps the delimitation of the nucleus from the adjacent internal capsule, dorsal thalamus, and zona in- certa (Celio, '90). We then verified by means of double retrograde tracing whether the commissural connection was established by RT cells that bifurcate bilaterally or by separate RT neurons projecting unilaterally to either side of the dorsal thalamus. Finally, in order to investigate the distribution of RT commissural fibers, injections of the anterograde tracer biocytin within RT were devised on the basis of the retrograde labeling findings.
MATERIALS AND METHODS Surgical and histological procedures
A total of 19 adult Wistar rats (260-330 g) were used. All surgical procedures, including perfusions, were performed
AD AM AV CeM CL fr G Gu Hb IAM ic LD LB MD mt Pc Pf Po Pt Pv Re Rh RT sm sn, VM VP
Abbreviations
anterodorsal n. anteromedial n. anteroventral n. central medial n. central lateral n. fasciculus retroflexus n. gelatinosus gustatory n. habenular complex interanteromedial n. internal capsule laterodorsal n. lateroposterior n . mediodorsal n. marnmillothalamic tract paracentral n. parafascicular n. posterior complex paratenial n. paraventricular n . n. reuniens rhomboid n. reticular n. stria medullaris ventrolateral n. ventromedial n. ventroposterior complex
TABLE 1. Experimental Data of the Retrograde Tracing Experiments
Case Injected Core of the code Tracer' volumes injection areasi
Single retrograde labeling Group A V6 2% DY 0 I) 111 VL, VP, Po v 3 2% FG 0.4 wI VL, VP, Po VR 19 FG 0.4 111 VL, VP, RT, ZI v 9 1% FG 0.4 rrl VL, VP, Po v11 2% DY 1.0 111 VL, VP, Po Group R v10 2% FG (1.3 wI CL, Pc, VL, VP, Po, VM V15 2? DY 0.2 WLI CL, Pr , VL. VM v7 X i DY 0.2 W l CL, Pc, VP v 5 2<b DY 0.2 W l CL, Pc v12 2% DY 0.2 W l CI,, P r V13 2'6 DY 0.2 +I CL
Double retrograde labeling VDL6 L: 2% DY 0.6 111 CL, Pc
VDLI L: 2% DY 0.6 111 CL, Pc
VDLR L: 2% DY 1 4 111 CL, Pc
'DY, diamidino yellow; FW, fast blue; FG, fluoragoold. T L , central lateral n . , MD, mediodorsal n ; Pc, paracentral n.. Po, posterior complex. RT, reticular n.; VL, ventrolateral n.; VM, ventromedial n.. VP, ventroposterior complex; ZI, zona incerta.
R. 2% FR 0 fi 111 CL, Pc, MD
R. 2% FB 0.6 111 CL, Pc, MD
R: 2% FR 1.2 111 CL, Pc, MD
under deep barbiturate anesthesia (sodium pentobarbital, 40 mg/kg i.p.). The tracers were injected stereotaxically by using the atlas of Paxinos and Watson ('86). The total injected volumes (Table 1) were delivered with Hamilton microsyringes through multiple penetrations.
All animals were perfused transcardially with saline followed by 4% phosphate-buffered paraformaldehyde (pH 7.4). The brains were dissected out, briefly postfixed in the fixative solution, and soaked in 30% buffered sucrose until they sank. Serial coronal sections were then cut at a 30 Frn thickness through the diencephalon on a freezing micro- tome. Every third and fourth section was collected in phosphate-buffered saline (PBS) in two adjacent series: One was used to study the distribution of labeled cell bodies or fibers, and the other was counterstained with cresyl violet for cytoarchitectonic control of the injection areas and loca- tion of the labeling. The sections from cases injected with fluorescent tracers were mounted on gelatinized slides, air dried, rapidly dipped in xylol, and coverslipped with Entellan.
Retrograde single- and double-labeling experiments
The retrograde tracers Fast Blue (FB, Illing), Diamidino Yellow dihydrochloride (DY, Illing), and Fluorogold (FG, Fluorochrome Inc.) were dissolved or suspended in distilled water. The experimental parameters of the injection proce- dure are summarized in Table 1. DY or FG were used for single-labeling experiments in 11 cases, whereas FB and DY were combined in 3 additional double retrograde labeling experiments. The survival period ranged from 3 to 5 days. The sections were studied with a Leitz Ploempack fluores- cence microscope, using filter-mirror systems A (360 nm excitation wavelength) and D (390 nm excitation wave- length). Both filter systems elicit the blue fluorescence of FB, which mainly labels the neuronal cytoplasm, the yellow- green DY fluorescence, confined to the neuronal nucleus, and the yellow-gold fluorescence of FG, which produces intense perikaryal labeling. The extent of the injection areas and the distribution of retrogradely labeled cells in RT were charted with a computerized X-Y plotter attached to the microscope stage by means of transducers.

RAT THALAMUS CROSSTALK VIA RETICULAR NUCLEUS 147
In order to assess a quantitative parameter suitable for the evaluation of the contralateral cell density, the neurons labeled in the contralateral RT were counted in the DY- injected cases. The percent value was then calculated in relation to the total labeled neuronal population [(contralat- eral RT cells)/(ipsilateral + contralateral RT cells) . 1001 in the two sides of RT through the same anteroposterior levels, i.e., throughout the sections in which contralateral neurons had been detected.
Immunocytochemical procedure In two cases injected with DY (V12 and V13, Table l),
most of the sections that contained labeled cells in the contralateral thalamus were processed for immunocyto- chemistry by using monoclonal antibodies raised in mouse against parvalbumin (kindly provided and characterized by Dr. M. Celio). The sections were first washed in PBS, pretreated with 5% normal goat serum in PBS for 45 minutes, and incubated overnight with primary antibody. The immune complex was visualized either with the avidin- biotin peroxidase (ABC) protocol (dilution of primary anti- body 1:10,000) following the procedure by Weinberg et al. ('85) or with immunofluorescence (dilution of primary antibody 1: 1,000) using secondary antibodies conjugated with rhodamine isothiocyanate (Jackson; 1:50 in PBS). After washing in PBS, all of the immunoreacted sections were mounted on g$latinized slides, air dried, quickly dipped in xylol, coverslipped with Entellan, and examined with the fluorescence microscope. In the sections processed with the ABC protocol, the dark-brown granules of the diaminobenzidine (DAB) reaction product could be detected in the cytoplasm of fluorescent DY-labeled cell bodies. The tissue processed with immunofluorescence was examined by switching during the observation from filter systems A and D to filter system N2 (550 nm excitation wavelength) in order to detect both the DY yellow labeling and the rhodamine red immunostaining.
Anterograde tracing experiments A freshly prepared 5% solution of biocytin (Sigma) in 1%
ethanol was injected in RT in 5 cases. The injections (0.3 ~1 each, delivered in one or two penetrations) were aimed at involving the rostral pole of the nucleus and/or the ventral sector of its middle third. The survival period ranged from 24 to 28 hours. Free-floating serial sections were processed for the visualization of biocytin by using the ABC protocol. After rinsing in PBS, the sections were treated with a DAB-H202 solution containing cobalt chloride and nickel ammonium (Adams, '81). After careful rinsing in PBS, the sections were mounted on gelatin-coated slides, air dried, dehydrated, coverslipped, and studied with brightfield illu- mination.
RESULTS Retrograde tracing experiments
Single retrograde labeling. The injection areas involved lateral or medial thalamic regions. In none of the cases was tracer diffusion to the opposite side observed.
As indi- cated in Table 1 and illustrated in the representative cases V6, V3, and V8 (Fig. I) , the extent and anteroposterior location of the injection areas varied in the different experiments. The central core consistently involved the ventrolateral (VL) and ventroposterior (VP) nuclei, as well
Injections in the lateral thalamus (group A).
as the posterior (Po) complex. In caseV8 (Fig. 11, the core of the injection area extended into the ventromedial portion of RT and the zona incerta. A halo of tracer diffusion was observed in the anterior nuclei (cases V6 and V3), rostral portion of RT (case V3), laterodorsal nucleus (cases V3, V8, V9), and the lateral part of the ventromedial (VM) nucleus (cases V8, V9, V11).
Retrogradely labeled cells were densely packed within the ipsilateral RT in all of the experiments of group A. Labeled neurons were distributed in various sectors of the nucleus throughout its extent. However, no labeled cells were observed in the contralateral thalamus in any of the cases of group A.
Injections in the anterior intralaminar and ventromedial nuclei (group B). In six additional animals, the injections were placed more medially than in group A. In the cases injected with DY (Table 1; Figs. 1, 2), the central core of the injection areas involved the anterior intralaminar central lateral (CL) and paracentral (Pc) nuclei at different antero- posterior levels, and in case V15 (Fig. 2) it also extended ventrally into VM. In case V13 (Table 1) the injection area was confined within CL with no involvement of Pc. A restricted halo of DY diffusion was observed in the lateral part of MD (cases V12, Fig. 1, and V15, Fig. a), or in VL (case V5, Fig. 2). In the experiment injected with FG (VIO, Fig. 11, the injection area involved VL, VP, Po, the lateral part of VM, the anterior intralaminar nuclei, and in partic- ular Pc.
In all of the group B experiments, numerous cells were retrogradely labeled in the ipsilateral RT. In addition, labeled neurons were observed in RT on the contralateral side, concentrated in portions of the nucleus that displayed a high density of labeled cells ipsilaterally. However, the reverse was not obvious, in that heavily labeled portions of the ipsilateral RT were not always accompanied by 'mirror' loci of contralateral labeling.
In cases V5 (Fig. 2) and V12, labeled neurons were located throughout the anterior portion of the contralateral RT, mainly distributed ventrally and medially, with a predominance in its rostral pole. In case V13, the contralat- eral neurons were concentrated in the ventral sector of the rostral portion of RT.
The contralateral labeled cells were instead located in the ventromedial part of the RT middle third after injections in the posterior part of CL and Pc (case V7), extending also into VM (case V15, Fig. 2). The FG injections (V10, Fig. 1) resulted in contralateral labeled cells scattered through both the rostral portion of RT and the ventromedial part of its middle third.
The combination of immunocytochemistry with DY retro- grade tracing, performed in cases V12 and V13, revealed that the neurons labeled in the contralateral thalamus, including those located at the most medial edge of RT, were parvalbumin-immunoreactive. It was thus confirmed that the contralateral retrogradely labeled cells were confined within RT.
The percentage of labeled contralateral RT cells, evalu- ated in the cases injected with DY in relation to the neuronal population labeled on both sides in the same levels of RT, appeared to be related to the location of the injection areas rather than to their extent. A relatively high density of contralateral cells (21% and 26%) was detected after injections in the anterior part of CL and Pc (cases V5 and V12). The density of contralateral neurons was also high (25%) after injection in VM and the posterior part of CL and
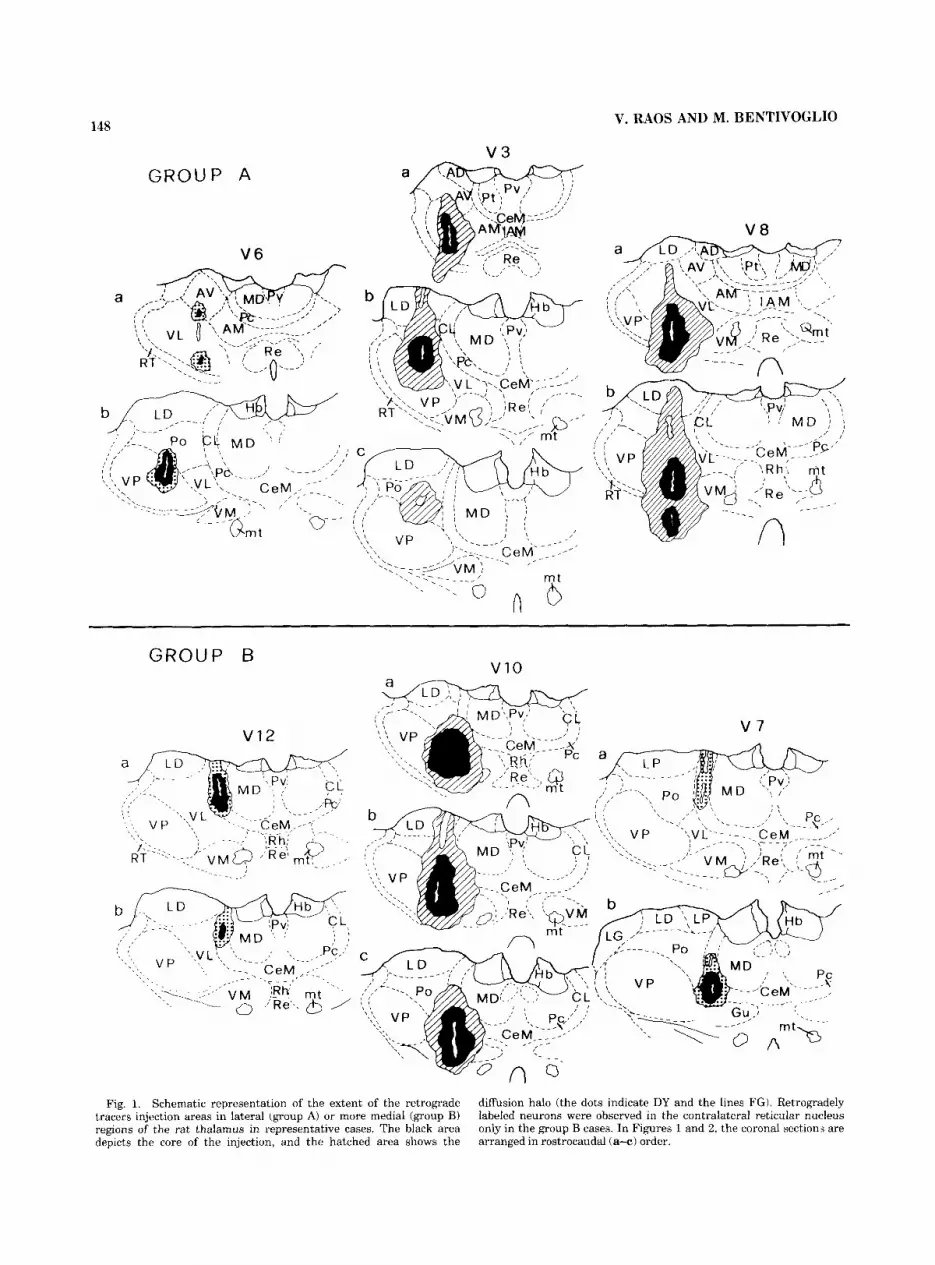
148 V. RAOS AND M. BENTIVOGLIO
GROUP A
V 6
v3 a
GROUP B
v12
v10
Fig. 1. Schematic representation of the extent of the retrograde tracers injection areas in lateral (group A) or more medial (group B) regions of the rat thalamus in representative cases. The black area depicts the core of the injection, and the hatched area shows the
diffusion halo (the dots indicate DY and the lines FG). Retrogradely labeled neurons were observed in the contralateral reticular nucleus only in the group B cases. In Figures 1 and 2, the coronal section,; are arranged in rostrocaudal (a+) order.
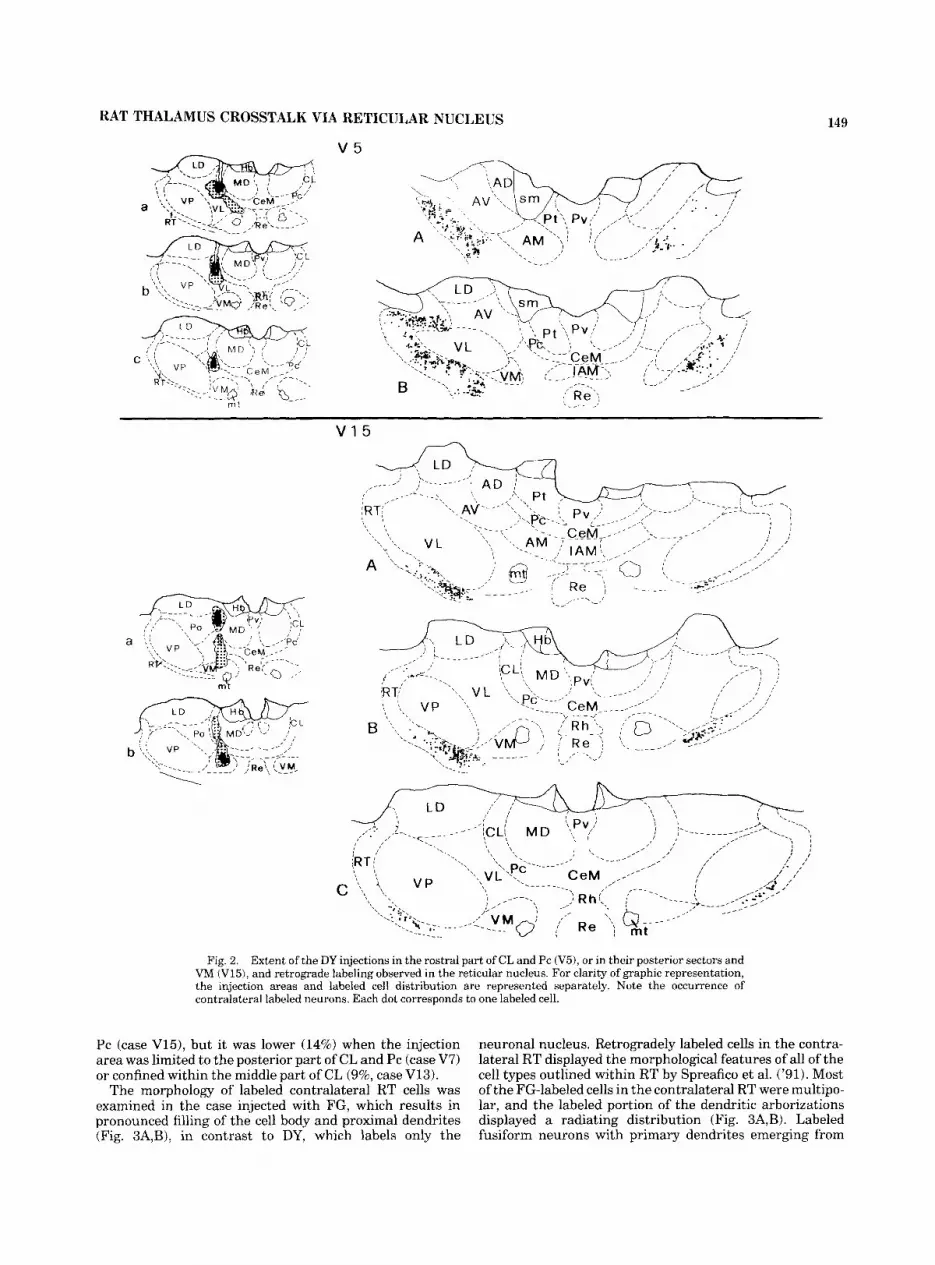
RAT THALAMUS CROSSTALK VIA RETICULAR NUCLEUS
v 5
C
Fig. 2. Extent of the DY injections in the rostra1 part of CL and Pc ( V 3 , or in their posterior sectors and VM (V15), and retrograde labeling observed in the reticular nucleus. For clarity of graphic representation, the injection areas and labeled cell distribution are represented separately. Note the occurrence of contralateral labeled neurons. Each dot corresponds to one labeled cell.
149
Pc (case V15), but it was lower (14%) when the injection area was limited to the posterior part of CL and Pc (case V7) or confined within the middle part of CL (9%, case V13).
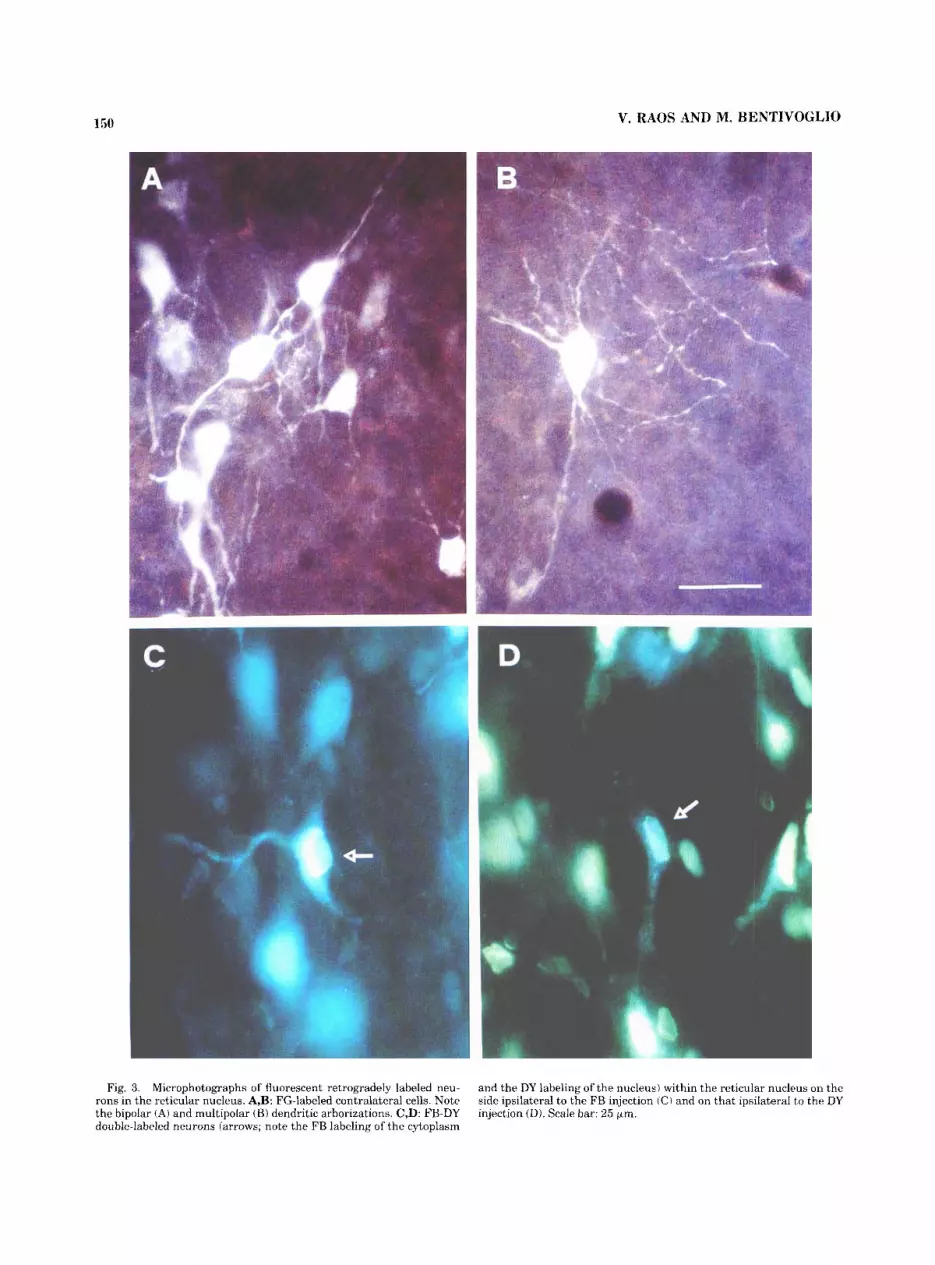
The morphology of labeled contralateral RT cells was examined in the case injected with FG, which results in pronounced filling of the cell body and proximal dendrites (Fig. 3A,B), in contrast to DY, which labels only the
neuronal nucleus. Retrogradely labeled cells in the contra- lateral RT displayed the morphological features of all of the cell types outlined within RT by Spreafico et al. ('91). Most of the FG-labeled cells in the contralateral RT were multipo- lar, and the labeled portion of the dendritic arborizations displayed a radiating distribution (Fig. 3A,B). Labeled fusiform neurons with primary dendrites emerging from
150 V. RAOS AND M. BENTIVOGLIO
Fig. 3. Microphotographs of fluorescent retrogradely labeled neu- rons in the reticular nucleus. A,B: FG-labeled contralateral cells. Note the bipolar (A) and multipolar (B) dendritic arborizations. C,D: FB-DY double-labeled neurons (arrows; note the FB labeling of the cytoplasm
and the DY labeling of the nucleus) within the reticular nucleus on the side ipsilateral to the FB injection (C) and on that ipsilateral to the DY injection (D). Scale bar: 25 pm.

151 RAT THALAMUS CROSSTALK VIA RETICULAR NUCLEUS
A n
Fig. 4. A-C: Schematic representation of the biocytin injection sites (hatched areas) in the anterior part of the reticular nucleus in three cases.
opposite sides of the cell body were also observed (Fig. 3A), and the long axis of these neurons was parallel to the dorsoventral axis of RT.
In the double tracing exper- iments (Table 11, the FB and DY injections were strictly unilateral. The injection areas were located dorsally and their core involved CL, extending into the dorsal part of Pc without invading the structures of the ventral nuclear group. The FB injection area was consistently larger than that of DY, and involved a wider rostrocaudal extent of the anterior intralaminar and MD nuclei. In cases VDL6 and VDL7, the core of the DY injection area was symmetrical to that of FB on the other side, whereas in case VDLS it was located slightly more laterally than the FB injection core.
Neurons retrogradely labeled from each injection were very numerous in the ipsilateral RT. In all cases, labeled neurons were also detected in the contralateral RT, concen- trated in the rostral part of the nucleus. On both sides, RT also contained double-labeled neurons (Fig. 3C,D). The three cell populations (single-labeled ipsilateral, single- labeled contralateral, and double labeled) were intermin- gled on each side of RT. In addition, FB-single-labeled cells were distributed in portions of the contralateral RT (mainly the ventral sector of its anterior portion), which did not contain neurons labeled from the DY ipsilateral injection.
The number of double-labeled cells in the two sides of RT varied in the different cases and in each experiment. In particular, the proportion of double-labeled neurons ap- peared related to the symmetrical location of the injection areas. In experiments VDL6 and VDL7, with a relatively restricted DY injection symmetrical to part of the larger FB injection core, the vast majority of the DY-labeled contralat- era1 neurons were double labeled. whereas 8% (VDLG) and
Double retrograde labeling.
In four cases the injection area involved the rostral portion of RT-extending into either the basal forebrain or the anteroventral nucleus (AV)-as well as the ventral sector of RT, slightly invading the internal capsule (Fig. 4). In one of these cases (V32, Fig. 4) the injected area involved a large portion of the rostral pole of RT. Finally, in the remaining case, in which a single injection of biocytin was made, the injected area was more posterior than in the previous experiments and was confined within VP and the sector of RT lateral to it.
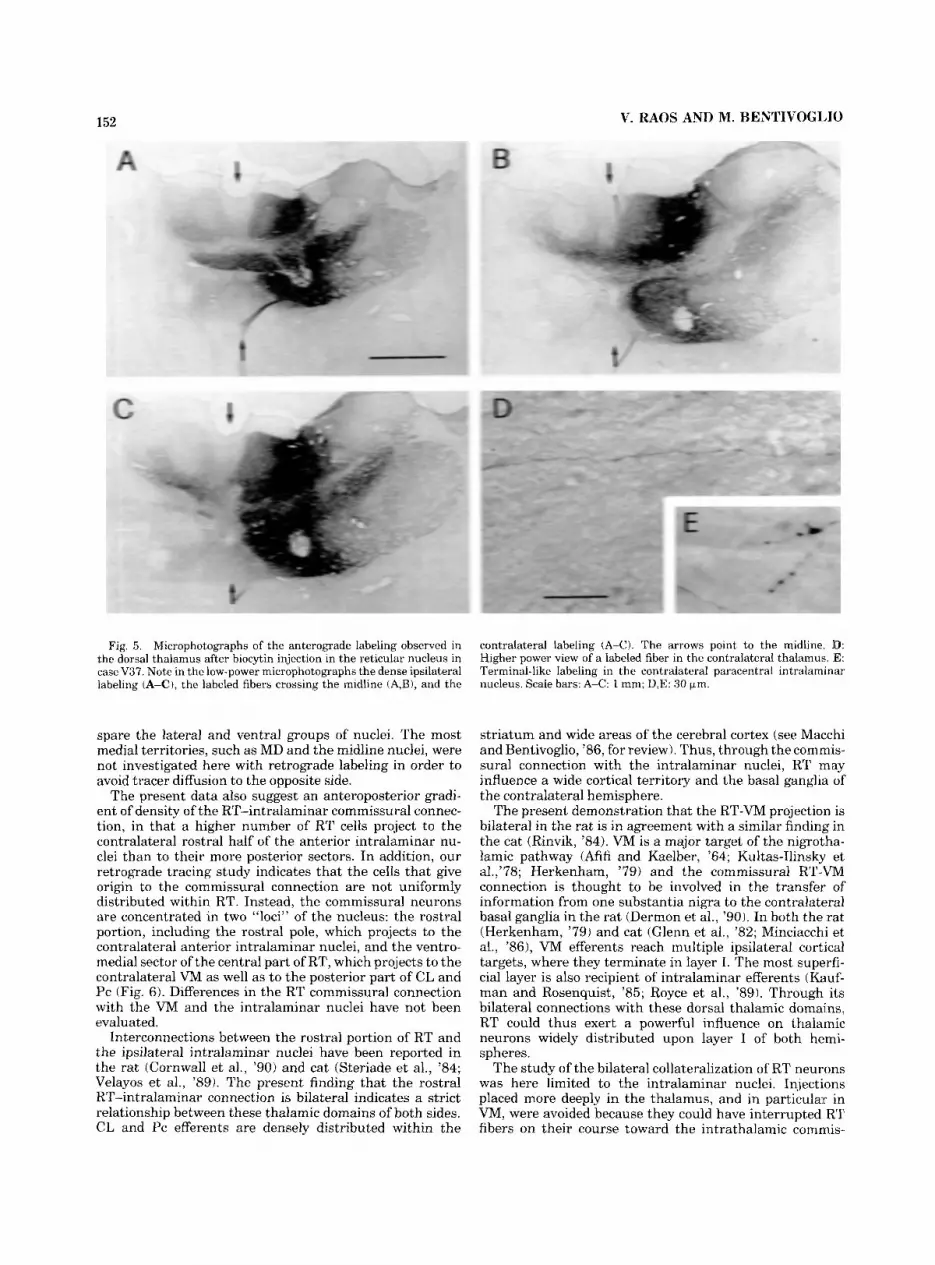
In this latter experiment, anterogradely labeled fibers and terminals were only observed within VP ipsilaterally, with no contralateral labeling. Dense anterograde labeling of fibers and preterminal and terminal-like elements was observed in all the other cases in the ipsilateral AV, anteromedial and interanteromedial nuclei, MD, VL, VM, and nucleus gelatinosus (Fig. 5A-C). In addition, in all these cases anterogradely labeled fibers were grouped in a bundle coursing in the internal medullary lamina and crossing the midline mainly at anterior levels (Fig. 5A,B). The labeled commissural fibers displayed a relatively thin caliber (Fig. 5D). Fine preterminal and terminal-like axonal arborizations could be detected in the central medial nu- cleus on both sides of the midline and were distributed contralaterally within CL and Pc (Fig. 5E), invading the lateral part of MD, as well as the dorsal part of VM. The contralateral labeling was especially dense in the cases in which a relatively large portion of the rostral part of RT had been involved in the injection area.
DISCUSSION Retrograde labeling findings
13% (VDL7) of the FB-labeled contralateral ones were double labeled. In experiment VDLS, in which the FB and DY injections were not entirely symmetrical, a few double- labeled neurons were observed on either side of RT.
The data obtained in the present study with the retro- grade transport of one fluorescent tracer indicate that RT neurons which project to contralateral dorsal thalamic structures establish a commissural svstem in the rat thala-
Anterograde tracing experiments mus. Other possible targets of the-RT projection to the contralateral thalamus cannot be ruled out on the basis of
Biocytin injections were placed in sectors of RT that were seen to give origin to commissural projections in the retrograde tracing experiments.
the present findings. However, our data indicate that the anterior intralaminar and VM nuclei are preferential tar- gets of RT fibers distributed contralaterally, which largely
152 V. RAOS AND M. BENTIVOGLJO
Fig. 5. Microphotographs of the anterograde labeling observed in the dorsal thalamus after biocytin injection in the reticular nucleus in case V37. Note in the low-power microphotographs the dense ipsilateral labeling (A-C), the labeled fibers crossing the midline iA,B), and the
contralateral labeling (A-C). The arrows point to the midline. ID: Higher power view of a labeled fiber in the contralateral thalamus. 'E: Terminal-like labeling in the contralateral paracentral intralarninar nucleus. Scale bars: A-C: 1 nun; D,E: 30 pm.
spare the lateral and ventral groups of nuclei. The most medial territories, such as MD and the midline nuclei, were not investigated here with retrograde labeling in order to avoid tracer diffusion to the opposite side.
The present data also suggest an anteroposterior gradi- ent of density of the RT-intralaminar commissural connec- tion, in that a higher number of RT cells project to the contralateral rostral half of the anterior intralaminar nu- clei than to their more posterior sectors. In addition, our retrograde tracing study indicates that the cells that give origin to the cornmissural connection are not uniformly distributed within RT. Instead, the commissural neurons are concentrated in two "loci" of the nucleus: the rostral portion, including the rostral pole, which projects to the contralateral anterior intralaminar nuclei, and the ventro- medial sector of the central part of RT, which projects to the contralateral VM as well as to the posterior part of CL and Pc (Fig. 6) . Differences in the RT commissural connection with the VM and the intralaminar nuclei have not been evaluated.
Interconnections between the rostral portion of RT and the ipsilateral intralaminar nuclei have been reported in the rat (Cornwall et al., '90) and cat (Steriade et al., '84; Velayos et al., '89). The present finding that the rostral RT-intralaminar connection is bilateral indicates a strict relationship between these thalamic domains of both sides. CL and Pc efferents are densely distributed within the
striatum and wide areas of the cerebral cortex (see Macchi and Bentivoglio, '86, for review). Thus, through the cornmis- sural connection with the intralaminar nuclei, R T may influence a wide cortical territory and the basal ganglia of the contralateral hemisphere.
The present demonstration that the RT-VM projection is bilateral in the rat is in agreement with a similar finding in the cat (Rinvik, '84). VM is a major target of the nigrotha- lamic pathway (Afifi and Kaelber, '64; Kultas-Ilinsky et a1.,'78; Herkenham, '79) and the commissural RT-VM connection is thought to be involved in the transfer of information from one substantia nigra to the contralateral basal ganglia in the rat (Dermon et al., '90). In both the rat (Herkenham, '79) and cat (Glenn et al., '82; Minciacchi et al., '861, VM efferents reach multiple ipsilateral cortical targets, where they terminate in layer 1. The most superfi- cial layer is also recipient of intralaminar efferents (Kauf- man and Rosenquist, '85; Royce et al., '89). Through its bilateral connections with these dorsal thalamic domains, RT could thus exert a powerful influence on thalaimic neurons widely distributed upon layer I of both hemi- spheres.
The study of the bilateral collateralization of RT neurons was here limited to the intralaminar nuclei. Injections placed more deeply in the thalamus, and in particular in VM, were avoided because they could have interrupted R T fibers on their course toward the intrathalamic cornrnis-

RAT THALAMUS CROSSTALK VIA RETICULAR NUCLEUS 153
- _ _ _ -
Fig. 6. Summary diagram of the bilateral projections of the reticular nucleus to the CL and Pc intralaminar nuclei and VM, shown in horizontal sections through the thalamus. The contralateral pathway is formed at least in part by axonal bifurcations of the ipsilateral connection, as demonstrated in the present study for the projections to the intralaminar nuclei.
sure, thus resulting in false-positive double labeling. The present data indicate that commissural RT cells are repre- sented, a t least in part, by neurons that bifurcate bilaterally upon the anterior intralaminar nuclei (Fig. 6). The relative proportion of the branched cell population displayed such a variability in the present experiments as to preclude any definite conclusion. However, the finding that most of the contralateral DY-labeled RT cells were double labeled in the cases in which the DY injection area was entirely symmetri- cal to that of FB on the other side suggests that the commissural pathway may be highly branched bilaterally. The dorsal placement of the injections, and the consistent finding of intermingled contralateral single-labeled and double-labeled cells on both sides of RT, indicate that tracer uptake from fibers of passage on their way to the contralat- era1 thalamus should not have occurred in these experi- ments.
Our study indicates that RT cells bifurcate upon symmet- rical portions of the anterior intralaminar nuclei of both sides (Fig. 6). These structures are composed in the cat of a mosaic of cell populations characterized by a different input-output organization (Bentivoglio et al., '88). Al- though this pattern has not been hitherto verified in the rat, the present findings suggest that single RT cells do not interconnect different portions of the intralaminar mosaic, and interact instead with the same intralaminar territory bilaterally.
Anterograde tracing experiments The targets of the RT commissural system inferred from
the retrograde tracing data were largely confirmed by the anterograde tracing experiments. RT fibers cross at all levels, with anterior prevalence, in the intrathalamic com- missure.
Most of the fiber bundles that enter the thalamus course through RT, so that the biocytin injections within this structure might have also involved fibers of passage. It has been reported that biocytin is not taken up by fibers of passage (King et al., '89; Izzo, '911, but this parameter was not specifically controlled in the present study. Therefore, we cannot exclude that some biocytin uptake from damaged axons might have occurred under the present experimental circumstances. Although most thalamic inputs are distrib- uted unilaterally, efferents from the frontal cortex reach the contralateral thalamus (Molinari et al., '85). However, biocytin injection in the ventrolateral sector of RT did not result in contralateral anterograde labeling, despite the fact that fibers of passage might also have been involved in these experiments. False-positive anterograde labeling could also arise from collaterals of neurons whose cell bodies could have taken up the tracer retrogradely. However, very few cell bodies in our experimental conditions were retrogradely labeled outside the thalamus. More importantly, the fact that the anterograde tracing data were entirely consistent with those revealed by retrograde methods provides in the present study a control of eventual false-positive antero- grade labeling.
Concluding remarks The RT commissural connection provides a channel for
crosstalk between the two sides of the dorsal thalamus at diencephalic level. Thus, RT mediates both ipsilaterally and contralaterally the exchange of information among dorsal thalamic structures, subserving unilateral intrathalamic segregation and bilateral interthalamic interaction of infor- mation processing in the dorsal thalamus. The anterior intralaminar and VM nuclei are crucial structures in the RT commissural pathway. On this basis, it seems reason-

154
able to suppose that the commissural RT cells would not subserve a filtering role of sensory and motor information processed in primary relay nuclei. Instead, the RT commis- surd system could be involved in global modulatory func- tions, mediating the interhemispheric balance of the mech- anisms subserving the regulation of thalamocortical spindle oscillations (Steriade et al., '90) and other aspects defining the states of wakefulness and sleep.
V. RAOS AND M. BENTIVOGLIO
ACKNOWLEDGMENTS The authors express their gratitude to Dr. S. Chen for his
help during all stages of this work, to Dr. M. Steriade for the stimulating discussions, and to Dr. G. Battaglia for his critical advice in the initial steps of this study. This work was supported by grants of the Italian Ministry of the University and Research, National Research Council, and Fidia Research Laboratories, and by grant E03111989 of the Greek Ministry of Health.
LITERATURE CITED Adams, J.C. (19811 Heavy metal intensification of DAB-based HRP reaction
product. J. Histochem. Cytochem. 29:775. Afifi, A,, and W.W. Kaelber (1964) Efferent connections of the substantia
nigra in the cat. Exp. Neurol. I 1 :474-482. Bentivoglio, M., D. Minciacchi, M. Molinari, A. Granato, R. Spreafico, and G.
Macchi (1988) The intrinsic and extrinsic organization of the thalamic intralaminar nuclei. In M. Bentivoglio and R. Spreafico (eds): Cellular Thalamic Mechanisms. Amsterdam: Elsevier, pp. 221-237.
Cello, M.R. (1990) Calbindin D-28k and parvalbumin in the rat nervous system. Neuroscience 35:375475.
Chen, S., V. Raos, and M. Bentivoglio (1992) Connections of the thalamic reticular nucleus with the contralateral thalamus in the rat. Neurosci. Lett. 14735-88.
Cornwall, J., J.D. Cooper, and O.T. Phillipson (1990) Projections to the rostral reticular thalamic nucleus in the rat. Exp. Brain Res. 80:157- 171.
Dermon, R.C., P. Pizarro, P. Georgopoulos, and E.H. Savaki (1990) Bilateral alterations in local cerebral glucose utilization following intranigral application of the GABAergic agonist muscimol. J. Neurosci. 10:2861- 2878.
Frassoni, C., M. Bentivoglio, R. Spreafico, M.P. Sanchez, L. Puelles, and A. Fairen ( 19911 Postnatal development of calbindin and parvalbumin immunoreactivity in the thalamus of the rat. Dev. Brain Res. 58243% 249.
Glees, P., and P.D. Wall (1948) Commissural fibers of the macaque thala- mus. An experimental study. J. Comp. Neurol. 88:129-137.
Glenn, L.L., J. Hada, J.P. Roy, M. Deschenes, and M. Steriade (1982) Anterograde tracer and field potential analysis of the neocortical layer I projection from nucleus ventralis medialis of the thalamus in cat. Neuroscience 7: 186 1-18 77.
Herkenham, M. (19791 The afferent and efferent connections of the ventro- medial thalamic nucleus in the rat. J. Comp. Neurol. 183:487-518.
Izzo, P.N. (1991) A note on the use of biocytin in anterograde tracing studies
in the central nervous system: Application at both light and electron microscopic level. J. Neurosci. Meth. 36:155-166.
Jones, E.G. (1985) The Thalamus. New York: Plenum. Kaufman, E.F.S., and A.C. Rosenquist (1985) Efferent projections or the
thalamic intralaminar nuclei in the cat. Brain Res. 335957-279. King, M.A., P.M. Louis, B.E. Hunter, and D.W. Walker (1989) Biocytin: A
versatile anterograde neuroanatomical tract-tracing alternative. Brain Res. 497:361-367.
Kultas-Ilinsky, K., LA. Ilinsky, I.C. Massopust, P A Young, and K.R. Smith (1978) Nigrothalamic pathway in the cat demonstrated by autoradiogra- phy and electron microscopy. Exp. Brain Res. 33481-492.
Macchi, G., and M. Bentivoglio (19861 The thalamic intralaminar nuclei and the cerebral cortex. In E.G. Jones and A. Peters (eds): Cerebral Cortex, Vol. 5 . New York: Plenum, pp. 355401.
Minciacchi, D., M. Bentivoglio, M. Molinari, K. Kultas-Ilinsky, LA. Ilinsk:?, and G. Macchi (1986) Multiple targets of one thalamic nucleus: The projections of the ventral medial nucleus in the cat studied with retrograde tracers. J. Comp. Neurol. 252:106-129.
Molinari, M., D. Minciacchi, M. Bentivoglio, and G. Macchi (1985) Efferent fibers from the motor cortex terminate bilaterally in the thalamus of rats and cats. Exp. Brain Res. 57:305-312.
Nakamura, H., and S. Kawamura (1988) The ventral lateral geniculate nucleus in the cat: Thalamic and commissural connections revealed by the use ofWGA-HRP transport. J. Comp. Neurol. 277509-528.
Paxinos, G., and C. Watson (1986) The Rat Brain in Stereotaxic Coordinatos. Sydney: Academic Press.
Rinvik, E. (1984) Thalamic commisural connections in the cat. Neurosci. Lett. 44:311-316.
Romo, R., A. Cheramy, G. Godeheu, and J. Glowinski (1984) Distinct commissural pathways are involved in the enhanced release of dopamine induced in the contralateral caudate nucleus and substantia nigra by unilateral application of GABA in the cat thalamic motor nuclei. Brain Res. 308:43-52.
Royce, G.J., S. Bromley, C. Gracco, and R.M. Beckstead (1989) Thalamocor- tical connections of the rostral intralaminar nuclei: An autoradiographic analysis in the cat. J. Comp. Neurol. 288:555-582.
Savaki, E.H., J.-A. Girault, M. Desban, J. Glowinski, and M.J . Besson (1984) Local cerebral metabolic effects induced by nigral stimulation following ventromedial thalamic lesions. I: Basal ganglia and related motor structures. Brain Res. Bull. 12609-616.
Spreafico, R., G. Battaglia, and C. Frassoni (1991) The reticular thalamic nucleus (RTN) of the rat: Cytoarchitectural, Golgi, immunocytochemi- cal, and horseradish peroxidase study. J. Conip. Neurol. 304:478490.
Steriade, M., A. Parent, and J. Hada (1984) Thalamic projections of nucleus reticularis thalami of cat: A study using retrograde transport of horserad- ish peroxidase and double fluorescent tracers. J. Comp. Neurol. 229:531- 547.
Steriade, M.. E.G. Jones, and R.R. Llinas (1990) Thalamic Oscillations and Signaling. New York: John Wiley & Sons.
Velayos, J.L., and F. Reinoso-Suarez (1985) Prosencephalic afferents to the mediodorsal thalamic nucleus. J. Comp. Neurol. 242:161-181.
Velayos, J.L., J. Jimenez-Castellanos, Jr., and F. Reinoso-Suarez (1,989) Topographical organization of the projections from the reticular tha- lamic nucleus to the intralaminar and medial thalamic nuclei in the cat. J. Comp. Neurol. 279:457-469.
Weinberg, R. , M. Bentivoglio, K. Phend, D.E. Schmechel, and A. Rustioni ( 1985) A new double-labeling method demonstrates transmitter-specific projections. Neurosci. Lett. 55:349-353.