head direction cell activity in the anterodorsal thalamus requires intact supragenual nuclei
TRANSCRIPT

doi: 10.1152/jn.00295.2012108:2767-2784, 2012. First published 8 August 2012;J Neurophysiol
Benjamin J. Clark, Joel E. Brown and Jeffrey S. Tauberequires intact supragenual nucleiHead direction cell activity in the anterodorsal thalamus
You might find this additional info useful...
69 articles, 27 of which you can access for free at: This article citeshttp://jn.physiology.org/content/108/10/2767.full#ref-list-1
1 other HighWire-hosted articles: This article has been cited by http://jn.physiology.org/content/108/10/2767#cited-by
including high resolution figures, can be found at: Updated information and serviceshttp://jn.physiology.org/content/108/10/2767.full
can be found at: Journal of Neurophysiology about Additional material and informationhttp://www.the-aps.org/publications/jn
This information is current as of May 7, 2013.
http://www.the-aps.org/. 20814-3991. Copyright © 2012 the American Physiological Society. ESSN: 1522-1598. Visit our website attimes a year (twice monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD
publishes original articles on the function of the nervous system. It is published 24Journal of Neurophysiology
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

Head direction cell activity in the anterodorsal thalamus requires intactsupragenual nuclei
Benjamin J. Clark, Joel E. Brown, and Jeffrey S. TaubeDepartment of Psychological and Brain Sciences, Dartmouth College, Hanover, New Hampshire
Submitted 6 April 2012; accepted in final form 5 August 2012
Clark BJ, Brown JE, Taube JS. Head direction cell activity in theanterodorsal thalamus requires intact supragenual nuclei. J Neuro-physiol 108: 2767–2784, 2012. First published August 8, 2012;doi:10.1152/jn.00295.2012.—Neural activity in several limbic areasvaries as a function of the animal’s head direction (HD) in thehorizontal plane. Lesions of the vestibular periphery abolish this HDcell signal, suggesting an essential role for vestibular afference in HDsignal generation. The organization of brain stem pathways conveyingvestibular information to the HD circuit is poorly understood; how-ever, recent anatomical work has identified the supragenual nucleus(SGN) as a putative relay. To test this hypothesis, we made lesions ofthe SGN in rats and screened for HD cells in the anterodorsalthalamus. In animals with complete bilateral lesions, the overall numberof HD cells was significantly reduced relative to control animals. Inanimals with unilateral lesions of the SGN, directional activity waspresent, but the preferred firing directions of these cells wereunstable and less influenced by the rotation of an environmentallandmark. In addition, we found that preferred directions displayedlarge directional shifts when animals foraged for food in a dark-ened environment and when they were navigating from a familiarenvironment to a novel one, suggesting that the SGN plays a criticalrole in projecting essential self-motion (idiothetic) information to theHD cell circuit.
head direction; orientation; place cell; spatial navigation; vestibularsystem
THE ABILITY TO NAVIGATE ACCURATELY is dependent on a networkof cells and pathways that determine an animal’s spatial loca-tion and orientation within an environment (Moser et al. 2008;Taube 2007). A critical component of this network is the headdirection (HD) cells, which specifically fire as a function of theanimal’s HD in the horizontal plane (Taube 2007). Each HDcell is preferentially tuned to a single direction (preferred firingdirection) such that each directional heading is representedwithin a small population of cells, collectively forming a neuralcompass that tracks the moment-to-moment changes in ananimal’s orientation. Preferred firing directions can be an-chored to external cues such that the rotation of salient land-marks can evoke corresponding rotations in the preferred firingdirections (Taube 1995; Taube et al. 1990b). Moreover, pre-ferred directions can be maintained in the absence of visualinformation and when the animal is locomoting into unfamiliarenvironments, suggesting that idiothetic information, such asvestibular and motor cues (efference copy and/or proprioception),can also control directional tuning (Goodridge et al. 1998; Taubeand Burton 1995; Yoder et al. 2011b). HD cell activity hasbeen studied most extensively in the anterodorsal thalamus
(ADN) (Taube 1995), but large populations of HD cells havebeen observed in several other brain regions particularly withinthe limbic system, including the para- and postsubiculum(Boccara et al. 2010; Taube et al. 1990a), medial entorhinalcortex (Sargolini et al. 2006), lateral mammillary nuclei(LMN) (Blair et al. 1998; Stackman and Taube 1998), retro-splenial cortex (Chen et al. 1994; Cho and Sharp 2001), anddorsal tegmental nuclei (DTN) (Sharp et al. 2001b).
Because directional heading can be derived from angularhead velocity information, models of the HD signal havepointed to the vestibular system as a critical input (Mc-Naughton et al. 1991; Redish et al. 1996; reviewed in Taubeand Bassett 2003). Supporting this view, lesions of the periph-eral vestibular system severely disrupt directional firing in theADN (Muir et al. 2009; Stackman and Taube 1997; Yoder andTaube 2009). The organization of brain stem pathways fromthe vestibular system to HD cell circuitry is poorly understood,but anatomical work has identified pathways stemming fromthe medial vestibular nucleus ¡ supragenual nucleus (SGN)and nucleus prepositus hypoglossi (NPH) ¡ DTN ¡ LMN ¡ADN (see Fig. 1; Biazoli et al. 2006; Brown et al. 2005; Taube2007). The DTN and LMN contain neurons that are modulatedby angular head velocity (Bassett and Taube 2001; Blair et al.1998; Stackman and Taube 1998), and lesion studies show thatdamage to either structure abolishes directional activity in theADN (Bassett et al. 2007; Blair et al. 1998). Nonetheless,whether the DTN and LMN receive angular head velocityinformation via the SGN, the nucleus prepositus, or both ispresently unclear.
The SGN, which is located ventral to the cerebellum andfourth ventricle, sends topographically organized output to theDTN and LMN such that each SGN nucleus sends contralateralprojections to the DTN and ipsilateral projections to the LMN(Fig. 1). One interesting feature of this connectivity is thatSGN input to the DTN is much stronger than input to LMN(Biazoli et al. 2006). Thus it is possible that projections to theDTN are more significant with respect to the HD circuit. In twoexperiments, we tested the functional relationship betweenbilateral SGN output to the HD cell circuit, as well as theimportance of the distinct ipsilateral/contralateral output to thetegmento-mammillary circuit. In experiment 1, we producedbilateral lesions electrolytically, thereby completely discon-necting the HD cell circuit of ascending projections from theSGN, and monitored cellular activity downstream (i.e., effer-ently) in the rodent ADN. In experiment 2, we producedunilateral neurotoxic lesions aimed either to the SGN locatedcontralateral to the ADN electrode implant, thereby discon-necting SGN output to DTN, or ipsilateral to the ADN implant,thereby disconnecting output to the LMN (Fig. 1). It was
Address for reprint requests and other correspondence: J. S. Taube, Dept. ofPsychological and Brain Sciences, Dartmouth College, Hanover, NH 03755(e-mail: [email protected]).
J Neurophysiol 108: 2767–2784, 2012.First published August 8, 2012; doi:10.1152/jn.00295.2012.
27670022-3077/12 Copyright © 2012 the American Physiological Societywww.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

hypothesized that lesions to the contralateral SGN would havea greater impact on HD signal processing than ipsilaterallesions. In experiment 1 we show that large bilateral lesions ofthe SGN abolish the HD signal in the ADN, while in experi-ment 2 we show that unilateral lesions of the SGN significantlyimpair the stability of their preferred firing directions. Impor-tantly, the latter result demonstrates that unilateral SGN le-sions, either ipsilateral or contralateral to the ADN electrodeimplant, equally disrupt HD cell activity, particularly the ca-pacity of the HD cell system to accurately update directionalorientation based on idiothetic information. Previous work hasshown that disruption of ascending vestibular input to the HDcell system produces “bursting” cellular activity in the ADN(Muir et al. 2009; Yoder and Taube 2009), which has theappearance of HD cells passing through their preferred direc-tions but is completely uncorrelated with the animal’s currentHD (see Clark and Taube 2012 for review). Here, we confirmthat a subpopulation of ADN neurons also exhibit burstingactivity in animals with SGN lesions, further implicating thisstructure in the processing of idiothetic information. Takentogether, these findings provide the first evidence that the SGNplays a pivotal role in processing the limbic HD cell signal and
its influence over HD signal processing is distributed across theDTN-LMN generative circuit.
METHODS
Subjects
Adult female Long-Evans rats (n � 42) weighing 300–350 g wereused in this experiment. Rats were group housed preoperatively andindividually housed postoperatively in plastic cages and kept on a12:12-h light-dark cycle, with food and water available ad libitum.Subjects were divided into SGN-lesioned (n � 21) and unlesionedcontrol (n � 21) groups. Some of the data from the control group werereported in recent studies (Clark and Taube 2011; Clark et al. 2010)but were collected simultaneously with the present study. All animalprocedures were performed with an institutionally approved Institu-tional Animal Care and Use Committee (IACUC) protocol and incompliance with standards described by the National Institutes ofHealth Guide for the Care and Use of Laboratory Animals.
Lesions and Electrode Implantation
In the present study, the anatomical boundaries of the SGN weredefined on the basis of the plates provided by Paxinos and Watson(1998). It is important to point out that the SGN is sometimesclassified as a rostral extension of the NPH (Klop et al. 2005).However, the anatomical study of Klop et al. was conducted in cats,and because the present study used rodents we confine our discussionof SGN to the anatomical studies that have been conducted in rodents.Furthermore, some recent work has confirmed that the input/outputrelationship of the SGN with HD cell circuitry is particularly distinctfrom the NPH. Specifically, output from the NPH is restricted to theipsilateral DTN (Biazoli et al. 2006; Brown et al. 2005), while outputfrom the SGN is largely directed to the contralateral DTN andipsilateral LMN. Other anatomical studies in rodents (Liu et al. 1984)and rabbits (Hayakawa and Zyo 1985) and other rodent brain atlases(Swanson 1998), as well as a review on the NPH (McCrea 2006), areall in agreement with the anatomical parcellation of SGN and NPH.
To avoid unintentional damage to the DTN or NPH, and tosufficiently damage the SGN, lesions were placed at the followingstereotaxic coordinates: �2 mm posterior to lambda (roughly between10 and 10.3 posterior to bregma) (anterior-posterior), �0.45 mm frommidline (medial-lateral), and �6.9 mm ventral to the surface of thecerebellum (dorsal-ventral) (Paxinos and Watson 1998). Rats receivedbilateral electrolytic (n � 9) lesions of the SGN in experiment 1 andunilateral neurotoxic (n � 12) lesions in experiment 2. We triedperforming several bilateral SGN lesions with a neurotoxin. Unfortu-nately, however, we were unsuccessful, as many of the animalsdied—presumably from spread of the toxin to adjacent brain areascritical for sustaining vital physiological processes. Prior to surgery,animals were anesthetized with Nembutal (40 mg/kg ip) and givenatropine sulfate (5 mg/kg ip) to prevent respiratory distress. Animalswere then placed in a Kopf stereotaxic instrument (David KopfInstruments, Tujunga, CA), and an incision was made to expose theskull. A craniotomy was made over the SGN, and an insect pin orsyringe was lowered to the SGN on one side and left in place for 1min. Electrolytic lesions were produced by passing 0.3 mA of anodaldirect current for 25 s via no. 0 stainless steel insect pins that wereinsulated to within 1 mm of the tip with Epoxylite. The insect pin wasthen removed and placed in the contralateral SGN, where the proce-dure was repeated. Unilateral neurotoxic lesions (left SGN: n � 8;right SGN: n � 4) were produced by injecting 0.15 �l of an 100 mMsolution of N-methyl D-aspartate (NMDA; dissolved in 0.9% saline)into each SGN at a rate of 0.05 �l/min through a blunt-tipped 1-�lHamilton syringe (Hamilton, Reno, NV). To aid diffusion, the syringewas left in place for 3 min. The needle was wiped with distilled water
Fig. 1. Schematic diagram showing the principal connections between thesupragenual nucleus (SGN) and the head direction (HD) cell circuit (reviewedin Taube 2007). The SGN, which is depicted in black, sends projections to thecontralateral dorsal tegmental nucleus (DTN) and the ipsilateral lateral mam-millary nuclei (LMN; Biazoli et al. 2006; Brown et al. 2005), both of whichcontain neurons sensitive to HD and angular head velocity. The connectionsbetween the DTN and LMN are central to the generation of HD cell activity inthe ADN, as well as other cortical and hippocampal areas containing HD cells(Bassett et al. 2007; Blair et al. 1999; Sharp and Koester 2008). Because theSGN receives inputs from the nucleus prepositus hypoglossi (NPH) and themedial vestibular nuclei (MVN), the SGN is a likely source of angular headvelocity information to the DTN. Arrows represent direction of informationflow between connected areas. Although most connections are excitatory,projections from the NPH ¡ DTN and DTN ¡ LMN are thought to beinhibitory and are indicated by broken lines. Note that in the present studyelectrodes were implanted in the ADN of the right hemisphere, which isdepicted in gray.
2768 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

between each injection and was tested to ensure that it was notclogged before being lowered to the next injection site.
As described in previous work from our laboratory (Taube 1995),recording electrodes were implanted in the right ADN of lesioned andcontrol animals. Briefly, a craniotomy was made over the right ADN,and the electrode was implanted at the following coordinates: �1.7 or�1.9 mm posterior to bregma, 1.3 mm right of the midline, and �3.7mm from the cortical surface (Paxinos and Watson 1998). Electrodeswere built as described previously (Kubie 1984) and consisted of abundle of ten 25-�m nichrome wires insulated to the tip. The wireswere threaded through a 26-gauge stainless steel cannula and con-nected to a modified Augat plug that was fixed to the skull with dentalacrylic but could be manipulated in the dorsal/ventral plane. Rats weregiven postsurgical analgesia with buprenorphine (0.015 mg/kg).
Cell Screening Procedures
To allow comparison with prior studies from our laboratory (e.g.,Clark and Taube 2011; Clark et al. 2010; Goodridge and Taube 1997;Taube 1995), a 7-day recovery period was provided before electrodeswere screened for cellular activity. Cell screening was performedwhile rats foraged for 20-mg food pellets (Bio-Serv, Frenchtown, NJ)scattered on the floor of a high-walled gray cylinder (76-cm diameter,50-cm height). A white cue card attached to the inside wall of thecylinder (�100° of arc) served as the sole visual landmark, and blackfloor-to-ceiling curtains surrounding the cylinder minimized extra-maze cues. The cylinder floor was lined with gray paper and waschanged between screening sessions. To detect cell activity, electricalsignals were passed through a field-effect transistor in a source-follower configuration. Signals were amplified by a factor of 10,000–50,000 (Grass Instruments, West Warwick, RI), band-pass filtered(300–10,000 Hz, �3 dB/octave), and sent through a dual-windowdiscriminator (BAK Electronics) for spike discrimination. A colorvideo camera was centered above the cylinder (model XC-711; Sony,Tokyo, Japan). The position and directional orientation of the rat weredetermined by an automated video tracking system (Ebtronics, El-mont, NY). This video tracking hardware provided x and y coordi-nates of red and green light-emitting diodes (LEDs) secured 10 cmapart above the head and back of the animal, respectively. The HD ofthe animal was determined by the relative position of the red andgreen LEDs. Monitoring of cellular activity continued until each ofthe 10 implanted wires had been examined for HD cells or othercellular activity. If no isolatable cells were detected, the electrode wasadvanced 25–50 �m and the animal was returned to its home cage andmonitored again either 4 h later or the next day. If an isolatable cellwas identified, the cell was recorded for an 8-min session and its firingproperties were evaluated. In cases in which HD cells were recorded,the animal was removed from the cylinder and placed in a cardboardbox located outside the curtained-off area (without detaching therecording cable) and the apparatus was prepared for three differenttests of landmark and idiothetic cue processing (Fig. 2). For eachexperiment below, the floor paper was replaced between testingsessions to control for olfactory cues, and white noise was playedfrom an overhead speaker to help mask surrounding auditory cues.
Landmark rotation test. The experiment consisted of three separate8-min sessions (Fig. 2A). First, a standard session (standard 1) wasperformed in which HD cell activity was recorded while the ratforaged for food pellets in the cylinder. This session was followed bya rotation session in which the visual landmark was rotated 90° ineither the clockwise (CW) or counterclockwise (CCW) direction. Therotation of the cue always occurred while the animal was in thecardboard box. Finally, the third session consisted of returning the cuecard to its original position during the first session (standard 2).Animals were given disorientation treatments prior to each recordingsession, which consisted of placing the animal in a cardboard box androtating it slowly (0.2–0.5 Hz) while the experimenter simultaneouslywalked around the testing room twice. This disorientation was used to
prevent the animal from using idiothetic cues to maintain orientationfrom one session to the next.
Dark test. This test consisted of recording HD cell activity with thevisual landmark removed and the room lights turned off (Fig. 2A).Prior to the dark test, the animals were removed from the cylinder andplaced in a cardboard box while the floor paper was replaced and thecue card removed from the cylinder. To start the test, the room lightswere turned off and the rat was gently placed in the cylinder andallowed to forage for food pellets for 8 min. Animals were notdisoriented prior to being placed in the cylinder for dark testing. Totrack the HD of the rat, the red and green LEDs remained on duringthe dark session. Although rats cannot see well in the red colorspectrum (Neitz and Jacobs 1986), they can see green light. It isimportant to note, however, that the gray cylinder was largely homo-geneous and did not contain obvious environmental cues that could beused for orientation. Thus it is likely that animals, in large part, reliedon idiothetic cues to maintain orientation in darkened conditions(Goodridge et al. 1998).
Standard 3. After the 8-min dark test was complete, animals wereremoved from the cylinder and a final 8-min standard session (stan-dard 3) was conducted in which the room lights were turned back on,the floor paper replaced, and the cue card returned to its originalposition from the previous standard sessions (Fig. 2A). Rats weregiven a disorientation treatment before being placed in the cylinder forstandard 3.
Dual-chamber test. Figure 2B illustrates an overhead view of thedual-chamber apparatus, which is composed of two gray-coloredcompartments, a cylinder similar to those used in the previous exper-iments (76 cm in diameter) and a rectangle (51 cm � 69 cm),interconnected by a narrow U-shaped passageway (41 cm � 15 cm).The walls along one side of the alleyway were slanted by �15° inorder for the overhead video camera to view all areas of the passage-way. A section of wall in each enclosure could be removed to controlaccess between the two compartments and passageway. The walls ofthe rectangle were featureless except for a white cue card positionedalong the wall such that it was rotated 90° CCW with respect to thecue card in the cylindrical compartment. The floors of the cylinder andrectangle were composed of gray photographic backdrop paper thatwas changed between animals; however, the passageway contained agray painted wooden floor.
Landmark Rotation and Dark test
Dual-Chamber Test
A
B
Rectangle (Novel)
Cylinder 2 (Familiar)
Cylinder 1 (Familiar)
Standard 1 Standard 2 Dark test Rotation Standard 3
Fig. 2. A: overhead view of the landmark rotation and dark test sessions. Eachsession, except the dark test, was separated by disorientation treatment.B: overhead view of the dual-chamber test sessions. The door to the rectanglewas closed when the animal entered the rectangle. The door to the cylinder wasclosed during both cylinder sessions (cylinder 1 and cylinder 2). Because therats had no prior experience in the rectangle enclosure, this environment wasconsidered novel.
2769IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

The cylinder enclosure in the dual-chamber apparatus was similarto the cylinder in which the rats had been trained and screened in forseveral weeks; thus it was considered a familiar environment to theanimals. The passageway and the rectangle represented a novelenvironment because the animals had no previous exposure to them.HD cells in control animals typically maintain a similar preferredfiring direction between both compartments of the dual-chamberapparatus (Taube and Burton 1995), and it is has been shown thatidiothetic cues available during the animal’s journey from the cylinderto the rectangle are crucial for maintaining a stable preferred direction(Stackman et al. 2003). The procedure for the dual-chamber experi-ment consisted of three phases. First, the animal was placed in thecylinder portion of the apparatus with the door to the passagewayclosed (cylinder 1 session). After the animal was allowed to forage forfood pellets scattered on the floor for 4 min, the cylinder compartmentdoor was opened and the animal was free to walk into the passagewayand enter the rectangle. Upon the animal entering the rectangle, thehidden door was closed, trapping the animal inside the rectangleenvironment while the activity of the HD cell was monitored for 10min (rectangle session). After the rectangle session, the door to therectangular compartment was opened and the rats were allowed tolocomote back through the passageway to the cylinder, where theywere again trapped while HD cell activity was monitored for another4 min (cylinder 2 session). The dual-chamber apparatus experimentwas conducted only once per animal since the task requires therectangular compartment to be novel. Thus only one HD cell record-ing session was collected per animal in this task.
Data Analysis
An automated video computer tracking system, sampling at 60 Hz,recorded the number of spikes occurring during each sample periodwhile simultaneously recording the x, y positions of each LED. Dataanalysis was accomplished off-line with custom software (LabVIEW,National Instruments, Austin, TX). The animal’s HD was sorted intosixty 6° bins. Neural activity was classified as HD cell activity whenthe firing clearly correlated with a single direction, independent of theanimal’s ongoing behavior. To determine whether putative HD cellswere significantly modulated by the animal’s directional heading, weperformed Rayleigh tests (Batschelet 1981) on the recorded cell’sfiring rate vs. HD tuning curve (Bassett et al. 2007; Muir et al. 2009;Yoder and Taube 2009). This statistical test involves computing themean vector length, r, based on the cell’s firing over 360° (Batschelet1981). Mean vector lengths range between 0 and 1, with higher valuesindicating that spike occurrence is clustered around a particulardirection. The critical significance level of r is then determined by thenumber of observations (which was defined as the sum of all firingrates from the 60 directional bins), and if the r value meets thissignificance level, the distribution is considered to be directional.Similar to recent studies, we adopted a criterion of r � 0.40 for a cellto be considered directionally modulated, although r values from“classic” HD cells typically fall in the range of 0.5–0.9 (Muir et al.2009; Yoder and Taube 2009). HD cells from lesioned and controlanimals were compared on several basic directional characteristicsmeasured during the standard 1 session: peak firing rate, backgroundfiring rate, directional firing range, directional information content,and anticipatory time interval. The preferred direction was defined asthe directional bin with the highest firing rate. The peak firing rate wasthe firing rate corresponding to the preferred direction. The directionalfiring range was defined as the width at the base of a triangle fit to afiring rate vs. HD plot. The background firing rate was the mean firingrate of all bins �18° outside the cell’s directional firing range.Directional information content is a measure of how many bits of HDinformation is conveyed by each spike (Skaggs et al. 1993) and wascalculated by the following formula: directional information content ��pi(li/l)log2(li/l), where pi is the probability that the head pointed in
the ith directional bin, li is the mean firing rate for bin i, and l is themean firing rate across all directional bins.
The anticipatory time interval is a measure of the amount of timethat cell firing best predicts where the animal will be pointing its headin the future. Previous work has estimated that the activity of ADNHD cells anticipates future head directions by �25 ms (Blair andSharp 1995; Taube and Muller 1998). We compared the anticipatorytime interval of cells from control and lesioned animals, using themethods of Blair and Sharp (1995). First, firing rate vs. HD plots wereconstructed for each HD cell by dividing the 360° directional rangeinto sixty 6° bins and then calculating the average firing rate for eachbin. Firing rate vs. HD plots were then constructed for CW and CCWdirections, and the difference between the preferred firing directionsfor the two functions (i.e., separation angle) was determined. Thespike record was then shifted forward and backward in time in stepsof 16.67 ms (the maximum temporal resolution of the recordinghardware), and the separation angle between the CW and CCWfunctions for head movements �90°/s was computed for each shift.The spike series was shifted incrementally �6 times (�100 ms)relative to the HD series, providing 13 values of CW-CCW separationangles. A scattergram was then constructed from the 13 CW-CCWseparation angles and their corresponding time shift. The x-interceptof the best-fit line of this plot is referred to as the anticipatory timeinterval and is equivalent to the amount of time that the spike serieshas to be shifted to achieve overlapping CW and CCW functions.Cells with insufficient CW and CCW sampling (�1 directional binwith no data) were not included in this analysis. With this criterion,nine cells were removed from the control group and six cells wereremoved from the lesion group for the analysis conducted on session1 data in experiment 2.
The spike trains of HD cells and in some cases nondirectional cellsgenerally included periods of high activity resembling bursts punctu-ated by periods of low activity. In the case of HD cells, the occurrenceof bursting activity is nonperiodic because the spikes are modulatedby the animal’s HD. Thus to quantify the bursting characteristics ofHD cells as well as nondirectional cells, we used a “burst index”measure introduced by Yoder and Taube (2009). The burst indexcomputes the amount of time a cell fires in high-frequency bursts oris inactive. Burst index scores are measured on a scale of 0 to 1, withhigher values representing cells that are “bursting” and lower valuesrepresenting cells that fire at a relatively constant rate. For thismeasure, spikes were sorted into 1-s bins from the beginning to theend of the recording session. The burst index was calculated using thefollowing formula: burst index � (no. of bins � 1.75 � mean firingrate) � (no. of bins � 0.25 � mean firing rate)/no. of total bins. Themean firing rate was defined as the total number of spikes in thesession divided by the total time.
Circular statistics (Batschelet 1981) were used to determine thestability of the directional signal between the landmark rotationsessions, standard sessions, and the three dual-chamber sessions.Angular shift scores across testing sessions were calculated with across-correlation method. This approach involves shifting the firingrate vs. HD function of the first session in 6° increments whilecorrelating this shifted function with the nonshifted function from theother session. The amount of shift required to produce the maximalPearson r correlation between the two sessions is defined as theangular shift score between the sessions. These values were thensubjected to Rayleigh tests to determine whether the scores weredistributed randomly or the preferred directions tended to shift in thesame direction and amount. ANOVAs and t-tests were utilized forgroup comparisons of the basic directional characteristics and abso-lute angular shift scores produced during the different test sessions.For ANOVAs, violations of homogeneity of variance were correctedwith Welch’s F-test. All statistical analyses were performed withSPSS (Statistical Package for the Social Sciences) version 20.0.
2770 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

Histology
Animals were deeply anesthetized with an overdose of pentobar-bital sodium. The locations of the recording electrodes were markedby passing anodal current through an electrode wire (20 �A, 15 s) fora Prussian blue reaction. The rats were perfused intracardially withsaline followed by a 10% formalin solution. Brains were removedfrom the skulls and postfixed in a 10% formalin solution with 2%potassium ferrocyanide for 24 h. The brains were cryoprotected in a20% sucrose solution for at least 48 h and then frozen and sectionedin the coronal plane in 30-�m slices with a cryostat. Every sectionthrough the SGN was taken, as well as every other section through theADN. Sections were mounted on glass slides, stained with thionin,and examined under a microscope to determine electrode location andextent of the lesion.
Electrolytic damage to the SGN resulted in the complete loss oftissue residing above the dorsal surface of the VIIth cranial nerve, andwas therefore easily quantified (i.e., if no tissue was observed in thisregion then the SGN was considered fully lesioned). SuccessfulNMDA lesions were determined on the basis of a marked loss ofneurons and a significant increase in gliosis in the SGN region. Toquantify the volume of remaining healthy SGN tissue (i.e., thepresence of large neurons and absence of gliosis), digital images werecaptured at two rostral-caudal levels (10 mm and 10.3 mm posteriorto bregma). The area of undamaged tissue in the SGN was calculated ateach rostral-caudal level with ImageJ software (http://rsb.info.nih.gov/ij/index.html). Once the area of undamaged tissue was calculated, thearea of spared tissue was summed across the two sections andcompared with the average area measured in control rats. The totalamount of damage was calculated with the following formula: tissuedamaged (%) � [average area of SGN in control rats (pixels2) � totalarea of spared SGN tissue in lesioned rats (pixels2)/average area ofSGN in control rats (pixels2)] � 100.
The extent of the recording electrode tracks through the ADN wasvisible from the Prussian blue marks at the electrode tips. The positionof the ventral-most point relative to the boundaries of the ADN wasused to estimate the portion of the screening record that could haveconceivably fallen within the ADN. All isolated cells were evaluated
for directional modulation, but only cells recorded inside the �1 to1.5 mm dorsal-ventral sampling range of the ADN were consideredpart of the experimental sample for the purposes of estimating theincidence rate of directional modulation.
RESULTS
Experiment 1: Cellular Activity in ADN of Rats withBilateral Electrolytic Lesions of SGN
Histology. Figure 3 depicts a coronal section through theSGN of representative control (Fig. 3B) and electrolytic le-sioned (Fig. 3, C and D) animals. Overall, lesions of the SGNwere large, ranging from 77.3% to 100% of the total volume ofSGN (mean � SE: 95.5 � 2.6%). Four of nine lesionedanimals showed evidence of healthy SGN tissue, which inthree cases was confined unilaterally (ipsilateral to the elec-trode implant � 1; contralateral to the electrode implant � 2)and in one case bilaterally. Lesions frequently produced unin-tentional damage to brain stem structures located ventral to theSGN, including the central gray of the pons, olivocochlearbundle, abducens nuclei, genu of the facial nerve, and themedial longitudinal fasciculus (mlf). More rarely, the lesionsextended dorsal to the SGN and into the cerebellum, specifi-cally the first cerebellar lobule (e.g., Fig. 3C). Importantly,damage to the mlf was large (e.g., Fig. 3D) and possiblycomplete in five animals. Because the mlf is known to conveyvestibular information to several brain stem and thalamicnuclei, it is possible that fibers carrying vestibular informationto the DTN were damaged. Nevertheless, Frohardt et al. (2006)described two rats that received complete transection of the mlfand observed that the animals were unimpaired in a navigationtask that requires an intact HD cell circuit. Although this resultsuggests functional independence between the HD cell systemand the mlf, below we provide some qualitative comparisonsbetween animals with and without mlf damage.
Fig. 3. A: plate reproduced with permissionfrom Paxinos and Watson (1998) showing theSGN at 10.3 mm posterior to bregma. B: en-larged view of the boxed area in A showingthe SGN (circled region). Also visible areother structures that were sometimes damagedwith the lesion: the abducens nuclei (6), genuof the facial nerve (g7), and the first cerebellarlobule (c1). mlf, Medial longitudinal fascicu-lus. Scale bar, 500 �m. C and D: representa-tive sections from electrolytic lesioned rats(sg1 and bc228, respectively) with completeloss of tissue in the immediate vicinity of theSGN. The horizontal arrow in C shows gliosisextending ventrally into the abducens nu-cleus, and the vertical arrow in C points todamage of the first cerebellar lobule. Thevertical arrows in D show the extent of dam-age ventral to the SGN, which included theentire mlf in this animal. The first cerebellarlobule is not visible in this section but was notdamaged in this animal.
2771IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

SGN lesions did not extend laterally into the vestibularnuclei, but they often included structures located rostral andcaudal. Most important of the rostral structures is the posteriorDTN, which in some cases (n � 3) received significant damage(�50%). Unlike the central zone of the DTN, vestibularinformation does not converge on the posterior DTN becauseof the absence of direct inputs from SGN or vestibular nuclei(Biazoli et al. 2006). However, it does provide a strong pro-jection to the HD circuit (Brown et al. 2005) and reportedlycontains neurons sensitive to angular head velocity (Sharp etal. 2001b). In light of this consideration, we also provide belowsome qualitative comparisons of this small subgroup of ani-mals with other rats in the lesioned group. The NPH, which ispositioned caudal to the SGN, received minor damage (�10–20%) in most animals. Table 1 summarizes the extent oflesions to the SGN, the amount of damage to posterior DTNand mlf, and the firing characteristics of ADN cells for eachanimal included in the experiment. Finally, it is important tomention that we did not observe any noticeable changes inposture, locomotion, or oculomotor function after SGN lesions.
Electrode arrays were judged to have advanced completelythrough the ADN in all animals. In lesioned rats, electrodesgenerally passed through the ADN between 1.3 and 1.8 mmposterior to bregma (Paxinos and Watson 1998). Similar elec-trode placements were observed in control animals. Figure 4shows the electrode location in a representative section from anelectrolytic lesioned rat.
HD cells. Figure 5 plots the firing rate vs. HD functions ofrepresentative HD cells recorded in the control group and allHD cells recorded in the lesioned group. Overall, only 4 of 76neurons (5.3%) were identified as HD cells within the dorsal-ventral extent of the ADN in the lesioned group (Fig. 5B)(Rayleigh r � 0.4). The remaining cells were classified asnondirectional. In contrast to the small overall percentage ofHD cells recorded in lesioned rats, 79 of 163 (48.5%) wereidentified as HD cells in control animals (Fig. 5A; meanRayleigh r � 0.731 � 0.017). A mean comparison of thepercentage of HD cells/rat confirmed a significant differencebetween lesioned (7.3 � 3.77%) and control [49.8 � 4.19%;t(26) � �6.39, P � 0.001] groups. One cell that was notinitially classified as directional, and did not pass the Rayleighcriterion (r � 0.35), showed some evidence of directionalmodulation (Fig. 5C, far left); however, even with this cellincluded, the group differences remained highly significant[t(26) � 6.31, P � 0.001]. Importantly, each HD cell recorded
in the lesioned group came from animals with incompletelesions (see Table 1), suggesting that small populations of SGNneurons can sustain directional activity in the ADN. Neverthe-less, even with small amounts of remaining SGN tissue, therewas a significant difference between this partial lesion sub-group and control animals in the percentage of HD cells/rat[t(21) � 3.36, P � 0.003]. With respect to individual differ-ences in the lesioned group, there was no obvious influence inthe amount or location (contra- or ipsilateral) of spared SGNtissue. Furthermore, there was no obvious influence of damageto the mlf or posterior DTN. For instance, rat 2 had a completeSGN lesion with no significant damage to mlf or posterior
Table 1. Summary of histological results and incidence of HD and bursting cell activity in animals with bilateral electrolytic lesions ofthe SGN
Rat Lesion Size, % Tissue Sparing Additional Damage Total Cells HD Cells Bursting Cells
jb41 100 pDTN, mlf 12 0 1sg1 100 9 0 0sg2 96.3 Right SGN 6 1 1sg3 77.3 Left SGN 7 2 0sg4 100 mlf 7 0 1sg6 96.4 Left SGN mlf 5 1 2bc227 100 pDTN, mlf 7 0 0bc228 100 pDTN, mlf 7 0 0bc229 89.7 Bilateral 16 0 10
All recordings were conducted in the right anterodorsal thalamus (ADN). Column listing additional damage lists only the regions that may have an influenceon head direction (HD) cell activity because of confirmed connectivity. SGN, supragenual nucleus; mlf, medial longitudinal fasciculus; pDTN, posterior dorsaltegmental nucleus.
Fig. 4. A: plate reproduced with permission from Paxinos and Watson (1998)showing the ADN at �1.4 mm relative to bregma. B: enlarged view of theboxed region in A showing the electrode wire tracks through the ADN (arrowspointing left) and the Prussian blue mark below the ADN (arrow pointingright) in a representative electrolytically lesioned rat (sg1). Scale bar, 500 �m.
2772 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

DTN, and, importantly, no HD cells were recorded in theADN. Taken together, damage specific to the SGN is the mostlikely cause of the abolished HD cell signal.
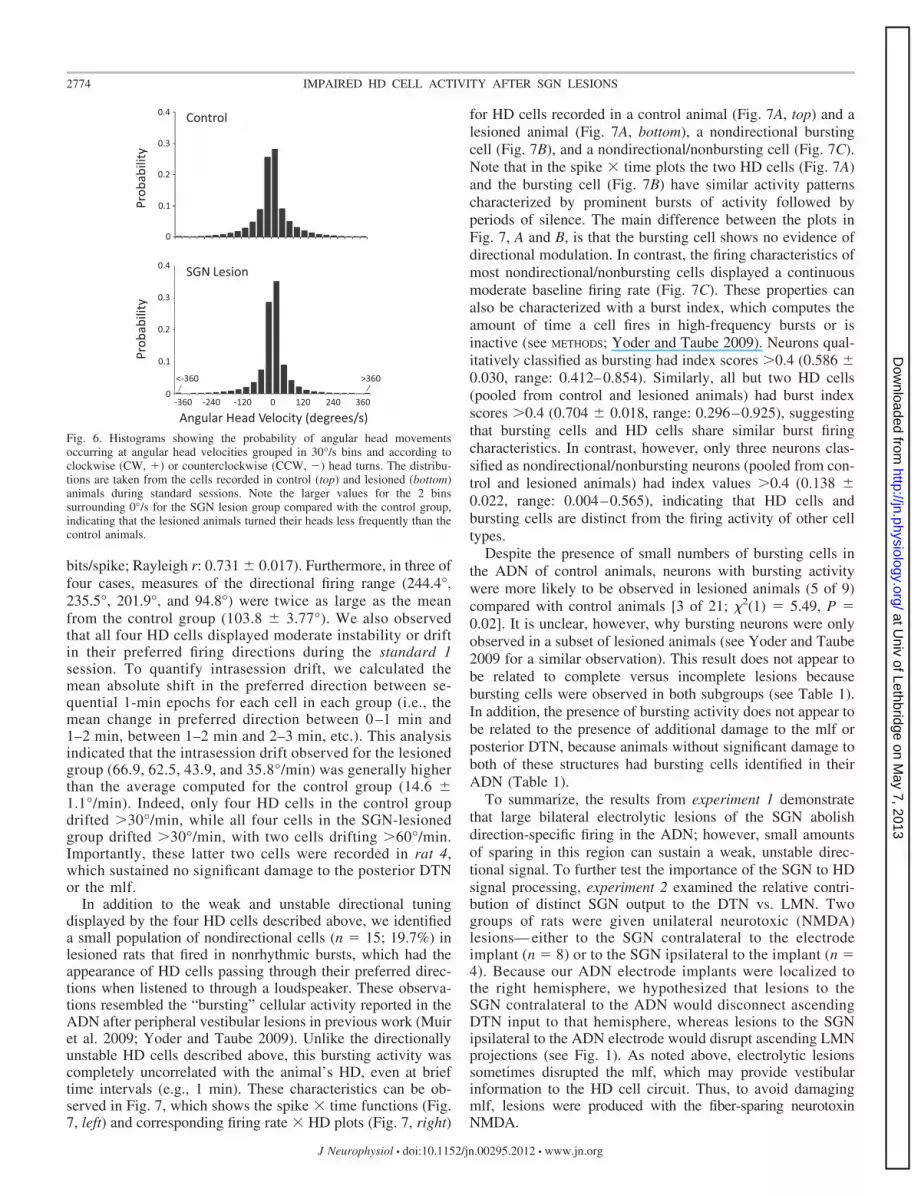
To determine whether differences in the frequency of dif-ferent angular head velocities could account for the weakdirectional signal, we examined the distribution of angularhead velocities over the course of the standard session for cells(HD and nondirectional cells) recorded in each group (Bassettet al. 2005). Figure 6 plots this distribution and shows thatanimals with SGN lesions had a higher probability of lowangular head velocities (between �30°/s and �30°/s) com-pared with control animals. This observation is confirmed by asignificant group difference in the instantaneous angular headvelocity between control (34.7 � 1.09°/s) and lesioned (24.6 �2.29°/s) animals [t(112) � 0.721, P � 0.001]. Although thisfinding supports the conclusion that differences in angular headvelocity may have contributed to the results of the present
study, previous work has shown that angular head velocityaccounts for only a small fraction of the firing properties of HDcells (Taube 1995). Furthermore, it is important to note thatalthough the lesioned group generally had lower angular headvelocities than the control group, all angular head velocities,including high ones, were present in the lesioned group. Thusit is more likely that the loss of directional tuning stems fromthe disruption of SGN input rather than the slower angular headmovements of lesioned animals.
Although a small sample of HD cells was observed inlesioned rats, the directional specificity displayed by theseneurons was disrupted (compare Fig. 5, A and B). In particular,the four HD cells in the lesioned group had measures ofdirectional information content (0.798, 0.772, 0.553, and 0.391bits/spike) and Rayleigh r values (0.608, 0.589, 0.549, and0.418) that were lower than the mean values computed for thecontrol group (directional information content: 1.33 � 0.07
Fig. 5. A: firing rate vs. HD plots for representative HD cells recorded in control animals. Cells shown have Rayleigh r values (inset) similar in magnitude tothe group average (r � 0.731). B: firing rate vs. HD plots are shown for all cells recorded in lesioned animals that were classified as directionally modulatedaccording to a Rayleigh r value of �0.40. C: firing rate vs. HD functions for cells from lesioned animals that approached, but did not surpass, the Rayleigh cutoff.Note that 1 cell had some modest directional modulation (C, top left); however, directional modulation was largely absent for the remaining cells.
2773IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from
bits/spike; Rayleigh r: 0.731 � 0.017). Furthermore, in three offour cases, measures of the directional firing range (244.4°,235.5°, 201.9°, and 94.8°) were twice as large as the meanfrom the control group (103.8 � 3.77°). We also observedthat all four HD cells displayed moderate instability or driftin their preferred firing directions during the standard 1session. To quantify intrasession drift, we calculated themean absolute shift in the preferred direction between se-quential 1-min epochs for each cell in each group (i.e., themean change in preferred direction between 0 –1 min and1–2 min, between 1–2 min and 2–3 min, etc.). This analysisindicated that the intrasession drift observed for the lesionedgroup (66.9, 62.5, 43.9, and 35.8°/min) was generally higherthan the average computed for the control group (14.6 �1.1°/min). Indeed, only four HD cells in the control groupdrifted �30°/min, while all four cells in the SGN-lesionedgroup drifted �30°/min, with two cells drifting �60°/min.Importantly, these latter two cells were recorded in rat 4,which sustained no significant damage to the posterior DTNor the mlf.
In addition to the weak and unstable directional tuningdisplayed by the four HD cells described above, we identifieda small population of nondirectional cells (n � 15; 19.7%) inlesioned rats that fired in nonrhythmic bursts, which had theappearance of HD cells passing through their preferred direc-tions when listened to through a loudspeaker. These observa-tions resembled the “bursting” cellular activity reported in theADN after peripheral vestibular lesions in previous work (Muiret al. 2009; Yoder and Taube 2009). Unlike the directionallyunstable HD cells described above, this bursting activity wascompletely uncorrelated with the animal’s HD, even at brieftime intervals (e.g., 1 min). These characteristics can be ob-served in Fig. 7, which shows the spike � time functions (Fig.7, left) and corresponding firing rate � HD plots (Fig. 7, right)
for HD cells recorded in a control animal (Fig. 7A, top) and alesioned animal (Fig. 7A, bottom), a nondirectional burstingcell (Fig. 7B), and a nondirectional/nonbursting cell (Fig. 7C).Note that in the spike � time plots the two HD cells (Fig. 7A)and the bursting cell (Fig. 7B) have similar activity patternscharacterized by prominent bursts of activity followed byperiods of silence. The main difference between the plots inFig. 7, A and B, is that the bursting cell shows no evidence ofdirectional modulation. In contrast, the firing characteristics ofmost nondirectional/nonbursting cells displayed a continuousmoderate baseline firing rate (Fig. 7C). These properties canalso be characterized with a burst index, which computes theamount of time a cell fires in high-frequency bursts or isinactive (see METHODS; Yoder and Taube 2009). Neurons qual-itatively classified as bursting had index scores �0.4 (0.586 �0.030, range: 0.412–0.854). Similarly, all but two HD cells(pooled from control and lesioned animals) had burst indexscores �0.4 (0.704 � 0.018, range: 0.296–0.925), suggestingthat bursting cells and HD cells share similar burst firingcharacteristics. In contrast, however, only three neurons clas-sified as nondirectional/nonbursting neurons (pooled from con-trol and lesioned animals) had index values �0.4 (0.138 �0.022, range: 0.004–0.565), indicating that HD cells andbursting cells are distinct from the firing activity of other celltypes.
Despite the presence of small numbers of bursting cells inthe ADN of control animals, neurons with bursting activitywere more likely to be observed in lesioned animals (5 of 9)compared with control animals [3 of 21; �2(1) � 5.49, P �0.02]. It is unclear, however, why bursting neurons were onlyobserved in a subset of lesioned animals (see Yoder and Taube2009 for a similar observation). This result does not appear tobe related to complete versus incomplete lesions becausebursting cells were observed in both subgroups (see Table 1).In addition, the presence of bursting activity does not appear tobe related to the presence of additional damage to the mlf orposterior DTN, because animals without significant damage toboth of these structures had bursting cells identified in theirADN (Table 1).
To summarize, the results from experiment 1 demonstratethat large bilateral electrolytic lesions of the SGN abolishdirection-specific firing in the ADN; however, small amountsof sparing in this region can sustain a weak, unstable direc-tional signal. To further test the importance of the SGN to HDsignal processing, experiment 2 examined the relative contri-bution of distinct SGN output to the DTN vs. LMN. Twogroups of rats were given unilateral neurotoxic (NMDA)lesions— either to the SGN contralateral to the electrodeimplant (n � 8) or to the SGN ipsilateral to the implant (n �4). Because our ADN electrode implants were localized tothe right hemisphere, we hypothesized that lesions to theSGN contralateral to the ADN would disconnect ascendingDTN input to that hemisphere, whereas lesions to the SGNipsilateral to the ADN electrode would disrupt ascending LMNprojections (see Fig. 1). As noted above, electrolytic lesionssometimes disrupted the mlf, which may provide vestibularinformation to the HD cell circuit. Thus, to avoid damagingmlf, lesions were produced with the fiber-sparing neurotoxinNMDA.
Fig. 6. Histograms showing the probability of angular head movementsoccurring at angular head velocities grouped in 30°/s bins and according toclockwise (CW, �) or counterclockwise (CCW, �) head turns. The distribu-tions are taken from the cells recorded in control (top) and lesioned (bottom)animals during standard sessions. Note the larger values for the 2 binssurrounding 0°/s for the SGN lesion group compared with the control group,indicating that the lesioned animals turned their heads less frequently than thecontrol animals.
2774 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

Experiment 2: Cellular Activity in Rats with UnilateralNeurotoxic Lesions of SGN
Histology. Overall, neurotoxic lesions produced significantdamage (�60%) to the unilateral SGN in all animals (n � 12)(Fig. 8). Unilateral damage was large in six animals, rangingfrom 90% to 100% (4 contralateral and 2 ipsilateral). In theremaining six animals, the lesions were smaller, ranging from60% to 80% (4 contralateral and 2 ipsilateral). For mostanimals in this latter group, the lesions were smaller becausethe syringe was unintentionally placed either slightly rostral orcaudal (approximately �0.75 mm), which in three cases hadthe effect of producing unilateral damage to the posterior DTN(�20%) and in two cases damaged the NPH unilaterally(�20%). In other animals, damage to the posterior DTN andNPH was minimal (�10%). In addition, partial unilateraldamage (�20–30%) was observed in the abducens nucleus inmost animals. Damage to the central gray of the pons wasobserved unilaterally in most animals; however, this damagewas unavoidable because of its close proximity to the SGN.Importantly, the mlf was not damaged in any of the animals,thereby precluding the possibility that ascending vestibularaxons through this pathway contributed to the results reportedbelow. Electrode arrays were judged to have advanced com-pletely through the ADN in all animals. Table 2 summarizesthe extent and hemisphere of SGN lesions, damage to otherbrain areas, and firing characteristics of ADN cells for eachanimal included in experiment 2.
HD cells. A total of 73 cells were isolated in rats withcontralateral lesions, and 33 cells were isolated in ipsilaterallylesioned rats. Of these cells, 34 cells were classified as direc-tionally modulated in contralaterally lesioned animals (46.6%)
and 17 cells in ipsilaterally lesioned animals (51.5%). Aone-way ANOVA on the percentage of HD cells/rat failed toreveal a significant group effect between contralateral (41.7 �7.8%) and ipsilateral (51.0 � 25.0%) lesion groups and be-tween the lesion and control groups [49.8 � 4.2%; F(2,6.5) �0.39, P � 0.69].
Measures of HD cell firing characteristics failed to identifysignificant group differences on measures of directional infor-mation content [contralateral: 1.39 � 0.26, ipsilateral: 1.03 �0.12, control: 1.33 � 0.07 bits/spike; F(2,96) � 0.610, P �0.545], peak firing rate [contralateral: 45.7 � 5.5, ipsilateral:30.7 � 5.0, control: 42.6 � 2.8 spikes/s; F(2,96) � 1.47, P �0.235], and background firing rate [contralateral: 2.51 � 0.48,ipsilateral: 1.75 � 0.32, control: 1.61 � 0.20 spikes/s;F(2,28.3) � 1.45, P � 0.252]. However, an ANOVA revealeda significant difference between the lesioned and controlgroups on measures of directional firing range [contralateral:122.2 � 7.2°, ipsilateral: 105.4 � 8.5°, control 103.8 � 3.8°;F(2,96) � 3.27, P � 0.042]. Subsequent t-tests revealed thatthis effect was largely mediated by the mean difference be-tween the contralateral lesion and control groups [t(94) � 2.52,P � 0.013] and not the difference between ipsilateral lesionand control groups [t(94) � �0.144, P � 0.89] or between thecontralateral and ipsilateral groups [t(94) � 1.43, P � 0.16].We also identified a significant difference between control andlesion groups on anticipatory time interval measures [contralat-eral: 64.2 � 11.3, ipsilateral: 0.23 � 28.4, control 30.8 � 7.0ms; F(2,81) � 5.16, P � 0.008]. Again, subsequent t-testsdetermined that this effect was largely mediated by the meandifference between the contralateral lesion and control groups[t(79) � �2.41, P � 0.018] and not the difference between
Fig. 7. A, left: plot depicting a spike vs. time (s) functionfor a representative HD cell in a control (top) and anSGN-lesioned (bottom) rat. Right: each cell’s corre-sponding firing rate vs. HD plot (inset, Rayleigh r value).B: spike vs. time (left) and firing rate vs. HD (right) plotsfor a cell classified as bursting. Note that this cell has atemporal firing pattern similar to the HD cells in A;however, the cell was not directionally modulated, asindicated by its low Rayleigh r value. C: spike vs. time(left) and firing rate vs. HD (right) plots for a cellidentified as nondirectional/nonbursting. Note that thiscell’s burst index score is lower than the HD cells in Aand the bursting cell in B.
2775IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

ipsilateral lesion and control groups [t(79) � �1.52, P �0.13]. There was a significant mean difference between thecontralateral and ipsilateral groups [t(79) � 2.95, P � 0.004],indicating that HD cells in contralaterally lesioned animalsanticipated to a greater extent compared with ipsilaterallylesioned animals. Similar to experiment 1, we also observedseveral cells that contained burst firing patterns in unilaterallylesioned animals (n � 13; 12.3%). The percentage of burstingcells in contralaterally lesioned animals (n � 12; 16.4%) wasgreater than in ipsilaterally lesioned animals (n � 1; 3.0%);however, a �2-test failed to indicate a significant differencebetween groups [�2(1) � 1.07, P � 0.30].
In summary, the analyses on the basic firing propertiesindicate that small unilateral lesions, in particular lesionscontralateral to the recording hemisphere, significantly in-crease the range of directions cells fire in. It is possible that thisreduction in directional specificity reflects a general deficit inspatial updating by environmental landmarks and/or by idio-thetic cues. These possibilities are tested below.
Landmark control. To determine whether unilateral SGNlesions impaired landmark control over ADN HD cells, wemonitored cell activity after a white cue card was rotated 90°CW or CCW along the wall of the testing cylinder (Fig. 2A). Atotal of 16 cells in the contralateral lesion group, 10 in theipsilateral lesion group, and 20 from the control group wererecorded during the landmark test. The results are plotted incircular histograms in Fig. 9A, showing the amount of angularshift in the cell’s preferred firing direction between the stan-dard 1 and rotation sessions and between the standard 1 andstandard 2 sessions. Overall, the white cue card demonstratedstrong control over HD cells in control animals, where theirpreferred directions shifted a similar angular distance andalways in the same direction as the landmark. In contrast,however, unilateral lesions of the SGN appeared to reduce thecontrol by the landmark. For instance, over half of the 26 HDcells recorded in lesioned animals underrotated their preferreddirections �30° (n � 6), did not rotate (�12°; n � 4), rotatedin the wrong direction (n � 3), or were clearly directionallyunstable for large portions of the 8-min rotation session (n �3). Figure 10 shows three representative firing rate vs. HDfunctions from rats with SGN lesions recorded during thelandmark rotation series, as well as an HD � time plot for acell recorded in a lesioned animal that displayed directionalinstability during the rotation session. Mean comparisons onthe amount of deviation from the expected 90° shifts (com-puted only for directionally stable cells; n � 23) confirmed asignificant group difference [F(2,13.8) � 4.46, P � 0.032],which, in large part, was influenced by the difference betweenthe contralateral lesion (58.8 � 16.1°) and control (15.3 �2.6°) groups [t(14.7) � 2.65, P � 0.02]. Although the meandeviation did not significantly differ between control andipsilaterally lesioned animals [t(8) � 1.60, P � 0.15], it isimportant to note that deviation values were much larger forHD cells recorded in ipsilaterally lesioned animals (31.5 �9.8°) compared with control animals. Moreover, over half ofthe 10 HD cells recorded in ipsilaterally lesioned animalsshowed either directional instability during the rotation ses-sions (n � 2) or underrotation in their preferred directions (n �4) or the cell’s preferred direction did not rotate at all (n � 1).Rayleigh analyses on the angular shift values confirm theseobservations showing that the mean vector length is signifi-cantly longer for the control group (r � 0.952) compared withthe lesioned group (r � 0.623; P � 0.05). However, it isimportant to point out that the distributions of angular shiftvalues were significantly clustered for both groups (P �0.001). Finally, a t-test did not indicate a significant differencein the mean shifts between the contralateral and ipsilaterallesion groups [t(20.6) � 1.45, P � 0.16].
We further tested the amount of control the cue card hadover HD cells in standard 2 by returning the cue card to itsoriginal angular orientation in the testing cylinder. In general,the preferred directions of HD cells recorded in control animalsshifted back to their original orientation (Fig. 9B). Similarly,
Fig. 8. A: section at �10.3 mm relative to bregma reproduced with permissionfrom Paxinos and Watson (1998). B: corresponding section from a rat withcomplete neurotoxic damage to the right SGN (ipsilateral to the electrodeimplant; jb25). Note that complete sparing of the left SGN occurred in thisanimal. Scale bar, 500 �m. C: section from an animal with complete neuro-toxic damage to the left SGN (contralateral to the electrode implant; jb28).Note that the right SGN is completely spared. Horizontal arrows in B and Cindicate regions where the lesion extended ventrally.
2776 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

the preferred direction of HD cells recorded in lesioned ani-mals also showed a high degree of landmark control byreturning to their original orientation in the majority of thetesting sessions (e.g., Fig. 10A, right, and Fig. 10B, right). Thisobservation is supported by a nonsignificant ANOVA conductedon the deviation from the expected return shift [F(2, 15) � 2.40,
P � 0.12]. Despite this nonsignificant result between thecontrol and lesioned groups, it is noteworthy that on averagethe mean deviation was larger in lesioned animals (contralat-eral lesion: 31.7 � 12.8°, ipsilateral lesion: 33.6 � 12.4°)compared with control animals (12.0 � 2.5°). In addition, twoHD cells in the lesioned group shifted to orientations that wereclearly unpredicted by landmark rotation (see Fig. 9B), and twoHD cells displayed directional instability. Rayleigh tests on theangular shift distributions confirm these observations, indicat-ing that the mean vector length is longer for the control group(r � 0.965) compared with the lesioned group (r � 0.765; P �0.05). However, similar to the rotation experiment, both dis-tributions were found to be significantly clustered (P � 0.001).
In sum, the results of the landmark rotation experimentindicate that unilateral lesions reduced the control exerted by asalient landmark. However, there was no impairment in land-mark control when the cue card was returned to its originalorientation in the testing cylinder.
Dark test. To determine whether SGN lesions disrupt thenormal processing of idiothetic cues in the HD cell circuit, wemonitored HD cell activity while rats foraged for food in theabsence of the cue card and in darkness. It was expected that ifSGN lesions impaired idiothetic cue processing, then the pre-ferred directions of HD cells would display a greater magni-tude of drift in darkness. We tested this prediction by con-structing HD � time plots for each HD cell recorded in thedark test (control: n � 19, contralateral: n � 15, ipsilateral:n � 8). Overall, HD cells in control animals demonstrated peakfiring rates that were clustered within a narrow range ofdirections throughout the entire 8-min dark test (e.g., Fig. 11A,left). In contrast, however, many of the HD cells recorded inSGN animals drifted throughout the session, and in some cases,oriented through all 360° directions (n � 7; e.g., Fig. 11A,right). To quantify these observations, we computed the angu-lar deviation for the peak firing rate points in the HD � timeplots. It was hypothesized that the drifting nature of HD cellsin the lesioned group would result in greater angular deviationvalues compared with the control group. Consistent with thisprediction, the mean angular deviation for HD cells in thelesioned groups (contralateral: 41.3 � 3.6°, ipsilateral: 56.2 �6.9°) was greater than those recorded in control animals(31.2 � 2.4°). This difference was confirmed by a significantANOVA [F(2,16.1) � 7.10, P � 0.006] as well as significantfollow-up comparisons between the contralateral lesion and
Table 2. Summary of histological results and incidence of HD and bursting cell activity in unilateral neurotoxic lesions of the SGN
Rat Lesion Site (Volume) Additional Damage Total Cells HD Cells Bursting Cells
jb23 Contralateral (60%) 6 2 2jb28 Contralateral (100%) 7 2 1jb40 Contralateral (80%) pDTN 7 1 0bc171 Contralateral (80%) pDTN 15 10 3bc172 Contralateral (90%) 8 1 4bc198 Contralateral (90%) 5 3 0bc199 Contralateral (80%) Rostral NPH 9 5 1bc200 Contralateral (100%) pDTN 16 10 1jb24 Ipsilateral (60%) Rostral NPH 6 1 0jb25 Ipsilateral (100%) 10 0 0bc167 Ipsilateral (100%) 9 9 1bc169 Ipsilateral (60%) 8 7 0
All recordings were conducted in the right ADN. Column listing additional damage lists only the regions that may have an influence on HD cell activity becauseof confirmed connectivity. NPH, nucleus prepositus hypoglossi.
Fig. 9. A: polar plots showing the angular shift in the preferred firing directionbetween the standard 1 and rotation sessions for control (left) and unilateralSGN-lesioned (right) rats. Each dot on the periphery represents the magnitudeof shift in the preferred firing direction for 1 HD cell. The arrows denote theobserved mean vector angle, and the broken line denotes the expected 90°vector if the angular shift values are perfectly controlled by the cue card. Meanvector length values (r) are indicated within the plot and are depicted by thelength of the arrow. The length of r represents the variability in the shift anglesfor the group, with values of 1.0 falling on the circle and indicating an absenceof variability. Asterisks mark r values that are significantly directional asindicated by the Rayleigh test (*P � 0.001). Note that the shifts for bothgroups are significantly clustered; however, for the lesioned group (contralat-eral, black dots; ipsilateral, gray dots) there was a tendency for underrotationas well as rotation in the wrong direction. B: polar plots showing the angularshift in the preferred firing direction between the standard 1 and standard 2sessions for control (left) and unilateral SGN-lesioned (right) rats. The 0° pointindicates the expected vector if the shift values are perfectly controlled by thecue card position. Although the shifts for both groups are significantlyclustered and concentrated around 0°, there was a tendency for cells in thelesion group to overrotate when the cue card was returned to its originalorientation.
2777IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

control groups [t(25.3) � �2.33, P � 0.038] and betweenipsilateral lesion and control groups [t(8.8) � �3.44, P �0.008]. However, there was no significant difference betweenthe contralateral and ipsilateral groups. The distribution ofangular deviation values for each group is shown in Fig. 11B.
We also investigated whether the magnitude of preferreddirection drift was related to variability in locomotor charac-teristics during the dark test. Specifically, theoretical and be-havioral studies suggest that the accuracy of idiothetic cueintegration (i.e., path integration) can degrade over time (Eti-enne and Jeffery 2004; Goodridge et al. 1998; McNaughton etal. 1991); thus it is possible that animals in the lesioned grouptaxed their path integration system to a greater extent thancontrol animals, possibly by making more head turns andtraveling a greater distance in the cylinder. We tested thishypothesis by measuring the cumulative distance traveled,number of head turns, and average angular head velocity. Meancomparisons between lesioned (contralateral and ipsilateralgroups pooled) and control animals failed to indicate a signif-icant difference for each measure [t(40) � 0.26, 1.07, and 0.96for cumulative distance traveled, angular head velocity, andtotal number of head turns, respectively; all P � 0.05]. Thusmovement characteristics alone cannot account for the direc-tional instability of HD cells in lesioned animals, supportingthe view that the SGN plays an important role in idiothetic cueprocessing.
Standard 3 (cue card return). To further test landmarkcontrol over HD cells in the lesion group, we turned the room
lights back on and returned the white cue card to its originalorientation in the testing cylinder after the dark test wascompleted. If the cue card exerted stimulus control over HDcells, their preferred directions should return to their originalstandard 2 orientations. Figure 11C plots the amount of angu-lar shift in the cells’ preferred directions between the standard2 and standard 3 sessions for control and lesioned animals. Ingeneral, the cue card exerted control over HD cells in bothgroups. Although the mean deviation from the expected shiftwas much greater for the ipsilateral lesion group (43.5 � 20.2°)compared with the contralateral lesion group (22.0 � 7.5°) andcontrol rats (19.7 � 5.9°), the ANOVA did not indicatesignificant group differences [F(2,15.9) � 0.617, P � 0.55]. Inaddition, the length of the mean vector was slightly shorter forthe lesioned group (r � 0.756) compared with control animals(r � 0.878), but this difference also did not reach significance(P � 0.05). Moreover, the Rayleigh test indicated that bothdistributions of angular shift values were significantly clusteredaround 0° (P � 0.001). Taken together, the results abovesuggest that landmark control was largely intact in SGNlesioned rats during the standard 3 sessions.
Dual-chamber apparatus. To further test whether SGN le-sions disrupt idiothetic information processing, we evaluatedthe stability of HD cell tuning while rats locomoted from thefamiliar cylinder to a novel, rectangular-shaped enclosure.Stable updating of the cell’s preferred direction should occurwhen a control rat enters the novel environment. A total of 10HD cells from control and 11 HD cells from lesioned animals
Fig. 10. A: representative firing rate � HD tuning curves for HD cells from unilateral SGN-lesioned rats during the landmark rotation and return series. Eachpanel depicts an HD cell recorded from different lesioned animals. The black line in each plot indicates the standard 1 session, the gray lines represent the rotationsession, and the red lines indicate the standard 2 sessions. The dashed vertical line in each plot shows the expected HD if cell tuning is perfectly controlled bythe cue card. Left: an HD cell in an ipsilaterally lesioned animal that underrotated in response to a CCW cue card rotation. Note that the preferred directionoverrotated when the cue card was positioned in its original orientation, suggesting that landmark control was largely intact in standard 2. Right: an HD cell froma contralaterally lesioned animal that rotated in the wrong direction after a CCW cue card rotation. Note, however, that the cell’s preferred direction returnedto its original orientation in standard 2. B, left: a representative “directionally unstable” cell in a lesion animal during the landmark rotation series. Note that thiscell lost its direction-specific firing during the landmark rotation session but showed a clear preferred direction during the standard 1 and standard 2 sessions.Right: an HD � time plot for the cell on left. Each dot depicts the time when the HD cell fired �50% of its maximum firing rate (black dot, standard 1; graydots, rotation; red dots, standard 2). Sampling resolution was 1 s. Note that the cell fired within a narrow range of directions in the standard 1 and standard 2sessions but drifted significantly during the landmark rotation session.
2778 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from
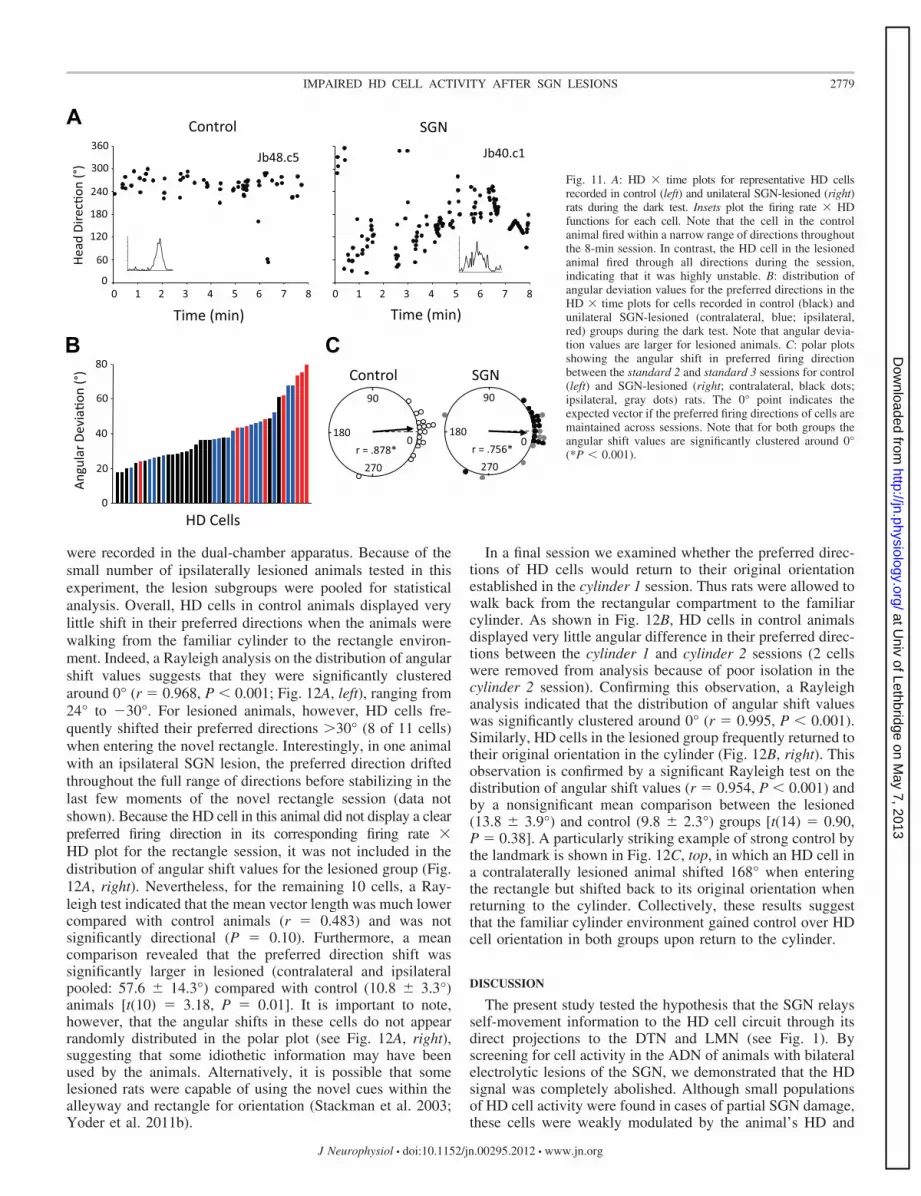
were recorded in the dual-chamber apparatus. Because of thesmall number of ipsilaterally lesioned animals tested in thisexperiment, the lesion subgroups were pooled for statisticalanalysis. Overall, HD cells in control animals displayed verylittle shift in their preferred directions when the animals werewalking from the familiar cylinder to the rectangle environ-ment. Indeed, a Rayleigh analysis on the distribution of angularshift values suggests that they were significantly clusteredaround 0° (r � 0.968, P � 0.001; Fig. 12A, left), ranging from24° to �30°. For lesioned animals, however, HD cells fre-quently shifted their preferred directions �30° (8 of 11 cells)when entering the novel rectangle. Interestingly, in one animalwith an ipsilateral SGN lesion, the preferred direction driftedthroughout the full range of directions before stabilizing in thelast few moments of the novel rectangle session (data notshown). Because the HD cell in this animal did not display a clearpreferred firing direction in its corresponding firing rate �HD plot for the rectangle session, it was not included in thedistribution of angular shift values for the lesioned group (Fig.12A, right). Nevertheless, for the remaining 10 cells, a Ray-leigh test indicated that the mean vector length was much lowercompared with control animals (r � 0.483) and was notsignificantly directional (P � 0.10). Furthermore, a meancomparison revealed that the preferred direction shift wassignificantly larger in lesioned (contralateral and ipsilateralpooled: 57.6 � 14.3°) compared with control (10.8 � 3.3°)animals [t(10) � 3.18, P � 0.01]. It is important to note,however, that the angular shifts in these cells do not appearrandomly distributed in the polar plot (see Fig. 12A, right),suggesting that some idiothetic information may have beenused by the animals. Alternatively, it is possible that somelesioned rats were capable of using the novel cues within thealleyway and rectangle for orientation (Stackman et al. 2003;Yoder et al. 2011b).
In a final session we examined whether the preferred direc-tions of HD cells would return to their original orientationestablished in the cylinder 1 session. Thus rats were allowed towalk back from the rectangular compartment to the familiarcylinder. As shown in Fig. 12B, HD cells in control animalsdisplayed very little angular difference in their preferred direc-tions between the cylinder 1 and cylinder 2 sessions (2 cellswere removed from analysis because of poor isolation in thecylinder 2 session). Confirming this observation, a Rayleighanalysis indicated that the distribution of angular shift valueswas significantly clustered around 0° (r � 0.995, P � 0.001).Similarly, HD cells in the lesioned group frequently returned totheir original orientation in the cylinder (Fig. 12B, right). Thisobservation is confirmed by a significant Rayleigh test on thedistribution of angular shift values (r � 0.954, P � 0.001) andby a nonsignificant mean comparison between the lesioned(13.8 � 3.9°) and control (9.8 � 2.3°) groups [t(14) � 0.90,P � 0.38]. A particularly striking example of strong control bythe landmark is shown in Fig. 12C, top, in which an HD cell ina contralaterally lesioned animal shifted 168° when enteringthe rectangle but shifted back to its original orientation whenreturning to the cylinder. Collectively, these results suggestthat the familiar cylinder environment gained control over HDcell orientation in both groups upon return to the cylinder.
DISCUSSION
The present study tested the hypothesis that the SGN relaysself-movement information to the HD cell circuit through itsdirect projections to the DTN and LMN (see Fig. 1). Byscreening for cell activity in the ADN of animals with bilateralelectrolytic lesions of the SGN, we demonstrated that the HDsignal was completely abolished. Although small populationsof HD cell activity were found in cases of partial SGN damage,these cells were weakly modulated by the animal’s HD and
Fig. 11. A: HD � time plots for representative HD cellsrecorded in control (left) and unilateral SGN-lesioned (right)rats during the dark test. Insets plot the firing rate � HDfunctions for each cell. Note that the cell in the controlanimal fired within a narrow range of directions throughoutthe 8-min session. In contrast, the HD cell in the lesionedanimal fired through all directions during the session,indicating that it was highly unstable. B: distribution ofangular deviation values for the preferred directions in theHD � time plots for cells recorded in control (black) andunilateral SGN-lesioned (contralateral, blue; ipsilateral,red) groups during the dark test. Note that angular devia-tion values are larger for lesioned animals. C: polar plotsshowing the angular shift in preferred firing directionbetween the standard 2 and standard 3 sessions for control(left) and SGN-lesioned (right; contralateral, black dots;ipsilateral, gray dots) rats. The 0° point indicates theexpected vector if the preferred firing directions of cells aremaintained across sessions. Note that for both groups theangular shift values are significantly clustered around 0°(*P � 0.001).
2779IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

were less stable relative to control animals. Consistent withprevious work (Muir et al. 2009; Yoder and Taube 2009), weidentified a population of ADN cells in bilaterally lesionedanimals that had nonperiodic “bursting” characteristics, whichresembled the activity of HD cells passing through their pre-ferred directions (Fig. 7). Because our SGN lesions in exper-iment 1 often included damage to the mlf, which also carriesascending vestibular information, in experiment 2 we furthertested the specific role of the SGN by unilaterally lesioning itwith the fiber-sparing neurotoxin NMDA and evaluating bothlandmark control and the use of idiothetic cues by ADN HDcells. We found that unilateral neurotoxic SGN lesions im-paired landmark control, as evidenced by the inconsistent
responses of HD cells to shift their preferred firing directions inalignment with the cue card after cue card rotations (Fig. 9A).However, this impairment was transient, as HD cells werecapable of reorienting in relation to the cue card in the standardsessions (Figs. 9B and 11C) and in the cylinder 2 sessions ofthe dual-chamber task (Fig. 12B). In contrast to the weakimpairments in landmark control, we observed large impair-ments in idiothetic processing. In particular, unilateral SGNlesions significantly reduced the stability of preferred direc-tions in the dark test (Fig. 11B). Furthermore, HD cells inunilaterally lesioned rats displayed large shifts in their pre-ferred firing directions when locomoting into a novel rectangleenvironment (Fig. 12A). Contralateral lesions of the SGNappeared to have a mildly greater impact on the directionalspecificity and landmark updating of HD tuning relative to theipsilateral group. Nevertheless, ipsilateral lesions producedcomparable impairments in idiothetic processing, suggestingthat bilateral output from each SGN nuclei is required forspatial updating.
What Is the Role of SGN Projections in HDSignal Processing?
The precise role of the SGN in processing the HD cell signalis not entirely understood; however, a number of lines ofevidence support the interpretation that the structure is cru-cially involved in conveying ascending vestibular signals to theHD cell circuit. First, the SGN receives a dense direct projec-tion from the medial vestibular nuclei (Biazoli et al. 2006), anarea that receives a high percentage of its input from thehorizontal semicircular canal, which is primarily sensitive toangular head acceleration in the horizontal (azimuthal) plane.The SGN also receive a more moderate projection from theNPH (Biazoli et al. 2006), which may indirectly convey ves-tibular information to the SGN (see Fig. 1). Second, the SGNsends large projections to the HD cell circuitry, specificallytargeting the DTN and LMN (Biazoli et al. 2006; Brown et al.2005), which are thought to be the site of HD signal generation(Bassett and Taube 2001; Sharp et al. 2001a; Taube 2007).Third, similar to lesions of the vestibular system (Muir et al.2009; Stackman and Taube 1997) or tegmento-mammillarycircuit (Bassett et al. 2007; Blair et al. 1999), bilateral damageof the SGN completely abolished HD tuning in the ADN.Finally, we identified bursting activity in the ADN after bilat-eral and unilateral SGN damage, which is consistent withprevious studies showing similar burst characteristics in theADN of animals with semicircular canal occlusion (Muir et al.2009) or transgenic otolith disruption (Yoder and Taube 2009).Although the points mentioned above suggest that SGN’srole in generating the HD signal relates to its interactionswith the vestibular system, it is possible that its role may bemore related to nonvestibular functions, such as maintaininga frame of reference or suppressing an eye-related positionsignal from a gaze signal to yield head position information.
It is also notable that partial lesions of the SGN, especiallycontralateral to the electrode implant, significantly increasedthe amount of time that ADN HD cells anticipate futureorientations (i.e., the anticipatory time interval of HD cells).Although it is not clear whether the anticipatory time intervalserves a particular function or if it is simply an epiphenomenonthat arises from vestibular system projections (Bassett et al.
Fig. 12. A: polar plots showing the angular shift in the preferred directionsbetween the cylinder 1 and rectangle sessions of the dual-chamber task forcontrol (left) and unilateral SGN-lesioned (right) rats. The broken line at 0°indicates the expected vector if the preferred directions of cells are maintainedacross sessions. The plots reveal that the angular shift values are significantlyclustered around 0° for control animals (*P � 0.001); however, angular shiftswere larger and not significantly clustered for lesioned animals. B: polar plotsshowing the angular shift in preferred firing direction between the cylinder 1and cylinder 2 sessions for control (left) and unilateral SGN-lesioned (right)rats. Again, the broken line at 0° points to the expected vector if the angularshift values maintain their orientation between sessions. Note that the angularshifts are significantly clustered around 0° for both groups (*P � 0.001).C: representative firing rate � HD tuning curves for HD cells from unilateralSGN-lesioned rats recorded in the dual-chamber apparatus. Black lines in eachplot indicate the cylinder 1 session, gray lines represent the rectangle session,and red lines indicate the cylinder 3 sessions. Note that each cell shifted itspreferred firing direction when the rat was entering the novel rectangleenclosure, but each cell’s preferred direction returned to its original orientationafter the rat locomoted to the familiar cylinder.
2780 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

2005; van der Meer et al. 2007), the general increase inanticipation reported here is similar to that shown in other workin which ascending self-motion information has been disrupted(Clark et al. 2009; Shinder and Taube 2011).
It is important to point out, however, that the SGN sendsprojections to the cerebellum, in particular the floccular nod-ular lobe (Blanks et al. 1983), which is often linked to gazecontrol (McCrea 1988). Thus it is possible that impairments ingaze control, rather than impairments in vestibular processing,may have contributed to the present results. We believe thispossibility is unlikely for several reasons. First, although ratshave a prominent vestibular-ocular reflex, they do not makefrequent saccades or smooth pursuit eye movements. Further-more, eye movements do not play as central a role in theirbehavior as in cats or primates. Second, one study conducted innonhuman primates (Robertson et al. 1999) found that the HDsignal was independent of eye position, suggesting that eyemovements were unlikely to be important in deriving the HDsignal. Third, it is believed that the HD signal represents theanimal’s perceived directional heading in space—not its gazeposition. Thus interfering with the animal’s ability to make eyemovements should not impact its perceived directional head-ing. Finally, in unpublished experiments we have lesionedparts of the cerebellar floccular nodular lobe—areas that arethought to play an important role in eye movements—and theselesions did not disrupt the HD signal in the sense that itabolished it (M. Glickstein and J. S. Taube, unpublishedobservations). Because several previous studies have shownthat interfering with the vestibular system disrupts the HDsignal (e.g., Muir et al. 2009; Stackman and Taube 1997), themost parsimonious interpretation of our findings is that the lossof vestibular information via the SGN to the HD circuit iscritical for enabling the generation of the HD signal. Finally, itis important to note that the SGN also receives moderateprojections from the paragigantocellular reticular nucleus (Bia-zoli et al. 2006). Very little is known about this structure, andfuture work should address this gap in our knowledge.
It is also important to address the possibility that damage tothe adjacent mlf fiber pathway may have contributed to ourresults. In experiment 1, electrolytic lesions were used toremove the SGN bilaterally, but this procedure also damagedadjacent fiber pathways such as the mlf. The mlf conveysascending vestibular information from not only the vestibularnuclei but also several midbrain structures involved in process-ing vestibular information; thus it is possible that damaging themlf may have contributed to the results of experiment 1. Asnoted in RESULTS, Frohardt et al. (2006) tested the possibilitythat the mlf plays a role in processing idiothetic cues byselectively damaging the mlf in two animals and testing themin a task requiring the accurate use of self-movement cues toreach a goal location. They reported that transection of the mlfdid not impair navigation in the task, suggesting that damage tothe mlf alone does not disrupt path integration processesdownstream (i.e., in efferent areas). Regardless, in experiment2, we addressed the possibility that the mlf contributed to ourresults by damaging the SGN with a neurotoxin (NMDA) thatdoes not damage axons passing near the injection site (Jarrard1989). As summarized above, neurotoxic lesions of the SGNdisrupted the use of idiothetic cues to maintain a stable HDsignal in the dark and when entering a novel environment,
indicating that the SGN plays an important role in angular pathintegration.
Given the conclusion that the SGN conveys idiothetic infor-mation to the HD cell circuit for angular path integration, it isunclear why mild impairments were observed during the land-mark rotation series, since it would seem that the idiotheticsystem would have little to do with the landmark system at thelevel of the brain stem. One possible explanation is that theSGN may influence HD signal processing through the tecto-pulvinar pathway because it sends moderate projections to thesuperior colliculus (i.e., SGN ¡ superior colliculus ¡ lateraldorsal thalamus ¡ postsubiculum) (Klop et al. 2005; Yoder etal. 2011a). However, previous work by Golob et al. (1998)established that lesions of this pathway at the level of thelateral dorsal thalamic nuclei do not impair landmark controlover HD tuning in the postsubiculum. An alternative hypoth-esis is that environmental landmarks are associated with anidiothetic-based reference frame (Alyan and Jander 1994; Mc-Naughton et al. 1996); thus lesions disrupting idiothetic pro-cessing would also disrupt landmark control. This hypothesis issupported by other studies showing that disruption of accuratevestibular processing impairs landmark control over HD cells(e.g., Knierim et al. 1995; Stackman and Taube 1997) and canlead to impairments in landmark-based navigation (Clark andTaube 2009; Dudchenko et al. 1997b).
In experiment 2 we investigated the relative role of topo-graphically organized SGN projections to the tegmento-mam-millary pathway such that each nucleus sends axons to thecontralateral DTN and ipsilateral LMN (Biazoli et al. 2006).One interesting feature of this connectivity is that SGN input tothe DTN is much stronger than input to LMN. As a conse-quence, it could be argued that projections to the DTN aremore significant with respect to HD signal processing. Theresults of experiment 2, however, suggest that both inputs arenecessary for generating an accurate signal based on spatialupdating via idiothetic cues. This result is particularly apparentin the dark test, in which HD cells were recorded in animalswith ipsilateral lesions of the SGN (thereby disconnectinginput to LMN) or contralateral lesions (thereby disconnectinginput to DTN). In general, the preferred directions of HD cellsrecorded in each group drifted a similar magnitude across the8-min test session (Fig. 11B). Similarly, in the rectanglesession of the dual-chamber task, cells in both groups demon-strated significant shifts in their preferred firing directions (Fig.12A). In sum, these findings suggest that inputs to the DTN andLMN from both SGN nuclei together are required for HD cellstability, and suggest that the influence of SGN output iswidespread across the HD cell generative circuit.
It is presently unknown what physiological information theSGN may convey to the HD cell circuit, since no study hasrecorded activity from this structure. However, it is likely thatthe SGN plays a role in processing angular head velocityinformation (Sharp et al. 2001a; Taube 2007). As noted above,the SGN receives input from the medial vestibular nuclei,which contain neurons modulated by angular head velocity(Angelaki and Cullen 2008). In addition to containing neuronsmodulated by HD, the DTN and LMN have large populationsof neurons sensitive to angular head velocity (Bassett andTaube 2001; Blair et al. 1998; Sharp et al. 2001b; Stackmanand Taube 1998). The DTN also sends a strong projection backto the contralateral SGN (Biazoli et al. 2006); thus it is possible
2781IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

that SGN neurons are also responsive to current HD because asmall population of HD cells have been reported in the DTN(Sharp et al. 2001b).
Other projections likely provide vestibular information to theHD cell system. For instance, the NPH receives strong inputsfrom the vestibular system and projects directly to the DTN (Fig.1) (Brown et al. 2005). SGN and NPH projections to the HD cellcircuit differ slightly in that the NPH does not project directly tothe LMN and, instead of projecting contralaterally, it sends mas-sive input to the ipsilateral DTN (Liu et al. 1984). Like the SGN,very little is known about NPH neural activity in freely movingrats. One study in immobilized rats found neurons that wereresponsive to optokinetic as well as sinusoidal vestibular stimu-lation in the horizontal plane (Lannou et al. 1984). In cats andnonhuman primates, NPH neurons are strongly modulated by eyemovements (Lopez-Barneo et al. 1982; Delgado-García et al.1989; McFarland and Fuchs 1992), and it has been argued that thisregion is the site of the neural integrator for the vestibulooccularreflex (Robinson 1989). Interestingly, the activity of some NPHneurons has been correlated with the animal’s angular headmovements (McFarland and Fuchs 1992), leaving open the pos-sibility that the NPH may provide an alternative vestibular path-way for HD cell computations.
Implications for Computational Models of HDCell Generation
McNaughton et al. (1991) provided the first computationalmodel that captured the notion that the HD cell signal could bederived by tracking an animal’s angular head velocity over time.Subsequent models (e.g., Redish et al. 1996; Skaggs et al. 1995;Zhang 1996) have expanded on this basic principle, using contin-uous attractor networks (reviewed in Taube and Bassett 2003). Inthese models, HD cells are conceptually arranged in a ringcorresponding to their preferred firing directions. Cells with pre-ferred directions that have overlapping directional firing rangesshare excitatory connections, and cells with greatly differentpreferred directions inhibit each other. This neural architectureforms a sustained “hill” of excitation centered on the animal’scurrent HD. Information about the animal’s angular head velocityis derived from vestibular inputs that are typically used to movethe hill of activity around to different directions depending onchanges to the animal’s HD. Consequently, the removal of angu-lar head velocity information results in an activity hill that nolonger moves along the ring in register with an animal’s HD.Instead, the activity hill moves around the ring at different ratesdepending on the remaining inputs that convey information aboutcurrent heading (e.g., visual and motor). Consistent with this viewis our observation that some ADN cells in SGN-lesioned rats firedin characteristic short, nonrhythmic “bursts,” which generally hadthe appearance of a cell periodically passing through its preferredfiring direction. These “bursting” cells could be interpreted as anactivity hill moving along the attractor ring at varying rates (seeClark and Taube 2012 for review). Bursting cell correlates havealso been identified in the ADN after disruption of the vestibularsystem (Muir et al. 2009; Stackman and Taube 1997; Yoder andTaube 2009) but not after lesions to the DTN or LMN (Bassett etal. 2007), suggesting that lesions of the vestibular system or SGNdo not disconnect the internal dynamics of the ring attractornetwork, whereas lesions of the DTN or LMN apparently disruptthe network at a more fundamental level. Interestingly, the ves-
tibular lesion studies have also reported that the temporal order ofsimultaneously recorded bursting cells is generally preserved(Muir et al. 2009), again suggesting that inputs from the vestibularsystem are not necessary for generating the organized structure ofthe HD cell (attractor) network. Given these findings, the HDattractor network most likely resides within the tegmental-mam-millary circuit (Bassett et al. 2007; Blair et al. 1999).
Summary
The present study evaluated the hypothesis that the SGNconveys vestibular information critical for the generation of theHD cell signal. Consistent with this proposal, we found thatcomplete lesions of the SGN abolished HD cell activity in theADN and unilateral lesions disrupted the capacity to updatedirectional orientation based on idiothetic input. This latter findingin particular suggests that unilateral SGN input to the HD circuitcan sustain directional activity but idiothetic processing of thedirectional signal requires input from both nuclei. Future studiesshould be directed at understanding the correlates of neural activ-ity within the SGN as well as understanding the contributions ofthe closely associated NPH. Such work will broaden our under-standing of the signal transformations involved in generating theHD cell signal and provide greater insight into the neural andcomputational mechanisms underlying spatial orientation.
ACKNOWLEDGMENTS
J. E. Brown received National Institutes of Health (NIH) postdoctoratefellowship DC-008773. B. J. Clark received a postgraduate fellowship from theNational Sciences and Engineering Research Council of Canada.
Present address of B. J. Clark: Canadian Centre for Behavioural Neurosci-ence, University of Lethbridge, Lethbridge, AB, Canada T1K 3M4.
GRANTS
This work was supported through NIH Grants NS-053907 and DC-009318to J. S. Taube.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: B.J.C., J.E.B., and J.S.T. conception and design ofresearch; B.J.C. and J.E.B. performed experiments; B.J.C. and J.S.T. analyzeddata; B.J.C. and J.S.T. interpreted results of experiments; B.J.C. preparedfigures; B.J.C. drafted manuscript; B.J.C. and J.S.T. edited and revised man-uscript; B.J.C., J.E.B., and J.S.T. approved final version of manuscript.
REFERENCES
Alyan S, Jander R. Short-range homing in the house mouse, Mus musculus:stages in the learning of directions. Anim Behav 48: 285–298, 1994.
Angelaki DE, Cullen KE. Vestibular system: the many facets of a multimodalsense. Annu Rev Neurosci 31: 125–150, 2008.
Bassett JP, Taube JS. Neural correlates for angular head velocity in the ratdorsal tegmental nucleus. J Neurosci 21: 5740–5751, 2001.
Bassett JP, Tullman ML, Taube JS. Lesions of the tegmentomammillarycircuit in the head direction system disrupt the head direction signal in theanterior thalamus. J Neurosci 27: 7564–7577, 2007.
Bassett JP, Zugaro MB, Muir GM, Golob EJ, Muller RU, Taube JS.Passive movements of the head do not abolish anticipatory firing propertiesof head direction cells. J Neurophysiol 93: 1304–1316, 2005.
Batschelet E. Circular Statistics in Biology. New York: Academic, 1981.Biazoli CE Jr, Goto M, Campos AM, Canteras NS. The supragenual
nucleus: a putative relay station for ascending vestibular signs to headdirection cells. Brain Res 1094: 138–148, 2006.
2782 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

Blair HT, Sharp PE. Anticipatory head direction signals in anterior thalamus:evidence for a thalamocortical circuit that integrates angular head motion tocompute head direction. J Neurosci 15: 6260–6270, 1995.
Blair HT, Cho J, Sharp PE. Role of the lateral mammillary nucleus in the rathead direction circuit: a combined single-unit recording and lesion study.Neuron 21: 1387–1397, 1998.
Blair HT, Cho J, Sharp PE. The anterior thalamic head-direction signal isabolished by bilateral but not unilateral lesions of the lateral mammillarynucleus. J Neurosci 19: 6673–6683, 1999.
Blanks RH, Precht W, Torigoe Y. Afferent projections to the cerebellarflocculus in the pigmented rat demonstrated by retrograde transport ofhorseradish peroxidase. Exp Brain Res 52: 293–306, 1983.
Boccara CN, Sargolini F, Thoresen VH, Solstad T, Witter MP, Moser EI,Moser MB. Grid cells in pre- and parasubiculum. Nat Neurosci 13:987–994, 2010.
Brown JE, Card JP, Yates BJ. Polysynaptic pathways from the vestibularnuclei to the lateral mammillary nucleus of the rat: substrates for vestibularinput to head direction cells. Exp Brain Res 161: 47–61, 2005.
Chen LL, Lin LH, Barnes CA, McNaughton BL. Head-direction cells in therat posterior cortex. II. Contributions of visual and ideothetic information tothe directional firing. Exp Brain Res 101: 24–34, 1994.
Cho J, Sharp PE. Head direction, place, and movement correlates for cells inthe rat retrosplenial cortex. Behav Neurosci 115: 3–25, 2001.
Clark BJ, Taube JS. Deficits in landmark navigation and path integrationafter lesions of the interpeduncular nucleus. Behav Neurosci 123: 490–503,2009.
Clark BJ, Taube JS. Intact landmark control and angular path integration byhead direction cells in the anterodorsal thalamus after lesions of the medialentorhinal cortex. Hippocampus 21: 767–782, 2011.
Clark BJ, Taube JS. Vestibular and attractor network basis of the headdirection cell signal in subcortical circuits. Front Neural Circuits 6: 7, 2012.
Clark BJ, Sarma A, Taube JS. Head direction cell instability in the anteriordorsal thalamus after lesions of the interpeduncular nucleus. J Neurosci 29:493–507, 2009.
Clark BJ, Bassett JP, Wang S, Taube JS. Impaired head direction cellrepresentation in the anterodorsal thalamus after lesions of the retrosplenialcortex. J Neurosci 30: 5289–5302, 2010.
Delgado-García JM, Vidal PP, Gómez C, Berthoz A. A neurophysiologicalstudy of prepositus hypoglossi neurons projecting to oculomotor and pre-oculomotor nuclei in the alert cat. Neurosci 29: 291–307, 1989.
Dudchenko PA, Goodridge JP, Taube JS. Effects of repeated disorientationon the acquisition of spatial tasks in rats: dissociation between the appetitiveradial arm maze and aversive water maze. J Exp Psych Anim Behav Process23: 194–210, 1997a.
Dudchenko PA, Goodridge JP, Taube JS. The effects of disorientation onvisual landmark control of head direction cell orientation. Exp Brain Res115: 375–380, 1997b.
Etienne AS, Jeffery KJ. Path integration in mammals. Hippocampus 14:180–192, 2004.
Frohardt RJ, Bassett JP, Taube JS. Path integration and lesions within thehead direction cell circuit: comparison between the roles of the anterodorsalthalamus and dorsal tegmental nucleus. Behav Neurosci 120: 135–149,2006.
Golob EJ, Wolk DA, Taube JS. Recordings of postsubicular head directioncells following lesions of the lateral dorsal thalamic nucleus. Brain Res 780:9–19, 1998.
Goodridge JP, Taube JS. Interaction between the postsubiculum and anteriorthalamus in the generation of head direction cell activity. J Neurosci 17:9315–9330, 1997.
Goodridge JP, Dudchenko PA, Worboys KA, Golob EJ, Taube JS. Cuecontrol and head direction cells. Behav Neurosci 112: 749–761, 1998.
Hayakawa T, Zyo K. Afferent connections of Gudden’s tegmental nuclei inthe rabbit. J Comp Neurol 235: 169–181, 1985.
Jarrard LE. On the use of ibotenic acid to lesion selectively differentcomponents of the hippocampal formation. J Neurosci Methods 29: 251–259, 1989.
Klop EM, Mouton LJ, Ehling T, Holstege G. Two parts of the nucleusprepositus hypoglossi project to two different subdivisions of the dorsolat-eral periaqueductal gray in cat. J Comp Neurol 3: 303–322, 2005.
Knierim JJ, Kudrimoti HS, McNaughton BL. Place cells, head directioncells, and the learning of landmark stability. J Neurosci 15: 1648–1659,1995.
Kubie JL. A driveable bundle of microwires for collecting single-unit datafrom freely-moving rats. Physiol Behav 32: 115–118, 1984.
Lannou J, Cazin L, Precht W, Le Taillanter M. Responses of prepositushypoglossi neurons to optokinetic and vestibular stimulations in the rat.Brain Res 301: 39–45, 1984.
Liu R, Chang L, Wickern G. The dorsal tegmental nucleus: an axoplasmictransport study. Brain Res 310: 123–132, 1984.
Lopez-Barneo J, Darlot C, Berthoz A, Baker R. Neural activity in preposi-tus nucleus correlated with eye movement in the alert cat. J Neurophysiol47: 329–352, 1982.
McCrea RA. The nucleus prepositus. Rev Oculomotor Res 2: 1–22, 1988.McCrea RA. Nucleus prepositus. Prog Brain Res 151: 205–230, 2006.McFarland JL, Fuchs AF. Discharge patterns in nucleus prepositus hypo-
glossi and adjacent medial vestibular nucleus during horizontal eye move-ment in behaving macaques. J Neurophysiol 68: 319–332, 1992.
McNaughton BL, Chen LL, Markus EJ. “Dead reckoning,” landmarklearning, and the sense of direction: a neurophysiological and computationalhypothesis. J Cogn Neurosci 3: 190–202, 1991.
McNaughton BL, Barnes CA, Gerrard JL, Gothard K, Jung MW, Kn-ierim JJ, Kudrimoti H, Qin Y, Skaggs WE, Suster M, Weaver KL.Deciphering the hippocampal polyglot: the hippocampus as a path integra-tion system. J Exp Biol 199: 173–185, 1996.
Moser EI, Kropff E, Moser MB. Place cells, grid cells, and the brain’s spatialrepresentation system. Annu Rev Neurosci 31: 69–89, 2008.
Muir GM, Brown JE, Carey JP, Hirvonen TP, Della Santina CC, MinorLB, Taube JS. Disruption of the head direction cell signal after occlusionof the semicircular canals in the freely moving chinchilla. J Neurosci 29:14521–14533, 2009.
Neitz J, Jacobs GH. Reexamination of spectral mechanisms in the rat (Rattusnorvegicus). J Comp Psychol 100: 21–29, 1986.
Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates (4th ed.).San Diego, CA: Academic, 1998.
Redish AD, Elga AN, Touretzky DS. A coupled attractor model of the rodenthead direction system. Netw Comput Neural Syst 7: 671–685, 1996.
Robertson RG, Rolls ET, Georges-François P, Panzeri S. Head directioncells in the primate pre-subiculum. Hippocampus 9: 206–219, 1999.
Robinson DA. Integrating with neurons. Annu Rev Neurosci 12: 33–45, 1989.Sargolini F, Fyhn M, Hafting T, McNaughton BL, Witter MP, Moser MB,
Moser EI. Conjunctive representation of position, direction, and velocity inentorhinal cortex. Science 312: 758–762, 2006.
Sharp PE, Koester K. Lesions of the mammillary body region severelydisrupt the cortical head direction, but not place cell signal. Hippocampus18: 766–784, 2008.
Sharp PE, Blair HT, Cho J. The anatomical and computational basis of therat head-direction cell signal. Trends Neurosci 24: 289–294, 2001a.
Sharp PE, Tinkelman A, Cho J. Angular velocity and head direction signalsrecorded from the dorsal tegmental nucleus of Gudden in the rat: implica-tions for path integration in the head direction cell circuit. Behav Neurosci115: 571–588, 2001b.
Shinder ME, Taube JS. Active and passive movement are encoded equally byhead direction cells in the anterodorsal thalamus. J Neurophysiol 106:788–800, 2011.
Skaggs WE, McNaughton BL, Gothard KM, Markus EJ. An information-theoretic approach to deciphering the hippocampal code. In: Advances inNeural Information Processing Systems, Vol. 5, edited by Hanson SJ,Cowan JD, Giles CL. San Mateo, CA: Morgan Kaufmann, 1993, p. 1030–1037.
Skaggs WE, Knierim JJ, Kudrimoti HS, McNaughton BL. A model of theneural basis of the rat’s sense of direction. In: Advances in Neural Infor-mation Processing Systems, Vol. 7, edited by Tesauro G, Touretzky DS,Leen TK. Cambridge, MA: MIT Press, 1995, p. 173–180.
Stackman RW, Taube JS. Firing properties of head direction cells in ratanterior thalamic neurons: dependence upon vestibular input. J Neurosci 17:4349–4358, 1997.
Stackman RW, Taube JS. Firing properties of rat lateral mammillary singleunits: head direction, head pitch, and angular head velocity. J Neurosci 18:9020–9037, 1998.
Stackman RW, Clark AS, Taube JS. Hippocampal spatial representationsrequire vestibular input. Hippocampus 12: 291–303, 2002.
Stackman RW, Golob EJ, Bassett JP, Taube JS. Passive transport disruptsdirectional path integration by rat head direction cells. J Neurophysiol 90:2862–2874, 2003.
Swanson LW. Brain Maps: Structure of the Rat Brain (2nd ed.). Amsterdam:Elsevier Science, 1998.
Taube JS. Head direction cells recorded in the anterior thalamic nuclei offreely moving rats. J Neurosci 15: 70–86, 1995.
2783IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from

Taube JS. The head direction signal: origins and sensory-motor integration.Annu Rev Neurosci 30: 181–207, 2007.
Taube JS, Bassett JP. Persistent neural activity in head direction cells. CerebCortex 13: 1162–1172, 2003.
Taube JS, Burton HL. Head direction cell activity monitored in a novelenvironment and during a cue conflict situation. J Neurophysiol 74: 1953–1971, 1995.
Taube JS, Muller RU. Comparisons of head direction cell activity in thepostsubiculum and anterior thalamus of freely moving rats. Hippocampus 8:87–108, 1998.
Taube JS, Muller RU, Ranck JB Jr. Head-direction cells recorded from thepostsubiculum in freely moving rats. I. Description and quantitative analy-sis. J Neurosci 10: 420–435, 1990a.
Taube JS, Muller RU, Ranck JB Jr. Head-direction cells recorded from thepostsubiculum in freely moving rats. II. Effects of environmental manipu-lations. J Neurosci 10: 436–447, 1990b.
van der Meer MA, Knierim JJ, Yoganarasimha D, Wood ER, van RossumMC. Anticipation in the rodent head direction system can be explained byan interaction of head movements and vestibular firing properties. J Neu-rophysiol 98: 1883–1897, 2007.
Yoder R, Taube JS. Head direction cell activity in mice: robust directionalsignal depends on intact otolith organs. J Neurosci 29: 1061–1076,2009.
Yoder RM, Clark BJ, Taube JS. Origins of landmark encoding in the brain.Trends Neurosci 34: 561–571, 2011a.
Yoder RM, Clark BJ, Brown JE, Lamia MV, Valerio S, Shinder ME,Taube JS. Both visual and idiothetic cues contribute to head direction cellstability during navigation along complex routes. J Neurophysiol 105:2989–3001, 2011b.
Zhang K. Representation of spatial orientation by the intrinsic dynamics ofthe head-direction cell ensemble: a theory. J Neurosci 16: 2112–2126,1996.
2784 IMPAIRED HD CELL ACTIVITY AFTER SGN LESIONS
J Neurophysiol • doi:10.1152/jn.00295.2012 • www.jn.org
at Univ of Lethbridge on M
ay 7, 2013http://jn.physiology.org/
Dow
nloaded from