characterization of glyceraldehyde-3-phosphate dehydrogenase as a novel transferrin receptor
TRANSCRIPT

Ct
SMa
b
a
ARRAA
KTGdITM
1
citlapwrftecT(
f
1d
The International Journal of Biochemistry & Cell Biology 44 (2012) 189– 199
Contents lists available at SciVerse ScienceDirect
The International Journal of Biochemistry& Cell Biology
journa l h o me page: www.elsev ier .com/ locate /b ioce l
haracterization of glyceraldehyde-3-phosphate dehydrogenase as a novelransferrin receptor
antosh Kumara, Navdeep Sheokanda, Mayur Anant Mhadeshwarb, Chaaya Iyengar Rajeb,∗,anoj Rajea,∗∗
Institute of Microbial Technology, CSIR, Sector 39A, Chandigarh 160036, IndiaNational Institute of Pharmaceutical Education and Research, Phase X, SAS Nagar, Punjab 160062, India
r t i c l e i n f o
rticle history:eceived 23 June 2011eceived in revised form 1 October 2011ccepted 5 October 2011vailable online 2 November 2011
eywords:ransferrin receptorlyceraldehyde-3-phosphate
a b s t r a c t
A majority of cells obtain of transferrin (Tf) bound iron via transferrin receptor 1 (TfR1) or by transferrinreceptor 2 (TfR2) in hepatocytes. Our study establishes that cells are capable of acquiring transferrin ironby an alternate pathway via GAPDH.
These findings demonstrate that upon iron depletion, GAPDH functions as a preferred receptor fortransferrin rather than TfR1 in some but not all cell types. We utilized CHO-TRVb cells that do not expressTfR1 or TfR2 as a model system. A knockdown of GAPDH in these cells resulted in a decrease of not onlytransferrin binding but also associated iron uptake. The current study also demonstrates that, unlike TfR1and TfR2 which are localized to a specific membrane fraction, GAPDH is located in both the detergent
ehydrogenaseronransferrinultifunctional protein
soluble and lipid raft fractions of the cell membrane. Further, transferrin uptake by GAPDH occurs by morethan one mechanism namely clathrin mediated endocytosis, lipid raft endocytosis and macropinocytosis.By determining the kinetics of this pathway it appears that GAPDH–Tf uptake is a low affinity, highcapacity, recycling pathway wherein transferrin is catabolised. Our findings provide an explanation forthe detailed role of GAPDH mediated transferrin uptake as an alternate route by which cells acquire iron.
. Introduction
Iron is an essential nutrient for all organisms as a criticalomponent of hemoproteins, iron–sulphur proteins and proteinsnvolved in vital housekeeping functions including the electronransport chain. Mammalian iron metabolism is a tightly regu-ated process consisting of three basic processes of uptake, storagend recycling. Almost all extracellular iron circulating in thelasma is bound to transferrin, an abundant iron transport proteinith high affinity for iron. Cellular iron uptake occurs chiefly via
eceptor-mediated uptake involving two well documented trans-errin receptors namely TfR1 and TfR2. Both these receptors areransmembrane, glycoproteins. TfR1 is a high affinity ubiquitouslyxpressed receptor while expression of TfR2 is restricted to certain
ell types and is unaffected by intracellular iron concentrations.fR2 binds to transferrin with a 25–30-fold lower affinity than TfR1Kawabata et al., 2000; West et al., 2000).Abbreviations: GAPDH, glyceraldehyde-3-phosphate dehydrogenase; Tf, trans-errin; TfR, transferrin receptor; DFO, desferrioxamine; CTX, cholera toxin.∗ Corresponding author.
∗∗ Corresponding author. Fax: +91 172 2690585/132.E-mail address: [email protected] (M. Raje).
357-2725/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.biocel.2011.10.016
© 2011 Elsevier Ltd. All rights reserved.
Previous reports have indicated that transferrin uptake couldbe mediated by receptors other than TfR1 and 2. Studies usingmelanoma, hepatoma and CHO-TRVb cells have suggested thatalternate routes for uptake of Tf bound iron exist (Chan et al., 1992;Levy et al., 1999; McGraw et al., 1987; Richardson and Baker, 1990,1994; Trinder and Morgan, 1997; Trinder et al., 1996), though thesepathways have not been elucidated. Previously, we reported thatthe multifunctional enzyme glyceraldehyde-3-phosphate dehy-drogenase (GAPDH), functions as a low affinity TfR in mammalianmacrophages (Raje et al., 2007). The current study provides evi-dence that GAPDH is this additional mechanism of transferrin ironuptake in some cell types particularly during iron depletion.
Using a cell line that lacks both TfRs as well as by the knockdownof GAPDH itself we demonstrate significant differences in terms ofits localization, internalization, recycling and kinetics of transferrinuptake. The current findings provide an insight into a completelyunexplored aspect of cellular iron metabolism and homeostasis.
2. Materials and methods
2.1. Media and cell lines
Multiple cell lines were obtained from National Center for CellScience (NCCS), Pune, India. These included L1210, BV2, J774,

1 f Bioch
Ra(M1lef1iCpa
2
vwc21C
2
sfiaGipG
2
(pi
niwNCaTc
Ticst
2
Fpf
90 S. Kumar et al. / The International Journal o
AW and N2A (mouse origin, CHO (Chinese hamster ovary), K562nd Hela (human). The CHO-TRVb, (derived from CHO) and NSmouse fibroblast) cell lines were kindly provided by Prof. T.
cGraw (McGraw et al., 1987) and Prof. A. Takashima (Xu et al.,995) respectively. All cells were cultured in RPMI 1640 and ana-
yzed by FACS for cell surface GAPDH expression. Iron depletionxperiments were done using complete medium with 100 �M Des-errioxamine (DFO) (Sigma) as described (Richardson and Baker,994). Controls were set up in parallel with normal media. Cellular
ron depletion was confirmed by atomic absorption spectroscopy.ell viability was confirmed by trypan blue, propidium iodide, sul-harhodamine and neutral red (Sigma) staining as well as MTTssay.
.2. FACS analysis
All FACS experiments were done essentially as described pre-iously (Raje et al., 2007). Briefly, 2 × 105 cells were stained eitherith 1 �g of monoclonal �GAPDH antibody (Advanced Immuno-
hemicals), followed by Sheep �mouse-FITC (Fab)2 (Sigma) or with �g of Tf-FITC (Sigma) or �CD71(TfR1)-FITC/PE (BD). Analysis of04 cells was done for each sample, using a FACS Calibur Flowytometer (BD).
.3. GAPDH silencing
CHO-TRVb cells were transfected with either control or GAPDHiRNA using Silencer transfection kit (Ambion). Silencing was con-rmed by KDalert GAPDH Assay Kit (Ambion), by western blottingnd also by measuring the decrease in intracellular, cell surfaceAPDH and transferrin binding by Tf-Alexa633 (Invitrogen). For
ron depletion experiments, transfected cells were shifted to com-lete or DFO containing media for an additional 24 h. Cell surfaceAPDH or the binding of Tf-Alexa633 was analyzed by FACS.
.4. 55Fe–transferrin uptake
55Fe–NTA (Sarkar et al., 2003) and 25 mg/ml human Apo-TfSigma) dissolved in 20 mM NaHCO3, 40 mM Tris, pH 7.4 were pre-ared separately and equilibrated in a 3:2 v/v ratio for 30 min. Free
ron was removed on a desalting column.CHO-TRVb cells (4 × 104 cells per well) were transfected with
egative control siRNA or GAPDH siRNA as per manufacturer’snstructions. Cells were shifted to normal media after 24 h. The plate
as centrifuged after 72 h and washed with 20 mM HEPES, 150 mMaCl, 5 mM KCl and 1 mM each of CaCl2, MgCl2 (neutral buffer).ells were then incubated in SFM containing 10 �M 55Fe–Tf for 6 ht 37 ◦C and lysed in 100 �l of 0.05% Triton X–100, 2 mg/ml BSA.he total cell associated radioactivity was measured by scintillationounting.
For measuring iron incorporation, into cellular proteins, CHO-RVb/J774 cells were processesed for 55Fe–Tf incorporation forndicated time points as above. Proteins in cell lysate were pre-ipitated using 15% TCA. Pellet (protein incorporated 55Fe) orupernatant (free 55Fe) was measured. Total cell associated radioac-ivity was calculated as the sum of fractions.
.5. Modulation of GAPDH/TfR1 expression
Iron starved J774 cells were stained for surface GAPDH or TfR1.or intracellular GAPDH/TfR1 staining, cells were fixed with 1%araformaldehyde, and permeablized using 0.1% saponin at 37 ◦Cor 15 min before staining and analysis by FACS.
emistry & Cell Biology 44 (2012) 189– 199
2.6. Characterization of cell surface and cytosolic GAPDH
Membrane and cytosolic protein fractions (Vinay et al., 1995)from J774 cells were dialyzed against PBS and GAPDH was purifiedusing 5′-AMP-Sepharose4B (Amersham Pharmacia), purificationwas confirmed by western blot.
2.7. N-terminal sequencing and glycosylation
Approximately, 10–15 �g of GAPDH from both sources wastransferred to PVDF and N-terminal sequencing was done usingApplied Biosystems PROCISE 491CLC Protein Sequencer. Cytoso-lic and membrane protein fractions of J774 cells were checked forglycosylation using GelCodeR Glycoprotein staining kit (Pierce).
2.8. GAPDH is not a peripherally associated membrane protein
J774 cells were washed with chilled neutral bufferfollowed by treatment with 2 M urea and 20 mM 2-(N-morpholino)ethanesulfonic acid (MES), pH 6.5, or a solutioncontaining 10 mM Tris–Cl, pH 8.0 and 0.5 M NaCl for 15 min at 4 ◦C(Schwab et al., 2000). Residual cell surface GAPDH was quantitatedby FACS. Additionally, membrane fractions were purified and theassociated GAPDH was detected by western blotting.
2.9. Co-immunoprecipitation of GAPDH from Lipid raft bytransferrin
CHO-TRVb cells (4.0 × 107) were incubated with 12.5 �Mbiotinylated Tf at 4 ◦C for 2 h and lipid raft fraction was puri-fied as described (Brown, 2002). Fractions were resuspendedand co-immunoprecipitated using Streptavidin Magnabeads (Poly-sciences) followed by detection of GAPDH.
2.10. Purification and analysis of DRM and non DRM fractions
Membrane fractions from control or iron depleted CHO-TRVbcells, were solubilized overnight in buffer containing 0.2% Tri-ton X-100 and centrifuged at 1,00,000 × g for 1 h. The non-DRM(non-detergent resistant membrane or detergent soluble) and DRMfractions were prepared and the GAPDH content was evaluated bywestern blot as before.
2.11. Flow cytometric differential detergent resistant ratio (FCDR)of GAPDH and Tf
Control or iron depleted CHO-TRVb, CHO and J774 cells werestained with Tf-Alexa633 or �GAPDH to determine specific binding.Stained cells were then left untreated or treated with 0.2% TritonX-100 at 25 ◦C for 5 min and analyzed by FACS to quantitate theamount of transferrin/GAPDH remaining (Gombos et al., 2004).
2.12. GAPDH–transferrin binding and uptake is specific andpronase sensitive
CHO-TRVb cells were treated with 0.1% pronase (Roche) for30 min at 4 ◦C, and washed with SFM, stained with holo-Tf-Alexa633 at 4 ◦C or 37 ◦C for 1 h alone or in the presence of 200×unlabeled holo-transferrin. All samples were analyzed by FACS.
2.13. GAPDH–transferrin affinity on cell surface
CHO-TRVb cells were incubated with different concentrationsof biotinylated Tf (Sigma) alone or in the presence of 200× unla-beled transferrin for 2 h at 4 ◦C. This was followed by incubation

f Bioch
wtGG
2i
wMwwFt2(Fis
2
c3fbaats
wstmsscoa
2
tAmre
2
Toatr(
2
w
S. Kumar et al. / The International Journal o
ith 1:2000 diluted Streptavidin–HRP for 45 min at 4 ◦C. The reac-ion was developed with TMB H2O2 substrate for ELISA (Bangaloreenei). Data was fitted for one site specific saturable binding usingraph Pad software.
.14. GAPDH associated transferrin is localized to andnternalized by lipid rafts
The rat GAPDH–EGFP/pcDNA3 vector and pmCherry-N1 vectorere received as gifts from Prof. S. Sealfon, Mount Sinai School ofedicine, and Dr. J. Lippincott Schwartz, NIH, respectively. GAPDHas amplified and cloned in frame with mCherry. CHO-TRVb cellsere transfected with this vector using Ex-Gen500 (Fermentas).
or confocal microscopy, cells were seeded overnight in glass bot-omed petri dishes, rinsed with SFM, blocked for 30 min at 4 ◦C with% BSA and incubated with Tf–FITC and Cholera toxin – Alexa647CTX–Alexa647, Invitrogen, a lipid raft marker) for 30 min at 37 ◦C.inally, cells were resuspended in media containing 10% FCS andmaged live at 37 ◦C on a Zeiss LSM 510META Laser Confocal Micro-cope, using an airflow incubator (Raje et al., 2007).
.15. Internalization assays of GAPDH and transferrin
To measure the kinetics of transferrin internalization, 2 × 105
ells per tube CHO-TRVb cells were incubated in SFM for 1 h at7 ◦C. After addition of 10 �g of Tf-Alexa633, cells were culturedor different periods of time at 37 ◦C or at 4 ◦C for 2 h. 37 ◦C incu-ated cells were additionally treated with 0.1% pronase for 15 mint 4 ◦C to remove surface label. All samples were analyzed by FACSnd the internal to surface ratio was calculated and plotted versusime (McGraw and Subtil, 1999). All values were corrected for non-pecific signal.
To measure cell surface GAPDH internalization, CHO-TRVb cellsere stained with �GAPDH antibody or isotype control on ice. Sub-
equently cells were incubated in pre-warmed SFM for differentime points at 37 ◦C to allow for internalization. The amount of pri-
ary antibody remaining on the cell surface was quantitated bytaining with Sheep �mouse-FITC (Fab)2 followed by FACS analy-is. Values were corrected for non-specific signal using an isotypeontrol. The MFI for each time point was expressed as % of the valuebtained for cells that had been kept on ice for the duration of thessay which represents 100% surface signal (Walseng et al., 2008).
.16. GAPDH–transferrin uptake is saturable
CHO-TRVb cells were incubated for 1hr in SFM, followed by addi-ion of increasing concentrations of 125I-transferrin at 37 ◦C for 2 h.fter removal of surface associated transferrin by pronase treat-ent, cells were solubilized using 0.5 M NaOH and internalized
adioactivity measured. To determine non-specific uptake a parallelxperiment was set up using 200× unlabeled Tf.
.17. Degradation of internalized transferrin
CHO-TRVb cells were incubated in SFM containing 500 nM 125I-f at 37◦C for 2 h. Cells were then washed and incubated in 250 �lf pre warmed SFM for different times points at 37 ◦C. The cellssociated radioactivity was determined. Separately proteins inhe culture supernatant were precipitated as described earlier andadioactivity in this pellet (protein associated 125I) and supernatantdegraded Tf) were measured (McGraw and Subtil, 1999).
.18. Recycling of GAPDH
Recycling of cell surface GAPDH in CHO-TRVb and J774 cellsas compared by antibody based method (Walseng et al., 2008).
emistry & Cell Biology 44 (2012) 189– 199 191
In addition, the kinetics of internalization was measured by ligandbased assay (McGraw and Subtil, 1999) using Alexa633 labelledtransferrin in CHO-TRVb cells.
2.19. Inhibition of Tf internalization
Transferrin internalization in CHO-TRVb cells was measured inthe presence of inhibitors of endocytosis. The inhibitors selectedwere used essentially as described previously, these were (i)lipid raft endocytosis inhibitors nystatin and filipin (Ivanov, 2008;Orlandi and Fishman, 1998), (ii) inhibitors of macropinocytosis,i.e. nocadazole, cytochalasin D, wortmanin and amiloride (Anientoand Gruenberg, 2003; Harding, 1997; Malide and Cushman, 1997;Muro et al., 2003), (iii) clathrin-pit mediated endocytosis inhibitorspotassium depletion, chlorpromazine and monodansyl cadaverine(McGraw and Subtil, 1999; Pho et al., 2000; Schutze et al., 1999),(iv) sodium azide and 2-deoxy-glucose to determine if the processis energy dependent (Reid and Watts, 1992)
3. Results
3.1. Iron depletion modulates cell surface expression of GAPDH,TfR1 and transferrin binding
Previously, we had observed that GAPDH is a transferrin recep-tor in macrophages and its cell surface expression is upregulatedon iron starvation (Raje et al., 2007). To ascertain whether this is ageneralized mechanism of transferrin uptake several cell lines werescreened.
After confirmation of iron depletion by atomic absorption spec-troscopy (Supplementary material Fig. S1) the change in cell surfaceGAPDH, TfR1 and transferrin binding were assessed by FACS anal-ysis. GAPDH was found to be present on a majority of the cell linestested. Iron depletion resulted in a significant increase in surfaceGAPDH, with the maximal increase in L1210, BV2, NS, J774, RAWand N2A cells (p < 0.0005, n = 3) (Supplementary material Figs. S2and S3). No expression was observed in K562 and Hela cells cells(Supplementary material Fig. S3).
Interestingly, in cells that showed the maximal increase inGAPDH, TfR1 levels decreased upon iron depletion (Supplementarymaterial Figs. S2 and S3). In contrast K562 and CHO cells exhib-ited increased cell surface TfR1 (Supplementary material Fig. S3).The increase in transferrin binding correlated significantly witheither the observed increase in GAPDH (p < 0.0002, n = 3) or TfR1 (inK562 cells). In CHO-TRVb cells, no TfR1 was detected, however bothcell surface GAPDH and Tf binding increased with DFO treatment(p < 0.0001, n = 3) (Supplementary material Fig. S2). These cells havebeen reported to acquire iron via transferrin dependent but “trans-ferrin receptor-independent” mechanisms (Chan et al., 1992).
3.2. GAPDH mediates transferrin binding and iron uptake in cells
In order to establish that GAPDH is a novel mechanism by whichcells acquire iron, the CHO-TRVb cell line which does not expressTfR1 for transferrin iron uptake was utilized. Cellular GAPDHwas silenced by 66%, as compared to siRNA control transfections(Supplementary material Figs. S4A and S4B). Both surface GAPDHand transferrin binding were reduced in these cells as compared tocontrols (Supplementary material Figs. S4C and S4D). Upon GAPDHknockdown, DFO treatment no longer resulted in an increase ineither surface GAPDH or transferrin binding (Fig. 1A and C). How-ever in controls (Fig. 1B and D) a 74% increase in GAPDH levels and
34% increase of transferrin binding was observed. GAPDH silencingresulted in a 30% decrease in Tf mediated 55Fe uptake upon DFOtreatment as compared to controls (Fig. 1E). Knockdown was alsoconfirmed by western blotting (Fig. 1F).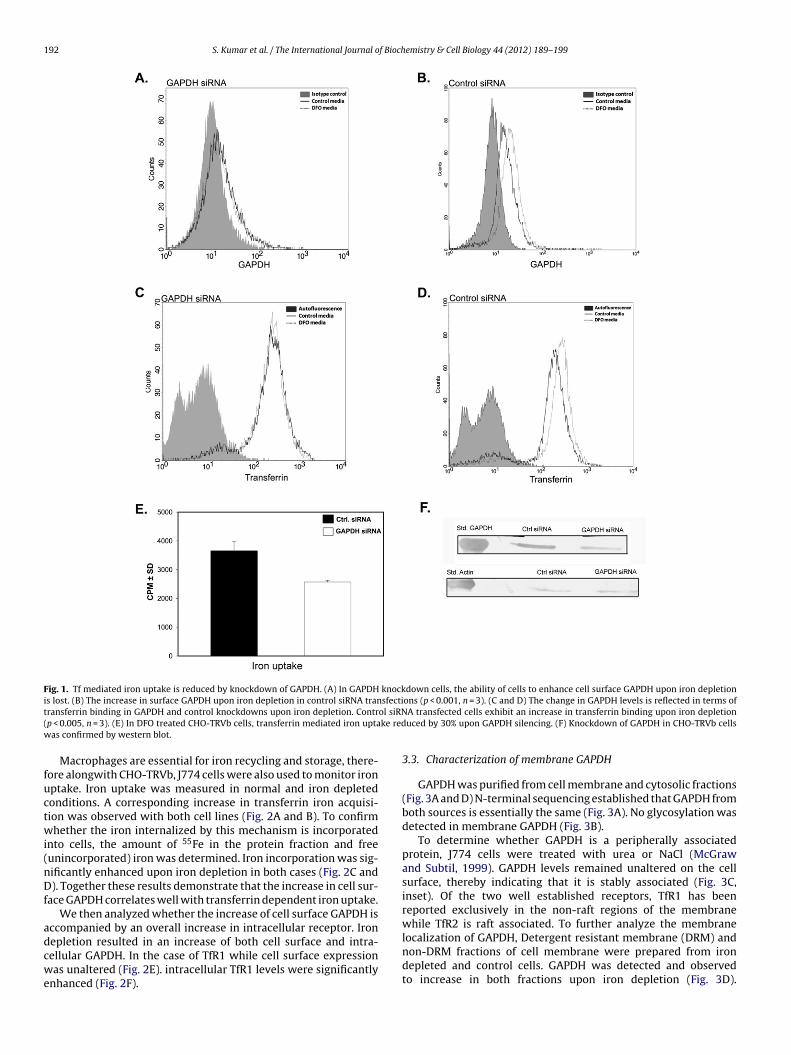
192 S. Kumar et al. / The International Journal of Biochemistry & Cell Biology 44 (2012) 189– 199
Fig. 1. Tf mediated iron uptake is reduced by knockdown of GAPDH. (A) In GAPDH knockdown cells, the ability of cells to enhance cell surface GAPDH upon iron depletioni sfectit ol siRN( ke redw
fuctwi(nDf
adcwe
s lost. (B) The increase in surface GAPDH upon iron depletion in control siRNA tranransferrin binding in GAPDH and control knockdowns upon iron depletion. Contrp < 0.005, n = 3). (E) In DFO treated CHO-TRVb cells, transferrin mediated iron uptaas confirmed by western blot.
Macrophages are essential for iron recycling and storage, there-ore alongwith CHO-TRVb, J774 cells were also used to monitor ironptake. Iron uptake was measured in normal and iron depletedonditions. A corresponding increase in transferrin iron acquisi-ion was observed with both cell lines (Fig. 2A and B). To confirmhether the iron internalized by this mechanism is incorporated
nto cells, the amount of 55Fe in the protein fraction and freeunincorporated) iron was determined. Iron incorporation was sig-ificantly enhanced upon iron depletion in both cases (Fig. 2C and). Together these results demonstrate that the increase in cell sur-
ace GAPDH correlates well with transferrin dependent iron uptake.We then analyzed whether the increase of cell surface GAPDH is
ccompanied by an overall increase in intracellular receptor. Iron
epletion resulted in an increase of both cell surface and intra-ellular GAPDH. In the case of TfR1 while cell surface expressionas unaltered (Fig. 2E). intracellular TfR1 levels were significantlynhanced (Fig. 2F).
ons (p < 0.001, n = 3). (C and D) The change in GAPDH levels is reflected in terms ofA transfected cells exhibit an increase in transferrin binding upon iron depletionuced by 30% upon GAPDH silencing. (F) Knockdown of GAPDH in CHO-TRVb cells
3.3. Characterization of membrane GAPDH
GAPDH was purified from cell membrane and cytosolic fractions(Fig. 3A and D) N-terminal sequencing established that GAPDH fromboth sources is essentially the same (Fig. 3A). No glycosylation wasdetected in membrane GAPDH (Fig. 3B).
To determine whether GAPDH is a peripherally associatedprotein, J774 cells were treated with urea or NaCl (McGrawand Subtil, 1999). GAPDH levels remained unaltered on the cellsurface, thereby indicating that it is stably associated (Fig. 3C,inset). Of the two well established receptors, TfR1 has beenreported exclusively in the non-raft regions of the membranewhile TfR2 is raft associated. To further analyze the membrane
localization of GAPDH, Detergent resistant membrane (DRM) andnon-DRM fractions of cell membrane were prepared from irondepleted and control cells. GAPDH was detected and observedto increase in both fractions upon iron depletion (Fig. 3D).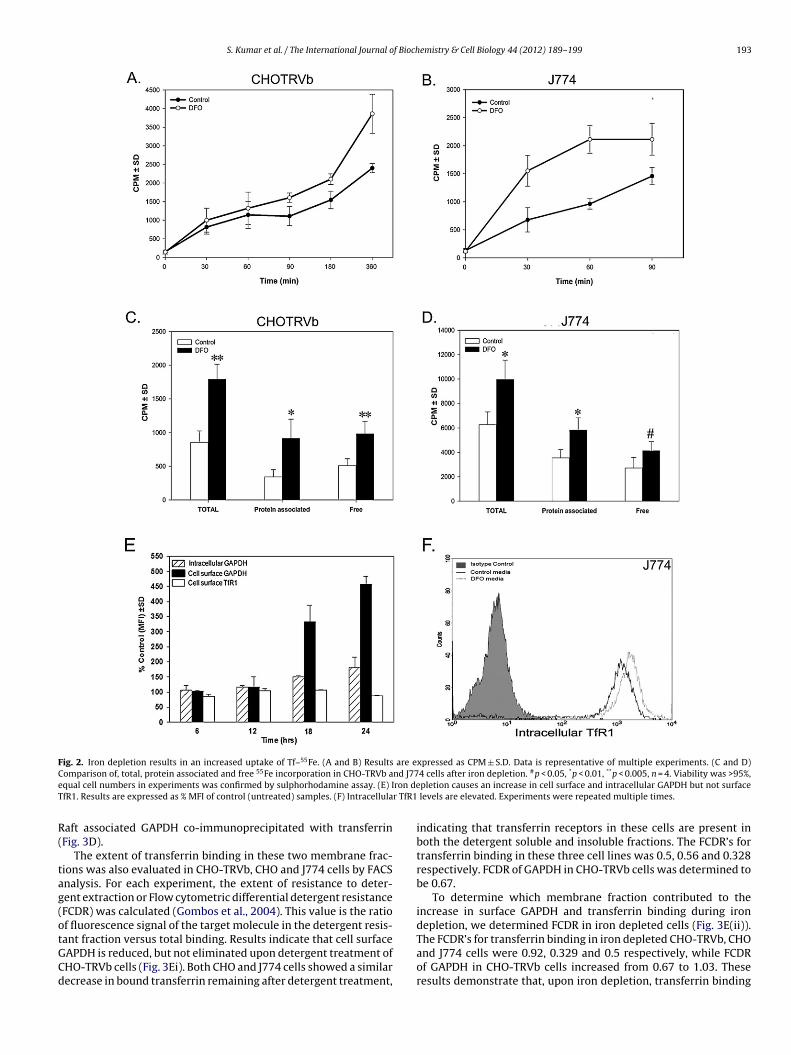
S. Kumar et al. / The International Journal of Biochemistry & Cell Biology 44 (2012) 189– 199 193
Fig. 2. Iron depletion results in an increased uptake of Tf–55Fe. (A and B) Results are expressed as CPM ± S.D. Data is representative of multiple experiments. (C and D)C nd J77e ron deT r TfR1
R(
tag(otGCd
omparison of, total, protein associated and free 55Fe incorporation in CHO-TRVb aqual cell numbers in experiments was confirmed by sulphorhodamine assay. (E) IfR1. Results are expressed as % MFI of control (untreated) samples. (F) Intracellula
aft associated GAPDH co-immunoprecipitated with transferrinFig. 3D).
The extent of transferrin binding in these two membrane frac-ions was also evaluated in CHO-TRVb, CHO and J774 cells by FACSnalysis. For each experiment, the extent of resistance to deter-ent extraction or Flow cytometric differential detergent resistanceFCDR) was calculated (Gombos et al., 2004). This value is the ratiof fluorescence signal of the target molecule in the detergent resis-
ant fraction versus total binding. Results indicate that cell surfaceAPDH is reduced, but not eliminated upon detergent treatment ofHO-TRVb cells (Fig. 3Ei). Both CHO and J774 cells showed a similarecrease in bound transferrin remaining after detergent treatment,4 cells after iron depletion. #p < 0.05, *p < 0.01, **p < 0.005, n = 4. Viability was >95%,pletion causes an increase in cell surface and intracellular GAPDH but not surface
levels are elevated. Experiments were repeated multiple times.
indicating that transferrin receptors in these cells are present inboth the detergent soluble and insoluble fractions. The FCDR’s fortransferrin binding in these three cell lines was 0.5, 0.56 and 0.328respectively. FCDR of GAPDH in CHO-TRVb cells was determined tobe 0.67.
To determine which membrane fraction contributed to theincrease in surface GAPDH and transferrin binding during irondepletion, we determined FCDR in iron depleted cells (Fig. 3E(ii)).
The FCDR’s for transferrin binding in iron depleted CHO-TRVb, CHOand J774 cells were 0.92, 0.329 and 0.5 respectively, while FCDRof GAPDH in CHO-TRVb cells increased from 0.67 to 1.03. Theseresults demonstrate that, upon iron depletion, transferrin binding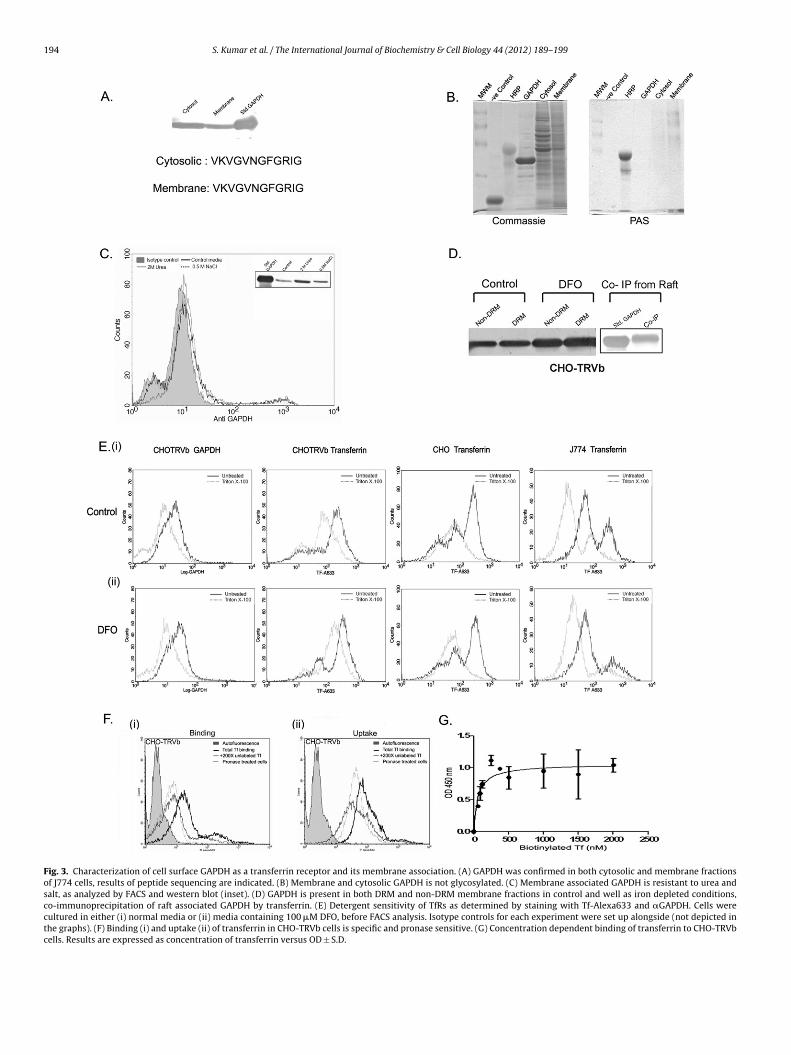
194 S. Kumar et al. / The International Journal of Biochemistry & Cell Biology 44 (2012) 189– 199
Fig. 3. Characterization of cell surface GAPDH as a transferrin receptor and its membrane association. (A) GAPDH was confirmed in both cytosolic and membrane fractionsof J774 cells, results of peptide sequencing are indicated. (B) Membrane and cytosolic GAPDH is not glycosylated. (C) Membrane associated GAPDH is resistant to urea andsalt, as analyzed by FACS and western blot (inset). (D) GAPDH is present in both DRM and non-DRM membrane fractions in control and well as iron depleted conditions,co-immunoprecipitation of raft associated GAPDH by transferrin. (E) Detergent sensitivity of TfRs as determined by staining with Tf-Alexa633 and �GAPDH. Cells werecultured in either (i) normal media or (ii) media containing 100 �M DFO, before FACS analysis. Isotype controls for each experiment were set up alongside (not depicted inthe graphs). (F) Binding (i) and uptake (ii) of transferrin in CHO-TRVb cells is specific and pronase sensitive. (G) Concentration dependent binding of transferrin to CHO-TRVbcells. Results are expressed as concentration of transferrin versus OD ± S.D.

f Bioch
igifcdi
dsem
ptcnGst
cfs(
3
c0ra
u1(
cri
3
itTcfirCffTmrtwwi
nbeo
S. Kumar et al. / The International Journal o
n both CHO-TRVb and J774 cells is enhanced mostly in the deter-ent insoluble membrane fraction. Although J774 cells express TfR1n addition to GAPDH, it is solely present in the detergent solubleraction and therefore the observed increase in transferrin FCDRan only be due to the presence of more GAPDH receptor in theetergent insoluble fraction. This increase was much more evident
n the CHO-TRVb cell line.The decrease in FCDR for transferrin in CHO cells upon iron
epletion is due to the increased presentation of TfR1 on the cellurface (Supplementary material Fig. S3) which is known to bexclusively localized to the detergent soluble membrane compart-ent.To establish whether GAPDH fulfils the criteria of a specific
rotein receptor we measured the binding and uptake of labelledransferrin in the presence of 200× unlabelled transferrin, and inells pre-treated with pronase. Under both these conditions, a sig-ificant inhibition was observed, indicating that the process ofAPDH mediated transferrin delivery is specific and pronase sen-itive (Fig. 3F). The in vivo KD values of this interaction was foundo be 60 nM (Fig. 3G).
CHO-TRVb cells expressing GAPDH-mCherry, were analyzed byonfocal microscopy, for the binding and internalization of trans-errin along with Cholera toxin (CTX). We observed that all threeignals co-localized on the cell surface and the early endosomesFig. 4A).
.4. Kinetics of GAPDH–transferrin uptake
The internalization rate constant of transferrin in CHO-TRVbells (which lack other known TfRs) was estimated to be.08 ± 0.023 min−1 (Fig. 4B). Cell surface GAPDH internalizationate constant was also determined directly by antibody based FACSnalysis. This gave a closely matching value of 0.1 min−1 (Fig. 4C).
Concentration dependent 125I-labelled transferrin (125I–Tf)ptake analysis demonstrates that the process is saturable (Fig. 5A).5–25% of the internalized transferrin is degraded within 15 minFig. 5B).
Using monoclonal anti-GAPDH we found that the receptor recy-ling is similar in both CHO-TRVb and J774 cells (Fig. 5C). Theecycling rate of the ligand (transferrin) was found to be 0.08 min−1
n CHO-TRVb cells (Fig. 5D).
.5. Inhibition of GAPDH–Tf internalization
To understand the mechanism of transferrin–GAPDH internal-zation, we utilized the CHO-TRVb and CHO cell lines and studiedhe effect of various inhibitors on transferrin uptake (Fig. 5E). CHO-RVb cells acquire transferrin only via GAPDH while CHO cellsan utilize both TfR1 and GAPDH. Treatment with nystatin andlipin (that affect cholesterol sequestration within membranes)esulted in a 40–60%, inhibition of transferrin internalization inHO-TRVb cells. On the other hand, transferrin uptake was unaf-ected in CHO cells since these cells can continue to utilize TfR1. Thisurther confirms that GAPDH–Tf uptake is also lipid raft mediated.he PI3K inhibitor, wortmannin blocks constitutive and stimulatedacropinocytosis (Araki et al., 1996). In our experiments transfer-
in uptake in CHO-TRVb was reduced by 70% upon wortmanninreatment while CHO cells remained unaffected. Both cell linesere sensitive to treatment with Cytochalasin D (p < 0.0001, n = 3),ith CHO-TRVb showing almost 90% inhibition as compared to 50%
nhibition in CHO cells.Although originally no effect of amiloride was found on Tf inter-
alization by the classical clathrin pathway, subsequently it haseen reported that this inhibitor may also block clathrin-mediatedndocytosis (Ivanov, 2008). In our experiments, 20% inhibition wasbserved in CHO-TRVb cells with no effect on CHO cells.
emistry & Cell Biology 44 (2012) 189– 199 195
The microtubule disrupting agent nocodazole, caused a deple-tion of 90%, in transferrin uptake by CHO-TRVb cells with 10%inhibition in CHO cells. In some cell types nocadazole treatmenthas reported to have no effect on endocytosis suggesting thatthe cell specific cytoskeletal organization may influence trafficking(Gaidarov et al., 1999). Inhibitors for clathrin mediated endocytosissignificantly inhibited transferrin internalization (p< 0.0001, n = 3)for both cell lines. Treatment with monodansyl cadaverine (MDC),chlorpromazine and potassium depletion resulted in a 30–40% inhi-bition, suggesting that GAPDH–Tf uptake also occurs by clathrinmediated endocytosis. Finally, treatment with sodium azide and2-deoxy glucose resulted in a 60–80% inhibition which correlateswith previous studies on TfR1 (p < 0.0001, n = 3) (Klausner et al.,1983), indicating that it is an energy dependent process.
4. Discussion
Despite the discovery of TfR1 and TfR2 (Jandl et al., 1959;Kawabata et al., 1999) several unanswered questions regardingreceptor mediated transferrin uptake remain. The existence of thisadditional TfR is supported by previous reports wherein TfR1−/−
knockout mice embryos were observed to survive until E10-12.5indicating that though TfR1 is an essential pathway, alternateroutes of transferrin uptake do exist (Levy et al., 1999). Numerousstudies with, melanoma, CHO-TRVb and hepatoma cell lines haveindicated the presence of an additional saturable, pH dependent,pronase sensitive non transferrin receptor mediated mechanismfor transferrin uptake (Chan et al., 1992; McGraw et al., 1987;Richardson and Baker, 1990, 1994; Trinder and Morgan, 1997;Trinder et al., 1996). With the discovery of TfR2 it was initiallythought that this receptor was involved in the non-TfR1 pathway,however, TfR2 is not expressed on numerous cell lines includingCHO-TRVb (Calzolari et al., 2007; Kawabata et al., 1999). The mostconvincing results are from extensive studies done in the humanhepatoma cell line HuH7 that express both TfR1 and TfR2. WhenTfR1 expression was knocked down, TfR2 by itself could not accountfor the extent of transferrin uptake observed (Trinder and Morgan,1997). In this study we knocked down GAPDH in CHO-TRVb cellsthat lack TfR1 and 2, to demonstrate that transferrin uptake bythis receptor can account for the above mentioned discrepancies.We also describe the detailed mechanism of Tf uptake via GAPDH,which differs significantly from the two known TfRs.
To determine whether GAPDH is involved in the acquisition andincorporation of transferrin iron, we chose the mouse macrophagecell line J774 as a model system. As these cells express TfR1in addition to GAPDH on their surface we carried out parallelexperiments in the CHO-TRVb cell line, where there would beno interference from other receptors of transferrin. In agreementwith previous reports (Bottomley et al., 1985; Mattia et al., 1984;Rao et al., 1985; Rudolph et al., 1985; Testa et al., 1985), we tooobserved an enhanced transferrin binding and incorporation ofiron upon iron depletion. However, in contrast, we observed thatthis was due to the increase in cell surface GAPDH rather thanincreased surface expression of TfR1. To some extent this can beexplained by analyzing the experimental protocols. Several studieshave measured the enhanced binding of labelled transferrin andattributed this to TfR1 expression alone without considering theexistence of an alternate receptor. An earlier study has infactshown that when hepatocytes were incubated with an antibodythat blocks transferrin binding to the transferrin receptor, cellswere still able to acquire iron from transferrin (Trinder et al., 1988).
In other studies, the N-terminal half of the transferrin moleculewas shown to be incapable of binding TfR, but it could still donateiron to hepatocytes (Thorstensen et al., 1995; Zak et al., 1994). Amajority of studies on transferrin iron acquisition have utilized the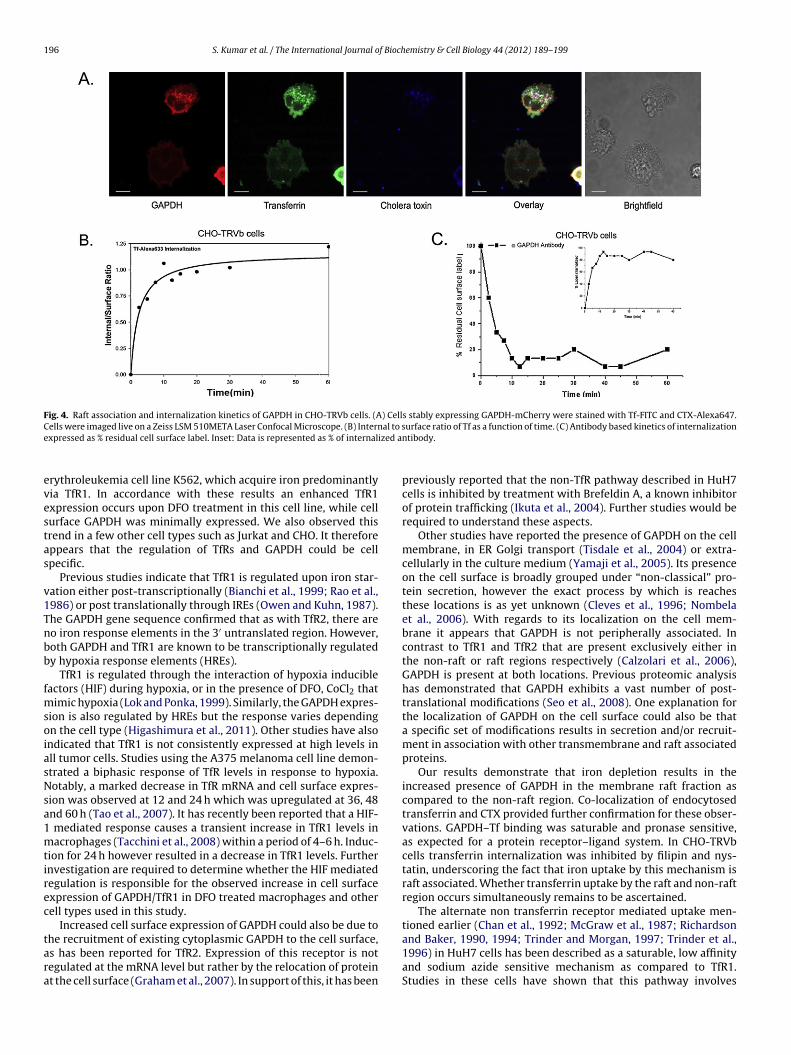
196 S. Kumar et al. / The International Journal of Biochemistry & Cell Biology 44 (2012) 189– 199
Fig. 4. Raft association and internalization kinetics of GAPDH in CHO-TRVb cells. (A) Cells stably expressing GAPDH-mCherry were stained with Tf-FITC and CTX-Alexa647.C al to se ized a
evestas
v1Tnbb
fmsoiasNsa1mtirec
tara
ells were imaged live on a Zeiss LSM 510META Laser Confocal Microscope. (B) Internxpressed as % residual cell surface label. Inset: Data is represented as % of internal
rythroleukemia cell line K562, which acquire iron predominantlyia TfR1. In accordance with these results an enhanced TfR1xpression occurs upon DFO treatment in this cell line, while cellurface GAPDH was minimally expressed. We also observed thisrend in a few other cell types such as Jurkat and CHO. It thereforeppears that the regulation of TfRs and GAPDH could be cellpecific.
Previous studies indicate that TfR1 is regulated upon iron star-ation either post-transcriptionally (Bianchi et al., 1999; Rao et al.,986) or post translationally through IREs (Owen and Kuhn, 1987).he GAPDH gene sequence confirmed that as with TfR2, there areo iron response elements in the 3′ untranslated region. However,oth GAPDH and TfR1 are known to be transcriptionally regulatedy hypoxia response elements (HREs).
TfR1 is regulated through the interaction of hypoxia inducibleactors (HIF) during hypoxia, or in the presence of DFO, CoCl2 that
imic hypoxia (Lok and Ponka, 1999). Similarly, the GAPDH expres-ion is also regulated by HREs but the response varies dependingn the cell type (Higashimura et al., 2011). Other studies have alsondicated that TfR1 is not consistently expressed at high levels inll tumor cells. Studies using the A375 melanoma cell line demon-trated a biphasic response of TfR levels in response to hypoxia.otably, a marked decrease in TfR mRNA and cell surface expres-
ion was observed at 12 and 24 h which was upregulated at 36, 48nd 60 h (Tao et al., 2007). It has recently been reported that a HIF-
mediated response causes a transient increase in TfR1 levels inacrophages (Tacchini et al., 2008) within a period of 4–6 h. Induc-
ion for 24 h however resulted in a decrease in TfR1 levels. Furthernvestigation are required to determine whether the HIF mediatedegulation is responsible for the observed increase in cell surfacexpression of GAPDH/TfR1 in DFO treated macrophages and otherell types used in this study.
Increased cell surface expression of GAPDH could also be due to
he recruitment of existing cytoplasmic GAPDH to the cell surface,s has been reported for TfR2. Expression of this receptor is notegulated at the mRNA level but rather by the relocation of proteint the cell surface (Graham et al., 2007). In support of this, it has beenurface ratio of Tf as a function of time. (C) Antibody based kinetics of internalizationntibody.
previously reported that the non-TfR pathway described in HuH7cells is inhibited by treatment with Brefeldin A, a known inhibitorof protein trafficking (Ikuta et al., 2004). Further studies would berequired to understand these aspects.
Other studies have reported the presence of GAPDH on the cellmembrane, in ER Golgi transport (Tisdale et al., 2004) or extra-cellularly in the culture medium (Yamaji et al., 2005). Its presenceon the cell surface is broadly grouped under “non-classical” pro-tein secretion, however the exact process by which is reachesthese locations is as yet unknown (Cleves et al., 1996; Nombelaet al., 2006). With regards to its localization on the cell mem-brane it appears that GAPDH is not peripherally associated. Incontrast to TfR1 and TfR2 that are present exclusively either inthe non-raft or raft regions respectively (Calzolari et al., 2006),GAPDH is present at both locations. Previous proteomic analysishas demonstrated that GAPDH exhibits a vast number of post-translational modifications (Seo et al., 2008). One explanation forthe localization of GAPDH on the cell surface could also be thata specific set of modifications results in secretion and/or recruit-ment in association with other transmembrane and raft associatedproteins.
Our results demonstrate that iron depletion results in theincreased presence of GAPDH in the membrane raft fraction ascompared to the non-raft region. Co-localization of endocytosedtransferrin and CTX provided further confirmation for these obser-vations. GAPDH–Tf binding was saturable and pronase sensitive,as expected for a protein receptor–ligand system. In CHO-TRVbcells transferrin internalization was inhibited by filipin and nys-tatin, underscoring the fact that iron uptake by this mechanism israft associated. Whether transferrin uptake by the raft and non-raftregion occurs simultaneously remains to be ascertained.
The alternate non transferrin receptor mediated uptake men-tioned earlier (Chan et al., 1992; McGraw et al., 1987; Richardson
and Baker, 1990, 1994; Trinder and Morgan, 1997; Trinder et al.,1996) in HuH7 cells has been described as a saturable, low affinityand sodium azide sensitive mechanism as compared to TfR1.Studies in these cells have shown that this pathway involves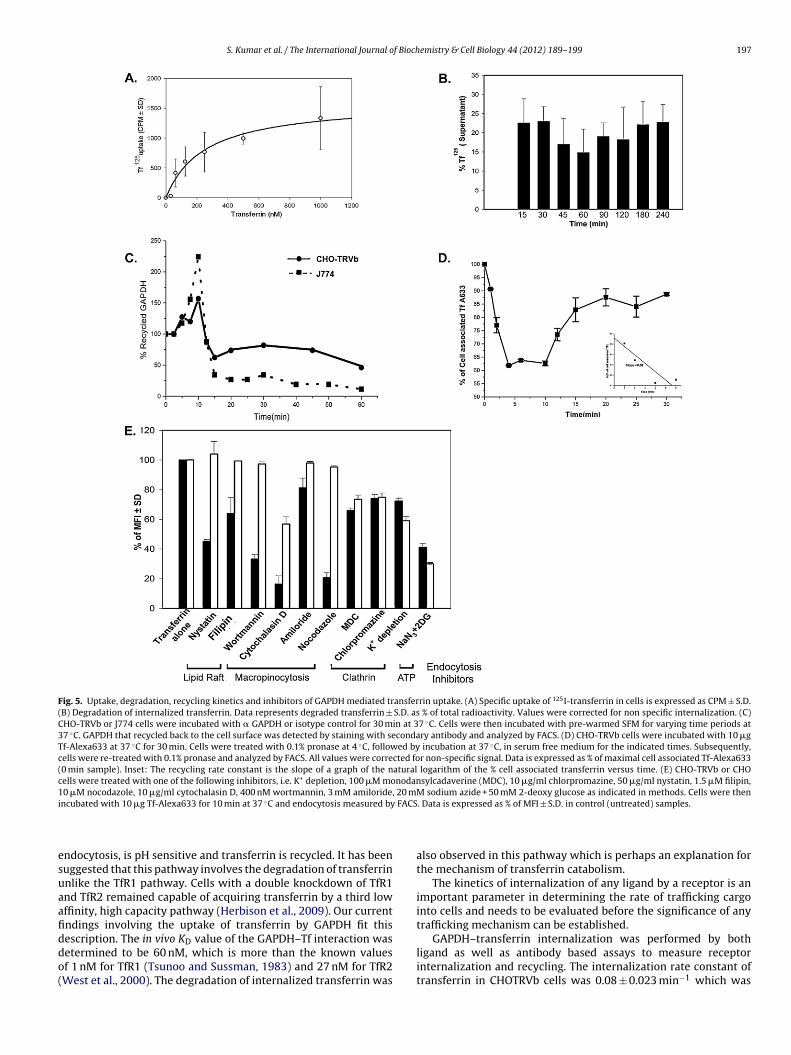
S. Kumar et al. / The International Journal of Biochemistry & Cell Biology 44 (2012) 189– 199 197
Fig. 5. Uptake, degradation, recycling kinetics and inhibitors of GAPDH mediated transferrin uptake. (A) Specific uptake of 125I-transferrin in cells is expressed as CPM ± S.D.(B) Degradation of internalized transferrin. Data represents degraded transferrin ± S.D. as % of total radioactivity. Values were corrected for non specific internalization. (C)CHO-TRVb or J774 cells were incubated with � GAPDH or isotype control for 30 min at 37 ◦C. Cells were then incubated with pre-warmed SFM for varying time periods at37 ◦C. GAPDH that recycled back to the cell surface was detected by staining with secondary antibody and analyzed by FACS. (D) CHO-TRVb cells were incubated with 10 �gTf-Alexa633 at 37 ◦C for 30 min. Cells were treated with 0.1% pronase at 4 ◦C, followed by incubation at 37 ◦C, in serum free medium for the indicated times. Subsequently,cells were re-treated with 0.1% pronase and analyzed by FACS. All values were corrected for non-specific signal. Data is expressed as % of maximal cell associated Tf-Alexa633(0 min sample). Inset: The recycling rate constant is the slope of a graph of the natural logarithm of the % cell associated transferrin versus time. (E) CHO-TRVb or CHOcells were treated with one of the following inhibitors, i.e. K+ depletion, 100 �M monodansylcadaverine (MDC), 10 �g/ml chlorpromazine, 50 �g/ml nystatin, 1.5 �M filipin,10 �M nocodazole, 10 �g/ml cytochalasin D, 400 nM wortmannin, 3 mM amiloride, 20 mM sodium azide + 50 mM 2-deoxy glucose as indicated in methods. Cells were theni FACS
esuaafiddo(
ncubated with 10 �g Tf-Alexa633 for 10 min at 37 ◦C and endocytosis measured by
ndocytosis, is pH sensitive and transferrin is recycled. It has beenuggested that this pathway involves the degradation of transferrinnlike the TfR1 pathway. Cells with a double knockdown of TfR1nd TfR2 remained capable of acquiring transferrin by a third lowffinity, high capacity pathway (Herbison et al., 2009). Our currentndings involving the uptake of transferrin by GAPDH fit this
escription. The in vivo KD value of the GAPDH–Tf interaction wasetermined to be 60 nM, which is more than the known valuesf 1 nM for TfR1 (Tsunoo and Sussman, 1983) and 27 nM for TfR2West et al., 2000). The degradation of internalized transferrin was. Data is expressed as % of MFI ± S.D. in control (untreated) samples.
also observed in this pathway which is perhaps an explanation forthe mechanism of transferrin catabolism.
The kinetics of internalization of any ligand by a receptor is animportant parameter in determining the rate of trafficking cargointo cells and needs to be evaluated before the significance of anytrafficking mechanism can be established.
GAPDH–transferrin internalization was performed by bothligand as well as antibody based assays to measure receptorinternalization and recycling. The internalization rate constant oftransferrin in CHOTRVb cells was 0.08 ± 0.023 min−1 which was

1 f Bioch
sco1
maoGcrmtt
trrafvrtati
A
gJcaMRbg
A
t
R
A
A
B
B
B
C
C
C
C
C
G
98 S. Kumar et al. / The International Journal o
imilar to that for GAPDH by the antibody based assay. The recy-ling rate for GAPDH was 0.08 min−1. The recycling rate constantf TfR1 has earlier been reported to be 0.06 min−1 (Johnson et al.,994).
Both TfR1–Tf and TfR2–Tf endocytosis occurs by the clathrinediated pathway (Chen et al., 2009; Trowbridge et al., 1993)
dditionally TfR2 trafficking also occurs by lipid raft or cave-lin mediated pathways (Calzolari et al., 2006). Our results proveAPDH–Tf internalization occurs by both clathrin mediated endo-ytosis and lipid raft endocytosis. Diverging from the other twoeceptors GAPDH–Tf appears to be trafficked to some extent byacropinocytosis. Transferrin internalization by all three recep-
ors, i.e. TfR1 (Klausner et al., 1983), TfR2 and GAPDH are sensitiveo the metabolic inhibitor sodium azide.
Taken together, our findings suggest that depending on the cellype, transferrin bound iron can be acquired by three transferrineceptors TfR1, TfR2 or GAPDH. The discovery of this third transfer-in receptor is unusual given that it is a glycolytic enzyme lackingny similarity to the previously identified receptors. As a trans-errin receptor, GAPDH has properties that are common to and atariance with existing TfRs. The discovery of GAPDH as a transferrineceptor offers an explanation for the “non-TfR mediated” uptake ofransferrin iron that has repeatedly been discussed, but has lacked
complete explanation. Further analysis of GAPDH and its regula-ion is essential to uncover the significance of this new receptor inron metabolism.
cknowledgements
We are extremely grateful to Dr. Satyajit Mayor, NCBS for hisenerous help with cell lines and reagents. Drs. A. Basu and N.R.ana, NBRC and S. Majumdar, IMTECH are thanked for providingell lines. Mr. Anil Theophilus and Mr. Himanshu Malhotra arecknowledged for assistance with experiments. Thanks are due tor. Vishant Boradia for proof reading. S.K. and N.S. received CSIR,
esearch Fellowships. Financial support for M.A.M. was providedy NIPER, SAS Nagar. The financial support by DST and DST WOS(A)rants is acknowledged. IMTECH communication no. 054/2010.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.biocel.2011.10.016.
eferences
niento F, Gruenberg J. Current Protocols in Immunology. New York: John Wiley &Sons; 2003. pp. 8.1C.1–8.1C.21.
raki N, Johnson MT, Swanson JA. A role for phosphoinositide 3-kinase in thecompletion of macropinocytosis and phagocytosis by macrophages. J Cell Biol1996;135(5):1249–60.
ianchi L, Tacchini L, Cairo G. HIF-1-mediated activation of transferrin receptor genetranscription by iron chelation. Nucleic Acids Res 1999;27(21):4223–7.
ottomley SS, Wolfe LC, Bridges KR. Iron metabolism in K562 erythroleukemic cells.J Biol Chem 1985;260(11):6811–5.
rown DA. Current Protocols in Immunology. New York: John Wiley & Sons; 2002.pp. 11.10.1–11.10.23.
alzolari A, Oliviero I, Deaglio S, Mariani G, Biffoni M, Sposi NM, et al. Transferrinreceptor 2 is frequently expressed in human cancer cell lines. Blood Cells MolDis 2007;39(1):82–91.
alzolari A, Raggi C, Deaglio S, Sposi NM, Stafsnes M, Fecchi K, et al. TfR2 localizesin lipid raft domains and is released in exosomes to activate signal transductionalong the MAPK pathway. J Cell Sci 2006;119(Pt 21):4486–98.
han RY, Ponka P, Schulman HM. Transferrin-receptor-independent but iron-dependent proliferation of variant Chinese hamster ovary cells. Exp Cell Res1992;202(2):326–36.
hen J, Wang J, Meyers KR, Enns CA. Transferrin-directed internalization and cycling
of transferrin receptor 2. Traffic 2009;10(10):1488–501.leves AE, Cooper DN, Barondes SH, Kelly RB. A new pathway for protein export inSaccharomyces cerevisiae. J Cell Biol 1996;133(5):1017–26.
aidarov I, Santini F, Warren RA, Keen JH. Spatial control of coated-pit dynamics inliving cells. Nat Cell Biol 1999;1(1):1–7.
emistry & Cell Biology 44 (2012) 189– 199
Gombos I, Bacso Z, Detre C, Nagy H, Goda K, Andrasfalvy M, et al. Cholesterolsensitivity of detergent resistance: a rapid flow cytometric test for detectingconstitutive or induced raft association of membrane proteins. Cytometry A2004;61(2):117–26.
Graham RM, Chua AC, Herbison CE, Olynyk JK, Trinder D. Liver iron transport. WorldJ Gastroenterol 2007;13(35):4725–36.
Harding CV. Current Protocols in Immunology. New York: John Wiley & Sons; 1997.pp. 16.2.1–16.2.15.
Herbison CE, Thorstensen K, Chua AC, Graham RM, Leedman P, Olynyk JK, et al. Therole of transferrin receptor 1 and 2 in transferrin-bound iron uptake in humanhepatoma cells. Am J Physiol Cell Physiol 2009;297(6):C1567–75.
Higashimura Y, Nakajima Y, Yamaji R, Harada N, Shibasaki F, Nakano Y, et al. Up-regulation of glyceraldehyde-3-phosphate dehydrogenase gene expression byHIF-1 activity depending on Sp1 in hypoxic breast cancer cells. Arch BiochemBiophys 2011;509(1):1–8.
Ikuta K, Zak O, Aisen P. Recycling, degradation and sensitivity to the synergisticanion of transferrin in the receptor-independent route of iron uptake by humanhepatoma (HuH-7) cells. Int J Biochem Cell Biol 2004;36(2):340–52.
Ivanov AI. Methods in Molecular Biology, vol 440. Totowa: Humana Press; 2008. pp.15–33.
Jandl JH, Inman JK, Simmons RL, Allen DW. Transfer of iron from serum iron-bindingprotein to human reticulocytes. J Clin Invest 1959;38(1 (Part 1)):161–85.
Johnson LS, Presley JF, Park JC, McGraw TE. Slowed receptor trafficking in mutantCHO lines of the End1 and End2 complementation groups. J Cell Physiol1994;158(1):29–38.
Kawabata H, Germain RS, Vuong PT, Nakamaki T, Said JW, Koeffler HP. Transferrinreceptor 2-alpha supports cell growth both in iron-chelated cultured cells andin vivo. J Biol Chem 2000;275(22):16618–25.
Kawabata H, Yang R, Hirama T, Vuong PT, Kawano S, Gombart AF, et al. Molecularcloning of transferrin receptor 2. A new member of the transferrin receptor-likefamily. J Biol Chem 1999;274(30):20826–32.
Klausner RD, Van Renswoude J, Ashwell G, Kempf C, Schechter AN, Dean A,et al. Receptor-mediated endocytosis of transferrin in K562 cells. J Biol Chem1983;258(8):4715–24.
Levy JE, Jin O, Fujiwara Y, Kuo F, Andrews NC. Transferrin receptor is neces-sary for development of erythrocytes and the nervous system. Nat Genet1999;21(4):396–9.
Lok CN, Ponka P. Identification of a hypoxia response element in the transferrinreceptor gene. J Biol Chem 1999;274(34):24147–52.
Malide D, Cushman SW. Morphological effects of wortmannin on the endosomalsystem and GLUT4-containing compartments in rat adipose cells. J Cell Sci1997;110(Pt 22):2795–806.
Mattia E, Rao K, Shapiro DS, Sussman HH, Klausner RD. Biosynthetic regulation ofthe human transferrin receptor by desferrioxamine in K562 cells. J Biol Chem1984;259(5):2689–92.
McGraw TE, Greenfield L, Maxfield FR. Functional expression of the human trans-ferrin receptor cDNA in Chinese hamster ovary cells deficient in endogenoustransferrin receptor. J Cell Biol 1987;105(1):207–14.
McGraw TE, Subtil A. Current Protocols in Cell Biology. New York: John Wiley & Sons;1999. pp. 15.3.1–15.3.23.
Muro S, Wiewrodt R, Thomas A, Koniaris L, Albelda SM, Muzykantov VR, et al. A novelendocytic pathway induced by clustering endothelial ICAM-1 or PECAM-1. J CellSci 2003;116(Pt 8):1599–609.
Nombela C, Gil C, Chaffin WL. Non-conventional protein secretion in yeast. TrendsMicrobiol 2006;14(1):15–21.
Orlandi PA, Fishman PH. Filipin-dependent inhibition of cholera toxin: evidence fortoxin internalization and activation through caveolae-like domains. J Cell Biol1998;141(4):905–15.
Owen D, Kuhn LC. Noncoding 3′ sequences of the transferrin receptor gene arerequired for mRNA regulation by iron. EMBO J 1987;6(5):1287–93.
Pho MT, Ashok A, Atwood WJ. JC virus enters human glial cells by clathrin-dependentreceptor-mediated endocytosis. J Virol 2000;74(5):2288–92.
Raje CI, Kumar S, Harle A, Nanda JS, Raje M. The macrophage cell surfaceglyceraldehyde-3-phosphate dehydrogenase is a novel transferrin receptor. JBiol Chem 2007;282(5):3252–61.
Rao K, Harford JB, Rouault T, McClelland A, Ruddle FH, Klausner RD. Transcrip-tional regulation by iron of the gene for the transferrin receptor. Mol Cell Biol1986;6(1):236–40.
Rao KK, Shapiro D, Mattia E, Bridges K, Klausner R. Effects of alterations in cellu-lar iron on biosynthesis of the transferrin receptor in K562 cells. Mol Cell Biol1985;5(4):595–600.
Reid PA, Watts C. Constitutive endocytosis and recycling of major histocompatibilitycomplex class II glycoproteins in human B-lymphoblastoid cells. Immunology1992;77(4):539–42.
Richardson DR, Baker E. The uptake of iron and transferrin by the human malignantmelanoma cell. Biochim Biophys Acta 1990;1053(1):1–12.
Richardson DR, Baker E. Two saturable mechanisms of iron uptake from trans-ferrin in human melanoma cells: the effect of transferrin concentration,chelators, and metabolic probes on transferrin and iron uptake. J Cell Physiol1994;161(1):160–8.
Rudolph NS, Ohlsson-Wilhelm BM, Leary JF, Rowley PT. Regulation of K562 cell
transferrin receptors by exogenous iron. J Cell Physiol 1985;122(3):441–50.Sarkar J, Seshadri V, Tripoulas NA, Ketterer ME, Fox PL. Role of ceruloplasmin inmacrophage iron efflux during hypoxia. J Biol Chem 2003;278(45):44018–24.
Schutze S, Machleidt T, Adam D, Schwandner R, Wiegmann K, Kruse ML, et al.Inhibition of receptor internalization by monodansylcadaverine selectively

f Bioch
S
S
T
T
T
T
T
T
S. Kumar et al. / The International Journal o
blocks p55 tumor necrosis factor receptor death domain signaling. J Biol Chem1999;274(15):10203–12.
chwab RB, Okamoto T, Scherer PE, Lisanti MP. Analysis of the association of pro-tein with membrane. In: Cathleen S, Morgan, editors. Current Protocols in CellBiology, vol. 5. John Wiley & Sons; 2000. pp. 5.4.1–5.4.17.
eo J, Jeong J, Kim YM, Hwang N, Paek E, Lee KJ. Strategy for comprehensive iden-tification of post-translational modifications in cellular proteins, including lowabundant modifications: application to glyceraldehyde-3-phosphate dehydro-genase. J Proteome Res 2008;7(2):587–602.
acchini L, Gammella E, De Ponti C, Recalcati S, Cairo G. Role of HIF-1 and NF-kappaBtranscription factors in the modulation of transferrin receptor by inflammatoryand anti-inflammatory signals. J Biol Chem 2008;283(30):20674–86.
ao J, Liu YQ, Li Y, Peng JL, Li L, Liu J, et al. Hypoxia: dual effect on the expres-sion of transferrin receptor in human melanoma A375 cell line. Exp Dermatol2007;16(11):899–904.
esta U, Louache F, Titeux M, Thomopoulos P, Rochant H. The iron-chelatingagent picolinic acid enhances transferrin receptors expression in human ery-throleukaemic cell lines. Br J Haematol 1985;60(3):491–502.
horstensen K, Trinder D, Zak O, Aisen P. Uptake of iron from N-terminalhalf-transferrin by isolated rat hepatocytes. Evidence of transferrin-receptor-independent iron uptake. Eur J Biochem 1995;232(1):129–33.
isdale EJ, Kelly C, Artalejo CR. Glyceraldehyde-3-phosphate dehydrogenaseinteracts with Rab2 and plays an essential role in endoplasmic reticu-
lum to Golgi transport exclusive of its glycolytic activity. J Biol Chem2004;279(52):54046–52.rinder D, Morgan E. Inhibition of uptake of transferrin-bound iron byhuman hepatoma cells by nontransferrin-bound iron. Hepatology 1997;26(3):691–8.
emistry & Cell Biology 44 (2012) 189– 199 199
Trinder D, Morgan EH, Baker E. The effects of an antibody to the rat transferrinreceptor and of rat serum albumin on the uptake of diferric transferrin by rathepatocytes. Biochim Biophys Acta 1988;943(3):440–6.
Trinder D, Zak O, Aisen P. Transferrin receptor-independent uptake of differic trans-ferrin by human hepatoma cells with antisense inhibition of receptor expression.Hepatology 1996;23(6):1512–20.
Trowbridge IS, Collawn JF, Hopkins CR. Signal-dependent membrane protein traf-ficking in the endocytic pathway. Annu Rev Cell Biol 1993;9:129–61.
Tsunoo H, Sussman HH. Placental transferrin receptor. Evaluation of the presence ofendogenous ligand on specific binding. J Biol Chem 1983;258(7):4118–22.
Vinay DS, Raje M, Verma RK, Mishra GC. Characterization of novel costimulatorymolecules. A protein of 38–42 kDa from B cell surface is concerned with T cellactivation and differentiation. J Biol Chem 1995;270(40):23429–36.
Walseng E, Bakke O, Roche PA. Major histocompatibility complex class II-peptidecomplexes internalize using a clathrin- and dynamin-independent endocytosispathway. J Biol Chem 2008;283(21):14717–27.
West Jr AP, Bennett MJ, Sellers VM, Andrews NC, Enns CA, Bjorkman PJ. Com-parison of the interactions of transferrin receptor and transferrin receptor 2with transferrin and the hereditary hemochromatosis protein HFE. J Biol Chem2000;275(49):38135–8.
Xu S, Ariizumi K, Edelbaum D, Bergstresser PR, Takashima A. Cytokine-dependentregulation of growth and maturation in murine epidermal dendritic cell lines.Eur J Immunol 1995;25(4):1018–24.
Yamaji R, Chatani E, Harada N, Sugimoto K, Inui H, Nakano Y. Glyceraldehyde-3-phosphate dehydrogenase in the extracellular space inhibits cell spreading.Biochim Biophys Acta 2005;1726(3):261–71.
Zak O, Trinder D, Aisen P. Primary receptor-recognition site of human transferrin isin the C-terminal lobe. J Biol Chem 1994;269(10):7110–4.