cerebellar network plasticity
TRANSCRIPT

R
CO
Ga
vBb
gc
Fd
F
ArtbikymepttwoassanmcfsptL
TSK
C
*MN�EA[cl
Neuroscience 153 (2008) 1–19
0d
EVIEW
EREBELLAR NETWORK PLASTICITY: FROM GENES TO FAST
SCILLATIONG
F
I
P
C
P
CAR
Tia(idTaaaeitml(cpod(Choe
. CHERON,a,b* L. SERVAISb,d AND B. DANa,c
Laboratory of Neurophysiology and Movement Biomechanics, Uni-ersité Libre de Bruxelles, CP640, 50 Av F Roosevelt, Brussels,elgium
Laboratory of Electrophysiology, Université de Mons-Hainaut, Bel-ium
Department of Neurology, Hopital Universitaire des Enfants Reineabiola, Université Libre de Bruxelles, Belgium
Service de Neurologie Pédiatrique, Hôpital Robert Debré, Paris,rance
bstract—The role of the cerebellum has been increasinglyecognized not only in motor control but in sensory, cogni-ive and emotional learning and regulation. Purkinje cells,eing the sole output from the cerebellar cortex, occupy an
ntegrative position in this network. Plasticity at this level isnown to critically involve calcium signaling. In the last fewears, electrophysiological study of genetically engineeredice has demonstrated the topical role of several genes
ncoding calcium-binding proteins (calretinin, calbindin,arvalbumin). Specific inactivation of these genes results inhe emergence of a fast network oscillation (ca. 160 Hz)hroughout the cerebellar cortex in alert animals, associatedith ataxia. This oscillation is produced by synchronizationf Purkinje cells along the parallel fiber beam. It behavess an electrophysiological arrest rhythm, being blocked byensorimotor stimulation. Pharmacological manipulationshowed that the oscillation is blocked by GABAA and NMDAntagonists as well as gap junction blockers. This cerebellaretwork oscillation has also been documented in mouseodels of human conditions with complex developmental
erebellar dysfunction, such as Angelman syndrome andetal alcohol syndrome. Recent evidence suggests a relation-hip between fast oscillation and cerebellar long term de-ression (LTD). This may have major implications for futureherapeutic targeting. © 2008 IBRO. Published by Elseviertd. All rights reserved.
Contentshe cerebellar module circuitry 2imple and complex spikes dialogue 3nockout of Ca2� binding-proteins as a tool for studying
neural network excitability 4ontrasting in vivo and in vitro firing behavior of pcs in
Cr�/� mice 4
Correspondence to: G. Cheron Laboratory of Neurophysiology andovement Biomechanics, Université Libre de Bruxelles, CP640,.5.109 (5th floor) 808, Route de Lennik, 1070 Brussels, Belgium. Tel:32-26502477; fax: �32-65373566.-mail address: [email protected] (G. Cheron).bbreviations: CaMKII, Ca2�/calmodulin-dependent protein kinase II;
Ca2�]i, concentration of intracellular Ca2�; Cb, calbindin D-28 K; CBX,
parbenoxolone; Cr, calretinin; FAS, fetal alcohol syndrome; LFPO,ocal field potential oscillation; PC, Purkinje cell; Pv, parvalbumin.
306-4522/08$32.00�0.00 © 2008 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2008.01.074
1
ranule cells of Cr�/� mice as a model ofcalcium-dependent control of neuronal excitability 4
rom PC firing alteration to emergence of fast oscillation(160–200 Hz) in alert mutant mice 5
n vivo firing behavior of individual PCs in differentCa2�-binding proteins mutants 5
harmacological properties of the 160 Hz oscillation 7Involvement of GABAA inhibitory network to 160 Hz
oscillation 7Involvement of gap junctions in 160 Hz oscillation 7ausal links between oscillation, firing rate, rhythmicity and
synchronicity 8hysiological significance of fast cerebellar oscillation 10Restoration of normal cerebellar electrophysiology
and motor behavior by targeted Cr expression ingranule cells of Cr�/� mice 11
Fast cerebellar oscillation in Angelman syndrome 11Fast oscillation and learning deficit in fetal alcohol
syndrome 13onclusions 14cknowledgments 14eferences 14
he cerebellum has always exerted a fascinating mysteryn neuroscience. There is now growing evidence that inddition to its well-documented role in motor coordinationThach et al., 1992; Bastian, 2006), the cerebellum isnvolved in sensory (Gao et al., 1996), cognitive (An-reasen et al., 1998; Booth et al., 2007; Gordon, 2007;hach, 2007) and emotional processing (Schmahmannnd Sherman, 1998; Schmahmann, 2004; Schmahmannnd Caplan, 2006). The timing control has been proposeds a common denominator of these multiple functions (Ivryt al., 2002; Spencer et al., 2003). However, recent exper-
ments that attempted to dissociate timing from coordina-ion control suggested that time-dependent control ofovements did not produce robust activity in the cerebel-
um with respect to state-dependent control conditionDiedrichsen et al., 2007). Nevertheless, precise control ofoordination signals in cerebellum critically requires tem-oral processing on the time scale of milliseconds to sec-nds and must also integrate the dynamic synchrony ofifferent neuronal populations and their interactionsWelsh et al., 1995; Kazantsev et al., 2003; Sanchez-ampusano et al., 2007). In this context, oscillatory activityas beeen shown to be a fundamental emergent propertyf neural networks (Destexhe and Contreras, 2006; Bartost al., 2007). As demonstrated previously in the hippocam-
us the recording of local field potential oscillation (LFPO)ved.

io
ibCKD2mctpoec
scfl[ii1It1sttpttcct
ciastpdh
Tpnntsvbt
m
d(Rfr1s
fsMacdH
lcafif1
Fm�agicGPu
G. Cheron et al. / Neuroscience 153 (2008) 1–192
n the cerebellum may contribute to a better understandingf its function.
Since the initial proposal of Ito (1972, 1982) the phys-ological role of the cerebellum in behavioral learning hasecome a central issue (Miles and Lisberger, 1981;heron, 1990; Raymond et al., 1996; Lisberger, 1998;ahlon and Lisberger, 2000; Carey and Lisberger, 2004;elgado-Garcia and Gruart, 2002, 2006; Medina et al.,002; De Zeeuw and Yeo, 2005). It has been studied atultiple levels, including behavioral, electrophysiological,
ellular and molecular aspects. The proper functioning ofhe cerebellum requires complex genetic and epigeneticrograms that ensure structural and functional propertiesf the neural network. Input–output interactions with thenvironment refine the synaptic weights of the neural cir-uitry throughout life.
These multiple plasticity processes are regulated at theingle cell level by changes in the concentration of intra-ellular Ca2� ([Ca2�]i) and are also dependent on Ca2�
ux shaping (Hansel et al., 2006; Coesmans et al., 2004).Ca2�]i regulates a large variety of neuronal functions,ncluding neurotransmitter release, ion channel permeabil-ty, enzyme activity, and gene transcription (Berridge,998). The control of [Ca2�]i is thus crucial for the neuron.t is dependent on pumps, exchangers, organelle seques-ration, and cytoplasmic Ca2�-binding proteins (Clapham,995). Among the different functions of Ca2� neuronalignaling (Usowicz et al., 1992; Llinás and Moreno, 1998)he control of the spontaneous firing of neuron and theransition from tonic spiking to bursting occupy a centralosition. Therefore, understanding the functional link be-ween Ca2� homeostasis at the single neuronal level andhe cerebellar neural network dynamics will not only in-rease our understanding of the basic mechanisms impli-ated in the network control but possibly also highlightherapeutic targets in neurological diseases.
This review will outline current understanding of theerebellar network based on the conjunction of in vivo andn vitro investigations in genetically engineered mice. Thispproach was initially developed in order to better under-tand the role played by the different Ca2�-binding pro-eins in Ca2� signaling. Its expanding use has revealedeculiar abnormalities in a number of models of humaniseases, such as Angelman syndrome and the fetal alco-ol syndrome.
THE CEREBELLAR MODULE CIRCUITRY
he cerebellar cortex appears particularly well suited forerforming in vivo/in vitro correlation in genetically engi-eered mice. It is formed by relatively simple array ofeural network structured around the Purkinje cell (PC),he sole output of the cerebellar cortex forming inhibitoryynapses with their target neurons in the cerebellar andestibular nuclei. Only two major afferent systems formedy the climbing and the mossy fibers determine the input ofhe cerebellar cortex (Fig. 1).
Climbing fibers arise from the inferior olive. They pri-
arily contact the more proximal portion of the PC den- iritic tree and provide the major excitatory action on PC1500 climbing fiber-PC synaptic contacts, Strata andossi, 1998). The mature PC receives this excitatory input
rom only one inferior olive neuron, giving rise to the wellecognizable complex spike (Eccles et al., 1966c; Thach,968) and the induced pause in the spontaneous simplepike firing of the same PC.
Transmission by mossy fibers conveying informationrom numerous regions of the cerebrum, brain stem andpinal cord, is more complex than that of climbing fibers.ossy fibers first excite the numerous granule cells whosexons give rise to the parallel fibers, which form synapticontacts at the spines of the distal branch of the PCendritic tree (150,000 parallel fiber–PC synaptic contacts,arvey and Napper, 1991).
Parallel fibers also excite interneurons in the molecularayer which in turn inhibit PCs (feedforward inhibitory cir-uit). Mossy fiber inputs are modulated by the inhibitoryction of Golgi cells, which are excited by both mossybers and parallel fibers, assuming a feedforward and aeedback inhibitory loop, respectively (Eccles et al.,966b). The Golgi cell is also excited by climbing fiber
++
++
+
+
++
+
++ +
+
+
-
-
-
-
--
-
-
--
--
--
DCN
PN
pf
Pc
gc
ba
st
mf
cf
1
2
GoC
R
C
L
R
mol L
Pc L
gr L
CrPvPv-Cb
Lc
ubc
IO
3
4
ig. 1. Scheme illustrating the neuronal elements of the cerebellarodule circuitry and the expression of different Ca2� binding proteins.and � signs indicate the excitatory or inhibitory action of the syn-
pse, respectively. The resistance symbol indicates the presence ofap junctions. Arrows indicate (1) climbing fiber input, (2) mossy fiber
nput, (3) PC output, (4) deep nuclear output. Abbreviations: ba, basketell; cf, climbing fiber; DCN, deep cerebellar nuclei; gc, granule cell;oC, Golgi cell; Lc, Lugaro cell; IO inferior olive; mf, mossy fiber; PC,urkinje cell; pf, parallel fiber; PN, pontine nuclei; st, stellate cell; ubc,nipolar brush cell.
nput (Hamori and Szentagothai, 1966; Eccles et al.,
1ubsLc(snf(c
bo1t(fc
cst12icp2mIcvlgs
Soaislhtfitssmaapdsi
cSfivLa2miasd
icdsTfit
Fot(da2
G. Cheron et al. / Neuroscience 153 (2008) 1–19 3
966a,d; Desclin, 1976) which indirectly modulates gran-le cell discharge. Two additional types of neurons muste taken into account in the treatment of the mossy fiberignals: the Lugaro cells and the unipolar brush cells. Theugaro cell provides synchronous inhibition of the Golgiell along a transverse axis parallel to the parallel fiberDieudonné and Dumoulin, 2000). The unipolar brush cellspecifically expressed in the vestibulo-cerebellum (Mug-aini and Floris, 1994) are excited by mossy fibers andorm giant glutamatergic synapses with granule cellsRossi et al., 1995). Like granule cells, the unipolar brushells are also inhibited by Golgi cells (Dugué et al., 2005).
Morphologically, the parallel fibers form longitudinaleams running for several millimeters along the main axisf the folium (Eccles et al., 1967; Cohen and Yarom,998). The parallel fibers beams are almost orthogonal tohe parasagittal organization of the cerebellar moduleVoogd and Glickstein, 1998), which is subdivided in dif-erent microzones considered as the operational unit of theerebellum (Oscarsson, 1979; Schonewille et al., 2006).
The unique origin of the complex spike firing controlontrasts with the multiple factors implicated in simplepike discharge. The latter results from the combination ofhe pacemaker activity of the PC (Häusser and Clark,997; Raman and Bean, 1997, 1999; Häusser et al.,004), their direct excitation from the parallel fibers, the
nhibition exerted by the basket, stellate, Golgi and Lugaroells (Suter and Jaeger, 2004) and finally the modulationerformed by the climbing fiber influence (McKay et al.,007; Mittmann and Häusser, 2007). This latter influenceay occur as long-term potentiation of spontaneous
PSCs (Kano et al., 1992, 1996) and participates in theomplex control of the feedforward inhibitory circuit acti-ated by the parallel fibers (Mittmann et al., 2005). Stimu-
ation of individual granule cells shows that at least 50ranule cells must fire simultaneously to produce a singleimple spike (Barbour, 1993).
SIMPLE AND COMPLEX SPIKES DIALOGUE
pontaneous climbing fibers occur at an average firing ratef �1 Hz (Thach, 1968; Lang et al., 1999). The functionaldvantage of this low frequency firing has been illustrated
n GluR�2 knockout mice (Yoshida et al., 2007) with re-pect to the timing of the optokinetic response. Short- andong-term climbing fiber regulation of simple spike firingas been described. For example, after chemical destruc-ion of the inferior olive the PC doubled their simple spikering rate, the spike rate became more regular and linkedo oscillation of the background (Colin et al., 1980). In thehort time range the antiphasic behavior of the complexpike and simple spike firing during vestibular stimulationay be explained by climbing fiber action on the Golgi cellcting as an inhibitory gate on mossy fiber input (Barmacknd Yakhnitsa, 2003). Simple spike firing has been re-orted to partially predict the occurrence of complex spikeuring tonic mode periods (Miall et al., 1998). Complexpike waveform reflects the compound excitation in prox-
mal PC dendrite by the climbing fiber and is classically ponsidered as invariant in vivo for a given PC (Ito andimpson, 1971). It remains controversial whether mossybers may cause specific modifications in this waveform inivo (Campbell and Hesslow, 1986a,b; Gilbert, 1976).ong- and short-term plasticity has been described in vitrot the climbing fiber–PC synapse (Hansel and Linden,000; Hashimoto and Kano, 1998). They cause specificodifications in the complex spike waveform recorded by
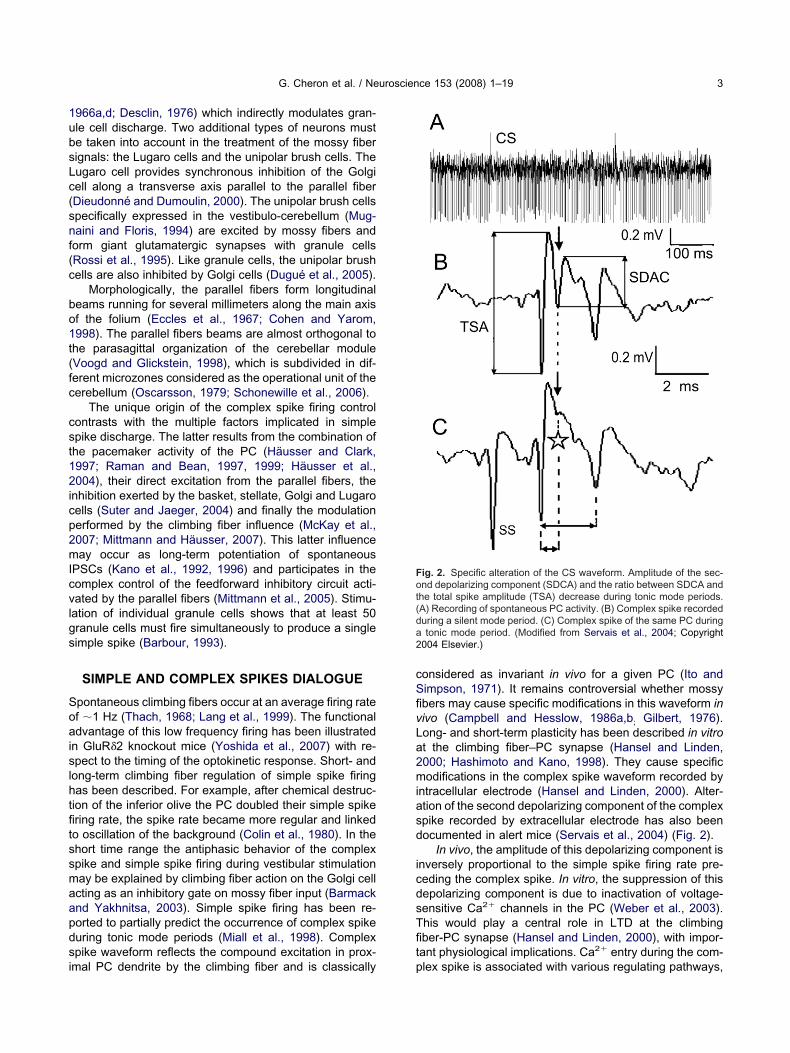
ntracellular electrode (Hansel and Linden, 2000). Alter-tion of the second depolarizing component of the complexpike recorded by extracellular electrode has also beenocumented in alert mice (Servais et al., 2004) (Fig. 2).
In vivo, the amplitude of this depolarizing component isnversely proportional to the simple spike firing rate pre-eding the complex spike. In vitro, the suppression of thisepolarizing component is due to inactivation of voltage-ensitive Ca2� channels in the PC (Weber et al., 2003).his would play a central role in LTD at the climbingber-PC synapse (Hansel and Linden, 2000), with impor-ant physiological implications. Ca2� entry during the com-
ig. 2. Specific alteration of the CS waveform. Amplitude of the sec-nd depolarizing component (SDCA) and the ratio between SDCA and
he total spike amplitude (TSA) decrease during tonic mode periods.A) Recording of spontaneous PC activity. (B) Complex spike recordeduring a silent mode period. (C) Complex spike of the same PC duringtonic mode period. (Modified from Servais et al., 2004; Copyright
004 Elsevier.)
lex spike is associated with various regulating pathways,
s1Av(mdtcreu
Ot1eeslt
bwnefpotTccmfPmCwi(iaC(p
Ttp1t
(qa(
s1uiaptCa
Bt1iecaa
FivfetotoSU
G. Cheron et al. / Neuroscience 153 (2008) 1–194
uch as upregulation of GABAA receptor (Kano et al.,992) and LTD at parallel fiber–PC synapse (Ito, 1989).lthough some authors have suggested that in vitro and inivo recordings of the complex spike are comparableSchmoleski et al., 2002), the association of in vivo decre-ent of complex spike secondary depolarizing peak and aecrease in Ca2� entry during the complex spike remains
o be demonstrated. If this association, described in vitro isonfirmed in vivo, it will indicate that parallel fiber pathwaysealize a feedforward control of its firing through Ca2�
ntry modulation, which would be of crucial importance innderstanding cerebellar function.
KNOCKOUT OF Ca2� BINDING-PROTEINS AS ATOOL FOR STUDYING
NEURAL NETWORK EXCITABILITY
ne advantage of using Ca-binding gene knockouts is thathey preserve cerebellar morphology. (Airaksinen et al.,997; Schiffmann et al., 1999; Cheron et al., 2000). Differ-nt types of cerebellar neurons specifically express differ-nt Ca2�-binding proteins. This offers the possibility ofelectively altering Ca2� homeostasis and thereby modu-
ate excitability of selected neuronal types in order to studyheir effect on the whole network.
Calretinin (Cr) is present in Golgi, Lugaro and unipolarrush cells. Calbindin D-28 K (Cb) is present only in PC,hile parvalbumin (Pv) is present in both molecular inter-eurons and PC (Fig. 1). However, in addition to cellxpression specificity, these Ca2�-binding proteins are dif-erently specialized, allowing subtle physiological pur-oses (Schwaller et al., 2002). Cr, Cb and Pv are membersf the EF-hand family of Ca2�-binding proteins. They con-
ain five, four and three EF-hand domains, respectively.he metal-binding sites of the Cr and Cb are Ca2�-spe-ific, whereas those of Pv bind to Mg2� in the restingondition. These binding characteristics explain one of theajor differences in Ca2�-buffering kinetics: Cr and Cb are
ast buffers, while Pv is relatively slow. Moreover, Cr andv are considered pure Ca2� buffers, acting as passiveodulators of the cytosolic calcium levels lacking anya2� sensor or trigger function (Schmidt et al., 2007),hile Cb acts as a sensor that regulates the degradation of
nositol messengers in an activity-dependent mannerSchmidt et al., 2005). As high frequency synaptic activitys often associated with IP3-mediated Ca2� transients, thectivation of myo-inositol monophosphatase (IMPase) byb is strategically situated for the synaptic integration
Finch and Augustine, 1998; Takechi et al., 1998) and PClasticity (Wang et al., 2000).
CONTRASTING IN VIVO AND IN VITRO FIRINGBEHAVIOR OF PCS IN Cr�/� MICE
he initial step in this series of investigations concernedhe role played by Cr, which is the only Ca2�-bindingrotein expressed in granule cells (Résibois and Rogers992; Marini et al., 1997), the most numerous neurons in
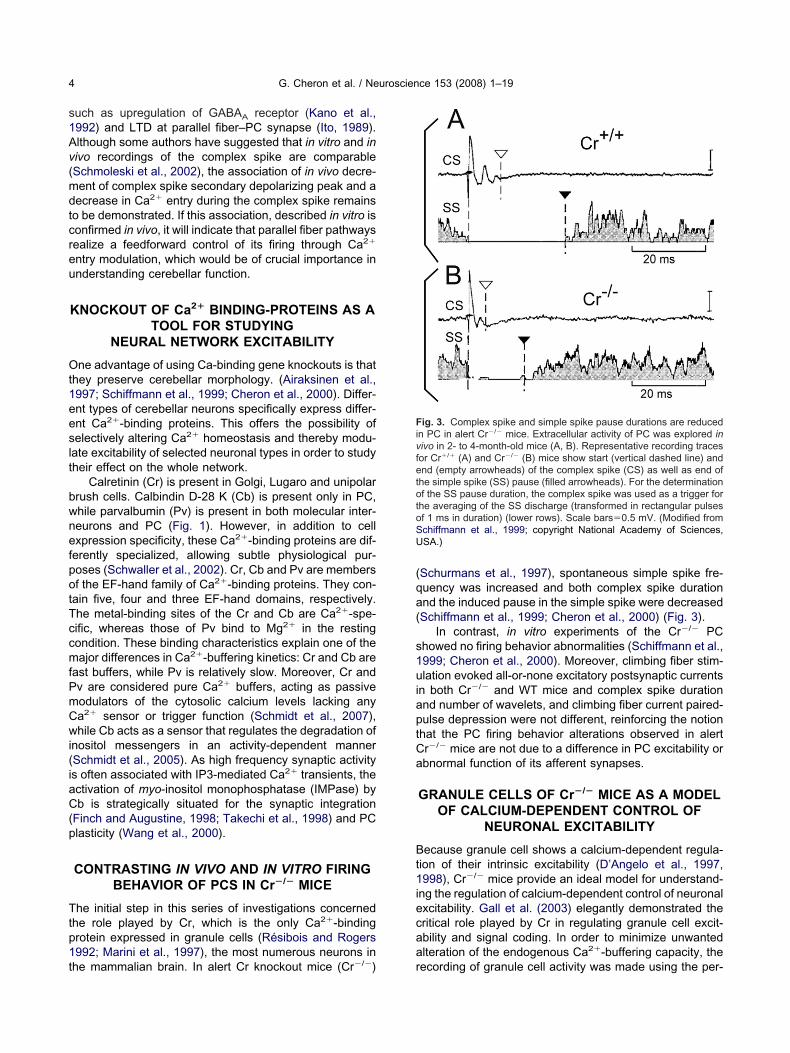
he mammalian brain. In alert Cr knockout mice (Cr�/�) rSchurmans et al., 1997), spontaneous simple spike fre-uency was increased and both complex spike durationnd the induced pause in the simple spike were decreasedSchiffmann et al., 1999; Cheron et al., 2000) (Fig. 3).
In contrast, in vitro experiments of the Cr�/� PChowed no firing behavior abnormalities (Schiffmann et al.,999; Cheron et al., 2000). Moreover, climbing fiber stim-lation evoked all-or-none excitatory postsynaptic currents
n both Cr�/� and WT mice and complex spike durationnd number of wavelets, and climbing fiber current paired-ulse depression were not different, reinforcing the notionhat the PC firing behavior alterations observed in alertr�/� mice are not due to a difference in PC excitability orbnormal function of its afferent synapses.
GRANULE CELLS OF Cr�/� MICE AS A MODELOF CALCIUM-DEPENDENT CONTROL OF
NEURONAL EXCITABILITY
ecause granule cell shows a calcium-dependent regula-ion of their intrinsic excitability (D’Angelo et al., 1997,998), Cr�/� mice provide an ideal model for understand-
ng the regulation of calcium-dependent control of neuronalxcitability. Gall et al. (2003) elegantly demonstrated theritical role played by Cr in regulating granule cell excit-bility and signal coding. In order to minimize unwantedlteration of the endogenous Ca2�-buffering capacity, the
ig. 3. Complex spike and simple spike pause durations are reducedn PC in alert Cr�/� mice. Extracellular activity of PC was explored inivo in 2- to 4-month-old mice (A, B). Representative recording tracesor Cr�/� (A) and Cr�/� (B) mice show start (vertical dashed line) andnd (empty arrowheads) of the complex spike (CS) as well as end of
he simple spike (SS) pause (filled arrowheads). For the determinationf the SS pause duration, the complex spike was used as a trigger for
he averaging of the SS discharge (transformed in rectangular pulsesf 1 ms in duration) (lower rows). Scale bars�0.5 mV. (Modified fromchiffmann et al., 1999; copyright National Academy of Sciences,SA.)
ecording of granule cell activity was made using the per-

ftrccTC
iittmsecc2
Tfprow
me((Pe
toaup1w(msoob(
traapt
to
wpr
fctstaq
Teuer2
FtL(rn5tcwotaaSts
G. Cheron et al. / Neuroscience 153 (2008) 1–19 5
orated patch technique. They found no morphological al-erations in Cr�/� mice and no changes in membraneesistance or capacitance. They also demonstrated in-reased granule cell excitability accompanied by a de-rease in the action potential half-width in the Cr�/� mice.hese effects were reversed by perfusion of an exogenousa2� buffer.
Based on their findings, the authors suggested a min-mal model that allows a qualitative understanding of thempact of variations in cytosolic Ca2�-buffering capacity onhe electroresponsiveness of an excitable cell. Accordingo this model, increased granule cell excitability in Cr�/�
ice can be mimicked by a fourfold decrease in the cyto-olic Ca2�-buffering capacity. The action potential short-ning reflects a more pronounced activation of IK-Ca be-ause of faster Ca2� dynamics taking place when theytosolic Ca2�-buffering capacity is decreased (Gall et al.,005).
FROM PC FIRING ALTERATION TOEMERGENCE OF FAST OSCILLATION(160–200 Hz) IN ALERT MUTANT MICE
he spike-generating mechanism of PC in vivo resultsrom a complex interaction between intrinsic and networkroperties. The analysis of the spontaneous firing rate,hythmicity and synchronicity constitutes an efficient wayf representing different functional states in a neural net-ork (Fellous et al., 2003; Nelson et al., 2003).
As discussed above, Cr in granule cells induces re-ote changes on the PC firing rate suggesting a networkffect. This is also reflected in cerebellar network activityidentified neurons and local field potentials) in both singleCr�/�, Cb�/�, Pv�/�) and double (Cr�/�-Cb�/�, Cb�/�-v�/�) knockout mice (Cheron et al., 2004a,b; Servaist al., 2005).
The most remarkable electrophysiological feature inhese Ca2�-binding protein-deficient mice is the presencef a spontaneous oscillation around 160–200 Hz (Fig. 4And B). This oscillatory activity can be classified in theltrafast (90–200 Hz) range of physiological oscillationreviously described in the hippocampus (Buzsáki et al.,992; Buzsáki and Draguhn, 2004.) Ultrafast oscillationas recorded throughout the explored cerebellar regions
vermis, uvula and nodulus) of single and double knockoutice for each of these genes. It appeared as episodes of
pindle-shaped oscillation with a mean rate of occurrencef 4.5 spindles/s. It is noteworthy that network 160 Hzscillation also emerged in slices prepared from the cere-ellum of mature rats when bathed in zero-calcium mediaTraub et al., 2007).
Spatiotemporal mapping of the oscillation was facili-ated by the cerebellar architecture characterized by highlyepetitive, well-defined circuitry (Eccles et al., 1967; Voogdnd Glickstein, 1998). In all mutant mice, LFPO amplitudelways increased when the recording electrode ap-roached the PC layer, reaching its maximum just beneath
he PC bodies (Fig. 4C) (Cheron et al., 2004), suggesting mhat PC populations are the major generator of the 160 Hzscillation.
Spatial coherence analysis demonstrated that LFPOsere perfectly synchronized at sites along the coronallane (i.e. along a parallel fiber beam) and not alongostro-caudal axis (Fig. 4B).
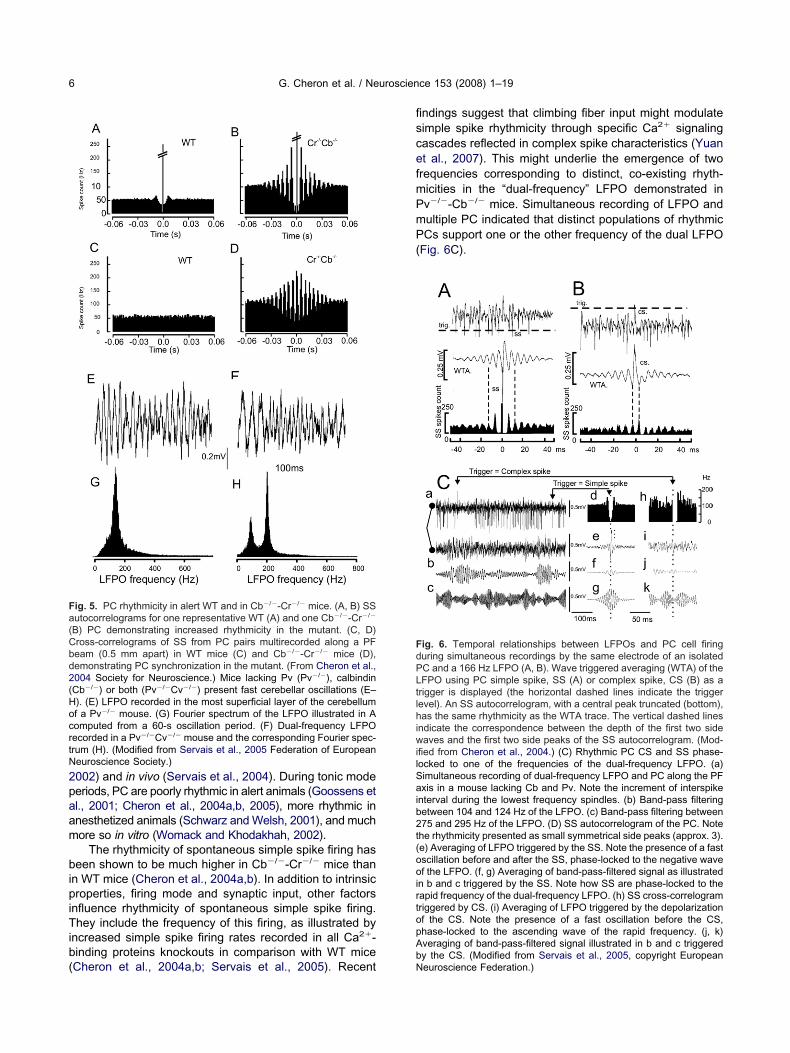
The frequency of oscillation showed no significant dif-erences between the different single knockout mice. Inontrast to mice deficient for either one of the genes andhe double Cr�/�-Cb�/�, LFPO in Pv�/�Cb�/� micehowed two frequency peaks during approximately 50% ofhe oscillation recording time (Servais et al., 2005) (Fig. 5Fnd H). The difference between the two oscillation fre-uencies was always larger than 100 Hz.
IN VIVO FIRING BEHAVIOR OF INDIVIDUALPCs IN DIFFERENT Ca2�-BINDING PROTEINS
MUTANTS
o understand how ultrafast oscillations contribute to cer-bellum functions, it is essential to first consider the basicnderlying mechanisms. Two key requirements for themergence of network oscillation are the presence of neu-onal rhythmicity and synchronicity (Servais and Cheron,005). PCs alternate between tonic, silent and bursting
ig. 4. Emergence and spatial coherence of high-frequency oscilla-ion in the cerebellum of Cr�/�-Cb�/�mice. (A) Sample records ofFPOs from wild-type mice (wt; top trace) and Cr�/�-Cb�/� micebottom trace). (B) Spatial coherence of LFPOs was analyzed byecording simultaneously from electrodes aligned along the longitudi-al [left, tracts 2, 1, 3 (0.5 mm apart)] or rostrocaudal (right, tracts 4, 1,) axis of a folium. Pair traces (left), using LFPO recording two as a
rigger (Trig) for a wave-trigger averaging (WTA), show coherent os-illations of the same period and without any significant phase delay,hereas for pair traces (right), using channel one as a trigger, noscillatory pattern was visible. (C) Depth profile (Ca-Cf) of WTA traces
hroughout the last PC layer of lobule 10 indicates that the maximummplitude was found just beneath the PC bodies. The arrow indicatesn electrolytic lesion at the recording depth of the WTA trace (Cb).cale bar�0.2 mm. The dashed line represents the recording elec-
rode tract. (From Cheron et al., 2004 Copyright Society for Neuro-cience.)
odes of firing both in vitro (Womack and Khodakhah,
2paam
bipiTib(
fiscefmPmP(
FdPLtlhiwilSaib2t(ooirtopA
Fa(Cbd2(HocrtN
G. Cheron et al. / Neuroscience 153 (2008) 1–196
002) and in vivo (Servais et al., 2004). During tonic modeeriods, PC are poorly rhythmic in alert animals (Goossens etl., 2001; Cheron et al., 2004a,b, 2005), more rhythmic innesthetized animals (Schwarz and Welsh, 2001), and muchore so in vitro (Womack and Khodakhah, 2002).
The rhythmicity of spontaneous simple spike firing haseen shown to be much higher in Cb�/�-Cr�/� mice than
n WT mice (Cheron et al., 2004a,b). In addition to intrinsicroperties, firing mode and synaptic input, other factors
nfluence rhythmicity of spontaneous simple spike firing.hey include the frequency of this firing, as illustrated by
ncreased simple spike firing rates recorded in all Ca2�-inding proteins knockouts in comparison with WT mice
ig. 5. PC rhythmicity in alert WT and in Cb�/�-Cr�/� mice. (A, B) SSutocorrelograms for one representative WT (A) and one Cb�/�-Cr�/�
B) PC demonstrating increased rhythmicity in the mutant. (C, D)ross-correlograms of SS from PC pairs multirecorded along a PFeam (0.5 mm apart) in WT mice (C) and Cb�/�-Cr�/� mice (D),emonstrating PC synchronization in the mutant. (From Cheron et al.,004 Society for Neuroscience.) Mice lacking Pv (Pv�/�), calbindinCb�/�) or both (Pv�/�Cv�/�) present fast cerebellar oscillations (E–). (E) LFPO recorded in the most superficial layer of the cerebellumf a Pv�/� mouse. (G) Fourier spectrum of the LFPO illustrated in Aomputed from a 60-s oscillation period. (F) Dual-frequency LFPOecorded in a Pv�/�Cv�/� mouse and the corresponding Fourier spec-rum (H). (Modified from Servais et al., 2005 Federation of Europeaneuroscience Society.)
Cheron et al., 2004a,b; Servais et al., 2005). RecentbN
ndings suggest that climbing fiber input might modulateimple spike rhythmicity through specific Ca2� signalingascades reflected in complex spike characteristics (Yuant al., 2007). This might underlie the emergence of tworequencies corresponding to distinct, co-existing rhyth-icities in the “dual-frequency” LFPO demonstrated inv�/�-Cb�/� mice. Simultaneous recording of LFPO andultiple PC indicated that distinct populations of rhythmicCs support one or the other frequency of the dual LFPO
Fig. 6C).
ig. 6. Temporal relationships between LFPOs and PC cell firinguring simultaneous recordings by the same electrode of an isolatedC and a 166 Hz LFPO (A, B). Wave triggered averaging (WTA) of theFPO using PC simple spike, SS (A) or complex spike, CS (B) as a
rigger is displayed (the horizontal dashed lines indicate the triggerevel). An SS autocorrelogram, with a central peak truncated (bottom),as the same rhythmicity as the WTA trace. The vertical dashed lines
ndicate the correspondence between the depth of the first two sideaves and the first two side peaks of the SS autocorrelogram. (Mod-
fied from Cheron et al., 2004.) (C) Rhythmic PC CS and SS phase-ocked to one of the frequencies of the dual-frequency LFPO. (a)imultaneous recording of dual-frequency LFPO and PC along the PFxis in a mouse lacking Cb and Pv. Note the increment of interspike
nterval during the lowest frequency spindles. (b) Band-pass filteringetween 104 and 124 Hz of the LFPO. (c) Band-pass filtering between75 and 295 Hz of the LFPO. (D) SS autocorrelogram of the PC. Note
he rhythmicity presented as small symmetrical side peaks (approx. 3).e) Averaging of LFPO triggered by the SS. Note the presence of a fastscillation before and after the SS, phase-locked to the negative wavef the LFPO. (f, g) Averaging of band-pass-filtered signal as illustrated
n b and c triggered by the SS. Note how SS are phase-locked to theapid frequency of the dual-frequency LFPO. (h) SS cross-correlogramriggered by CS. (i) Averaging of LFPO triggered by the depolarizationf the CS. Note the presence of a fast oscillation before the CS,hase-locked to the ascending wave of the rapid frequency. (j, k)veraging of band-pass-filtered signal illustrated in b and c triggered
y the CS. (Modified from Servais et al., 2005, copyright Europeaneuroscience Federation.)
ptcEz2d2(si�
fIta(tdnaaaa2
I1
AnGkoeblnc(amp
pcg12Hcth2fc
(cw
picific2tenaotw
nicaaD
I
AnLebjvopCagpctW
tlManMb2tt(je1
G. Cheron et al. / Neuroscience 153 (2008) 1–19 7
The synchronization of simple spike firing along thearallel fiber beam echoes Braitenberg’s original proposal
hat parallel fiber line transmission ensures timing andoordination control of movement (Braitenberg, 1967;ccles et al., 1967). In WT animals, on beam-synchroni-ation does not occur spontaneously (Goossens et al.,001; Cheron et al., 2004a,b) but emerges without anyelay when animals perform reaching tasks (Heck et al.,007). The absence of delay between distant PC�600 �m) suggests the involvement of highly efficientynchronization mechanisms as discussed below. Fig. 5llustrates cross-correlograms between PC pairs in Cb�/
Cr�/� mice showing remarkable synchrony of the high-requency simple spike firing along the parallel fiber beam.n contrast, cross-correlograms were flat in WT mice. Theemporal relationship between the LFPO and simple spikend complex spike firing was shown to be phase-lockedFig. 6). Simple spike discharges occurred at the depth ofhe LFPO, whereas the complex spike discharge appeareduring the LFPO ascending phase. In contrast, there waso phase locking between LFPO and Golgi cell spikes,lthough increased Golgi cell firing rate was correlated withsignificant reduction of concomitant LFPOs (Cheron et
l., 2004a). Simple spike firing phase-locking to LFPO haslso been demonstrated in cerebellar slices (Traub et al.,007).
PHARMACOLOGICAL PROPERTIES OF THE160 Hz OSCILLATION
nvolvement of GABAA inhibitory network to60 Hz oscillation
major difference between cerebral and cerebellar cortexeurotransmission resides in the preponderant role ofABAergic inhibition exerted by PCs considered as theey element of the cerebellar cortex. This implies that theutput from the cerebellar cortex (i.e. GABAergic PC) ex-rts inhibition not only on target neurons in the deep nuclei,ut also on the GABAergic interneurons of the molecular
ayer. In addition, like in the cerebral cortex, these inter-eurons are organized in a network of mutually inhibitoryonnections through fast GABAA receptor synapsesKondo and Marty, 1998; Carter and Regehr, 2002; Puia etl., 1994). A similarly organized network is known to pro-ote the emergence of gamma oscillation in the hip-ocampus (Whittington et al., 1995).
In all knockout mice for genes encoding Ca2�-bindingroteins in which 160 Hz oscillations were recorded, mi-roinjection of GABAA antagonist, such as bicuculline orabazine (SR95531), reversibly reduced the power of the60 Hz oscillation (Cheron et al., 2004a; Servais et al.,005) indicating the implication of inhibitory synapses.owever, this does not demonstrate that the 160 Hz os-illation is directly attributable to a phasic action of inhibi-ory synaptic currents, as bicuculline and gabazine couldave non-phasic actions, perhaps on axons (Traub et al.,003a). These authors demonstrated that GABA can elicitast oscillations when it is puffed on to hippocampal tissue
ontaining axons, and this effect is blocked by bicuculline (Traub et al., 2003a). It is possible that GABA may underertain conditions exert a tonic action in the cerebellum asell.
The highly stereotyped arrangement of closely ap-osed PCs and their orientation as well as their physiolog-
cal functioning as an open field make PC assemblies idealandidates for generating coherent LFPs when their activ-
ty is enhanced and synchronized. In addition, parallelbers make powerful synapses on stellate cells. This latterells acted as “coincident detectors” (Carter and Regehr,002) which may facilitate synchronization of the oscilla-ion along the folial axis. The finding that inhibitory andxcitatory GABA synapses coexist in the cerebellar inter-euron network (Chavas and Marty, 2003) may provide andditional mechanism whereby PC collaterals projectingn molecular interneurons would trigger a rhythmic inhibi-ion-excitation sequence in the coupled interneuron net-ork.
Finally, because the 160 Hz oscillations result frometwork activity, other types of neurons might also be
nvolved, such as basket cells, whose axons are electri-ally interconnected in the “pinceau” ensheathing the initialxon segment of PCs, or Lugaro cells, whose myelinatedxons can span 2 mm along a folium (Dieudonné andumoulin, 2000).
nvolvement of gap junctions in 160 Hz oscillation
s the inhibitory molecular interneurons are densely con-ected through dendrodendritic gap junctions (Sotelo andlinás, 1972; Mann-Metzer and Yarom, 1999), we hypoth-sized that this nonsynaptic mode of transmission mighte involved in the LFPO. We tested the effect of microin-
ection of gap-junction blocker carbenoxolone (CBX) inivo (Cheron et al., 2004a,b, 2005a,b), given the findingsf 200 Hz oscillation blockade with CBX in the hippocam-us in vitro (Draguhn et al., 1998). The blocking effect ofBX on the 160 Hz oscillation was potent, suggesting thatgap junction–coupled network could support the emer-
ence of this fast oscillation, as demonstrated in the hip-ocampus (Draguhn et al., 1998; Traub et al., 1999). Re-ently, Traub et al. (2007) reported that CBX also blockshe 160 Hz oscillation recorded in slices of cerebellum of
T rats bathed in zero-calcium media.There are at least two possible sites where gap junc-
ions would be relevant to the 160 Hz oscillation: at theevel of stellate and basket cells (Sotelo and Llinás, 1972;
ann-Metzer and Yarom, 1999) and at the level of thexonal plexus of PC collaterals. Several subtypes of con-exins, namely connexin 36 (Teubner et al., 2000; but seeeier et al., 2002), connexin 43 (Simburger et al., 1997;ut see Rash et al., 2005) and connexin 47 (Teubner et al.,001), are expressed in different neuronal populations inhe cerebellum, including molecular interneurons and PCs,hough expression of connexins in PCs has been disputedOdermatt et al., 2003). Recently, a new class of gap-unction proteins, namely pannexins, has been found to bexpressed in cerebellum (Bruzzone et al., 2003). Pannexinand pannexin 2 are expressed in PC and some cells
Golgi and Lugaro) in the granule cell layer. Pannexin 2 is

aia2P2ete
lidh2uPTdiboisIo(crabi1cjai
ttfiTacts2tote
DLb
crfswefsfsrlmLltc
tdrGCFsLpmsLfnNrcafiamlaartwcecoenfkPaeaa
G. Cheron et al. / Neuroscience 153 (2008) 1–198
lso expressed in stellate cells (Ray et al., 2006). Interest-ngly, pannexins exhibit a remarkable sensitivity to block-de by CBX, exceeding that of connexins (Bruzzone et al.,005). Moreover, important enzymes such as PKC, PKA,KG and CKII are able to modify pannexins (Barbe et al.,006). As pannexins are expressed in the majority of cer-bellar neurons, they appear as plausible candidates forhe coordination of gap-junction communication in the cer-bellum (Litvin et al., 2006).
The blocking action of CBX could switch off the oscil-ating circuit between molecular interneurons by decreas-ng the synchronicity inside disinhibition loops or throughirect action on axoaxonal junctions, as suggested in theippocampus (Traub et al., 2003a,b; Hamzei-Sichani et al.,007). Nonspecific effects of CBX at the concentrationsed in vivo are ruled out, because it does not affect theC firing rate while the concomitant LFPO is suppressed.his is in line with the preservation of antidromically andirectly evoked action potentials in the presence of CBX on
n vitro preparations (Schmitz et al., 2001). CBX has alsoe utilized recently (Blenkinsop and Lang, 2006) to dem-nstrate the role played by the electrotonic coupling of
nferior olivary neurons in the synchronization of complexpike activity as initially proposed by Llinás et al. (1974).njection of CBX in the inferior olive resulted in a decreasef complex spike firing rate, synchrony, and rhythmicityBlenkinsop and Lang, 2006). The CBX effect on syn-hrony and rhythmicity of inferior olive neurons corrobo-ates our CBX effect on the 160 Hz oscillation (Cheron etl., 2004a, 2005) and underlines the important role playedy gap junctions in brain oscillation. In Cb�/� mice, the
ncrease in Ca2� transient influx in cells (Airaksinen et al.,997; Barski et al., 2003) could form a link betweenhanges in Ca2� homeostasis and activation of a gapunction-coupled-network producing 160 Hz oscillations,s gating of gap junctions can be modified by increased
ntracellular [Ca2�] (De Pina-Benabou et al., 2001).Experimental and theoretical studies demonstrated
hat the collective behavior of gap-junction coupling be-ween axons of pyramidal cells results in high-frequencyeld oscillation in the hippocampus (Traub et al., 2003a,b).his fast oscillation occurs as a consequence of randomctivity within the axonal plexus and is uncovered when allhemical synapses are blocked. It was proposed that ec-opic spike generation at a low rate (0.05–1/s) is able toustain such high-frequency oscillation (Traub et al.,003a,b). Similar modeling of cerebellar slice fast oscilla-ion suggested that somatic prepotentials arose from ax-nal spikelets that were induced as coupling potentials inhe respective axon by spikes in neighboring axons (Traubt al., 2007).
CAUSAL LINKS BETWEEN OSCILLATION,FIRING RATE, RHYTHMICITY AND
SYNCHRONICITY
ifferent hypotheses attempt to explain the emergence ofFPO and PC synchronicity in mice lacking calcium-
inding proteins. First, increased excitability of granule bells (Gall et al., 2003) could lead to increased PC firingate, rhythmicity and synchronicity, possibly giving rise toast oscillation. Fast LFPO would thus be considered as aide effect of increased simple spike firing rate, whichould be the primary cause of ataxia and other cerebellarlectrophysiological abnormalities. Another view is thatast oscillation itself would be the primary cause of PCynchronization in a high rhythmic frequency range. Thisast field oscillation would then “trap” PCs into a rhythmic,ynchronous discharge pattern. This entrapment wouldender their firing less adaptable, which might possiblyead to clinical manifestations such as ataxia. These two
echanisms are not mutually exclusive. In neocortical fastFPO, Grenier et al. (2003) proposed a “vicious” feedback
oop in which very fast LFPO reflect the synchronous ac-ion of neocortical neurons and help to generate and syn-hronize action potentials in adjacent neurons.
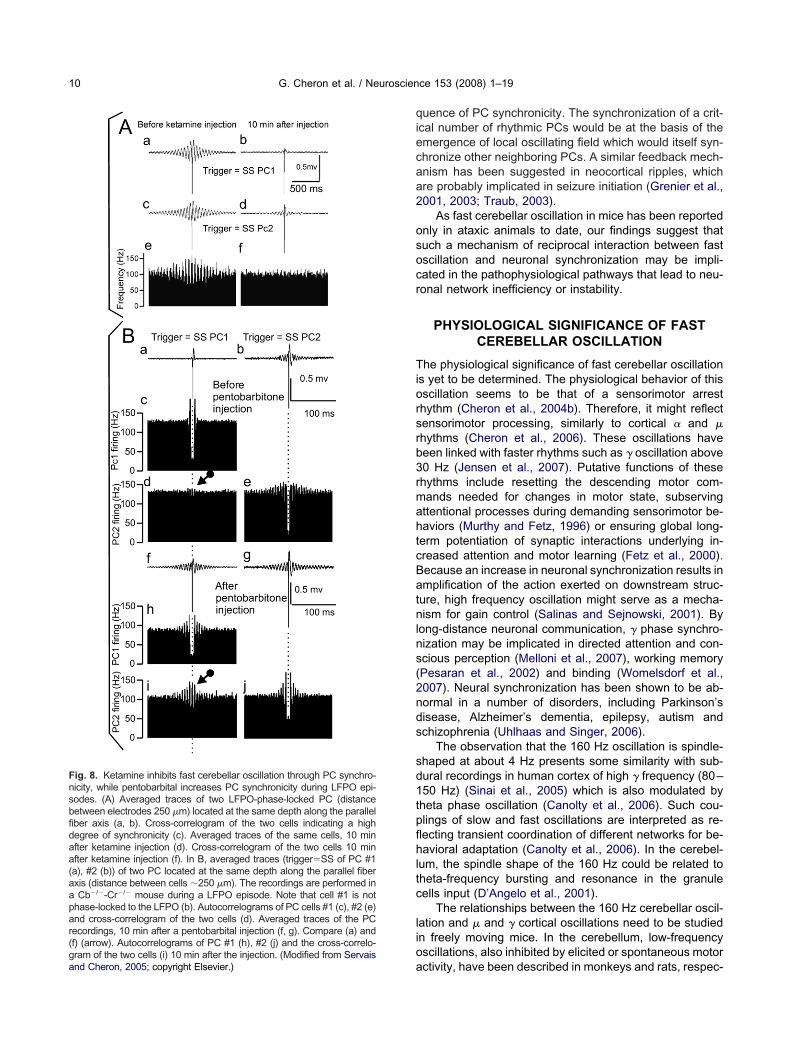
Insights into these questions have been gainedhrough the study of fast LFPO and PC firing before anduring general anesthesia induced by ketamine (a NMDAeceptor antagonist) (Fig. 7A, C, D) or pentobarbital (aABAA receptor positive modulator) (Fig. 7B) (Servais andheron, 2005). After ketamine injection, the peak of theourier spectrum is decreased, indicating inhibition, andhifted toward the left, indicating slowing down of theFPO (Fig. 7A). Paradoxically, in spite of this LFPO sup-ression, ketamine induced an increase of the PC rhyth-icity as illustrated in the autocorrelogram of the simple
pike firing (Fig. 7C versus Fig. 7D). The frequency of theFPO was reduced following ketamine injection. This ef-ect was related to decrease simple spike-firing synchro-icity (Fig. 7C, D and Fig. 8A). This could be explained byMDA receptor blockade. In the cerebellar cortex, NMDA
eceptors are located on interneurons, granule and brushells, and they mediate the mossy fibers excitation. Theyre also present at the presynaptic level, at the parallelber–PC and at the interneuron-PC synapse (Casado etl., 2002 and Duguid and Smart, 2004). Despite the im-unolabeling of some NMDA receptor subunits at the PC
evel in adult mice (Thompson et al., 2000), it is generallyssumed that PC excitation by parallel or climbing fiberction is not mediated by NMDA receptors. However, veryecent data demonstrated that mature PC express func-ional NMDA receptors taking part to the complex spikeave form (Piochon et al., 2007). The fact that the NMDAurrents are relatively small compared with the total CF-xcitatory currents and that the basic elements of theomplex spike response was maintained in the presencef APV (Piochon et al., 2007) seems to indicate that theffect of ketamine on simple spike-firing synchronicity wasot related to an effect on the climbing fiber input. There-ore, the main inhibiting and desynchronizing action ofetamine at the PC level is probably related to reduction inF activity due to reduced granule cell excitation. Ket-mine has also a number of other pharmacological prop-rties, including effects on acetylcholinesterase (Cohen etl., 1974), monoamine transporters (Smith et al., 1981)nd opiate receptors (Smith et al., 1980), which remain to
e investigated in this context.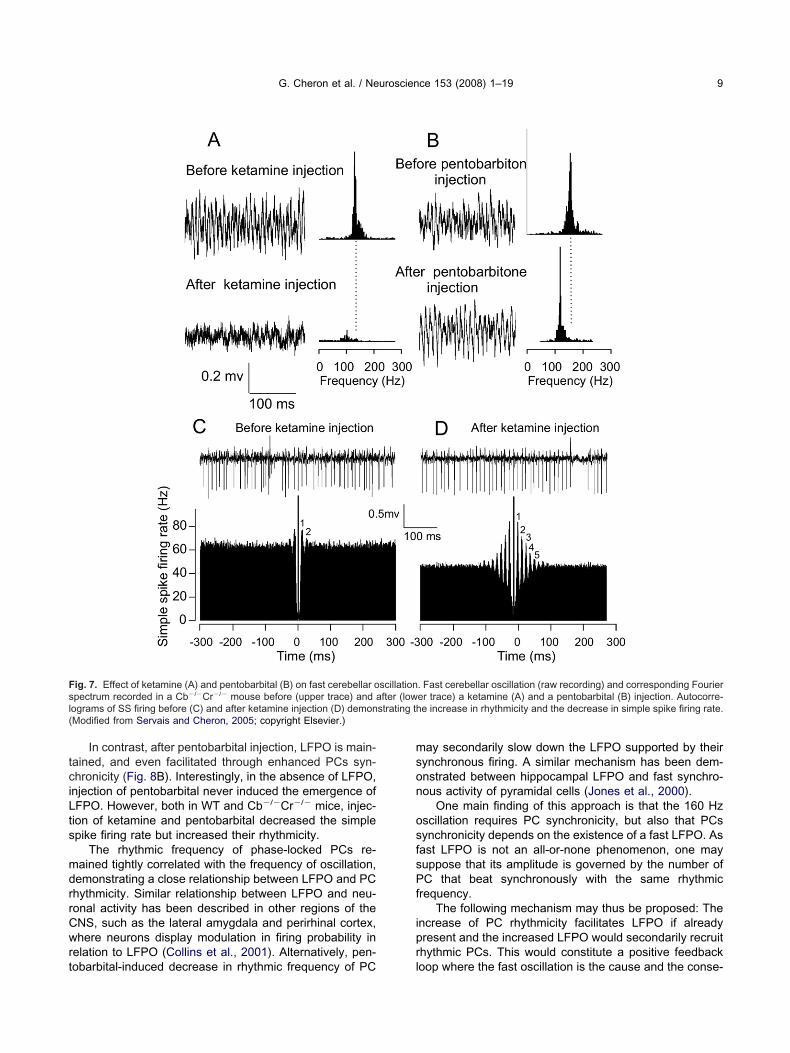
tciLts
mdrrCwrt
mson
osfsPf
ipr
Fsl(
G. Cheron et al. / Neuroscience 153 (2008) 1–19 9
In contrast, after pentobarbital injection, LFPO is main-ained, and even facilitated through enhanced PCs syn-hronicity (Fig. 8B). Interestingly, in the absence of LFPO,
njection of pentobarbital never induced the emergence ofFPO. However, both in WT and Cb�/�Cr�/� mice, injec-ion of ketamine and pentobarbital decreased the simplepike firing rate but increased their rhythmicity.
The rhythmic frequency of phase-locked PCs re-ained tightly correlated with the frequency of oscillation,emonstrating a close relationship between LFPO and PChythmicity. Similar relationship between LFPO and neu-onal activity has been described in other regions of theNS, such as the lateral amygdala and perirhinal cortex,here neurons display modulation in firing probability in
elation to LFPO (Collins et al., 2001). Alternatively, pen-
ig. 7. Effect of ketamine (A) and pentobarbital (B) on fast cerebellar opectrum recorded in a Cb�/�Cr�/� mouse before (upper trace) and aograms of SS firing before (C) and after ketamine injection (D) demonModified from Servais and Cheron, 2005; copyright Elsevier.)
obarbital-induced decrease in rhythmic frequency of PC l
ay secondarily slow down the LFPO supported by theirynchronous firing. A similar mechanism has been dem-nstrated between hippocampal LFPO and fast synchro-ous activity of pyramidal cells (Jones et al., 2000).
One main finding of this approach is that the 160 Hzscillation requires PC synchronicity, but also that PCsynchronicity depends on the existence of a fast LFPO. Asast LFPO is not an all-or-none phenomenon, one mayuppose that its amplitude is governed by the number ofC that beat synchronously with the same rhythmic
requency.The following mechanism may thus be proposed: The
ncrease of PC rhythmicity facilitates LFPO if alreadyresent and the increased LFPO would secondarily recruithythmic PCs. This would constitute a positive feedback
. Fast cerebellar oscillation (raw recording) and corresponding Fourierer trace) a ketamine (A) and a pentobarbital (B) injection. Autocorre-e increase in rhythmicity and the decrease in simple spike firing rate.
scillationfter (low
strating th
oop where the fast oscillation is the cause and the conse-
qiecaa2
osocr
Tiorsrb3rmahtcBatnlns(2nds
sd1tpflhltc
lioa
Fnsbfidaa(aapar(gand Cheron, 2005; copyright Elsevier.)
G. Cheron et al. / Neuroscience 153 (2008) 1–1910
uence of PC synchronicity. The synchronization of a crit-cal number of rhythmic PCs would be at the basis of themergence of local oscillating field which would itself syn-hronize other neighboring PCs. A similar feedback mech-nism has been suggested in neocortical ripples, whichre probably implicated in seizure initiation (Grenier et al.,001, 2003; Traub, 2003).
As fast cerebellar oscillation in mice has been reportednly in ataxic animals to date, our findings suggest thatuch a mechanism of reciprocal interaction between fastscillation and neuronal synchronization may be impli-ated in the pathophysiological pathways that lead to neu-onal network inefficiency or instability.
PHYSIOLOGICAL SIGNIFICANCE OF FASTCEREBELLAR OSCILLATION
he physiological significance of fast cerebellar oscillations yet to be determined. The physiological behavior of thisscillation seems to be that of a sensorimotor arresthythm (Cheron et al., 2004b). Therefore, it might reflectensorimotor processing, similarly to cortical � and �hythms (Cheron et al., 2006). These oscillations haveeen linked with faster rhythms such as � oscillation above0 Hz (Jensen et al., 2007). Putative functions of thesehythms include resetting the descending motor com-ands needed for changes in motor state, subservingttentional processes during demanding sensorimotor be-aviors (Murthy and Fetz, 1996) or ensuring global long-erm potentiation of synaptic interactions underlying in-reased attention and motor learning (Fetz et al., 2000).ecause an increase in neuronal synchronization results inmplification of the action exerted on downstream struc-ure, high frequency oscillation might serve as a mecha-ism for gain control (Salinas and Sejnowski, 2001). By
ong-distance neuronal communication, � phase synchro-ization may be implicated in directed attention and con-cious perception (Melloni et al., 2007), working memoryPesaran et al., 2002) and binding (Womelsdorf et al.,007). Neural synchronization has been shown to be ab-ormal in a number of disorders, including Parkinson’sisease, Alzheimer’s dementia, epilepsy, autism andchizophrenia (Uhlhaas and Singer, 2006).
The observation that the 160 Hz oscillation is spindle-haped at about 4 Hz presents some similarity with sub-ural recordings in human cortex of high � frequency (80–50 Hz) (Sinai et al., 2005) which is also modulated byheta phase oscillation (Canolty et al., 2006). Such cou-lings of slow and fast oscillations are interpreted as re-ecting transient coordination of different networks for be-avioral adaptation (Canolty et al., 2006). In the cerebel-
um, the spindle shape of the 160 Hz could be related toheta-frequency bursting and resonance in the granuleells input (D’Angelo et al., 2001).
The relationships between the 160 Hz cerebellar oscil-ation and � and � cortical oscillations need to be studiedn freely moving mice. In the cerebellum, low-frequencyscillations, also inhibited by elicited or spontaneous motor
ig. 8. Ketamine inhibits fast cerebellar oscillation through PC synchro-icity, while pentobarbital increases PC synchronicity during LFPO epi-odes. (A) Averaged traces of two LFPO-phase-locked PC (distanceetween electrodes 250 �m) located at the same depth along the parallelber axis (a, b). Cross-correlogram of the two cells indicating a highegree of synchronicity (c). Averaged traces of the same cells, 10 minfter ketamine injection (d). Cross-correlogram of the two cells 10 minfter ketamine injection (f). In B, averaged traces (trigger�SS of PC #1a), #2 (b)) of two PC located at the same depth along the parallel fiberxis (distance between cells �250 �m). The recordings are performed inCb�/�-Cr�/� mouse during a LFPO episode. Note that cell #1 is not
hase-locked to the LFPO (b). Autocorrelograms of PC cells #1 (c), #2 (e)nd cross-correlogram of the two cells (d). Averaged traces of the PCecordings, 10 min after a pentobarbital injection (f, g). Compare (a) andf) (arrow). Autocorrelograms of PC #1 (h), #2 (j) and the cross-correlo-ram of the two cells (i) 10 min after the injection. (Modified from Servais
ctivity, have been described in monkeys and rats, respec-

t1rgtatabap2
eTad2isendApaakdslalstcsd
itcmawtet
Rag
Obtinhr
ebnt1ttca
ikm(esdoac2
trabi(tm
F
TmcipamtwCihaCpd
gUc2pceps
G. Cheron et al. / Neuroscience 153 (2008) 1–19 11
ively (Pellerin and Lamarre, 1997; Hartmann and Bower,998). In these 8–18 Hz oscillations, the granular layerather than the PC layer was presumed to be the waveenerator, and PC firing was never in a clear phasic rela-ionship. In contrast, the phase locking of both simple spikend complex spike to the 160 Hz oscillation suggests thathe synchronous activities of PCs generate the oscillationnd exert influence on olivary neurons via the deep cere-ellar nucleus inhibitory input (Lang et al., 1996; Medina etl., 2002) and hence probably also on other cerebellarrojection areas (Kelly and Strick, 2003; Akkal et al.,007).
As in the hippocampus (Draguhn et al., 1998; Moortgatt al., 2000; Traub and Bibbig, 2000; Bartos et al., 2002;raub et al., 2002), fast oscillations point to a new mech-nism for synchronization that might take part in the coor-inating and timing function of the cerebellum (Mauk et al.,000; Medina et al., 2000). Networks of excitatory and
nhibitory neurons have been shown to display discreteilent, irregular (chaotic) or oscillatory states (Loewensteint al., 2005; Fernandez et al., 2007). According to Shan-on’s principle, the irregular state is optimal with respect toiffusion of information (Destexhe and Contreras, 2006).lthough the PC behavior recorded in alert animal alsoresents different states such as silent, irregularly tonicnd bursting (Womack and Khodakhah, 2002; Servais etl., 2004), the persistence of the 160 Hz oscillation innockout mice may represent the expression of cerebellareficit. The optimal functional state of PC populationeems to be situated between excessive irregularitieseading to motor deficits (Hoebeek et al., 2005; Walter etl., 2006) and persistent oscillations (i.e. excessive regu-
arity) resulting in ataxia (Cheron et al., 2005). The recenttudy of Medina and Lisberger (2007) demonstrated thathe optimal control of pursuit exerted by PC populationorresponds to transition from highly covariant simplepike firing during movement initiation to more indepen-ent firing later on.
Various approaches will be necessary to solve thisssue of whether 160 Hz directly reflects pathological func-ioning of the cerebellum or represents an adaptive pro-ess to overcome deficits in cerebellar function. Insightsay be gained from the study of genetically engineerednimals with specific rescuing of gene knockout. Insightsill also be provided by animal models of human condi-
ions with cerebellar ataxia or other manifestations of cer-bellar dysfunction. In this context, developmental condi-
ions are likely to be of particular interest.
estoration of normal cerebellar electrophysiologynd motor behavior by targeted Cr expression inranule cells of Cr�/� mice
ne interesting way to definitely prove the functional linketween the knockout of one Ca2� binding proteins in one
ype of cerebellar neuron and the reported functional deficits to genetically rescue this protein only in this type ofeuron. Bearzatto et al. (2006) generated Cr-rescue micearboring a specific rescue of Cr expression in these neu-
ons. This selective rescue restored (1) a normal neuronal (xcitability of granule cells in slices, (2) a normal PC firingehavior leading to the absence of oscillation and (3) aormal motor coordination (runway test) as compared withhe alterations detected in Cr�/� mice (Schiffmann et al.,999). These results confirm the functional links betweenhe initial protein deletion, the intrinsic alteration of theargeted neuron, the resulting network action on the prin-ipal cell of the network, the emergence of fast oscillationnd the resulting ataxia.
The coexistence of ataxia and changes of PC behav-or, including the 160 Hz LFPO in Ca2�-binding proteinsnockout mice may point to novel pathophysiologicalechanisms underlying cerebellar ataxia. In Cb�/� mice
Airaksinen et al., 1997; Farré-Castany et al., 2007), forxample, ataxia can be selectively attributed to the ab-ence of Cb in PC, since PC-specific Cb knockout miceisplay the same type of ataxia (Barski et al., 2003). More-ver, fine tuning of Ca2� signals in granule cells by Crppears to be required for correct network stability andomputation in the cerebellar cortex (Bearzatto et al.,006).
In this context, very recent data from our group showedhe emergence of beta oscillation phase-locked with ultra-hythmic Purkinje and Golgi cells in alert BK�/� mice. Welso demonstrated that this PC firing pattern and the ataxicehavior of BK�/� mice were reproduced in vivo by micro-
njection of BK channels blocker in WT mice cerebellumCheron et al., 2007). This reinforces the hypothesis thathe temporal coding modification of PC spike firing is theain cause of ataxia.
ast cerebellar oscillation in Angelman syndrome
he relationship between cerebellar fast oscillation andotor disorder can also be studied in models of human
onditions with ataxia. For example, Angelman syndromes characterized by mental retardation, a typical behavioralhenotype and motor dysfunction including ataxia (Dannd Cheron, 2003; Dan et al., 2004). All patients with aolecular diagnosis of Angelman syndrome have a func-
ional absence of the maternally inherited UBE3A gene,hich is located on chromosome 15 (Kishino et al., 1997).erebellar dysfunction has been suggested since the orig-
nal description of the syndrome (Angelman, 1965) andas been confirmed by functional imaging (Holopainen etl., 2001) and motor analysis (Dan et al., 2001; Dan andheron, 2004). In addition to ataxia, speech and cognitiveroblems have been hypothesized to result from cerebellarysfunction (Dan and Cheron, 2003).
Like Ca2�-binding proteins knockout mice the An-elman syndrome mouse models with knockout maternalbe3a (Ube3a m�/p�) showed ataxia but no morphologicerebellar abnormalities (Jiang et al., 1998; Miura et al.,002). However, these mice showed lack of Ube3a ex-ression specifically in PC layer (Miura et al., 2002) orytoplasmic accumulation of Ube3a substrate in PC (Jiangt al., 1998). Unexpectedly, but as in the Ca2�-bindingroteins knockout mice, the cerebellum of AS mice pre-ents very similar emergence of fast (160 Hz) oscillation
Cheron et al., 2005) (Fig. 9). This oscillation was related to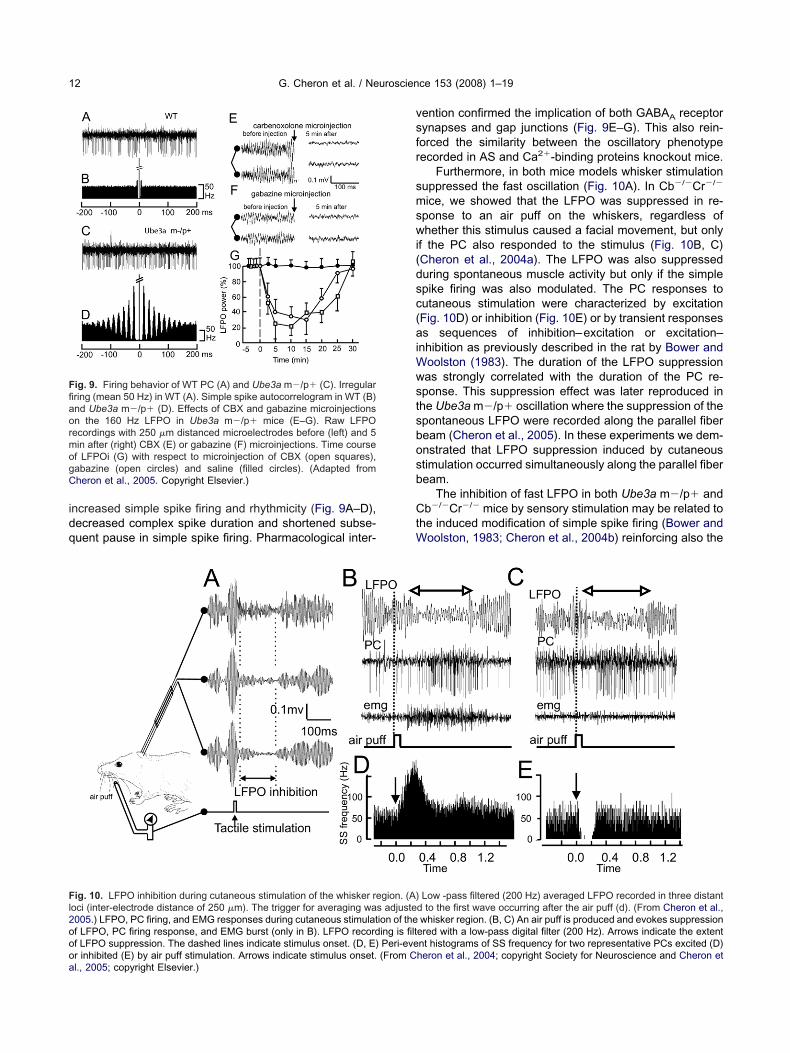
idq
vsfr
smswi(dsc(aiWwstsbosb
CtW
FfiaormogC
Fl2oooa
G. Cheron et al. / Neuroscience 153 (2008) 1–1912
ncreased simple spike firing and rhythmicity (Fig. 9A–D),ecreased complex spike duration and shortened subse-uent pause in simple spike firing. Pharmacological inter-
ig. 9. Firing behavior of WT PC (A) and Ube3a m�/p� (C). Irregularring (mean 50 Hz) in WT (A). Simple spike autocorrelogram in WT (B)nd Ube3a m�/p� (D). Effects of CBX and gabazine microinjectionsn the 160 Hz LFPO in Ube3a m�/p� mice (E–G). Raw LFPOecordings with 250 �m distanced microelectrodes before (left) and 5in after (right) CBX (E) or gabazine (F) microinjections. Time coursef LFPOi (G) with respect to microinjection of CBX (open squares),abazine (open circles) and saline (filled circles). (Adapted fromheron et al., 2005. Copyright Elsevier.)
ig. 10. LFPO inhibition during cutaneous stimulation of the whisker reoci (inter-electrode distance of 250 �m). The trigger for averaging wa005.) LFPO, PC firing, and EMG responses during cutaneous stimulatf LFPO, PC firing response, and EMG burst (only in B). LFPO recordf LFPO suppression. The dashed lines indicate stimulus onset. (D, E)
r inhibited (E) by air puff stimulation. Arrows indicate stimulus onset. (From Cl., 2005; copyright Elsevier.)ention confirmed the implication of both GABAA receptorynapses and gap junctions (Fig. 9E–G). This also rein-orced the similarity between the oscillatory phenotypeecorded in AS and Ca2�-binding proteins knockout mice.
Furthermore, in both mice models whisker stimulationuppressed the fast oscillation (Fig. 10A). In Cb�/�Cr�/�
ice, we showed that the LFPO was suppressed in re-ponse to an air puff on the whiskers, regardless ofhether this stimulus caused a facial movement, but only
f the PC also responded to the stimulus (Fig. 10B, C)Cheron et al., 2004a). The LFPO was also suppresseduring spontaneous muscle activity but only if the simplepike firing was also modulated. The PC responses toutaneous stimulation were characterized by excitationFig. 10D) or inhibition (Fig. 10E) or by transient responsess sequences of inhibition–excitation or excitation–
nhibition as previously described in the rat by Bower andoolston (1983). The duration of the LFPO suppressionas strongly correlated with the duration of the PC re-ponse. This suppression effect was later reproduced inhe Ube3a m�/p� oscillation where the suppression of thepontaneous LFPO were recorded along the parallel fibeream (Cheron et al., 2005). In these experiments we dem-nstrated that LFPO suppression induced by cutaneoustimulation occurred simultaneously along the parallel fibeream.
The inhibition of fast LFPO in both Ube3a m�/p� andb�/�Cr�/� mice by sensory stimulation may be related to
he induced modification of simple spike firing (Bower andoolston, 1983; Cheron et al., 2004b) reinforcing also the
Low -pass filtered (200 Hz) averaged LFPO recorded in three distantd to the first wave occurring after the air puff (d). (From Cheron et al.,whisker region. (B, C) An air puff is produced and evokes suppression
ered with a low-pass digital filter (200 Hz). Arrows indicate the extentnt histograms of SS frequency for two representative PCs excited (D)
gion. (A)s adjusteion of theing is filtPeri-eve
heron et al., 2004; copyright Society for Neuroscience and Cheron et
cg
tmficsTraU2hnmel
ta(ip2Uraprr1ctdUCc
Fs
Tdaiadtfpni
cs(toi
ccbc2nptccctrPac
cbcp
FsstpcvF(eac
G. Cheron et al. / Neuroscience 153 (2008) 1–19 13
ommon origin of the fast oscillation in these differentenetics models.
The neurophysiological LFPO similarities between mu-ant mice lacking Ube3a and calcium binding proteinsight suggest that these calcium binding proteins are de-
cient in Ube3a m�/p� mice. However, immunocyto-hemical staining demonstrated normal calbindin expres-ion in the PC of Ube3a m�/p� mice (Jiang et al., 1998).he molecular mechanisms by which Ube3a inactivationesults in Angelman syndrome and in cerebellar LFPOre still unclear. Different mechanisms may relatebe3a deficiency and GABAA transmission (Dan et al.,004a, b). Another mechanism might implicate Ca2�
omeostasis and GABAA transmission. Intrinsic neuro-al Ca2�conductances and intracellular [Ca2�] ho-eostasis are considered as crucial partners in themergence of neuronal network activity, including oscil-
ations (Llinás, 1988).In Ube3a m�/p� mice, deficits in hippocampal long-
erm potentiation have recently been related to diminishedctivity of the Ca2�/calmodulin-dependent protein kinase IICaMKII; Weeber et al., 2003). In PC, CaMKII is criticallynvolved in the signaling cascade regulating the longtermotentiation of GABAA receptors (Kawaguchi and Hirano,002). The increased spontaneous PC firing found inbe3a m�/p� mice may result from impairment in this
egulation process. Given the specific reduction in CaMKIIctivity demonstrated in Ube3a m�/p� mouse hippocam-us (Weeber et al., 2003), similar derangement may beeasonably suspected in PC, leading to alteration of theebound potentiation of the GABAA receptors (Kano et al.,992, 1996). CaMKII is also essential for PC plasticity anderebellar motor learning (Hansel et al., 2006). Recently,he group of Weeber (van Woerden et al., 2007) hasemonstrated that the hippocampal LTP deficit in thebe3a m�/p� mice can be rescued by decreasing aaMKII inhibitory phosphorylation. The effect of this res-uing on the cerebellum is the object of current studies.
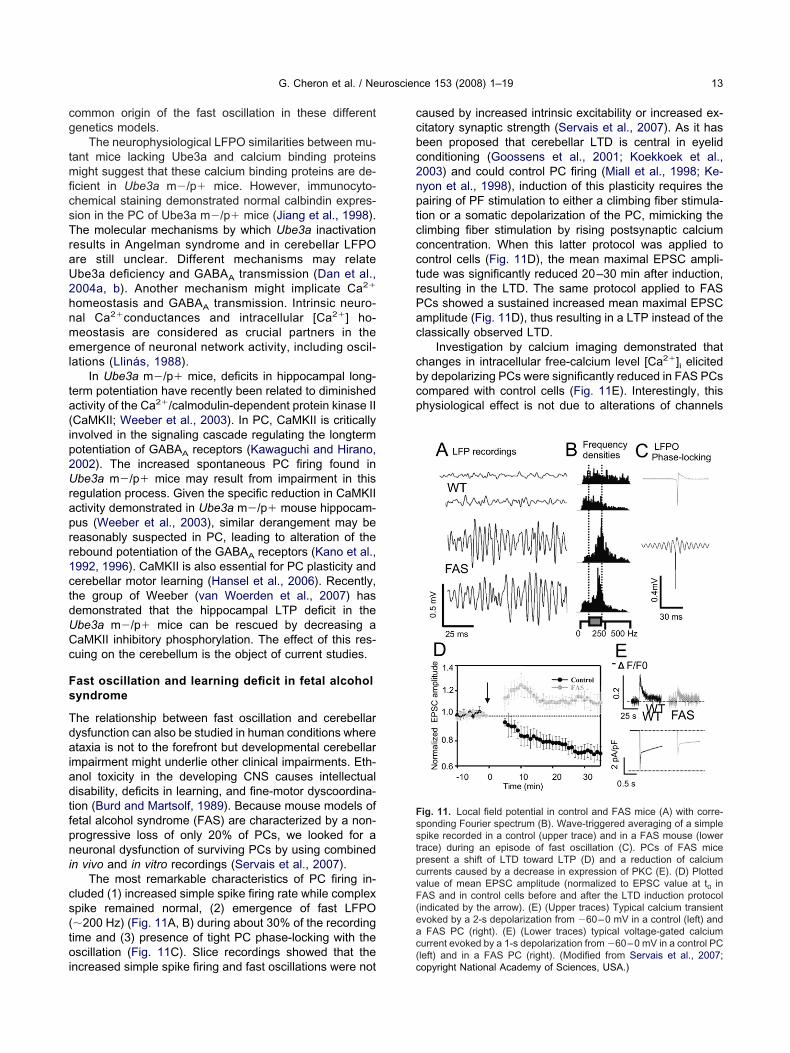
ast oscillation and learning deficit in fetal alcoholyndrome
he relationship between fast oscillation and cerebellarysfunction can also be studied in human conditions wheretaxia is not to the forefront but developmental cerebellar
mpairment might underlie other clinical impairments. Eth-nol toxicity in the developing CNS causes intellectualisability, deficits in learning, and fine-motor dyscoordina-ion (Burd and Martsolf, 1989). Because mouse models ofetal alcohol syndrome (FAS) are characterized by a non-rogressive loss of only 20% of PCs, we looked for aeuronal dysfunction of surviving PCs by using combined
n vivo and in vitro recordings (Servais et al., 2007).The most remarkable characteristics of PC firing in-
luded (1) increased simple spike firing rate while complexpike remained normal, (2) emergence of fast LFPO�200 Hz) (Fig. 11A, B) during about 30% of the recordingime and (3) presence of tight PC phase-locking with thescillation (Fig. 11C). Slice recordings showed that the
ncreased simple spike firing and fast oscillations were not(c
aused by increased intrinsic excitability or increased ex-itatory synaptic strength (Servais et al., 2007). As it haseen proposed that cerebellar LTD is central in eyelidonditioning (Goossens et al., 2001; Koekkoek et al.,003) and could control PC firing (Miall et al., 1998; Ke-yon et al., 1998), induction of this plasticity requires theairing of PF stimulation to either a climbing fiber stimula-ion or a somatic depolarization of the PC, mimicking thelimbing fiber stimulation by rising postsynaptic calciumoncentration. When this latter protocol was applied toontrol cells (Fig. 11D), the mean maximal EPSC ampli-ude was significantly reduced 20–30 min after induction,esulting in the LTD. The same protocol applied to FASCs showed a sustained increased mean maximal EPSCmplitude (Fig. 11D), thus resulting in a LTP instead of thelassically observed LTD.
Investigation by calcium imaging demonstrated thathanges in intracellular free-calcium level [Ca2�]i elicitedy depolarizing PCs were significantly reduced in FAS PCsompared with control cells (Fig. 11E). Interestingly, thishysiological effect is not due to alterations of channels
ig. 11. Local field potential in control and FAS mice (A) with corre-ponding Fourier spectrum (B). Wave-triggered averaging of a simplepike recorded in a control (upper trace) and in a FAS mouse (lowerrace) during an episode of fast oscillation (C). PCs of FAS miceresent a shift of LTD toward LTP (D) and a reduction of calciumurrents caused by a decrease in expression of PKC (E). (D) Plottedalue of mean EPSC amplitude (normalized to EPSC value at t0 inAS and in control cells before and after the LTD induction protocolindicated by the arrow). (E) (Upper traces) Typical calcium transientvoked by a 2-s depolarization from �60–0 mV in a control (left) andFAS PC (right). (E) (Lower traces) typical voltage-gated calcium
urrent evoked by a 1-s depolarization from �60–0 mV in a control PC
left) and in a FAS PC (right). (Modified from Servais et al., 2007;opyright National Academy of Sciences, USA.)
eip
smt(itghPire
tpdf
p(tioottAfieeKamFasirbvcmeoMlsdl
teadd
avfcoF
Tli(
oLamfpatsatdtsrmHaa
dPmnCSsshipme
AS1atDa
A
G. Cheron et al. / Neuroscience 153 (2008) 1–1914
xpression or activation/inactivation kinetics. In contrast, its related to a reduced expression of the � isotype of therotein kinase C (PKC �) (Servais et al., 2007).
Co-immunostaining for calbindin and PKC�, a neuron-pecific calcium-activated enzyme involved in receptor-ediated phospho-inositide signaling cascades, confirmed
hat expression of this isoform was confined within PCsBareggi et al., 1996; Barmack et al., 2000). The functionalmportance of PKC� is illustrated in spinocerebellar ataxiaype 14, which is secondary to a missense mutation in itsene (Yabe et al., 2003). Although, PKC� knockout miceave normal LTD, they have distorted cerebellar circuitry.Cs in PKC� knockout mice receive multiple climbing fiber
nnervation. This multiple climbing fiber innervation may beesponsible for the ataxia expressed by these mice (Chent al., 1995).
FAS mice demonstrated significant impairment in mo-or coordination in both rotarod and runway assays. Com-arison of the daily percentage of conditioned responseuring eyelid conditioning demonstrated a significant dif-
erence in motor learning between FAS and control mice.LTD at the parallel fiber–PC synapse has been pro-
osed as the mechanism that regulates PC firing in vivoMiall et al., 1998; Kenyon et al., 1998). It can be arguedhat the abolition of LTD in PCs does not cause alterationn PC firing in vivo or motor coordination impairment (Go-ssens et al., 2001). However, in FAS mice LTD was notnly absent but replaced by LTP. Thus, we hypothesizehat this shift results in a chronic overstimulation of PCs byhe parallel fibers, resulting in increased PC firing in vivo.ccording to the view that an intact LTD at the parallelber–PC synapse is required for the optimal efficiency ofyelid conditioning studied as a prototypical reflect of cer-bellar learning (Koekkoek et al., 2003; Thompson andrupa, 1994), the shift of LTD toward LTP in FAS couldlso contribute to the deficits in eyelid conditioning andotor learning. Whether the replacement of LTD by LTP inAS mice constitutes the only cause of LFPO emergencend PCs firing behavior in FAS has not yet been demon-trated, and similar approach would also be accomplishedn the other mice showing 160 Hz oscillation. Whatever theesult of these future investigations examining the linketween LTD, fast oscillations and other mechanisms in-olving PCs, other cerebellar neurons might also be impli-ated. For instance, increased simple spike firing in FASice could partially be due to granule cells alteration bythanol (Offenhäuser et al., 2006) or from hyperexcitationf extracerebellar areas that project on the cerebellum.oreover, the role of LTD in motor learning was chal-
enged by Welsh et al. (2005), as they demonstrated thatuppression of parallel fiber–PC LTD by the ethanol hy-rochloride (T-588) in alert rat does not affect rotorod
earning or adaptation of eyeblink conditioning.Finally, LTD has been suggested to represent a pro-
ection mechanism against excitotoxic cell death (Kimurat al., 2005). This may be particularly relevant as PCs aremong the most vulnerable neurons to excitotoxic celleath. In particular they are killed by climbing fibers input
uring states when the inferior olive is pathologically over-ctive (Mameli et al., 2005). As ethanol is known to acti-ate inferior olive neurons increasing the complex spikerequency, the PC of the FAS mice unprotected by LTDould be more vulnerable against apoptosis in the devel-pment stage explaining the 20% of PC deaths in matureAS.
CONCLUSIONS
he role of the 160 Hz LFPO is currently unclear. It couldead to ataxia or reflect a compensatory mechanism allow-ng the cerebellum to function despite abnormal PC firingCheron et al., 2004a,b).
Enhanced PC synchronization may facilitate recordingf cerebellar input into the phase discharges of the 160 HzFPO, increasing the precision of timing control in face oflteration calcium signaling or other basic processes. Thisight restore some degree of network functioning. The
ocal mossy fiber input related to cutaneous stimulationrobably overcomes the generalized state of synchronousctivation of PC populations. LFPO arrest is produced byransient excitation–inhibition or inhibition–excitation re-ponse of PC (Cheron et al., 2004a). Occurrence of thisrrest while the mouse can show a physiological responseo sensory stimulation suggests that the fast oscillationoes not completely sever the cerebellar from environmen-al information, at least in some contextual settings. LFPOynchrony could be hypothesized to act as a spatiotempo-al filter sharpening the action of selected rostrocaudalodules of the cerebellum (Voogd and Glickstein, 1998).owever, the topography of synchronization along the par-llel fiber beam rather than along the rostro-caudal axisrgues against this putative role.
To date, 160 Hz oscillation has been reported in sixifferent knockout mice (Cb�/�, Cr�/�, Cb�/�Cr�/�,v�/�, Pv�/�Cb�/� and Ube3a m�/p�) and in the FASice. All these animals showed cerebellar ataxia (Airaksi-en et al., 1997; Schiffmann et al., 1999; Miura et al., 2002;heron et al., 2004, 2005; Servais et al., 2005, 2007).uch high-frequency oscillation sustained by increasedimple spike firing and rhythmicity has never been de-cribed in WT or in non-ataxic mice. LFPO could then beypothesized to disrupt cerebellar physiological rhythms
nvolved in motor coordination. This may have major im-lications for future therapeutic targeting, as fast LFPOay be inhibited by different pharmacological and geneticsngineering options.
cknowledgments—This work was funded by the Belgian Federalcience Policy Office, the European Space Agency, (AO-2004,18), the Belgian National Fund for Scientific Research (FNRS)nd research funds of the Université Libre de Bruxelles (ULB) andhe Université de Mons-Hainaut (Belgium). The authors thank P.emaret and M. Dufief and E. Hortmanns for expert technicalssistance.
REFERENCES
iraksinen MS, Eilers J, Garaschuk O, Thoenen H, Konnerth A, Meyer
M (1997) Ataxia and altered dendritic calcium signaling in mice
A
A
A
B
B
B
B
B
B
B
B
B
B
BB
B
B
B
B
B
B
B
B
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
CC
C
C
C
C
G. Cheron et al. / Neuroscience 153 (2008) 1–19 15
carrying a targeted null mutation of the calbindin D28k gene. ProcNatl Acad Sci U S A 94:1488–1493.
kkal D, Dum RP, Strick PL (2007) Supplementary motor area andpresupplementary motor area: targets of basal ganglia and cere-bellar output. J Neurosci 27:10659–10673.
ndreasen NC, Paradiso S, O’Leary DS (1998) “Cognitive dysmetria”as an integrative theory of schizophrenia: a dysfunction in cortical-subcortical-cerebellar circuitry? Schizophr Bull 24:203–218.
ngelman H (1965) “Puppet” children: a report on three cases. DevMed Child Neurol 7:681–688.
arbe MT, Monyer H, Bruzzone R (2006) Cell-cell communicationbeyond connexins: the pannexin channels. Physiology (Bethesda)21:103–114.
arbour B (1993) Synaptic currents evoked in Purkinje cells by stim-ulating individual granule cells. Neuron 11:759–769.
areggi R, Narducci P, Grill V, Lach S, Martelli AM (1996) Selectivedistribution of multiple protein kinase C isoforms in mouse cere-bellar cortex. Biol Cell 7:55–63.
armack NH, Qian Z, Yoshimura J (2000) Regional and cellular dis-tribution of protein kinase C in rat cerebellar Purkinje cells. J CompNeurol 427:235–254.
armack NH, Yakhnitsa V (2003) Cerebellar climbing fibers modulatesimple spikes in Purkinje cells. J Neurosci 23:7904–7916.
arski JJ, Hartmann J, Rose CR, Hoebeek F, Morl K, Noll-Hussong M,De Zeeuw CI, Konnerth A, Meyer M (2003) Calbindin in cerebellarPurkinje cells is a critical determinant of the precision of motorcoordination. J Neurosci 23:3469–3477.
artos M, Vida I, Frotscher M, Meyer A, Monyer H, Geiger JR, JonasP (2002) Fast synaptic inhibition promotes synchronized gammaoscillationsin hippocampal interneuron networks. Proc Natl AcadSci U S A 99:13222–13227.
artos M, Vida I, Jonas P (2007) Synaptic mechanisms of synchro-nized gamma oscillations in inhibitory interneuron networks. NatRev Neurosci 8:45–56.
astian AJ (2006) Learning to predict the future: the cerebellumadapts feedforward movement control. Curr Opin Neurobiol16:645–649.
earzatto B, Servais L, Roussel C, Gall D, Baba-Aïssa F, SchurmansS, de Kerchove d’Exaerde A, Cheron G, Schiffmann SN (2006)Targeted calretinin expression in granule cells of calretinin-nullmice restores normal cerebellar functions. FASEB J 20:380–382.
erridge MJ (1998) Neuronal calcium signaling. Neuron 21:13–26.lenkinsop TA, Lang EJ (2006) Block of inferior olive gap junctional
coupling decreases Purkinje cell complex spike synchrony andrhythmicity. J Neurosci 26:1739–1748.
ooth JR, Wood L, Lu D, Houk JC, Bitan T (2007) The role of the basalganglia and cerebellum in language processing. Brain Res1133:136–144.
ower JM, Woolston DC (1983) Congruence of spatial organization oftactile projections to granule cell and Purkinje cell layers of cere-bellar hemispheres of the albino rat: vertical organization of cer-ebellar cortex. J Neurophysiol 49:745–766.
raitenberg V (1967) Is the cerebellar cortex a biological clock in themillisecond range? Prog Brain Res 25:334–346.
ruzzone R, Barbe MT, Jakob NJ, Monyer H (2005) Pharmacologicalproperties of homomeric and heteromeric pannexin hemichannelsexpressed in Xenopus oocytes. J Neurochem 92:1033–1043.
ruzzone R, Hormuzdi SG, Barbe MT, Herb A, Monyer H (2003)Pannexins, a family of gap junction proteins expressed in brain.Proc Natl Acad Sci U S A 100:13644–13649.
urd L, Martsolf JT (1989) Fetal alcohol syndrome: diagnosis andsyndromal variability. Physiol Behav 46:39–43.
uzsáki G, Horvath Z, Urioste R, Hetke J, Wise K (1992) High-frequency network oscillation in the hippocampus. Science 256:1025–1027.
uzsáki G, Draguhn A (2004) Neuronal oscillations in cortical net-
works. Science 304:1926–1929.ampbell NC, Hesslow G (1986a) The secondary spikes of climbingfibre responses recorded from Purkinje cell somata in cat cerebel-lum. J Physiol (Lond) 377:207–224.
ampbell NC, Hesslow G (1986b) The secondary spikes of climbingfibre responses recorded from Purkinje cell axons in cat cerebel-lum. J Physiol (Lond) 377:225–235.
anolty RT, Edwards E, Dalal SS, Soltani M, Nagarajan SS, KirschHE, Berger MS, Barbaro NM, Knight RT (2006) High gamma poweris phase-locked to theta oscillations in human neocortex. Science313:1626–1628.
arey MR, Lisberger SG (2004) Signals that modulate gain control forsmooth pursuit eye movements in monkeys. J Neurophysiol91:623–631.
arter AG, Regehr WG (2002) Quantal events shape cerebellar inter-neuron firing. Nat Neurosci 5:1309–1318.
asado M, Isope P, Ascher P (2002) Involvement of presynapticN-methyl-D-aspartate receptors in cerebellar long-term depres-sion. Neuron 33:123–130.
havas J, Marty A (2003) Coexistence of excitatory and inhibitoryGABA synapses in the cerebellar interneuron network. J Neurosci23:2019–2031.
hen C, Kano M, Abeliovich A, Chen L, Bao S, Kim JJ, Hashimoto K,Thompson RF, Tonegawa S (1995) Impaired motor coordinationcorrelates wit persistent multiple climbing fiber innervation in PKC�
mutant mice. Cell 83:1233–1242.heron G (1990) Effect of incisions in the brainstem commissural
network on the short-term vestibulo-ocular adaptation of the cat. JVestib Res 1:223–239.
heron G, Gall D, Servais L, Dan B, Maex R, Schiffmann SN (2004a)Inactivation of calcium-binding protein genes induces 160 Hz os-cillations in the cerebellar cortex of alert mice. J Neurosci 24:434–441.
heron G, Servais L, Dan B, Gall D, Roussel C, Schiffmann SN(2004b) Fast oscillation in the cerebellar cortex of calcium bindingprotein-deficient mice: a new sensorimotor arrest rhythm. ProgBrain Res 148:167–180.
heron G, Leroy A, De Saedeleer C, Bengoetxea A, Lipshits M,Cebolla A, Servais L, Dan B, Berthoz A, McIntyre J (2006) Effect ofgravity on human spontaneous 10-Hz electroencephalographicoscillations during the arrest reaction. Brain Res 1121:104–116.
heron G, Schurmans S, Lohof A, d’Alcantara P, Meyer M, Draye JP,Parmentier M, Schiffmann SN (2000) Electrophysiological behav-ior of Purkinje cells and motor coordination in calretinin knock-outmice. Prog Brain Res 124:299–308.
heron G, Servais L, Cebolla A, Dufief M, Dan B, Sausbier M, StormJ, Ruth P (2007) Altered Purkinje cell firing phase-locked to betaoscillation in the cerebellar cortex of mice lacking BK channels.Soc Neurosci Abstr 512.14/RR18.
heron G, Servais L, Wagstaff J, Dan B (2005) Fast cerebellar oscil-lation associated with ataxia in a mouse model of Angelman syn-drome. Neuroscience 130:631–637.
lapham DE (1995) Calcium signaling. Cell 80:259–268.oesmans M, Weber JT, De Zeeuw CI, Hansel C (2004) Bidirectional
parallel fiber plasticity in the cerebellum under climbing fiber con-trol. Neuron 44:691–700.
ohen ML, Chan SL, Bhargave HN, Trevor AJ (1974) Inhibition ofmammalian brain acetylcholinesterase by ketamine. BiochemPharmacol 23:1647–1652.
ohen D, Yarom Y (1998) Patches of synchronized activity in the cere-bellar cortex evoked by mossy-fiber stimulation: questioning the roleof parallel fibers. Proc Natl Acad Sci U S A 95:15032–15036.
olin F, Manil J, Desclin JC (1980) The olivocerebellar system I.Delayed and slow inhibitory effects: an overlooked salient featureof cerebellar climbing fibers. Brain Res 187:3–27.
ollins DR, Pelletier JG, Pare D (2001) Slow and fast (gamma) neu-ronal oscillations in the perirhinal cortex and lateral amygdala.
J Neurophysiol 85:1661–1672.
D
D
D
D
D
D
D
D
D
D
D
D
D
D
D
D
D
D
D
E
E
E
E
E
F
F
F
F
F
G
G
G
G
G
G
G
G
H
H
H
H
H
H
H
H
H
G. Cheron et al. / Neuroscience 153 (2008) 1–1916
an B, Bouillot E, Bengoetxea A, Boyd SG, Cheron G (2001) Distinctivemultijoint control strategies in spastic diplegia associated with prema-turity or Angelman syndrome. Clin Neurophysiol 112:1618–1625.
an B, Boyd SG, Cheron G (2004a) From genomic imprinting todevelopmental physiology: identifying stepping stones. Curr Phar-macogenom 2:3.
an B, Servais L, Boyd SG, Wagstaff J, Cheron G (2004b) Fromelectrophysiology to chromatin: a bottom-up approach to An-gelman syndrome. Ann N Y Acad Sci 1030:599–611.
an B, Cheron G (2003) Angelman syndrome: clinical and geneticmodel. Rev Neurol (Paris) 159:499–510.
an B, Cheron G (2004) Postural rhythmic muscle bursting activity inAngelman syndrome. Brain Dev 26:389–393.
’Angelo E, De Filippi G, Rossi P, Taglietti V (1997) Synaptic activa-tion of Ca2� action potentials in immature rat cerebellar granulecells in situ. J Neurophysiol 78:1631–1642.
’Angelo E, De Filippi G, Rossi P, Taglietti V (1998) Ionic mechanismof electroresponsiveness in cerebellar granule cells implicates theaction of a persistent sodium current. J Neurophysiol 80:493–503.
’Angelo E, Nieus T, Maffei A, Armano S, Rossi P, Taglietti V, FontanaA, Naldi G (2001) Theta-frequency bursting and resonance incerebellar granule cells: experimental evidence and modeling of aslow K�-dependent mechanism. J Neurosci 21:759–770.
elgado-Garcia JM, Gruart A (2002) The role of interpositus nucleus ineyelid conditioned responses. Cerebellum 1:289–308.
elgado-Garcia JM, Gruart A (2006) Building new motor responses:eyelid conditioning revisited. Trends Neurosci 29:330–338.
e Pina-Benabou MH, Srinivas M, Spray DC, Scemes E (2001) Cal-modulin kinase pathway mediates the K�-induced increase in gapjunctional communication between mouse spinal cord astrocytes.J Neurosci 21:6635–6643.
esclin JC (1976) Early terminal degeneration of cerebellar climbingfibers after destruction of the inferior olive in the rat. Synapticrelationships in the molecular layer. Anat Embryol (Berl) 149:87–112.
estexhe A, Contreras D (2006) Neuronal computations with stochas-tic network states. Science 314:85–90.
e Zeeuw CI, Yeo CH (2005) Time and tide in cerebellar memoryformation. Curr Opin Neurobiol 15:667–674.
iedrichsen J, Criscimagna-Hemminger SE, Shadmehr R (2007) Dis-sociating timing and coordination as functions of the cerebellum.J Neurosci 27:6291–6301.
ieudonné S, Dumoulin A (2000) Serotonin-driven long-range inhibitoryconnections in the cerebellar cortex. J Neurosci 20:1837–1848.
raguhn A, Traub RD, Schmitz D, Jefferys JG (1998) Electrical cou-pling underlies high-frequency oscillations in the hippocampus invitro. Nature 394:189–192.
ugué GP, Dumoulin A, Triller A, Dieudonné S (2005) Target-depen-dent use of co-released inhibitory transmitters at central synapses.J Neurosci 25:6490–6498.
uguid IC, Smart TG (2004) Retrograde activation of presynapticNMDA receptors enhances GABA release at cerebellar interneu-ron-Purkinje cell synapses. Nat Neurosci 7:525–533.
ccles JC, Ito M, Szentagothai J (1967) The cerebellum as a neuronalmachine. Berlin: Springer-Verlag.
ccles JC, Llinás R, Sasaki K (1966a) The inhibitory interneuroneswithin the cerebellar cortex. Exp Brain Res 1:1–16.
ccles JC, Llinás R, Sasaki K (1966b) The mossy fibre-granule cellrelay of the cerebellum and its inhibitory control by Golgi cells. ExpBrain Res 1:82–101.
ccles JC, Llinás R, Sasaki K (1966c) The excitatory synaptic action ofclimbing fibres on the Purkinje cells of the cerebellum. J Physiol182:268–296.
ccles JC, Llinás R, Sasaki K, Voorhoeve PE (1966d) Interactionexperiments on the responses evoked in Purkinje cells by climbingfibres. J Physiol 182:297–315.
arré-Castany MA, Schwaller B, Gregory P, Barski J, Mariethoz C,
Eriksson JL, Tetko IV, Wolfer D, Celio MR, Schmutz I, Albrecht U,Villa AE (2007) Differences in locomotor behavior revealed in micedeficient for thecalcium-binding proteins parvalbumin, calbindinD-28k or both. Behav Brain Res 178:250–261.
ellous JM, Rudolph M, Destexhe A, Sejnowski TJ (2003) Synapticbackground noise controls the input/output characteristics of singlecells in an in vitro model of in vivo activity. Neuroscience 122:811–829.
ernandez FR, Engbers JD, Turner RW (2007) Firing dynamics ofcerebellar purkinje cells. J Neurophysiol 98:278–294.
etz EE, Chen D, Murthy VN, Matsumura M (2000) Synaptic interac-tions mediating synchrony and oscillations in primate sensorimotorcortex. J Physiol (Paris) 94:323–331.
inch EA, Augustine GJ (1998) Local calcium signalling by inositol-1,4,5-trisphosphate in Purkinje cell dendrites. Nature 396:753–756.
all D, Roussel C, Nieus T, Cheron G, Servais L, D’Angelo E, Schiff-mann SN (2005) Role of calcium binding proteins in the control ofcerebellar granule cell neuronal excitability: experimental and mod-eling studies. Prog Brain Res 148:321–328.
all D, Roussel C, Susa I, D’Angelo E, Rossi P, Bearzatto B, GalasMC, Blum D, Schurmans S, Schiffmann SN (2003) Altered neuro-nal excitability in cerebellar granule cells of mice lacking calretinin.J Neurosci 23:9320–9327.
ao JH, Parsons LM, Bower JM, Xiong J, Li J, Fox PT (1996) Cere-bellum implicated in sensory acquisition and discrimination ratherthan motor control. Science 272:545–547.
ilbert PF (1976) Simple spike frequency and the number of second-ary spikes in the complex spike of the cerebellar Purkinje cell. BrainRes 114:334–338.
oossens J, Daniel H, Rancillac A, van der Steen J, Oberdick J,Crepel F, De Zeeuw, CI , Frens MA (2001) Expression of proteinkinase C inhibitor blocks cerebellar long-term depression withoutaffecting Purkinje cell excitability in alert mice. J Neurosci 21:5813–5823.
ordon N (2007) The cerebellum and cognition. Eur J Paediatr Neurol11:232–234.
renier F, Timofeev I, Steriade M (2001) Focal synchronization ofripples (80–200 Hz) in neocortex and their neuronal correlates.J Neurophysiol 86:1884–1898.
renier F, Timofeev I, Steriade M (2003) Neocortical very fast oscil-lations (ripples 80–200 Hz) during seizures: intracellular corre-lates. J Neurophysiol 89:841–852.
amori J, Szentagothai J (1966) Identification under the electronmicroscope of climbing fibers and their synaptic contacts. ExpBrain Res 1:65–81.
amzei-Sichani F, Kamasawa N, Janssen WG, Yasumura T, David-son KG, Hof PR, Wearne SL, Stewart MG, Young SR, WhittingtonMA, Rash JE, Traub RD (2007) Gap junctions on hippocampalmossy fiber axons demonstrated by thin-section electron micros-copy and freeze fracture replica immunogold labeling. Proc NatlAcad Sci U S A 104:12548–12553.
ansel C, de Jeu M, Belmeguenai A, Houtman SH, Buitendijk GH,Andreev D, De Zeeuw CI, Elgersma Y (2006) alphaCaMKII Isessential for cerebellar LTD and motor learning. Neuron 51:835–843.
ansel C, Linden DJ (2000) Long-term depression of the cerebellarclimbing fiber-Purkinje neuron synapse. Neuron 26:473–482.
artmann MJ, Bower JM (1998) Oscillatory activity in the cerebellarhemispheres of unrestrained rats. J Neurophysiol 80:1598–1604.
arvey RJ, Napper RM (1991) Quantitative studies on the mammaliancerebellum. Prog Neurobiol 36(6):437–463.
ashimoto K, Kano KM (1998) Presynaptic origin of paired-pulsedepression at climbing fibre-Purkinje cell synapses in the rat cer-ebellum. J Physiol (Lond) 506:391–405.
äusser M, Clark BA (1997) Tonic synaptic inhibition modulates neu-ronal output pattern and spatiotemporal synaptic integration. Neu-ron 19:665–678.
äusser M, Raman IM, Otis T, Smith SL, Nelson A, du Lac S, Loe-
wenstein Y, Mahon S, Pennartz C, Cohen I, Yarom Y (2004) The
H
H
H
I
I
II
I
J
J
J
K
K
K
K
K
K
K
K
K
K
K
L
L
L
L
L
L
L
L
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
G. Cheron et al. / Neuroscience 153 (2008) 1–19 17
beat goes on: spontaneous firing in mammalian neuronalmicrocircuits. J Neurosci 24:9215–9219.
eck DH, Thach WT, Keating JG (2007) On-beam synchrony in thecerebellum as the mechanism for the timing and coordination ofmovement. Proc Natl Acad Sci U S A 104(18):7658–7663.
oebeek FE, Stahl JS, van Alphen AM, Schonewille M, Luo C, Rut-teman M, van den Maagdenberg AM, Molenaar PC, Goossens HH,Frens MA, De Zeeuw CI (2005) Increased noise level of purkinjecell activities minimizes impact of their modulation during sensori-motor control. Neuron 45:953–965.
olopainen IE, Metsahonkala EL, Kokkonen H, Parkkola RK, MannerTE, Nagren K, Korpi ER (2001) Decreased binding of [11C]fluma-zenil in Angelman syndrome patients with GABA(A) receptor beta3subunit deletions. Ann Neurol 49:110–113.
to M (1972) Neuron design of the cerebellar motor control system.Brain Res 40:81–4.
to M (1982) Cerebellar control of the vestibulo-ocular reflex aroundthe flocculus hypothesis. Annu Rev Neurosci 5:275–296.
to M (1989) Long-term depression. Annu Rev Neurosci 12:85–102.to M, Simpson J (1971) Discharges in Purkinje cell axons during
climbing fibre activation. Brain Res 31:215–219.vry RB, Spencer RM, Zelaznik HN, Diedrichsen J (2002) The cere-
bellum and event timing. Ann N Y Acad Sci 978:302–317.ensen O, Kaiser J, Lachaux JP (2007) Human gamma-frequency
oscillations associated with attention and memory. Trends Neuro-sci 30:317–324.
iang Y, Armstrong D, Albrecht U, Atkins CM, Noebels JL, Eichele G,Sweatt JD, Beaudet AL (1998) Mutation of the Angelman ubiquitinligase in mice causes increased cytoplasmic p53 and deficits ofcontextual learning and long-term potentiation. Neuron 21:799–811.
ones MS, MacDonald KD, Choi B, Dudek FE, Barth DS (2000)Intracellular correlates of fast (200 Hz) electrical oscillations in ratsomatosensory cortex. J Neurophysiol 84:1505–1518.
ahlon M, Lisberger SG (2000) Changes in the responses of Purkinjecells in the floccular complex of monkeys after motor learning insmooth pursuit eye movements. J Neurophysiol 84:2945–2960.
ano M, Kano M, Fukunaga K, Konnerth A (1996) Ca(2�)-inducedrebound potentiation of gamma-aminobutyric acid-mediated cur-rents requires activation of Ca2�/calmodulin-dependent kinase II.Proc Natl Acad Sci U S A 93:13351–13356.
ano M, Rexhausen U, Dreessen J, Konnerth A (1992) Synaptic excita-tion produces a long-lasting rebound potentiation of inhibitory synapticsignals in cerebellar Purkinje cells. Nature 356:601–604.
awaguchi SY, Hirano T (2002) Signaling cascade regulating long-term potentiation of GABA(A) receptor responsiveness in cerebel-lar Purkinje neurons. J Neurosci 22:3969–3976.
azantsev VB, Nekorkin VI, Makarenko VI, Llinás R (2003) Olivo-cerebellar cluster-based universal control system. Proc Natl AcadSci U S A 100:13064–13068.
elly RM, Strick PL (2003) Cerebellar loops with motor cortex and pre-frontal cortex of a nonhuman primate. J Neurosci 23:8432–8444.
enyon GT, Medina JF, Mauk MD (1998) A mathematical model of thecerebellar-olivary system II: motor adaptation through systematic dis-ruption of climbing fiber equilibrium. J Comput Neurosci 5:71–90.
imura T, Sugimori M, Llinás RR (2005) Purkinje cell long-term de-pression is prevented by T-588, a neuroprotective compound thatreduces cytosolic calcium release from intracellular stores. ProcNatl Acad Sci U S A 102:17160–17165.
ishino T, Lalande M, Wagstaff J (1997) UBE3A/E6-AP mutationscause Angelman syndrome. Nat Genet 15:70–73.
oekkoek SK, Hulscher HC, Dortland BR, Hensbroek RA, Elgersma Y,Ruigrok TJ, De Zeeuw CI (2003) Cerebellar LTD and learning-dependent timing of conditioned eyelid responses. Science 301:1736–1739.
ondo S, Marty A (1998) Synaptic currents at individual connections
among stellate cells in rat cerebellar slices. J Physiol 509:221–232.ang EJ, Sugihara I, Llinás R (1996) GABAergic modulation of com-plex spike activity by the cerebellar nucleoolivary pathway in rat.J Neurophysiol 76:255–275.
ang EJ, Sugihara I, Welsh JP, Llinás RR (1999) Patterns of sponta-neous Purkinje cell complex spike activity in the awake rat. J Neu-rosci 19:2728–2739.
isberger SG (1998) Cerebellar LTD: a molecular mechanism of be-havioral learning? Cell 92:701–704.
itvin O, Tiunova A, Connell-Alberts Y, Panchin Y, Baranova A (2006)What is hidden in the pannexin treasure trove: the sneak peek andthe guesswork. J Cell Mol Med 10:613–634.
linás R, Baker R, Sotelo C (1974) Electronic coupling between neu-rons in cat inferior olive. J Neurophysiol 37:560–571.
linás RR (1988) The intrinsic electrophysiological properties of mam-malian neurons: insights into central nervous system function.Science 242:1654–1664.
linás RR, Moreno H (1998) Local Ca2� signaling in neurons. CellCalcium 24:359–366.
oewenstein Y, Mahon S, Chadderton P, Kitamura K, Sompolinsky H,Yarom Y, Häusser M (2005) Bistability of cerebellar Purkinje cellsmodulated by sensory stimulation. Nat Neurosci 8:202–211.
ameli M, Zamudio PA, Carta M, Valenzuela CF (2005) Developmen-tally regulated actions of alcohol on hippocampal glutamatergictransmission. J Neurosci 25:8027–8036.
ann-Metzer P, Yarom Y (1999) Electrotonic coupling interacts withintrinsic properties to generate synchronized activity in cerebellarnetworks of inhibitory interneurons. J Neurosci 19:3298–3306.
arini AM, Strauss KI, Jacobowitz DM (1997) Calretinin-containingneurons in rat cerebellar granule cell cultures. Brain Res Bull42(4):279–288.
auk MD, Medina JF, Nores WL, Ohyama T (2000) Cerebellarfunction: coordination, learning or timing? Curr Biol 10:522–525.
cKay BE, Engbers JD, Mehaffey WH, Gordon GR, Molineux ML,Bains JS, Turner RW (2007) Climbing fiber discharge regulatescerebellar functions by controlling the intrinsic characteristics ofpurkinje cell output. J Neurophysiol 97:2590–2604.
edina JF, Garcia KS, Nores WL, Taylor NM, Mauk MD (2000) Timingmechanisms in the cerebellum: testing predictions of a large-scalecomputer simulation. J Neurosci 20:5516–5525.
edina JF, Lisberger SG (2007) Variation, signal, and noise in cere-bellar sensory-motor processing for smooth-pursuit eye move-ments. J Neurosci 27:6832–6842.
edina JF, Nores WL, Mauk MD (2002) Inhibition of climbing fibres isa signal for the extinction of conditioned eyelid responses. Nature416:330–333.
eier C, Petrasch-Parwez E, Habbes HW, Teubner B, Guldenagel M,Degen J, Sohl G, Willecke K, Dermietzel R (2002) Immunohisto-chemical detection of the neuronal connexin36 in the mouse cen-tral nervous system in comparison to connexin36-deficient tissues.Histochem Cell Biol 117:461–471.
elloni L, Molina C, Pena M, Torres D, Singer W, Rodriguez E (2007)Synchronization of neural activity across cortical areas correlateswith conscious perception. J Neurosci 27:2858–2865.
iall RC, Keating JG, Malkmus M, Thach WT (1998) Simple spikeactivity predicts occurrence of complex spikes in cerebellar Pur-kinje cells. Nat Neurosci 1:13–15.
iles FA, Lisberger SG (1981) Plasticity in the vestibulo-ocular reflex:a new hypothesis. Annu Rev Neurosci 4:273–299.
ittmann W, Koch U, Häusser M (2005) Feed-forward inhibitionshapes the spike output of cerebellar Purkinje cells. J Physiol563:369–378.
ittmann W, Häusser M (2007) Linking synaptic plasticity and spikeoutput at excitatory and inhibitory synapses onto cerebellar Pur-kinje cells. J Neurosci 27:5559–5570.
iura K, Kishino T, Li E, Webber H, Dikkes P, Holmes GL, Wagstaff J(2002) Neurobehavioral and electroencephalographic abnormali-
ties in Ube3a maternal-deficient mice. Neurobiol Dis 9:149–159.
M
M
M
N
O
O
O
P
P
P
P
R
R
R
R
R
R
R
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
G. Cheron et al. / Neuroscience 153 (2008) 1–1918
oortgat KT, Bullock TH, Sejnowski TJ (2000) Gap junction effects onprecision and frequency of a model pacemaker network. J Neuro-physiol 83:984–997.
ugnaini E, Floris A (1994) The unipolar brush cell: a neglectedneuron of the mammalian cerebellar cortex. J Comp Neurol339:174–180.
urthy VN, Fetz EE (1996) Oscillatory activity in sensorimotor cortexof awake monkeys: synchronization of local field potentials andrelation to behavior. J Neurophysiol 76:3949–3967.
elson AB, Krispel CM, Sekirnjak C, du Lac S (2003) Long-lastingincreases in intrinsic excitability triggered by inhibition. Neuron40:609–620.
dermatt B, Wellershaus K, Wallraff A, Seifert G, Degen J, Euwens C,Fuss B, Bussow H, Schilling K, Steinhauser C, Willecke K (2003)Connexin 47 (Cx47)-deficient mice with enhanced green fluores-cent protein reporter gene reveal predominant oligodendrocyticexpression of Cx47 and display vacuolized myelin in the CNS.J Neurosci 23:4549–4559.
ffenhäuser N, Castelletti D, Mapelli L, Soppo BE, Regondi MC, RossiP, D’Angelo E, Frassoni C, Amadeo A, Tocchetti A, Pozzi B,Disanza A, Guarnieri D, Betsholtz C, Scita G, Heberlein U, Di FiorePP (2006) Increased ethanol resistance and consumption in Eps8knockout mice correlates with altered actin dynamics. Cell127:213–226.
scarsson O (1979) Functional units of the cerebellum. Sagittal zonesand microzones. Trends Neurosci 2:143–145.
ellerin JP, Lamarre Y (1997) Local field potential oscillations inprimate cerebellar cortex during voluntary movement. J Neuro-physiol 78:3502–3507.
esaran B, Pezaris JS, Sahani M, Mitra PP, Andersen RA (2002)Temporal structure in neuronal activity during working memory inmacaque parietal cortex. Nat Neurosci 5:805–811.
iochon C, Irinopoulou T, Brusciano D, Bailly Y, Mariani J, Levenes C(2007) NMDA receptor contribution to the climbing fiber responsein the adult mouse Purkinje cell. J Neurosci 27:10797–10809.
uia G, Costa E, Vicini S (1994) Functional diversity of GABA-acti-vated Cl-currents in Purkinje versus granule neurons in rat cere-bellar slices. Neuron 12:117–126.
aman IM, Bean BP (1997) Resurgent sodium current and actionpotential formation in dissociated cerebellar Purkinje neurons.J Neurosci 17:4517–4526.
aman IM, Bean BP (1999) Ionic currents underlying spontaneousaction potentials in isolated cerebellar Purkinje neurons. J Neuro-sci 19:1663–1674.
ash JE, Davidson KG, Kamasawa N, Yasumura T, Kamasawa M,Zhang C, Michaels R, Restrepo D, Ottersen OP, Olson CO, NagyJI (2005) Ultrastructural localization of connexins (Cx36, Cx43,Cx45), glutamate receptors and aquaporin-4 in rodent olfactorymucosa, olfactory nerve and olfactory bulb. J Neurocytol 34:307–341.
ay A, Zoidl G, Wahle P, Dermietzel R (2006) Pannexin expression inthe cerebellum. Cerebellum 5(3):189–192.
aymond JL, Lisberger SG, Mauk MD (1996) The cerebellum: aneuronal learning machine? Science 272:1126–1131.
ésibois A, Rogers JH (1992) Calretinin in rat brain: an immunohis-tochemical study Neuroscience 46:101–134.
ossi DJ, Alford S, Mugnaini E, Slater NT (1995) Properties of trans-mission at a giant glutamatergic synapse in cerebellum: the mossyfiber-unipolar brush cell synapse. J Neurophysiol 74:24–42.
alinas E, Sejnowski TJ (2001) Correlated neuronal activity and theflow of neural information. Nat Rev Neurosci 2:539–550.
anchez-Campusano R, Gruart A, Delgado-Garcia JM (2007) Thecerebellar interpositus nucleus and the dynamic control of learnedmotor responses. J Neurosci 27:6620–6632.
chiffmann SN, Cheron G, Lohof A, d’Alcantara P, Meyer M, Parmen-tier M, Schurmans S (1999) Impaired motor coordination and Pur-kinje cell excitability in mice lacking calretinin. Proc Natl Acad Sci
U S A 96:5257–5262.chmahmann JD (2004) Disorders of the cerebellum: ataxia, dysme-tria of thought, and the cerebellar cognitive affective syndrome.J Neuropsychiatry Clin Neurosci 16:367–378.
chmahmann JD, Caplan D (2006) Cognition, emotion and the cere-bellum. Brain 129:290–292.
chmahmann JD, Sherman JC (1998) The cerebellar cognitive affec-tive syndrome. Brain 121:561–579.
chmidt H, Arendt O, Brown EB, Schwaller B, Eilers J (2007) Parval-bumin is freely mobile in axons, somata and nuclei of cerebellarPurkinje neurones. J Neurochem 100:727–735.
chmitz D, Schuchmann S, Fisahn A, Draguhn A, Buhl EH, Petrasch-Parwez E, Dermietzel R, Heinemann U, Traub RD (2001) Axo-axonal coupling. A novel mechanism for ultrafast neuronal com-munication. Neuron 31:831–840.
chmidt H, Schwaller B, Eilers J (2005) Calbindin D28k targets myo-inositol monophosphatase in spines and dendrites of cerebellarPurkinje neurons. Proc Natl Acad Sci U S A 102:5850–5855.
chonewille M, Luo C, Ruigrok TJ, Voogd J, Schmolesky MT, Rutte-man M, Hoebeek FE, De Jeu MT, De Zeeuw CI (2006) Zonalorganization of the mouse flocculus: physiology, input, and output.J Comp Neurol 497:670–682.
chmoleski MT, Weber JT, De Zeeuw CI, Hansel C (2002) Themaking of a complex spike: ionic composition and plasticity. Ann NY Acad Sci 978:359–390.
churmans S, Schiffmann SN, Gurden H, Lemaire M, Lipp HP,Schwam V, Pochet R, Imperato A, Bohme GA, Parmentier M(1997) Impaired long-term potentiation induction in dentate gyrusof calretinin-deficient mice. Proc Natl Acad Sci U S A 94:10415–10420.
chwaller B, Meyer M, Schiffmann S (2002) “New” functions for “old”proteins: the role of the calcium-binding proteins calbindin D-28k,calretinin and parvalbumin, in cerebellar physiology. Studies withknockout mice. Cerebellum 1:241–258.
chwarz C, Welsh JP (2001) Dynamic modulation of mossy fibersystem throughput by inferior olive synchrony: a multielectrodestudy of cerebellar cortex activated by motor cortex. J Neuro-physiol 86:2489–2504.
ervais L, Bearzatto B, Hourez R, Dan B, Schiffmann SN, Cheron G(2004) Effect of simple spike firing mode on complex spike firingrate and waveform in cerebellar Purkinje cells in non-anesthetizedmice. Neurosci Lett 367:171–176.
ervais L, Bearzatto B, Schwaller B, Dumont M, De Saedeleer C, DanB, Barski JJ, Schiffmann SN, Cheron G (2005) Mono- and dual-frequency fast cerebellar oscillation in mice lacking parvalbuminand/or calbindin D-28k. Eur J Neurosci 22:861–870.
ervais L, Cheron G (2005) Purkinje cell rhythmicity and synchronicityduring modulation of fast cerebellar oscillation. Neuroscience134:1247–1259.
ervais L, Hourez R, Bearzatto B, Gall D, Schiffmann SN, Cheron G(2007) Purkinje cell dysfunction and alteration of long-term synap-tic plasticity in fetal alcohol syndrome. Proc Natl Acad Sci U S A104:9858–9863.
imburger E, Stang A, Kremer M, Dermietzel R (1997) Expression ofconnexin43 mRNA in adult rodent brain. Histochem Cell Biol107:127–137.
inai A, Bowers CW, Crainiceanu CM, Boatman D, Gordon B, LesserRP, Lenz FA, Crone NE (2005) Electrocorticographic high gammaactivity versus electrical cortical stimulation mapping of naming.Brain 128:1556–1570.
mith DJ, Azzaro AJ, Zaldivar SB, Palmer S, Lee HS (1981) Proper-ties of the optical isomers and metabolites of ketamine on the highaffinity transport and catabolism of monoamines. Neuropharmacol-ogy 120:391–396.
mith DJ, Pekoe GM, Martin LL, Coalgate B (1980) The interaction ofketamine with the opiate receptor. Life Sci 26:789–795.
otelo C, Llinás R (1972) Specialized membrane junctions between
neurons in the vertebrate cerebellar cortex. J Cell Biol 53:271–289.
S
S
S
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
U
U
v
V
W
W
W
W
W
W
W
W
W
Y
Y
Y
G. Cheron et al. / Neuroscience 153 (2008) 1–19 19
pencer RM, Zelaznik HN, Diedrichsen J, Ivry RB (2003) Disruptedtiming of discontinuous but not continuous movements by cerebel-lar lesions. Science 300:1437–1439.
trata P, Rossi F (1998) Plasticity of the olivocerebellar pathway.Trends Neurosci 21:407–413.
uter KJ, Jaeger D (2004) Reliable control of spike rate and spiketiming by rapid input transients in cerebellar stellate cells. Neuro-science 124:305–317.
akechi H, Eilers J, Konnerth A (1998) A new class of synapticresponse involving calcium release in dendritic spines. Nature396:757–760.
hach WT (1968) Discharge of Purkinje and cerebellar nuclear neu-rons during rapidly alternating arm movements in the monkey.J Neurophysiol 31:785–797.
hach WT (2007) On the mechanism of cerebellar contributions tocognition. Cerebellum 6:163–167.
hach WT, Goodkin HP, Keating JG (1992) The cerebellum and theadaptive coordination of movement. Annu Rev Neurosci 15:403–442.
hompson CL, Drewery DL, Atkins HD, Stephenson FA, Chazot PL(2000) Immunohistochemical localization of N-methyl-D-aspartatereceptor NR1, NR2A, NR2B and NR2C/D subunits in the adultmammalian cerebellum. Neurosci Lett 283:85–88.
hompson RF, Krupa DJ (1994) Organization of memory traces in themammalian brain. Annu Rev Neurosci 17:519–549.
eubner B, Degen J, Sohl G, Guldenagel M, Bukauskas FF, TrexlerEB, Verselis VK, De Zeeuw CI, Lee CG, Kozak CA, Petrasch-Parwez E, Dermietzel R, Willecke K (2000) Functional expressionof the murine connexin 36 gene coding for a neuron-specific gapjunctional protein. J Membr Biol 176:249–262.
eubner B, Odermatt B, Guldenagel M, Sohl G, Degen J, BukauskasF, Kronengold J, Verselis VK, Jung YT, Kozak CA, Schilling K,Willecke K (2001) Functional expression of the new gap junctiongene connexin47 transcribed in mouse brain and spinal cord neu-rons. J Neurosci 21:1117–1126.
raub RD (2003) Fast oscillations and epilepsy. Epilepsy Curr 3:77–79.
raub RD, Bibbig A (2000) A model of high-frequency ripples in thehippocampus based on synaptic coupling plus axon-axon gapjunctions between pyramidal neurons. J Neurosci 20:2086–2093.
raub RD, Cunningham MO, Gloveli T, LeBeau FE, Bibbig A, Buhl EH,Whittington MA (2003a) GABA-enhanced collective behavior inneuronal axons underlies persistent gamma-frequency oscilla-tions. Proc Natl Acad Sci U S A 100:11047–11052.
raub RD, Pais I, Bibbig A, LeBeau FE, Buhl EH, Monyer H, HormuzdiSG, Whittington MA (2003b) Contrasting roles of axonal (pyramidalcell) and dendritic (interneuron) electrical coupling in the genera-tion of neuronal network oscillations. Proc Natl Acad Sci U S A100:1370–1374.
raub RD, Draguhn A, Whittington MA, Baldeweg T, Bibbig A, BumhlEH, Schmitz D (2002) Axonal gap junctions between principalneurons: a novel source of network oscillations, and perhaps epi-leptogenesis. Rev Neurosci 13:1–30.
raub RD, Middleton SJ, Knopfel T, Whittington MA (2007) Very fast(�100 Hz) low-calcium induced synchronized oscillations,amongst Purkinje neurons in vitro, are replicated with a networkmodel based on electrical coupling via gap junctions betweenaxons. Soc Neurosci Abstr 793.10/L23.
raub RD, Schmitz D, Jefferys JG, Draguhn A (1999) High-frequency
population oscillations are predicted to occur in hippocampal py-ramidal neuronal networks interconnected by axoaxonal gapjunctions. Neuroscience 92:407–426.
hlhaas PJ, Singer W (2006) Neural synchrony in brain disorders:relevance for cognitive dysfunctions and pathophysiology. Neuron52:155–168.
sowicz MM, Sugimori M, Cherksey B, Llinás R (1992) P-type calciumchannels in the somata and dendrites of adult cerebellar Purkinjecells. Neuron 9:1185–1199.
an Woerden GM, Harris KD, Hojjati MR, Gustin RM, Qiu S, de AvilaFreire R, Jiang YH, Elgersma Y, Weeber EJ (2007) Rescue ofneurological deficits in a mouse model for Angelman syndrome byreduction of alphaCaMKII inhibitory phosphorylation. Nat Neurosci10:280–282.
oogd J, Glickstein M (1998) The anatomy of the cerebellum. TrendsNeurosci 21:370–375.
alter JT, Alviña K, Womack MD, Chevez C, Khodakhah K (2006)Decreases in the precision of Purkinje cell pacemaking causecerebellar dysfunction and ataxia. Nat Neurosci 9:389–397.
ang SS, Denk W, Häusser M (2000) Coincidence detection in singledendritic spines mediated by calcium release. Nat Neurosci12:1266–1273.
eber JT, De Zeeuw CI, Linden DJ, Hansel C (2003) Long-termdepression of climbing fiber-evoked calcium transients in Purkinjecell dendrites. Proc Natl Acad Sci U S A 100:2878–2883.
eeber EJ, Jiang YH, Elgersma Y, Varga AW, Carrasquillo Y, Brown SE,Christian JM, Mirnikjoo B, Silva A, Beaudet AL, Sweatt JD (2003)Derangements of hippocampal calcium/calmodulin-dependent pro-tein kinase II in a mouse model for Angelman mental retardationsyndrome. J Neurosci 23:2634–2644.
elsh JP, Lang EJ, Suglhara I, Llinás R (1995) Dynamic organizationof motor control within the olivocerebellar system. Nature374:453–457.
elsh JP, Yamaguchi H, Zeng XH, Kojo M, Nakada Y, Takagi A,Sugimori M, Llinás RR (2005) Normal motor learning during phar-macological prevention of Purkinje cell long-term depression. ProcNatl Acad Sci U S A 102:17166–17171.
hittington MA, Traub RD, Jefferys JG (1995) Synchronized oscilla-tions in interneuron networks driven by metabotropic glutamatereceptor activation. Nature 373:612–615.
omack MD, Khodakhah K (2002) Active contribution of dendrites tothe tonic and trimodal patterns of activity in cerebellar Purkinjeneurons. J Neurosci 22:10603–10612.
omelsdorf T, Schoffelen JM, Oostenveld R, Singer W, Desimone R,Engel AK, Fries P (2007) Modulation of neuronal interactionsthrough neuronal synchronization. Science 316:1609–1612.
abe I, Sasaki H, Chen DH, Raskind WH, Bird TD, Yamashita I, TsujiS, Kikuchi S, Tashiro K (2003) Spinocerebellar ataxia type 14caused by a mutation in protein kinase C gamma. Arch Neurol60:1749–1751.
oshida T, Funabiki K, Hirano T (2007) Increased occurrence ofclimbing fiber inputs to the cerebellar flocculus in a mutant mouseis correlated with the timing delay of optokinetic response. EurJ Neurosci 25:1467–1474.
uan Q, Qiu DL, Weber JT, Hansel C, Knöpfel T (2007) Climbingfiber-triggered metabotropic slow potentials enhance dendritic cal-cium transients and simple spike firing in cerebellar Purkinje cells.
Mol Cell Neurosci 35:596–603.(Accepted 25 January 2008)(Available online 19 February 2008)