caenorhabditis elegans survives atmospheric breakup of sts107, space shuttle columbia
TRANSCRIPT

ASTROBIOLOGYVolume 5, Number 6, 2005© Mary Ann Liebert, Inc.
Research Paper
Caenorhabditis elegans Survives Atmospheric Breakup ofSTS-107, Space Shuttle Columbia
NATHANIEL J. SZEWCZYK,1,* ROCCO L. MANCINELLI,1 WILLIAM MCLAMB,2,†
DAVID REED,2 BARUCH S. BLUMBERG,3 and CATHARINE A. CONLEY1
ABSTRACT
The nematode Caenorhabditis elegans, a popular organism for biological studies, is being de-veloped as a model system for space biology. The chemically defined liquid medium, C. el-egans Maintenance Medium (CeMM), allows axenic cultivation and automation of experi-ments that are critical for spaceflight research. To validate CeMM for use during spaceflight,we grew animals using CeMM and standard laboratory conditions onboard STS-107, spaceshuttle Columbia. Tragically, the Columbia was destroyed while reentering the Earth’s at-mosphere. During the massive recovery effort, hardware that contained our experiment wasfound. Live animals were observed in four of the five recovered canisters, which had sur-vived on both types of media. These data demonstrate that CeMM is capable of supportingC. elegans during spaceflight. They also demonstrate that animals can survive a relatively unprotected reentry into the Earth’s atmosphere, which has implications with regard to thepackaging of living material during space flight, planetary protection, and the interplanetarytransfer of life. Key Words: Caenorhabditis elegans—Nematode—Spaceflight research—Atmospheric reentry—Space shuttle Columbia. Astrobiology 5, 690–705.
690
INTRODUCTION
JOHNSON AND NELSON (1991) suggested thatCaenorhabditis elegans could serve as a model
for biological studies in space and be particu-larly relevant for studies in development, ra-diobiology, and aging. Three spaceflight exper-iments involving live C. elegans flew prior to the experiment reported here. On STS-42, C. el-
egans completed two life cycles in space with-out gross developmental abnormalities whengrown on standard laboratory media, and mating was successful (Nelson et al., 1994a,b).On STS-76, an observed increase in the rate of mutation was shown to be the direct effect of cosmic radiation (Hartman et al., 2001). OnSTS-95, both the flown and ground control an-imals died, probably from anoxia, and no re-
1NASA Ames Research Center, Moffett Field, California.2Bionetics Corporation, Kennedy Space Center, Florida.3Fox Chase Cancer Center, Philadelphia, Pennsylvania.*Present address: Department of Biological Sciences, University of Pittsburgh, Pittsburgh, Pennsylvania.†Present address: Dynamac Corporation, Kennedy Space Center, Florida.
5996_02_p690-705 12/20/05 4:03 PM Page 690

sults were obtained (E. Moss, personal commu-nication).
In the laboratory, C. elegans is cultured on agarplates using Escherichia coli as a food source [Nematode Growth Medium (NGM)] (Brenner,1974). Animals grown on NGM require weeklytransfers to new plates, which renders NGM cul-ture suboptimal for studies in space because as-tronaut time is limited. On NGM, as in the wild,animals are covered in a thin film of water andthus exposed to surface tension (Viglierchio,1992). The force exerted by surface tensionaround a cylinder with a diameter equal to thatof an adult worm (100 �m) can be calculated tobe approximately 10,000 times the force of grav-ity on Earth. Surface tension forces are appliedonly to the external surface cuticle of the animals,while gravity causes sedimentation of internalmaterials, both tissues and nutrients, in a density-dependent manner. In space, the force of surfacetension on C. elegans may mask effects inducedby the loss of gravity (microgravity). To increasethe possibility of automation and minimize theinfluence of surface tension during spaceflight,growth under the surface of liquid media ishighly desirable.
Recently, a chemically defined axenic mediumfor cultivation of C. elegans has been established.C. elegans Maintenance Medium (CeMM) (Lu andGoetsch,1993) is usable in liquid and solid forms,and animals can be maintained in a reproductivestate considerably longer than on NGM (Szew-czyk et al., 2003). Use of CeMM allows automa-tion of culturing and experimentation, as well asdirect testing of the hypothesis that surface ten-sion masks the effects of microgravity on C. ele-gans. As a first step toward these goals, it is im-portant to establish that CeMM supports growthand reproduction in the spaceflight environment.This test is essential because living systems exposed to spaceflight experience metabolicchanges (Oser and Battrick, 1989), and all exist-ing data that show that C. elegans can undergomultiple generations during spaceflight werebased upon growth on a medium similar to NGM(Nelson et al., 1994a,b). The Conley laboratorywas selected to perform comparative tests be-tween CeMM and NGM in December 2002, on-board space shuttle Columbia during missionSTS-107.
During the course of our experiment, spaceshuttle Columbia was tragically destroyed as shereentered the Earth’s atmosphere for landing. De-
bris was recovered during an intense and exten-sive search (Columbia Accident InvestigationBoard [CAIB], 2003). Among the recovered itemswere five of the six canisters that contained thespaceflight portion of this experiment. Data ob-tained from the recovered canisters demonstratethat CeMM is capable of supporting C. elegansduring spaceflight, which suggests that CeMMwill be useful for biological studies of C. elegansin space. Of broader interest and perhaps of moresignificance, C. elegans survived reentry into theEarth’s atmosphere relatively unprotected, as thecanisters that housed the animals exited thespacecraft at a height of 42–32 km above theEarth’s surface and at velocities of 660–1,050km/h (CAIB, 2003). This discovery has significantimplications for the packaging of living materialduring space flight, planetary protection, and theinterplanetary transfer of life.
MATERIALS AND METHODS
Nematode handling
Standard procedures were utilized for nema-tode handling. Wild-type C. elegans strain N2 wasmaintained on NGM (Brenner, 1974) using UV-killed E. coli strain OP50 as a food source (Gemsand Riddle, 2000). Wild-type C. elegans strain CC1was created by growing N2 animals in CeMM (Luand Goetsch, 1993) for several years (Szewczyk etal., 2003), and was maintained on 1.7% agar platescontaining CeMM obtained on contract fromFisher Scientific (Santa Clara, CA). As describedpreviously, age-synchronized first-stage larval(L1) animals were obtained by settling culturesunder gravity (Zdinak et al., 1997), growth rateswere determined by measuring and averagingthe lengths of 10 developing animals (Bolanowskiet al., 1981), and brood sizes were determined bymeasuring and averaging the number of eggs laidby 20 animals (Byerly et al., 1976).
Animals flown on STS-107 Columbia
Wild-type N2 animals grown on NGM andCC1 animals grown on CeMM were flown in sixanodized aluminum BRIC-60 canisters on boardColumbia as shown in Fig. 1 and detailed in Table1. BRIC-60 canisters are ventilated with four �2-mm-diameter holes machined into their lids,which are reinforced in such a way that the tubelength is slightly less than 1 cm. Clear polystyrene
C. ELEGANS SURVIVES COLUMBIA BREAKUP 691
5996_02_p690-705 12/20/05 4:03 PM Page 691
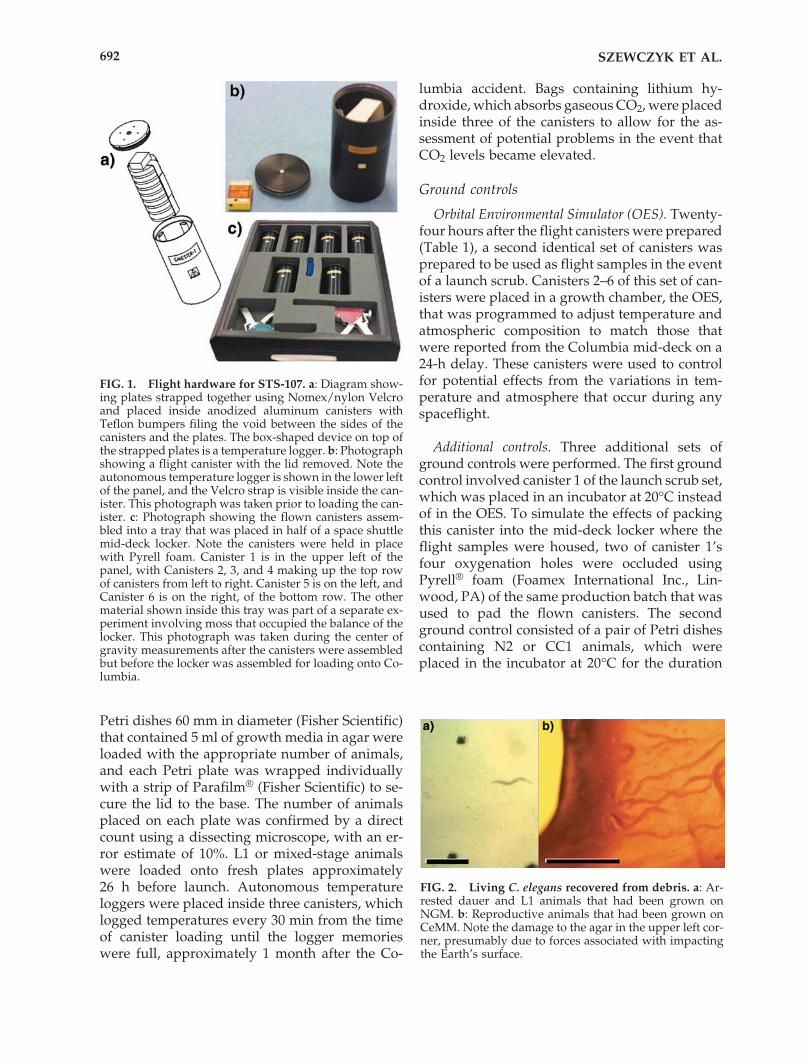
Petri dishes 60 mm in diameter (Fisher Scientific)that contained 5 ml of growth media in agar wereloaded with the appropriate number of animals,and each Petri plate was wrapped individuallywith a strip of Parafilm® (Fisher Scientific) to se-cure the lid to the base. The number of animalsplaced on each plate was confirmed by a directcount using a dissecting microscope, with an er-ror estimate of 10%. L1 or mixed-stage animalswere loaded onto fresh plates approximately 26 h before launch. Autonomous temperatureloggers were placed inside three canisters, whichlogged temperatures every 30 min from the timeof canister loading until the logger memorieswere full, approximately 1 month after the Co-
lumbia accident. Bags containing lithium hy-droxide, which absorbs gaseous CO2, were placedinside three of the canisters to allow for the as-sessment of potential problems in the event thatCO2 levels became elevated.
Ground controls
Orbital Environmental Simulator (OES). Twenty-four hours after the flight canisters were prepared(Table 1), a second identical set of canisters wasprepared to be used as flight samples in the eventof a launch scrub. Canisters 2–6 of this set of can-isters were placed in a growth chamber, the OES,that was programmed to adjust temperature andatmospheric composition to match those thatwere reported from the Columbia mid-deck on a24-h delay. These canisters were used to controlfor potential effects from the variations in tem-perature and atmosphere that occur during anyspaceflight.
Additional controls. Three additional sets ofground controls were performed. The first groundcontrol involved canister 1 of the launch scrub set,which was placed in an incubator at 20°C insteadof in the OES. To simulate the effects of packingthis canister into the mid-deck locker where theflight samples were housed, two of canister 1’sfour oxygenation holes were occluded usingPyrell® foam (Foamex International Inc., Lin-wood, PA) of the same production batch that wasused to pad the flown canisters. The secondground control consisted of a pair of Petri dishescontaining N2 or CC1 animals, which wereplaced in the incubator at 20°C for the duration
SZEWCZYK ET AL.692
FIG. 1. Flight hardware for STS-107. a: Diagram show-ing plates strapped together using Nomex/nylon Velcroand placed inside anodized aluminum canisters withTeflon bumpers filing the void between the sides of thecanisters and the plates. The box-shaped device on top ofthe strapped plates is a temperature logger. b: Photographshowing a flight canister with the lid removed. Note theautonomous temperature logger is shown in the lower leftof the panel, and the Velcro strap is visible inside the can-ister. This photograph was taken prior to loading the can-ister. c: Photograph showing the flown canisters assem-bled into a tray that was placed in half of a space shuttlemid-deck locker. Note the canisters were held in placewith Pyrell foam. Canister 1 is in the upper left of thepanel, with Canisters 2, 3, and 4 making up the top rowof canisters from left to right. Canister 5 is on the left, andCanister 6 is on the right, of the bottom row. The othermaterial shown inside this tray was part of a separate ex-periment involving moss that occupied the balance of thelocker. This photograph was taken during the center ofgravity measurements after the canisters were assembledbut before the locker was assembled for loading onto Co-lumbia.
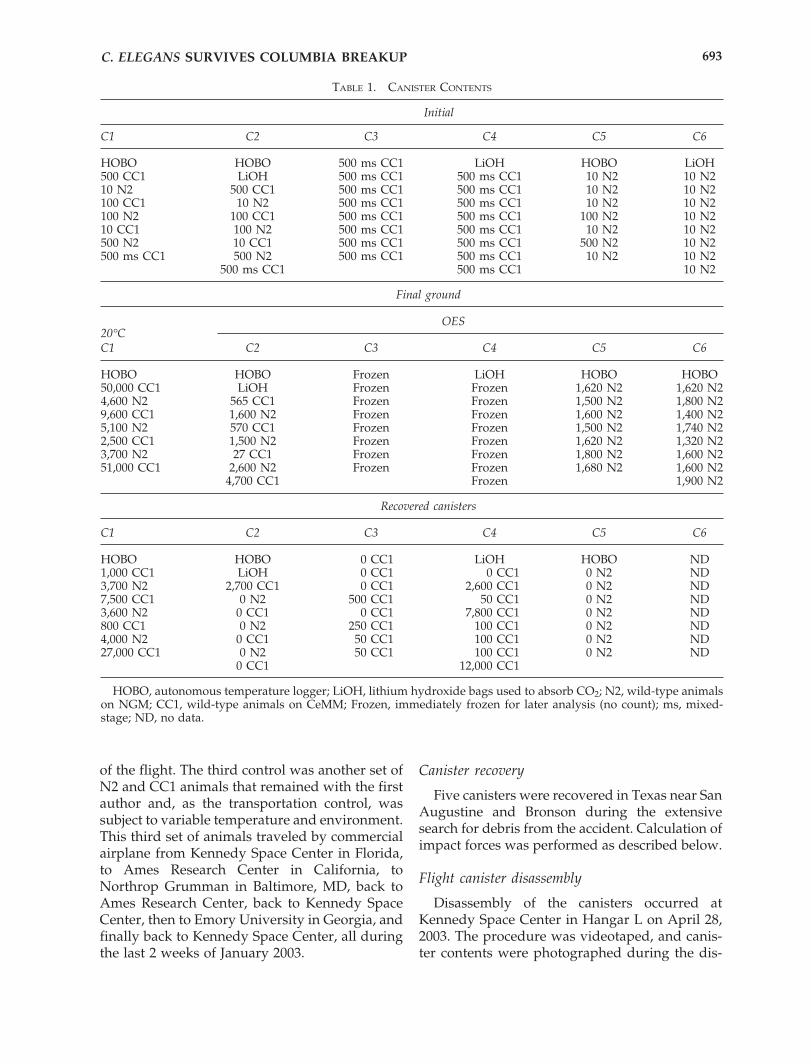
FIG. 2. Living C. elegans recovered from debris. a: Ar-rested dauer and L1 animals that had been grown onNGM. b: Reproductive animals that had been grown onCeMM. Note the damage to the agar in the upper left cor-ner, presumably due to forces associated with impactingthe Earth’s surface.
5996_02_p690-705 12/20/05 4:03 PM Page 692
of the flight. The third control was another set ofN2 and CC1 animals that remained with the firstauthor and, as the transportation control, wassubject to variable temperature and environment.This third set of animals traveled by commercialairplane from Kennedy Space Center in Florida,to Ames Research Center in California, toNorthrop Grumman in Baltimore, MD, back toAmes Research Center, back to Kennedy SpaceCenter, then to Emory University in Georgia, andfinally back to Kennedy Space Center, all duringthe last 2 weeks of January 2003.
Canister recovery
Five canisters were recovered in Texas near SanAugustine and Bronson during the extensivesearch for debris from the accident. Calculation ofimpact forces was performed as described below.
Flight canister disassembly
Disassembly of the canisters occurred atKennedy Space Center in Hangar L on April 28,2003. The procedure was videotaped, and canis-ter contents were photographed during the dis-
C. ELEGANS SURVIVES COLUMBIA BREAKUP 693
TABLE 1. CANISTER CONTENTS
Initial
C1 C2 C3 C4 C5 C6
HOBO HOBO 500 ms CC1 LiOH HOBO LiOH500 CC1 LiOH 500 ms CC1 500 ms CC1 10 N2 10 N210 N2 500 CC1 500 ms CC1 500 ms CC1 10 N2 10 N2100 CC1 10 N2 500 ms CC1 500 ms CC1 10 N2 10 N2100 N2 100 CC1 500 ms CC1 500 ms CC1 100 N2 10 N210 CC1 100 N2 500 ms CC1 500 ms CC1 10 N2 10 N2500 N2 10 CC1 500 ms CC1 500 ms CC1 500 N2 10 N2500 ms CC1 500 N2 500 ms CC1 500 ms CC1 10 N2 10 N2
500 ms CC1 500 ms CC1 10 N2
Final ground
20°COES
C1 C2 C3 C4 C5 C6
HOBO HOBO Frozen LiOH HOBO HOBO50,000 CC1 LiOH Frozen Frozen 1,620 N2 1,620 N24,600 N2 565 CC1 Frozen Frozen 1,500 N2 1,800 N29,600 CC1 1,600 N2 Frozen Frozen 1,600 N2 1,400 N25,100 N2 570 CC1 Frozen Frozen 1,500 N2 1,740 N22,500 CC1 1,500 N2 Frozen Frozen 1,620 N2 1,320 N23,700 N2 27 CC1 Frozen Frozen 1,800 N2 1,600 N251,000 CC1 2,600 N2 Frozen Frozen 1,680 N2 1,600 N2
4,700 CC1 Frozen 1,900 N2
Recovered canisters
C1 C2 C3 C4 C5 C6
HOBO HOBO 0 CC1 LiOH HOBO ND1,000 CC1 LiOH 0 CC1 12,000 CC1 0 N2 ND3,700 N2 2,700 CC1 0 CC1 2,600 CC1 0 N2 ND7,500 CC1 0 N2 500 CC1 12,050 CC1 0 N2 ND3,600 N2 0 CC1 0 CC1 7,800 CC1 0 N2 ND800 CC1 0 N2 250 CC1 12,100 CC1 0 N2 ND4,000 N2 0 CC1 50 CC1 12,100 CC1 0 N2 ND27,000 CC1 0 N2 50 CC1 12,100 CC1 0 N2 ND
0 CC1 12,000 CC1
HOBO, autonomous temperature logger; LiOH, lithium hydroxide bags used to absorb CO2; N2, wild-type animalson NGM; CC1, wild-type animals on CeMM; Frozen, immediately frozen for later analysis (no count); ms, mixed-stage; ND, no data.
5996_02_p690-705 12/20/05 4:03 PM Page 693

assembly process. After all canisters were disas-sembled, the Petri dishes were scored for live an-imals in the presence of NASA quality controltechnicians. Animals were photographed in theflown Petri dishes prior to opening the dishes for subculturing. Strain CC2 (flown N2) andstrain CC3 (flown CC1) were deposited with theCaenorhabditis Genetics Center (http://biosci.umn.edu/CGC/CGChomepage.htm).
Terminal velocity and impact for a BRIC-60 class canister
A full discussion of these calculations is givenin the Appendix. Briefly, our calculations suggestthat the terminal velocity for a BRIC-60 canister atsea level should be in the vicinity of 45 m/s, or 100miles/h. The atmosphere is much less dense at 61km, where disintegration of the vehicle occurred,but it is unknown at what altitude the canisterswere released from the locker in which they werecontained. Also, the canisters likely would haveexperienced significant angular acceleration due totumbling and potential impact with other debrisduring the descent, as the atmosphere thickenedand slowed their fall. The inertial acceleration athigh altitude is somewhat less than gravity at thesurface of the Earth, but steady state is assumedat low altitude as well. The calculated terminal ve-locity of 45 m/s is reasonable in comparison withterminal velocities reported for other objects.
To calculate deceleration, a deceleration timeor a deceleration distance must be estimated. Wechose a deceleration distance of 4 cm, approxi-
mately one-half the diameter of a BRIC, based onanecdotal descriptions that the canisters were em-bedded halfway in the ground before recovery(unfortunately, in situ images were not available).The time required to decelerate from 45 m/s to 0 m/s over 4 cm is 0.002 s, as calculated in theAppendix.
The values of 45 m/s for terminal velocity and0.002 s for deceleration time were used to calcu-late the acceleration in G:
acceleration in G � 45 m/s divided by 0.002 sdivided by 9.8 m/s/s, or 2,295 G
RESULTS
Flight samples in flight
Three hours after the recovered canisters wereopened, live C. elegans nematodes were pho-tographed (Fig. 2). The number of live animals ineach recovered canister was determined, with theresult that live animals were found in all canis-ters except canister 5 (Table 1). Live animals werefound on both NGM and CeMM plates, so sur-vival was not dependent on media type. Animalsgrown on CeMM were still reproductive, whileanimals on NGM were arrested as dauers or L1s(Fig. 2). Recultured animals from both media dis-played normal growth rates, normal brood sizes,and no heritable mutant phenotypes (Table 2).One Uncoordinated animal (Brenner, 1974) andone Egg Laying Defective animal (Trent et al.,
SZEWCZYK ET AL.694
TABLE 2. SURVIVING C. ELEGANS HAD NORMAL GROWTH RATES AND BROODSIZES
Rate of development (mm)
Growth rate, length (mm) Post-flight 25°C Unflown 25°C
On NGMAfter 1 day 0.398 � 0.06 0.38 � 0.03After 2 days 1.011 � 0.038 1.025 � 0.039
On CeMMAfter 1 day 0.264 � 0.037 0.251 � 0.019After 2 days 0.441 � 0.034 0.439 � 0.059After 3 days 0.633 � 0.032 0.634 � 0.036After 4 days 0.835 � 0.035 0.835 � 0.051After 5 days 1.008 � 0.041 1.003 � 0.024
Egg laying (number of eggs)
Post-flight 25°C Unflown 25°C
On NGM 152 � 22 147 � 34On CeMM 43 � 17 42 � 12
5996_02_p690-705 12/20/05 4:03 PM Page 694

1983) were initially subcloned, but neither pheno-type was transmitted. We expected to observe noheritable mutations in the number of animals ob-served, based on the previously determined mu-tation rate from STS-42 and STS-76 (Nelson et al.,1994b; Hartman et al., 2001). These data demon-strate that most of the live animals present in therecovered canisters were not dramatically alteredin heritable phenotype by the spaceflight, the reen-try accident, the post-accident exposure to theTexas environment, or recovery storage. However,their physiological responses to spaceflight couldnot be assessed because of the following con-founding factors.
Evaluation of inflight growth on NGM versusCeMM was complicated by several factors, in-cluding the long delay between reentry andanalysis and a contaminating mold that grewonly on CeMM plates. The N2 animals grown onNGM were expected to arrest during the flightbecause of exhaustion of food supply and werein fact arrested (Table 1). We cannot be certain,however, that this occurred inflight. The numberof N2 dauer animals recovered from flight canis-ter 1 was consistent with normal growth and ar-rest inflight, as expected from reported growthrates (Byerly et al., 1976; Szewczyk et al., 2003).
Back-extrapolation from the final number ofCC1 animals was more difficult. Dauers are moreresistant to a number of stresses, including tem-perature, than reproductive animals (Cassadaand Russell, 1975; Anderson, 1978). CC1 animalswere probably reproductive at the time of the ac-cident, so any stress associated with reentry couldhave affected CC1 more than N2. Upon recultur-ing the CeMM plates, it was evident that a moldhad contaminated some of the CeMM plates re-covered after the crash. The contaminant musthave gained access while the hardware was ex-posed to the Texas environment, since nothingsimilar was observed in any of the controls. Thecontaminating mold was capable of killing a largenumber of CC1 animals; however, the mold re-quired 3–4 weeks of growth at 25°C to kill an en-tire population. This temperature was not expe-rienced for at least the first month after impact.The presence of the mold, the accident itself, andthe long interval between impact and analysis(Table 3) preclude evaluating how well C. elegansgrew on CeMM in flight. The animals were keptalive on CeMM, but it is impossible to evaluatepopulation growth or dauer diapause in flight.These data are essential before CeMM can be fully
validated for automated spaceflight experimen-tation.
OES
Animals grown in the OES failed to reach population sizes consistent with predictions forgrowth at 25°C and with observed growth at 20°C(Table 1). Staged NGM- or CeMM-grown animalswere sterile adults, while the initially mixed-stageCeMM-grown animals were a mix of healthyadults and dauers. The autonomous temperaturerecorders inside the canisters and the tempera-ture recorder inside the OES registered tempera-tures in the 27°C range on the third and fourthday of the experiment. On NGM, growth of C. el-egans at high (27°C and higher) temperature re-sults in reduced fecundity and dauer formation(Byerly et al., 1976; Ailion and Thomas, 2000). Toconfirm that 27°C exposure was sufficient to in-duce sterility and dauer formation, we placedmixed-stage animals on both media types at 27°Cand subsequently observed sterile adults anddauers. Dauers recovered upon shift to 20°C.
The CO2 level rose throughout the experimentbut did not reach 0.5%. Inclusion of lithium hy-droxide in canisters in the OES did not have a sta-tistically significant effect on final population size(Table 1) and did not correlate with the numberof sterile or dauer animals. In a post-flight con-trol experiment, growth on NGM at 25°C with 5%CO2 did not result in sterile or dauer animals.However, we have no other data to suggestwhether the levels of CO2 experienced were detri-mental. C. elegans can recognize dissolved CO2 asa chemoattractant (Dusenbery, 1974), and move-ment is altered in the presence of 4% CO2 (Dusen-bery, 1985). To our knowledge, however, no other
C. ELEGANS SURVIVES COLUMBIA BREAKUP 695
TABLE 3. TIMELINE OF EVENTS
Date Event
January 15, 2003 Animals loaded into flighthardware
January 16, 2003 Columbia launchJanuary 26, 2003 On orbit video of canistersFebruary 01, 2003 Breakup of ColumbiaFebruary 07, 2003 Canister 2 recoveredFebruary 10, 2003 Canister 3 recoveredFebruary 14, 2003 Canister 4 recoveredFebruary 17, 2003 Canister 5 recoveredMarch 21, 2003 Canister 1 recoveredApril 24, 2003 Canister access granted by CAIBApril 28, 2003 Canisters opened
5996_02_p690-705 12/20/05 4:03 PM Page 695
reports describing the effect of elevated CO2 lev-els on C. elegans biology have been published.
The growth curves of animals in the OES canbe explained entirely by a temperature spikereplicated from reported cabin ambient early inthe flight. No temperature spike was noted by the internal recorders from our flown canisters,which could indicate that the thermal mass of ourlocker protected the flight samples from a spikein temperature at the location of the cabin sensorattached to the wall of the mid-deck. The num-ber of arrested N2 animals in the recovered flightcanister 1 was more similar to the number inground control 1, which remained at 20°C, thanin canister 2, which was in the OES (Table 1). Thisfinding demonstrates the need for careful evalu-ation with regard to temperatures used forground controls versus actual flight data, and theneed to use autonomous temperature loggersduring flight as opposed to relying solely on thecabin temperature recordings, which were usedto drive the OES.
Other ground controls
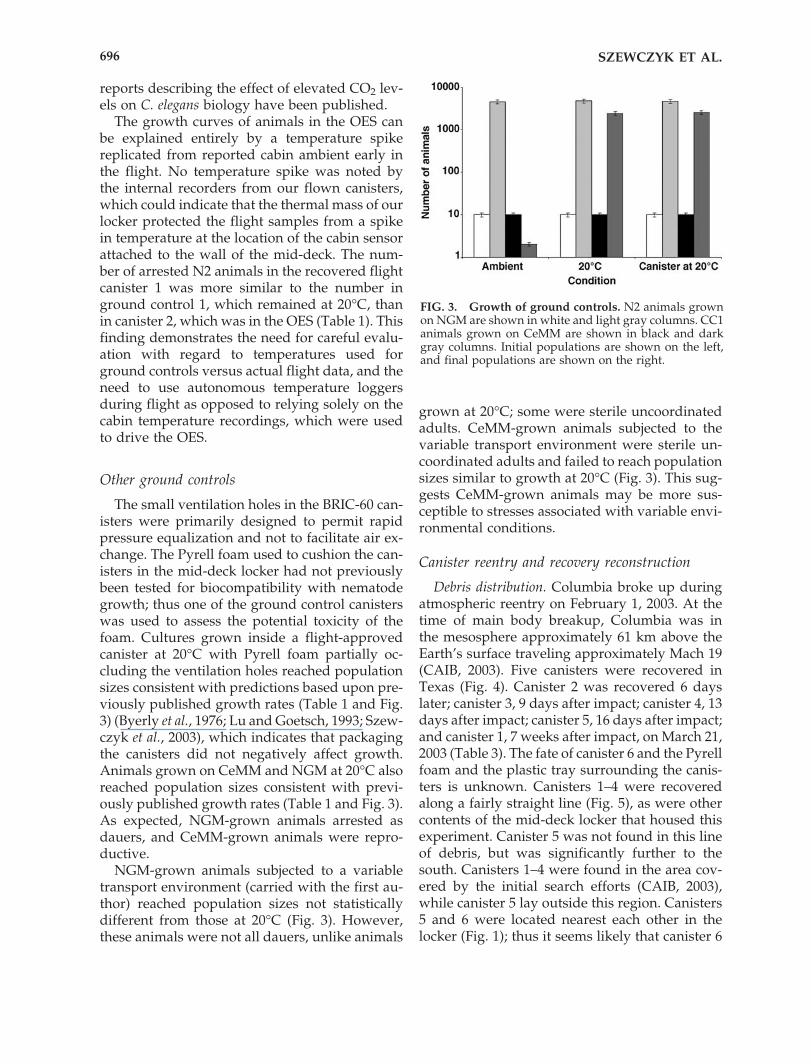
The small ventilation holes in the BRIC-60 can-isters were primarily designed to permit rapidpressure equalization and not to facilitate air ex-change. The Pyrell foam used to cushion the can-isters in the mid-deck locker had not previouslybeen tested for biocompatibility with nematodegrowth; thus one of the ground control canisterswas used to assess the potential toxicity of thefoam. Cultures grown inside a flight-approvedcanister at 20°C with Pyrell foam partially oc-cluding the ventilation holes reached populationsizes consistent with predictions based upon pre-viously published growth rates (Table 1 and Fig.3) (Byerly et al., 1976; Lu and Goetsch, 1993; Szew-czyk et al., 2003), which indicates that packagingthe canisters did not negatively affect growth.Animals grown on CeMM and NGM at 20°C alsoreached population sizes consistent with previ-ously published growth rates (Table 1 and Fig. 3).As expected, NGM-grown animals arrested asdauers, and CeMM-grown animals were repro-ductive.
NGM-grown animals subjected to a variabletransport environment (carried with the first au-thor) reached population sizes not statisticallydifferent from those at 20°C (Fig. 3). However,these animals were not all dauers, unlike animals
grown at 20°C; some were sterile uncoordinatedadults. CeMM-grown animals subjected to thevariable transport environment were sterile un-coordinated adults and failed to reach populationsizes similar to growth at 20°C (Fig. 3). This sug-gests CeMM-grown animals may be more sus-ceptible to stresses associated with variable envi-ronmental conditions.
Canister reentry and recovery reconstruction
Debris distribution. Columbia broke up duringatmospheric reentry on February 1, 2003. At thetime of main body breakup, Columbia was in the mesosphere approximately 61 km above theEarth’s surface traveling approximately Mach 19(CAIB, 2003). Five canisters were recovered inTexas (Fig. 4). Canister 2 was recovered 6 dayslater; canister 3, 9 days after impact; canister 4, 13days after impact; canister 5, 16 days after impact;and canister 1, 7 weeks after impact, on March 21,2003 (Table 3). The fate of canister 6 and the Pyrellfoam and the plastic tray surrounding the canis-ters is unknown. Canisters 1–4 were recoveredalong a fairly straight line (Fig. 5), as were othercontents of the mid-deck locker that housed thisexperiment. Canister 5 was not found in this lineof debris, but was significantly further to thesouth. Canisters 1–4 were found in the area cov-ered by the initial search efforts (CAIB, 2003),while canister 5 lay outside this region. Canisters5 and 6 were located nearest each other in thelocker (Fig. 1); thus it seems likely that canister 6
SZEWCZYK ET AL.696
FIG. 3. Growth of ground controls. N2 animals grownon NGM are shown in white and light gray columns. CC1animals grown on CeMM are shown in black and darkgray columns. Initial populations are shown on the left,and final populations are shown on the right.
5996_02_p690-705 12/20/05 4:03 PM Page 696

also fell outside the area in which other locker de-bris was found.
Evaluation of recovered crew module compo-nents allowed for an extensive trajectory recon-struction (CAIB, 2003). Breakup of the crew mod-ule appears to have been the result of ballisticheating and dynamic loading. Cabin items, suchas the tray housing the canisters, were releasedat heights of approximately 42–32 km and equiv-alent velocities of approximately 660–1,050 km/h.From the debris path (Fig. 5), we estimated theground order of canister impact, which correlatesweakly with animal survival. Canister 1 impactedfirst with live animals on all dishes, canister 3 im-pacted next and contained some dishes with liveand some dead animals, next was canister 4 withmost dishes containing live animals, followed bycanister 2 with only one dish of live animals, andcanister 5 with no dishes containing live animals.Fortuitous location within the spacecraft as wellas within individual debris may have been an im-portant factor with regard to survival. The cont-aminating mold provides an alternative explana-tion for the reduced survival in canisters 3 and 4,both of which contained only CC1 animals onCeMM plates.
Reentry heating. As shown in Fig. 4a and d–g,the recovered canisters displayed external dam-age but were structurally intact. Some loss of theanodized surface and external pitting of the alu-minum were present, with differing amounts ofdamage and in different locations. Pitting is con-sistent with the canisters falling at high velocitythrough other fragmented debris, though onecannot tell whether the canisters or the debriswere traveling faster at the time of each impact.Plasma etching of the external surface may haveoccurred from exposure at high velocity to hotgasses associated with the breakup. No evidenceexists to suggest that the canisters were impactedby large fragments of debris. Despite obvioussigns of heating, portions of Kapton® (E.I. DuPontde Nemours & Co., Wilmington, DE) and papertape labels on the exterior of the canisters werestill present (Fig. 4). Anodization loss appears tohave been an ablative process associated with de-scent and impact. Scorching alone could have dis-colored the anodization, as seen around the Kap-ton tape label of canister 1 (Fig. 4a).
The melting point of the aluminum canisters isin the range of 582–652°C. Given the external but
not internal heat damage to all of the recoveredcanisters, regions of the exterior surface but notthe interior of the canisters reached temperaturesin excess of 582°C. Although aluminum inside thecanisters was not melted, internal heating was ev-ident. The plastic temperature logger housings,made of acrylonitrile butadiene styrene (ABS),were partially melted, and the Teflon® (DuPont)bumpers and Velcro® (Velcro Inc., Manchester,NH) straps were scorched (Fig. 4). The tempera-ture logger housings were in contact with the can-ister lids, and the bumpers were in contact withthe canister sides. Thus, at least some portions ofthe interior surface of the canisters were heatedto the melting point of ABS plastic, 103–128°C,and of Teflon, 260°C. Damage to the Teflonbumpers, ABS plastic, and Velcro straps only oc-curred at points of canister contact. With tem-peratures as high as 260°C at the internal wall ofthe canisters, it is remarkable that fewer than halfof the polystyrene Petri dishes, with a meltingpoint of 64–80°C, were damaged by heating.These observations suggest that, though the ex-terior of the canisters became quite hot, heattransfer toward the center of each canister was in-efficient. An additional cooling component couldarise from the adiabatic expansion of gas throughthe canister vent holes, since the canisters wereoriginally at cabin ambient pressure and the at-mospheric pressure at the altitude of the breakupwas considerably lower.
Unfortunately, data from the three recoveredtemperature loggers do not document the maxi-mum temperature reached within the canisters.Temperature was logged autonomously every 30min, but breakup occurred between logged datapoints. Ten minutes before breakup, canister tem-peratures were consistent with cabin ambient(22°C). Twenty minutes after breakup, internaltemperatures were about twice recovery site am-bient (canister 1, 26°C vs. 12°C; canister 2, 23°Cvs. 11°C; canister 5, 28°C vs. 14°C). Canister tem-peratures were consistent with ambient there-after. The elevated temperatures measured afterimpact probably represent residual heat pro-duced during the descent that had not completelydissipated into the surrounding environment.
C. elegans nematodes are killed rapidly by tem-peratures above 40°C (Anderson, 1978); thus itseems reasonable that survival, as describedabove, should correlate with maximum internalcanister temperature. It is also possible that toxic
C. ELEGANS SURVIVES COLUMBIA BREAKUP 697
5996_02_p690-705 12/20/05 4:03 PM Page 697

breakdown products were released by the heat-ing of the canister contents, though the Parafilmwrapping on each Petri plate would have pre-vented some exposure. Consistent with this, can-ister 1 contained all live animals and had nomelted plates. Canister 2 contained only one platewith live animals, while canister 4 contained onlyone plate without live animals (Table 1). In bothcases, the plate in question was located at an endof the canister, which suggests that locationwithin the canister was important to survival. Itis possible that one end of a canister was heatedmore than the other, which resulted in killingtemperatures at only one end. Canister 5 con-
tained no live animals and had partially meltedTeflon bumpers, Nomex® (DuPont)/nylon Vel-cro, and Petri dishes (Fig. 4a) as well as the high-est post-impact temperature recorded. These datasupport the conclusion that the animals in canis-ter 5 were killed by high temperatures duringreentry and/or toxic heating products. Unlikecanisters 1–4, however, canister 5 experiencedpost-impact pre-recovery freezing (data notshown), which provides for an alternative expla-nation.
Acceleration forces. At first glance, it may ap-pear surprising that C. elegans withstood an ac-
SZEWCZYK ET AL.698
FIG. 4. Canisters recovered from the Columbia debris. a: Ground control canister 1 is shown on the left, and flowncanister 1 is shown on the right. Note the ablation of anodization and visible pitting on the exterior of the canister.Damage occurred principally on the side, with some also present on the lid and bottom. Scorching is evident in thearea surrounding where the Kapton label had been, though the tape label remained intact. b: Flown canister 1 isshown along with all Teflon bumpers and Petri dishes. This photograph was taken immediately after the canister wasopened while disassembling the contents for further analysis. The temperature logger is out of the field of view.Melted plastic from the temperature logger housing is visible inside the canister. Partial melting and scorching of themiddle Teflon bumper are evident, with scorch marks on the side in contact with the canister wall at locations cor-responding to points of Petri dish contact on the opposite side of the bumper. Scorching of the Nomex/nylon Velcrostrap is evident in the upper right corner of the stack. The discoloration inside Petri dishes is not burnt agar; ratherit is the result of mold growth on the CeMM plates. c: Flown autonomous temperature loggers. All show melting ofthe plastic housing in points that were in direct contact with canister lids. The logger on the left is from canister 5,the middle from canister 1, and the right from canister 2. d: Flown canister 2 is shown with all Teflon bumpers andPetri dishes. This photograph was taken immediately after the canister was opened while disassembling the contentsfor further analysis. The temperature logger is out of the field of view. Dishes were unstrapped to allow examinationof the Petri dish containing lithium hydroxide, shown in front of canister. Note the ablation of anodization and vis-ible pitting on the exterior of the canister. Damage is noted to be fairly uniform around the canister and to a lesserextent than of some canisters. Scorching is evident in the area surrounding where the Kapton label had been, andportions are retained. The paper on the canister is a label applied during recovery efforts. Partial melting of the Teflonbumpers is evidenced by compression of the middle and right bumpers. Scorching of the Nomex/nylon Velcro strapis evident in the upper right corner of the strap. The Petri dish containing lithium hydroxide is melted at the pointof Nomex/nylon Velcro strap contact, with some melted strap present. The bottom-most Petri dish is also partlymelted. e: Flown canister 3 is shown with all Teflon bumpers and Petri dishes. This photograph was taken immedi-ately after the canister was opened while disassembling the contents for further analysis. Note the ablation of an-odization and visible pitting on the exterior of the canister. Damage is noted to be principally on the side displayedand top of the canister. A larger impact than on some canisters was noted on the top of this canister. The paper onthe canister is a label applied during recovery efforts. Partial melting of the Teflon bumpers is evidenced by com-pression of the middle and especially the right bumper. Scorching of the Nomex/nylon Velcro strap is evident in thebottom-right corner of the strap. The top two Petri dishes are partly melted, and the top one is cracked. f: Flown can-ister 4 is shown with all Teflon bumpers and Petri dishes. This photograph was taken immediately after the canisterwas opened while disassembling the contents for further analysis. Note the ablation of anodization and visible pit-ting on the exterior of the canister. Damage is noted to be fairly uniform around the canister and to a larger extentthan on other canisters. Larger impacts than on some other canisters were noted on the bottom. Scorching is evidentin the area surrounding where the Kapton label and paper tape label had been, and portions of both are retained.Partial melting of the Teflon bumpers is evidenced by compression of the middle and the right bumper. Scorchingand partial melting of the Nomex/nylon Velcro strap are evident on the top and side of the strap, respectively. ThePetri dish containing lithium hydroxide, shown in front of the stack, is melted at points of canister contact, with plas-tic retained on the inside of the canister lid. The top three and second from bottom Petri dishes in the stack are partlymelted. g: Flown canister 5 is shown with all Teflon bumpers and Petri dishes. The temperature logger is out of thefield of view. This photograph was taken immediately after the canister was opened while disassembling the con-tents for further analysis. Note the ablation of anodization and visible pitting on the exterior of the canister. Meltedplastic from the temperature logger housing is visible inside the canister. Partial melting of the Teflon bumpers is ev-idenced by compression of the middle bumper. Scorching and partial melting of the Nomex/nylon Velcro strap areevident on the top the strap. The top and bottom two Petri dishes in the stack are partly melted, and the bottom Petridish is cracked, as evidenced by the obvious lean of the stack of dishes.
➛
5996_02_p690-705 12/20/05 4:03 PM Page 698

C. ELEGANS SURVIVES COLUMBIA BREAKUP 699
5996_02_p690-705 12/20/05 4:03 PM Page 699

celeration due to impact with the ground thatwould probably have been lethal for larger or-ganisms. Two Petri dishes showed damage con-sistent with significant impact forces: the bot-tom-most Petri dish in canister 5 and thetop-most dish in canister 3 were cracked. Impactdamage to the agar was noted in all dishes (Fig.2), which is much more fragile than either poly-styrene or Parafilm. Neither the agar in thedishes nor the Parafilm used to wrap the Petridishes showed signs of scorching. Our theoreti-cal calculations of terminal velocity for the can-isters suggest that the transient acceleration ex-perienced upon impact would be nearly 2,500times Earth gravity. However, transient cen-trifugation at up to 10,000 g is a common ma-nipulation in standard C. elegans laboratory pro-cedures (e.g., Epstein and Shakes, 1995). Theimpact acceleration experienced during thistragic accident was well within their physiolog-ical tolerances.
DISCUSSION
C. elegans and biology in space
Experimentation with C. elegans is relativelyeasy and inexpensive. Animals have a short ges-tation period and produce large numbers of prog-eny, and mutant strains are easily isolated andcharacterized. The anatomy is simple, and a com-plete anatomical description of the nervous sys-tem is available (White et al., 1986). C. elegans wasthe first animal to have a fully sequenced genome(C. elegans Sequencing Consortium, 1998), and ex-cellent bioinfomatics tools are available, such as Wormbase (www.wormbase.org) (Stein et al.,2001). There is a large community of C. elegans re-searchers, many of whom study processes specif-ically relevant to astronaut health, such as radia-tion sensitivity and resistance; muscle structure,function, and atrophy; immune response; andstress responses. Using space-flown model or-ganisms such as C. elegans would complement ex-isting efforts and allow for the study of adapta-tion to spaceflight environments.
Conducting experiments in space is challeng-ing, in that power, stowage space, and astronauttime are limited (Space Sciences Board, 1998).Therefore, the development of experimental sys-tems that require minimal astronaut interven-tion, space, and power is important. Three fea-tures of CeMM culturing minimize the need forastronaut intervention when culturing C. elegans.Animals on CeMM will grow and reproduce considerably longer than on NGM without sub-culturing (Szewczyk et al., 2003). Culture auto-mation is feasible using liquid CeMM, and au-tomated analysis of liquid-grown animals ispossible using either video recordings or fluo-rescence sensors.
With respect to stowage, it is possible to trans-port large numbers of animals in minimal spaceas C. elegans is microscopic. The present study hasnot demonstrated the ability to transport largenumbers of animals because it was designed tomaximize redundancy in the event that any par-ticular canister was subject to unfavorable growthconditions. Future studies will likely involve or-ders of magnitude more animals in a fraction ofthe volume used in this study.
The issue of power is more complex. The pres-ent study demonstrated that C. elegans can sur-vive in passive onboard stowage. However, the
SZEWCZYK ET AL.700
FIG. 5. Location of recovered canisters. The Global Po-sitioning System locations of recovery provided by theCAIB were plotted in red against a map of the area. La-beling of the recovery sites is on the left of the site for can-isters 1, 3, and 5 and on the right for the site for canisters2 and 4. For reference, San Augustine is located at313147N, 0940621W; Bronson at 312038N, 0940048W; andthe CAIB numbering of recovered debris was as follows:canister 1, 68292; canister 3, 1676; canister 4, 2463; canis-ter 2, 1487; and canister 5, 8144.
5996_02_p690-705 12/20/05 4:03 PM Page 700

results from the OES demonstrated that space-craft may reach higher temperatures at which C.elegans become dauer or sterile. Most aspects ofC. elegans biology are temperature dependent, in-cluding growth rate (Byerly et al., 1976), egg lay-ing (Byerly et al., 1976), life span (Klass, 1977),dauer larvae formation (Ailion and Thomas,2000), oxygen consumption (Anderson, 1978),and tolerance to acute heat stress (Anderson,1978). Therefore, careful consideration of powerneeds for thermal control is required. Passivestowage may be acceptable for transport to andfrom space, but it would be prudent to conductexperiments in a thermally controlled environ-ment.
CeMM has advantages over NGM, but also has limitations. Animals grown on CeMM hadsignificantly lower broodsizes and decreasedgrowth rates than animals grown on NGM (Table2). Animals grown on CeMM were more suscep-tible to stressors associated with ambient growth(Fig. 3). It appears the benefits of using CeMMcome at the expense of rapid culture increase,though for longer-duration experiments this maynot be a significant detriment. In total, the datademonstrate that animals grown on CeMM aredifferent than animals grown on NGM, so cau-tion should be used when applying data from an-imals on NGM to animals grown on or in CeMM.Another caveat is that this study used only agarplates, and the surface tension to which the ani-mals were exposed might mask some adverse ef-fects of reduced gravity. A demonstration thatliquid CeMM can support growth during space-flight is still required before developing auto-mated CeMM culture systems for space-based re-search.
Packaging of living material for spaceflight
The survival of C. elegans despite the hazardsdescribed here and elsewhere (Szewczyk andConley, 2003; Szewczyk and McLamb, 2005)demonstrates that the packaging and placementof the specimens were sufficient for survival. TheBRIC canisters were stored inside a locker in themid-deck area of the orbiter. Additional studywould provide information on how to packagespecimens that are to be returned to Earth fromrobotic or human space vehicles during thecourse of a mission. Our findings indicate thatmultiple levels of physical containment, com-
bined with a thermal or ablative protection sys-tem, would be adequate. Specimens could be re-turned successfully in modules such as those em-ployed to return film by the U.S. CORONAproject and the Russian Zenit spacecraft (Day etal., 1998).
Planetary protection
With the commencement of human spaceflight,concerns have been raised regarding the possi-bility of contaminating other celestial bodies byterrestrial organisms (Rummel, 2001). In this re-gard, our data support the argument that plane-tary probes should be sterilized meticulously.Multiple spacecraft reentry events have resultedin spacecraft destruction, but a significant num-ber of spacecraft components have survived thesereentry events (Patera and Ailor, 1998). C. eleganssurvival of Columbia’s breakup demonstratesthat living organisms inside spacecraft debris cansurvive atmospheric reentry. This suggests thatdisposal of a spacecraft in the atmosphere of an-other celestial body may not incinerate the space-craft completely (Patera and Ailor, 1998), or anyliving organisms within it.
The recent Genesis mission accident accentu-ates this possibility. The Genesis sample returncanister did not release its drogue parachutes andimpacted the ground at a speed of 311 kph (NASA,2004a). From released images of the canister andimpact crater, it can be estimated that the canisterdecelerated over approximately 0.5 m. Using thecalculations presented in the Appendix, we de-termined that, for a deceleration time of 0.012 s,the deceleration force for the sample return can-ister of the Genesis mission was 731 G, consider-ably less than the deceleration calculated for oursmaller canisters. The sample collection hardwarewithin the return canister was surprisingly intactand is expected to yield useful samples for analy-sis (NASA, 2004b).
Permanent transfer of life requires the survivalof organisms during the trip to a particular ce-lestial body, as well as survival and reproductionin the new environment. It is possible for organ-isms to survive an interplanetary trip, but fail toreproduce after arrival. If the C. elegans onboardColumbia had not been returned to the authorsthe animals would not have survived. The N2 an-imals were starved and in stasis, while the CC1animals were reproductive but were being killed
C. ELEGANS SURVIVES COLUMBIA BREAKUP 701
5996_02_p690-705 12/20/05 4:03 PM Page 701

by the contaminating mold. For planets, moons,and other celestial objects where there is no evi-dence of life, no oxygen atmosphere, and no liq-uid water, it is unlikely that terrestrial organismscould survive.
Interplanetary transfer of life
Speculation on the interplanetary transfer oflife predates human spaceflight (e.g., Conan-Doyle, 1912; Dose, 1986). Life on Earth may haveevolved from a universal ancestor (Woese, 1998),but there are several theories of origin (Dose,1986). Life may have originated on another planetand survived escape, space travel, atmosphericentry, and impact on Earth, where it successfullyreproduced (Mileikowsky et al., 2000; Nicholsonet al., 2000). Our results demonstrate that livingorganisms can survive reentry into Earth’s at-mosphere during breakup of the surroundingspacecraft. As has been the case with flight hard-ware that survived reentry events (Patera andAilor, 1998), spacecraft breakup around the can-isters on board Columbia was likely to have dis-sipated some of the destructive forces (CAIB,2003; Szewczyk and Conley, 2003; Szewczyk andMcLamb, 2005). This process is analogous to theone that occurs when meteor breakup protectsmeteoroid fragments (Bronshten, 1983). Our datademonstrate that terrestrial organisms can sur-vive one of the three steps necessary for inter-planetary transfer of life—the other two essentialsteps are ejection from the planet of origin, andorbital passage between planets.
Available data are consistent with the possi-bility that terrestrial organisms can survive bothejection and space travel. Bacterial Bacillus subtilisspores and Deinococcus radiodurans cells can sur-vive the acceleration and jerk forces associatedwith ejection from a planetary surface (Mastrapaet al., 2001). The internal temperature of meteoriteALH84001 most likely did not exceed 40°C dur-ing or after ejection from Mars (Weiss et al., 2000),which is a survivable temperature for many or-ganisms. A number of nematodes are known to live in inhospitable environments, such asHaughton Crater well above the Arctic Circle onEarth (Mullin et al., 2002). During a transit, or-ganisms would need to withstand long-durationfreezing and possibly dessication. C. elegans cansurvive “indefinitely” in liquid nitrogen (Epsteinand Shakes, 1995), and other nematodes such asAnguina tritici can be revived by rehydration af-
ter 28 years (Fielding, 1951), conditions similar tothose possible inside a frozen meteoroid core.
This report has demonstrated that animals cansurvive a relatively unprotected reentry event,which is the last of the three legs required for aninterplanetary trip. Given these data, it is rea-sonable to speculate that organisms more com-plex than prokaryotes could be transferred be-tween planets by natural means. We do not knowwhether terrestrial life began here, and our re-sults suggest it is possible that even animals havebeen ejected from Earth and transported else-where in our Solar System.
APPENDIX
Terminal velocity and impact calculation for a BRIC-60 class canister
For a body falling through a fluid, three forcesact on a body: gravity, buoyancy, and drag (Hal-liday and Resnick, 1986; Denn, 1980):
FG � ��p�Dp3g/6 (1)
FB � ��Dp3g/6 (2)
FD � (�/8)�Vp2Dp
2CD (3)
where FG � force due to gravity, FB � force dueto buoyancy, FD � force due to drag, �p � den-sity of the body (in kg/m3), � � density of thesurrounding fluid (in kg/m3), � is dimensionless,Dp � diameter of the body (in m), g � accelera-tion due to gravity (9.81 m/s2), Vp � velocity ofthe body (in m/s), and CD � drag coefficient (di-mensionless). At steady state, as experienced atterminal velocity, Eqs. 1–3 sum to 0.
0 � (��p�Dp3g/6) � (��Dp
3g/6)
� ((�/8)�Vp2Dp
2CD) (4)
For a body moving at terminal velocity through arelatively non-dense and non-viscous fluid like air,the Reynolds number (Re) is anticipated to be high.For 103 � Re � 2 � 105, the so-called Newtonianregime of the Re, CD � 0.44. Equation 4 yields:
Vp � [3Dpg((�p/�) � 1)]0.5 (5)
The BRIC canister is an irregular object. Calculatea sphere of equivalent surface area of a BRIC can-
SZEWCZYK ET AL.702
5996_02_p690-705 12/20/05 4:03 PM Page 702

ister. The BRIC is a cylinder, where Scyl � cylin-der surface area (in m2), rc � cylinder radius �0.041 m, and h � cylinder height � 0.159 m:
Scyl � 2(�rc2) � 2�rch (6)
Scyl � 2 (3.1415)(0.041)2
� 2 (3.1415)(0.041)(0.159)
Scyl � 2 (3.1415)(0.00681)� 2 (3.1415)(0.041)(0.159)
Scyl � 0.010562 � 0.04096
Scyl � 0.052 m2 (7)
Determine the diameter of a sphere of equivalentsurface area as in Eq. 7, where Ssph � sphere sur-face area (in m2) and rs � sphere radius (in m):
Ssph � 4�rs2 (8)
rs � (Ssph/4�)0.5
rs � [0.052/(4 � 3.1415)]0.5
rs � 0.064 m (9)
Dp � 2rs � 2(0.064) � 0.13 m (10)
Determine the volume of the equivalent sphere,where Vp � spherical particle volume (in m3):
Vp � (4/3)�rs3 (11)
Vp � (4/3)(3.1415)(0.064)3
Vp � 0.001 m3 (12)
Convert the mass of the BRIC canister:
m � 1.5 lbs � (1 kg/2.2 lbs) � 0.68 kg (13)
Determine the density of the equivalent spherefrom Eqs. 11 and 12:
�p � m/Vp (14)
�p � 0.68/0.001 � 680 kg/m3 (15)
From Denn (1980), the density of the atmospherenear sea level is:
� � 1.3 kg/m3
This is a reasonable value to use. Even though theatmosphere is much thinner at 61 km where dis-integration of the vehicle occurred, the steady-state velocity of the canister would have changedas it descended into the more dense lower at-mosphere. In addition, the canisters likely wouldhave experienced significant angular accelerationdue to tumbling and potential impact with otherdebris during the descent. Also, it is unknown atwhat altitude the canisters were released from thelocker in which they were contained. The inertialacceleration at high altitude is somewhat lessthan gravity at the surface of the Earth. However,again, steady state is assumed at low altitude aswell so the value near the surface of the Earth isused. Solving Eq. 5:
Vp � [3(0.13)(9.81)((680/1.3) � 1)]0.5
Vp � [3.826 (522)]0.5
Vp � [1,997.5]0.5
Vp � 44.7 m/s or 100 miles/h (16)
Verify the Re assumption to determine whetheror not a BRIC canister traveling at velocity (Eq.16) is in the Newtonian regime. Viscosity of air istaken from Denn (1980), where Re is dimension-less) and � � viscosity of air (2 � 10�5 Pa s):
Re � DpVp�/� (17)
Re � (0.13)(44.7)(1.3)/0.00002 � 377,715 (18)
This result is somewhat troublesome because itsuggests the Re is beyond the range appropriatefor the use of Eq. 5. At Re values greater than ap-proximately 350,000, a marked drop in CD is ob-served, and calculations become more difficult(Denn, 1980; Perry and Green, 1984). However,our calculated terminal velocity of slightly lessthan 45 m/s is reasonable given terminal veloci-ties reported for other objects.
Assuming that the calculations above are stillvalid, then deceleration may be calculated. Eithera deceleration time or a deceleration distancemust be estimated to perform this calculation. Wechoose a deceleration distance of 4 cm, approxi-mately one-half the diameter of a BRIC, based onanecdotal descriptions that canisters were em-bedded halfway in the earth before recovery (im-ages not available). The equation from which we
C. ELEGANS SURVIVES COLUMBIA BREAKUP 703
5996_02_p690-705 12/20/05 4:03 PM Page 703

can calculate the time required to decelerate overa given distance is, where V � final velocity (0m/s), Vo � initial velocity (44.7 m/s, terminal ve-locity), d � deceleration distance (in m), and t �deceleration time (in s):
d � 0.5(Vo � V)t (19)
0.04 � 0.5(44.7 � 0)t
t � 0.002 s (20)
The values of 45 m/s for terminal velocity and0.002 s for deceleration time were used to calcu-late the acceleration in G:
acceleration (in G) � 45 m/s divided by 0.002 sdivided by 9.8 m/s/s, or 2,295 G
IN MEMORIAM
R. Husband, W. McCool, D. Brown, L. Clark, I.Ramon, M. Anderson, K. Chawla, K. Forbes-Dun-field.
ACKNOWLEDGMENTS
Strain N2 was provided by Stuart Kim, andstrains CC2 (recovered N2) and CC3 (recoveredCC1) were deposited with the Caenorhabditis Ge-netics Center, which is funded by the National In-stitutes of Health’s National Center for ResearchResources. Thanks to the rescue and recoveryworkers and the Columbia Accident InvestigationBoard for recovery and recovery details of flighthardware. Thanks to J. Barfus, Earth Systems Mod-eling & Data Management Lab, Dynamac Corpo-ration, for generation of the ArcGIS figures de-picting recovered canister locations. Thanks to S.Winfree, J. Sunga, G. O’Brien, J. Cohen, K. Cullings,C. Wade, K. Norwood, A. Boody, F. Ahmay, andG. Etheridge for technical assistance. Thanks to G.Benian, D. Flaherty, H. Joshi, and the Space StationBiological Research Project for reagents. Thanks to G. Nelson, M. Hughes-Fulford, L. Jacobson, T. Powers, M. Favero, J. Rummel, D. Williams, C.Swain, J. Allton, D. Des Marais, and D. Beaty foruseful discussions. B. Little, G. Stephenson, and T.Pohle provided library support. This research wasfunded by NASA Fundamental Biology and theNASA Astrobiology Institute.
ABBREVIATIONS
ABS, acrylonitrile butadiene styrene; CAIB,Columbia Accident Investigation Board; CeMM,C. elegans Maintenance Medium; L1, first-stagelarval; NGM, Nematode Growth Medium; OES,Orbital Environmental Simulator.
REFERENCES
Ailion, M. and Thomas, J.H. (2000) Dauer formation in-duced by high temperatures in Caenorhabditis elegans.Genetics 156, 1047–1067.
Anderson, G.L. (1978) Responses of dauer larvae of C. el-egans (Nematoda: Rhabitidae) to thermal stress andoxygen deprivation. Can. J. Zool. 56, 1786–1791.
Bolanowski, M.A., Russell, R.L., and Jacobson, L.A. (1981)Quantitative measures of aging in the nematodeCaenorhabditis elegans. I. Population and longitudinalstudies of two behavioral parameters. Mech. AgeingDev. 15, 279–295.
Brenner, S. (1974) The genetics of Caenorhabditis elegans.Genetics 77, 71–94.
Bronshten, V.A. (1983) Physics of Meteoric Phenomena, D. Reidel Publishing Co., Dordrecht, The Netherlands.
Byerly, L., Cassada, R.C., and Russell, R.L. (1976) The lifecycle of the nematode Caenorhabditis elegans. I. Wild-type growth and reproduction. Dev. Biol. 51, 23–33.
C. elegans Sequencing Consortium (1998) Genome se-quence of the nematode C. elegans: a platform for in-vestigating biology. Science 282, 2012–2018.
Cassada, R.C. and Russell, R.L. (1975) The dauerlarva, apost-embryonic developmental variant of the nema-tode Caenorhabditis elegans. Dev. Biol. 46, 326–342.
Columbia Accident Investigation Board (2003) ColumbiaAccident Investigation Board Report, Columbia AccidentInvestigation Board, Washington, DC.
Conan-Doyle, A. (1912) The Lost World, Oxford UniversityPress, Oxford.
Day, D.A., Logsdon, J.M., and Latell, B. (1998) Eye in theSky: The Story of the CORONA Spy Satellites, Smithson-ian Institution Press, Washington, DC.
Denn, M. (1980) Process Fluid Mechanics, Prentice-Hall,Upper Saddle River, NJ.
Dose, K. (1986) Hypotheses on the appearance of life onEarth. Adv. Space Res. 6, 181–186.
Dusenbery, D.B. (1974) Analysis of chemotaxis in the ne-matode C. elegans by countercurrent separation. J. Exp.Zool. 188, 41–47.
Dusenbery, D.B. (1985) Video camera-computer trackingof nematode C. elegans to record behavioral responses.J. Chem. Ecol. 11, 1239–1247.
Epstein, H. and Shakes, D., eds. (1995) Methods in Cell Biology, Vol. 48: Caenorhabditis elegans: Modern Bio-logical Analysis of an Organism, Academic Press, SanDiego, CA.
Fielding, M.J. (1951) Observations on the length of dor-
SZEWCZYK ET AL.704
5996_02_p690-705 12/20/05 4:03 PM Page 704

mancy in certain plant infecting nematodes. Proc.Helminth. Soc. Wash. 18, 110–112.
Gems, D. and Riddle, D.L. (2000) Genetic, behavioral andenvironmental determinants of male longevity inCaenorhabditis elegans. Genetics 154, 1597–1610.
Halliday, D. and Resnick, R. (1986) Fundamentals ofPhysics, 2nd ed., John Wiley & Sons, New York.
Hartman, P.S., Hlavacek, A., Wilde, H., Lewicki, D., Schu-bert, W., Kern, R.G., Kazarians, G.A., Benton, E.V., Ben-ton, E.R., and Nelson, G.A. (2001) A comparison of mu-tations induced by accelerated iron particles versusthose induced by low earth orbit space radiation in theFEM-3 gene of Caenorhabditis elegans. Mutat. Res. 474,47–55.
Johnson, T.E. and Nelson, G.A. (1991) Caenorhabditis ele-gans: a model system for space biology studies. Exp.Gerontol. 26, 299–309.
Klass, M.R. (1977) Aging in the nematode Caenorhabditiselegans: major biological and environmental factors in-fluencing life span. Mech. Ageing Dev. 6, 413–429.
Lu, N.C. and Goetsch, K.M. (1993) Carbohydrate re-quirement of Caenorhabditis elegans and the final devel-opment of a chemically defined medium. Nematologica39, 303–331.
Mastrapa, R.M.E., Glanzberg, H., Head, J.N., Melosh, H.J.,and Nicholson, W.L. (2001) Survival of bacteria exposedto extreme acceleration: implications for panspermia.Earth Planet. Sci. Lett. 189, 1–8.
Mileikowsky, C., Cucinotta, F.A., Wilson, J.W., Gladman,B., Horneck, G., Lindegren, L., Melosh, J., Rickman, H.,Valtonen, M., and Zheng, J.Q. (2000) Natural transferof viable microbes in space. Icarus 145, 391–427.
Mullin, P., Shuerger, A., and Powers, T. (2002) Nematodesof Haughton Crater. Nematology 4, 270.
NASA (2004a). Genesis Mission Status Report on Sept. 8,2004, press release #04-292, available at http://www.nasa.gov/home/hqnews/2004/sep/HQ_04292_Gene-sis_status.html
NASA (2004b). Genesis Mission Status Report on Sept. 30, 2004, press release #2004-245, available at http://www.nasa.gov/mission_pages/genesis/media/gene-sis-093004.html
Nelson, G.A., Schubert, W.W., Kazarians, G.A., andRichards, G.F. (1994a) Development and chromosomemechanics in nematodes: results from IML-1. Adv. SpaceRes. 14, 209–214.
Nelson, G.A., Schubert, W.W., Kazarians, G.A., Richards,G.F., Benton, E.V., Benton, E.R., and Henke, R. (1994b)Radiation effects in nematodes: results from IML-1 ex-periments. Adv. Space Res. 14, 87–91.
Nicholson, W.L., Munakata, N., Horneck, G., Melosh, H.J.,and Setlow, P. (2000) Resistance of Bacillus endosporesto extreme terrestrial and extraterrestrial environments.Microbiol. Mol. Biol. Rev. 64, 548–572.
Oser, H. and Battrick, B. (1989) Life Sciences Research inSpace, ESA Publications Division, Noordwijk, TheNetherlands.
Patera, R.P. and Ailor, W.H. (1998) The realities of reen-try disposal. Adv. Astronaut. Sci. 99, 1059–1071.
Perry, R. and Green, D. (1984) Perry’s Chemical Engineers’Handbook, 6th ed., McGraw Hill, New York.
Rummel, J.D. (2001) Planetary exploration in the time ofastrobiology: protecting against biological contamina-tion. Proc. Natl. Acad. Sci. USA 98, 2128–2131.
Space Science Board (1998) A Strategy for Research in SpaceBiology and Medicine in the New Century, National Re-search Council, Washington, DC.
Stein, L., Sternberg, P., Durbin, R., Thierry-Mieg, J., andSpieth, J. (2001) WormBase: network access to thegenome and biology of Caenorhabditis elegans. NucleicAcids Res. 29, 82–86.
Szewczyk, N.J. and Conley, C.A. (2003) NASA TechnicalReport: Surviving Atmospheric Spacecraft Breakup, doi20030710, NASA Center for AeroSpace Information,Washington, DC.
Szewczyk, N.J. and McLamb, W. (2005) Surviving at-mospheric spacecraft breakup. Wilderness Environ. Med.16, 27–32.
Szewczyk, N.J., Kozak, E., and Conley, C.A. (2003) Chem-ically defined medium and Caenorhabditis elegans. BMCBiotechnol. 3, 19.
Trent, C., Tsung, N., and Horvitz, H.R. (1983) Egg-layingdefective mutants of the nematode C. elegans. Genetics104, 619–647.
Viglierchio, D.R. (1992) The World of Nematodes, AgAccess,Davis, CA.
Weiss, B.P., Kirschvink, J.L., Baudenbacher, F.J., Vali, H.,Peters, N.T., Macdonald, F.A., and Wikswo, J.P. (2000)A low temperature transfer of ALH84001 from Mars toEarth. Science 290, 791–795.
White, J.G., Southgate, E., Thomson, J.N., and Brenner, S.(1986) The structure of the nervous system of Caenorhab-ditis elegans. Philos. Trans. R. Soc. Ser. B 314, 1–340.
Woese, C. (1998) The universal ancestor. Proc. Natl. Acad.Sci. USA 95, 6854–6859.
Wright, I.P., Grady, M.M., and Pillinger, C.T. (1989) Or-ganic materials in a martian meteorite. Nature 340,220–222.
Zdinak, L.A., Greenberg, I.B., Szewczyk, N.J., Barmada,S.J., Cardamone Rayner, M., Hartman, J.J., and Jacob-son, L.A. (1997) Transgene-coded chimeric proteins asreporters of intracellular proteolysis: starvation-in-duced catabolism of a lacZ fusion protein in musclecells of Caenorhabditis elegans. J. Cell. Biochem. 67, 143–153.
Address reprint requests to:Catharine A. Conley
NASA Ames Research CenterM/S 239-11
Moffett Field, CA 94035-1000
E-mail: [email protected]
C. ELEGANS SURVIVES COLUMBIA BREAKUP 705
5996_02_p690-705 12/20/05 4:03 PM Page 705