brain stem afferents to the festigial nucleus in the cat demonstrated by transport of horseradish...
TRANSCRIPT

Brain Stem Afferents to the Fastigial Nucleus in the Cat Demonstrated by Transport of Horseradish Peroxidasel
DAVID RUGGIERO, ROBERT R. BATTON I I I , ~ , ~ A. JAYARAMAN 2 AND MALCOLM B. CARPENTER Department of Anatomy, College of Physicians and Surgeons, Columbia Unioersity, New York, New York 10032
ABSTRACT Although retrograde and anterograde degeneration studies have provided important information concerning brain stem afferents to the fastigial nu- cleus (FN), these data may be incomplete and should be confirmed by axonal trans- port methods. Attempts were made to inject horseradish peroxidase (HRP) unilaterally into the FN in a series of adult cats. Animals were perfused with dex- tran and a fixative solution of paraformaldehyde and glutaraldehyde in 0.1 M phos- pate buffer. Representative sections were treated by the Graham and Karnovsky (‘66) method.
Selective HRP injections in one FN resulted in retrograde transport of the marker to Purkinje cells of the ipsilateral vermis and distinctive appendages of the contralateral medial accessory olivary (MAO) nucleus (nucleus p and the dorso- medial cell column). Retrograde transport of the label was found bilaterally in cells of the medial (MVN) and inferior (IVN) vestibular nuclei, in cell group x and in the nucleus prepositus (PP). Labeled vestibular neurons, most numerous in MVN, were identified in dorsal, caudal and lateral regions, with a slight ipsilateral prepon- derance. Only a few neurons in caudal, dorsal and lateral regions of the IVN were labeled and none of these included cells of group f. Labeled cells in the caudal third of PP were greatest ipsilaterally. Rostra1 and caudal injections of FN labeled smaller numbers of cells in MVN, IVN, cell group x and PP.
HRP injections of FN and portions of lobules VIII and IX resulted in bilateral retrograde labeling of larger numbers of cells in MVN, IVN and cell group x, and ipsilateral labeling of cells in group y and the interstitial nucleus of the vestibular nerve. Injections of HRP into basal folia of lobules V and VI resulted in retrograde transport of the marker to cells of the medial and dorsal accessory olivary nuclei contralaterally, and to cells of the ipsilateral accessory cuneate nucleus. Transport of label injected into portions of the pyramis was detected in parts of the con- tralateral MA0 and bilaterally in parts of the pontine and reticulotegmental nuclei.
This study suggests that the principal afferents of the fastigial nucleus arise from: (1) Purkinje cells of the ipsilateral vermis, (2) restricted portions of the con- tralateral M A 0 (nucleus /3 and dorsomedial cell column), (3) portions of the MVN and IVN (bilaterally) and (4) caudal parts of the PP. Secondary vestibular inputs to the fastigial nucleus probably are relayed mainly by Purkinje cells in the cerebellar cortex.
In attempts to determine the Origins and terminations of secondary vestibulocere- bellar fibers by degeneration technics two
have been employed: (1) the study of retrograde cell changes after cer- ebellar lesions, and (2 ) the study of anterograde degeneration following lesions in the vestibular nuclei. Using the modified
J. CUMP. NEUR., 172: 189-210. 189
’ This investigation was supported by Research Grant NS- 01538-18 and NS-13747-01 from the National Institute of Neurological and Communicative Disorders and Stroke of the National Institutes of Health, Bethesda, Maryland.
* Postdoctoral trainees in neuroanatomy supported by Training Grant 5T01-NS-05242-17 from the National Institute of Neurological and Communicative Disorders and Stroke.
Current address: Department of Anatomy, Hershey Col- lege of Medicine, Pennsylvania State University, Hershey, Pennsylvania 17033.

190 D. RUGGIERO, R. BATTON 111, A.
Gudden method, Brodal and Torvik (’57) reported that lesions involving “vestibular parts” of the cerebellum produced un- e uivocal cell changes in three locations: (3 ventrolateral and caudal parts of the inferior vestibular nucleus (IVN), including cell group f (Brodal and Pompeiano, ’57), (2) ventral and caudal parts of the medial vestibular nucleus (MVN), and (3) cell group x, situated along the lateral border of IVN. Since these lesions were not re- stricted to particular subdivisions of the “vestibular part” of the cerebellum (i.e., nodulus, uvula or fastigial nucleus) and most lesions of the fastigial nucleus (F”) encroached upon portions of the uvula and nodulus, these data can not be regarded as definitive for the fastigial nucleus. Isolated lesions of the fastigial nucleus in the adult cat suggested that cells in the IVN pro- jected both crossed and uncrossed fibers to the fastigial nucleus (Carpenter et al., ’59).
Attempts to trace degeneration from lesions in the vestibular nuclei into the cer- ebellum indicated terminations in the ipsi- lateral nodulus, uvula, flocculus and the fastigial nucleus. (Dow, ’36). Silver staining methods for degenerated fibers confirmed this observation but revealed only scant degeneration in rostral arts of the fastigial nucleus (Carpenter, ’60 7 .
While both of the above methods pro- vided certain information concerning sec- ondary vestibulocerebellar fibers, neither method produced specific data concerning afferents to the fastigial nuclei. The method of retrograde axonal transport of horseradish peroxidase (HRP) (LaVail and LaVail, ’72; LaVail, ’75) appeared to offer possibilities for the determination of brain stem afferents to the fastigial nucleus. This method was used.
MATERIALS AND METHODS
In a series of adult cats attempts were made to inject horseradish peroxidase (HRP, Sigma type VI) into portions of one fastigial nucleus. Injections of HRP were made stereotaxically via a suboccipital ap- proach with a 1 p1 Hamilton syringe. Volumes of a 30% solution of HRP (dis-
JAYARAMAN AND M. CARPENTER
tilled water) injected ranged from 0.1-0.7 p1; these volumes were injected slowly over a 15 to 30 minute interval by a micro- meter drive.
After postoperative survival times of 24 to 72 hours, animals were anesthetized with Nembutal and perfused via the left ventricle of the heart. Some animals were given 500 units of heparin a half hour before they were perfused. Animals were perfused with a liter of 6% dextran with 5% dextrose, followed by 1 or 2 1 of a fixative solution containing either 1 or 2% paraformaldehyde and 1.25 or 2.5% glutar- aldeh de in a 0.1 M phosphate buffer (pH 7.4) i;( im and Strick, ’76). Brains were immediately dissected out and sectioned perpendicular to the axis of the brain stem. Blocks of brain were immersed in the above described fixative and stored over- night at 4°C. On the following day the tis- sue blocks were transferred to fresh 0.1 M phosphate buffer containing 30% sucrose and stored another 24 hours at 4°C.
Blocks of tissue from the brain stem, cer- ebellum and upper cervical spinal cord were cut on a freezing microtome at 40 pm and collected in representative groups of five to six sections in distilled water. These sections were treated by the Graham and Karnovsky (’66) method to produce the reaction for the identification of HRP activity (LaVail, ’75). Sections were mounted on slides with gelatin, and approximately half of the sections were lightly stained with cresyl violet. Both stained and unstained sections were studied microscopically with bright and dark field illumination.
RESULTS
In the stereotaxic approach used, the injection needle usually traversed portions of the pyramis and entered the fastigial nu- cleus in its longitudinal axis. The principal structure labled by this enzyme marker was the fastigial nucleus, but frequently there was diffusion of HRP into the sur- rounding white matter, folia of the pyramis and uvula and occasionally into basal folia of lobules V and VI (Larsell, ’53) rostral to

BRAIN STEM FASTIGIAL AFFERENTS PLATE 191
the fastigial nucleus. While it was virtually impossible to precisely define the regions of HRP uptake, two important bits of evi- dence were used to support the premise that axon terminals within the fastigial nu- cleus had taken up the protein marker, namely: (1) retrograde transport of HRP granules to the somata and dendritic ex- pansions of Purkinje cells in the ipsilateral cerebellar vermis, and (2) retrograde transport of HRP granules to cells of the dorsomedial cell column and nucleus /3 of the contralateral medial accessory olivary nucleus (Brodal, '40, '67, '76). Labeling of portions of the cerebellar cortex, either alone, or in addition to the fastigial nucleus, produced a different pattern of retrograde transport of label to cells of the contralat- era1 inferior olivary complex, as well as la- beling of cells in other cerebellar relay nu- clei.
Unilateral fastigial injections Entire fastigial nucleus
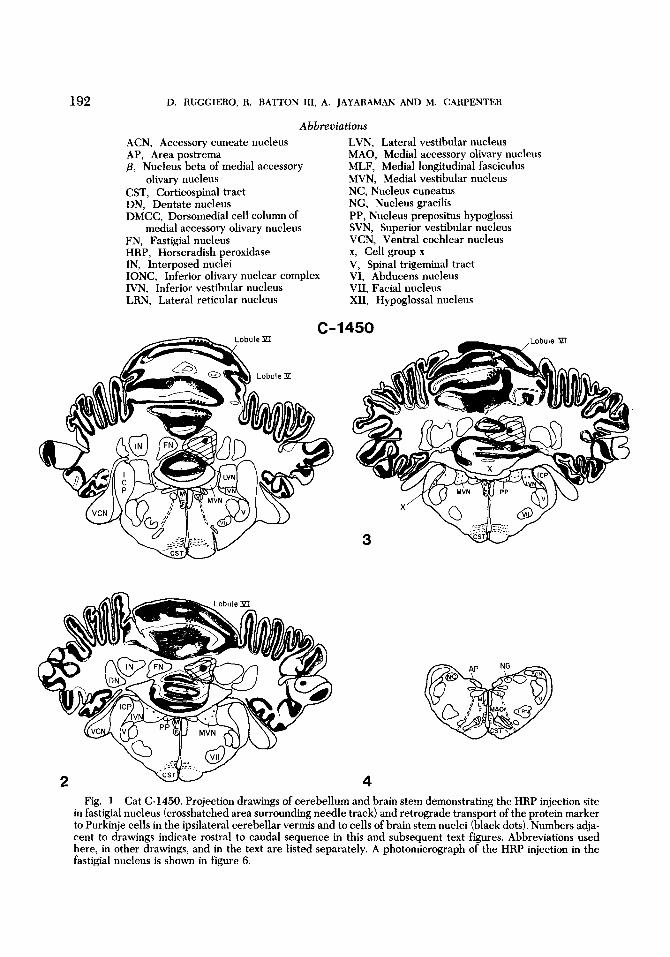
Attempted HRP injections in two ani- mals resulted in apparent selective labeling of the entire fastigial nucleus on one side (figs. 1,5,6). There was no evidence of pri- mary HRP label in the cerebellar cortex or in other deep cerebellar nuclei. In one ani- mal (C-1450) retrograde transport of the HRP label to Purkin'e cells was seen in a
the injected side in folia of lobules I through X (Larsell, '53). Retrograde la- beling of Purkinje cells was maximal in folia of lobules VI, VII, V and X, in that order (fig. 12). Similar, but less widely dis- tributed, retrograde labeling of the somata and dendritic expansions of Purkinje cells was seen in cat C-1411; many of these dendritic profiles of Purkinje cells ex- tended from the somata to the pial surface (fig. 11). Labeling of Purkinje cells in this animal was evident only in folia of lobules V and VI.
Retrograde transport of the HRP label was seen in portions of the vestibular nu- clei, the nucleus prepositus and the inferior olivary nuclear complex (fig. 1). Consider-
narrow longitudinal i and of the vermis on
ing the massive retrograde transport of the label to vermal Purkinje cells in cat C- 1450, label in the contralateral medial accessory olivary nucleus (MAO) was mini- mal. Label was found laterally in caudal parts of the MAO, in nucleus j3 and in sur- rounding dorsomedial parts of the MAO. In cat C-1411 retrograde transport of label to cells of the contralateral inferior olivary nuclear complex (IONC) was seen in nu- cleus /3, the dorsomedial cell column, and the dorsal cap of Kooy. All of these cell groups except the dorsal cap are distinctive appendages of the MA0 (Brodal, '40, '76). No labeled neurons were seen in the ipsilateral IONC.
HRP label transported to portions of the vestibular nuclei and the nucleus pre- positus was bilateral (figs. 1, 13, 14). In cat C-1450 label was seen primaril in parts of the medial vestibular nuclei [MVN). La- beled cells in the MVN, identified in dorsal, caudal and lateral regions, were nearly equal on the two sides. Only a very few cells in the inferior vestibular nucleus (IVN) were labeled and none of these in- cluded the large-celled group located ven- trolaterally and designated as cell group f (Brodal and Pompeiano, '57). Small cells located lateral to the IVN and rostra1 to the accessory cuneate nucleus, known as cell group x, were labeled bilaterally (fig. 1).
In cat C-1411 labeled neurons in the vestibular nuclei were seen bilaterally in caudal and dorsal parts of the IVN, but not in cell group f. Retrograde transport of la- bel also was seen bilaterally in some neu- rons of cell group x. The scattered cells composing this nucleus appeared small and spindle-shaped with branched radiating dendrites. In the MVN numerous medium- sized neurons were labeled bilaterally, predominately in dorsal regions of the nu- cleus (figs, 13, 14). Although labeled neu- rons were most numerous in caudal parts of the MVN, cells with label were identified throughout the rostro-caudal extent of the nucleus. At all levels labeled cells were appreciably greater on the injected side and more numerous than those in the IVN. No neurons in the lateral vestibular nucleus
192 D. RUGGIERO, R. BATTON 111, A. JAYARAMAN AND M. CARPENTER
Abbreviations ACN, Accessory cuneate nucleus AP, Area postrema p, Nucleus beta of medial accessory
olivary nucleus CST, Corticospinal tract DN, Dentate nucleus DMCC, Dorsomedial cell column of
medial accessory olivary nucleus FN, Fastigial nucleus HRP, Horseradish peroxidase IN, Interposed nuclei IONC, Inferior olivary nuclear complex IVN, Inferior vestibular nucleus LRN, Lateral reticular nucleus
LVN, Lateral vestibular nucleus MAO, Medial accessory olivary nucleus MLF, Medial longitudinal fasciculus MVN, Medial vestibular nucleus NC, Nucleus cuneatus NG, Nucleus gracilis PP, Nucleus prepositus hypoglossi SVN, Superior vestibular nucleus VCN, Ventral cochlear nucleus x, Cell group x v, Spinal trigeminal tract VI, Abducens nucleus VII, Facial nucleus XII, Hypoglossal nucleus
C-1450
y&J/ CST 3
4 Fig. 1 Cat C-1450. Projection drawings of cerebellum and brain stem demonstrating the HRP injection site
in fastigial nucleus (crosshatched area surrounding needle track) and retrograde transport of the protein marker to Purkinje cells in the ipsilateral cerebellar vermis and to cells of brain stem nuclei (black dots). Numbers adja- cent to drawings indicate rostra1 to caudal sequence in this and subsequent text figures. Abbreviations used here, in other drawings, and in the text are listed separately. A photomicrograph of the HRP injection in the fastigial nucleus is shown in figure 6.

BRAIN STEM FASTIGIAL AFFERENTS PLATE 193
(LVN), the superior vestibular nucleus (SVN), cell group y, or the interstitial nuc- leus of the vestibular nerve were labeled in either of these animals.
Neurons in the nucleus prepositus were labeled with HRP granules bilaterally in the caudal third of the nucleus, with the largest number of identified cells ip- silateral to the injection. No other brain stem nuclei were labeled as a consequence of these fastigial injections, except for a few cells in the medial pontine nuclei at isthumus levels in cat C-1411.
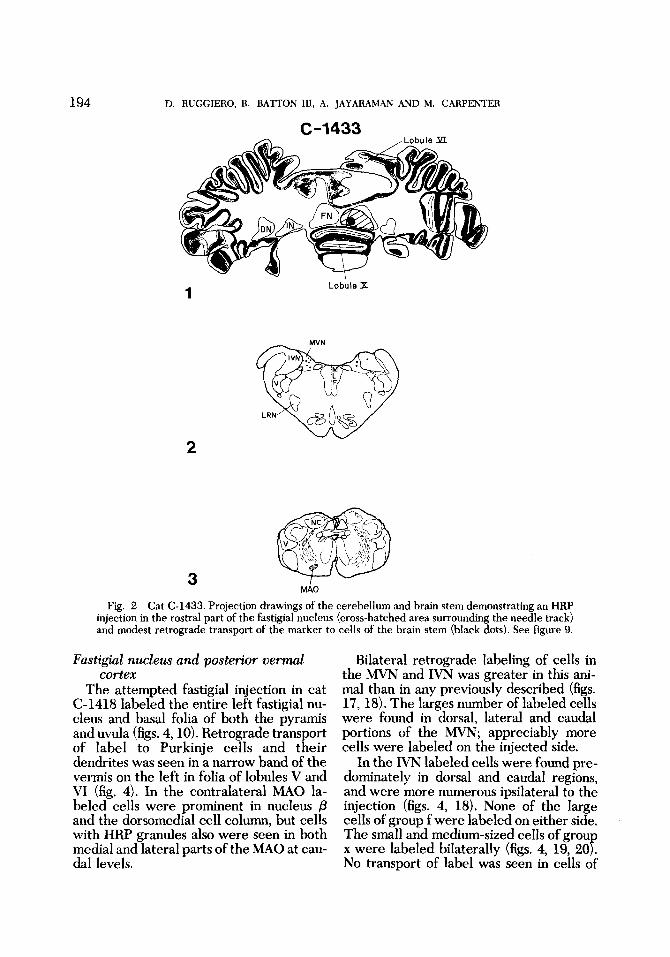
Rostra1 fastigial nucleus Attempts were made to inject the right
fastigial nucleus (C-1433) with 0.1 pl of HRP. The injection needle tranversed por- tions of the uvula, passed medial to the caudal pole of the fastigial nucleus and en- tered the rostral half of the fastigial nu- cleus where an intense brownish-yellow stain was evident (figs. 2, 9). The needle did not extend into the rostral pole of the fastigial nucleus and very little HRP was seen in the white matter rostral to it. Thus, the HRP injection was confined largely to the rostral half of the fastigial nucleus, excluding the rostral pole. Retrograde transport of the label was seen in the Purkinje cells in the overlying vermal cor- tex (fig. 2). Label also was seen in cells of the caudal part of the left MAO.
Retrograde transport of the injected la- bel to brain stem nuclei was modest (fig. 2). Only one or two cells in the IVN ipsilateral to the injection contained HRP granules. Label in cells of the caudal parts of the MVN was bilateral and greatest contralat- erally. Only one or two labeled cells were seen in the contralateral nucleus preposi- tus. No labeled cells were seen in other vestibular nuclei, the accessory cuneate nucleus (ACN) or in the lateral reticular nucleus (LRN) of the medulla.
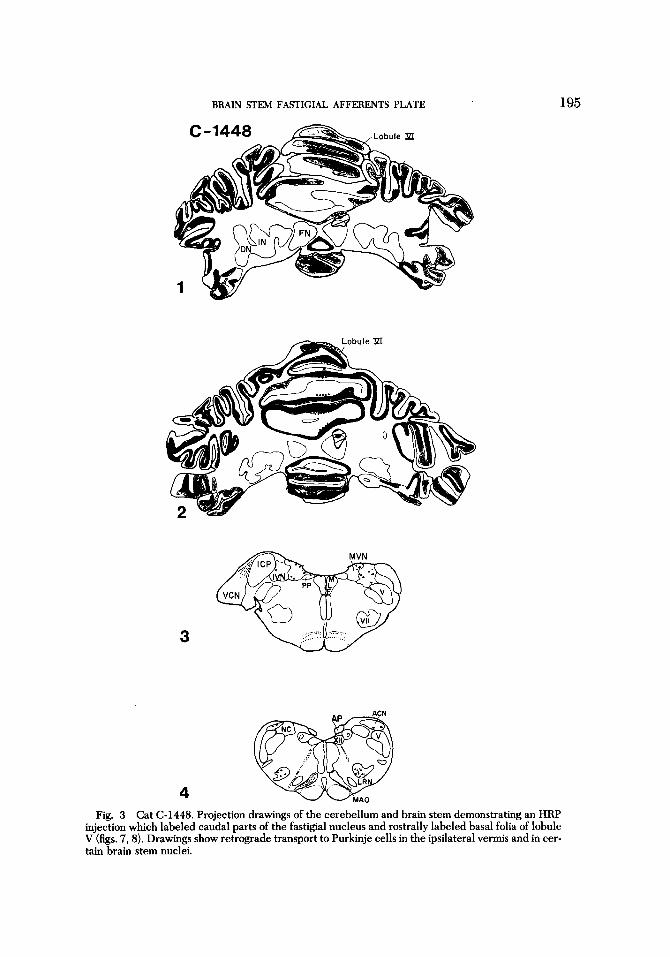
Caudal fastigial nucleus In one animal (C-1448) the injection
needle traversed dorsal parts of the caudal two-thirds of the fastigial nucleus before emerging from the nucleus to enter basal
folia of lobule V (figs 3,7, 8). HRP stain was intense in the dorsal third of the caudal part of the fastigial nucleus. There was no diffusion of HRP into other deep cerebellar nuclei.
Purkinje cells were labeled by retro- grade transport in lobules V and VI of the ipsilateral vermis (fig. 3). A few cells of the left fastigial nucleus exhibited HRP gran- ules in their somata and dendrites (fig. 22). One or two similarly labeled cells in ventral parts of the right fastigial nucleus suggested retrograde transport of HRP from the cortex of lobule V. Retrograde transport of HRP was seen mainly in the MA0 on the left side. Labeled cells were present in nucleus /3 and in the dorsal cap of Kooy.
Labeled cells in portions of the MVN and IVN were less numerous and exhibited a different pattern than that seen with more extensive labeling of the fastigial nucleus (fig. 3). Cells of the IVN were labeled pre- dominately in dorsal and lateral regions, but no label was seen in cells of group f; la- beled cells were most numerous ipsilateral- ly. Although cells of group x were labeled bilaterally by retrograde transport, their number was small.
The number of labeled cells in the MVN was less than that found in animals with more extensive injections of the fastigial nucleus. Cells labeled in the h4VN were found predominately ipsilaterally in dorsal and caudal parts of the nuclei. No neurons of the LVN were labeled on either side, and only one or two neurons in peripheral portions of the SVN were labeled ipsilat- erally. None of the neurons of cell group y or the interstitial nucleus of the vestibular nerve was labeled.
Cells of the nucleus prepositus were la- beled in the same fashion as previously de- scribed, but in addition a few cells of the nucleus of Roller and the dorsal parame- dian reticular nuclei also contained HRP granules. Other brain stem nuclei labeled bilaterally were parts of the ACN, caudal parts of the LRN (fig. 21), caudal parts of the reticulotegmental nucleus and portions of the pontine nuclei.
194 D. RUGGIERO, R. BATTON 111, A. JAYARAMAN AND M. CARPENTER
,Lobule 91 c-1433
A
Lobule X 1
2
Fig. 2 Cat C-1433. Projection drawings of the cerebellum and brain stem demonstrating an HRP injection in the rostra1 part of the fastigial nucleus (cross-hatched area surrounding the needle track) and modest retrograde transport of the marker to cells of the brain stem (black dots). See figure 9.
Fastigial nucleus and posterior vernal cortex
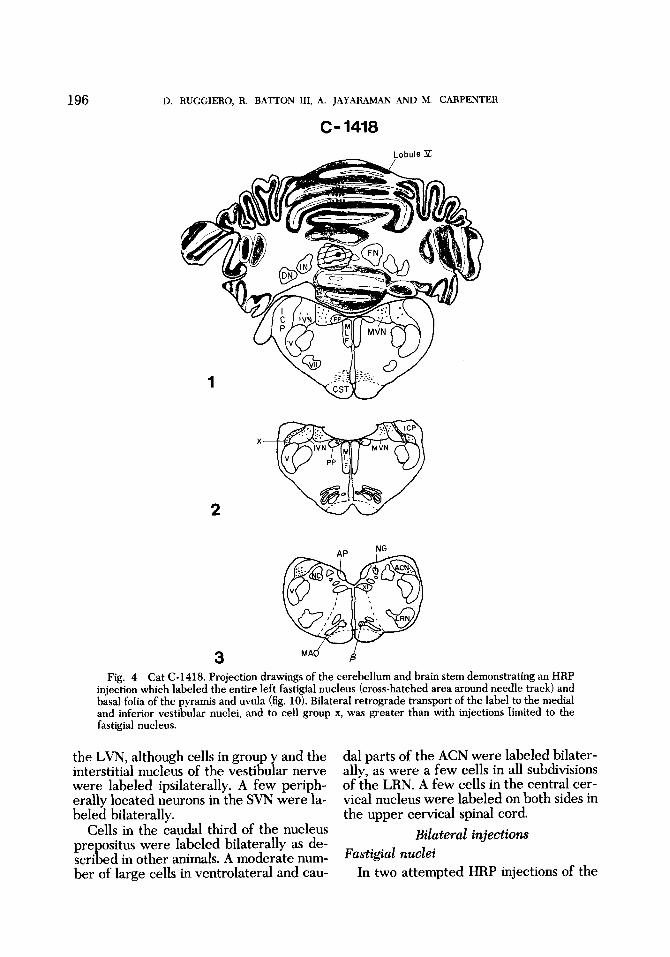
The attempted fastigial injection in cat (2-1418 labeled the entire left fastigial nu- cleus and basal folia of both the pyramis and uvula (figs. 4,lO). Retrograde transport of label to Purkinje cells and their dendrites was seen in a narrow band of the vermis on the left in folia of lobules V and VI (fig. 4). In the contralateral M A 0 la- beled cells were prominent in nucleus p and the dorsomedial cell column, but cells with HRP granules also were seen in both medial and lateral parts of the M A 0 at cau- dal levels.
Bilateral retrograde labeling of cells in the MVN and IVN was greater in this ani- mal than in any previously described (figs. 17, 18). The larges number of labeled cells were found in dorsal, lateral and caudal portions of the MVN; appreciably more cells were labeled on the injected side.
In the IVN labeled cells were found pre- dominately in dorsal and caudal regions, and were more numerous ipsilateral to the injection (figs. 4, 18). None of the large cells of group f were labeled on either side. The small and medium-sized cells of grou x were labeled bilaterally (figs. 4, 19, ZOf: No transport of label was seen in cells of
BRAIN STEM FASTIGIAL AFFERENTS PLATE
3
4
195
Fig. 3 Cat (2-1448. Projection drawings of the cerebellum and brain stem demonstrating an HRP injection which labeled caudal parts of the fastigial nucleus and rostrally labeled basal folia of lobule V (figs. 7,8). Drawings show retrograde transport to Purkinje cells in the ipsilateral vermis and in cer- tain brain stem nuclei.
196 D. RUGGIERO, R. BA'ITON 111, A. JAYARAMAN AND M. CARPENTER
C- 1418 Lobule P
1
2
X
3 Fig. 4 Cat C-1418. Projection drawings of the cerebellum and brain stem demonstrating an HRP
injection which labeled the entire left fastigial nucleus (cross-hatched area around needle track) and basal folia of the pyramis and uvula (fig. 10). Bilateral retrograde transport of the label to the medial and inferior vestibular nuclei, and to cell group x, was greater than with injections limited to the fastigial nucleus.
the LVN, although cells in group y and the interstitial nucleus of the vestibular nerve were labeled ipsilaterally. A few periph- erally located neurons in the SVN were la- beled bilaterally.
Cells in the caudal third of the nucleus prepositus were labeled bilaterally as de- scribed in other animals. A moderate num- ber of large cells in ventrolateral and cau-
dal parts of the ACN were labeled bilater- ally, as were a few cells in all subdivisions of the LRN. A few cells in the central cer- vical nucleus were labeled on both sides in the upper cervical spinal cord.
Bilateral injections Fastigial nuclei
In two attempted HRP injections of the

BRAIN STEM FASTIGIAL AFFERENTS PLATE 197
fastigial nucleus, the needle entered the ventral part of the pyramis slightly to the left of the midline and traversed only the medial part of the fastigial nucleus, or the area between the fastigial nuclei. There was HRP stain in medial portions of both fastigial nuclei. In cat C-1417 HRP was evident through the extent of the fastigial nuclei, but in cat C-1419 label was evident only in the caudal regions. Basal folia of the pyramis and uvula showed diffusion of HRP in both animals. Ret- rograde transport of the injected label was seen bilaterally in the somata and dendrites of Purkinje cells in folia of lobules V and VI in cat C-1417. In cat C- 1419, a few Purkinje cells were labeled only in portions of the uvula.
Brain stem nuclei The retrograde transport of HRP label
to brain stem nuclei following bilateral injections of the fastigial nuclei was similar to that described for unilateral injections.
Labeled cells in the IVN, predominately in dorsal and caudal regions, were dis- tributed symmetrically on the two sides in both animals. No transport of the enzyme was seen to cells in rostral parts of the IVN. Many cells of group x were labeled bilater- ally in an impressive fashion in both ani- mals. Neurons in dorsal parts of the h4VN were labeled bilaterally and symmetrically throughout their rostrocaudal extent. The largest number of labeled vestibular neu- rons was found in the MVN. Although no label was detected in cells of the LVN or SVN in one animal, labeled neurons were observed bilaterally in ventral parts of these nuclei in cat C-1417. Neurons la- beled bilaterally in the SVN were numer- ous and showed right predominance. No la- beled cells were seen in the interstitial nu- cleus of the vestibular nerve, but HRP granules were found in cell group y on the right in one animal (C-1419).
In the nucleus prepositus labeled cells were found bilaterally and symmetrically distributed in the caudal thirds of the nu- clei. When only the caudal halves of the fastigial nuclei were injected (C-1419), transport of the enzyme to cells of these
nuclei was scant, suggesting a regional dis- tribution of terminals in the fastigial nu- cleus.
Transport of HRP to cells in the inferior olivary complex in these animals was simi- lar to that in animals with unilateral fas- tigial injections, but was not always sym- metrical. Label was seen bilaterally in cell group /3 (figs. 15, 16) and the dorsomedial cell column in cat C-1417, but was seen only on the right side in cat C-1419.
Large cells in ventrolateral parts of the ACN contained HRP granules bilaterally. In cat C-1417 labeled cells in the ACN were numerous at all rostrocaudal levels. In the other animal labeled cells in the ACN were found mainly on the right side. Although some retrograde transport of la- bel was seen bilaterally in cells of all sub- divisions of the LRN in cat C-1417, no label was present in this nucleus in cat C-1419. A few cells of the central cervical nucleus were labeled bilaterally.
Cortical injections Attempted injections of the fastigial nu-
cleus in two cats resulted in labeling of re- gions of the cortex of the cerebellar ver- mis. In cat C-1449 the HRP injection la- beled basal folia in lobules V and VI and a small rostral part of the right fastigial nu- cleus. Direct uptake of HRP by Purkinje and granule cell somata was observed in a narrow band of cortex; a few cells of the fastigial nuclei on both sides exhibited retrograde transport of the enzyme.
Retrograde transport of the injected la- bel was found primarily in medial portions of the medial and dorsal accessory olivary nuclei on the left. None of the cells in the right IONC were labeled. The largest num- ber of labeled neurons were found in the caudal two-thirds ipsilateral ACN. Only a few labeled cells were seen in ventral por- tions of the opposite ACN. Central portions of the contralateral MVN and IVN con- tained only a few labeled cells. No labeled cells were identified in other nuclei of the vestibular complex on either side except for a few cells in the ipsilateral SVN. No transport of the enzyme was detected in the nucleus prepositus. Labeled pontine

198 D. RUGGIERO, R. BATTON 111, A. JAYARAMAN AND M. CARPENTER
nuclei, seen in both medial and lateral cell columns, were most numerous on the left side.
The attempted fastigial injection in cat C-1436 resulted in localized HRP labeling of several adjacent folia of the pyramis on the right side. Retrograde transport from these portions of the pyramis was detected contralaterally in medial and dorsal parts of the M A 0 adjacent to nucleus p , and bilaterally in portions of the pontine and reticulotegmental nuclei.
DISCUSSION
There is ample evidence that the retro- grade axonal transport of horseradish peroxidase (HRP) can be used to identify the cells of origin of afferent fibers in the central nervous system (LaVail and LaVail, '72, '74, '75; LaVail et al., '73; LaVail, '75; Jones and Leavitt, '74; Nauta et al., '74; Rinvik and Walberg, '75; P. Brodal, '75; Brodal et al., '75; Walberg et al., '76; Bro- dal, '76). Electron microscopic studies have demonstrated that at the injection site HRP is taken up by pinocytotic activity of nerve terminals and boutons, incorporated into small vesicles and membrane bound organelles, and transported in a retrograde fashion to somata and dendrites (Holtzman and Petersen, '69; LaVail and LaVail, '72, '74; LaVail, '75; Nauta et al., '75).
Injections of HRP into the cerebellum present unique problems because the nee- dle must traverse multiple folial interfaces before reaching the deep cerebellar nu- clei. These circumstances make it difficult to avoid inadvertent labeling of cortical areas. Similar difficulties have been de- scribed in attempts to label specific regions of the cerebellar cortex (Wallberg et al., '76; Brodal, '76). Even when these difficul- ties seem to be avoided by fortuitous cir- cumstances, the precise definition of the injection site is elusive in spite of the visi- ble brownish-yellow stain. Attempts to determine the size of the injection site by examining the tissue several days after injection yield only a rough estimate, since the process of clearing the enzyme begins immediately (Brightman, '65; LaVail and
LaVail, '74; LaVail, '75; Walberg et al., '76). It is widely acknowledged that the actual region from which HRP is trans- ported is considerably smaller than the stained zone, but this region is even more difficult to delimit. In the present study two criteria were used to indicate uptake of the HRP marker by terminals in the fastigial nucleus, namely, retrograde trans- port of the label to: (1)Purkinje cells in the ipsilateral vermis (figs. 1, 11, 12), and (2) specific portions of the contralateral inferior olivary complex (figs. 15, 16). Several experimental studies support the thesis that the vermal cortex projects to the fastigial nuclei (Eager, '66; Larsell and Jansen, '72) with the most profuse projec- tions arising from the anterior lobe of the cerebellum and from lobules VIII and IX. Studies of olivocerebellar fibers based upon the modified Gudden technic and retrograde axonal transport of HRP indi- cate that distinctive portions of the medial accessory olive (i.e., nucleus p and the dor- somedial cell column) and the dorsal cap of Kooy (a caudal medial part of the ventral lamella of the principal olivary nucleus) project fibers to the contralateral fastigial nucleus (Brodal, '40, '67, '76).
The most discrete and selective labeling of terminals in one fastigial nucleus (cats C- 1411 and C-1450) indicates that brain stem afferents to this nucleus are not numerous and arise largely from parts of the medial and inferior vestibular nuclei, parts of the inferior olivary nuclear complex and the nucleus prepositus (figs. 1, 13, 14). The lit- erature concerning secondary vestibulo- cerebellar fibers is inconsistent and con- tradictory (Brodal et al., '62). Following lesions in the fastigial nuclei retrograde cell changes have been described in the lateral, medial and superior vestibular nu- clei (Kuzume, '26), or in the superior vestibular nucleus only (Spaier, '36). The most definitive study of secondary vestib- ulocerebellar fibers suggested that these fibers arise predominately from ventrolat- era1 regions of the IVN, including cell group f, and from cell group x (Brodal and Torvik, '57). Some secondary vestibulocer-

BRAIN STEM FASTICIAL AFFERENTS PLATE 199
ebellar fibers also were described from the ventrolateral part of the MVN. The obser- vations of these authors indicated that sec- ondary vestibulocerebellar fibers pro- jected to the nodulus, the uvula, the flocculus and the fastigial nuclei. Evidence in the cat based upon isolated fastigial lesions indicated that crossed and un- crossed afferent fibers originated from the IVN (Carpenter et al., '59). Silver impregnation studies in the cat based upon discrete lesions in the IVN tended to confirm this observation (Carpenter, '60).
Current findings appear strikingly differ- ent from those based upon anterograde and retrograde degeneration studies in that: (1) the largest number of afferents from the vestibular nuclei arise from dorsal and cau- dal regions of the MVN, (2) the IVN ap- pears to give rise to relatively few fastigial afferent fibers and none of these arise from cell group f, (3) only a few neurons of cell group x appear to pro'ect to the ipsilateral fastigial nucleus, and t 4) none of the cells in the LVN, SVN, cell group y or the interstitial nucleus of the vestibular nerve appear to project to the fastigial nucleus.
Aside from the above described vestibu- lar projections and those from the con- tralateral IONC, the only additional pro- jection to the fastigial nucleus arises bilat- erally from the caudal third of the nucleus prepositus. Observations concerning the nucleus prepositus are consistent with the findings of Brodal ('52), but serve to distin- guish this nucleus from the other perihypo- glossal nuclei which apparently do not pro- ject terminals to the fastigial nucleus. This finding is different from prior studies which suggested that all subdivisions of the perihypoglossal nuclei projected fibers to the entire cerebellar receiving area, said to consist of the anterior lobe, the p ramis, the uvula and the fastigial nuclei (korvik and Brodal, '54).
Limited data concerning brain stem af- ferents to particular parts of the fastigial nucleus suggest that the rostral half of this nucleus (C-1433) receives fibers bilaterally mainly from the caudal regions of the MVN (fig. 2). Judging from the transport of the
marker to cells of the MVN, these fibers are not numerous and have a contralateral pre- dominance. Caudal portions of the fastigial nucleus ((2-1448) receive more numerous fibers bilaterally from dorsal regions of both the MVN and IVN.
In several animals with unilateral HRP injections of the fastigial nucleus a few cells in the contralateral nucleus exhibited retrograde transport of the marker (fig. 22). These cells were found mainly in ros- tral parts of the fastigial nucleus. While it is tempting to regard these cells as giving rise to possible commissural fibers, it seems more likely that retrograde axonal trans- port might be via collaterals. Collaterals of this type were not described in a detailed Golgi study of the fastigial nucleus (Mat- sushita and Iwahori, '71). It is possible that label in these contralateral fastigial neu- rons, as well as in some ipsilateral neurons, may be a consequence of uptake of HRP by cortical neurons, as reported by several au- thors (Walberg, '76; Tolbert et al., '76; Gould and Graybiel, '76). Since there is no evidence that HRP can be transported across synapses (LaVail et al., '73; LaVail, '75a), it seems likely that cells of the deep cerebellar nuclei may project fibers, or at least collaterals, to the cerebellar cortex. Systematic studies of the deep cerebellar nuclei after HRP injections of superficial regions of the cortex tend to confirm this finding. Injections of lobules V, VI and VII reportedly produced bilateral labeling of fastigial neurons, especially at rostral levels (Gould and Graybiel, '76). In general there appears to be an orderly arrangement of projections from the deep nuclei to the cerebellar cortex as determined by retro- grade axonal transport of HRP and antero-
rade transport of C3H] amino acids 7 Tolbert et al., '76). There are no data sug- gesting that the deep cerebellar nuclei exhibit endogenous preoxidase activity.
Unilateral injections of the fastigial nu- cleus and basal folia of the pyramis and uvu- la (C-1418) resulted in bilateral retrograde labeling of cells in the MVN, IVN and cell group x (figs. 4,17-20). Virtually all cells of group x were labeled bilaterally in a sym-

200 D. RUGGIERO, R. BATTON 111, A. JAYARAMAN AND M. CARPENTER
metrical fashion. This paravestibular nu- cleus does not receive primary vestibular fibers (Walberg et al., '58) but it receives crossed fastigial projections (Walberg et al., '62) and fibers that ascend in the lateral funiculus of the spinal cord (Pompeiano and Brodal, '57). Data from this study sug- gest that while a few fibers from cell group x probably project to the fastigial nucleus, one of the major projections of this nucleus appears to be the vermal cortex of lobules VIII and IX.
Combined HRP labeling of the fastigial nucleus and parts of lobules VIII and IX re- sults in far more extensive transport of the marker to cells of the MVN and IVN than seen with discrete injections of the fastigial nucleus. This suggests that a larger number of vestibular neurons project to the vermal cortex of lobules VIII and IX. None of the large cells of group f contained the enzyme marker after these combined injections. Cell group f, like cell group x, does not receive primary vestibular fibers (Walberg et al., '58), but it receives a massive crossed projection from the fastigial nucleus via the uncinate fasciculus (Walberg et al., '62). Al- though cell group f has been considered one of the principal sources of secondary vestibulocerebellar fibers (Brodal and Trovik, '57), current observations indicate that this nucleus probably does not project fibers to the fastigial nucleus, or to lobules VIII and IX.
Observations from inadvertent bilateral HRP injections of the fastigial nuclei confirm the finding that the principal ves- tigial afferents to the fastigial nucleus arise from the MVN. The identification of a large number of cells in the superior vestibular nucleus labeled bilaterally in another animal ((2-1417) offers an unex- plained inconsistency, perhaps related to diffusion of HRP.
Although cortical injections of HRP do not provide data concerning the origins of fastigial afferents, they serve as a kind of control and supply comparative data. La- beling of basal folia of lobules V and VI re- sulted in retrograde labeling of neurons in the accessory cuneate nucleus, as well as in
parts of the medial and dorsal accessory olivary nuclei contralaterally. Labeling of cells in the accessory cuneate nucleus also was seen in other animals in which HRP diffused into parts of lobules V and VI (fig. 4). The above findings are consistent with the observations of Rinvik and Walberg ('75) concerning the cerebellar projections of the accessory cuneate nucleus. These authors also reported that cells of the main cuneate nucleus project axons to the super- ficial parts of folia in lobule V. In this study no neurons of the cuneate nuclei were la- beled.
Several animals in this study showed retrograde transport of HRP label to some cells in all subdivisions of the lateral retic- ular nucleus (LRN), but the largest number of such cells was found in the magnocellu- lar division (Walberg, '52). Per Brodal ('75) determined that the magnocellular part of the LRN projects to caudal portions of the anterior lobe of the cerebellum and to ros- tral parts of the paramedian lobule. An autoradiographic study of the LRN sug- gests an extensive projection to lobules I through VIII (Kunzle, '75). While label in cells of the LRN clearly was related to injections in caudal parts of the anterior lobe in some animals, the possibility re- mains that part of the transported label may have come from portions of lobule VIII which was traversed by the injection needle (figs. 4, 21). Labeled cells in por- tions of the pontine and reticulotegmental nuclei were seen in several animals in which portions of the vermal cortex had taken up the enzyme. Bilateral transport to portions of the pontine nuclei was most fre- quent. Available evidence did not suggest that these nuclei project to the fastigial nu- cleus.
There is evidence from this study that the central cervical nucleus projects fibers to vermal cortex, although few of these fibers appear to terminate in the fastigial nucleus. The largest number of fibers from the central cervical nucleus appear to ter- minate in the anterior lobe of the cerebel- lum (Wiksten, '75; Matsushita and Ikeda, '75), but their precise distribution in the

BRAIN STEM FASTIGIAL AFFERENTS PLATE 201
cortex might best be determined by auto- radiographic studies.
This study of the fastigial nucleus sug- gests that the principal afferents arise from: (1) Purkinje cells of the ipsilateral cerebellar vermis, (2) restricted portions of the contralateral medial accessory olivary nucleus (i.e., nucleus f3 and the dorsomedial cell column), (3) the nucleus prepositus (bilateral), and (4) portions of the medial and inferior vestibular nuclei. Secondary vestibular fibers projecting to the fastigial nucleus arise bilaterally, primarily from dorsal and caudal portions of the medial vestibular nucleus. Smaller numbers of fastigial afferents arise from dorsal regions of the inferior vestibular nucleus. None of the large neurons of cell group f project to the fastigial nucleus. Although secondary vestibular fibers passing to the fastigial nu- cleus arise bilaterally, the bulk of the fibers appear to be uncrossed. A few neurons in cell group x project to the fastigial nucleus, but a large number of these cells project to the cortex of lobules VIII and IX.
Although Dow ('36) estimated that sec- ondary vestibulocerebellar fibers were three times as numerous as primary vestib- ulocerebellar fibers, it is apparent that sec- ondary fibers which end in the fastigial nu- cleus are modest in number. Primary vestibular fibers terminating in the fastigial nucleus have not been demonstrated con- vincingly by degeneration technics (Brodal and Haivik, '64; Carpenter et al., '72) or in GoIgi studies (Larsell, '70). These observa- tions suggest that primary and secondary vestibular inputs to the fastigial nucleus probably are relayed by Purkinje cells in the cerebellar cortex.
LITERATURE CITED
Brightman, M. W. 1965 The distribution within the brain of ferritin injected into the cerebrospinal fluid compartments. 11. Parenchymal distribution. Am. J. Anat., 11 7: 193-200.
Brodal, A. 1940 Experimentelle Untersuchungen uber die olivocerebellare Lokalisation. Z. ges. Neur. Psychiat., 169: 1-153.
1952 Experimental demonstration of cere- bellar connexions from the peri-hypoglossal nuclei (nucleus intercalatus; nucleus praepositus hypo-
glossi and nucleus of Roller) in the cat. J. Anat., 86:
Anatomical studies of cerebellar fibre connections with special reference to problems of functional localization. In: Progress in Brain Re- search, The Cerebellum. C. A. Fox and R. S. Snider, eds. Elsevier Publishing Co., Amsterdam, 23: 135- 173.
1976 The olivocerebellar projection in the cat studied with the method of retrograde axonal transport of horseradish peroxidase. 11. The projec- tion to the uvula. J. Comp. Neur., 166: 417-426.
Brodal, A., and B. Haivik 1964 Site and mode of ter- mination of primary vestibulocerebellar fibres in the cat. An experimental study with silver impreg nation methods. Arch. ital. Biol., 102: 1-21.
Brodal, A., and 0. Pompeiano 1957 The vestibular nuclei in the cat. J. Anat., 91: 438-454.
Brodal, A., 0. Pompeiano and F. Walberg 1962 The Vestibular Nuclei and their Connections, Anatomy and Functional Correlations. C. C Thomas, Springfield, Ill., 193 pp.
Brodal, A., and A. Torvik 1957 Ueber den Ursprung der sekundaren vestibulocerebetlaren Fasern bei der Katze. Eine experimentell-anatomische Studie. Arch. Psychiat. Ztschr. ges. Neur., 195: 550-567.
Brodal, A., F. Walberg and G. H. Hoddevik 1975 The olivocerebellar projection in the cat studied with the method of retrograde axonal transport of horse- radish peroxidase. J. Comp. Neur., 164: 449-470.
Brodal, P. 1975 Demonstration of a somatotopical- ly organized projection onto the paramedian lobule and the anterior lobe from the lateral reticular nu- cleus: An experimental study with the horseradish peroxidase method. Brain Res., 95: 221-239.
Carpenter, M. B., 1960 Fiber projections from the descending and lateral vestibular nuclei in the cat. Am. M. Anat., 107: 1-22.
Carpenter, M. B., D. S. Bard and F. A. Alling 1959 Anatomical connections between the fastigial nu- clei, the labyrinth and the vestibular nuclei in the cat. J. Comp. Neur., 1 1 1 : 1-26.
Carpenter, M. B., B. M. Stein and P. Peter 1972 Pri- mary vestibulocerebellar fibers in the monkey: Dis- tribution of fibers arising from distinctive cell groups of the vestibular ganglia. Am. J. Anat., 135:
Dow, R. S. 1936 The fiber connections of the pos- terior parts of the cerebellum in the cat and rat. J. Comp. Neur.. 63: 527-548.
Eager, R. P. 1966 Patterns and mode of termina- tion of cerebellar corticonuclear pathways in the monkey. J. Camp. Neur., 126: 551-566.
Could, B. B., and A. M. Graybiel 1976 Afferents to the cerebellar cortex in the cat: Evidence for an intrinsic pathway leading from the deep nuclei to the cortex. Brain Res., 110: 601-611.
Graham, R. C. Jr., and M. J. Karnovsky 1966 The ear- ly stages of absorption of injected horseradish peroxidase in the proximal tubules of mouse kidney: Ultrastructural cytochemistry by a new technique. J. Histochem. Cytochem., 14: 291-302.
110-129. 1967
221-250.

202 D. RUGGIERO, R. BATTON 111, A.
Holtzman, E., and E. R. Petersen 1969 Uptake of protein by mammalian neurons. J. Cell Biol., 40:
Jones, E. G., and R. Y. Leavitt 1974 Retrograde axonal transport and the demonstration of non- specific projections to the cerebral cortex and striatum from thalamic intralaminar nuclei in the rat, cat and monkey. J. Comp. Neur., 154: 349-378.
Kim, C. C., and P. L. Strick 1976 Critical factors involved in the demonstration of horseradish perox- idase retrograde transport. Brain Res., 103: 356- 361.
Khzle, H. 1975 Autoradiographic tracing of the cerebellar projections from the lateral reticular nu- cleus in the cat. Exper. Brain Res., 22: 255-266.
Kuzume, G. 1926 Experimentell-anatomische Un- tersuchungen uber die inneren und ausseren ver- bindungen des Flocculus und der Kleinhirnkerne (Hauptsachlich des Dachkerns). Folia anat. Jap., 4:
Larsell, 0. 1953 The cerebellum of the cat and the monkey. J. Comp. Neur., 99: 135-200.
Larsell, 0. 1970 The Comparative Anatomy and Histology of the Cerebellum from Monotremes through Apes. J. Jansen (ed.), University of Min- nesota Press, Minneapolis, Minnesota, p. 18.
Larsell, O., and J. Jansen 1972 The Comparative Anatomy and Histology of the Cerebellum. The Human Cerebellum, Cerebellar Connections and Cerebellar Cortex. University of Minnesota Press, Minneapolis, Minnesota.
La Vail, J. H. 1975 Retrograde cell degeneration and retrograde transport techniques. In: The Use of Axonal Transport for Studies of Neuronal Connec- tivity. W. M. Cowan and M. CuCnod (eds.), Elsevier Scientific Publishing Co., Amsterdam, Chap. 10, pp.
La Vail, J. H. 1975a The retrograde transport method. Fed. Proc., 34: 1618-1624.
La Vail, J. H., and M. M. La Vail 1972 Retrograde axonal transport in the central nervous system. Science, 176: 1416-1417.
La Vail, J. H., and M. M. La Vail 1974 The retrograde intraaxonal transport of horseradish peroxidase in the chick visual system. A light and electron micro- scopic study. J. Comp. Neur., 157: 303-358.
La Vail., M. M., and J. H. La Vail 1975 Retrograde intraaxonal transport of horseradish peroxidase in retinal ganglion cells of the chick. Brain Res., 85:
La Vail., J. H., K. R. Winston and A. Tish 1973 A method based on retrograde intraaxonal transport of protein for identification of cell bodies of origin of axons terminating within the CNS. Brain Res., 58:
Matsushita, M., and M. Ikeda 1976 The central cervi- cal nucleus as cell origin of a spinocerebellar tract
863-869.
75-110.
217-247.
273-280.
470-477.
JAYARAMAN AND M. CARPENTER
arising from the cervical cord: A study in the cat using horseradish peroxidase. Brain Res., 100: 412- 417.
Matsushita, M., and N. Iwahori 1971 Structural organization of the fastigial nucleus. I. Dendrites and axonal pathways. Brain Res., 25: 597-610.
Nauta, H. J. W., I. R. Kaiserman-Abramof and R. J. Lasek 1975 Electron microscopic observations of horseradish peroxidase transported from the caudoputamen to the substantia nigra in the rat: Possible involvement of the agranular reticulum. Brain Res., 85: 373-384.
1974 Afferents to the rat caudoputamen studied with horseradish peroxidase. An evaluvation of a retrograde neuroanatomical research method. Brain Res., 67: 219-238.
Pompeiano, O., and A. Brodal 1957 Spino-vestibular fibers in the cat. An experimental study. J. Comp. Neur., 108: 353-382.
Rinvik, E., and F. Walberg 1975 Studies on the cere- bellar projections from the main and external cune- ate nuclei in the cat by means of retrograde axonal transport of horseradish peroxidase. Brain Res., 95:
Spaier, E. L. 1936 Zur Frage der Verbindungen zwischen dem Vestibularisverven und dem Klein- him. Quoted by Abl. ges Neur. Psychiat., 82: 555.
Tolbert, D. L., H. Bantli and J. R. Bloedel 1976 A cer- ebellar nucleo-cortical projection in the cat. Anat. Rec., 184: 547.
Torvik, A., and A. Brodal 1954 The cerebellar pro- jection of the perihypoglossal nuclei (nucleus inter- calatus, nucleus praepositus and nucleus of Roller) in the cat. J. Neuropath. and Exper. Neur., 13: 515- 527.
Walberg, F. 1952 The lateral reticular nucleus of the medulla oblongata in mammals. A comparative- anatomical study. J. Comp. Neur., 96: 283-343.
Walberg, F., D. Bowsher and A. Brodal 1958 The termination of primary vestibular fibers in the vestibular nuclei in the cat. An experimental study with silver methods. J. Comp. Neur., 110: 391-419.
Walberg, F., A. Brodal and G. H. Hoddevik 1976 A note on the method of retrograde transport of horseradish peroxidase as a tool in studies of afferent cerebellar connections, particularly those from the inferior olive; with comments on the orthograde transport in Purkinje cell axons. Exp. Brain Res., 24: 383-401.
Walberg, F., 0. Pompeiano, A. Brodal and J. Jansen 1962 The fastigiovestibular projection in the cat. An experimental study with silver impregnation methods. J. Comp. Neur., 118: 49-76.
Wiksten, B. 1975 The central cervical nucleus-A source of spinocerebellar fibres, demonstrated by retrograde transport of horseradish peroxidase. Neurosci. Letters, I : 81-84
Nauta, H. J. W., M. B. Pritz and R. J. Lasek
371-381.

PLATES

PLATE 1
EXPLANATION OF FIGURES
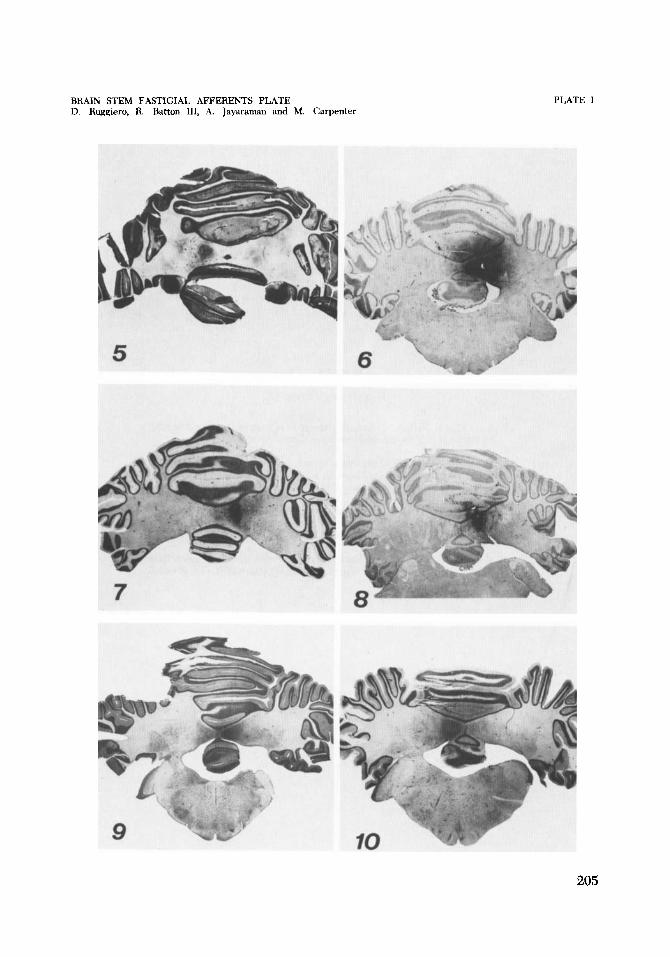
5 Cat C-1411. Photomicrograph of HRP injection that selectively labeled the entire left fastigial nucleus. Cresyl violet. x 5.
6 Cat (2-1450. Photomicrograph of a fairly selective HRP injection of the entire right fastigial nucleus. See figure 1 for drawings of brain stem neu- rons labeled by retrograde transport. Cresyl violet. X 2.5.
7 , 8 Cat (2-1448. Photomicrographs of an HRP injection that labeled mainly the caudal half of the right fastigial nucleus. Rostrally the needle entered the basal folio of lobule V. See figure 3. Cresyl violet. x 5, x 5.
9 Cat C-1433. Photomicrograph of an HRP injection in the rostra1 half of the right fastigial nucleus. See figure 2. Cresyl violet. x 2.5.
10 Cat C-1418. Photomicrograph of an HRP injection in the left fastigial nu- cleus that also labeled basal folio of the pyramis and uvula. See figure 4 for diagrams of retrograde transport of the label. Cresyl violet. x 2.5.
204
BRAIN STEM FASTIGIAL AFFERENTS PLATE D. Ruggiero, R. Batton 111, A. Jayaraman and M. Carpenter
PLATE 1
205

PLATE 2
EXPLANATION OF FIGURES
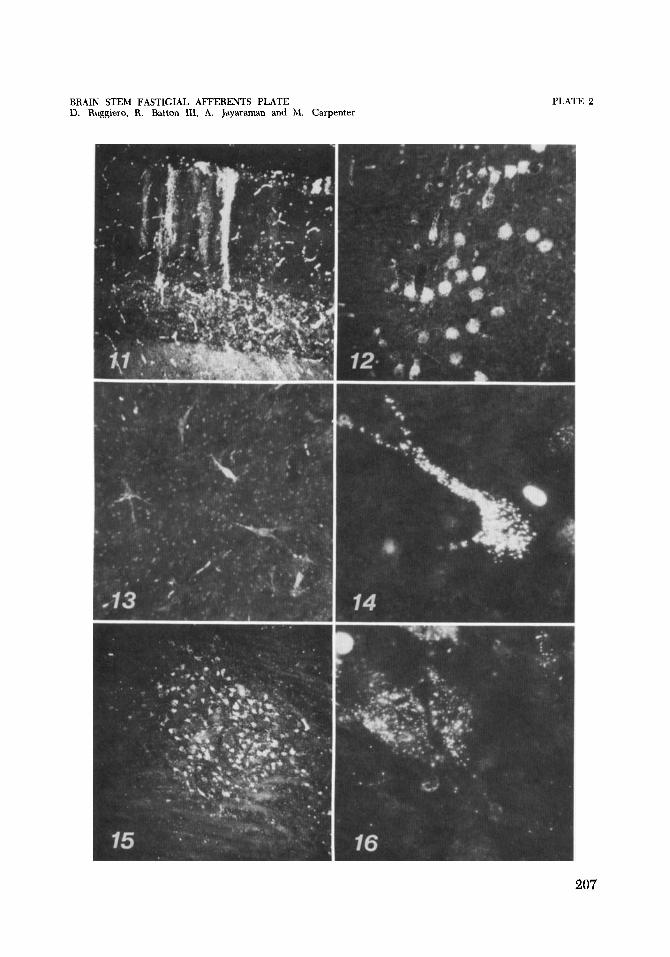
11 Cat C-1411. Dark field photomicrograph of retrograde transport of HRP to Purkinje cell somata and dendrites. Cresyl violet. x 52.
12 Cat C-1450. Dark field photomicrograph of a narrow band of the cere- bellar vermis in which Purkinje cell somata have been labeled following an ipsilateral fastigial injection of HRP. Cresyl violet. X 133.
13,14 Cat C-1411. Dark field photomicrographs demonstrating retrograde transport of the marker to neurons in the ipsilateral medial vestibular nucleus. Cresyl violet. X 133, X 325.
Cat C-1417. Dark field photomicrographs of label transported to cells of nucleus p following an HRP injection of the fastigial nuclei. Cresyl violet. X 52. X 325.
15, 16
206
BRAIN STEM FASTIGIAL AFFERENTS PLATE 1). Ruggiero, R. Ratton 111, A. fayamman and M. Carpenter
PLATE 2
207

PLATE 3
EXPLANATION OF FIGURES
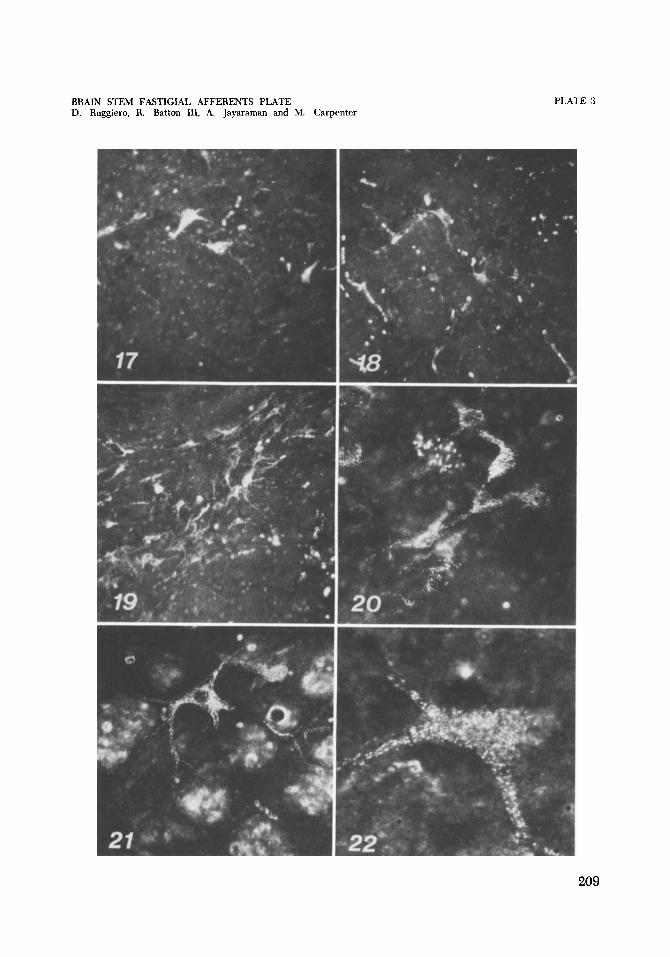
17, 18 Cat C-1418. Dark field photomicrographs of retrograde transport of HRP to cells in dorsal regions of the medial (17) and inferior (18) vestibular nuclei. See figure 4. Cresyl violet. x 133, x 133.
Cat C-1418. Dark field photomicrographs of retrograde transport of la- bel to cell group x on the left following an HRP injection of the left fastigial nucleus and basal folia of the pyramis and uvula. Cresyl violet. X 133, X 210.
Cat C-1448. Dark field photomicrograph of retrograde transport of the protein marker to large cells of the lateral reticular nucleus of the me- dulla. Cresyl violet. x 500.
Cat C-1448. Dark field photomicrograph of retrograde transport of label to a cell in the contralateral fastigial nucleus following an injection of the fastigial nucleus and basal folia of lobule V. Cresyl violet. x 325.
19,20
21
22
208
BRAIN STEM FASTIGIAL AFFERENTS PLATE D. Ruggiero, R. Batton 111, A. Jayaraman and M. Carpenter
PLATE 3
209