bovine viral diarrhea virus induced apoptosis correlates with increased intracellular viral rna...
TRANSCRIPT

Virus Research 69 (2000) 95–107
Bovine viral diarrhea virus induced apoptosis correlates withincreased intracellular viral RNA accumulation�
Ventzislav B. Vassilev, Ruben O. Donis *Department of Veterinary and Biomedical Sciences, Uni6ersity of Nebraska, 202 VBS, Lincoln, NE 68583-0905, USA
Received 19 January 2000; received in revised form 26 May 2000; accepted 6 June 2000
Abstract
Non-cytopathic (NCP) and cytopathic (CP) parent–daughter pairs are often isolated from cattle with bovine viraldiarrhea virus (BVDV) induced mucosal disease. Alignment of these pair genomes revealed that genetic changes in CPBVDV involve the NS2-3 coding region and correlate with expression of NS3. However, additional mutations arepresent elsewhere in the genomes of these natural pairs, precluding unambiguous mapping of this function to theNS2-3 region. To evaluate this phenomenon in identical genetic backgrounds, we have constructed an NCP isogenicpair of the NADL by deletion of the cIns from NS2 region. The levels of viral protein synthesis in infected cellsrevealed no marked difference between the CP and the isogenic NCP BVDV mutant. In contrast, RNA accumulationin cells infected with CP virus was up to 25 times higher than that in cells infected with NCP BVDV. No significantdifference in growth kinetics and viral yields were observed between the CP BVDV and the isogenic NCP pair.Analyses of additional NCP/CP parent–daughter field BVDV isolates revealed a similar pattern of macromolecularsynthesis, suggesting the generality of this phenomenon. These results implicate increased levels of RNA accumulationin CP BVDV infected cells, along with the production of NS3 as potential contributors to viral cytopathogenicity.© 2000 Published by Elsevier Science B.V.
Keywords: Bovine viral diarrhea virus; Replication; Pestivirus; RNA
www.elsevier.com/locate/virusres
1. Introduction
Bovine viral diarrhea virus (BVDV) is the pro-totype species in the Pesti6irus genus of the Fla-viviridae. The single stranded RNA genome of
BVDV has mRNA polarity and is approximately12 500 nucleotides (nt) in length. The RNA isneither polyadenylated nor capped (reviewed inDonis, 1995). The genomic RNA is infectious;cells transfected with RNA produce viableprogeny virions. The BVDV genome comprises asingle large open reading frame (ORF) flanked byuntranslated regions (5%-UTR and 3%-UTR, re-spectively). The 385-nt long 5%-UTR is essentialfor viral protein synthesis by virtue of its functionas an internal ribosomal entry site (IRES), direct-
� Publication 12812 of the Agricultural Research Division,University of Nebraska.
* Corresponding author. Tel.: +1-402-4726063; fax: +1-402-4729690.
E-mail address: [email protected] (R.O. Donis).
0168-1702/00/$ - see front matter © 2000 Published by Elsevier Science B.V.
PII: S0168-1702(00)00176-3

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–10796
ing internal translation initiation (Poole et al.,1995; Ali and Siddiqui, 1997; Fukushi et al., 1997;Rijnbrand et al., 1997; Vassilev et al., 1997; Chonet al., 1998). The 5% terminus is also involved inRNA replication functions (Lin et al., 1994b;Frolov et al., 1998).
Upon synthesis, the BVDV polyprotein is pro-cessed into mature functional proteins by a com-bination of host and viral proteases. The order ofmature proteins is as follows, from N-terminus toC-terminus: Npro, C, E0, E1, E2, p7, NS2-3,NS4A, NS4B, NS5A, NS5B. The first protein,Npro, is an autoprotease, which generates its ownC terminus by cleaving itself away from thepolyprotein. The structural proteins (C, E0, E1and E2), as well as p7, which can not be found inthe virions, are generated by proteolytic cleavagemediated by host signal peptidases (Lin et al.,1994a; Elbers et al., 1996). The serine proteasedomain of NS3 or NS2-3 completes the process-ing of the non-structural proteins.
The non-structural proteins catalyze transcrip-tion of the positive-strand RNA genome intofull-length negative strand intermediate and sub-sequent synthesis of positive-strand genomicRNA progeny. Analysis of RNA accumulation incells infected with different BVDV strains hasrevealed a wide spectrum of RNA synthesis levels(Vassilev, unpublished data). It is not known howBVDV RNA steady state levels are regulated incells infected with CP or NCP isolates. In prelim-inary studies we observed significantly differentlevels of RNA accumulation between the mem-bers of BVDV NCP/CP pairs isolated from casesof mucosal disease, suggesting a linkage betweenthese two phenotypic characters. Previous workhas established that bovine cells infected withcytopathic BVDV undergo apoptosis (Hoff andDonis, 1997). In this study we explored the rela-tionship between the putative pro-apoptotic phe-notype of CP BVDV and levels of viralmacromolecular synthesis in cells infected withisogenic CP and NCP strains of BVDV derived byreverse genetics. The recent successes with reversegenetic systems for pestiviruses (Meyers et al.,1996; Moormann et al., 1996; Ruggli et al., 1996;Vassilev et al., 1997; Xu et al., 1997) provided thetools for a direct approach to address the issue.
Out data indicate that expression of CP pheno-type by BVDV coincides with a remarkable in-crease of RNA synthesis as compared with NCPBVDV, whereas viral protein synthesis levels andgrowth kinetics remained unchanged. Thus, in-creased levels of RNA synthesis, along with theproduction of NS3 can contribute to induction ofapoptosis and expression of the CP phenotype bycertain BVDV strains.
2. Materials and methods
2.1. Cells, 6iruses, 6ectors and hosts
Embryonic bovine trachea (EBTr) and CV-1cells were from American Type Culture Collection(CCL-44 and CCL-70, respectively). Bovine testi-cle fibroblasts (BT cells) were derived as described(Donis and Dubovi, 1987a). Bovine cells werecultured in minimum essential medium (MEM)supplemented with fetal equine serum (FES) (No-vatech). Vaccinia virus MVA-T7 was provided byB. Moss and propagated in CV-1 cells (Wyatt etal., 1995). BVDV strain 9a-CP/8b-NCP, JV4-CP/JV4-NCP was obtained from Edward Dubovi(Cornell University, Ithaca, NY) (Tautz et al.,1996; Corapi et al., 1988). Strains Thompson-CPand Thompson-NCP were kindly provided byFernando Osorio (Nebraska Veterinary Diagnos-tic Center, University of Nebraska-Lincoln).
Escherichia coli K12 GM2163 [F- ara-14, leuB6,thi-1, fhuA31, lacY1, tsx-78, galK2, galT22,supE44, hisG4, rpsL136, (Strr), xyl-5, mtl-1,dam13::Tn9(Camr), dcm-6, mcrB1, hsdR2(rk
−mk+),
mcrA] is from New England Biolabs, XL-10Gold [D(mcrA)183, D(mcrCB-hsdSMR-mrr)173,endA1, supE44, thi-1, recA1, gyrA96, relA1, lac,Tetr, Hte[F’ proAB, lacIqZDM15, Tn10(Tetr),Amy, Camr] and Sure [e14−(mcrA), D(mcrCB-hs-dSMR-mrr)171, endA1, supE44, thi-1, gyrA96,relA1, lac, recB, recJ, sbcC, umuC:Tn5(Kanr),uvrC, [F’ proAB, lacIqZDM15, Tn10, (Tetr)] arefrom Stratagene, DH5a [F-, f80dlacZDM15,D(lacZYA-argF)U169, endA1, recA1, hsdR17(rk
−
mk+), deoR, thi1, supE44, l-, gyrA96, relA1] is
from Stratagene and MC1061 [DE(araA-leu)7697,DE(codB-lacI)3, araD139, galU, galK16, hsdR2,

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107 97
rpsL, l-, mcrA0, relA1, spoT1, mcrB9999] is fromE. coli Genetic Stock Center at Yale University.pACYC177 cloning vector was obtained fromNew England Biolabs (Rose, 1988).
2.2. Plasmid construction
To construct pNADLp15 we excised a HaeIIfragment from the plasmid pVV NADL (Vassilevet al., 1997), containing entire cDNA of theBVDV strain NADL genome and the b-lactamasegene of the vector pGEM-4 (Promega). This frag-ment was ligated into the large HaeII fragment ofpACYC177 cloning vector, containing p15Aorigin of replication and b-lactamase (Rose,1988). The construct in which both b-lactamasegenes were ligated ‘head to tail’ was namedpNADLp15 and chosen for all further manipula-tions. To delete a 270- nucleotide segment extend-ing from position 4994 to 5263 encoding thecellular insert (cIns) in the NS2 region, we sub-cloned the region of interest from pVVNADL asan RsrII-HpaI fragment into a window created inthe pCV77 vector with the same endonucleases(the nucleotide numbering used is as in Colett etal. (1988). This intermediate plasmid, namedpCV-HR, was used as a template for inverse PCRusing the following phosphorylated primers:
FBVD5263 5%-CCTTTCAGGCAGGAATA-CAATG 3% andRBVD4974 5%-ACCCTCAAAGTTGCCTTC-CC-3%.Ligation of the PCR product and transforma-
tion into JM109 E. coli competent cells gave riseto pCV-HRdIns. The RsrII-HpaI fragment frompNADLp15 was exchanged with RsrII-HpaI frag-ment from pCV-HR dIns, resulting in pN-dIns.
2.3. Plaque and focus-forming assays
BVDV infectivity titers were determined byplaque or focus forming assays. Briefly, CPBVDV was serially diluted in MEM plus 5% FES.BT cells plated in 6 well tissue culture plates(1.6×105 per well), were infected at 37°C with 0.5ml of each dilution. One hour after infection BTmonolayers were overlaid with 1% low-melting-point agarose in MEM containing 5% FES.
Plaques were visualized at 96 h post-infection byfixing and staining cell monolayers with 0.05%crystal violet in 10% formalin. For quantitation ofNCP BVDV foci of infected cells were visualizedby immunostaining. To this end, the agarose over-lay was carefully removed, and cells were fixed byincubation for 15 min at 22°C in a phosphatebuffered saline (PBS) (pH 7.4) containing 30%acetone and supplemented with 100 mg/ml bovineserum albumin (BSA). Fixed monolayers were airdried, rehydrated in PBS and blocked by incuba-tion for 1 h in PBS containing 4% skim milk.Monolayers were sequentially incubated with goatanti-BVDV polyvalent antiserum (VMRL Inc.)followed by HRPO-conjugated Protein G. Bothwere diluted in PBS containing 0.01% Tween-20and 0.5 M NaCl. Bound conjugates were revealedwith the peroxidase substrate 3-amino-9-ethylcar-bazole (Sigma).
2.4. RNA transcription and transfection
Full-length NADL RNA transcripts were pre-pared from pNADLp15 or pN-dIns plasmidDNA linearized with SacII or from full-lengthNADL PCR products generated from the aboveplasmids by using T7 RNA polymerase system asdescribed previously according to recommenda-tions of the manufacturer (Vassilev et al., 1997)(MEGAscript, Ambion). Electroporation of invitro transcribed and purified RNA into EBTRcells was performed using 1 mg of in vitro-tran-scribed RNA and 2×106 EBTR cells in 200 mlDulbecco’s MEM in a 0.2-cm cuvette as describedpreviously (ECM-600, BTX) (Vassilev et al.,1997).
Liposome mediated transfection of PCR ampli-con DNA was carried out with cationic lipidaccording to the manufacturer’s protocol (Lipo-fectAMINE, Life Technologies).
2.5. Long PCR
PCR amplification of the full-length cDNA ofBVDV from medium-copy number plasmids withp15A replicons, was performed in 25 ml reactionmix using KlenTaq-LA polymerase mix (Clon-tech) (Barnes, 1994; Cheng et al., 1994). The

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–10798
reaction mix contained 1× KlenTaq-LA poly-merase buffer, 1.3% DMSO, 0.4 M betaine (Sigma),200 mM of each dNTP, 1U KlenTaq-LA poly-merase mix, 10 pmol of each primer (Forward,BVDV-T7, 5%-TAATACGACTCACTATAGTA-TACG-3% and reverse, NADL-3%, 5%-GGGGGCT-GTTAGAGGTCTTC-3%), and 5 fmol plasmidDNA. The PCR consisted of an initial 4-mindenaturation step at 94°C, followed by 27 cycles,each composed of 1 min denaturation at 93°C, 1min annealing at 57°C, 13 min extension, and afinal 20 min extension at 72°C.
2.6. Western blotting
Proteins were separated by SDS-PAGE (8%polyacrylamide), transferred to a Hybond™-C sup-ported nitrocellulose membrane (Amersham LifeSciences) in a semi-dry apparatus (Trans-Blot,Bio-Rad) and processed as described previously(Ausubel et al., 1987). For detection of NS3 (orNS2-3), filters were probed first with murine mon-oclonal antibody 20.10.6. (1:10 000), which is spe-cific for the pestivirus non-structural protein p80(NS3), and the bound antibodies were detected byHR peroxidase-conjugated rabbit anti-mouse im-munoglobulin (1:500) (Corapi et al., 1990; Floreset al., 1996). A chemiluminescent substrate systemwas used to detect the anti-Ig conjugates by captur-ing the images on X-ray film (ECL, Amersham).
2.7. Northern blotting
Total RNA was isolated from BT cells by themethod of Chomczynski and Sacchi (1987) (LifeTechnologies). RNA was resuspended in 1×DNase buffer (20 mM Tris–HCl, pH 8.4, 2 mMMdCl2, 50 mM KCl) and incubated for 30 min at37°C with DNase I (10 mg/ml, USB). RNA wasquantified using RiboGreentper RNA quantitationreagent (Molecular Probes) and a fluorometer(TD700 Turner Designs) (Le Pecq and Paoletti,1966). Thirty micrograms of total RNA was elec-trophoresed for Northern blotting as describedpreviously, transferred to a Hybond™-N+charged nylon membrane (Amersham) and immo-bilized by UV cross-linking (Ausubel et al., 1987).A 32P-labeled hybridization probe, consisting of
cDNA spanning from nucleotide 1 to 468 of theBVDV NADL genome, was prepared by randompriming and extension with Klenow fragment andwas used for probing the Northern blot shown onFig. 6A. The hybridization probes for detectionof i-NADL/i-N-dIns, Thompson-CP/Thompson-NCP, 9a-CP/8b-NCP, JV4-CP/JV4-NCP pairsshown on Fig. 7A were prepared by RT-PCR (nt.90-386) of the 5%UTR fragment derived from i-NADL, Thompson-CP, 9a-CP and JV4-NCP, re-spectively. The RT-PCR products were sub-sequently used as templates in single-stranded-PCRreactions in the presence of [a-32P]dATP for am-plification of the negative strand. Prehybridizationwas performed 2 h at 42°C in a buffer containing50% formamide, 5× SSPE (stock 20× SSPE is 3M NaCl, 0.2 M NaH2PO4, and 20 mM EDTA, pH7.4), Denhardt’s solution (2 g/l of Ficoll 400, 2 g/lof polyvinylpyrrolidone, and 2 g/l of bovine serumalbumin), 2% SDS, denatured salmon sperm DNA(100 mg/ml). Hybridization was performedovernight at 42°C in fresh prehybridizaton solutionto which a probe labeled as described above (106
dpm/ml of hybridization solution) was added. Fil-ters were washed twice with 1× SSC–0.1% SDSat room temperature for 5 min and once with 0.1×SSC–0.1% SDS at 50°C for 10 min. Washed filterswere exposed in phosphoimager cassettes for 6 h at22–25°C. Detection and quantitation of radioac-tivity were performed with phosphoimager usingImageQuant software (Molecular Dynamics).
2.8. Nucleotide sequencing
Sequencing was performed as described (Hankeand Wink, 1994). The sequence of complete BVDVgenome cDNA in plasmid pNADLp15 and pN-dIns has been deposited in GenBank/EMBL datalibraries, under accession no. AJ33738 andAJ33739, respectively.
3. Results
3.1. Deri6ation of an isogenic non-cytopathicmutant of BVDV NADL
High copy number plasmids bearing large viralcDNA inserts are relatively unstable in E. coli as

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107 99
compared with plasmids of similar size withmedium-to-low copy replicons (Lerner and Inouye,1990; Cunningham et al., 1993). Thus, themedium-to-low copy number vector pACYC177was used as a vector for the cloning andpropagation of the full-length cDNA sequence ofBVDV strain NADL (Vassilev et al., 1997). We andothers (Mendez et al., 1998) have observed that theinstability problems of certain large high copynumber plasmids is often correlated with limitedexpression of the selectable marker enzymeb-lactamase. Thus, we designed a strategy toconstruct a low copy number vector, containingtwo independent b-lactamase genes flanking theorigin of replication (Fig. 1). The cloning strategyyielded two types of full-length BVDV cDNA lowcopy plasmids differing in the orientation of thetwo b-lactamase genes; ‘head to head’ or ‘head totail’. These constructs allowed rapid growth of theE. coli host at relatively high concentrations of
ampicilin, between 100 and 150 mg/ml (data notshown). The plasmid with the head-to-tailarrangement of b-lactamases was namedpNADLp15 (Fig. 1). We performed nucleotidesequence analysis of the BVDV cDNA cloned inpNADLp15 and the results were deposited into theGenBank database as described in Section 2. Thisplasmid was propagated in E. coli recA (JM109,SURE, DH5A); rec+ (MC1061) or dam−(GM2163) mutant strains without sequencedeletions and rearrangements after 2-l scaleovernight cultivation. Likewise, pNADLp15bacterial transformants had a uniform colonymorphology when plated in solid media (data notshown). Plasmid DNA from several differentbacterial strains transformed with pNADLp15 wasused to amplify the entire cDNA sequence ofBVDV NADL along with the adjacent T7promoter by PCR. The PCR products wereprecipitated with ethanol and transfected using
Fig. 1. Schematic presentation of the plasmid pNADLp15. Features such as the BVDV ORF and its encoded proteins, 5% and 3%UTRs, b-lactamase genes and origin of replication are shown with filled boxes. The ‘head to tail’ orientation of b-lactamase genesis depicted by arrows.

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107100
Fig. 2. Nucleotide sequence analysis of a segment of theBVDV genome encoding for NS2 (nucleotide 4981 to 5271)depicting deletion of the cIns genetic element. The pN-dInsNS2 region sequences shown flank the arrow which marks thesite (nt 4993-4) where the cIns element would be found in CPNADL.
microscopy. Except for the absence of cIns in theNS2 region, the polypeptides of the i-N-dIns virusare predicted to be identical to the parentalNADL. We passed the i-N-dIns virus four consec-utive times on BT cells and evaluated the stabilityof the deletion by RT-PCR after each passage.The size of all RT-PCR products indicated ab-sence of the cIns; therefore we concluded that theengineered deletion is stably maintained (data notshown). Nucleotide sequence analyses of the re-maining BVDV cDNA in pN-dIns revealed theexpected absence of the cIns. The nucleotide se-quence of the BVDV cDNA in pN-dIns has beendeposited in GenBank.
3.2. Virus reco6ery from the full-length cDNAclones of BVDV
EBTr cells transfected by electroporation with 1mg of RNA transcribed in vitro from thepNADLp15 template DNA yielded infectiousBVDV, termed i-NADL, within 48 h of cultiva-tion. Virus was also recovered from BT or EBTrcells lipofected with in vitro transcribed RNA.Similar results were obtained with pN-dIns, ex-cept that no CPE was observed in these monolay-ers. To assess the identity of the BVDV recoveredfrom in vitro-synthesized transcripts, total RNAisolated from virus infected cells was amplified byRT-PCR. The nucleotide sequence of ampliconsfrom the NS2-3 region revealed that the onlydifference between i-NADL and i-N-dIns RNA’swas the absence of the 270 nt encoding for cIns ini-N-dIns (Fig. 2 and data not shown). Plaqueproduction by i-NADL and focus formation byi-N-dIns viruses was visualized by crystal violetstaining or immunoperoxidase assay with a chro-mogenic substrate, respectively. Both viruses gen-erated uniform plaques/foci morphology. Threedays after infection, plaque diameter of i-NADLwas �3.5 mm (Fig. 3A) and the size of the focigenerated from pN-dIns was �4 mm (Fig. 3B).This size difference is only apparent, as im-munoperoxidase staining of i-NADL-inducesplaques that have identical diameter to those ofi-N-dIns, due to staining of cells, which have notyet died. Interestingly, antigen concentrationwithin the foci produced by i-N-dIns was not
cationic lipid complexes into MVA-T7 vacciniavirus-infected BT cells (Vassilev and Donis, 1996).Supernatants from transfected cells were har-vested at 24 h post-transfection, filtered through0.1-mm filter to eliminate the vaccinia virus helper,and used to infect fresh BT monolayers. Infec-tious virus was rescued from all transfections,indicating that all bacterial strains tested propa-gated the pNADLp15 without deleterious rear-rangements or deletions (data not shown).
A previous report indicated that deletion of thecIns sequence from the NS2 coding region ofNADL resulted in a BVDV that failed to processthe NS3 protein, and did not cause apoptosis ininfected cells (Mendez et al., 1998). To explore thehypothesis that the non-cytopathic phenotype ofBVDV NADL can be correlated with decreasedlevels of macromolecular synthesis (RNA andprotein), we constructed a vector (pN-dIns) inwhich the coding sequence of cIns was excisedfrom the wt genome (Fig. 2). The resulting virus,termed i-N-dIns, completed its replication cycle inbovine cells without causing cell death or anyother morphological changes observable by light
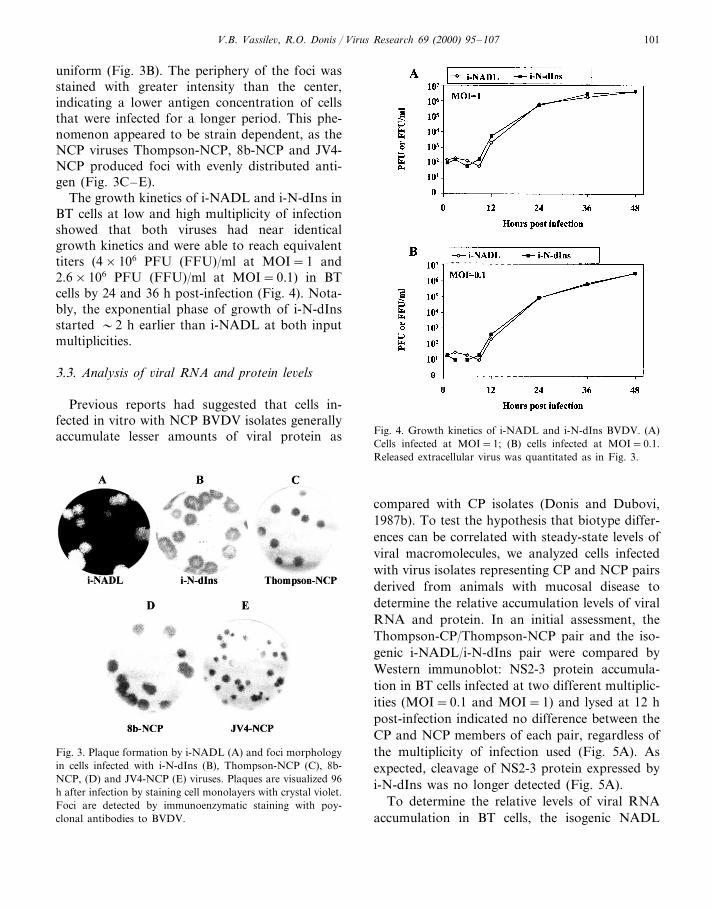
V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107 101
uniform (Fig. 3B). The periphery of the foci wasstained with greater intensity than the center,indicating a lower antigen concentration of cellsthat were infected for a longer period. This phe-nomenon appeared to be strain dependent, as theNCP viruses Thompson-NCP, 8b-NCP and JV4-NCP produced foci with evenly distributed anti-gen (Fig. 3C–E).
The growth kinetics of i-NADL and i-N-dIns inBT cells at low and high multiplicity of infectionshowed that both viruses had near identicalgrowth kinetics and were able to reach equivalenttiters (4×106 PFU (FFU)/ml at MOI=1 and2.6×106 PFU (FFU)/ml at MOI=0.1) in BTcells by 24 and 36 h post-infection (Fig. 4). Nota-bly, the exponential phase of growth of i-N-dInsstarted �2 h earlier than i-NADL at both inputmultiplicities.
3.3. Analysis of 6iral RNA and protein le6els
Previous reports had suggested that cells in-fected in vitro with NCP BVDV isolates generallyaccumulate lesser amounts of viral protein as Fig. 4. Growth kinetics of i-NADL and i-N-dIns BVDV. (A)
Cells infected at MOI=1; (B) cells infected at MOI=0.1.Released extracellular virus was quantitated as in Fig. 3.
Fig. 3. Plaque formation by i-NADL (A) and foci morphologyin cells infected with i-N-dIns (B), Thompson-NCP (C), 8b-NCP, (D) and JV4-NCP (E) viruses. Plaques are visualized 96h after infection by staining cell monolayers with crystal violet.Foci are detected by immunoenzymatic staining with poy-clonal antibodies to BVDV.
compared with CP isolates (Donis and Dubovi,1987b). To test the hypothesis that biotype differ-ences can be correlated with steady-state levels ofviral macromolecules, we analyzed cells infectedwith virus isolates representing CP and NCP pairsderived from animals with mucosal disease todetermine the relative accumulation levels of viralRNA and protein. In an initial assessment, theThompson-CP/Thompson-NCP pair and the iso-genic i-NADL/i-N-dIns pair were compared byWestern immunoblot: NS2-3 protein accumula-tion in BT cells infected at two different multiplic-ities (MOI=0.1 and MOI=1) and lysed at 12 hpost-infection indicated no difference between theCP and NCP members of each pair, regardless ofthe multiplicity of infection used (Fig. 5A). Asexpected, cleavage of NS2-3 protein expressed byi-N-dIns was no longer detected (Fig. 5A).
To determine the relative levels of viral RNAaccumulation in BT cells, the isogenic NADL

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107102
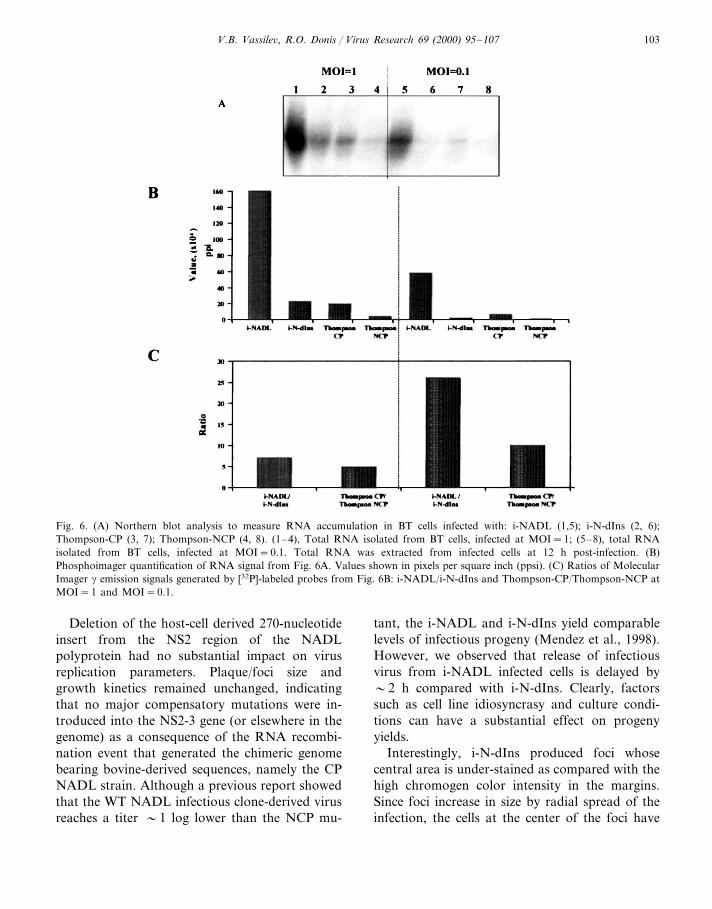
CP/NCP pair and Thompson-CP/NCP pair wereused to infect monolayers at MOI=0.1 andMOI=1. Total RNA isolated at 12 h post-infec-tion was analyzed by Northern blot hybridization(Fig. 6A) with a DNA probe derived from the 5%466 nucleotides the BVDV NADL genome. Quan-tification of the bound probe by phosphoimagerrevealed that viral RNA accumulation in i-NADLinfected cells at input multiplicity (MOI) of 1 was7 times higher than that in i-N-dIns virus infectedcells. Likewise, cells infected at this multiplicitywith the Thompson-CP BVDV accumulated fivetimes more viral RNA than the NCP BVDVcounterpart. This differential accumulation of vi-ral RNA was more pronounced when cells wereinfected at MOI=0.1. At this multiplicity, i-NADL-infected cells accumulated 26 times moreRNA than the i-N-dIns isogenic virus. At thelower MOI Thompson BVDV-infected cellsshowed a 10-fold difference between CP andNCP. These results suggest that expression ofNS3 alone correlates with dramatic increase ofRNA synthesis levels, which appear to be in-versely correlated to the MOI.
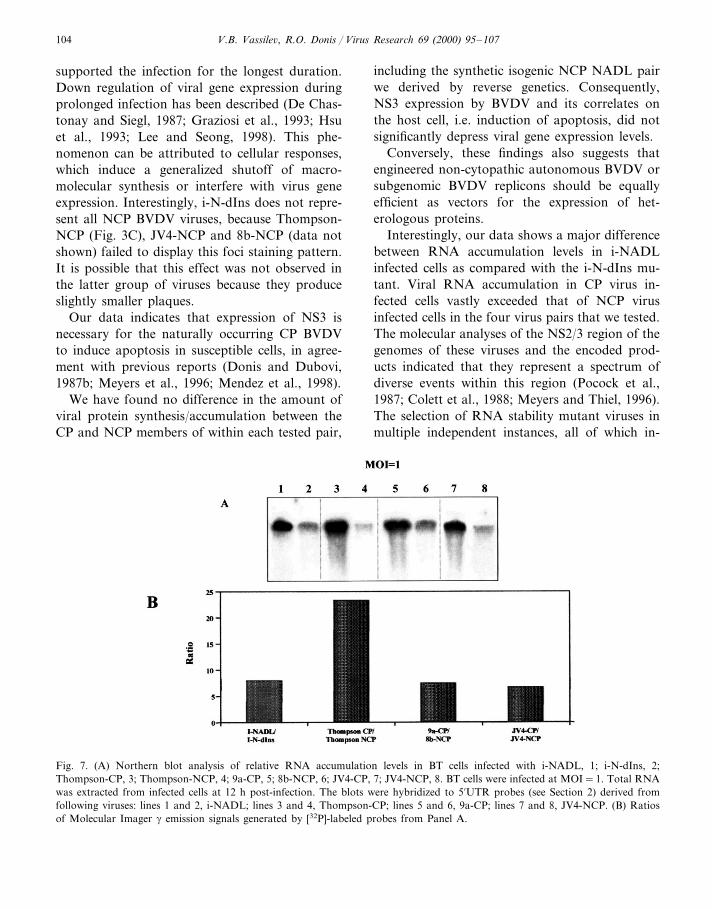
We extended our analyses of RNA and proteinaccumulation levels in cells to two additionalBVDV CP/NCP pairs — 9a-CP/8b-NCP, andJV4-CP/JV4-NCP (Corapi et al., 1988). Westernblot analysis (Fig. 5B) revealed that the previouslyobserved pattern was maintained; protein accu-mulation levels were virtually the same in cellsinfected with the CP or NCP member of theBVDV isolate pairs. For greater accuracy inquantification of RNA accumulation levels in-duced by each BVDV CP/NCP isolate pair tested,we prepared homologous probes specific for eachpair. Total RNA from infected BT cells was sepa-rated by electrophoresis and transferred to mem-branes to be hybridized to the homologousradiolabeled probes (see Section 2). The signalfrom the Northern blot hybridization (Fig. 7A)was quantified by phosphoimager and the ratiosfor each pair were plotted (Fig. 7B). The resultsconfirmed the great differences in RNA levelsobserved in the previous experiments. To normal-ize this additional CP/NCP data, the NADL pairwas included with the new pairs. The RNA accu-mulation ratio (at MOI=1) was 8 for NADL-CP/NCP, whereas for the 9b-CP/8a-NCP pair aratio of 7.5 was observed. Finally, the ratio RNAaccumulation in JV4-CP/JV4-NCP infected cells,was 6.5. Interestingly, cells infected with theThompson-CP/NCP pair yielded RNA accumula-tion ratios as high as 23, consistent with the use ofhomologous probes in this experiment.
4. Discussion
As noted previously, a low copy number episo-mal origin of DNA replication improves the sta-bility of plasmids bearing full-length BVDVgenome cDNA in E. coli (Moormann et al., 1996;Ruggli et al., 1996; Mendez et al., 1998). Bycombining a duplicated b-lactamase gene togetherwith the low copy number origin of replication,we obtained highly stable infectious clones ofBVDV in various hosts. While this result suggeststhat the b-lactamase gene duplication is beneficial,further analysis is required to understand thepotentially stabilizing mechanism.
Fig. 5. (A) Western blot analysis of NS3 and NS2-3 expres-sion. i-NADL, (1, 5); i-N-dIns, (2, 6); Thompson-CP, (3, 7);Thompson-NCP, (4, 8); cells were infected at MOI=0.1, (1-4)and at MOI=1, (5-8). (B) Western blot analysis of NS3 andNS2-3 expression. i-NADL, 1; i-N-dIns, 2; Thompson-CP, 3;Thompson-NCP, 4; 9a-CP, 5; 8b-NCP, 6; JV4-CP, 7; JV4-NCP, 8; Cells were infected at MOI=1. All cell lysates wereprepared 12 h post-infection. Electrophoresed and blottedproteins were detected using monoclonal antibody 20.10.6.,specific for the pestivirus non-structural protein p80 (NS3).
V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107 103
Fig. 6. (A) Northern blot analysis to measure RNA accumulation in BT cells infected with: i-NADL (1,5); i-N-dIns (2, 6);Thompson-CP (3, 7); Thompson-NCP (4, 8). (1–4), Total RNA isolated from BT cells, infected at MOI=1; (5–8), total RNAisolated from BT cells, infected at MOI=0.1. Total RNA was extracted from infected cells at 12 h post-infection. (B)Phosphoimager quantification of RNA signal from Fig. 6A. Values shown in pixels per square inch (ppsi). (C) Ratios of MolecularImager g emission signals generated by [32P]-labeled probes from Fig. 6B: i-NADL/i-N-dIns and Thompson-CP/Thompson-NCP atMOI=1 and MOI=0.1.
Deletion of the host-cell derived 270-nucleotideinsert from the NS2 region of the NADLpolyprotein had no substantial impact on virusreplication parameters. Plaque/foci size andgrowth kinetics remained unchanged, indicatingthat no major compensatory mutations were in-troduced into the NS2-3 gene (or elsewhere in thegenome) as a consequence of the RNA recombi-nation event that generated the chimeric genomebearing bovine-derived sequences, namely the CPNADL strain. Although a previous report showedthat the WT NADL infectious clone-derived virusreaches a titer �1 log lower than the NCP mu-
tant, the i-NADL and i-N-dIns yield comparablelevels of infectious progeny (Mendez et al., 1998).However, we observed that release of infectiousvirus from i-NADL infected cells is delayed by�2 h compared with i-N-dIns. Clearly, factorssuch as cell line idiosyncrasy and culture condi-tions can have a substantial effect on progenyyields.
Interestingly, i-N-dIns produced foci whosecentral area is under-stained as compared with thehigh chromogen color intensity in the margins.Since foci increase in size by radial spread of theinfection, the cells at the center of the foci have
V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107104
supported the infection for the longest duration.Down regulation of viral gene expression duringprolonged infection has been described (De Chas-tonay and Siegl, 1987; Graziosi et al., 1993; Hsuet al., 1993; Lee and Seong, 1998). This phe-nomenon can be attributed to cellular responses,which induce a generalized shutoff of macro-molecular synthesis or interfere with virus geneexpression. Interestingly, i-N-dIns does not repre-sent all NCP BVDV viruses, because Thompson-NCP (Fig. 3C), JV4-NCP and 8b-NCP (data notshown) failed to display this foci staining pattern.It is possible that this effect was not observed inthe latter group of viruses because they produceslightly smaller plaques.
Our data indicates that expression of NS3 isnecessary for the naturally occurring CP BVDVto induce apoptosis in susceptible cells, in agree-ment with previous reports (Donis and Dubovi,1987b; Meyers et al., 1996; Mendez et al., 1998).
We have found no difference in the amount ofviral protein synthesis/accumulation between theCP and NCP members of within each tested pair,
including the synthetic isogenic NCP NADL pairwe derived by reverse genetics. Consequently,NS3 expression by BVDV and its correlates onthe host cell, i.e. induction of apoptosis, did notsignificantly depress viral gene expression levels.
Conversely, these findings also suggests thatengineered non-cytopathic autonomous BVDV orsubgenomic BVDV replicons should be equallyefficient as vectors for the expression of het-erologous proteins.
Interestingly, our data shows a major differencebetween RNA accumulation levels in i-NADLinfected cells as compared with the i-N-dIns mu-tant. Viral RNA accumulation in CP virus in-fected cells vastly exceeded that of NCP virusinfected cells in the four virus pairs that we tested.The molecular analyses of the NS2/3 region of thegenomes of these viruses and the encoded prod-ucts indicated that they represent a spectrum ofdiverse events within this region (Pocock et al.,1987; Colett et al., 1988; Meyers and Thiel, 1996).The selection of RNA stability mutant viruses inmultiple independent instances, all of which in-
Fig. 7. (A) Northern blot analysis of relative RNA accumulation levels in BT cells infected with i-NADL, 1; i-N-dIns, 2;Thompson-CP, 3; Thompson-NCP, 4; 9a-CP, 5; 8b-NCP, 6; JV4-CP, 7; JV4-NCP, 8. BT cells were infected at MOI=1. Total RNAwas extracted from infected cells at 12 h post-infection. The blots were hybridized to 5%UTR probes (see Section 2) derived fromfollowing viruses: lines 1 and 2, i-NADL; lines 3 and 4, Thompson-CP; lines 5 and 6, 9a-CP; lines 7 and 8, JV4-NCP. (B) Ratiosof Molecular Imager g emission signals generated by [32P]-labeled probes from Panel A.

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107 105
volve the center of the viral genome, would beextremely unlikely. Thus, we postulate that accu-mulation differences are more likely due tobiosynthetic rate rather than degradation differ-ences. Hoff et al. as well as others demonstratedthat CP BVDV induces cell death by triggeringapoptosis (Zhang et al., 1996; Hoff and Donis,1997). This finding was supported by observationof typical morphological and biochemical changesof apoptosis; condensation of chromatin, appear-ance of apoptotic bodies and cleavage of poly(ADP-ribose) polymerase (PARP) (Hoff andDonis, 1997). Thus, increased viral RNA concen-tration within infected cells may trigger eventsleading to irreversible execution of the apoptoticcascade, including caspase activation, cleavage ofprotein targets and DNA fragmentation. It istempting to speculate that increased concentrationof viral RNA activates PKR (p68 kinase), whichis a known inducer of apoptosis. Mechanistically,PKR can induce apoptosis by activation of NF-kB, signaling via the Fas pathway or inactivationof eIF2a (Balachandran et al., 1998; Srivastava etal., 1998). dsRNA could also activate the 2%–5%oligoadenylate synthetase (Castelli et al., 1998),and the latter species lead to activation of RNaseL (Player and Torrence, 1998), both of which arepro-apoptotic (Diaz-Guerra et al., 1997). Indeed,dsRNA or its downstream effectors have beenproposed to mediate apoptosis in influenza andvaccinia virus infection (Takizawa et al., 1996;Kibler et al., 1997).
Although higher concentration of viral RNA inbovine cells infected with CP strains of BVDVmay contribute induction of cell death, it may notbe the only factor involved. Certainly, the loss ofNS2 from the N-terminus of NS3 can potentiallyalter the intracellular trafficking of this essentialBVDV protein. The presence of an enzymaticallyactive NS3 protease and helicase in certain intra-cellular compartments may by itself triggerapoptosis.
CP BVDV strains have been exclusively isolatedfrom cases of mucosal disease, virtually all fatal(Donis, 1995; Donis and Dubovi, 1987b). Regard-less of the genetic or metabolic path leading to itsexpression, NS3 is invariably produced by CPBVDV isolates, which elicit apoptosis in cultured
bovine cells. Thus, three elements correlate ex-tremely well (I) fatal BVDV disease in a persis-tently infected cow; (ii) expression of NS3, and(iii) apoptosis in cultured cells. This work addsviral RNA accumulation as a new parameter,which correlates with these three phenotypic char-acters of CP BVDV. Further studies are needed todetermine which of these elements (if any) areessential for induction of fatal mucosal diseaseand the relations between them. Manipulation ofthe viral genome by reverse genetics will facilitatethe construction of virus mutants in which thesecharacters are dissociated to evaluate their role inpathogenesis individually. Such studies may eluci-date the role, if any, of heightened CP BVDVRNA accumulation levels in mucosal disease.
Acknowledgements
This work was supported by NRI grant No.97-35204-5068 and 92-37204-7959 from theUSDA. We thank Dr Fernando Osorio forBVDV isolates and Dr Dubovi for providing uswith virus isolates and monoclonal antibodies.Our thanks to all the members of the laboratoryfor helpful discussions.
References
Ali, N., Siddiqui, A., 1997. The La antigen binds 5 noncodingregion of the hepatitis C virus RNA in the context of theinitiator AUG codon and stimulates internal ribosomeentry site-mediated translation. Proc. Natl. Acad. Sci. USA94 (6), 2249–2254.
Ausubel, F., Brent, R., Kingston, R., Moore, J., Siedman, J.,Smith, J., Struhl, K., 1987. Current Protocols in MolecularBiology. Wiley, New York.
Balachandran, S., Kim, C.N., Yeh, W.C., Mak, T.W., Bhalla,K., Barber, G.N., 1998. Activation of the dsRNA-depen-dent protein kinase, PKR, induces apoptosis throughFADD-mediated death signaling. EMBO J. 17 (23), 6888–6902.
Barnes, W.M., 1994. PCR amplification of up to 35-kb DNAwith high fidelity and high yield from lambda bacterio-phage templates. Proc. Natl. Acad. Sci. USA 91 (6), 2216–2220.
Castelli, J., Wood, K.A., Youle, R.J., 1998. The 2-5A systemin viral infection and apoptosis. Biomed. Pharmacother. 52(9), 386–390.

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107106
Cheng, S., Fockler, C., Barnes, W.M., Higuchi, R., 1994.Effective amplification of long targets from cloned insertsand human genomic DNA. Proc. Natl. Acad. Sci. USA 91(12), 5695–5699.
Chomczynski, P., Sacchi, N., 1987. Single-step method ofRNA isolation by acid guanidinium thiocyanate–phenol–chloroform extraction. Anal. Biochem. 162 (1), 156–159.
Chon, S.K., Perez, D.R., Donis, R.O., 1998. Genetic analysisof the internal ribosome entry segment of bovine viraldiarrhea virus. Virology 251 (2), 370–382.
Colett, M.S., Larson, R., Gold, C., Strick, D., Anderson,D.K., Purchio, A.F., 1988. Molecular cloning and nucle-otide sequence of the pestivirus bovine viral diarrhea virus.Virology 165 (1), 191–199.
Corapi, W.V., Donis, R.O., Dubovi, E.J., 1988. Monoclonalantibody analyses of cytopathic and noncytopathic virusesfrom fatal bovine viral diarrhea virus infections. J. Virol.62 (8), 2823–2827.
Corapi, W.V., Donis, R.O., Dubovi, E.J., 1990. Characteriza-tion of a panel of monoclonal antibodies and their use inthe study of the antigenic diversity of bovine viral diarrheavirus. Am. J. Vet. Res. 51 (9), 1388–1394.
Cunningham, T.P., Montelaro, R.C., Rushlow, K.E., 1993.Lentivirus envelope sequences and proviral genomes arestabilized in Escherichia coli when cloned in low-copy-number plasmid vectors. Gene 124 (1), 93–98.
De Chastonay, J., Siegl, G., 1987. Replicative events in hepati-tis A virus-infected MRC-5 cells. Virology 157 (2), 268–275.
Diaz-Guerra, M., Rivas, C., Esteban, M., 1997. Activation ofthe IFN-inducible enzyme RNase L causes apoptosis ofanimal cells. Virology 236 (2), 354–363.
Donis, R.O., 1995. Molecular biology of bovine viral diarrheavirus and its interactions with the host. Vet. Clin. NorthAm. Food Anim. Pract. 11 (3), 393–423.
Donis, R.O., Dubovi, E.J., 1987a. Glycoproteins of viral di-arrhoea-mucosal disease virus in infected bovine cells. J.Gen. Virol. 68 (Pt 6), 1607–1616.
Donis, R.O., Dubovi, E.J., 1987b. Differences in virus-inducedpolypeptides in cells infected by cytopathic and noncyto-pathic biotypes of bovine virus diarrhea-mucosal diseasevirus. Virology 158 (1), 168–173.
Elbers, K., Tautz, N., Becher, P., Stoll, D., Rumenapf, T.,Thiel, H.J., 1996. Processing in the pestivirus E2-NS2region: identification of proteins p7 and E2p7. J. Virol. 70(6), 4131–4135.
Flores, E.F., Kreutz, L.C., Donis, R.O., 1996. Swine andruminant pestiviruses require the same cellular factor toenter bovine cells. J. Gen. Virol. 77 (Pt 6), 1295–1303.
Frolov, I., McBride, M.S., Rice, C.M., 1998. cis-acting RNAelements required for replication of bovine viral diarrheavirus-hepatitis C virus 5% nontranslated region chimeras.Rna 4 (11), 1418–1435.
Fukushi, S., Kurihara, C., Ishiyama, N., Hoshino, F.B., Oya,A., Katayama, K., 1997. The sequence element of theinternal ribosome entry site and a 25-kilodalton cellularprotein contribute to efficient internal initiation of transla-tion of hepatitis C virus RNA. J. Virol. 71 (2), 1662–1666.
Graziosi, C., Pantaleo, G., Butini, L., Demarest, J.F., Saag,M.S., Shaw, G.M., Fauci, A.S., 1993. Kinetics of humanimmunodeficiency virus type 1 (HIV-1) DNA and RNAsynthesis during primary HIV-1 infection. Proc. Natl.Acad. Sci. USA 90 (14), 6405–6409.
Hanke, M., Wink, M., 1994. Direct DNA sequencing ofPCR-amplified vector inserts following enzymatic degrada-tion of primer and dNTPs. Biotechniques 17 (5), 858–860.
Hoff, H.S., Donis, R.O., 1997. Induction of apoptosis andcleavage of poly(ADP-ribose) polymerase by cytopathicbovine viral diarrhea virus infection. Virus Res. 49 (1),101–113.
Hsu, M.C., Dhingra, U., Earley, J.V., Holly, M., Keith, D.,Nalin, C.M., Richou, A.R., Schutt, A.D., Tam, S.Y.,Potash, M.J., et al., 1993. Inhibition of type 1 humanimmunodeficiency virus replication by a tat antagonist towhich the virus remains sensitive after prolonged exposurein vitro. Proc. Natl. Acad. Sci. USA 90 (14), 6395–6399.
Kibler, K.V., Shors, T., Perkins, K.B., Zeman, C.C., Ba-naszak, M.P., Biesterfeldt, J., Langland, J.O., Jacobs, B.L.,1997. Double-stranded RNA is a trigger for apoptosis invaccinia virus- infected cells. J. Virol. 71 (3), 1992–2003.
Le Pecq, J.B., Paoletti, C., 1966. A new fluorometric methodfor RNA and DNA determination. Anal. Biochem. 17 (1),100–107.
Lee, K.H., Seong, B.L., 1998. The position 4 nucleotide at the3% end of the influenza virus neuraminidase vRNA isinvolved in temporal regulation of transcription and repli-cation of neuraminidase RNAs and affects the repertoireof influenza virus surface antigens. J. Gen. Virol. 79 (Pt 8),1923–1934.
Lerner, C.G., Inouye, M., 1990. Low copy number plasmidsfor regulated low-level expression of cloned genes in Es-cherichia coli with blue/white insert screening capability.Nucleic Acids Res. 18 (15), 4631.
Lin, C., Lindenbach, B.D., Pragai, B.M., McCourt, D.W.,Rice, C.M., 1994a. Processing in the hepatitis C virusE2-NS2 region: identification of p7 and two distinct E2-specific products with different C termini. J. Virol. 68 (8),5063–5073.
Lin, Y.J., Liao, C.L., Lai, M.M., 1994b. Identification of thecis-acting signal for minus-strand RNA synthesis of amurine coronavirus: implications for the role of minus-strand RNA in RNA replication and transcription. J.Virol. 68 (12), 8131–8140.
Mendez, E., Ruggli, N., Collett, M.S., Rice, C.M., 1998.Infectious bovine viral diarrhea virus (strain NADL) RNAfrom stable cDNA clones: a cellular insert determines NS3production and viral cytopathogenicity. J. Virol. 72 (6),4737–4745.
Meyers, G., Thiel, H.J., 1996. Molecular characterization ofpestiviruses. Adv. Virus Res. 47, 53–118.
Meyers, G., Tautz, N., Becher, P., Thiel, H.J., Kummerer,B.M., 1996. Recovery of cytopathogenic and noncyto-pathogenic bovine viral diarrhea viruses from cDNA con-structs. J. Virol. 70 (12), 8606–8613.

V.B. Vassile6, R.O. Donis / Virus Research 69 (2000) 95–107 107
Moormann, R.J., van Gennip, H.G., Miedema, G.K., Hulst,M.M., van Rijn, P.A., 1996. Infectious RNA transcribedfrom an engineered full-length cDNA template of thegenome of a pestivirus. J. Virol. 70 (2), 763–770.
Player, M.R., Torrence, P.F., 1998. The 2-5A system: modula-tion of viral and cellular processes through acceleration ofRNA degradation. Pharmacol. Ther. 78 (2), 55–113.
Pocock, D.H., Howard, C.J., Clarke, M.C., Brownlie, J., 1987.Variation in the intracellular polypeptide profiles fromdifferent isolates of bovine virus diarrhoea virus. Arch.Virol. 94 (1-2), 43–53.
Poole, T.L., Wang, C., Popp, R.A., Potgieter, L.N., Siddiqui,A., Collett, M.S., 1995. Pestivirus translation initiationoccurs by internal ribosome entry. Virology 206 (1), 750–754.
Rijnbrand, R., van der Straaten, T., van Rijn, P.A., Spaan, W.J.,Bredenbeek, P.J., 1997. Internal entry of ribosomes isdirected by the 5% noncoding region of classical swine fevervirus and is dependent on the presence of an RNA pseudo-knot upstream of the initiation codon. J. Virol. 71 (1),451–457.
Rose, R.E., 1988. The nucleotide sequence of pACYC177.Nucleic Acids Res. 16 (1), 356.
Ruggli, N., Tratschin, J.D., Mittelholzer, C., Hofmann, M.A.,1996. Nucleotide sequence of classical swine fever virusstrain Alfort/187 and transcription of infectious RNA fromstably cloned full-length cDNA. J. Virol. 70 (6), 3478–3487.
Srivastava, S.P., Kumar, K.U., Kaufman, R.J., 1998. Phospho-rylation of eukaryotic translation initiation factor 2 mediatesapoptosis in response to activation of the double-strandedRNA-dependent protein kinase. J. Biol. Chem. 273 (4),2416–2423.
Takizawa, T., Ohashi, K., Nakanishi, Y., 1996. Possible in-volvement of double-stranded RNA-activated protein ki-nase in cell death by influenza virus infection. J. Virol.70 (11), 8128–8132.
Tautz, N., Meyers, G., Stark, R., Dubovi, E.J., Thiel, H.J.,1996. Cytopathogenicity of a pestivirus correlates with a27-nucleotide insertion. J. Virol. 70 (11), 7851–7858.
Vassilev, V.B., Donis, R.O. In vivo rescue of infectiousBVDV by transfection of plasmid DNA into cells in-fected with vaccinia virus expressing T7 RNA poly-merase, Proceedings of the third ESVV Symposium onPestivirus Infections Lelystad, The Netherlands, 19–20September, 1996, pp. 1–7.
Vassilev, V.B., Collett, M.S., Donis, R.O., 1997. Authenticand chimeric full-length genomic cDNA clones of bovineviral diarrhea virus that yield infectious transcripts. J.Virol. 71 (1), 471–478.
Wyatt, L.S., Moss, B., Rozenblatt, S., 1995. Replication-deficient vaccinia virus encoding bacteriophage T7 RNApolymerase for transient gene expression in mammaliancells. Virology 210 (1), 202–205.
Xu, J., Mendez, E., Caron, P.R., Lin, C., Murcko, M.A.,Collett, M.S., Rice, C.M., 1997. Bovine viral diarrheavirus NS3 serine proteinase: polyprotein cleavage sites,cofactor requirements, and molecular model of an en-zyme essential for pestivirus replication. J. Virol. 71 (7),5312–5322.
Zhang, G., Aldridge, S., Clarke, M.C., McCauley, J.W.,1996. Cell death induced by cytopathic bovine viral di-arrhoea virus is mediated by apoptosis. J. Gen. Virol. 77(Pt 8), 1677–1681.
.