blood circulation as source for osteopontin in acellular extrinsic fiber cementum and other...
TRANSCRIPT

http://jdr.sagepub.com/Journal of Dental Research
http://jdr.sagepub.com/content/78/11/1688The online version of this article can be found at:
DOI: 10.1177/00220345990780110501
1999 78: 1688J DENT REST. VandenBos, A.L.J.J. Bronckers, H.A. Goldberg and W. Beertsen
Mineralizing TissuesBlood Circulation as Source for Osteopontin in Acellular Extrinsic Fiber Cementum and Other
Published by:
http://www.sagepublications.com
On behalf of:
International and American Associations for Dental Research
can be found at:Journal of Dental ResearchAdditional services and information for
http://jdr.sagepub.com/cgi/alertsEmail Alerts:
http://jdr.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://jdr.sagepub.com/content/78/11/1688.refs.htmlCitations:
at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

J Dent Res 78(11): 1688-1695, November, 1999
Blood Circulation as Source for Osteopontinin Acellular Extrinsic Fiber Cementumand Other Mineralizing TissuesT. VandenBos1, A.L.J.J. Bronckers2, H.A. Goldberg3, and W. Beertsen'4
'Department of Periodontology, Academic Center for Dentistry Amsterdam (ACTA), University of Amsterdam, Louwesweg 1, 1066 EAAmsterdam, The Netherlands; 2Department of Oral Cell Biology, Academic Center for Dentistry Amsterdam (ACTA), Free University; 3Divisionof Oral Biology, School of Dentistry, University of Westem Ontario, London, Ontario, Canada; 4corresponding author
Abstract. Osteopontin (OPN) is one of the major non-collage-nous proteins in root cementum and other mineralized tis-sues. Although most of this mineral-seeking protein isthought to be produced by local tissue cells, some of it mightenter the mineralizing matrix from the blood. To test thishypothesis, we followed the distribution of a single dose ofpurified porcine or rat 1251-labeled OPN injected i.v. in rats, inmineralizing and non-mineralizing tissues and in subcuta-neously implanted collagenous implants. The animals werekilled 30 or 48 hrs after injection. Tissues (calvaria, tibia, lowerand upper jaws) were harvested and processed forradioautography and biochemical analysis. Tissues as well ascalcifying collagenous implants proved to have taken upradiolabel. In EDTA extracts of long bones, the majority of theradiolabel was demonstrated to be associated with intactOPN. The iodinated protein was also found in the acellularextrinsic fiber cementum (acellular cementum) layer investingthe continuously growing incisors, in laminae limitantes,cement lines, and in forming bone near the mineralizationfront. Further, the label was present in the circumpulpaldentin of the incisors, and some of it appeared to have beenincorporated into developing enamel. It is concluded thatOPN in acellular cementum and other mineralizing tissuesmay-at least partially-originate from sources outside thedirect environment following its transportation via serum.
Key words: osteopontin, collagen, mineralization, rootcementum, bone, dentin.
Received January 12, 1998; Last Revision February 22, 1999;Accepted February 23, 1999
Introduction
In mammals, all hard tissues contain a mineral phase in theform of carbonated hydroxyapatite crystallites. With theexception of enamel and also acellular afibrillar cementum,the matrix of these tissues contains both collagen and a vari-ety of non-collagenous substances, which include growth fac-tors and phosphorylated proteins (for reviews, see Fisher andTermine, 1985; Linde, 1989; Roach, 1994; Butler and Ritchie,1995). Least studied of all mineralized tissues is the cemen-tum layer covering the roots of our teeth. This tissue helps toprovide anchorage of the teeth to the socket wall viaSharpey's fibers. Several types of cementum can be distin-guished, including (1) acellular extrinsic fiber cementum(which is usually found in the occlusal half of the root) and(2) cellular cementum (which invests the more apical portionof the root surface) (Schroeder, 1986). The cementum containsa host of non-collagenous proteins, such as fibronectin(Connor et al., 1984), tenascin (Zhang et al., 1993), osteonectin(Reichert et al., 1992), osteopontin (OPN; Bronckers et al., 1994;McKee et al., 1996), bone sialoprotein ll (MacNeil et al., 1994),osteocalcin (Bronckers et al., 1994), proteoglycans (Bartold etal., 1990), growth factors (Nakae et al., 1991), cementumattachment protein (specific for cementum; Arzate et al.,1992), and or2HS-glycoprotein (McKee et al., 1996).
In bone, dentin, and cellular cementum, the mineralizedmatrix-associated proteins (with the exception of albuminand a2HS-glycoprotein, which appear to arise from the circu-lation) are synthesized and deposited by osteoblasts, odonto-blasts, and cementoblasts, respectively. However, the sourcefor these non-collagenous proteins in acellular cementum isnot presently known. With respect to the phosphorylatedsialoprotein OPN, for example, it was noted that this mole-cule is found in relatively high concentrations in acellularcementum (Bronckers et al., 1994; McKee et al., 1996). Whileperiodontal ligament fibroblasts are able to synthesize OPNin culture (Ramakrishnan et al., 1995; Nohutcu et al., 1996), theprotein is already detected in the cementum layer before it isexpressed in the adjacent periodontal ligament cells
1688 at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

Blood Circulation as Sourcefor Osteopontin
(Bronckers et al., 1994; Lekic et al., 1996). At later stages ofdevelopment and also during wound healing, periodontalligament fibroblasts stain slightly positive for OPN (Chen etal., 1993; Bronckers et al., 1994; Takano-Yamamoto et al., 1994;Lekic et al., 1996). The observations of Bronckers et al. (1994)suggested to us that some of the OPN in acellular extrinsicfiber cementum is derived from a source outside the directenvironment. The present study was undertaken to deter-mine whether OPN can enter acellular cementum and othercalcifying tissues directly from the circulatory system.
Materials and methods
Binding of alkaline phosphatase to collagen slices
Collagen sheets from demineralized bovine bone and dentin werecomplexed with intestinal alkaline phosphatase of bovine origin(Boehringer Mannheim, Germany) as described previously(VandenBos and Beertsen, 1994). They were stored in 0.1 mol/Lglycine buffer, pH 10.5, at 4°C.
Radio-iodination of OPNOPN (0.5 mg; 98 to 99% pure), isolated from porcine bone asdescribed by Goldberg and Sodek (1994), was radiolabeled with 2mCi 1251 after oxidation with Chloramin T (Amersham protocol;Guide to Radioiodination Techniques). We removed non-bound labelby de-salting the protein on a PD-10 column (Sephadex G-25;Pharmacia, Uppsala, Sweden). The specific radioactivity of theprotein was approximately 815,000 cpm/pu.g; 95% of the label wasprecipitable with 5% TCA. Similarly, 0.5 mg OPN purified fromrat bone (99% pure) was radiolabeled. The specific radioactivity ofthis rat OPN was 900,000 cpm/>ig protein, whereas 85% of thelabel was precipitable with 5% TCA.
Purity of OPN was assessed by amino acid analysis, and bythe apparent homogeneity of stained protein bands on polyacry-lamide gels, before and after thrombin digestion, according to thesensitive Stains-all/silver staining protocol (see Fig. 3A), whichcan detect protein in the subnanogram range (Goldberg andWarner, 1997). No other protein bands were evident on the SDS-PAGE at the concentrations of protein that was loaded.
In vitro mineralizationDemineralized bone slices complexed with alkaline phosphatase(containing 1.6 ± 0.3 mU alkaline phosphatase/p.g hydroxypro-line; n = 4) and slices without the enzyme were incubated in 1 mLIscove's Modified Dulbecco's Medium containing 10 mmol/L P-glycerophosphate and 125I-OPN (13,000 cpm/mL). After 1, 3, 7,and 14 days, collagen slices were harvested and washed in water,after which the amount of bound 1251 was determined in aPackard y-counter. Also, the amount of phosphate taken up bythe collagen slices was assessed and expressed per hydroxyprolinecontent. Statistical analysis was carried out by linear regressionanalysis techniques.
Immunolocalization of OPNThe protocol for the use of animals was approved by theCommittee of Animal Experiments of the Academic Medical
Center (Amsterdam). Ten-week-old male Wistar rats were anes-thetized with Hypnorm® (Janssen Pharmaceutical Ltd, Oxford,UK), and an incision was made through the skin overlying theskull and in the right and left sides of the abdominal wall.Demineralized dentin and bone slices were then implanted intoskin pockets in 20 rats. Each animal received 8 implants, 4 overthe skull (supraperiosteally) and 4 in the dorsolateral aspect ofthe abdominal wall. At each site, two dentin slices (- or + alkalinephosphatase in a concentration of 1.8 ± 0.1 mU/vig hydroxypro-line) and two bone slices (- or + alkaline phosphatase in a concen-tration of 1.6 ± 0.3 mU/,ug hydroxyproline) were inserted. After2, 4, 7, 14, or 21 days, the implants, including their surroundingsoft connective tissue capsules, were removed and fixed byimmersion in freshly prepared 5% paraformaldehyde in 0.1mol/L sodium phosphate buffer (pH 7.3) containing 2% sucrose(w/v) at 4°C for 24 hrs.
Light and electron microscopy. After fixation, part of each specimenwas embedded in LX-112 epoxy resin (Ladd", Burlington, VT,USA) and processed for electron microscopy. Ultrathin sectionswere cut with a diamond knife, stained with uranyl acetate andlead citrate, and examined in an EM1OC Zeiss electron micro-scope. The remaining parts of the implants were rinsed severaltimes in 0.1 mol/L sodium phosphate buffer (pH 7.3) containing50 mmol/L ammonium chloride, dehydrated in an ascendingseries of ethanols, cleared in xylene, and embedded in low-melt-ing paraffin. Serial sections were cut, 5 pLm thick, mounted onglass slides, dried overnight at 37°C, and stored at room tempera-ture. Some sections were deparaffinized and stained with methyl-ene blue, or according to the Von Kossa method.
Immunohistochemistry. Immediately before use, sections were de-waxed in xylene, rinsed twice in 100% ethanol, incubated in 3%H202 in 100% ethanol, followed by a rehydration step and rinsedin 0.1 mol/L sodium phosphate buffer. Dr. W.T. Butler (Houston,TX, USA) kindly donated OPN antibodies. They were raised to ratbone OPN in goats, affinity-purified, and cross-absorbed to 60-kDa glycoprotein (cx2HS-glycoprotein) as reported (Mizuno et al.,1991). It was shown, on Western immunoblots, that these affinity-purified, polyclonal antibodies react only with OPN (Pinero et al.,1995). For immunostaining, the avidin-biotin-complex peroxidasemethod was applied as reported before (Bronckers et al., 1987), bymeans of a rabbit anti-goat ABC-kit from Vector Labs(Burlingame, CA, USA). For controls, normal rabbit IgG (Miles,Yeda, Israel) was used in the same concentration as the primaryantibodies. Sections were mounted in 30% glycerol, covered witha glass cover slip, and examined.
In vivo administration of 1251-OPNPorcine 125I-OPN (0.3 mg, 120 p.Ci, 0.5 mL in PBS) was injectedinto one of the tail veins of two male Wistar rats (rats la and lb,250 g b.w.). In an additional experiment, two other rats (rats 2aand 2b) were each injected with rat 125I-OPN (0.25 mg, 100 pLCi).Two control rats (rats 3a and 3b) received the same amount of 125Iin PBS. One week prior to injection, 4 demineralized dentin slices(containing 4.4 ± 1.1 mU alkaline phosphatase/pg hydroxypro-line) and 4 control implants (no enzyme) had been installed in theback skin of rats la and lb. Rat la was killed after 30 hrs, all theother rats after 48 hrs. The implants were harvested and analyzed
j Dent Res 7801) 1999 1689
at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

1690 VandenBos et al.
for uptake of radioactivity and mineral. In addition, skin, muscle,liver, kidney, calvaria, tibia, and upper and lower jaws were takenout and fixed by immersion in 4% paraformaldehyde and 1% glu-taraldehyde in 0.1 mol/L Na-cacodylate buffer (pH 7.4). Theamount of radioactivity was determined as a function of the wetweight of these tissues. Mineralized tissues of rats la and lb werethen decalcified in 0.1 mol/L EDTA, dehydrated through a seriesof ethanols, and embedded in LX-112 epoxy resin (Ladd®).Sections 1 ,um thick were prepared for radioautography by beingdipped in L4 liquid emulsion (Ilford Ltd., Essex, England) diluted1:1 with distilled water, dried, and stored at 4°C. After 4 to 12weeks' exposure time, specimens were developed in Kodak D-19developer. They were then fixed, rinsed, and stained with methyl-ene blue. All tissues were examined, and the distribution of Aggrains was studied at a magnification of 1000x, under an oil-immersion lens. Special attention was given to the acellularcementum layer investing the continuously growing lower andupper incisors. By means of an ocular grid, grain counts weremade in standardized areas (2.5 ,um x 40 p,m) over the cementumlayer and also over the adjacent mantle dentin and periodontalligament at a distance of 10 ,um from the root surface. The concen-tration of Ag grains was also determined in standardized areasover the developing circumpulpal dentin and enamel layers in theapical part of the incisor.
Biochemical determinationsHydroxyproline was determined colorimetrically (Stegemann andStalder, 1967), phosphate as described by Hess and Derr (1975),and alkaline phosphatase activity as described previously(Beertsen and VandenBos, 1992).
To determine whether the radiolabel taken up by the hard tis-sues was still linked to OPN, we dissected the long bones of theOPN and control rats free of adhering soft tissues, pulverizedthem, and washed them twice in PBS containing proteinaseinhibitors (Steinfort et al., 1989). The residue was extracted twicewith 4 mol/L guanidine.HCl and decalcified in 0.4 mol/L EDTA,
4000
3000
a 2000
1000
00 1 2
pg P04/,jg Hyp3
Figure 1. Relation between in vitro mineralization and 125I-OPN accu-mulation. Demineralized bone sheets, either complexed or not com-plexed with alkaline phosphatase, were incubated in Iscove's ModifiedDulbecco's Medium containing radiolabeled OPN. The amount of min-eral (expressed as pLg phosphate/uWg hydroxyproline) and bound OPNwas determined after 1, 3, 7, and 15 days of incubation.
4 mol/L guanidine.HCI. After centrifugation, the EDTA-super-natant was precipitated with 5% TCA.
Samples of the EDTA extract of the long bones of rats 2a and2b were precipitated with 5% TCA, taken up in sample buffercontaining SDS and 4 M urea, and run on 4 to 15% gradient gels(BioRad Ready gels, Hercules, CA, USA) together withKaleidoscope pre-stained standards (BioRad) and rat-1251-OPN[whether or not digested by incubation for 1 hr at 37°C withthrombin (1 unit in 0.1 mol/L Tris, pH 8.0, 0.02 mol/L CaCl2;Sigma, St. Louis, MO, USA)]. After electrophoresis, the gels werestained with Stains-All (Goldberg and Warner, 1997) and cut inslices of 2.5 mm, whereafter the radioactivity in the slices wasdetermined.
Results
Mineralizing collagen sheetsIn vitro observations. Mineralization of the demineralized boneslices complexed with alkaline phosphatase (expressed as ,uginorganic phosphate per pg hydroxyproline) and uptake of theradiolabel during the in vitro incubation started from day 3 on.On day 1, no 125I label was detected. In Fig. 1, the relationbetween mineral uptake and porcine-1251I-OPN accumulationis shown. As the carrier material mineralized, a progressiveinflux of 125I-OPN was observed (df = 29; r = 0.59; p < 0.001).
In vivo observations: Immunolocalization of OPN. In line withour previous results, all collagenous implants complexedwith active alkaline phosphatase had taken up mineral fol-lowing installation in the subcutis. However, adjacent to min-eralized areas (which were Von Kossa-positive), a rim ofnon-mineralized implant matrix could often be observed (Fig.2A). At the electron microscopic level, it appeared that min-eral aggregates were always deposited in close spatial rela-tion to the collagen fibrils of the implant materials (notshown). Mineral was not found in the peri-implant soft con-nective tissue capsule. Without attached alkaline phos-phatase, none of the collagenous carriers accumulatedmineral during the experimental period.
While all control implants proved to be OPN-negative,alkaline-phosphatase-complexed implants stained positivewith the OPN antibody. The immunostaining was restrictedto the mineralized areas (Fig. 2B). In the connective tissue sur-rounding the implants, no cells staining for OPN wereobserved. This would seem to suggest that OPN was derivedfrom sources outside the immediate area of the implants.
Biochemical analysis. All mineralizing implants (demineralizeddentin complexed with alkaline phosphatase) inserted intothe back skin of rats receiving radiolabeled OPN (rats la andlb) incorporated 1251-labeled protein. Control implants with-out attached alkaline phosphatase showed very little, if any,mineral and OPN uptake (Table 1).
Uptake of 1251-OPN in mineralizingand non-mineralizing tissuesBiochemical analysis. We noted a heterogeneous distributionof the radiolabel among the various tissues (Table 2). In the
A
A
.,sI A
-
I
AIA,A I
A -
AAAA
I A
j Dent Res 7801) 1999
at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

Blood Circulationi as Sourcefor Osteopontin1
experimental animals, kidney,liver, and bone contained (on aper wet weight basis) substantialamounts of 1215-labeled material.The other tissues, specificallyskin and muscle, showed lessuptake. Of note, there were noapparent differences in the dis-tribution of radiolabel in thesetissues whether rat or porcineOPN was used. In the control T,rats (that had received unbound1251), the tissues (liver, kidney,muscle, but especially the miner- . .
alized tissues) had taken up less Fiumineralized part of deof the radiolabel compared with implant complexed with athe experimental animals (Table The mineral-containing arc
2). The relatively high amount of stain witth methylene blue (1radioactivity associated with theskin was most likely due to contamination of hairs with 125jexcreted via urine and feces (see also below under"Radioautography").
With respect to the blood, in the experimental animals,about 75 to 85%, of the label could be precipitated with 5%/TCA, indicating that most of it was still bound to protein. Inthe control rats, the label was mainly (80',) in the TCAsupernatant, indicating that it was not firmly associated with
rr
wving mineralized andmineralized dentinIkaline phosphatase.'a (asterisk) does notOOOx).
Figure 2B. Same area as in 2A (consecutive sec-tion) stained with anti-rat OPN antibodies. Notethat the OP'N-positive area (asterisk) correspondsto the mineralized part of the implant (1100x).
to this 66-kDa band, a relatively small shoulder of approxi-mately 36 kDa was observed which corresponded to the posi-tion of rat-1251-OPN digested in vitro with thrombin (Fig. 3A).
Radioautography. Radioautography of the subcutis of porcine-125I-OPN labeled rats showed no label above background. Incontrast, bones and teeth showed distinct bands of Ag grainsin the mineralized compartment. In areas where bone was
proteins.
Detection of 125J-OPN in the long boncs. To testwhether the uptake of radioactivity in theskeleton represented radiolabeled OPN, weanalyzed long bones of the 48-hour animalsbiochemically. The distribution of the radio-label is presented in Table 3. The majority ofthe label was recovered in the EDTA-guani-dine.HCl extract, whereas the PBS andguanidine.HCI extracts contained 100, and15°/ of the radiolabel, respectively. Themajority of the radiolabel in the EDTA-guanidine.HCI extract (from 70 to 90'Yo) wasprecipitable in 50,, TCA, indicating that 12"Iwas still bound to protein. In contrast, thelow level of label found in the long bones ofthe control rats was mainly found in the PBSand guanidine.HCI extracts (78% and 19%,respectively), whereas 30o was recovered inthe EDTA-guanidine.HCI fraction.Radioactivity in the latter fraction could notbe precipitated with 5%, TCA: Ninety per-cent remained in the supernatant.
Samples of the EDTA-guanidine.HClextracts of rat-12-l-OPN-labeled animalswere precipitated with 5', TCA, taken up insample buffer, and analyzed by SDS-PAGE.As shown in Fig. 3B, the majority of theradiolabel (66%S) was found as a broad bandat approximately Mr 66,000. The migrationof this band corresponded to the position ofrat-1251-OPN before injection (Fig. 3A). Next
Table 1. Amount of OPN and mineral (mean + SD) taken up in demineralized dentinimplants
OPN Mineralcpm 1251/g hydroxyproline gg P?40/g hydroxyproline
Conditions n =6 n 6
No alkaline phosphatase 2.0 + 1.0 0.6 + 0.4Attached alkaline phosphatase 14.0 + 6.0' 15.4 4.7a
d p < 0.001 according to Student's t test.
Table 2 Amount of radiolabeled OPN (cpm 1251/mg wet weight) taken up in various tissues
Porcine-OPN Rats Rat-OPN Rats Control RatsRat 1a' Rat lbb Rat 2ab Rat 2b5 Rat 3a' Rat 3b1
Skin 15 10 24 42 138 153Muscle 10 15 10 8 3 5L iver 105 61 94 83 44 29Kidney 279 267 450 366 37 30Calvaria 17 50 40 45 10 8Tibia 16 116 63 68 7 6Lower jaw 29 49 54 47 7 6Upper jaw 31 63 63 75 12 8Serum (1 gLL) not done 22 35 44 32 35
bThe tissues of rat I a were analyzed 30 hrs after 1251 OPN injection.Forty-eight hrs after injection, the tissues were analyzed.The control rats received the same dose of 1251 as rats l and 2 and were analyzed 48 hrsafter injection.
j Deiit Rcs 7801) 1999 1 691
at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
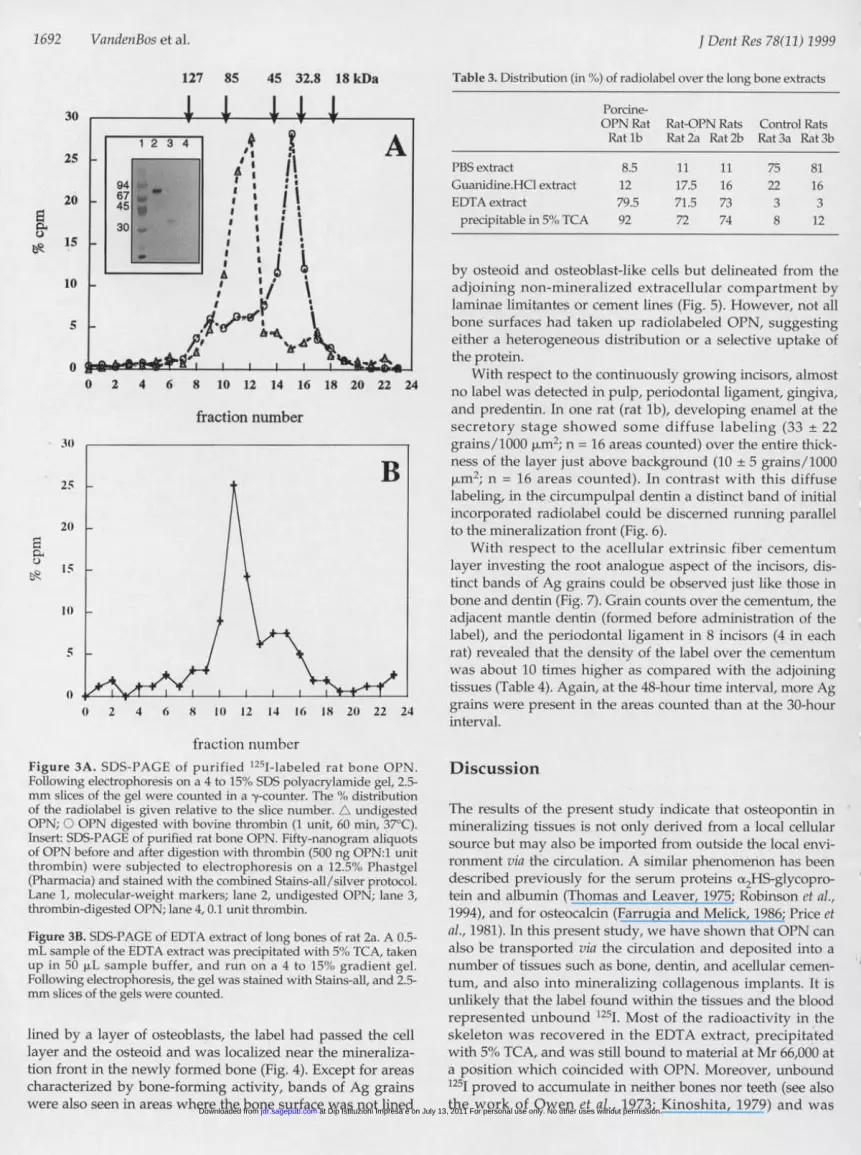
1692 VandenBos et al.
127 85 45 32.8 18 kDa
30 -
25 -
20 -
15 -
10
0
30 -
25 -
20-
5-
l -
5 -
2 4 6 8 10 12 14 16 18 20 22 24
fraction number
0 2 4 6 8 tO 12 14 16 18 20 22 24
fraction numberFigure 3A. SDS-PAGE of purified 1251-labeled rat bone OPN.Following electrophoresis on a 4 to 15'"o SDS polyacrylamide gel, 2.5-mm slices of the gel were counted in a -y-counter. The o,% distributionof the radiolabel is given relative to the slice number. A undigestedOPN; O OPN digested with bovine thrombin (1 unit, 60 min, 37°C).Insert: SDS-PAGE of purified rat bone OPN. Fifty-nanogram aliquotsof OPN before and after digestion with thrombin (500 ng OPN:1 unitthrombin) were subjected to electrophoresis on a 12.5", Phastgel(Pharmacia) and stained with the combined Stains-all/silver protocol.Lane 1, molecular-weight markers; lane 2, undigested OPN; lane 3,thrombin-digested OPN; lane 4, 0.1 unit thrombin.
Figure 3B. SDS-PAGE of EDTA extract of long bones of rat 2a. A 0.5-mL sample of the EDTA extract was precipitated with 50, TCA, takenup in 50 p.L sample buffer, and run on a 4 to 15%, gradient gel.Following electrophoresis, the gel was stained with Stains-all, and 2.5-mm slices of the gels were counted.
lined by a layer of osteoblasts, the label had passed the celllayer and the osteoid and was localized near the mineraliza-tion front in the newly formed bone (Fig. 4). Except for areascharacterized by bone-forming activity, bands of Ag grainswere also seen in areas where the bone surface was not lined
Table 3. Distribution (in co,) of radiolabel over the long bone extracts
Porcine-OPN Rat Rat-OPN Rats Control RatsRat lb Rat 2a Rat 2b Rat 3a Rat 3b
PBS extract 8.5 11 11 75 81Guanidine.HCl extract 12 17.5 16 22 16EDTA extract 79.5 71.5 73 3 3precipitable in 5%, TCA 92 72 74 8 12
by osteoid and osteoblast-like cells but delineated from theadjoining non-mineralized extracellular compartment bylaminae limitantes or cement lines (Fig. 5). However, niot allbone surfaces had taken up radiolabeled OPN, suggestingeither a heterogeneous distribution or a selective uptake ofthe protein.
With respect to the continuously growing incisors, almostno label was detected in pulp, periodontal ligament, gingiva,and predentin. In one rat (rat lb), developing enamel at thesecretory stage showed some diffuse labeling (33 + 22grains/1000 iLM2; n = 16 areas counted) over the entire thick-ness of the layer just above background (10 + 5 grains/1000gmi2; n = 16 areas counted). In contrast with this diffuselabeling, in the circumpulpal dentin a distinct band of initialincorporated radiolabel could be discerned running parallelto the mineralization front (Fig. 6).
With respect to the acellular extrinsic fiber cementumlayer investing the root analogue aspect of the incisors, dis-tinct bands of Ag grains could be observed just like those inbone and dentin (Fig. 7). Grain counts over the cementum, theadjacent mantle dentin (formed before administration of thelabel), and the periodontal ligament in 8 incisors (4 in eachrat) revealed that the density of the label over the cementumwas about 10 times higher as compared with the adjoiningtissues (Table 4). Again, at the 48-hour time interval, more Aggrains were present in the areas counted than at the 30-hourinterval.
Discussion
The results of the present study indicate that osteopontin inmineralizing tissues is not only derived from a local cellularsource but may also be imported from outside the local envi-ronment via the circulation. A similar phenomenon has beendescribed previously for the serum proteins (x2HS-glycopro-tein and albumin (Thomas and Leaver, 1975; Robinson et al.,1994), and for osteocalcin (Farrugia and Melick, 1986; Price etal., 1981). In this present study, we have shown that OPN canalso be transported via the circulation and deposited into anumber of tissues such as bone, dentin, and acellular cemen-tum, and also into mineralizing collagenous implants. It isunlikely that the label found within the tissues and the bloodrepresented unbound 1251. Most of the radioactivity in theskeleton was recovered in the EDTA extract, precipitatedwith 5% TCA, and was still bound to material at Mr 66,000 ata position which coincided with OPN. Moreover, unbound1251 proved to accumulate in neither bones nor teeth (see alsothe work of Owen et al., 1973; Kinoshita, 1979) and was
j Deiit Res 7801) 1999
at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

Blood Circuilation as Sourcefor Osteopontin
Figure 4. Radioautograph of tibia.Silver grains are localized as a wideband of radioactivity in mineralizedbone. The osteoblast layer (GB) andthe osteoid seam (OS) do not containradiolabel (950x).
Figure 5. Silver grains overlying lam-ina limitans (arrow; 1650x) at the bor-der of bone marrow and bone.
Figure 6. Silver grains (arrowheads)overlying dentin. Note that the pre-dentin (P) is free from label (1500x).
Figure 7. Silver grains over acellularextrinsic fiber cementum layer (C) ofthe lower incisor. Note that hardly anysilver grains are present in the dentin(D) and the adjoining periodontal liga-ment (P; 2000x).
rapidly excreted from the body.Thus, we believe that the greaterpart of radiolabeled OPN whichaccumulated in the mineralizedtissues had remained intact.However, a small proportion ofthe label migrated at a position ofapproximately 36 kDa and proba-bly represents OPN digested byproteolytic action. Partial breakdown could have occurredeither by blood thrombin after i.v. injection or during the bio-chemical preparation of the bone samples.
At present, we do not know the cellular origin of OPN incirculation. OPN, unlike many other serum proteins, is notsynthesized by the liver (Kerr et al., 1991; Nohutcu et al.,1996). Perhaps it is produced in excess by bone-associatedcells, as is also the case with osteocalcin (Price, 1985;Hauschka et al., 1989). Normal turnover of bone will alsorelease non-collagenous proteins into the circulation.Moreover, other cells-like activated macrophages (Senger etal., 1988), immune T-cells (Patarca et al., 1993), and vascularsmooth-muscle cells (Giachelli et al., 1993)-may be consid-ered as sources for serum OPN.We can only speculate on the relative amount of this mole-
cule in calcifying tissues that is derived from the blood. Givenan average circulating blood volume of about 10 mL in therat, the injected dose of OPN amounted to 30 Vig/mL.Although the concentration of constitutive OPN in rat serumis unknown, a comparison with man (30 Rg/L; Bautista et al.,1996) suggests that the injected dose was about three ordersof magnitude higher. However, a direct comparison withman is difficult, not only because the actual concentrations inthe tissues and blood may differ among the two species, butalso because of its dynamic behavior in terms of turnover.With respect to one of the other bone-seeking proteins, osteo-calcin, the concentration in rat serum is considerably higherthan that in man, indicating species differences (Hauschka,1986; Lian and Gundberg, 1988).We cannot exclude the possibility that the relatively high
dose of injected OPN could have led to a somewhat artificialdistribution pattern. An example of this may be its presence
in the developing enamel in rat lb. As far as we are aware,there are no reports in the literature indicating that the pro-tein is present in developing enamel matrix. However,immunohistochemical techniques may not be sensitiveenough to detect trace amounts of OPN in enamel. The rela-
Table 4. Ag grain counts (mean + SD) in acelLular extrinsic fibercementum and adjacent areas of mantle dentin and periodontalligament of rat incisors
Incisor Adjacent Area ofRat Location Areas Counted AEFC' Dentin PDL5
la upper left 10 10 + 3 2 ±1 2 +1upper right 10 28 ± 8 4 +2 4 ±1lower left 10 8+ 5 1+1 1 ±1lower right 10 6 + 2 1 +1 I ±1
mean 13+10 2+1 2+±1
lb upper left 9 33 ± 8 2 +1 1 ±1upper right 7 40 + 14 3± 1 3 +1lowerleft 11 17+13 3+1 1+1lower right 8 23 + 13 2 +1 4 +1mean 28+10 3--1 2±2
Rats were injected with 1251-OPN and killed 30 hrs (rat la) and 48hrs (rat lb) thereafter. Grain counts of radioautographs of theincisors were performed. The number of grains pl'1 unit area (100mm2) was corrected for background. From each incisor, 8 to 11areas were counted.Acellular extrinsic fiber cementum.
b Periodontal ligament.According to Wilcoxon signed-ranks test: AEFC vs. Dentin/PDL, p< 0.001.
1693I Dent Res 7801) 1999
at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

1694 VandenBos et al.
tively high dose of OPN we injected, in combination with thehigh sensitivity of the radioautographic method, has shownthat this protein can enter the enamel at the secretory stage.
The overall distribution pattern of the injected radiolabelwas quite similar to what is known for tissue OPN (McKee etal., 1996). Part of it could be detected in the liver and kidney,suggesting that these organs are involved in the clearance ofOPN, just as they do for osteocalcin (Price et al., 1981;Farrugia and Melick, 1986). Besides, OPN was clearly associ-ated with mineralizing tissues. Some of these were lined by acontinuous layer of formative cells (osteoblasts, odonto-blasts). However, we also observed that the labeled proteinwas present in mineralized tissues not lined by a continuouslayer of cells, such as in cement lines, laminae limitantes, andalso in the acellular extrinsic fiber cementum layer investingthe root-analogue portion of the continuously growingincisors. With respect to the cementum, it is uncertain towhat extent the blood functions as a major reservoir of OPN.However, we clearly have shown that proteins circulating inthe blood can be taken up by this tooth-supporting tissue. Inthis respect, our results lend support to the hypothesis(Beertsen et al., 1997) that some of the non-collagenous pro-teins in acellular cementum may come from the circulationand are deposited together with calcium-phosphate precipi-tates, and that their influx into the forming mineral does notnecessarily require synthetic activity of local cells. Influx ofOPN from the blood was also conspicuous in the minerali-zing collagenous carriers. Deposition of the moleculeoccurred synchronously with the deposition of the mineralphase. In carriers that did not mineralize, no 125I-OPN uptakewas noted. Also, the in vivo implants demonstrated uptake ofmineral together with OPN, whereas the surrounding cellsdid not express the protein. This may indicate that OPNincorporation follows mineralization and is not a prerequi-site for it.
An interesting observation was that radiolabeled OPNcan readily pass the osteoblast layer and' the osteoid seamand is deposited near the mineralization front. OPN was alsofound in cement lines. However, not all bone surfaces hadtaken up radiolabeled OPN. Perhaps this local variation inuptake reflects differences in the dynamics of the mineraliza-tion process and/or the exchange of molecules with theblood stream. Selective uptake of OPN from the blood alsolikely occurs along the surfaces of implanted biomaterialssuch as hydroxyapatite particles and titanium (Kawaguchi etal., 1993; Ramakrishnan et al., 1995) and in calcifying athero-sclerotic plaques (Giachelli et al., 1993).
Next to acellular cementum and bone, radiolabeled OPNwas detected in dentin. Our observations suggest for the firsttime that OPN in dentin can originate directly from the circu-lation, a possibility put forward previously by Helder et al.(1993), who reported immunostaining of OPN in (pre)dentinwithout mRNA being expressed in the odontoblasts.
Recently, OPN was identified in human dental calculusmatrix. It was suggested that this OPN originated from salivaand/or crevicular fluid (Kido et al., 1995). This finding is inline with our data and supports the conclusion that OPN inacellular cementum and other mineralizing tissues may-atleast partially-originate from sources outside the immediateenvironment following its transportation via serum.
Acknowledgments
We thank Kevin Wamer for preparation of rat OPN, Dr. W.T.Butler for the OPN antibody, Mrs. Anneke Niehof for excel-lent technical support, and Dr. Vincent Everts for critical read-ing of the manuscript. This study was supported by grantsfrom the Netherlands Institute for Dental Sciences (TvdB,WB), the Royal Netherlands Academy for Arts and Sciences(AB), and the Medical Research Council of Canada (HAG).Part of this work has been presented at the 3rd Joint Meetingof the Continental European Division and the ScandinavianAssociation of the IADR, 1996, Berlin, Germany.
References
Arzate H, Olson SW, Page RC, Gown AM, Narayanan AS (1992).Production of a monoclonal antibody to an attachment pro-tein derived from human cementum. FASEB 1 6:2990-2995.
Bartold PM, Reinboth B, Nakae H, Narayanan AS, Page RC(1990). Proteoglycans of bovine cementum: isolation andcharacterization. Matrix 10:10-19.
Bautista DS, Saad Z, Chambers AF, Tonkin KS, O'Malley FP,Singhal H, et al. (1996). Quantification of osteopontin inhuman plasma with an ELISA: basal levels in pre- and post-menopausal woman. Clin Biochem 29:231-239.
Beertsen W, VandenBos T (1992). Alkaline phosphatase inducesmineralization of sheets of collagen implanted subcuta-neously in the rat. J Clin Invest 89:1974-1980.
Beertsen W, VandenBos T, Everts V (1997). Continuous growthof acellular extrinsic fiber cementum. Acta Med Dent Helv2:103-115.
Bronckers AL, Gay S, Finkelman RD, Butler WT (1987).Immunolocalization of Gla proteins (osteocalcin) in rat toothgerms: comparison between indirect immunofluorescence,peroxidase-antiperoxidase, avidin-biotin-peroxidase com-plex, and avidin-biotin-gold complex with silver enhance-ment. J Histochem Cytochem 35:825-830.
Bronckers AL, Farach-Carson MC, Van Waveren E, Butler WT(1994). Immunolocalization of osteopontin, osteocalcin, anddentin sialoprotein during dental root formation and earlycementogenesis in the rat. J Bone Miner Res 9:833-841.
Butler WT, Ritchie H (1995). The nature and functional signifi-cance of dentin extracellular matrix proteins. Int J Dev Biol39:169-179.
Chen J, Singh K, Mukherjee BB, Sodek J (1993). Developmentalexpression of osteopontin (OPN) messenger RNA in rat tis-sues: evidence for a role for OPN in bone formation andresorption. Matrix 13:113-123.
Connor NS, Aubin JE, Melcher AH (1984). The distribution offibronectin in rat tooth and periodontal tissues: an immuno-fluorescence study using a monoclonal antibody. I HistochemCytochem 32:565-572.
Farrugia W, Melick RA (1986). Metabolism of osteocalcin. CalcifTissue Int 39:234-238.
Fisher LW, Termine JD (1985). Noncollagenous proteins influen-cing the local mechanisms of calcification. Clin Orthop Rel Res200:362-385.
Giachelli CM, Bae N, Almeida M, Denhardt DT, Alpers CE,Schwartz SM (1993). Osteopontin is elevated during neo-
j Dent Res 78(11) 1999
at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

Blood Circulation as Sourcefor Osteopontin
intima formation in rat arteries and is a novel component ofhuman atherosclerotic plaques. I Clin Invest 92:1686-1696.
Goldberg HA, Sodek J (1994). Purification of mineralized tissue-associated osteopontin. J Tissue Culture Meth 16:211-215.
Goldberg HA, Wamer KJ (1997). The staining of acidic proteins onpolyacrylamide gels: enhanced sensitivity and stability of"Stains-All" staining in combination with silver nitrate. AnalBiochem 251:227-233.
Hauschka PV (1986). Osteocalcin: the vitamin K-dependent Ca2l-binding protein of bone matrix. Haemostasis 16:258-272.
Hauschka PV, Lian JB, Cole DE, Gundberg CM (1989). Osteocalcinand matrix Gla protein: vitamin K-dependent proteins inbone. Physiol Rev 69:990-1045.
Helder MN, Bronckers AL, Woltgens JH (1993). Dissimilar expres-sion patterns for the extracellular matrix proteins osteopontin(OPN) and collagen type I in dental tissues and alveolar boneof the neonatal rat. Matrix 13:415-425.
Hess HH, Derr JE (1975). Assay of inorganic and organic phos-phorus in the 0.1-5 nanomole range. Anal Biochem 63:607-613.
Kawaguchi H, McKee MD, Okamoto H, Nanci A (1993).Immunocytochemical and lectin-gold characterization of theinterface between alveolar bone and implanted hydroxy-apatite in the rat. Cells Mater 3:337-350.
Kerr JM, Fisher LW, Termine JD, Young MF (1991). The cDNAcloning and RNA distribution of bovine osteopontin. Gene108:237-243.
Kido J, Kasahara C, Ohishi K, Nishikawa S, Ishida H, YamashitaK, et al. (1995). Identification of osteopontin in human dentalcalculus matrix. Arch Oral Biol 40:967-972.
Kinoshita Y (1979). Incorporation of serum albumin into develop-ing dentine and enamel matrix in the rabbit incisor. CalcifTissue Int 29:41-46.
Lekic P, Sodek J, McCulloch CA (1996). Osteopontin and bonesialoprotein expression in regenerating rat periodontal liga-ment and alveolar bone. Anat Rec 244:50-58.
Lian JB, Gundberg CM (1988). Osteocalcin: biochemical considera-tions and clinical applications. Clin Orthop Rel Res 226:267-291.
Linde A (1989). Dentin matrix proteins: composition and possiblefunctions in calcification. Anat Rec 224:154-166.
MacNeil RL, Sheng N, Strayhorn C, Fisher LW, Somerman MJ(1994). Bone sialoprotein is localized to the root surface dur-ing cementogenesis. I Bone Miner Res 9:1597-1606.
McKee MD, Zalzal S, Nanci A (1996). Extracellular matrix in toothcementum and mantle dentin: localization of osteopontinand other noncollagenous proteins, plasma proteins, and gly-coconjugates by electron microscopy. Anat Rec 245:293-312.
Mizuno M, Farach-Carson MC, Pinero GJ, Fujisawa R, Brunn JC,Seyer JM, et al. (1991). Identification of the rat bone 60K acidicglycoprotein as (x2HS-glycoprotein. Bone Miner 13:1-12.
Nakae H, Narayanan AS, Raines E, Page RC (1991). Isolation andpartial characterization of mitogenic factors from cementum.Biochemistry 30:7047-7052.
Nohutcu RM, McCauley LK, Shigeyama Y, Somerman MJ (1996).Expresssion of mineral-associated proteins by periodontalligament cells: in vitro vs. ex vivo. J Periodont Res 31:369-372.
Owen M, Triffitt JT, Melick RA (1973). Albumin in bone. Hard tis-sue growth, repair and remineralization. Ciba Fdn Symp11:263-293.
Patarca R, Saavedra RA, Cantor H (1993). Molecular and cellularbasis of genetic resistance to bacterial infection: the role of theearly T-lymphocyte activation-1/osteopontin gene. Crit RevImmunol 13:225-246.
Pinero GJ, Farach-Carson MC, Devoll RE, Aubin JC, Brunn JC,Butler WT (1995). Bone matrix proteins in osteogenesis andremodelling in the neonatal rat mandible as studied byimmunolocalization of osteopontin, bone sialoprotein,(x2HS-glycoprotein and alkaline phosphatase. Arch Oral Biol40:145-155.
Price PA (1985). Vitamin K-dependent formation of bone Gla pro-tein (osteocalcin) and its function. Vitamin Hormones 42:65-108.
Price PA, Williamson MK, Lothringer JW (1981). Origin of thevitamin K-dependent bone protein found in plasma and itsclearance by kidney and bone. I Biol Chem 256:12760-12766.
Ramakrishnan PR, Lin WL, Sodek J, Cho MI (1995). Synthesis ofnoncollagenous extracellular matrix proteins during devel-opment of mineralized nodules by rat periodontal ligamentcells in vitro. Calcif Tissue Int 57:52-59.
Reichert T, Stbrkel S, Becker K, Fisher LW (1992). The role ofosteonectin in human tooth development: an immunohisto-logical study. Calcif Tissue Int 50:468472.
Roach HI (1994). Why does bone matrix contain non-collagenousproteins? The possible roles of osteocalcin, osteonectin, osteo-pontin and bone sialoprotein in bone mineralisation andresorption. Cell Biol Int 18:617-628.
Robinson C, Brookes SJ, Kirkham J, Shore RC, Bonass WA (1994).Uptake and metabolism of albumin by rodent incisor enamelin vivo and postmortem: implications for control of mineral-ization by albumin. Calcif Tissue Int 55:467472.
Schroeder HE (1986). The periodontium. In: Handbook of micro-scopic anatomy. Vol. V/5. Oksche A, Vollrath L, editors.Berlin: Springer-Verlag, pp. 23-129.
Senger DR, Perruzzi CA, Gracey CF, Papadopoulos A, Tenen DG(1988). Secreted phosphoprotein associated with neoplastictransformation. Close homology with plasma proteinscleaved during blood coagulation. Cancer Res 48:5770-5774.
Stegemann H, Stalder K (1967). Determination of hydroxyproline.Clin Chim Acta 18:267-273.
Steinfort J, VandenBos T, Beertsen W (1989). Differences betweenenamel-related and cementum-related dentin in the ratincisor with special emphasis on the phosphoproteins. J BiolChem 264:2840-2845.
Takano-Yamamoto T, Takemura T, Kitamura Y, Nomura S (1994).Site-specific expression of mRNAs for osteonectin, osteocal-cin, and osteopontin revealed by in situ hybridization in rat
periodontal ligament during physiological tooth movement.
J Histochem Cytochem 42:885-896.Thomas M, Leaver AG (1975). Identification and estimation of
plasma proteins in human dentine. Arch Oral Biol 2:217-218.VandenBos T, Beertsen W (1994). Bound phosphoproteins
enhance mineralization of alkaline phosphatase-collagencomplexes in vivo. J Bone Miner Res 9:1205-1209.
Zhang X, Schuppan D, Becker J, Reichart P, Gelderblom HR(1993). Distribution of undulin, tenascin, and fibronectin inthe human periodontal ligament and cementum: compara-tive immunoelectron microscopy with ultra-thin cryosec-tions. J Histochem Cytochem 41:245-251.
J Dent Res 78(11) 1999 1695
at Dip Istituzioni Impresa e on July 13, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from