binding to complement factors and activation of the alternative pathway by acanthamoeba
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Immunobiology 216 (2011) 225–233
Contents lists available at ScienceDirect
Immunobiology
journa l homepage: www.e lsev ier .de / imbio
Binding to complement factors and activation of the alternativepathway by Acanthamoeba
Wilawan Pumidonminga, Julia Walochnika,∗, Elke Dauberb, Franz Petryb,1
a Department of Medical Parasitology, Institute of Specific Prophylaxis and Tropical Medicine, Medical University of Vienna, Kinderspitalgasse 15, 1090 Vienna, Austriab Institute of Medical Microbiology and Hygiene, University Medical Center of the Johannes Gutenberg University Mainz, Augustusplatz/Hochhaus, D-55101 Mainz, Germany
a r t i c l e i n f o
Article history:Received 26 January 2010Received in revised form 30 March 2010Accepted 2 May 2010
Keywords:AcanthamoebaAlternative pathwayComplementNormal human serum (NHS)Normal mouse serum (NMS)
a b s t r a c t
Acanthamoeba can cause severe ocular and cerebral diseases in healthy and immunocompromised indi-viduals, respectively. Activation of complement appears to play an important role in host defence againstinfection. The exact mechanism, however, is still unclear. The aim of the present study was to investigatethe effect of normal human serum (NHS) and normal mouse serum (NMS) on Acanthamoeba trophozoites,the binding of different complement factors to Acanthamoeba and the activation of the complementsystem. Moreover, we aimed to work out any possible differences between different strains of Acan-thamoeba. A virulent T4 strain, a non-virulent T4 strain and a virulent T6 strain were included in thestudy. It was shown that NHS, but not NMS clearly has amoebicidal properties. After 5 min of incubationwith NHS, amoebae showed plasma membrane disruption and extrusion of intracellular components.Cells were completely destroyed within 60 min of incubation in NHS but stayed intact after incubation inheat-inactivated serum. The binding of human C3 and C9 to amoebae was established by immunoblot-ting. Although incubation with mouse serum did not result in lysis of Acanthamoeba trophozoites animmunofluorescence assay (IFA) demonstrated a strong deposition of mouse complement factor C3 acti-vation products, moderate binding of C1q, but no binding of MBL-A and MBL-C. EDTA inhibited the bindingof C3 to acanthamoebae. Binding of amoebae to C3b was observed with sera from C1qa−/− and MBL-A/C−/− mice, but not with serum from Bf/C2−/− mice demonstrating an activation of complement viathe alternative pathway. There were no significant differences between the three Acanthamoeba strainsinvestigated. Altogether, our results prove that NHS is amoebolytic and that Acanthamoeba binds to C3and C9 and activates the complement system via the alternative pathway.
© 2010 Elsevier GmbH. All rights reserved.
Introduction
Acanthamoebae are free-living, but potentially pathogenicprotozoans. In healthy individuals, acanthamoebae can cause Acan-thamoeba keratitis (AK) predominantly occurring in contact lenswearers (Carvalho et al. 2009; Stapleton et al. 2009) and inimmunocompromised individuals, acanthamoebae can cause gran-ulomatous amoebic encephalitis (GAE) (Illingworth and Cook 1998;Martinez 1982; Walochnik et al. 2008). In addition, Acanthamoebaskin lesions and pneumonitis have been described occasionally(Gullett et al. 1979; Van Hamme et al. 2001).
Abbreviations: AK, Acanthamoeba keratitis; GAE, granulomatous amoebicencephalitis; IFA, immunofluorescence assay; NHS, normal human serum; NMS,normal mouse serum.
∗ Corresponding author. Tel.: +43 1 40490 79446; fax: +43 1 40490 79435.E-mail address: [email protected] (J. Walochnik).
1 Present address: Planalyze GmbH, Hermannstrasse 15, D-60318 Frankfurt, Ger-many.
Acanthamoebae are ubiquitously distributed in various habi-tats making contact with this organism almost unavoidable. Thus,exposure to Acanthamoeba antigens occurs frequently. Serologicalsurveys have demonstrated that 50–100% of healthy individualswithout known history of Acanthamoeba infection exhibit specificantibodies against Acanthamoeba (Cursons et al. 1980; Walochniket al. 2001a). However, the vast majority of healthy individuals donot develop disease in spite of contact to Acanthamoeba (Alizadeh etal. 2001; Brindley et al. 2009; Cursons et al. 1980). Adaptive as wellas innate immunity may play a role in Acanthamoeba infections.
In amoebae, including parasitic Entamoeba, but also faculta-tively pathogenic Acanthamoeba and Naegleria, direct contact withnormal human serum (NHS) results in complement activation(Ferrante and Rowan-Kelly 1983; Ortiz-Ortiz et al. 1974, 1978;Whiteman and Marciano-Cabral 1987). The complement systemcan be activated via the classical, the lectin and the alternativepathway, respectively and activation of any of these three path-ways can lead to formation of the membrane attack complexand lysis of the pathogen (Roestenberg et al. 2007; Petry et al.2008; Tsoni et al. 2009). Entamoebae, relatively closely related to
0171-2985/$ – see front matter © 2010 Elsevier GmbH. All rights reserved.doi:10.1016/j.imbio.2010.05.002

Author's personal copy
226 W. Pumidonming et al. / Immunobiology 216 (2011) 225–233
Acanthamoeba, activate the complement system via both, the alter-native and the classical pathway, resulting in cell lysis (Calderonand Schreiber 1985; Walderich et al. 1997). In initial studies, dif-ferences of complement susceptibility were observed betweenvirulent and non-virulent entamoebae, the pathogenic Entamoebahistolytica being more susceptible to complement lysis than thenon-pathogenic Entamoeba dispar (Hamelmann et al. 1992, 1993).However, Walderich et al. (1997) performed a large scale studyon altogether 77 clincal isolates and found both, E. histolyticaand E. dispar, susceptible to complement lysis. In the phyloge-netically distant genus Naegleria, the virulent Naegleria fowleriwere shown to be complement-resistant, and Naegleria gruberi, anon-virulent species, to be complement-sensitive (Whiteman andMarciano-Cabral 1987). In Acanthamoeba, both, virulent and non-virulent strains, were reported to be lysed by human complement(Ferrante 1991; Ferrante and Rowan-Kelly 1983). Nevertheless,certain Acanthamoeba strains can resist the lytic activity of com-plement, whereby this ability seems to correlate to the virulence ofthe particular strain (Toney and Marciano-Cabral 1998).
The genus Acanthamoeba has been classified into 15 genotypesbased on 18S rRNA gene sequencing (Gast 2001; Gast et al. 1996;Horn et al. 1999; Stothard et al. 1998). Genotype T4 is not only themost ubiquitous genotype in the environment, but also the majorgenotype associated with disease (Booton et al. 2009; Niyyati et al.2009). However, not all T4 strains are virulent, and several repre-sentatives from other genotypes can also cause disease (Walochniket al. 2008, 2000a,b). Thus, virulent and non-virulent acanthamoe-bae from different genotypes might show differences of bindingand activation of the complement system. The aim of this studywas to examine the effect of NHS on Acanthamoeba, in particular thebinding of different complement factors to Acanthamoeba tropho-zoites, the activation of the complement system by Acanthamoebaand any possible differences between three isolates belonging totwo different genotypes.
Materials and methods
Amoebae
A virulent Acanthamoeba genotype T4 strain (3ST), a non-virulent genotpye T4 strain (9GU), and a virulent genotype T6 strain(11DS) were used for this study. Strains 3ST and 11DS are iso-lates from clinical specimens of patients with severe AK. Strain9GU was isolated from the contact lens case of a patient withbacterial corneal infiltrates. Corneal scrapings were negative foramoebae and the infection completely healed after anti-bacterialtreatment – this amoebal strain is thus considered as non-virulent(Walochnik et al. 2000a,b). Sequences are available under the fol-lowing GenBank accession numbers: AF260723 (3ST), AF251939(11DS) and AF251938 (9GU). In in vitro pathogenicity tests, all threestrains show cytopathic potential at 34 ◦C, however, strains 3STand 11DS also grow at higher temperatures and migrate underagarose, while strain 9GU does not grow at high temperature andonly slightly migrates under agarose (Pumidonming et al. 2010;Walochnik et al. 2000b). All strains were freshly axenised prior tothe experiments in order to avoid long-term axenic culture effects;none of the strains had been passed through experimental ani-mals.
Amoebae were cultured in sterile-filtrated proteose peptoneyeast extract-glucose medium (Visvesvara and Balamuth 1975)in 150-cm2 tissue culture flasks (ASAI Glass, Osaka, Japan) atroom temperature. Acanthamoeba trophozoites were harvested atexponential growth phase by centrifugation at 500 × g for 10 min.Trophozoite pellets were washed twice with PBS, counted and usedfor further analysis.
Sera and antibodies
Normal human serum was produced by pooling 10 randomlyselected sera from healthy individuals. Heat-inactivated NHS wasprepared by incubating NHS at 56 ◦C for 30 min (Pillemer et al. 1953;Vogt et al. 1974).
Mouse serum is similar to human serum concerning comple-ment components (Borsos and Cooper 1961), however, has lowcomplement activity (Brown 1943, McGhee 1952). Thus, mouseserum does not result in rapid lysis of Acanthamoeba trophozoitesand can be used to demonstrate the deposition of mouse com-plement molecules on Acanthamoeba trophozoites. Sera from wildtype C57BL/6 mice as well as sera from MBL-A/C−/− (Shi et al.2004), Bf/C2−/− (Taylor et al. 1998), C1qa−/− (Botto et al. 1998)mice, all with the genetic background of the C57BL/6 strain, wereused in this study. All serum samples were stored separately at−70 ◦C. Monoclonal rat antibodies to mouse MBL-A (clone 8G6),mouse MBL-C (clone 14D12) and mouse C3b/iC3b/C3c (clone 3/26)were purchased from HyCult biotechnology (Uden, NL). Polyclonalantibodies to mouse C1q were produced in a goat (Petry et al.1991). Goat-anti-rat IgG-AlexaFluor 488 or donkey anti-goat IgG-AlexaFluor 488 (Molecular Probes, Invitrogen, Karlsruhe, Germany)was used as secondary antibodies.
Amoebicidal activity of human serum
Axenic cultures of amoeba trophozoites were harvested by cen-trifugation at 500 × g for 10 min, washed twice with PBS, countedand adjusted to a cell density of 1 × 106 cell/ml with PBS. Then100 �l of this suspension were mixed with 100 �l of NHS, NHS plus5 mM EDTA, or NHS plus 5 mM EGTA, respectively, in Eppendorfreaction tubes and incubated at 37 ◦C for 1 h. Trophozoite mor-phology was examined at 0, 5, 10, 20 and 60 min of incubationby differential interference contrast (DIC) microscopy (Axioskop2 equipped with an AxioCam Hrc digital camera, Zeiss). Livingand dead amoebae were counted after 60 min of incubation usinga hemacytometer. Living trophozoites appear bright and mov-ing, killed trophozoites are transparent and the cell membrane isdestroyed (Walderich et al. 1997). The percentage of killed tropho-zoites was calculated. Heat-inactivated serum or PBS served ascontrols.
Immunoblot analysis
For detection of bound human C3, trophozoites were incu-bated in PBS, heat-inactivated NHS, or diluted NHS (final serumdilutions: 1:4, 1:10 or 1:20) at 37 ◦C for 1 h. After incubation, sam-ples were washed thoroughly 2 times with PBS by centrifugation.Cells were resuspended in a minimal volume (below 25 �l) of1 M NH2OH in 0.2 M NaHCO2, pH 10 and incubated for 60 min at37 ◦C in order to release thioester-bound C3 activation fragments.Then the supernatants were separated from cells by centrifuga-tion at high speed at 12,000 rpm for 10 min and transferred tonew tubes. Reducing SDS sample buffer (4×) (63 mM Tris/HCl, pH6.8, 10% (v/v) glycerol, 2% (w/v) SDS, 5% (v/v) 2-mercaptoethanoland 0.01% (v/v) bromophenol blue) was added and samples wereheated at 95 ◦C for 10 min. Proteins were separated by SDS-PAGE(10%).
For the detection of bound C9, trophozoites were incubated withPBS, heat-inactivated NHS or diluted NHS (1:4, 1:10 or 1:20) at37 ◦C for 1 h. Samples were washed thoroughly with PBS by cen-trifugation. The cell pellets were supplemented with 4× reducingSDS buffer and heated at 95 ◦C for 10 min. Proteins were separatedby SDS-PAGE (10% SDS).
Proteins separated by SDS-PAGE were transferred to nitrocellu-lose (Schleicher and Schuell, Dassel, Germany) using semi-dry elec-

Author's personal copy
W. Pumidonming et al. / Immunobiology 216 (2011) 225–233 227
Table 1Amoebicidal activity of NHS.
Treatment Trophozoites killed (%)
3ST 9GU 11DS
PBSa 0 ± 0 0 ± 0 0 ± 0Heat-inactivated NHSa 0 ± 0 0 ± 0 0 ± 0NHSa 98.4 ± 0.9 98.9 ± 1.2 99.4 ± 1.1
a Incubation time: 60 min.
troblotting. The blot membranes were blocked with 2% skimmedmilk powder in Tris-buffered saline (TBS). Primary antibodies weregoat-anti-human C3 (C-7761, Sigma) or mouse-anti-human C9(cl.X197, HyCult biotechnology). Secondary antibodies were rabbit-anti-goat-alkaline phosphatase conjugate (AP) (A-4062, Sigma) orgoat-anti-mouse-AP (A-3562, Sigma). BCIP/NBT (Sigma/Fast tablet,Sigma) served as a substrate and chromogen for alkaline phos-phatase.
Immunofluorescence assay (IFA)
Trophozoite suspensions were applied to poly-l-lysine coatedslides (Adcell, Roth, Karlsruhe, Germany) followed by air-dryingand fixation with cold acetone (−20 ◦C). Slides were washed withPBS in a cuvette for 5 min, then blocked with 1% BSA/PBS for 1 hat room temperature and washed once again with PBS. After that,slides were incubated with mouse serum, mouse serum plus 20 mMEDTA, mouse serum plus 20 mM mannose or knockout mouseserum from MBL-A/C−/−, Bf/C2−/− or C1qa−/− mice for 1 h in amoist chamber at room temperature. After two washing steps withPBS, primary antibodies, rat anti-mouse MBL-A, anti-mouse MBL-C,rat monoclonal anti-mouse C3b/iC3b/C3c or polyclonal goat-anti-mouse C1q were applied to slides and incubated for 1 h in a moistchamber at room temperature. Negative controls without primaryantibodies were performed for each test. Slides were washed 3times with PBS and then incubated with secondary antibodies (anti-rat IgG-AlexaFluor 488 or anti-goat IgG-AlexaFluor 488). Slides
Fig. 1. Amoebicidal activity of NHS against Acanthamoeba. Differential interference contrast microscopy of Acanthamoeba trophozoites of the virulent T4 strain (3ST), incubatedwith PBS (Panel I), heat-inactivated NHS (Panel II) or NHS (Panel III) at 0 min (A), 5 min (B), 10 min (C), 20 min (D) and 60 min (E). Scale bar: 50 �m.

Author's personal copy
228 W. Pumidonming et al. / Immunobiology 216 (2011) 225–233
were examined under epifluorescence and differential interferencecontrast.
Results
Amoebicidal activity of human serum against Acanthamoeba
Normal human serum exhibited amoebicidal activities againstAcanthamoeba trophozoites. After 5 min of incubation with NHS,trophozoites exhibited plasma membrane disruption. After 20 minof incubation extrusion of intracellular components was observed.Forty minutes later, the cells were completely destroyed and theorganelles had disappeared. More than 98% of the cells of all threestrains were killed after incubation in NHS for 60 min (Table 1).Trophozoites that had been incubated for the same period of timewith PBS or heat-inactivated NHS remained viable, exhibiting pseu-dopodia, intact plasma membranes and organelles (Fig. 1). Theamoebicidal property of NHS was also inhibited by EDTA, but notby EGTA indicating a requirement for Mg2+. Acanthamoeba tropho-zoites that had been incubated with NHS in the presence of 5 mMEGTA were completely lysed within 60 min, while trophozoite cellsstayed intact after incubating with NHS plus 5 mM EDTA (notshown). Similar results were obtained for all three strains, 3ST(virulent T4), 9GU (non-virulent T4) and 11DS (virulent T6) (notshown).
Binding of human C3 and human C9 to Acanthamoeba
As incubation of Acanthamoeba trophozoites in NHS resultedin complete lysis of the cells, dilutions of NHS had to be usedto demonstrate the binding of human C3 and C9 to amoebae byimmunoblotting. After incubating the trophozoites with dilutedNHS and intense washing of the cells with PBS, thioester-boundC3 fragments on the amoebae were eluted with hydroxylamine.Immunoblotting showed anti-C3 reactive bands of approximately69 kDa and 40 kDa suggesting a further degradation of C3b to iC3b.The binding of C3 to the trophozoites was not observed afterincubating the trophozoites with PBS and heat-inactivated NHS(Fig. 2A). C9 was also detected on the immunoblots, with multi-ple bands of molecular weights in the 69–34 kDa range. Human C9binding did not occur in the incubations of the trophozoites withPBS and heat-inactivated NHS (Fig. 2B). This was observed for allthree investigated Acanthamoeba isolates.
Binding of mouse MBL-A, MBL-C, C3b and C1q to Acanthamoebatrophozoites
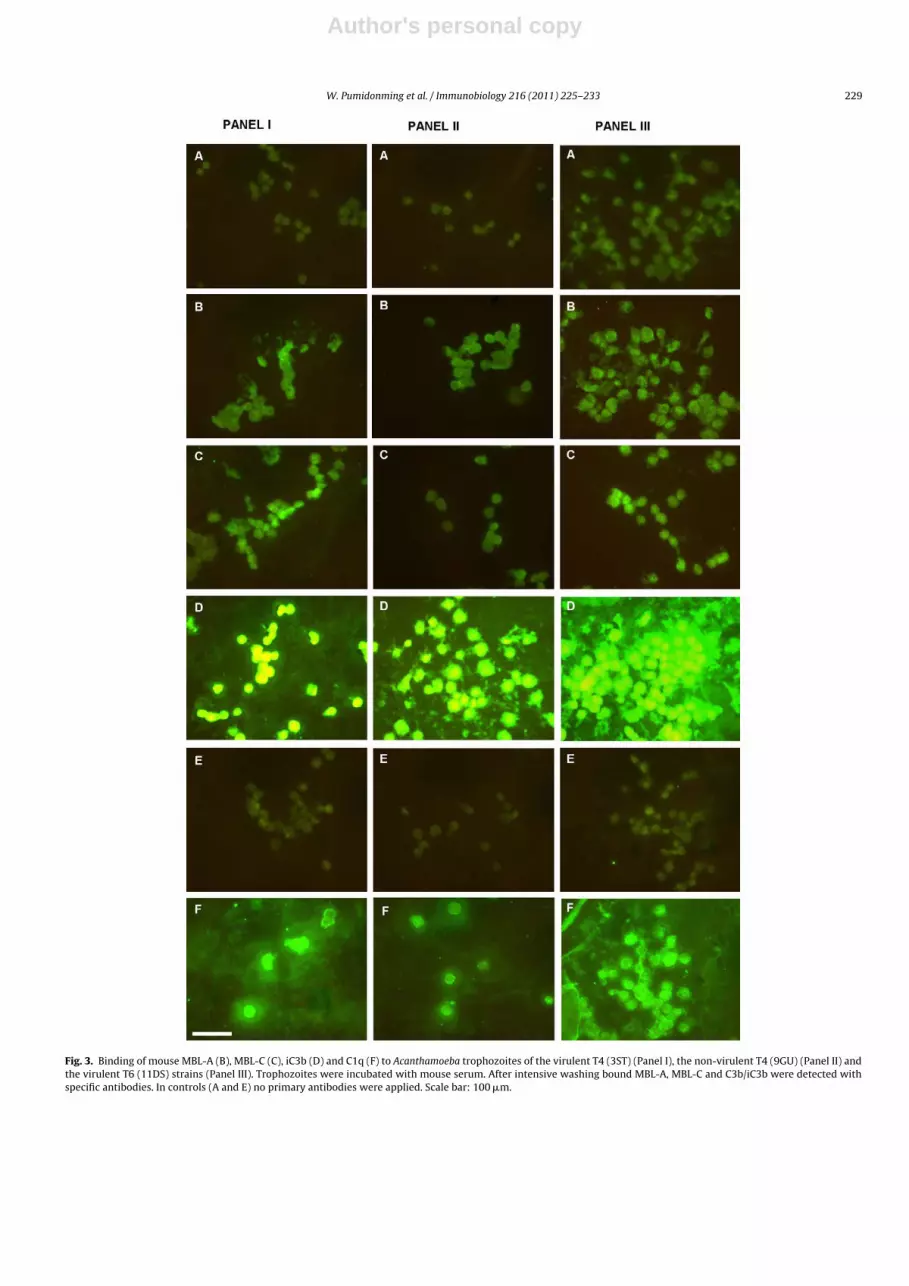
In order to investigate the effect of NMS on Acanthamoebatrophozoites experiments similar to the NHS incubations wereperformed. In contrast to human serum, mouse serum did notresult in lysis of the trophozoites. Therefore, we could demon-strate the binding of mouse MBL-A, MBL-C, C3b, and C1q toAcanthamoeba trophozoites by IFA. Strong binding of anti-C3b wasobserved, indicating a massive deposition of C3 activation products.Anti-C1q exhibited moderate binding to the trophozoites. Reactiv-ities of anti-MBL-A and anti-MBL-C with trophozoites were weak.Again, no differences could be detected between the three strains3ST (virulent T4), 9GU (non-virulent T4) and 11DS (virulent T6)(Fig. 3).
Binding of complement factors to Acanthamoeba trophozoites inthe presence of EDTA
In the presence of EDTA in NMS, there was no binding ofmouse C3b to the trophozoites, resulting in a negative signal inthe IFA, while the control without EDTA showed a strong binding
Fig. 2. Immunoblot of Acanthamoeba trophozoites after incubation with NHS. (A)Binding of human C3 to trophozoites. Immunoblotting showed human C3 activationproducts of approximately 69 kDa and 40 kDa molecular weight. C3 binding was notobserved after incubating the trophozoites with PBS or heat-inactivated NHS. Similarresults were obtained from all three strains, 3ST (lane 1), 9GU (lane 2) and 11DS (lane3). (B) Binding of human C9 to trophozoites. Anti-C9-reactive bands were detectedwith molecular weights in the 69–34 kDa range after incubating the trophozoiteswith NHS. C9 binding was not detected in incubations of trophozoites with eitherPBS or heat-inactivated NHS. All three strains, 3ST (lane 1), 9GU (lane 2) and 11DS(lane 3) exhibited similar results.
of mouse C3b to the trophozoites (Fig. 4). Inhibiting effects of EDTAagainst the binding of mouse MBL-A, MBL-C and C1q to the tropho-zoites were also found. All three strains gave similar results (notshown).
Binding of mouse MBL-A, MBL-C, iC3b and C1q to Acanthamoebatrophozoites in the presence of mannose
There were no differences between trophozoite binding ofmouse MBL-A and MBL-C, and binding was weak in both, pres-ence and absence of mannose in NMS. However, the presenceof mannose in NMS had no influence on the strong binding ofmouse C3b to amoebae and the weak binding of mouse C1q totrophozoites (Fig. 5). All three strains yielded similar results (notshown).
Author's personal copy
W. Pumidonming et al. / Immunobiology 216 (2011) 225–233 229
Fig. 3. Binding of mouse MBL-A (B), MBL-C (C), iC3b (D) and C1q (F) to Acanthamoeba trophozoites of the virulent T4 (3ST) (Panel I), the non-virulent T4 (9GU) (Panel II) andthe virulent T6 (11DS) strains (Panel III). Trophozoites were incubated with mouse serum. After intensive washing bound MBL-A, MBL-C and C3b/iC3b were detected withspecific antibodies. In controls (A and E) no primary antibodies were applied. Scale bar: 100 �m.

Author's personal copy
230 W. Pumidonming et al. / Immunobiology 216 (2011) 225–233
Fig. 4. Binding of mouse MBL-A (B), MBL-C (C), C3b (D) and C1q (F) to Acanthamoebatrophozoites of the virulent T4 strain (3ST) after incubation with NMS (Panel I) andNMS plus 5 mM EDTA (Panel II). Primary antibodies were not applied in controls (Aand E). Scale bar: 100 �m.
Fig. 5. Binding of mouse MBL-A (B), MBL-C (C), C3b (D) and C1q (F) to Acanthamoebatrophozoites of the virulent T4 strain (3ST), after incubating with NMS (Panel I) andNMS plus 5 mM mannose (Panel II). Primary antibodies were not applied in controls(A and E). Scale bar: 100 �m.

Author's personal copy
W. Pumidonming et al. / Immunobiology 216 (2011) 225–233 231
Fig. 6. Binding of C3b from knockout mouse sera, MBL-A/C−/− (A), C1qa−/− (B) and Bf/C2−/− (C) to Acanthamoeba trophozoites of the virulent T4 strain (3ST). In Panel IItrophozoites were incubated with knockout mouse sera. Bound C3 was detected with an anti-C3b/iC3b/C3c antibody. In Panel I no primary antibody was applied. Scale bar:100 �m.
Binding of C3b from C1q, MBL and Bf/C2 knockout mouse sera toAcanthamoeba trophozoites
Knockout sera from mouse strains C1qa−/−, MBL-A/C−/−, andBf/C2−/− were used to further investigate which of the threepathways of mouse complement is involved in C3 activation anddeposition on Acanthamoeba trophozoites. After incubating tropho-zoites with serum from Bf/C2−/− mice, there was no C3b depositionon the trophozoites detectable, while a strong binding of anti-C3bto the trophozoites was observed after incubating the trophozoiteswith the sera of either C1qa−/− or MBL-A/C−/− mice (Fig. 6).
Discussion
Amoebicidal activity of normal human serum (NHS)
The present study demonstrates that Acanthamoeba is sus-ceptible to complement-mediated lysis in NHS. As determined
microscopically, plasma membrane disruption, extrusion of intra-cellular components and complete cell lysis were observed as afunction of time of incubation in NHS. We did not observe any sig-nificant difference between the three strains investigated, whichis in accordance with Ferrante and Rowan-Kelly (1983), who alsoinvestigated three different strains, including virulent and non-virulent strains. Toney and Marciano-Cabral (1998) demonstratedcomplement resistance in Acanthamoeba, with the most pathogenicstrain being most resistant to complement lysis. However, asthey found all three strains investigated capable of depletingcomplement activity from human serum, they assumed that thiscomplement resistance is not an inability to activate the comple-ment pathway, but rather attributed to complement inhibitorysurface proteins and/or the release of a protective extracellularmatrix. Moreover, they showed that complement resistance isdependent on growth conditions, complement susceptibility ofstrains being lower after growth in media containing hemin andserum, and that the lytic activity of individual sera differs signifi-

Author's personal copy
232 W. Pumidonming et al. / Immunobiology 216 (2011) 225–233
cantly between donors, which was the reason, why we decided touse a pool serum of 10 individual sera in our study.
Although, of course, the eye and brain – the main localisa-tions of Acanthamoeba in the human body – are immune-privilegedsites, complement components are continuously active at low lev-els in the normal human eye and brain (Bora et al. 2008; Woodruffet al. 2010). Thus, in vivo, strains causing chronic infections dohave to circumvent the action of activated complement proteinsto invade and multiply. An earlier study indicated that particu-larly pathogenic acanthamoebae might be able to block the bindingsites of human C1q, the recognition protein of the classical path-way (Walochnik et al. 2001b), however, the classical pathwaypresumably only plays a minor role in Acanthamoeba infections.In E. histolytica and N. fowleri, CD59-like proteins are present inpathogenic, but obviously absent in non-pathogenic strains (Bragaet al. 1992; Flores-Romo et al. 1994; Fritzinger et al. 2006). Theseproteins protect the amoebae from lysis by inhibiting the insertionof first binding C5B-9 protein and preventing the polymerization ofC9 in the plasma membrane (Petri et al. 2002). Several pathogensutilize host complement factor H, central regulator of the alter-native pathway controlling early activation at the C3 convertaselevel, for immune evasion (Zipfel et al. 2008). For example, hydatidcysts of Echinococcus granulosus bind to host factor H resultingin complement resistance (Díaz et al. 1997), or the cell surfaceof Borrelia strongly binds to factor H resulting in resistance tocomplement-mediated killing (Hellwage et al. 2001; Kraiczy et al.2001).
The activation of the complement system requires Ca2+ andMg2+. EDTA inhibits complement activation via the classical and thealternative pathway by chelating Ca2+ and Mg2+ (Snyderman andPike 1975). EGTA preferentially chelating Ca2+, blocks complementactivation via the classical pathway (Ca2+ and Mg2+-dependent) butnot via the alternative pathway (requires Mg2+ only). In this study,Acanthamoeba cells lysed after incubation with NHS and with NHSincluding 5 mM EGTA, but trophozoites stayed intact after incu-bation with NHS including 5 mM EDTA. Heating NHS to 56 ◦C for30 min destroyed the complement enzyme activities such as thatof factor B of the alternative pathway. As a result, acanthamoe-bae stayed intact after incubation with heat-inactivated NHS. Thesefindings suggest that Acanthamoeba are killed by a complement-dependent process activated via the alternative pathway.
Binding of human C3 and C9 to Acanthamoeba
In order to support our results of complement-mediated lysisof acanthamoebae determined microscopically, immunoblot anal-yses were performed to demonstrate the deposition of complementactivation products such as C3 and C9 on the amoebae. Cova-lent binding of C3b via internal thioester was demonstrated byrelease of C3b from the cells after incubation with hydroxylamine.Immunoblotting showed the presence of C3 bands of molecularweights of approximately 69 kDa and 41 kDa in correspondencewith other studies (Kozel and Pfrommer 1986; Kozel et al. 1987,1989). The detection of C3 fragments bound to the trophozoitesindicates that acanthamoebae activate the complement systemvia the alternative pathway. This had already been suggested byFerrante and Rowan-Kelly (1983). The detection of bound C9 afterincubating the trophozoite with NHS indicates the forming ofthe membrane attack complex (C5b6789). Membrane attack com-plexes are the result of complement activation and typically formon the surface of pathogens leading to lysis of the target cell.
In a time course experiment with incubation times from 5 min to60 min, an increase of total C9 antigen and proteolytic C9 fragmentswas observed (not shown). Thrombin has been reported to cleaveC9 into fragments of 37 kDa and 34 kDa under reducing conditions(Yoden et al. 1988). However, immunoblotting showed multi-
ple anti-C9-reactive fragments with apparent molecular weightsbetween 69 kDa and 34 kDa. These data suggest that further pro-teolytic activities are active and these enzymes could be both, ofserum or amoebal origin.
Binding and activation of mouse complement to Acanthamoebatrophozoites
In contrast to human serum, incubation of amoebae with wildtype C57BL/6 mouse serum did not result in complement-mediatedlysis. Using monospecific antibodies to various complement com-ponents and sera from complement deficient strains of mice, thebinding and activation of mouse complement by acanthamoebaewas investigated. A strong binding of mouse C3b to Acanthamoebatrophozoites was demonstrated, but there was no binding of C3bin the presence of EDTA. These findings suggest that the alternativepathway plays the major role in Acanthamoeba lysis.
Mouse MBL-A, MBL-C and C1q only weakly bound, both, in thepresence or absence of mannose and EDTA, suggesting that the MBL,trigger molecule of the lectin pathway and C1q, trigger molecule ofthe classical pathway, are not essential in the complement activa-tion by Acanthamoeba.
Knockout sera from mouse strains C1qa−/−, MBL-A/C−/−, andBf/C2−/− were used to further investigate the binding of C3b tothe trophozoites. The binding of Acanthamoeba to complement fac-tors in knockout mouse sera can distinguish between pathways ofcomplement activation. In factor B- and C2-deficient mice all threepathways are blocked (Taylor et al. 1998). The C1qa−/− mouse lacksC1q in the serum, thus the classical pathway does not function, butalternative and lectin pathways remain intact (Botto 1998). MBL-A/C−/− mouse (Shi et al. 2004) lacks the lectin pathway but hasthe alternative and the classical pathways. There was no bindingof mouse C3b to the trophozoites after incubating trophozoiteswith serum from Bf/C2−/− mice but strong binding of C3b to thetrophozoites was seen after incubating the trophozoites with seraof C1qa−/− and MBL-A/C−/− mice. These results strongly indicatethat the alternative pathway plays a major role in complementactivation by Acanthamoeba.
In conclusion our results show that NHS is amoebolytic. Themechanism by which Acanthamoeba trophozoites were destroyedby sera of healthy individuals is a complement-dependent processactivated via the alternative pathway. In contrast to NHS, normalmouse serum did not lead to lysis of Acanthamoeba trophozoites,however, binding and activation of the mouse complement systemcould be demonstrated. No differences in binding and activation ofcomplement were observed between the different Acanthamoebastrains.
Acknowledgements
This work was funded by the Austrian Exchange Office (OEAD)and by the German Research Foundation (DFG grant SFB490-D7 toF.P.).
References
Alizadeh, H., Apte, S., El-Agha, M.S., Li, L., Hurt, M., Howard, K., Cavanagh, H.D.,McCulley, J.P., Niederkorn, J.Y., 2001. Tear IgA and serum IgG antibodies againstAcanthamoeba in patients with Acanthamoeba keratitis. Cornea 20, 622–627.
Booton, G.C., Joslin, C.E., Shoff, M., Tu, E.Y., Kelly, D.J., Fuerst, P.A., 2009. Genotypicidentification of Acanthamoeba sp. isolates associated with an outbreak of Acan-thamoeba keratitis. Cornea 28, 673–676.
Bora, N.S., Jha, P., Bora, P.S., 2008. The role of complement in ocular pathology. Semin.Immunopathol. 30, 85–95.
Borsos, T., Cooper, M., 1961. On the hemolytic activity of mouse complement. Proc.Soc. Exp. Biol. 107, 227.
Botto, M., 1998. C1q knock-out mice for the study of complement deficiency inautoimmune disease. Exp. Clin. Immunogenet. 15, 231–234.

Author's personal copy
W. Pumidonming et al. / Immunobiology 216 (2011) 225–233 233
Botto, M., Dell’Agnola, C., Bygrave, A., Thompson, E.M., Cook, H.T., Petry, F., Loos,M., Pandolfi, P.P., Walport, M.J., 1998. Homozygous C1q deficiency causesglomerulonephritis associated with multiple apoptotic bodies. Nature Genet. 19,56–59.
Braga, L.L., Ninomiya, H., McCoy, J.J., Eacker, S., Wiedmer, T., Pham, C., Wood, S., Sims,P.J., Petri Jr., W.A., 1992. Inhibition of the complement membrane attack complexby the galactose-specific adhesion of Entamoeba histolytica. J. Clin. Invest. 90,1131–1137.
Brindley, N., Matin, A., Khan, N.A., 2009. Acanthamoeba castellanii: high antibodyprevalence in racially and ethnically diverse populations. Exp. Parasitol. 121,254–256.
Brown, G.C., 1943. Complementary activity of mouse-serum. J. Immunol. 46,319–323.
Calderon, J., Schreiber, R.D., 1985. Activation of the alternative and classical comple-ment pathways by Entamoeba histolytica. Infect. Immun. 50, 560–565.
Carvalho, F.R., Foronda, A.S., Mannis, M.J., Hofling-Lima, A.L., Belfort Jr., R., de Freitas,D., 2009. Twenty years of Acanthamoeba keratitis. Cornea 28, 516–519.
Cursons, R.T., Brown, T.J., Keys, E.A., Moriarty, K.M., Till, D., 1980. Immunity topathogenic free-living amoebae: role of humoral antibody. Infect. Immun. 29,401–407.
Díaz, A., Ferreira, A., Sim, R.B., 1997. Complement evasion by Echinococcus granulo-sus: sequestration of host factor H in the hydatid cyst wall. J. Immunol. 158 (8),3779–3786.
Ferrante, A., 1991. Immunity to Acanthamoeba. Rev. Infect. Dis. 13 (Suppl. 5),S403–S409.
Ferrante, A., Rowan-Kelly, B., 1983. Activation of the alternative pathway ofcomplement by Acanthamoeba culbertsoni. Clin. Exp. Immunol. 54, 477–485.
Flores-Romo, L., Tsutsumi, V., Estrada-García, T., Shibayama, M., Aubry, J.P., Bacon,K.B., Martínez-Palomo, A., 1994. CD59 (protectin) molecule, resistance to com-plement, and virulence of Entamoeba histolytica. Trans. R. Soc. Trop. Med. Hyg.88, 116–117.
Fritzinger, A.E., Toney, D.M., MacLean, R.C., Marciano-Cabral, F., 2006. Identifica-tion of a Naegleria fowleri membrane protein reactive with anti-human CD59antibody. Infect. Immun. 74, 1189–1195.
Gast, R.J., 2001. Development of an Acanthamoeba-specific reverse dot-blot and thediscovery of a new ribotype. J. Eukaryot. Microbiol. 48, 609–615.
Gast, R.J., Ledee, D.R., Fuerst, P.A., Byers, T.J., 1996. Subgenus systematics of Acan-thamoeba: four nuclear 18S rDNA sequence types. J. Eukaryot. Microbiol. 43,498–504.
Gullett, J., Mills, J., Hadley, K., Podemski, B., Pitts, L., Gelber, R., 1979. Disseminatedgranulomatous acanthamoeba infection presenting as an unusual skin lesion.Am. J. Med. 67, 891–896.
Hamelmann, C., Foerster, B., Burchard, G.D., Horstmann, R.D., 1992. Lysis ofpathogenic and nonpathogenic Entamoeba histolytica by human complement:methodological analysis. Parasite Immunol. 14, 23–35.
Hamelmann, C., Urban, B., Foerster, B., Horstmann, R.D., 1993. Complement resis-tance of pathogenic Entamoeba histolytica mediated by trypsin-sensitive surfacecomponent(s). Infect. Immun. 61, 1636–1640.
Hellwage, J., Meri, T., Heikkilä, T., Alitalo, A., Panelius, J., Lahdenne, P., Seppälä, I.J.,Meri, S., 2001. The complement regulator factor H binds to the surface proteinOspE of Borrelia burgdorferi. J. Biol. Chem. 276, 8427–8435.
Horn, M., Fritsche, T.R., Gautom, R.K., Schleifer, K.H., Wagner, M., 1999. Novel bacte-rial endosymbionts of Acanthamoeba spp. related to the Paramecium caudatumsymbiont Caedibacter caryophilus. Environ. Microbiol. 1, 357–367.
Illingworth, C.D., Cook, S.D., 1998. Acanthamoeba keratitis. Surv. Ophthalmol. 42,493–508.
Kozel, T.R., Brown, R.R., Pfrommer, G.S., 1987. Activation and binding of C3 by Candidaalbicans. Infect. Immun. 55, 1890–1894.
Kozel, T.R., Pfrommer, G.S., 1986. Activation of the complement system by Cryp-tococcus neoformans leads to binding of iC3b to the yeast. Infect. Immun. 52,1–5.
Kozel, T.R., Wilson, M.A., Farrell, T.P., Levitz, S.M., 1989. Activation of C3 and bindingto Aspergillus fumigatus conidia and hyphae. Infect. Immun. 57, 3412–3417.
Kraiczy, P., Skerka, C., Kirschfink, M., Zipfel, P.F., Brade, V., 2001. Mechanismof complement resistance of pathogenic Borrelia burgdorferi isolates. Int.Immunopharmacol. 1, 393–401.
Martinez, A.J., 1982. Acanthamoebiasis and immunosuppression. Case report. J. Neu-ropathol. Exp. Neurol. 41, 548–557.
McGhee, R.B., 1952. Presence of complement in serum of the mouse. Proc. Soc. Exp.Biol. Med. 80, 419–420.
Niyyati, M., Lorenzo-Morales, J., Rezaie, S., Rahimi, F., Mohebali, M., Maghsood, A.H.,Motevalli-Haghi, A., Martin-Navarro, C.M., Farnia, S., Valladares, B., Rezaeian,M., 2009. Genotyping of Acanthamoeba isolates from clinical and environmentalspecimens in Iran. Exp. Parasitol. 121, 242–245.
Ortiz-Ortiz, L., Capin, R., Capin, N.R., Sepulveda, B., Zamacona, G., 1978. Activationof the alternative pathway of complement by Entamoeba histolytica. Clin. Exp.Immunol. 34, 10–18.
Ortiz-Ortiz, L., Sepulveda, B., Chevez, A., 1974. Action of normal and immune humanserums on trophozoites of Entamoeba histolytica. Arch. Invest. Med. (Mex) 5(Suppl. 2), 337–342.
Petri Jr., W.A., Haque, R., Mann, B.J., 2002. The bittersweet interface of parasite andhost: lectin-carbohydrate interactions during human invasion by the parasiteEntamoeba histolytica. Annu. Rev. Microbiol. 56, 39–64.
Petry, F., Jakobi, V., Wagner, S., Tessema, T.S., Thiel, S., Loos, M., 2008. Binding andactivation of human and mouse complement by Cryptosporidium parvum (Api-complexa) and susceptibility of C1q- and MBL-deficient mice to infection. Mol.Immunol. 45, 3392–3400.
Petry, F., Reid, K.B., Loos, M., 1991. Gene expression of the A- and B-chain of mouseC1q in different tissues and the characterization of the recombinant A-chain. J.Immunol. 147, 3988–3993.
Pillemer, L., Lepow, I.H., Blum, L., 1953. The requirement for a hydrazine-sensitiveserum factor and heat-labile serum factors in the inactivation of human C’3 byzymosan. J. Immunol. 71, 339–345.
Pumidonming, W., Koehsler, M., Walochnik, J., 2010. Acanthamoeba strains showreduced temperature tolerance after long-term axenic culture. Parasitol. Res.106, 553–559.
Roestenberg, M., McCall, M., Mollnes, T.E., van Deuren, M., Sprong, T., Klasen, I.,Hermsen, C.C., Sauerwein, R.W., van der Ven, A., 2007. Complement activationin experimental human malaria infection. Trans. R. Soc. Trop. Med. Hyg. 101,643–649.
Shi, L., Takahashi, K., Dundee, J., Shahroor-Karni, S., Thiel, S., Jensenius, J.C., Gad,F., Hamblin, M.R., Sastry, K.N., Ezekowitz, R.A., 2004. Mannose-binding lectin-deficient mice are susceptible to infection with Staphylococcus aureus. J. Exp.Med. 199, 1379–1390.
Snyderman, R., Pike, M.C., 1975. Interaction of complex polysaccharides with thecomplement system: effect of calcium depletion on terminal component con-sumption. Infect. Immun. 11, 273–279.
Stapleton, F., Ozkan, J., Jalbert, I., Holden, B.A., Petsoglou, C., McClellan, K., 2009.Contact lens-related Acanthamoeba Keratitis. Optom. Vis. Sci. (Epub ahead ofprint).
Stothard, D.R., Schroeder-Diedrich, J.M., Awwad, M.H., Gast, R.J., Ledee, D.R.,Rodriguez-Zaragoza, S., Dean, C.L., Fuerst, P.A., Byers, T.J., 1998. The evolutionaryhistory of the genus Acanthamoeba and the identification of eight new 18S rRNAgene sequence types. J. Eukaryot. Microbiol. 45, 45–54.
Taylor, P.R., Nash, J.T., Theodoridis, E., Bygrave, A.E., Walport, M.J., Botto, M., 1998. Atargeted disruption of the murine complement factor B gene resulting in loss ofexpression of three genes in close proximity, factor B, C2, and D17H6S45. J. Biol.Chem. 273, 1699–1704.
Toney, D.M., Marciano-Cabral, F., 1998. Resistance of Acanthamoeba species to com-plement lysis. J. Parasitol. 84, 338–344.
Tsoni, S.V., Kerrigan, A.M., Marakalala, M.J., Srinivasan, N., Duffield, M., Taylor, P.R.,Botto, M., Steele, C., Brown, G.D., 2009. Complement C3 plays an essential rolein the control of opportunistic fungal infections. Infect. Immun. 77, 3679–3685.
Van Hamme, C., Dumont, M., Delos, M., Lachapelle, J.M., 2001. Cutaneous acan-thamoebiasis in a lung transplant patient. Ann. Dermatol. Venereol. 128,1237–1240.
Visvesvara, G.S., Balamuth, W., 1975. Comparative studies on related free-living andpathogenic amebae with special reference to Acanthamoeba. J. Protozool. 22,245–256.
Vogt, W., Dieminger, L., Lynen, R., Schmidt, G., 1974. Alternative pathway for theactivation of complement in human serum. Formation and composition of thecomplex with cobra venom factor that cleaves the third component of comple-ment. Hoppe Seylers Z. Physiol. Chem. 355, 171–183.
Walderich, B., Weber, A., Knobloch, J., 1997. Sensitivity of Entamoeba histolytica andEntamoeba dispar patient isolates to human complement. Parasite Immunol. 19,265–271.
Walochnik, J., Aichelburg, A., Assadian, O., Steuer, A., Visvesvara, G., Vetter, N.,Aspock, H., 2008. Granulomatous amoebic encephalitis caused by Acanthamoebaamoebae of genotype T2 in a human immunodeficiency virus-negative patient.J. Clin. Microbiol. 46, 338–340.
Walochnik, J., Haller-Schober, E., Kolli, H., Picher, O., Obwaller, A., Aspock, H., 2000a.Discrimination between clinically relevant and nonrelevant Acanthamoebastrains isolated from contact lens-wearing keratitis patients in Austria. J. Clin.Microbiol. 38, 3932–3936.
Walochnik, J., Obwaller, A., Aspock, H., 2000b. Correlations between morphological,molecular biological, and physiological characteristics in clinical and non-clinical isolates of Acanthamoeba spp. Appl. Environ. Microbiol. 66, 4408–4413.
Walochnik, J., Obwaller, A., Aspöck, H., 2001a. Anti-acanthamoeba IgG, IgM, and IgAimmunoreactivity of Acanthamoeba keratitis patients and asymptomatic indi-viduals. Parasitol. Res. 87, 651–656.
Walochnik, J., Obwaller, A., Haller-Schober, E.M., Aspöck, H., 2001b. Differences inimmunoreactivities and capacities to bind human complement subcomponentC1q between a pathogenic and a nonpathogenic Acanthamoeba strain. In: IXthInternational Meeting on the Biology and Pathogenicity of Free-Living AmoebaeProceedings, pp. 59–65.
Whiteman, L.Y., Marciano-Cabral, F., 1987. Susceptibility of pathogenic and non-pathogenic Naegleria spp. to complement-mediated lysis. Infect. Immun. 55,2442–2447.
Woodruff, T.M., Ager, R.R., Tenner, A.J., Noakes, P.G., Taylor, S.M., 2010. The role ofthe complement system and the activation fragment C5a in the central nervoussystem. Neuromolecular Med (Epub ahead of print).
Yoden, A., Moriyama, T., Inoue, K., Inai, S., 1988. The role of the C9b domain in thebinding of C9 molecules to EAC1-8 defined by monoclonal antibodies to C9. J.Immunol. 140, 2317–2321.
Zipfel, P.F., Hallström, T., Hammerschmidt, S., Skerka, C., 2008. The complementfitness factor H: role in human diseases and for immune escape of pathogens,like pneumococci. Vaccine 26 (Suppl. 8), 67–74.