binding of cd40l to mac-1's i-domain involves the eqlkksktl motif and mediates leukocyte...
TRANSCRIPT

Christoph Bode, Edward F. Plow, Peter Libby, Karlheinz Peter and Andreas ZirlikWillecke, Daniel Duerschmied, Constantin von zur Muhlen, Dmitry A. Soloviev, Li Zhang, Hoppe, Alexandra Ortiz Rodriguez, Nadine Herr, Ingo Hilgendorf, Peter Stachon, Florian
Blankenbach, Timoteo Marchini, Katharina Gutte, Katharina Zeschky, Nicole Bassler, Natalie Dennis Wolf, Jan-David Hohmann, Ansgar Wiedemann, Kamila Bledzka, Hermann
Thrombosis in MiceBut Does Not Affect Immunity and−−Leukocyte Recruitment and Atherosclerosis
Binding of CD40L to Mac-1's I-Domain Involves the EQLKKSKTL Motif and Mediates
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2011 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/CIRCRESAHA.111.2476842011;109:1269-1279; originally published online October 13, 2011;Circ Res.
http://circres.ahajournals.org/content/109/11/1269World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2011/10/06/CIRCRESAHA.111.247684.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Integrative Physiology
Binding of CD40L to Mac-1’s I-Domain Involves theEQLKKSKTL Motif and Mediates Leukocyte Recruitmentand Atherosclerosis—But Does Not Affect Immunity and
Thrombosis in MiceDennis Wolf, Jan-David Hohmann, Ansgar Wiedemann, Kamila Bledzka, Hermann Blankenbach,
Timoteo Marchini, Katharina Gutte, Katharina Zeschky, Nicole Bassler, Natalie Hoppe,Alexandra Ortiz Rodriguez, Nadine Herr, Ingo Hilgendorf, Peter Stachon, Florian Willecke,
Daniel Duerschmied, Constantin von zur Muhlen, Dmitry A. Soloviev, Li Zhang, Christoph Bode,Edward F. Plow, Peter Libby, Karlheinz Peter,* Andreas Zirlik*
Rationale: CD40L figures prominently in chronic inflammatory diseases such as atherosclerosis. However, sinceCD40L potently regulates immune function and hemostasis by interaction with CD40 receptor and the plateletintegrin GPIIb/IIIa, its global inhibition compromises host defense and generated thromboembolic complicationsin clinical trials. We recently reported that CD40L mediates atherogenesis independently of CD40 and proposedMac-1 as an alternate receptor.
Objective: Here, we molecularly characterized the CD40L-Mac-1 interaction and tested whether its selectiveinhibition by a small peptide modulates inflammation and atherogenesis in vivo.
Methods and Results: CD40L concentration-dependently bound to Mac-1 I-domain in solid phase bindingassays, and a high-affinity interaction was revealed by surface-plasmon-resonance analysis. We identified themotif EQLKKSKTL, an exposed loop between the �1 helix and the �-sheet B, on Mac-1 as binding site forCD40L. A linear peptide mimicking this sequence, M7, specifically inhibited the interaction of CD40L andMac-1. A cyclisized version optimized for in vivo use, cM7, decreased peritoneal inflammation and inflammatorycell recruitment in vivo. Finally, LDLr�/� mice treated with intraperitoneal injections of cM7 developed smaller,less inflamed atherosclerotic lesions featuring characteristics of stability. However, cM7 did not interfere withCD40L-CD40 binding in vitro and CD40L-GPIIb/IIIa-mediated thrombus formation in vivo.
Conclusions: We present the novel finding that CD40L binds to the EQLKKSKTL motif on Mac-1 mediatingleukocyte recruitment and atherogenesis. Specific inhibition of CD40L-Mac-1 binding may represent an attractiveanti-inflammatory treatment strategy for atherosclerosis and other inflammatory conditions, potentially avoiding theunwanted immunologic and thrombotic effects of global inhibition of CD40L. (Circ Res. 2011;109:1269-1279.)
Key Words: atherosclerosis � inflammation � CD40L � Mac-1 � peptide inhibitor
Atherosclerosis is a chronic inflammatory disease gov-erned by a network of inflammatory cytokines and
immunologic phenomena.1,2 CD40L, a member of the tumornecrosis factor superfamily first described on T cells, potentlyregulates B and T cell function via interaction with its classicreceptor CD40.3 However, expression of CD40L is notconfined to T cells but extends to a variety of cell typesincluding those typically resident in atherosclerotic plaques
such as endothelial cells (ECs), smooth muscle cells (SMCs),and macrophages. Thus, CD40L has been implicated with avariety of different inflammatory pathologies including ath-erosclerosis.4–7 Functional blockade of CD40L not onlyreduced atherosclerotic plaque formation and progression, butalso attenuated monocyte and lipid content of these lesionswhile increasing numbers of collagen fibers and SMCs,6,8,9
features associated with more stable plaques in humans.10
Original received May 2, 2011; revision received September 22, 2011; accepted September 26, 2011. In August 2011, the average time from submissionto first decision for all original research papers submitted to Circulation Research was 16 days.
From the Atherogenesis Research Group, Department of Cardiology, University of Freiburg, Freiburg, Germany (D.W., A.W., H.B., T.M., K.G., K.Z., N.H.,A.O.R., N.H., I.H. P.S., F.W., D.D., C.M., C.B., A.Z.); Baker IDI Heart and Diabetes Institute, Melbourne, Australia (D.W., J.-D.H., N.B., K.P.); ClevelandClinic, Department of Molecular Cardiology, Cleveland, Ohio (K.B., D.A.S., E.F.P.); Center for Vascular and Inflammatory Diseases, University of Maryland,Baltimore (L.Z.); Cardiovascular Medicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, Massachusetts (P.L.).
*These authors share senior authorship.Correspondence to Prof. Dr. Karlheinz Peter, Atherothrombosis and Vascular Biology, Baker IDI Heart and Diabetes Institute, P.O. Box 6492, St. Kilda
Road Central, Melbourne, Victoria 8008, Australia. E-mail [email protected]© 2011 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.111.247684
1269 at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

CD40L also augments monocyte/macrophage expression ofcollagenases implicated in plaque rupture and of tissue factor,a trigger of thrombosis following plaque rupture. Beyondthat, CD40L stabilizes thrombi through interaction with theplatelet integrin GPIIb/IIIa (�IIb�3).11 While anti-CD40Lantibody treatment generated promising results in early clin-ical trials, elevated thromboembolic complications prohibitedthe pursuit of this strategy.11,12 In addition, long-term inhibi-tion of CD40L—as is most likely required for treatment ofchronic inflammatory diseases—severely compromises hostdefenses, rendering generalized inhibition of CD40L anunappealing treatment strategy.
We previously reported the surprising finding that CD40Lpromotes atherogenesis without participation of CD40L on bonemarrow–derived cells and independently of its classic receptorCD40.8,13 These findings point toward a role of CD40L onvascular cells such as ECs or SMCs interacting with an alternatereceptor. Indeed, we suggested a novel interaction of CD40Lwith the leukocyte integrin Mac-113 promoting inflammatorycell recruitment, a crucial step in inflammation and atherogen-esis. Mac-1 (�M�2, CD11b/CD18) belongs to the �2-family ofintegrins and functions as adhesive receptor mediating adhesionand transmigration of leukocytes. Mac-1 interacts with a varietyof different ligands such as C3bi,14 ICAM,15 fibrinogen,16
fibronectin,17 heparin,18 GPIb�,19 and RAGE.20 Thus, Mac-1has been implicated with immunity, inflammation, and hemo-stasis.21–23 Inhibition of Mac-1 by neutralizing antibodies atten-uated atherosclerotic lesion formation in mice by impairingmonocyte recruitment.13 Here, we aimed to characterize theinteraction of CD40L and Mac-1 on a molecular level, to designan inhibitory peptide specifically inhibiting this interaction, andto test whether this peptide ultimately limits inflammation andatherogenesis in mice without affecting CD40L’s immunologicand thrombotic properties.
MethodsAn expanded Methods section is available in the online DataSupplement available at http://circres.ahajournals.org.
Recombinant Protein ExpressionMac-1 I-domain was produced as His-tag fusion protein by insertingthe DNA-sequence coding for the Mac-1 amino acids R115 to S340 inpET20b (Novagen, Darmstadt, Germany), subsequent purificationby Ni-NTA immobilized metal affinity chromatography (Qiagen,
Hilden, Germany), and anion-exchange chromatography usingQ-Sepharose (GE Healthcare, Sydney, Australia). CD40L was pro-duced as His- and c-myc-tag fusion protein by inserting the codingDNA for amino acids E108 to L261 in pHOG-21.
Solid Phase Binding AssayRecombinant CD40L was incubated with immobilized Mac-1I-domain in the presence or absence of blocking peptides. Binding ofsCD40L was detected by addition of antic-myc-HRP (Invitrogen,Mulgrave, Australia), TMB-substrate (Pierce, Scoresby, Australia),and colorimetric reaction. Alternatively, CD40L (Provitro, Berlin,Germany) was immobilized and binding of the recombinantMac-1 I-domain was quantified by addition of anti-His-biotin(Qiagen) and HRP-coupled streptavidin (Pierce). For binding toimmobilized peptides, CD40L was biotinylated (Micro Biotiny-lation Kit, Sigma, St. Louis, MO).
Surface Plasmon ResonanceBinding was characterized on a BIAcore 3000 (BIAcore AB, GEHealthcare) by amine coupling of CD40L or Mac-1 I-domain onto aCM5 sensor chip. Experiments were performed by passing samplesolutions over the receptor coated and reference flow cells at RTin PBS �0.1% BSA at the flow rate 20 �L/min. Surfaces wereregenerated by 30-s pulses of 5 mmol/L NaOH. Kd values werecalculated using the kinetic rate constants using the softwareprovided by the manufacturer.
Dynamic and Static Adhesion AssaysWe coated 96-well plates (Nunc, New York, NY) with sCD40L andincubated with CHO cells expressing constitutively activated Mac-1,or THP-1 cells. HEK 293-cells expressing wildtype or chimeric�2-integrins have been previously described and have been used asindicated.24 Briefly, cells were allowed to adhere for 20 to 50minutes. Cells were preincubated with blocking antibodies (10�g/mL) or peptides (50 �mol/L) as indicated. Permeabilizationbuffer (6 mg/mL phosphatase substrate (Sigma), 1% Triton X-100,50 mmol/L sodium acetate, pH 5.5) was added for colorimetricquantification. Alternatively, adhering cells were counted or stainedfor detection in fluorometer. For dynamic adhesion assays, 35-mmdishes or glass capillaries were coated with 1% BSA, CD40L, GPIb�(Abnova, Heidelberg, Germany), fibrinogen (Sigma), or ICAM-1(R&D Systems, Minneapolis, MN). Adhering and rolling cells werequantified in a parallel flow chamber system (Glycotech, Gaithers-burg, MD) at the indicated shear rates and in the presence of theindicated peptides (1 �mol/L) or antibodies (10 �g/mL).
Flow CytometryQuantification of leukocyte subsets was performed by flow cytom-etry. Binding of cM7 to CD40L-expressing Murine fibroblasts wasdetermined by quantification of FITC-coupled cM7. Binding ofCD40L to Mac-1 expressing CHO-cells or human leukocytes wasperformed by incubation with CD40L (10 �g/mL) and subsequentdetection with antipenta-His antibody (Qiagen).
Serum CytokinesPlasma concentrations of cytokines and chemokines were deter-mined by cytometric bead array (BD Biosciences, San Jose, CA).
PharmacokineticsC57BL/6J mice received intraperitoneal injections with FITC orTAMRA-labeled peptides cM7 or scM7. Fluorescence in plasmasamples was measured at indicated time points. Endothelial bindingwas visualized by intravital microscopy.
Murine PeritonitisWT or CD40L�/� mice (Jackson Laboratories, Bar Harbor, ME)received an injection of 3 mL of 4% thioglycollate broth (Sigma). Aperitoneal lavage was performed after 15 hours. Peritoneal exudatecells (PECs) were quantified after red cell lysis.
Non-standard Abbreviations and Acronyms
C3bi complement receptor type 3
CD40L CD40 ligand, CD154
EC endothelial cell
GPIb� glycoprotein Ib alpha
GPIIb/IIIa glycoprotein IIb/IIIa, integrin �IIb�3
ICAM intercellular adhesion molecule
LDLr low-density lipoprotein (LDL) receptor
Mac-1 macrophage-1 antigen, integrin �M�2, CD11b/CD18
RAGE receptor for advanced glycation endproducts
SMC sSmooth muscle cell
1270 Circulation Research November 11, 2011
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Intravital MicroscopyMice received intraperitoneal injections of 200 ng of Murine TNF�(R&D Systems) and 100 �g of peptides 5 hours before surgery. Micewere anesthetized by intraperitoneal injection of ketamine hydro-chloride (Essex) and xylazin (Bayer, Leverkusen, Germany). Thecremaster muscle was exteriorized. Videos were taken with anintravital microscope (AxioScope Vario, Carl Zeiss, Sydney, Aus-tralia) fitted with a saline immersion objective (WPlan-APOCHROMAT 20x/1,0DIC IR, Carl Zeiss). Rolling leukocyte fluxwas defined as the number of leukocytes moving at a velocity lessthan erythrocytes. Adherent leukocytes were defined as cells thatremained stationary for at least 30 s.
Atherogenesis StudyEight-week-old male LDL-receptor– deficient (LDLr�/�) mice(Jackson Laboratories) consuming a high-cholesterol diet (HCD) for20 weeks were treated with intraperitoneal injections of the peptidescM7, scM7 (100 �g), or sterile saline 3 times a week. Atheroscle-rotic lesions were analyzed histologically as described previously.8,13
In Vivo Thrombosis ModelThree- to 4-week-old C57BL/6J mice received intraperitoneal injec-tions of either sterile saline (100 �L), the peptides cM7, scM7, or theindicated antibodies. A mesenteric arteriole was chosen and injuredwith ferrichloride. Platelets were stained by retro-orbital injection ofrhodamine 3G and visualized through an intravital microscope(AxioScope Vario, Carl Zeiss). Vessel occlusion time and thrombusembolization rate was analyzed. Tail bleeding time was determinedas previously reported.11
Structural ModelingMac-1 I-domain structure was visualized using Sirius visualizationsystem 1.2 (San Diego Supercomputer Center) and a crystallographicdataset for the Mac-1 I-domain (PDB ID: 1NA5).
Statistical AnalysisData are presented as mean�SEM. Statistical testing employedStudent unpaired t test or analysis of variance (ANOVA), followedby Newman–Keuls post hoc test or Mann–Whitney U test asindicated. Probability values �0.05 were considered significant.
ResultsCD40L Concentration-Dependently Binds to theMac-1 I-DomainSince most of Mac-1’s ligands—such as fibrinogen, ICAM-1,GPIb�, RAGE, C3bi, or heparin—bind to the Mac-1 I-do-main,18–20,25,26 a stretch of �220 amino acids within the �M
subunit of the integrin, we hypothesized that the I-domain alsoserves as binding partner for CD40L. As expected, �M�2
(Mac-1) expressing HEK cells strongly adhered to CD40L whileHEK cells expressing the �L�2 integrin (LFA-1) failed tomediate cell adhesion. However, binding of HEK cells express-ing �L�2 to CD40L could be rescued when the �M I-domainreplaced the �L I-domain in the �L�2 backbone, demonstratingthat CD40L binds to Mac-1’s I-domain (Figure 1A). Similarly,recombinantly produced variants of the I-domain and CD40L(Supplemental Figure I) specifically bound to each other in solidphase binding assays (Figure 1B, Supplemental Figure IIa).Binding was enhanced in the presence of integrin-activatingMn2� as observed for most �M ligands27 (Supplemental FigureIIb). Surface plasmon resonance (SPR) analysis revealed ahigh-affinity interaction between both molecules. Kd was214�78 nmol/L and 671�272 nmol/L for binding of chip-coupled I-domain to soluble CD40L and chip-coupled CD40L to
soluble I-domain, respectively (Figure 1C,D). Interestingly, theMac-1 antibody clone 2LPM19c blocked adhesion of Mac-1-expressing CHO cells to fibrinogen only while both antibodyclones, ICRF44 and 2LPM19c, abrogated adhesion of these cellsto CD40L, suggesting that CD40L binds to the I-domain, but toa binding site distinct from that of fibrinogen (SupplementalFigure IIc, IId). Anti-CD40L treatment abrogated adhesion toCD40L but not to fibrinogen, indicating specificity of the assay.
The EQLKKSKTL Motif Within Mac-1’sI-Domain Serves as Binding Site for CD40LTo identify the binding site used by CD40L, we employed apeptide mapping strategy using a set of linear peptides,M1-M8, originating from the hydrated surface of Mac-1’sI-domain (Supplemental Table I, Figure 2A),19 as competitive
Figure 1. CD40L concentration-dependently binds to Mac-1’s I-domain. HEK293 cells expressing integrin �M�2, �L�2, orchimeric �L�2 backbones substituted with �M I-domain wereallowed to adhere to immobilized CD40L or Fibrinogen recogni-tion peptide PC-2. Cell adhesion was quantified by fluorescentstaining (A). The isolated Mac-1 (�M) I-domain concentration-dependently bound to immobilized CD40L (B). Binding was alsoquantified in SPR by passing sample solutions over chip-coupled CD40L (C), Mac-1 I-domain (D), and reference flowcells in 0.1%BSA/PBS at the flow rate of 20 �L/min (C, D). Theinsert shows recombinant, purified CD40L and I-domain on acoomassie blue-stained acrylamide gel (B). Data are presentedas mean�SEM of at least 3 independent experiments. A repre-sentative experiment is shown (C, D). n.b.�no binding.
Wolf et al CD40L-Mac-1 and Atherosclerosis 1271
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from
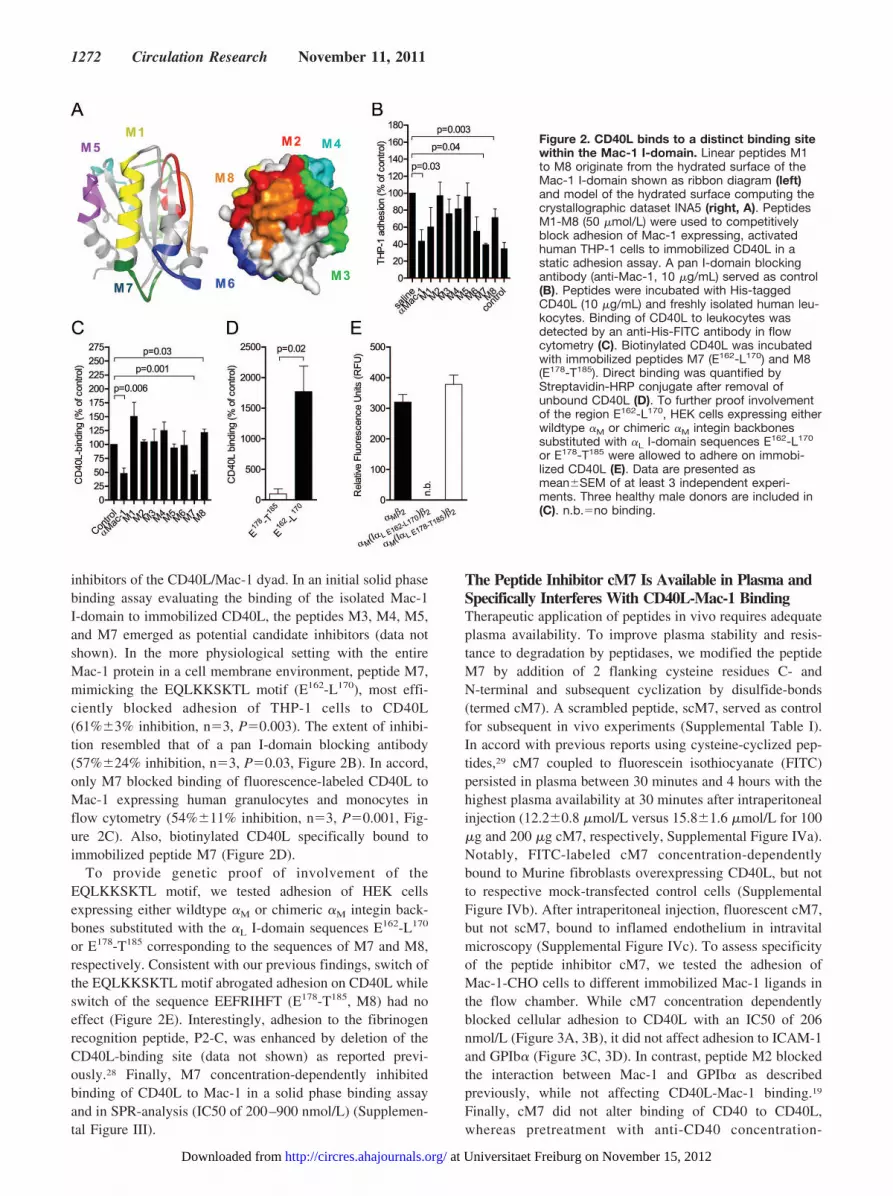
inhibitors of the CD40L/Mac-1 dyad. In an initial solid phasebinding assay evaluating the binding of the isolated Mac-1I-domain to immobilized CD40L, the peptides M3, M4, M5,and M7 emerged as potential candidate inhibitors (data notshown). In the more physiological setting with the entireMac-1 protein in a cell membrane environment, peptide M7,mimicking the EQLKKSKTL motif (E162-L170), most effi-ciently blocked adhesion of THP-1 cells to CD40L(61%�3% inhibition, n�3, P�0.003). The extent of inhibi-tion resembled that of a pan I-domain blocking antibody(57%�24% inhibition, n�3, P�0.03, Figure 2B). In accord,only M7 blocked binding of fluorescence-labeled CD40L toMac-1 expressing human granulocytes and monocytes inflow cytometry (54%�11% inhibition, n�3, P�0.001, Fig-ure 2C). Also, biotinylated CD40L specifically bound toimmobilized peptide M7 (Figure 2D).
To provide genetic proof of involvement of theEQLKKSKTL motif, we tested adhesion of HEK cellsexpressing either wildtype �M or chimeric �M integin back-bones substituted with the �L I-domain sequences E162-L170
or E178-T185 corresponding to the sequences of M7 and M8,respectively. Consistent with our previous findings, switch ofthe EQLKKSKTL motif abrogated adhesion on CD40L whileswitch of the sequence EEFRIHFT (E178-T185, M8) had noeffect (Figure 2E). Interestingly, adhesion to the fibrinogenrecognition peptide, P2-C, was enhanced by deletion of theCD40L-binding site (data not shown) as reported previ-ously.28 Finally, M7 concentration-dependently inhibitedbinding of CD40L to Mac-1 in a solid phase binding assayand in SPR-analysis (IC50 of 200–900 nmol/L) (Supplemen-tal Figure III).
The Peptide Inhibitor cM7 Is Available in Plasma andSpecifically Interferes With CD40L-Mac-1 BindingTherapeutic application of peptides in vivo requires adequateplasma availability. To improve plasma stability and resis-tance to degradation by peptidases, we modified the peptideM7 by addition of 2 flanking cysteine residues C- andN-terminal and subsequent cyclization by disulfide-bonds(termed cM7). A scrambled peptide, scM7, served as controlfor subsequent in vivo experiments (Supplemental Table I).In accord with previous reports using cysteine-cyclized pep-tides,29 cM7 coupled to fluorescein isothiocyanate (FITC)persisted in plasma between 30 minutes and 4 hours with thehighest plasma availability at 30 minutes after intraperitonealinjection (12.2�0.8 �mol/L versus 15.8�1.6 �mol/L for 100�g and 200 �g cM7, respectively, Supplemental Figure IVa).Notably, FITC-labeled cM7 concentration-dependentlybound to Murine fibroblasts overexpressing CD40L, but notto respective mock-transfected control cells (SupplementalFigure IVb). After intraperitoneal injection, fluorescent cM7,but not scM7, bound to inflamed endothelium in intravitalmicroscopy (Supplemental Figure IVc). To assess specificityof the peptide inhibitor cM7, we tested the adhesion ofMac-1-CHO cells to different immobilized Mac-1 ligands inthe flow chamber. While cM7 concentration dependentlyblocked cellular adhesion to CD40L with an IC50 of 206nmol/L (Figure 3A, 3B), it did not affect adhesion to ICAM-1and GPIb� (Figure 3C, 3D). In contrast, peptide M2 blockedthe interaction between Mac-1 and GPIb� as describedpreviously, while not affecting CD40L-Mac-1 binding.19
Finally, cM7 did not alter binding of CD40 to CD40L,whereas pretreatment with anti-CD40 concentration-
Figure 2. CD40L binds to a distinct binding sitewithin the Mac-1 I-domain. Linear peptides M1to M8 originate from the hydrated surface of theMac-1 I-domain shown as ribbon diagram (left)and model of the hydrated surface computing thecrystallographic dataset INA5 (right, A). PeptidesM1-M8 (50 �mol/L) were used to competitivelyblock adhesion of Mac-1 expressing, activatedhuman THP-1 cells to immobilized CD40L in astatic adhesion assay. A pan I-domain blockingantibody (anti-Mac-1, 10 �g/mL) served as control(B). Peptides were incubated with His-taggedCD40L (10 �g/mL) and freshly isolated human leu-kocytes. Binding of CD40L to leukocytes wasdetected by an anti-His-FITC antibody in flowcytometry (C). Biotinylated CD40L was incubatedwith immobilized peptides M7 (E162-L170) and M8(E178-T185). Direct binding was quantified byStreptavidin-HRP conjugate after removal ofunbound CD40L (D). To further proof involvementof the region E162-L170, HEK cells expressing eitherwildtype �M or chimeric �M integin backbonessubstituted with �L I-domain sequences E162-L170
or E178-T185 were allowed to adhere on immobi-lized CD40L (E). Data are presented asmean�SEM of at least 3 independent experi-ments. Three healthy male donors are included in(C). n.b.�no binding.
1272 Circulation Research November 11, 2011
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from
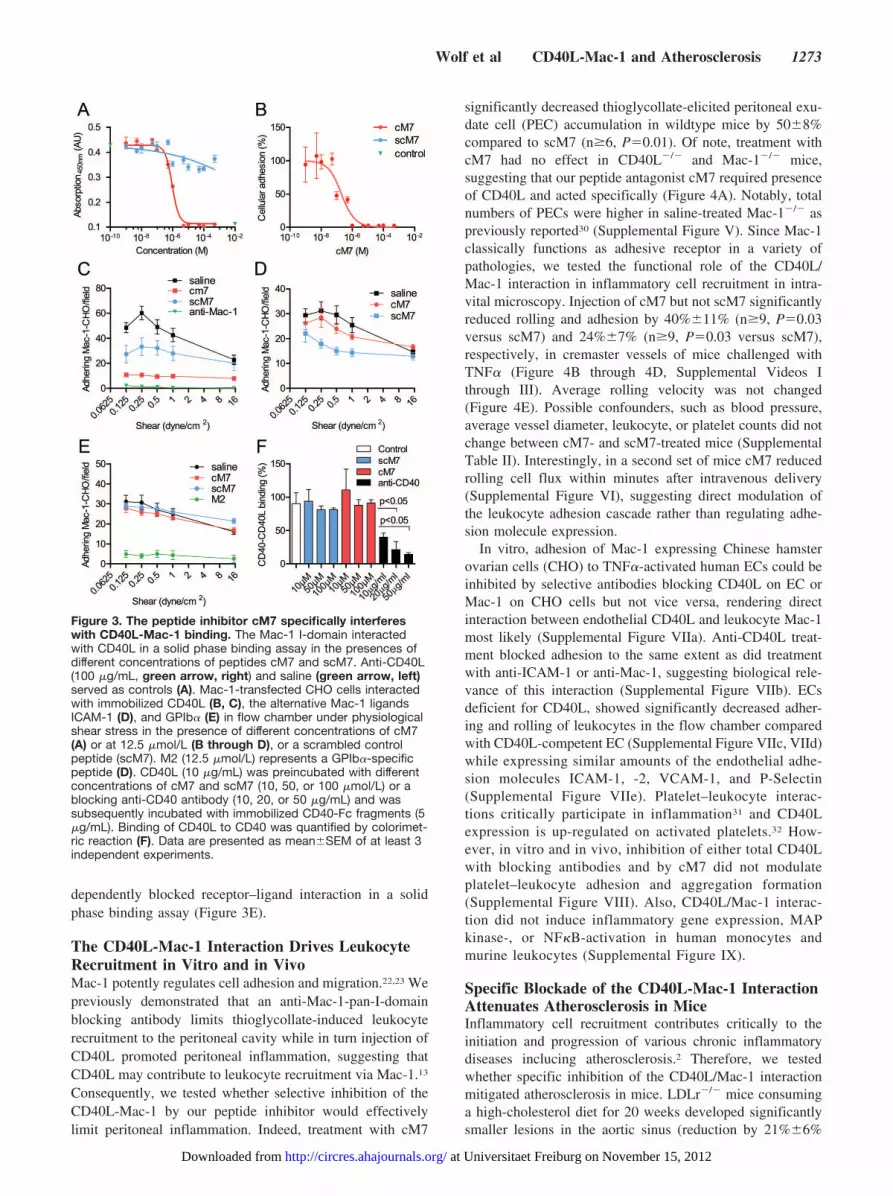
dependently blocked receptor–ligand interaction in a solidphase binding assay (Figure 3E).
The CD40L-Mac-1 Interaction Drives LeukocyteRecruitment in Vitro and in VivoMac-1 potently regulates cell adhesion and migration.22,23 Wepreviously demonstrated that an anti-Mac-1-pan-I-domainblocking antibody limits thioglycollate-induced leukocyterecruitment to the peritoneal cavity while in turn injection ofCD40L promoted peritoneal inflammation, suggesting thatCD40L may contribute to leukocyte recruitment via Mac-1.13
Consequently, we tested whether selective inhibition of theCD40L-Mac-1 by our peptide inhibitor would effectivelylimit peritoneal inflammation. Indeed, treatment with cM7
significantly decreased thioglycollate-elicited peritoneal exu-date cell (PEC) accumulation in wildtype mice by 50�8%compared to scM7 (n�6, P�0.01). Of note, treatment withcM7 had no effect in CD40L�/� and Mac-1�/� mice,suggesting that our peptide antagonist cM7 required presenceof CD40L and acted specifically (Figure 4A). Notably, totalnumbers of PECs were higher in saline-treated Mac-1�/� aspreviously reported30 (Supplemental Figure V). Since Mac-1classically functions as adhesive receptor in a variety ofpathologies, we tested the functional role of the CD40L/Mac-1 interaction in inflammatory cell recruitment in intra-vital microscopy. Injection of cM7 but not scM7 significantlyreduced rolling and adhesion by 40%�11% (n�9, P�0.03versus scM7) and 24%�7% (n�9, P�0.03 versus scM7),respectively, in cremaster vessels of mice challenged withTNF� (Figure 4B through 4D, Supplemental Videos Ithrough III). Average rolling velocity was not changed(Figure 4E). Possible confounders, such as blood pressure,average vessel diameter, leukocyte, or platelet counts did notchange between cM7- and scM7-treated mice (SupplementalTable II). Interestingly, in a second set of mice cM7 reducedrolling cell flux within minutes after intravenous delivery(Supplemental Figure VI), suggesting direct modulation ofthe leukocyte adhesion cascade rather than regulating adhe-sion molecule expression.
In vitro, adhesion of Mac-1 expressing Chinese hamsterovarian cells (CHO) to TNF�-activated human ECs could beinhibited by selective antibodies blocking CD40L on EC orMac-1 on CHO cells but not vice versa, rendering directinteraction between endothelial CD40L and leukocyte Mac-1most likely (Supplemental Figure VIIa). Anti-CD40L treat-ment blocked adhesion to the same extent as did treatmentwith anti-ICAM-1 or anti-Mac-1, suggesting biological rele-vance of this interaction (Supplemental Figure VIIb). ECsdeficient for CD40L, showed significantly decreased adher-ing and rolling of leukocytes in the flow chamber comparedwith CD40L-competent EC (Supplemental Figure VIIc, VIId)while expressing similar amounts of the endothelial adhe-sion molecules ICAM-1, -2, VCAM-1, and P-Selectin(Supplemental Figure VIIe). Platelet–leukocyte interac-tions critically participate in inflammation31 and CD40Lexpression is up-regulated on activated platelets.32 How-ever, in vitro and in vivo, inhibition of either total CD40Lwith blocking antibodies and by cM7 did not modulateplatelet–leukocyte adhesion and aggregation formation(Supplemental Figure VIII). Also, CD40L/Mac-1 interac-tion did not induce inflammatory gene expression, MAPkinase-, or NF�B-activation in human monocytes andmurine leukocytes (Supplemental Figure IX).
Specific Blockade of the CD40L-Mac-1 InteractionAttenuates Atherosclerosis in MiceInflammatory cell recruitment contributes critically to theinitiation and progression of various chronic inflammatorydiseases inclucing atherosclerosis.2 Therefore, we testedwhether specific inhibition of the CD40L/Mac-1 interactionmitigated atherosclerosis in mice. LDLr�/� mice consuminga high-cholesterol diet for 20 weeks developed significantlysmaller lesions in the aortic sinus (reduction by 21%�6%
Figure 3. The peptide inhibitor cM7 specifically interfereswith CD40L-Mac-1 binding. The Mac-1 I-domain interactedwith CD40L in a solid phase binding assay in the presences ofdifferent concentrations of peptides cM7 and scM7. Anti-CD40L(100 �g/mL, green arrow, right) and saline (green arrow, left)served as controls (A). Mac-1-transfected CHO cells interactedwith immobilized CD40L (B, C), the alternative Mac-1 ligandsICAM-1 (D), and GPIb� (E) in flow chamber under physiologicalshear stress in the presence of different concentrations of cM7(A) or at 12.5 �mol/L (B through D), or a scrambled controlpeptide (scM7). M2 (12.5 �mol/L) represents a GPIb�-specificpeptide (D). CD40L (10 �g/mL) was preincubated with differentconcentrations of cM7 and scM7 (10, 50, or 100 �mol/L) or ablocking anti-CD40 antibody (10, 20, or 50 �g/mL) and wassubsequently incubated with immobilized CD40-Fc fragments (5�g/mL). Binding of CD40L to CD40 was quantified by colorimet-ric reaction (F). Data are presented as mean�SEM of at least 3independent experiments.
Wolf et al CD40L-Mac-1 and Atherosclerosis 1273
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

versus scM7, n�11, P�0.03) and by tendency also inabdominal aortas (reduction by 30%�10% versus scM7,n�9, P�0.056) when treated with cM7 compared withscM7-treated controls (Figure 5A, 5B). Beyond a merereduction in size, atherosclerotic plaques from cM7-treatedanimals contained significantly fewer macrophages (decreaseby 37%�6% versus scM7, n�10, P�0.001, Figure 6B) andlower amounts of lipids (reduction by 32%�5% versus scM7,n�11, P�0.001, Figure 6A). In contrast, number of smooth-muscle cells (SMCs) were unchanged compared with saline-treated mice but increased by 88%�26% compared withscM7 treatment (n�11, P�0.01, Figure 6C), while infiltra-
tion of neutrophils was overall low and not changed withinthe groups (Supplemental Figure X). Collagen content in-creased in plaques of both, the treatment and the controlgroup (Figure 6D). However, plaques from cM7-treatedanimals contained more stable collagen fibers (increase by38%�9% versus scM7, n�11, P�0.014, Figure 6e, Supple-mental Figure X). Thus, features of plaques from cM7-treatedanimals resembled those of more stable plaques in humans.Importantly, lipid levels and weights remained unchangedbetween cM7 and scM7-terated mice (Table). Presence ofCD40L is critical for T-cell function, proliferation, andexpression of T-cell-specific cytokines.33–36 However, we did
Figure 4. CD40L-Mac-1 interactiondrives leukocyte recruitment in vivo.C57Bl/6 wildtype, CD40L�/�, and Mac-1�/� mice (n�6 per group) were injectedwith 3 mL 4% thioglycollate broth and 100�g of the peptides cM7, scM7, or salineas indicated. After 15 hours, numbers ofthioglycollate-elicited peritoneal exsudatecells were determined. Numbers of PECswere adjusted to percentage of treatmentwith saline (A). For intravital microscopy,C57Bl/6 mice were injected i.p. with TNF�(100 ng), peptides cM7, scM7 (100 �g), orsaline. After 5 hours, cremaster musclewas exteriorized, mounted under an intra-vital microscope (B), and leukocyte adhe-sion (C), rolling (D), and cumulative fre-quency of rolling velocity (E) werequantified. The inlay in (E) representsmean rolling velocity. Data are presentedas mean�SEM. At least 9 mice per groupwere used for B through D. Scalebar�20 �m (B).
Figure 5. Specific blockade of theCD40L-Mac-1 interaction attenuatesatherosclerosis in mice. LDLr�/� miceconsumed a high-cholesterol diet for 20weeks. Mice were injected i.p. with thespecific inhibitor (100 �g) of the CD40L-Mac-1 interaction, cM7 (n�13), the scram-bled control peptide, scM7 (n�12), orsaline (n�12), 3 times a week. Atheroscle-rotic lesion size was determined in sec-tions of the aortic root (A) or by quantifi-cation of Oil-red-O-positive staining of enface preparations of the abdominal aorta(B). Data are presented as mean�SEM.Scale bar�1000 �m (A, B).
1274 Circulation Research November 11, 2011
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

not observe changes in cytokine profiles indicative of aswitch in Th1-/Th-2 phenotype or T-cell subsets on long-termtreatment with cM7 in mice of the atherogenesis study(Supplemental Figure XI).
CD40L-Mac-1 Interaction Does Not AffectBleeding Time and Thrombus Formation in MiceHemostatic functioning of CD40L depends on interactionwith either CD40 or platelet integrin GPIIb/IIIa (�IIb�3).11,37
The inhibition of this interaction by former therapeuticstrategies employing antibodies neutralizing total CD40Lprovoked thromboembolic complications. Thus, confirmingprevious studies, treatment with an anti-CD40L blockingantibody significantly prolonged tail vein bleeding time by74%�12% (n�4, P�0.04) in our study. Interestingly, selec-tive blockade with cM7 did not modulate platelet activation(Supplemental Figure VIIIb) and bleeding time (Figure 7A),suggesting that CD40L-Mac-1 interaction is specific forCD40L’s inflammatory pathways. Accordingly, cM7 did notprolong vessel occlusion time in a model of arterial throm-bosis (Supplemental Videos IV through X), whereas anti-CD40L and anti-CD40 treatment impaired thrombus forma-tion in mesenterial arterioles resulting in a prolongation of theocclusion time by 113%�22% (n�5, P�0.005) and116%�22% (n�4, P�0.05), respectively (Figure 7B). Fur-thermore, disruption of the CD40L-Mac-1 interaction by cM7only caused a slight increase in thromboembolization rate (by67%�15%, n�5, P�0.005). However, this was a negligibleeffect compared with anti-CD40L and anti-CD40 treatmentincreasing embolization rate by 339%�38% (n�6,P�0.001), and 173%�40% (n�3, P�0.008), respectively.Interestingly, treatment with neutralizing anti-Mac-1 antibod-ies also increased the embolization rate—albeit mildly—by131%�41% (n�4, P�0.03, Figure 7C, 7D).
DiscussionWe previously made the surprising observation that CD40Lbinds to the leukocyte integrin Mac-1 and proposed this newinteraction as alternative pathway for CD40L-mediated in-flammation.13 In the present study, we characterized theCD40L-Mac-1 interaction on a molecular level and demon-strated that CD40L specifically binds to a distinct regionwithin Mac-1’s I-domain revealing a high-affinity interactioncomparable with that of CD40L’s interaction with �IIb�3.11
Our plasmon resonance data suggest a binding affinity in therange of 200 nmol/L of the �M I-domain for CD40L, whichis about 10-fold lower than the classical interaction betweenCD40L and CD40 as reported previously.38 However, since itis known that integrin activity is regulated by inflammation,it is likely that the binding affinity between CD40L andMac-1 regionally is much higher and could be furtherenhanced by additional interactions between CD40L andother regions of Mac-1 outside of the �M I-domain. Also,binding affinity of CD40L was �3-fold higher to the immo-bilized, oligomerized �M I-domain, suggesting that integrinclustering might critically strengthen binding as reportedpreviously.39
Employing a peptide-mapping strategy, we identified themotif EQLKKSKTL within Mac-1’s I-domain as a siteinvolved in binding of CD40L. Our data–that chimeric �M
integrin in which the �L I-domain sequence correspondingE162-L170 did not bind CD40L– strongly suggest that theEQLKKSKTL motif in fact represents a critical bindingsequence. However, we cannot rule out that EQLKKSKTL isonly part of the binding sequence or modulates bindingaffinity as previously observed for other Mac-1 ligands.24
Also, other segments of the integrin within or outside of thethe �M I-domain could be involved. A systematic mutationalanalysis will have to be undertaken to address these questions
Figure 6. Inhibition of the CD40L-Mac-1dyad reduces macrophage and lipidaccumulation in atherosclerotic lesions.LDLr�/� mice consumed a high-cholesteroldiet for 20 weeks. Mice were injected i.p.with the specific inhibitor of the CD40L-Mac-1 interaction, cM7 (n�13), the scram-bled control peptide, scM7 (n�12), or saline(n�12), 3 times a week. Composition of ath-erosclerotic plaques was determined bylipid-specific Oil-red-O staining (A), orimmunohistochemistry against the macro-phage antigen Mac-3 (B) and smooth-muscle cell specific �-actin (C). Collagendeposition was quantified by picrosirius-redstaining as total collagen (D) and distribu-tion of collagen fractions based on emittedcolor in polarizing light microscopy (E). Dataare presented as mean�SEM. *Indicatesprobability value �0.05.
Wolf et al CD40L-Mac-1 and Atherosclerosis 1275
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

in the future. We further demonstrated that small peptideinhibitors mimicking this sequence, M7 or cM7, its cyclisizedversion, selectively blocked binding of CD40L to Mac-1. Insolid phase binding assays and cellular adhesion assays, theIC50 ranged between 200 and 900 nmol/L. However, cM7did not affect some of the other receptor–ligand interactionsdescribed previously for both proteins. This is in accordancewith Ustinov et al, who proposed a mosaic binding modelwith distinct or overlapping regions within the I-domain aspotential binding sites.40
Interestingly, the region corresponding to M7 (E162-L170),located on an exposed loop between the �1 helix and �-sheetB in the tertiary structure of the I-domain, has not beenimplicated in binding of GPIb�, NIF, C3bi, ICAM-1, orfibrinogen to Mac-1 (Figure 2B).19,26,41–43 The peptide cM7blocked the interaction of CD40L to Mac-1 while leaving theinteractions of Mac-1 with GPIb�, ICAM-1, and fibrinogenunaffected. These data corroborate the concept that CD40Lbinds to a unique binding site on Mac-1. CD40L itself has atleast 4 different receptors, including CD40, GPIIb/IIIa(�IIb�3), Mac-1 (�M�2), and �5�1
44. Binding of CD40 toCD40L was previously mapped to CD40L’s amino residuesK143, R203, G220, Y145, and Y146.45,46 Andre et al showed that
binding of GPIIb/IIIa to CD40L involved CD40L’sKGD115–117 sequence.11 Whether binding of Mac-1 to CD40Lalso occurs at a distinct region within CD40L needs to beverified by future studies. However, our data strongly suggestthis notion: we show (1) that M7 and cM7 bind to CD40L, (2)that cM7 does not affect binding of CD40L to CD40, and (3)that cM7 does not affect bleeding time and thrombus forma-tion—CD40L’s hemostatic functions, which have beenshown to be dependent on interactions with both CD40 andGPIIb/IIIa. These findings support the notion of specificbinding sites for the CD40L-Mac-1 interaction on bothinteraction partners.
We and others previously implicated CD40L in leukocyterecruitment in inflammatory disease including atherosclerosis.6,8
In line with this concept, cM7 effectively limited recruitment ofinflammatory cells to the peritoneal cavity and the endotheliumin vivo, demonstrating that CD40L mediates inflammatory cellrecruitment by interaction with the EQLKKSKTL motif onMac-1. Mac-1 participates in inflammation as adhesion receptor,efficiently promoting both firm adhesion and slow rolling ofleukocytes.15,47 While this effect was thought to be solelydependent on the binding of Mac-1 to endothelial ICAM-1, Loet al proposed the existence of an alternative endothelial ligand
Table. Atherosclerosis Study Characteristics
Saline P * cM7 P† scM7 P‡
Weight (g)
BF 23.8�1.7 0.57 23.4�2.3 0.23 24.2�1.2 0.44
AF 36.4�3.8 0.65 35.7�3.8 0.74 35.3�2.2 0.37
Cholesterol (mg/dL)
AF 96.6�29.7 0.63 91.5�30.5 0.97 91.0�33.6 0.65
Triglycerides (mg/dL)
AF 228�97 0.18 277�107 0.20 201�190 0.63
Visceral fat pads (g)
BF 2.3�0.7 0.96 2.3�0.7 0.81 2.2�0.5 0.77
Systolic blood pressure (mm Hg)
AF 103�12 0.23 98�7 0.79 97�13 0.25
Heart rate (bpm)
AF 655�54 0.44 638�58 0.29 660�42 0.80
Leukocytes (�1000/�L)
BF 12.1�2.8 0.41 11.2�3.1 0.13 13.3�3.9 0.35
AF 5.23�1.31 0.17 4.54�1.28 0.90 4.62�1.68 0.29
Platelets (�1000/�L)
BF 557�153 0.51 529�53 0.25 562�91 0.93
AF 663�138 0.01 486�198 0.30 556�135 0.05
CD11b� (% of leukocytes)
AF 16.8�6.5 0.33 14.3�4.4 0.68 13.4�5.6 0.19
Granulocytes (% of leukocytes)
AF 13.9�4.3 0.60 13.0�3.3 0.93 13.2�4.8 0.70
Monocytes (% of leukocytes)
AF 9.8�3.6 0.06 7.2�2.1 0.50 6.4�3.1 0.03
Data are expressed as mean�SD. AF, after feeding; BF, before feeding; bpm, beats per minute.*P value saline vs cM7.†P value cM7 vs scM7.‡P value scM7 vs saline.
1276 Circulation Research November 11, 2011
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

for Mac-1.48 In accord, Lauterbach et al observed that theadhesion of neutrophils in intravital microscopy was moreseverely impaired in ICAM-1-Mac-1 double knock-out micethan in mice deficient in either of these factors alone.49 CD40Lcould be such an alternative endothelial binding partner forMac-1. Since platelet depositions occur at sites of inflamedEC,31 the interaction of leukocyte Mac-1 with platelets express-ing CD40L could also contribute to the effects observed in ourstudy. However, this is unlikely since we did not observemodulation of platelet activation or monocyte platelet aggregateformation by cM7. Interestingly, cM7 treatment also resulted inattenuated expression of TNF� and MCP-1 in a Murine cytokinechallenge model, but cM7 did not modulate inflammatorysignaling and gene expression in cell culture ex vivo. These datasuggest that cM7 does not modulate outside in signaling viaMac-1 but may rather affect inflammatory gene expression invivo via secondary mechanisms, eg, reduced leukocyterecruitment.
Since the recruitment of monocytes is considered a crucialstep in the initiation and progression of atherosclerosis,2 wetested whether specific disruption of the CD40L-Mac-1 dyadwould ultimately limit the development of atherosclerotic le-sions in vivo. Treatment with cM7 resulted in significantlysmaller, less inflamed, and potentially more stable atheroscle-rotic lesions. These findings agree with previous reports by usand others demonstrating that genetic deficiency or unselectiveinhibition of CD40L by neutralizing antibodies attenuates ath-erosclerotic lesion formation and favorably affects plaque mor-
phology.5,6,9 Beyond that, our data illustrate for the first time thatCD40L’s interaction with a distinct region on Mac-1’s I-domainmediates its proatherogenic properties. While impaired migra-tion of monocytes to atherosclerotic lesions may in fact beexplained by the anti-inflammatory, antiadhesive effects of cM7,the increased number of SMCs and stable collagen fibers afterCD40L-Mac-1 blockade needs to be further explored. SinceSMCs express CD40L,50 a cross-talk between SMCs and mac-rophages mediated by the CD40L/Mac-1 interaction mightexplain this effect.
While inflammation drives many chronic diseases includingatherosclerosis, few selective anti-inflammatory treatment op-tions currently exist. In the context of atherosclerosis, statinsallow a glimpse at the therapeutic potential of such strategies.51
Also cox-2 inhibitors exemplify the impressive therapeuticbenefits but also the difficulty in developing anti-inflammatorydrugs without side effects.52 Our peptide-based strategy mightovercome some of these limitations. Also, cM7 attenuatedinflammation and atherogenesis rather than abrogating it com-pletely. In fact, a more subtle inhibition of inflammatory activitymay be actually the more promising and fruitful strategy in thecombat of atherosclerosis where most likely long-term therapy isneeded and inflammatory regenerative pathways should stayintact. Our data add a new dimension to the understanding ofCD40L-mediated inflammation—for long thought to be solelydependent on the interaction with its classical receptor CD40.Our findings might revive the concept of a therapeutic blockadeof CD40L. Previous concepts aimed at the global inhibition of
Figure 7. CD40L-Mac-1 interaction does notmediate thrombus formation and stability inmice. C57Bl/6 wildtype mice were injected withthe peptides cM7, scM7 (100 �g), blockingantibodies against Mac-1, CD40L, CD40 (100�g), IgG isotype control (100 �g), or saline,before assessment of tail bleeding time (A) andin vivo thrombus formation (B through D) inmesenteric arterioles following injury with ferri-chloride. Thromboembolization rate wasdefined as frequency of emboli per minute (C,D). Data are presented as mean�SEM of atleast 4 animals per group. Scale bar�200 �m.
Wolf et al CD40L-Mac-1 and Atherosclerosis 1277
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

CD40L and failed due to acute or long-term side effects. Inparticular, clinical data revealed thromboembolic complicationsmost likely due to destabilization of thrombi.11 In accord, weshow that inhibition of CD40L and CD40 prolonged bleedingtime, inhibited thrombus formation, and promoted thrombusdestabilization. In contrast, specific inhibition of the CD40L-Mac-1 interaction hardly affected thrombus integrity. Moreover,cM7 did not interfere with CD40-CD40L binding in vitro anddid not induce changes in basic immunologic characteristicssuch as alteration of Th1/Th2-phenotype as previously demon-strated for CD40L-CD40-signaling.3 Taken together, these find-ings suggest the following scenario: CD40L mediates its inflam-matory actions preferably via the CD40L-Mac-1 dyad, whereasits immunologic and thrombotic features rather depend on itsinteraction with CD40 and GPIIb/IIIa.
Indeed, the use of selective strategies to block CD40L’sinteraction partners as suggested in this work might enabledevelopment of tailored drugs for different CD40L-dependentconditions. The peptide inhibitor cM7 effectively and specifi-cally inhibited CD40L-Mac-1-dependent inflammatory cell re-cruitment and atherogenesis. Therefore, selective inhibition ofthe CD40L–Mac-1 interaction as achieved by cM7 treatment inthis study may represent a fruitful novel strategy to combatchronic inflammatory diseases such as atherosclerosis.
AcknowledgmentsWe thank Daniel Topcic from the Baker IDI Heart and DiabetesInstitute Melbourne for advice on recombinant protein production,Patrick Stoll and Rene Schmidt from the University of Freiburg fortheir support with intravital microscopy. We thank Andreas Straubfrom the University of Tubingen for his assistance in intravitalmicroscopy and for proofreading of the manuscript, as well asSandra Ernst and Christian Munkel for technical support and SaraKarwacki for editorial assistance.
Sources of FundingThis work was supported by research grants from the DeutscheForschungsgemeinschaft (DFG ZI743/3-1 and 3-2) and from the Else-Kroner-Fresenius Stiftung (EKFS P30/10//A43/10) to Dr Zirlik, grantHL 34636 from the National Institutes of Health to Dr Libby, and grantsfrom the National Health and Medical Research Council of Australia toDr Peter. Dr Peter is a Future Fellow of the Australian ResearchCouncil. Dr Wolf was funded by a fellowship of the Bayer Science andEducation Foundation.
DisclosuresNone.
References1. Libby P. Inflammation in atherosclerosis. Nature. 2002;420:868–874.2. Swirski FK, Pittet MJ, Kircher MF, Aikawa E, Jaffer FA, Libby P,
Weissleder R. Monocyte accumulation in mouse atherogenesis is pro-gressive and proportional to extent of disease. Proc Natl Acad Sci U S A.2006;103:10340–10345.
3. Iezzi G, Sonderegger I, Ampenberger F, Schmitz N, Marsland BJ, KopfM. CD40-CD40L cross-talk integrates strong antigenic signals andmicrobial stimuli to induce development of IL-17-producing CD4� Tcells. Proc Natl Acad Sci U S A. 2009;106:876–881.
4. Heeschen C, Dimmeler S, Hamm CW, van den Brand MJ, Boersma E,Zeiher AM, Simoons ML. Soluble CD40 ligand in acute coronary syn-dromes. N Engl J Med. 2003;348:1104–1111.
5. Lutgens E, Gorelik L, Daemen MJ, de Muinck ED, Grewal IS, Kote-liansky VE, Flavell RA. Requirement for CD154 in the progression ofatherosclerosis. Nat Med. 1999;5:1313–1316.
6. Mach F, Schonbeck U, Sukhova GK, Atkinson E, Libby P. Reduction ofatherosclerosis in mice by inhibition of CD40 signalling. Nature. 1998;394:200–203.
7. Quezada SA, Jarvinen LZ, Lind EF, Noelle RJ. CD40/CD154 interactionsat the interface of tolerance and immunity. Annu Rev Immunol. 2004;22:307–328.
8. Bavendiek U, Zirlik A, LaClair S, MacFarlane L, Libby P, Schonbeck U.Atherogenesis in mice does not require CD40 ligand from bone marrow-derived cells. Arterioscler Thromb Vasc Biol. 2005;25:1244–1249.
9. Schonbeck U, Sukhova GK, Shimizu K, Mach F, Libby P. Inhibition ofCD40 signaling limits evolution of established atherosclerosis in mice.Proc Natl Acad Sci U S A. 2000;97:7458–7463.
10. Sukhova GK, Schonbeck U, Rabkin E, Schoen FJ, Poole AR, BillinghurstRC, Libby P. Evidence for increased collagenolysis by interstitial colla-genases-1 and -3 in vulnerable human atheromatous plaques. Circulation.1999;99:2503–2509.
11. Andre P, Prasad KS, Denis CV, He M, Papalia JM, Hynes RO, PhillipsDR, Wagner DD. CD40L stabilizes arterial thrombi by a beta3 integ-rin–dependent mechanism. Nat Med. 2002;8:247–252.
12. Kawai T, Andrews D, Colvin RB, Sachs DH, Cosimi AB. Thrombo-embolic complications after treatment with monoclonal antibody againstCD40 ligand. Nat Med. 2000;6:114.
13. Zirlik A, Maier C, Gerdes N, MacFarlane L, Soosairajah J, Bavendiek U,Ahrens I, Ernst S, Bassler N, Missiou A, Patko Z, Aikawa M, SchonbeckU, Bode C, Libby P, Peter K. CD40 ligand mediates inflammationindependently of CD40 by interaction with Mac-1. Circulation. 2007;115:1571–1580.
14. Ross GD, Lambris JD. Identification of a C3bi-specific membrane com-plement receptor that is expressed on lymphocytes, monocytes, neu-trophils, and erythrocytes. J Exp Med. 1982;155:96–110.
15. Dunne JL, Ballantyne CM, Beaudet AL, Ley K. Control of leukocyterolling velocity in TNF-alpha-induced inflammation by LFA-1 andMac-1. Blood. 2002;99:336–341.
16. Altieri DC, Agbanyo FR, Plescia J, Ginsberg MH, Edgington TS, PlowEF. A unique recognition site mediates the interaction of fibrinogen withthe leukocyte integrin Mac-1 (CD11b/CD18). J Biol Chem. 1990;265:12119–12122.
17. Kanse SM, Matz RL, Preissner KT, Peter K. Promotion of leukocyteadhesion by a novel interaction between vitronectin and the beta2 integrinMac-1 (alphaMbeta2, CD11b/CD18). Arterioscler Thromb Vasc Biol.2004;24:2251–2256.
18. Peter K, Schwarz M, Conradt C, Nordt T, Moser M, Kubler W, Bode C.Heparin inhibits ligand binding to the leukocyte integrin Mac-1 (CD11b/CD18). Circulation. 1999;100:1533–1539.
19. Ehlers R, Ustinov V, Chen Z, Zhang X, Rao R, Luscinskas FW, Lopez J,Plow E, Simon DI. Targeting platelet–leukocyte interactions: identifi-cation of the integrin Mac-1 binding site for the platelet counter receptorglycoprotein Ibalpha. J Exp Med. 2003;198:1077–1088.
20. Chavakis T, Bierhaus A, Al-Fakhri N, Schneider D, Witte S, Linn T,Nagashima M, Morser J, Arnold B, Preissner KT, Nawroth PP. Thepattern recognition receptor (RAGE) is a counterreceptor for leukocyteintegrins: a novel pathway for inflammatory cell recruitment. J Exp Med.2003;198:1507–1515.
21. Diamond MS, Staunton DE, Marlin SD, Springer TA. Binding of the integrinMac-1 (CD11b/CD18) to the third immunoglobulin-like domain of ICAM-1(CD54) and its regulation by glycosylation. Cell. 1991;65:961–971.
22. Flick MJ, Du X, Witte DP, Jirouskova M, Soloviev DA, Busuttil SJ, PlowEF, Degen JL. Leukocyte engagement of fibrin(ogen) via the integrinreceptor alphaMbeta2/Mac-1 is critical for host inflammatory response invivo. J Clin Invest. 2004;113:1596–1606.
23. Smith CW, Marlin SD, Rothlein R, Toman C, Anderson DC. Cooperativeinteractions of LFA-1 and Mac-1 with intercellular adhesion molecule-1in facilitating adherence and transendothelial migration of human neu-trophils in vitro. J Clin Invest. 1989;83:2008–2017.
24. Zhang L, Plow EF. Identification and reconstruction of the binding sitewithin alphaMbeta2 for a specific and high affinity ligand, NIF. J BiolChem. 1997;272:17558–17564.
25. Plow EF, Haas TA, Zhang L, Loftus J, Smith JW. Ligand binding tointegrins. J Biol Chem. 2000;275:21785–21788.
26. Diamond MS, Garcia-Aguilar J, Bickford JK, Corbi AL, Springer TA.The I domain is a major recognition site on the leukocyte integrin Mac-1(CD11b/CD18) for four distinct adhesion ligands. J Cell Biol. 1993;120:1031–1043.
27. Takagi J, Springer TA. Integrin activation and structural rearrangement.Immunol Rev. 2002;186:141–163.
1278 Circulation Research November 11, 2011
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

28. Zhang L, Plow EF. A discrete site modulates activation of I domains.Application to integrin alphaMbeta2. J Biol Chem. 1996;271:29953–29957.
29. Koenen RR, von Hundelshausen P, Nesmelova IV, Zernecke A, LiehnEA, Sarabi A, Kramp BK, Piccinini AM, Paludan SR, Kowalska MA,Kungl AJ, Hackeng TM, Mayo KH, Weber C. Disrupting functionalinteractions between platelet chemokines inhibits atherosclerosis in hy-perlipidemic mice. Nat Med. 2009;15:97–103.
30. Orlova VV, Choi EY, Xie C, Chavakis E, Bierhaus A, Ihanus E, Bal-lantyne CM, Gahmberg CG, Bianchi ME, Nawroth PP, Chavakis T. Anovel pathway of HMGB1-mediated inflammatory cell recruitment thatrequires Mac-1-integrin. EMBO J. 2007;26:1129–1139.
31. Gawaz M, Langer H, May AE. Platelets in inflammation and athero-genesis. J Clin Invest. 2005;115:3378–3384.
32. Henn V, Slupsky JR, Grafe M, Anagnostopoulos I, Forster R, Muller-Berghaus G, Kroczek RA. CD40 ligand on activated platelets triggers aninflammatory reaction of endothelial cells. Nature. 1998;391:591–594.
33. Guiducci C, Valzasina B, Dislich H, Colombo MP. CD40/CD40L inter-action regulates CD4�CD25� T reg homeostasis through dendritic cell-produced IL-2. Eur J Immunol. 2005;35:557–567.
34. Schoenberger SP, Toes RE, van der Voort EI, Offringa R, Melief CJ.T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40Linteractions. Nature. 1998;393:480–483.
35. Smook ML, Heeringa P, Damoiseaux JG, Daemen MJ, de Winther MP,Gijbels MJ, Beckers L, Lutgens E, Tervaert JW. Leukocyte CD40Ldeficiency affects the CD25(�) CD4 T cell population but does not affectatherosclerosis. Atherosclerosis. 2005;183:275–282.
36. Toubi E, Shoenfeld Y. The role of CD40-CD154 interactions in autoim-munity and the benefit of disrupting this pathway. Autoimmunity. 2004;37:457–464.
37. Henn V, Steinbach S, Buchner K, Presek P, Kroczek RA. The inflammatoryaction of CD40 ligand (CD154) expressed on activated human platelets istemporally limited by coexpressed CD40. Blood. 2001;98:1047–1054.
38. Khandekar SS, Silverman C, Wells-Marani J, Bacon AM, Birrell H,Brigham-Burke M, DeMarini DJ, Jonak ZL, Camilleri P, Fishman-Lobell J.Determination of carbohydrate structures N-linked to soluble CD154 andcharacterization of the interactions of CD40 with CD154 expressed in Pichiapastoris and Chinese hamster ovary cells. Protein Expr Purif. 2001;23:301–310.
39. Simon SI, Green CE. Molecular mechanics and dynamics of leukocyterecruitment during inflammation. Annu Rev Biomed Eng. 2005;7:151–185.
40. Ustinov VA, Plow EF. Delineation of the key amino acids involved inneutrophil inhibitory factor binding to the I-domain supports a mosaicmodel for the capacity of integrin alphaMbeta 2 to recognize multipleligands. J Biol Chem. 2002;277:18769–18776.
41. Rieu P, Sugimori T, Griffith DL, Arnaout MA. Solvent-accessibleresidues on the metal ion-dependent adhesion site face of integrin CR3mediate its binding to the neutrophil inhibitory factor. J Biol Chem.1996;271:15858–15861.
42. Yakubenko VP, Solovjov DA, Zhang L, Yee VC, Plow EF, Ugarova TP.Identification of the binding site for fibrinogen recognition peptidegamma 383–395 within the alpha(M)I-domain of integrin alpha(M)beta2.J Biol Chem. 2001;276:13995–14003.
43. Zhang L, Plow EF. Amino acid sequences within the alpha subunit ofintegrin alpha M beta 2 (Mac-1) critical for specific recognition of C3bi.Biochemistry. 1999;38:8064–8071.
44. Leveille C, Bouillon M, Guo W, Bolduc J, Sharif-Askari E, El-Fakhry Y,Reyes-Moreno C, Lapointe R, Merhi Y, Wilkins JA, Mourad W. CD40ligand binds to alpha5beta1 integrin and triggers cell signaling. J BiolChem. 2007;282:5143–5151.
45. Bajorath J, Chalupny NJ, Marken JS, Siadak AW, Skonier J, Gordon M,Hollenbaugh D, Noelle RJ, Ochs HD, Aruffo A. Identification of residueson CD40 and its ligand which are critical for the receptor–ligand inter-action. Biochemistry. 1995;34:1833–1844.
46. Bajorath J, Marken JS, Chalupny NJ, Spoon TL, Siadak AW, Gordon M,Noelle RJ, Hollenbaugh D, Aruffo A. Analysis of gp39/CD40 interactionsusing molecular models and site-directed mutagenesis. Biochemistry.1995;34:9884–9892.
47. Dunne JL, Collins RG, Beaudet AL, Ballantyne CM, Ley K. Mac-1, butnot LFA-1, uses intercellular adhesion molecule-1 to mediate slow leu-kocyte rolling in TNF-alpha-induced inflammation. J Immunol. 2003;171:6105–6111.
48. Lo SK, Van Seventer GA, Levin SM, Wright SD. Two leukocytereceptors (CD11a/CD18 and CD11b/CD18) mediate transient adhesion toendothelium by binding to different ligands. J Immunol. 1989;143:3325–3329.
49. Lauterbach M, O’Donnell P, Asano K, Mayadas TN. Role of TNFpriming and adhesion molecules in neutrophil recruitment to intravascularimmune complexes. J Leukoc Biol. 2008;83:1423–1430.
50. Mach F, Schonbeck U, Sukhova GK, Bourcier T, Bonnefoy JY, Pober JS,Libby P. Functional CD40 ligand is expressed on human vascular endo-thelial cells, smooth muscle cells, and macrophages: implications forCD40-CD40 ligand signaling in atherosclerosis. Proc Natl Acad SciU S A. 1997;94:1931–1936.
51. Ridker PM, Danielson E, Fonseca FA, Genest J, Gotto AM Jr, KasteleinJJ, Koenig W, Libby P, Lorenzatti AJ, Macfadyen JG, Nordestgaard BG,Shepherd J, Willerson JT, Glynn RJ. Reduction in C-reactive protein andLDL cholesterol and cardiovascular event rates after initiation of rosuv-astatin: a prospective study of the JUPITER trial. Lancet. 2009;373:1175–1182.
52. Patrono C, Rocca B. Nonsteroidal antiinflammatory drugs: past, presentand future. Pharmacol Res. 2009;59:285–289.
Novelty and Significance
What Is Known?
● CD40L, a member of the tumor necrosis factor superfamiliy, is apotent mediator of inflammation, immunity, and hemostasis.
● Therapeutic inhibition of CD40L by blocking antibodies was success-ful in attenuating inflammatory disease in clinical trials, butinhibition severely compromises immune and hemostatic function.
● Beyond its classical immune and platelet receptors CD40 andGPIIb/IIIa, CD40L also binds to the proinflammatory leukocyteintegrin Mac-1.
What New Information Does This Article Contribute?
● CD40L binds to a distinct region within the Mac-1 molecule. Apeptide mimicking this region, cM7, is capable of specificallyblocking binding of CD40L to Mac-1.
● The peptide inhibitor cM7 attenuated leukocyte recruitment, inflam-mation, and atherogenesis in mice, but did not interfere withCD40L’s immune and hemostatic functions.
● Specific inhibition of CD40L’s interaction with inflammatory partnersmight represent a selective, anti-inflammatory treatment strategy.
Atherosclerosis is an inflammatory disease involving a plethoraof proinflammatory mediators. Previous reports establishedCD40L as marker and mediator of atherosclerotic disease.However, global inhibition of CD40L generated unfavorable sideeffects in clinical trials. Here, we present a strategy to selec-tively inhibit proatherogenic function of CD40L in several murinemodels. Given the importance of inflammatory pathways inatherosclerosis, the selective inhibition of the CD40L-Mac-1interaction as characterized in our study may be a fruitfulstrategy to combat chronic inflammatory diseases such asatherosclerosis.
Wolf et al CD40L-Mac-1 and Atherosclerosis 1279
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-1-
SUPPLEMENTAL MATERIAL TO
Binding of CD40L to Mac-1’s I-domain involves the EQLKKSKTL motif and mediates leukocyte recruitment and atherosclerosis – but does not affect immunity and
thrombosis in mice
Dennis Wolf, MD1,2, Jan-David Hohmann, BS2, Ansgar Wiedemann, BS1, Kamila Bledzka, PhD3, Hermann Blankenbach, BS1, Timoteo Marchini, BS1, Katharina Gutte, BS1, Katharina
Zeschky, BS1, Nicole Bassler, BS2, Natalie Hoppe, BS1, Alexandra Ortiz Rodriguez, MS1, Nadine Herr, BS1, Ingo Hilgendorf, MD1, Peter Stachon, MD1, Florian Willecke, MD1, Daniel
Dürschmied, MD1, Constantin von zur Mühlen, MD1, Dmitry A. Soloviev, PhD3, Li Zhang, PhD4, Christoph Bode, MD1, Edward F. Plow, PhD3, Peter Libby, MD5, Karlheinz Peter, MD2,
and Andreas Zirlik, MD1
EXPANDED MATERIALS AND METHODS Antibodies and peptides. Epitope-specific antibodies were purchased as follows: anti-human CD11b, clone 2LPM19c (Santa Cruz Biotechnology); anti-human CD11b, clone ICRF44 (Biolegend); anti-human CD11a, clone HI111 (Biolegend); anti-human CD40L, clone 24-31 (Calbiochem); anti-human CD40L, clone 40804 (R&D systems); anti-human ICAM-1, clone BBIG-I1 (R&D systems). Peptides were synthesized from Peptide Specialty Laboratories (Heidelberg), purified by HPLC, and cyclisized, if applicable. Molecular mass was determined by mass spectrometry. Peptides had a purity >95%. Cell culture. Human umbilical vein endothelial cells (HUVECs) were purchased from Lonza and grown in M199, 20 % fetal calf serum (FCS), 1% Penicillin/Streptomycin, 0,1 % Fungizone, 1 % non-essential amino acids (NEAA), 1 % Na-Pyruvat, 1 % Heparin, 1 % ECGS. Monocytic THP-1 were cultured in RPMI 1640, 1 % Penicillin/Streptomycin, 10 % FCS, 0,05 mM 2-Mercaptoethanol. CHO cells expressing constitutively activated Mac-1 have been described previously1 and were cultured in DMEM, 1 % Penicillin/Streptomycin, 10 % FCS, 1 % NEAA, 1 % L-Glutamin, 125 μg/ml Zeocin, 70 μg/ml Geniticin. CD40L- and mock-transfected murine fibroblasts were a gift from Dr. K. Zirlik (University of Freiburg, Department for Hematology, Freiburg, Germany) and were cultured in DMEM, 1 % Penicillin/Streptomycin, 10 % FCS, 1 % NEAA, 1% L-glutamin, 125 μg/ml. HEK cells expressing chimeric β2-integrins have previously been described2. Isolation of murine endothelial cells. For isolation of murine endothelial cells corresponding wildtype or CD40L-/- mice (all C57BL/6J) were euthanized with CO2, and lungs, heart, brain, and liver were harvested employing sterile techniques, minced with a razor blade, and digested in 0.2 % collagenase type-1/1% BSA (Worthington, Lakewood, NJ and Sigma, St. Louis, MO) for 90 min at 37 °C. After washing with 0.1 % BSA and filtering through a 70 μm nylon mesh, cells were resuspended in 0.1 % BSA and incubated with an anti-mouse CD31 antibody conjugated to sheep anti-rat Dynabeads (Dynal Biotech, Oslo, Norway) for 10 min at room temperature. Cells were then separated and washed three times using a magnetic particle concentrator (Dynal Biotech) and seeded into gelatin-coated plates. After they reached confluence, a second magnetic sorting was performed with a rat anti-mouse ICAM-2 antibody (BD Pharmingen). Cells were grown in DMEM high glucose supplemented with 20 % fetal bovine serum (FBS), 1 % sodium pyruvate, 1 % heparin, 1 % bovine endothelial growth factor, 0.6 % NEAA, and 1 % penicillin/streptomycin. Cells were maintained in M-199 supplemented with 0.1 % FBS 24 h prior to experiments.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-2-
Recombinant protein expression. The Mac-1 I-domain was produced as fusion protein containing an C-terminal His-tag by inserting the DNA-sequence coding for the Mac-1 amino acids R115 to S340 in the expression vector pET20b (Novagen) by a PCR-based strategy using the following primers: 5’-AGAAGTTCCCAGAGGCCCT-3’ and 5’-GAGTGCGGCCGCGGCAGCGCTGAAGCCTTCCTG-3’. A CHO cell line constitutively expressing the entire human Mac-1 α-chain served as template. The resulting PCR-fragment was cloned in pGEMT (Promega), released by NcoI and NotI (New England Biolabs) and inserted into the NcoI-NotI-linearized pET20b. This expression vector was transformed in BL-21 DE Star (Invitrogen) and expressed by addition of 0.5 mM IPTG (Sigma). The protein was extracted by BugBuster lysis (Novagen) and subsequently purified by Ni-NTA immobilized metal affinity chromatography (Qiagen) in a standard FPLC-system (GE Healthcare). After elution of the target protein by 250 nM imidiazol (Sigma) the fraction containing the Mac-1 I-domain (∼28 kDa) was dialyzed against 20 mM Tris-Cl, 20 mM NaCl, pH 8.0 and further purified by anion-exchange chromatography on a Q-Sepharose-columns (GE Healthcare). CD40L was produced as fusion protein containing a N-terminal His- and c-myc-tag, as well as a trimerization domain as described previously3. The coding DNA sequence for amino acids E108 to L26 were amplified by PCR using the following primers: 5’-CCTAGGCGGCCGCTATCAGAGTTTGAGTAAGCCAAAGGAC-3’ and 5’- CTTCTAGA AAACAGCTTTGAAATGCAAAAAGA-3’. A cDNA clone coding for the human CD40L (Origene) served as template. The His- and c-myc-tag were amplified by the following primers: 5’-CCGGCCATGGCCGAACAAAAGCTGATCTCAGAAGAAG-3’ and 5’-TGAG GTACCTAGGTGATGGTGATGGTGATGTGAG-3’. As template for the trimerization domain served the primer 5’- ATGAAACAGATTGAAGATAAAATTGAAGAAATTCTG AGCAAAATTTATCATATTGAAAACGAAATTGCGCGTATTAAAAAACTGATTGGAGAA-3’. All PCR fragments were cloned into pGEMT and released by NcoI, KpnI (His- and c-myc-Tag), KpnI and XbaI (trimerization motif) and XbaI and NotI (CD40L). Fragments were subsequently cloned into the expression vector pHOG-214 and transformed into TG-1 bacteria (Promega). CD40L was expressed after induction with 1 mM IPTG. Proteins were extracted as insoluble inclusion bodies, solubilized in 7 M Urea, 100 mM NaH2PO4, 100 mM Tris-Cl, pH 8.0 and purified under denaturing conditions by Ni-NTA immobilized metal affinity chromatography. CD40L was refolded by dialysis against decreasing Urea-concentrations. Both proteins were finally dialyzed against PBS and stored at -80°C until further use. The purity of both recombinant proteins was >90% as assessed by SDS gel electrophoresis. Solid phase binding assay. The recombinant Mac-1 I-domain was immobilized in 96-well plates (Nunc) in PBS at 4°C overnight. After blocking in 2 % BSA/PBS and subsequent washing with PBS, recombinant CD40L was added to the wells in the indicated concentrations and incubated for 2 hours at 37 °C. Effect of the peptides M1-M8 was assessed by incubating CD40L (10 μg/ml) in the presence of peptides (50 μM). After removing of unbound CD40L by washing with 0.1 % Tween-20/PBS, anti-c-myc-HRP (Invitrogen) was added and incubated for 2 hours at room temperature. Binding was quantified by addition of TMB-substrate (Pierce), colorimetric reaction at 450 nm. To determine the IC50 of cM7, I-domain was immobilized (25μg/ml) in a 96-well plate (Nunc) in NaHCO3/Na2CO3 Coating Buffer for 2 hours, blocked in 1% BSA for 1 hour, then CD40L (5μg/ml) and cM7/scM7 was added at the desired concentrations. Wells were washed with 0.1 % Tween-20/PBS and incubated with anti-c-myc-HRP (Invitrogen) for 2 hours at 37oC. Binding was quantified by addition of TMB-substrate (Pierce), colorimetric reaction at 450 nm. Alternatively, CD40L without a His-tag (Provitro) was immobilized and blocked as described above. Binding of the recombinant Mac-1 I-domain was quantified by addition of anti-His-Biotin monoclonal antibody (Qiagen), HRP-coupled streptavidin (Pierce) and colorimetric reaction at 450 nm. For the specific binding of the Mac-1 I-domain BSA-coated wells were subtracted from the CD40L-coated. Kd was estimated using a one-site binding hyperbola nonlinear regression model with the Software Prism (Graphpad). For quantification of the binding of CD40L to peptides, peptides were immobilized in 96-well plates overnight at 4 °C in 50 mM sodium carbonate, pH 10.6. CD40L was biotinylated using the Micro-Biotinylation-
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-3-
Kit (Sigma) following the manufacturer’s instructions and detected by HRP-coupled streptavidin (Pierce) and colorimetric reaction. A mixture of equal molarities of all peptides served as positive control. Absorbance on BSA-coated wells served as negative control and was subtracted. Alternatively, Fisher 96-wells TC treated plate was coated overnight at +4ºC with 100 μl of 0.1 μM CD40L (Genomic Insight) and postcoated with 250 μl of 0.5% polyvinylpyrrolidone (PVP). The plates were washed and different dilutions of cM7, csM7 and cM8 were added in 100 μl of HBSS/HEPES, pH 7.4, containing 5 mM CaCl2 and 5 mM MgCl2, each concentration point in triplicate. The plate was incubated for 15 min at 37ºC then 10 μl of 0.1 μM FITC-labeled αM I-domain were added and the plate was incubated for additional 15 min. After incubation the well were two times washed with HEBSS/HEPES, pH 7.4, 5 mM Ca/Mg, and fluorescence was measured with Fluorescence Multy-well reader CytoFluor II (Applied Biotechnologies). Binding of FITC-I-domain to CD40L in the absence of peptides was used as control for 100% binding, and the binding to wells coated with PVP only was used as control for non-specific background binding (0%). Plasmon Surface Resonace. Binding was measured on a BIAcore 3000 (BIAcore AB). Recombinant CD40L or recombinant Mac-1 I-domain was immobilized onto the CM5 sensor chip via amine coupling using the application wizard for surface preparation according to the manufacturer's protocol. Experiments were performed by passing sample solutions over the receptor coated and reference flow cells at RT in PBS +0.1 % BSA at the flow rate 20 μl/min. Surfaces were regenerated by 30-s pulses of 5mM NaOH. In inhibition assays CD40L or a mixture of CD40L and the peptide (M7 or sM7) at a 1:100 molar ratio was passed over immobilized αM I-domain. To reduce nonspecific binding 0.1% BSA was included in the running buffer. Data transformation was performed and overlay plots were prepared using the BIAevaluation software. Kinetic data were analyzed in terms of the Langmuir 1:1 interaction model. Kd values were calculated using the kinetic rate constants using the software provided by the manufacturer. The calculated KD values are means± S.D. of three independent experiments. Static adhesion assays. 96-well plates (Nunc) were coated with sCD40L (10 μg/ml) in PBS overnight at 4 °C. After removal of unbound CD40L by washing with PBS, plates were blocked with 0.1 % agarose for 1 hour at room temperature and washed with PBS. Blocking antibodies against CD40L (10 μg/ml) were given to the w ells as indicated and incubated for 15min at room temperature, followed by subsequent washing with PBS. CHO cells expressing constitutively activated Mac-15 or THP-1 cells were pre-incubated with function blocking antibodies against CD11b or CD11a (10 µg/ml) for 15 min at room temperature. 5 x 104 cells/well were allowed to adhere for 20 to 50 min at 37 °C. As indicated, static adhesion assays were carried out in the presence of peptides at a concentration of 50 µM. After removal ob unbound cells by washing with PBS, permeabilization buffer (6 mg/ml phosphatase substrate (Sigma), 1 % Triton X-100, 50 mM sodium acetate, pH 5.5) was added for 1 hour at 37 °C and adhering cells were quantified by colorimetric reaction at 405 nm. Alternatively, adhering cells were counted under the microscope (Zeiss). Alternatively, human umbilical vein endothelial cells (HUVECs) were stimulated with 50 ng/ml TNF-α prior to the experiment. Mac-1 expressing CHO were loaded with carboxyfluorescein diacetate succinimidyl ester (CFDA, Invitrogen) according to the manufacturer’s protocol. HUVECs or CHO-cells were selectively incubated with blocking antibodies (10 µg/ml) as indicated, washed and cells were allowed to adhere on HUVECs for 35 min at 37°C. After removal of unbound cells by washing with PBS adhering cells were counted under the fluorescence microscope. For cell adhesion of chimeric integrin expressing HEK cells to CD40L, 48-well Costar TC treated plates (Corning) were coated overnight at 4°C with 200 ml of 1, 5 or 10 μg/ml CD40L, each concentration point is in triplicate and then post-coated with 0.5% polyvinylpyrrolidone (PVP) for 1 h at room temperature. Control wells (background adhesion to plastic) were coated with PVP only. Background adhesion was determined on the same plate for each of the cell type. Prior to use, the plates were rinsed three times with PBS. HEK 293 transfectant cells were harvested using Invitrogen HBSS-based enzyme free Cell
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-4-
Dissociation Buffer, washed three times with 50 ml HBSS/HEPES, pH 7.4, and resuspended in the HBSS/HEPES, containing 2 mM CaCl2, 2 mM MgCl2 and 5% BSA. The cells were then seeded at 105 cells/well onto the assay plates and incubated at 37°C, 5% CO2 for 30 min. To determine the extent of adhesion/attachment, the plates were washed three times with PBS, and the number of adherent cells in each well was quantified using the Cyquant Cell Proliferation Assay Kit (Molecular Probes) according to the manufacturer's instructions. Data are presented as Relative Fluorescence Units (RFU) and represent the results of single independent experiment in triplicate. Laminar flow chamber assay. For dynamic adhesion assays, 35 mm dishes or glass capillaries were coated overnight at 4 °C with 1 % BSA, CD40L, GPIbα (Abnova), ICAM-1 (R&D systems) or fibrinogen (Sigma), at a concentration of 10 µg/ml, and 30 µg/ml, respectively. Adhesion and rolling of Mac-1 expressing CHO-cells was tested in a parallel flow chamber system (Glycotech) using increasing flow rates from 0.5 dyne/cm² (venous flow) up to 15 dyne/cm² (arterial flow) or as indicated otherwise. Cells were quantified under the microscope (Olympus). As indicated, effects of inhibitors were tested at the indicated shear rates and in the presence of the indicated peptides (1 µM) or antibodies (10 µg/ml). Alternatively, murine endothelial cells were isolated and TNF-α stimulated as described above. Adhesion and rolling of peritoneal exudate cells on isolated murine endothelial cells was quantified as described above. Rolling velocity was computed employing Image Pro cell tracking tool (Media Cybernetics) Flow cytometry. Flow cytometric analyses, as well as platelet activation assays and quantification of leukocyte-platelet aggregates were performed as previously described6, 7. Briefly, murine blood samples were taken by intracardiac puncture. Red cells were lyzed in 155 mM NH4Cl, 5.7 mM K2HPO4, 0.1 mM EDTA, pH 7.3. Leukocytes were resuspendet in 0.1 % BSA/PBS and Fc-Receptors were blocked by anti-CD16/CD32 antibodies (Ebioscience). Antibodies for epitope specific fluorescence-activated cell sorting (FACS Calibur, BD) included anti-CD11b, anti-CD115, anti-Gr-1, anti-CD4, anti-CD8, anti-CD20, anti-CD41, anti-CD62P, anti-CD54, anti-CD102, and anti-CD106 (all from Ebioscience). Binding of cM7 to CD40L-or mock-transfected murine fibroblasts was determined by incubation of FITC-cM7 at the indicated concentrations with cells for 30 min at 37 °C and subsequent quantification of the fluorescence in the FL-1 channel. Binding of CD40L to Mac-1 expressing CHO-cells or human leukocytes was performed by incubation of the with the His-tag-CD40L fusion protein (10 µg/ml) for 30 min at 37 °C in PBS +Ca2+/Mg2+ and subsequent detection with Alexa488-labeled anti-PentaHis (Qiagen). Human monocytes and granulocytes were gated based on their properties in the forward- and sideward scatter. For the analysis of the endothelial expression of adhesion molecules, cells were TNF-α stimulated for 24 hours, detached using accutase (Sigma) and incubated with fluorochrome-coupled antibodies. Cytokine challenge. 8 weeks old C57BL/6J mice received an intraperitoneal injection of 200 ng of murine TNF-α (R&D systems) and 100 µg either of the peptides cM7, scM7 or an equal volume of sterile saline. After 5 hours mice were euthanized with CO2. The peritoneal cavity was flushed with 2 ml PBS and supernatant was screened for cytokines. Blood was collected by an intracardial puncture. Plasma concentrations of IL-6, IL-10, IL-12p70, TNF-α, IFN-γ, MCP-1, KC, and RANTES were determined by the Cytometric Bead Array (CBA, BD Biosciences) according to the manufacturer’s instructions. Activation of peripheral leukocytes and platelets was assessed by flow cytometry as described above. Signaling studies. Murine splenocytes or human monocytes cells were isolated as previously described8. After overnight starving, cells were stimulated with CD40L (10 µg/ml) for 24 h in the presence of peptides (10µM) or blocking antibodies against Mac-1 (10 µg/ml). Cells were lysed, proteins were separated by SDS-PAGE and blotted to polyvinylidene difluoride membranes. Primary antibodies for western blot included anti-total NFκB or anti-
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-5-
phopho NFκB (Cell Signaling). Alternatively, samples were tested with cytometric bead array for phosphor ERK1/2, JNK1/2 or p38. Pharmacokinetics. C57BL/6J mice were injected intraperitoneal with FITC-labeled cM7. Fluorescence in Plasma samples was measured at the indicated time points in Fluorescence Plate Reader (Spectramax). CM7-FITC diluted in plasma samples served as standard. Murine Peritonitis. WT, CD40L-/- or Mac-1-/- mice (Jackson Laboratories) received an injection of 3ml of 4 % thioglycollate broth (Sigma). A peritoneal lavage was performed after 15 hours by flushing the peritoneal cavity with PBS. Peritoneal exudate cells (PECs) were quantified after red cell lysis. Intravital microscopy. Mice received an intraperitoneal injection of 200ng of murine TNFα (R&D systems) and 100μg of peptides dissolved in sterile saline 5 hours before surgery. Mice were anesthetized with an intraperitoneal injection of ketamine hydrochloride (Essex) and xylazin (Bayer) at a dose at 187.5mg/kg of body weight and 62.5mg/kg of body weight, respectively. The cremaster muscle was exteriorized as described previously9, 10. For some experiments a catheter was placed in the jugular vein and peptides were administered during microscopy. The cremaster was superfused with thermo-controlled (36°C) saline. Mice were placed on a heating pad to maintain body temperature. Videos were taken with an intravital microscope (AxioScope Vario, Carl Zeiss) fitted with a saline immersion objective (WPlan-APOCHROMAT 20x/1,0DIC IR, Carl Zeiss) and a high sensitivity camera system (AxioCam MRm, Carl Zeiss) for 30 seconds each. Rolling leukocyte flux was defined as the number of leukocytes moving at a velocity less than erythrocytes. Leukocyte rolling velocity was measured by the average time required for leukocytes to roll over a defined length of the venule at each time point. Adherent leukocytes were defined as cells that remained stationary for at least 30s. Rolling leukocyte flux, adhering flux were quantified by blinded investigators. Atherogenesis study. Eight-week-old male LDL-receptor–deficient (LDLr-/-) mice (Jackson Laboratories) consuming high-cholesterol diet (HCD) were treated with intraperitoneal injections of the peptides cM7, scM7 (Peptide Specialty Laboratory) in a dose of 100μg, or sterile saline three times a week. After 20 weeks blood samples were taken for flow cytometric analysis of leukocyte subpopulations, cholesterol, and triglyceride plasma levels, as well as for the determination of plasma cytokines and chemokines. Blood pressure was determined by a non-invasive blood pressure measurement (NIBP, Harvard Apparatus). Mice were euthanized and aortic roots and arches were frozen in OCT (OCT compound; Tissue-Tek). Thoracic and abdominal aortas were fixed in 10% buffered formalin. Serial cryostat sections (6μm) of mouse aortic tissues were fixed in acetone and air-dried. Non-specific binding was blocked with 5% species-appropriate normal serum (Vector Laboratories). Sections were then incubated with primary antibodies diluted in phosphate-buffered saline, supplemented with 5 % species-appropriate normal serum. Incubation with secondary antibodies was followed by avidin-biotin complex (ABC, Vector Laboratories). Antibody binding was visualized with 3-amino-9-ethylcarbazole (AEC, Dako), followed by counterstaining with Gill’s hematoxylin solution (Sigma-Aldrich). Control stainings included staining with the respective IgG isotypes (Pharmingen, Dako). Antibodies used were rat anti-mouse Mac-3 for macrophage specific staining, anti α-actin for smooth muscle cell-specific staining (Dako). For the visualization of Type I Collagen, formalin-fixed frozen sections were incubated for 4 hours in a freshly prepared 0.1% solution of picrosirius red (Polysciences) in saturated aqueous picric acid. After rinsing in 0.01 N HCl and distilled water, sections were dehydrated in 70% ethanol and mounted in Permount (Vector Laboratories). Picrosirius red staining was analyzed by polarization microscopy. As the color of collagen fibers assessed in the picrosirius red staining depends on the thickness of collagen fibers and changes from green (thin fibers) to yellow, orange, and red (thick fibers)11, 12, color distribution in stained collagen sections was quantified. Deposition of lipids was determined by Oil red O staining after formalin fixation in aortic sections or in en face preparations of the abdominal aorta. To
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-6-
quantify the composition of the aortic lesions, sections of the aortic tissue were analyzed microscopically in all mice. Within the aortic root, lesion areas were analyzed in cross-sections obtained at the level of all 3 leaflets of the aortic valve, immediately proximal to the right coronary artery ostium. The total aortic wall area, lesion area in the aortic root, and the percentage of area stained for macrophages, lipids, SMCs, or collagen were determined via computer-assisted image quantification (ImagePro, Media Cybernetics). In vivo thrombosis model. 3-4 weeks old C57BL/6J mice received an intraperitoneal injection of either sterile saline (100µl), the peptides cM7, scM7, or the indicated antibodies. A mesenteric arteriole was chosen and injured with ferrichloride. Platelets were stained by retroorbital injection of rhodamine 3G and visualized through an intravital microscope (AxioScope Vario, Carl Zeiss). Vessel occlusion time was recorded and thrombus embolization rate analyzed at different time points and expressed as mean embolization rate/min. Tail bleeding time was determined as previously reported13. Structural modeling. Mac-1 I-domain structure was visualized using Sirius visualization system 1.2 (San Diego Supercomputer Center) and a crystallographic dataset for the Mac-1 I-domain (PDB ID: 1NA5). Statistical analysis. Data are presented as mean±SEM. Significance was measured with the unpaired Student’s t test or analysis of variance (ANOVA), followed by Newman-Keuls post hoc test or Mann-Whitney-U test as indicated. P values < 0.05 were considered significant.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from
Supplemental Material CD40L-Mac-1 and atherosclerosis
-7-
SUPPLEMENTAL FIGURES
a b
c M W1-3 E1-3 FT
M FT W E1-3
M Elution FractionsNaCl
37 kDa-
50 kDa-
25 kDa-
20 kDa-
37 kDa-
50 kDa-
25 kDa-20 kDa-
37 kDa-50 kDa-
25 kDa-20 kDa-
15 kDa-
Supplemental Figure I. Bacterial expression of recombinant variants of the Mac-1 I-domain and CD40L. A protein fragment composed of the amino residues R115 to S340, coding for the αM I-domain, was produced as soluble His-tag fusion protein (∼28kDa) in a bacterial expression system and purified by immobilized metal affinity chromatography (IMAC) (a). Contaminating bacterial proteins were further removed by anion-exchange chromatography and increasing concentrations of sodium chloride (NaCl). Elution fractions containing the isolated I-domain as assessed by Coomassie staining were pooled and dialyzed against PBS (b). The TNF homologous region of human CD40L (E108 to L261) was produced as c-myc- and His-tag-fusion protein. The protein (∼19kDa) was extracted from insoluble inclusion bodies, purified by IMAC and refolded by subsequent dialysis against PBS (c). The purity of both protein preparations was >95% as assessed by SDS-polyacrylamide gel. (W) washing fractions, (FT) column flow through, (E) elution fractions, (M) protein size marker.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-8-
a b
c d
0
100
200
300
400 +Ca2+
+Mn2+
CD40L Fib. ICAM-1
p<0.01 p=0.02 p=0.01
Bind
ing
(%)
BSA
I-dom
ain BSA
I-dom
ain0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7 12.5μg/ml 50μg/ml
p<0.001p<0.001
OD
450
Contro
l
4080
424
-31
2LPM19
c
ICRF44
HI111
0
20
40
60
80
100
120
140
160
180
p<0.001
p=0.02
p<0.001Ad
herin
g ce
lls (%
of c
ontro
l)
Contro
l
4080
424
-31
2LPM19
c
ICRF44
HI111
0
20
40
60
80
100
120
140
160
p=0.01
p=0.04
Adhe
ring
cells
(% o
f con
trol)
Supplemental Figure II. CD40L binds to the Mac-1 I-domain. Recombinant CD40L (12.5 or 50μg/ml) was incubated with the immobilized I-domain (10μg/ml) or BSA in a solid phase binding assay (a). Preparations of the isolated, recombinant αM I-domain were co-incubated with immobilized with Cd40L, Fibrinogen or ICAM-1 (each 10μg/ml) in the presence of 1mM Ca2+ or Mn2+. Binding is shown as % binding to control buffer containing Ca2+ (b) and was quantified by colorimetric reaction (a, b). Different antibody clones specifically blocking Mac-1’s I-domain (2LPM19c, ICRF44), CD40L (40804, 24-31), or LFA-1 (HI111) were tested for their capability to block adhesion of Mac-1 transfected CHO cells to immobilized fibrinogen (30μg/ml, c) or CD40L (10μg/ml, d). After pre-incubation with antibodies (10μg/ml), CHO cells (105/ml) were allowed to adhere for 30min. Unbound cells were removed by washing and adhering cells were counted. Data are presented as mean±SEM of at least three independent experiments. Fib.: Fibrinogen (b).
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-9-
50 100 150 200 250 300 350 Time (s)
14012010080604020
Resp
onse
(RU) CD40L
sM7M7
a
b0 5 10 15 20 25
0
500
1000
1500M7sM7
concentration (ug/ml)
Bind
ing
(AU)
Supplemental Figure III. Peptide M7 inhibits molecular interaction between the isolated Mac-1 I-domain and CD40L. Bound CD40L was preincubated for 10 min with peptides M7 and sM7 at 1.25-20μM, followed by incubation with 1mM I-domain-FITC conjugate for 30 min. Plates were washed and fluorescence of bound FITC-I-domain was detected (a). For surface plasmon resonance analysis recombinant I-domain was immobilized on the CM5 sensor chip by amine coupling. Binding was determined in the presence of M7 and sM7 (10μmol/l, b).
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from
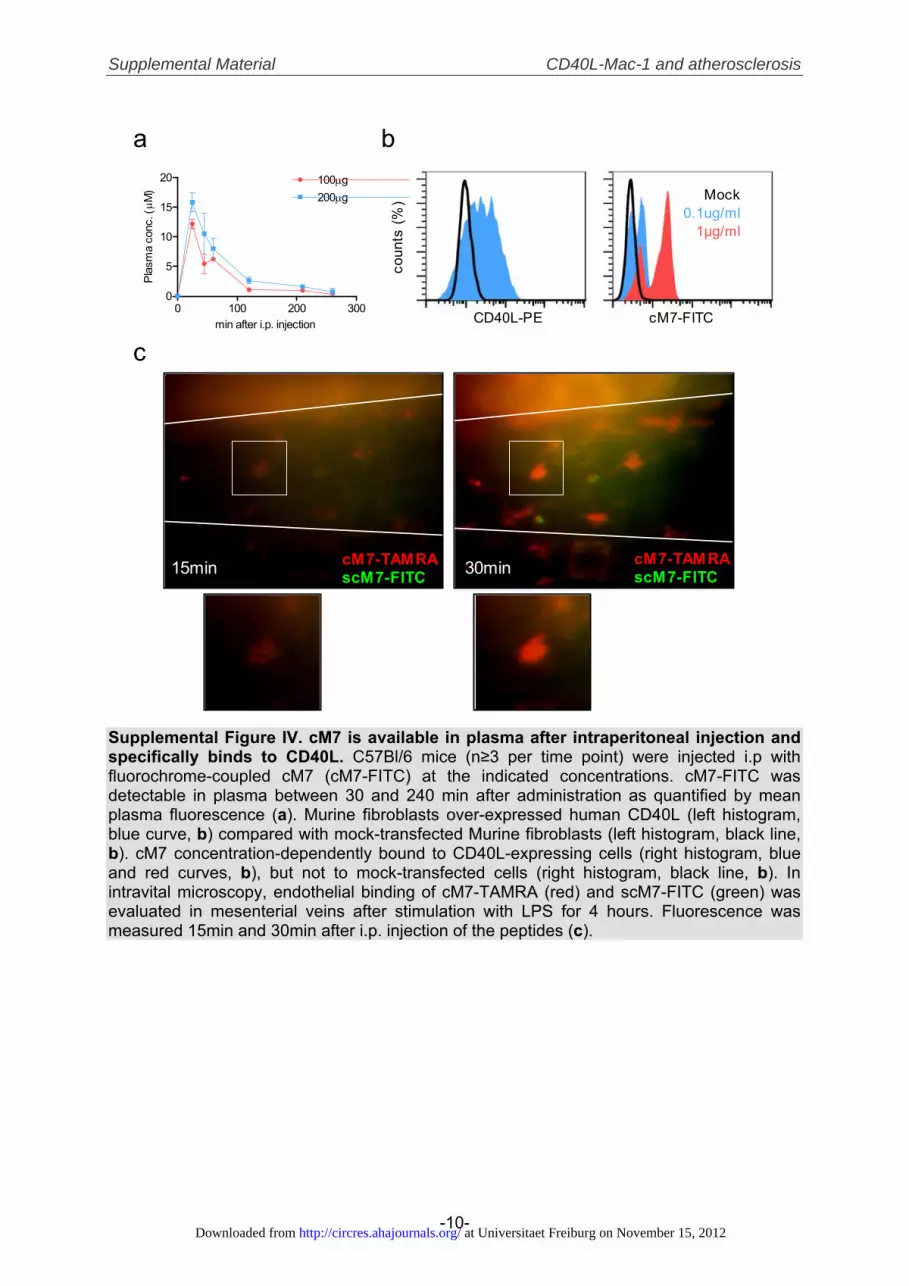
Supplemental Material CD40L-Mac-1 and atherosclerosis
-10-
a b
c
0 100 200 3000
5
10
15
20 100μg200μg
min after i.p. injection
Plas
ma
conc
. (μ M
)
cM7-TAMRAscM7-FITC15min 30min
coun
ts (%
) Mock0.1ug/ml
1µg/ml
CD40L-PE cM7-FITC
cM7-TAMRAscM7-FITC
Supplemental Figure IV. cM7 is available in plasma after intraperitoneal injection and specifically binds to CD40L. C57Bl/6 mice (n≥3 per time point) were injected i.p with fluorochrome-coupled cM7 (cM7-FITC) at the indicated concentrations. cM7-FITC was detectable in plasma between 30 and 240 min after administration as quantified by mean plasma fluorescence (a). Murine fibroblasts over-expressed human CD40L (left histogram, blue curve, b) compared with mock-transfected Murine fibroblasts (left histogram, black line, b). cM7 concentration-dependently bound to CD40L-expressing cells (right histogram, blue and red curves, b), but not to mock-transfected cells (right histogram, black line, b). In intravital microscopy, endothelial binding of cM7-TAMRA (red) and scM7-FITC (green) was evaluated in mesenterial veins after stimulation with LPS for 4 hours. Fluorescence was measured 15min and 30min after i.p. injection of the peptides (c).
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-11-
0
50
100
150
200
250 p=0.1 p=0.006Wildtype
CD40L-/-
Mac-1-/-PE
Cs
(% W
T)
a
b
saline
cM7
scM7
WT CD40L-/- Mac-1-/-
CD14+Gr-1-
CD14- Gr-1+
CD14- Gr-1-
saline
cM7
scM7
WT CD40L-/- Mac-1-/-
CD14+Gr-1-
CD14- Gr-1+
CD14- Gr-1-
Supplemental Figure V. Disruption of the CD40L/Mac-1 dyad does not modulate infiltration of specific leukocyte subsets to the inflamed peritoneal cavity. Baseline infiltration in WT, CD40L- and Mac-1-deficient mice (n=6 per group) was evaluated after injection with 3ml 4% thioglycollate broth followed by incubation for 15 hours. Cell infiltration was adjusted to percent of wildtype mice (a). Alternatively, 100μg of the peptides cM7, scM7 or saline were injected i.p. 15 hours after injection of thioglycollate, peritoneal exsudate cells were analyzed in FACS for expression of CD14 and Gr-1 (b). Granulocytes were defined as CD14- Gr-1+ cells, macrophages as CD14+ Gr-1- and lymphocytes as CD14+ Gr-1-.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-12-
0 10 20 300
20
40
60
80
100 cM7scM7
p=0.01
min after i.v. injection
Rol
ling
cell f
lux
(%)
Supplemental Figure VI. Specific inhibition of the CD40L/Mac-1 interaction modulates leukocyte recruitment within minutes after intravenous delivery. C57Bl/6 mice were injected with 200ng TNFα to induce peritoneal inflammation. Leukocyte rolling flux was quantified at the indicated time points in inflamed cremaster venules after intravenous delivery of 100μg of peptides cM7 or scM7.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-13-
Wildtyp
e -/-
CD40L
0
20
40
60
80
100
120 p=0.045
Adhe
ring
leuk
ocyt
es/fi
eld
Wild
type -/-
CD40L
0
20
40
60
80
100
120
140 p<0.001
Rol
ling
leuk
ocyt
e flu
x/fie
ld
WT CD40L-/-0
50
100
150 p=0.39
VCAM
-1 (%
)
0
50
100
150p=0.69
ICAM
-1 (%
)
WT CD40L-/-0
100200300400 p=0.30
CD
62P
(%)
0
50
100
150p=0.57
ICAM
-2 (%
)
contr
ol
CD40L
α ICAM-1
αCD11
b
α
0
20
40
60 p<0.05
p<0.05
p<0.05
Adhe
ring
mon
ocyt
es/fi
eld
anti-C
D40L
contr
ol
CHO
HU
VEC
anti-C
D11b
anti-C
D11a
a b
c d e
Supplemental Figure VII. Endothelial CD40L recruits inflammatory leukocytes by interaction with Mac-1. Mac-1-expressing CHO cells were allowed to adhere on TNFα-primed HUVECs (a), while both cell types were selectively blocked with antibodies either against Mac-1, CD40L, or LFA-1 (b). Anti-CD40L antibody blocked dynamic adhesion of human monocytes to HUVECs comparable with anti-ICAM-1 or anti-Mac-1 (n≥4). In intravital microscopy, numbers of adhering and rolling murine leukocytes decreased when interacting with CD40L-deficient endothelial cells compared with wild-type ECs (n=5 per group, c, d). CD40L deficiency did not regulate surface expression of the adhesion molecules ICAM-1, ICAM-2, VCAM-1, or P-selectin as determined by flow cytometry after stimulation with TNFα (e) for 24 hours. Data are presented as mean±SEM of at least three independent experiments.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from
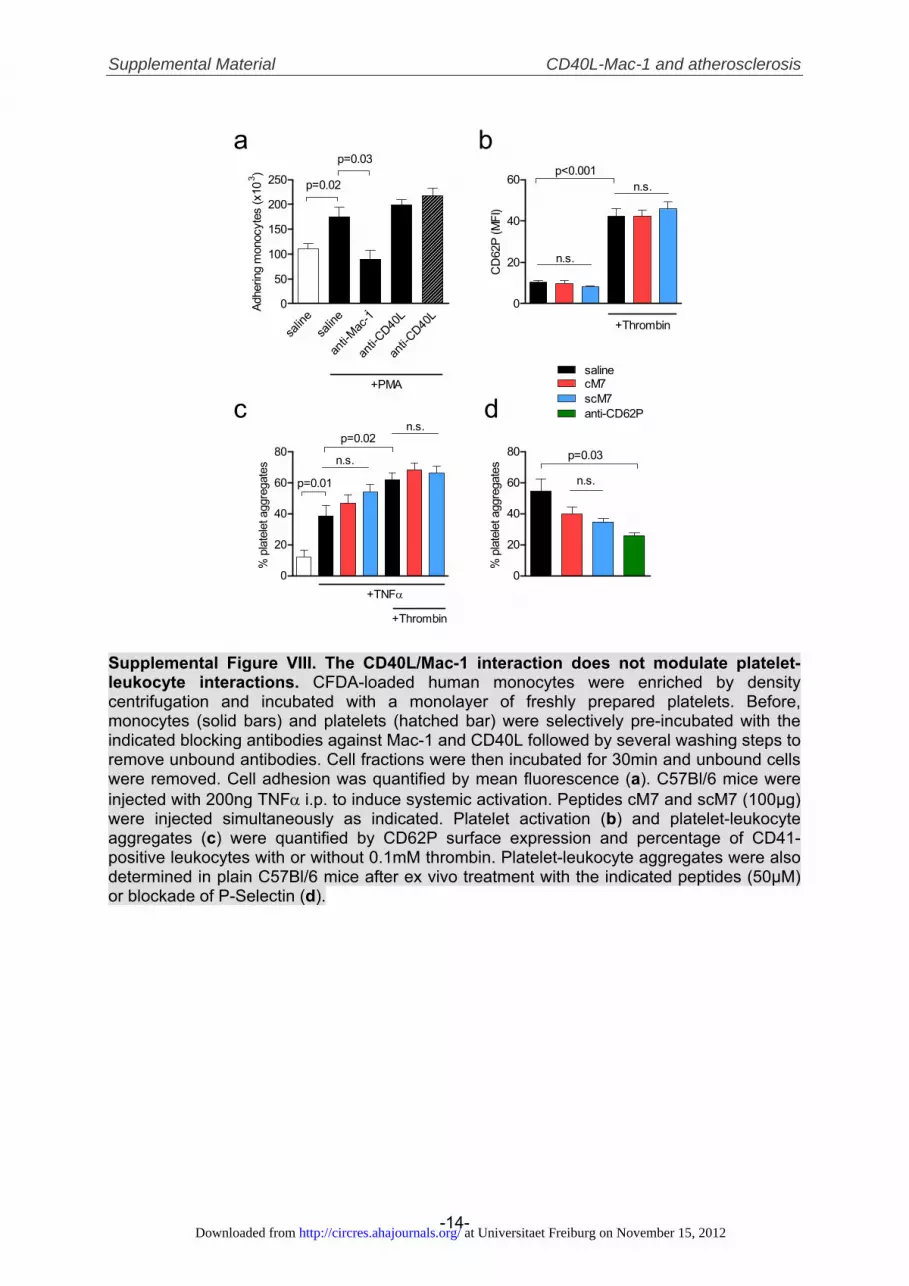
Supplemental Material CD40L-Mac-1 and atherosclerosis
-14-
a b
c d
0
20
40
60p<0.001
n.s.
n.s.
+Thrombin
CD
62P
(MFI
)
salin
esa
line
anti-M
ac-1
anti-C
D40L
anti-C
D40L
0
50
100
150
200
250
+PMA
p=0.02
p=0.03
´Adhe
ring
mon
ocyt
es (x
103 )
0
20
40
60
80
+TNFα
+Thrombin
p=0.01
p=0.02
n.s.
n.s.
% p
late
let a
ggre
gate
s
0
20
40
60
80
n.s.
p=0.03
salinecM7scM7anti-CD62P
% p
late
let a
ggre
gate
s
Supplemental Figure VIII. The CD40L/Mac-1 interaction does not modulate platelet-leukocyte interactions. CFDA-loaded human monocytes were enriched by density centrifugation and incubated with a monolayer of freshly prepared platelets. Before, monocytes (solid bars) and platelets (hatched bar) were selectively pre-incubated with the indicated blocking antibodies against Mac-1 and CD40L followed by several washing steps to remove unbound antibodies. Cell fractions were then incubated for 30min and unbound cells were removed. Cell adhesion was quantified by mean fluorescence (a). C57Bl/6 mice were injected with 200ng TNFα i.p. to induce systemic activation. Peptides cM7 and scM7 (100μg) were injected simultaneously as indicated. Platelet activation (b) and platelet-leukocyte aggregates (c) were quantified by CD62P surface expression and percentage of CD41-positive leukocytes with or without 0.1mM thrombin. Platelet-leukocyte aggregates were also determined in plain C57Bl/6 mice after ex vivo treatment with the indicated peptides (50μM) or blockade of P-Selectin (d).
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-15-
0
5
10
15
20p=0.39
p=0.02p=0.02
TNFα - + +
TNFα
(pg/
ml)
a b c
d e f
g
0
25
50
75
100
125p=0.004
p=0.046p=0.002
TNFα - + +
MC
P-1
(pg/
ml)
0
100
200
300n.s.
p=0.02
CD40L - + - + + +αMac-1 - - + + - - cM7 - - - - + - scM7 - - - - - +
pNFκB
total NFκB
pNFκ
B/to
tal N
Fκ B
(%)
0
50
100
150
200
250 p=0.90
p=0.21
p=0.03cM7
TNFα - + + +
salinescM7
p-p3
8 (M
FI)
0
20
40
60p=0.56p=0.045
CD40L - + +αMac-1 - - +
IL-8
(pg/
ml)
0
20
40
60
80
100
n.s.
p=0.03
CD40L - + + + cM7 - - + - scM7 - - - +
phos
po-E
RK1
/2 (U
/ml)
0
50
100
150
200
250
n.s.
p=0.01
CD40L - + + +cM7 - - + - scM7 - - - +
phos
po-J
NK1/
2 (U
/ml)
Supplemental Figure IX. CD40L induces activation of MAP-kinases and NFκB –but independently of interaction with Mac-1. C57Bl/6-mice were treated by intraperitoneal injection with 200ng TNFα to induce systemic inflammation. Peptides cM7 and scM7 (100μg) were injected simultaneously as indicated. Plasma cytokines TNFα (a), MCP-1 (b), and phosphorylated p38 (c) in peritoneal cell lysates were determined by cytometric bead array. To evaluate direct effects of CD40L on intracellular signalling, murine splenocytes were isolated, starved overnight and stimulated with CD40L (20μg/ml) in the presence of antibodies and peptides (all 10μg/ml). NFκB-activity was determined by phosphorylated portion of total NFκB as assessed by western blot (d). MAP-kinases ERK1/2 and JNK1/2 (e, f) were quantified by cytometric bead array. Human monocytes were enriched by density centrifugation, starved overnight and stimulated with CD40L (20μg/ml) in the presence or absence of blocking anti-Mac-1 antibody. Human IL-8 was determined by ELISA.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from
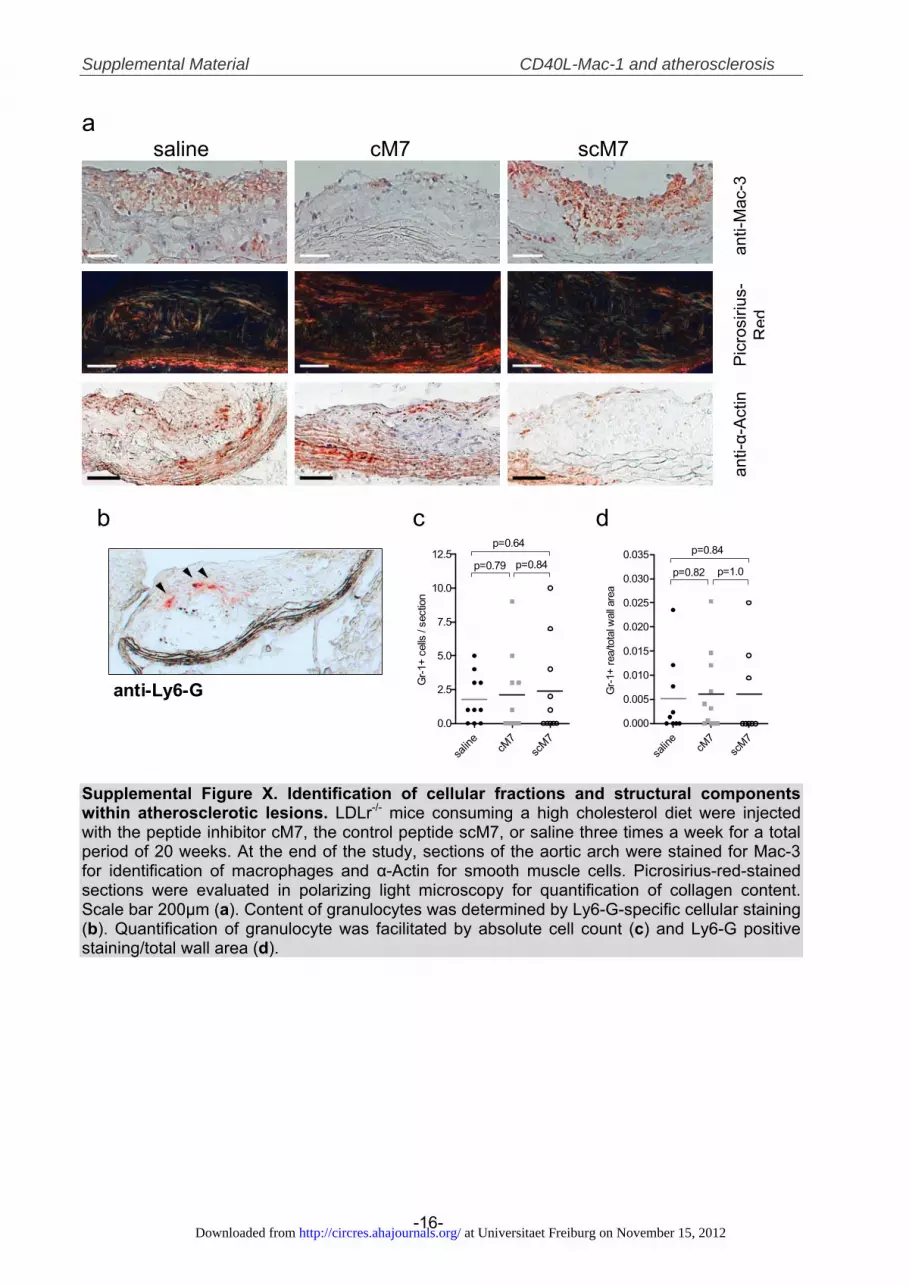
Supplemental Material CD40L-Mac-1 and atherosclerosis
-16-
a saline cM7 scM7
anti-
Mac
-3
Pic
rosi
rius-
Red
anti-α-
Act
in
salin
ecM
7sc
M70.0
2.5
5.0
7.5
10.0
12.5p=0.79 p=0.84
p=0.64
Gr-1
+ ce
lls /
sect
ion
salin
ecM
7sc
M70.000
0.005
0.010
0.015
0.020
0.025
0.030
0.035p=0.82 p=1.0
p=0.84
Gr-1
+ re
a/to
tal w
all a
rea
b c d
anti-Ly6-G
Supplemental Figure X. Identification of cellular fractions and structural components within atherosclerotic lesions. LDLr-/- mice consuming a high cholesterol diet were injected with the peptide inhibitor cM7, the control peptide scM7, or saline three times a week for a total period of 20 weeks. At the end of the study, sections of the aortic arch were stained for Mac-3 for identification of macrophages and α-Actin for smooth muscle cells. Picrosirius-red-stained sections were evaluated in polarizing light microscopy for quantification of collagen content. Scale bar 200μm (a). Content of granulocytes was determined by Ly6-G-specific cellular staining (b). Quantification of granulocyte was facilitated by absolute cell count (c) and Ly6-G positive staining/total wall area (d).
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-17-
saline cM7 scM70
100
200
300
p=0.53
p=0.22
p=0.43Il-
6 (%
of b
asel
ine)
saline cM7 scM70
25
50
75
100
125
150
175
p=0.18
p=0.62
p=0.50
MC
P-1
(% o
f bas
elin
e)
saline cM7 scM70
50
100
150
200
250
p=0.19
p=0.56
p=0.56
TNF-
α (%
of b
asel
ine)
saline cM7 scM70
50
100
150
200
p=0.37
p=0.30
p=0.82
IFN-
γ (%
of b
asel
ine)
saline cM7 scM70
50
100
150
200
250
p=0.10
p=0.05
p=0.86
IL-1
2 (%
of b
asel
ine)
saline cM7 scM70
5
10
15
20
p=0.18
p=0.11
p=0.70
CD
3+ C
D4+
CD
8- (%
lym
phoc
ytes
)
saline cM7 scM70
5
10
15
20
p<0.01
p=0.0001
p=0.10
CD
3+ C
D4-
CD
8+ (%
lym
phoc
ytes
)
saline cM7 scM70
20
40
60
80
100
120
140
Gr-1
high
G
r-1lo
w
p=0.24
p=0.24
p=0.77
Rat
io (%
CD
11b+
CD
115+
)
saline cM7 scM70
20
40
60
80
p=0.89
p=1.0
p=0.92
CD
19+
(% ly
mph
ocyt
es)
a b c
d
g h i
e f
Supplemental Figure XI. Inhibition of the CD40L-Mac-1 interaction does not cause a shift in Th1/Th2 polarization in vivo. LDLr-/- mice consuming a high cholesterol diet were injected with the peptide inhibitor cM7, the control peptide scM7, or saline three times a week for a total period of 20 weeks. Levels of plasma cytokines IL-6, IL-10, IL-12, TNFα, and IFNγ were quantified by a cytometric bead assay (a-f). T-cell subpopulations (f-g), B-cells (h), and Gr-1-positive inflammatory monocytes (i) were quantified by flow cytometry. Data are presented as mean±SEM.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-18-
SUPPLEMENTAL TABLES
Supplemental Table I: Peptides used in the study
Peptide Sequence I-domain* Structure MW (kDa)
M1 PHDFRRMKEFVST P147–T159 linear 1.649
M2 PITQLLGRTHTATGIRK P201–K217 linear 1.863
M3 KFGDPLGYEDVIPEADR K245–R261 linear 1.921
M4 DAFRSEKSRQELNTI D273–I287 linear 1.793
M5 FQVNNFEALKT F297–T307 linear 1.310
M6 QNNPNPRS Q190–S197 linear 0.925
M7 EQLKKSKTL E162–L170 linear 1.074
M8 EEFRIHFT E178–T185 linear 1.078
cM7 C-EQLKKSKTL-C E162–L170 cyclic 1.280
scM7 C-KLSLEKQTK-C n/a cyclic 1.280
FITC-cM7 C-EQLKKSKTL-C E162–L170 cyclic, FITC 1.638
TAMRA-cM7 C-EQLKKSKTL-C E162–L170 cyclic, TAMRA 1.690
FITC-scM7 C-KLSLEKQTK-C n/a cyclic, FITC 1.636
* indicates the stretch of the Mac-1 I-domain the peptide corresponds to
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from
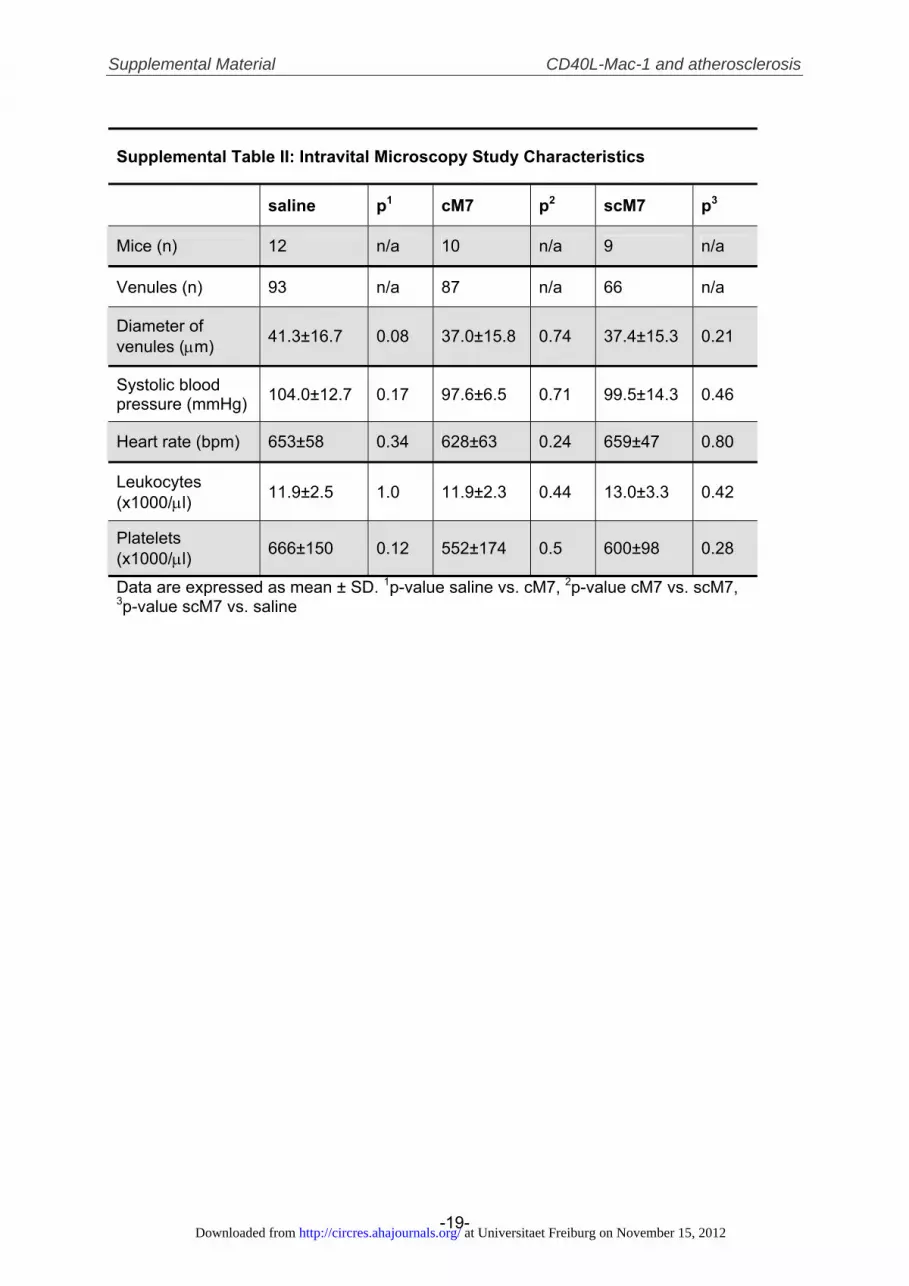
Supplemental Material CD40L-Mac-1 and atherosclerosis
-19-
Supplemental Table II: Intravital Microscopy Study Characteristics
saline p1 cM7 p2 scM7 p3
Mice (n) 12 n/a 10 n/a 9 n/a
Venules (n) 93 n/a 87 n/a 66 n/a
Diameter of venules (μm) 41.3±16.7 0.08 37.0±15.8 0.74 37.4±15.3 0.21
Systolic blood pressure (mmHg) 104.0±12.7 0.17 97.6±6.5 0.71 99.5±14.3 0.46
Heart rate (bpm) 653±58 0.34 628±63 0.24 659±47 0.80
Leukocytes (x1000/μl) 11.9±2.5 1.0 11.9±2.3 0.44 13.0±3.3 0.42
Platelets (x1000/μl) 666±150 0.12 552±174 0.5 600±98 0.28
Data are expressed as mean ± SD. 1p-value saline vs. cM7, 2p-value cM7 vs. scM7, 3p-value scM7 vs. saline
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-20-
SUPPLEMENTAL REFERENCES 1. Schuler P, Assefa D, Ylanne J, Basler N, Olschewski M, Ahrens I, Nordt T, Bode C,
Peter K. Adhesion of monocytes to medical steel as used for vascular stents is mediated by the integrin receptor mac-1 (cd11b/cd18; alpham beta2) and can be inhibited by semiconductor coating. Cell Commun Adhes. 2003;10:17-26
2. Zhang L, Plow EF. Identification and reconstruction of the binding site within alphambeta2 for a specific and high affinity ligand, nif. J Biol Chem. 1997;272:17558-17564
3. Morris AE, Remmele RL, Jr., Klinke R, Macduff BM, Fanslow WC, Armitage RJ. Incorporation of an isoleucine zipper motif enhances the biological activity of soluble cd40l (cd154). J Biol Chem. 1999;274:418-423
4. Schwarz M, Meade G, Stoll P, Ylanne J, Bassler N, Chen YC, Hagemeyer CE, Ahrens I, Moran N, Kenny D, Fitzgerald D, Bode C, Peter K. Conformation-specific blockade of the integrin gpiib/iiia: A novel antiplatelet strategy that selectively targets activated platelets. Circ Res. 2006;99:25-33
5. Eisenhardt SU, Schwarz M, Schallner N, Soosairajah J, Bassler N, Huang D, Bode C, Peter K. Generation of activation-specific human anti-alphambeta2 single-chain antibodies as potential diagnostic tools and therapeutic agents. Blood. 2007;109:3521-3528
6. Zirlik A, Maier C, Gerdes N, MacFarlane L, Soosairajah J, Bavendiek U, Ahrens I, Ernst S, Bassler N, Missiou A, Patko Z, Aikawa M, Schonbeck U, Bode C, Libby P, Peter K. Cd40 ligand mediates inflammation independently of cd40 by interaction with mac-1. Circulation. 2007;115:1571-1580
7. Straub A, Smolich J, d'Udekem Y, Brizard C, Peter K, Horton S. Activation of platelets in young infants during cardiopulmonary bypass. Thromb Haemost. 2010;103:466-469
8. Missiou A, Kostlin N, Varo N, Rudolf P, Aichele P, Ernst S, Munkel C, Walter C, Stachon P, Sommer B, Pfeifer D, Zirlik K, MacFarlane L, Wolf D, Tsitsikov E, Bode C, Libby P, Zirlik A. Tumor necrosis factor receptor-associated factor 1 (traf1) deficiency attenuates atherosclerosis in mice by impairing monocyte recruitment to the vessel wall. Circulation. 2010;121:2033-2044
9. Lister KJ, James WG, Hickey MJ. Immune complexes mediate rapid alterations in microvascular permeability: Roles for neutrophils, complement, and platelets. Microcirculation. 2007;14:709-722
10. Baez S. An open cremaster muscle preparation for the study of blood vessels by in vivo microscopy. Microvasc Res. 1973;5:384-394
11. Deguchi JO, Aikawa E, Libby P, Vachon JR, Inada M, Krane SM, Whittaker P, Aikawa M. Matrix metalloproteinase-13/collagenase-3 deletion promotes collagen accumulation and organization in mouse atherosclerotic plaques. Circulation. 2005;112:2708-2715
12. Whittaker P, Kloner RA, Boughner DR, Pickering JG. Quantitative assessment of myocardial collagen with picrosirius red staining and circularly polarized light. Basic Res Cardiol. 1994;89:397-410
13. Andre P, Prasad KS, Denis CV, He M, Papalia JM, Hynes RO, Phillips DR, Wagner DD. Cd40l stabilizes arterial thrombi by a beta3 integrin--dependent mechanism. Nat Med. 2002;8:247-252
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from

Supplemental Material CD40L-Mac-1 and atherosclerosis
-21-
LEGENDS FOR VIDEO FILES Videos 1-3 demonstrate how leukocyte recruitment in intravital microscopy is altered by treatment with saline (Supplemental Video 1), cM7 (Supplemental Video 2), scM7 (Supplemental Video 3). Videos 4-10 show effects of treatment with saline (Supplemental Video 4), an isotype control antibody (Supplemental Video 5), scM7 (Supplemental Video 6), cM7 (Supplemental Video 7), a blocking anti-CD40L antibody (Supplemental Video 8), a blocking anti-CD40 (Supplemental Video 9), and a blocking anti-Mac-1 antibody (Supplemental Video 10) on thrombus embolization rate in vivo.
at Universitaet Freiburg on November 15, 2012http://circres.ahajournals.org/Downloaded from