benzodiazepines and the potential trophic effect of antidepressants on dentate gyrus cells in mood...
TRANSCRIPT

Benzodiazepines and the potential trophic effectof antidepressants on dentate gyrus cells inmood disorders
Maura Boldrini1,2, Tanya H. Butt1, Adrienne N. Santiago1, Hadassah Tamir1,2,3,Andrew J. Dwork1,2,3, Gorazd B. Rosoklija1,2,4, Victoria Arango1,2, René Hen2,5,6,7
and J. John Mann1,2
1Division of Molecular Imaging and Neuropathology and New York State Psychiatric Institute, New York, NY, USA2Department of Psychiatry, Columbia University, New York, NY, USA3Department of Pathology and Cell Biology, Columbia University, New York, NY, USA4Macedonian Academy of Sciences & Arts, Republic of Macedonia5Division of Integrative Neuroscience, New York State Psychiatric Institute, New York, NY, USA6Department of Neuroscience and Columbia University, New York, NY, USA7Department of Pharmacology, Columbia University, New York, NY, USA
Abstract
Modest antidepressant response rates of mood disorders (MD) encourage benzodiazepine (BZD) co-medicationwith debatable benefit. Adult hippocampal neurogenesis may underlie antidepressant responses, but diazepamco-administration impairs murine neuron maturation and survival in response to fluoxetine. We counted neuralprogenitor cells (NPCs), mitotic cells, and mature granule neurons post-mortem in dentate gyrus (DG) fromsubjects with: untreated Diagnostic and Statistical Manual of Mental Disorders (DSM) IV MD (n=17);antidepressant-treated MD (MD*ADT, n=10); benzodiazepine-antidepressant-treated MD (MD*ADT*BZD, n=7);no psychopathology or treatment (controls, n=18).MD*ADT*BZD had fewer granule neurons vs. MD*ADT in anterior DG and vs. controls in mid DG, and did
not differ from untreated-MD in any DG subregion. MD*ADT had more granule neurons than untreated-MD inanterior and mid DG and comparable granule neuron number to controls in all dentate subregions. Untreated-MD had fewer granule neurons than controls in anterior and mid DG, and did not differ from any other group inposterior DG. MD*ADT*BZD had fewer NPCs vs.MD*ADT in mid DG. MD*ADT had more NPCs vs. untreated-MD and controls in anterior and mid DG. MD*ADT*BZD and MD*ADT had more mitotic cells in anterior DG vs.controls and untreated-MD. There were no between-group differences in mid DG in mitotic cells or in posteriorDG for any cell type.Our results in mid-dentate, and to some degree anterior dentate, gyrus are consistent with murine findings
that benzodiazepines counteract antidepressant-induced increases in neurogenesis by interfering with progenitorproliferation. We also confirmed, in this expanded sample, our previous finding of granule neuron deficit inuntreated MD.
Received 7 November 2013; Reviewed 4 January 2014; Revised 25 March 2014; Accepted 26 April 2014
Key words: Ki-67, Nestin, NeuN, Neurogenesis, Stereology.
Introduction
To manage the anxiety component of mood disorders(MD), a benzodiazepine (BZD) is often co-administeredwith antidepressant medication (Bandelow et al., 2008).Selective serotonin reuptake inhibitors (SSRIs) may in-crease anxiety in MD during the initial treatment of MD
and, as an alternative to lowering SSRI dosage, adjunctiveBZD use may be recommended (Edwards and Anderson,1999; Bandelow et al., 2008). Recently, there has beenincreasing concern that such BZD administration mayworsen depression, cause transient cognitive or motor im-pairment, and potentially lead to abuse or dependence(Lader, 2011; Dell’osso and Lader, 2013). Additionalstudies have also suggested longer-term cognitive impair-ment with prolonged BZD use (Klein et al., 2009;Wu et al., 2009).
New neurons develop from neural progenitor cells(NPCs) in adult human dentate gyrus (DG), and matureinto functional granule neurons (Eriksson et al., 1998)that contribute to hippocampal-dependent functions,such as learning and memory (Saxe et al., 2006) and
Address for correspondence: Maura Boldrini, Associate Professor ofNeurobiology in Psychiatry, Columbia University Medical Center –New York State Psychiatric Institute, 1051 Riverside drive, Box 42,New York, NY 10032, USA.Tel.: +1-646-774-7523 Fax: +1-212-543-6017Email: [email protected]
International Journal of Neuropsychopharmacology, Page 1 of 11. © CINP 2014doi:10.1017/S1461145714000844
ARTICLE

pattern discrimination (Clelland et al., 2009; Aimoneet al., 2011).
Increased neurogenesis and cell survival improvespattern discrimination (Sahay et al., 2011). Neurogenesismediates recovery from stress via environmental enrich-ment (Schloesser et al., 2010) and antidepressant effectson chronic unpredictable stress, novelty suppressed feed-ing (Surget et al., 2008) and contextual discrimination(Tronel et al., 2012). Nevertheless, adult neurogenesisis not necessary for all antidepressant responses inanimal models, suggesting the existence of neurogenesis-dependent and -independent mechanisms of antidepres-sant action, at least in rodent depression models (Davidet al., 2009).
Antidepressant treatment in major depression is asso-ciated with more mitotic cells, NPCs (Boldrini et al.,2009, 2012) and mature granule neurons (Boldrini et al.,2013) in human DG. This antidepressant effect wouldmitigate the defective neurogenesis that is hypothesizedto contribute to the pathogenesis of major depression(Kempermann and Kronenberg, 2003). In support of thishypothesis, we find fewer mature granule neurons inhippocampal DG of untreated subjects with major de-pression compared with controls, as well as a decreasein DG granule neurons associated with earlier onset ofmajor depression (Boldrini et al., 2013). In addition, wefound that more lifetime major depressive episodes arecorrelated with smaller DG volume (Boldrini et al.,2013), consistent with magnetic resonance (MR) volumefindings in vivo (Sheline et al., 2003; McKinnon et al., 2009).
In rodents, co-administration of diazepam and fluoxe-tine inhibits the neurogenesis effect of fluoxetine and thesuppression of anxious and depressive behavior (Wu andCastren, 2009; Sun et al., 2013). Although specific under-lying mechanisms of BZD function remain unknown,tonic and phasic gamma-amino-butyric acid (GABA) acti-vation regulates the synaptic integration of newbornneurons in murine DG (Ge et al., 2006). Additionally,a balance of glutamatergic and GABAergic transmissionclosely regulates adult hippocampal neurogenesis (Sunet al., 2009). BZDs, which act as GABAA receptor agonists(Rudolph et al., 1999) may impact hippocampal neuro-genesis by enhancing GABAergic signaling and causingan imbalance in neuronal activity.
The effect of BZDs on the relationship between adulthippocampal neurogenesis and antidepressant use hasnot been studied in the brain of depressed patients. Inthis study we assessed the relationship of BZD to antide-pressant co-treatment by quantifying neural progenitorcell, mitotic cell, and mature granule neuron number inhuman DG of subjects with mood disorders. We hypo-thesized that fewer mature granule neurons, NPCs, andmitotic cells would be observed in subjects with MDco-treated with BZD and antidepressants, comparedwith those treated with antidepressants alone. Other com-parison groups were untreated MD and non-psychiatriccontrols.
Method
Brain collection
International Review Board (IRB) approval was ob-tained for all research conducted. Post-mortem tissuewas acquired from the Macedonian/New York State Psy-chiatric Institute brain collection. We dissected the hippo-campus from 2-cm thick coronal blocks of the righthemisphere that were frozen in dichlorodifluoromethane(−30 °C) and stored at −80 °C at the time of autopsy.Samples of selected brain areas were formalin-fixed forneuropathology screening and brain pH determination.Toxicology tests were performed on cerebellar tissue,blood, and other body fluids.
Clinical measures
Subjects were diagnosed using a psychological autopsyand the Structured Clinical Interview for DSM-IV-TRAxis I Disorders (SCID I) or NP (Non-Patient edition)and Structured Clinical Interview for DSM-IV Axis IIPersonality Disorders (SCID II) (Lobbestael et al., 2011),using a method validated for Diagnostic and StatisticalManual of Mental Disorders (DSM) axis I and II diagnoses(Kelly and Mann, 1996). History of lifetime mood disor-ders, developmental history, and recentmedication historywere obtained. Other instruments and assessments in-cluded the Global Assessment Scale (Endicott et al.,1976). Cause of death, time to autopsy and freezer storagetime were noted.
Subjects
Four groups of subjects were studied: benzodiazepine-antidepressant-treated MDs (MD*ADT*BZD; N=7), MDstreated with antidepressants only (MD*ADT; N=10),untreated MDs (N=17) and controls without psychiatricdisease or treatment (N=18). The percentage of bipolarand major depressive disorder subjects was not differentbetween groups (Table 1). Subjects were included intreated groups if they received drug prescriptions in thelast three months of life and tested positive for such drugs(brain or blood toxicology) at autopsy. Groups werematched for sex and post-mortem interval (PMI) becauseof the influence of estrogen on neurogenesis (Saraviaet al., 2007) and the possible effect of PMI on antigen po-tency. Males and females were equally distributed in thedifferent subject groups (χ2=3.456; df=3; p=0.327). Therewas no difference between groups in terms of PMI (p=0.903). Age differed between groups (F=3.2230; df=3,48;p=0.030): MD*ADT were younger than MD*ADT*BZD(p=0.036) and untreated-MDs (p=0.030). No differenceswere found between other groups. Therefore, age wasincluded as a covariate in analyses of group effect.
Hippocampus preparation
The whole right hippocampus was dissected from con-secutive frozen coronal blocks, then fixed in 4%
2 M. Boldrini et al.

Table 1. Demographic and clinical characteristics of subjects
Group (n)Age (yr)(mean±S.E.M)
PMI (h)(mean±S.E.M)
Brain pH(mean±S.E.M)
Sex(M:F)
Suicide(n)
DSM axis Idiagnosis Blood and brain toxicology
GAS(mean±S.E.M)
N. episodesof MDE(mean±S.E.M)
Age of firstMDE(mean±S.E.M)
Medicationprescribedduring the last3 months of life
SmokingTobacco
C (n=18) 46±4.1 14.7±1.2 6.42±0.08 11:7 0 None (18) None (9) 84.3±2.51 – – None (18) Yes (7)Anesthetics (4)Carbon Monoxide (1)
No (10)
MD (n=17) 51±4.6 14.7±1.8 6.08±0.07 11:6 13 Primary:MDD (16)
None (14) 54.4±5.88 1.66±0.32 44.6±5.09 None (16) Yes (6)
BPD (1) Analgesic (1) Nitroglycerine (1) No (8)Comorbidity:OCD (1)
Atenolol (1)
GD (1) Albuterol (1)AA (1)
MD*ADT(n=10)
34±2.6 15.8±2.2 6.39±0.12 3:7 8 Primary:MDD (7)
Anesthetics (1) 35.3±7.24 3±0.96 25.3±2.69 Amitriptyline (1) Yes (3)
BPD (3) Antidepressants (BupropionFluoxetine NortriptylineParoxetine Sertraline) (10)
BenztropineMesylate (1)
No (7)
Comorbidity:CA (1)
Lithium (2) Bupropion (1)
BA (2) Opioids (1) Fluoxetine (3)Solvents (1) Fluphenazine (1)
Lithium (2)Nortriptyline (2)Paroxetine (1)Sertraline (2)Trazodone (3)Valproic Acid (1)
MD*ADT*BZD(n=7)
57±6.8 16.9±4.3 6.57±0.18 4:3 4 Primary:MDD (4)
Analgesic (2) 11.2±7.14 1 68.7±7.66 Sertraline (1) Yes (1)
BPD (3) Antidepressants (7)Antipsychotics (1)Barbiturates (2)
Clomipramine (1) No (3)
BZD (7)Fluoxetine (1) Mianserin (1)Opioids (2)Sertraline (2)
AA, Alcohol Abuse; BA, Bulimia/Anorexia; BPD, Bipolar Disorder; BZD, Benzodiazepines; C,controls without psychopathology or treatment; CA, Cannabis Abuse; DSM, Diagnostic and StatisticalManual of Mental Disorders; GAS, Global Assessment Scale; GD, Gambling Disorder; MDD, Major Depressive Disorder; MD, Untreated subjects with mood disorder; MD*ADT, Subjects withmood disorder treated with antidepressant only; MD*ADT*BZD, Subjects with mood disorder treated with antidepressants and benzodiazepines; MDE, major depressive episode; OCD, ObsessiveCompulsive Disorder; PMI, post-mortem interval; SEM, standard error of the mean.
Benzodiazepinesand
hippocampalneuroplasticity
3

paraformaldehyde phosphate buffer saline at 4 °C andcryoprotected in 30% sucrose. Sections were cut (at50 μm) using a freezing microtome (Microm HM440E)and stored in 40-well boxes at −20 °C in cryoprotectant(30% ethylene glycol in 0.1 M phosphate buffer). Whilesectioning, reference slides at 1-mm intervals werestained with Cresyl Violet, and were later used to alignsections processed for immunohistochemistry along theanterior–posterior axis of the hippocampal formation.
Immunohistochemistry and stereology
Sections were processed to identify mature granuleneurons (anti-Neuronal Nuclear antigen [NeuN] mousemonoclonal antibody, 1:100000; Chemicon, California),NPCs (anti-nestin mouse monoclonal antibody, 1:8000,Chemicon) and mitotic cells (anti-Ki-67 (Scholzen andGerdes, 2000) mouse monoclonal antibody, 1:200,Novocastra Clone-MM1, UK). Immunohistochemistryand stereology were performed as previously described(Boldrini et al., 2009, 2012, 2013). The choice of nestin asa marker to detect NPCs was determined by the factthat nestin (NEural STem cell proteIN) is a class VI inter-mediate filament protein expressed during development,until around post-natal day 11 in rat cortex and graduallyreplaced by intermediate filament proteins specific formature cells, such as glial fibrillary acidic protein(GFAP) in glial cells and other types of neurofilament inneurons (Kalman and Ajtai, 2001). In transgenic adultmice expressing green fluorescent protein under the con-trol of regulatory regions of the nestin gene, nestin-positive cells eventually express markers of neuroblasts:polysialated neuronal cell adhesion molecule, doublecor-tin and the transcription factor NeuroD (Yamaguchi et al.,2000). In culture, nestin-immunoreactive cell spheresdifferentiate into neurons and glia (Itoh et al., 2006).Moreover, nestin-positive type II NPCs, but not GFAP-positive type I NPCs, are targeted by antidepressants(Encinas et al., 2006) (15). Therefore, nestin seems anideal marker to examine NPCs and their response toantidepressants in the adult brain. However, nestinre-expression in reactive astrocytes is induced by cerebralischemia (Duggal et al., 1997), traumatic brain injury(Sahin et al., 1999), de-afferentation (Brook et al., 1999),and neurotoxicity (Yoo et al., 2005), but reactive astro-cytes can be distinguished by morphology and becausethey express both nestin and GFAP (Yoo et al., 2005).Nestin is also a marker of neovascularization, and itlabels capillaries and newly formed vessels after ischemiauntil the stroke is remodeling, while it is not expressedin absence of vascular response (Mokry et al., 2008;Salehi et al., 2008).
Statistical analysis
Regression analysis tested correlations between cell num-bers and continuous variables. Age was used as covariate
in a multivariate analysis of covariance (MANCOVA)to assess the effect of group on cell numbers. We usedanalysis of variance (ANOVA) with Tukey post-hoc testfor between-group (MD*ADT*BZD, MD*ADT, untreatedMDs and non-psychiatric controls) comparisons of con-tinuous dependent variables. Mitotic cell number wascompared using a non-parametric test (Mann–Whitney).For the comparison of qualitative variables, a χ2-testwas employed. We set p<0.05 for significance level.All statistics were analyzed using SPSS (18.0.3;Apache Software Foundation) and data were expressedas mean±S.E.M.
Results
Between-groups comparisons for DG granule neurons,NPCs and mitotic cells
There was a group effect on mature granule neuron, NPC,and mitotic cell numbers in anterior and mid DG, using aMANCOVA with group as the independent variable andage as covariate. No group differences were found inposterior DG for any cell type. For representative imagesof hippocampal tissue from each group showing NPCsand mitotic cells, see Fig. 1.
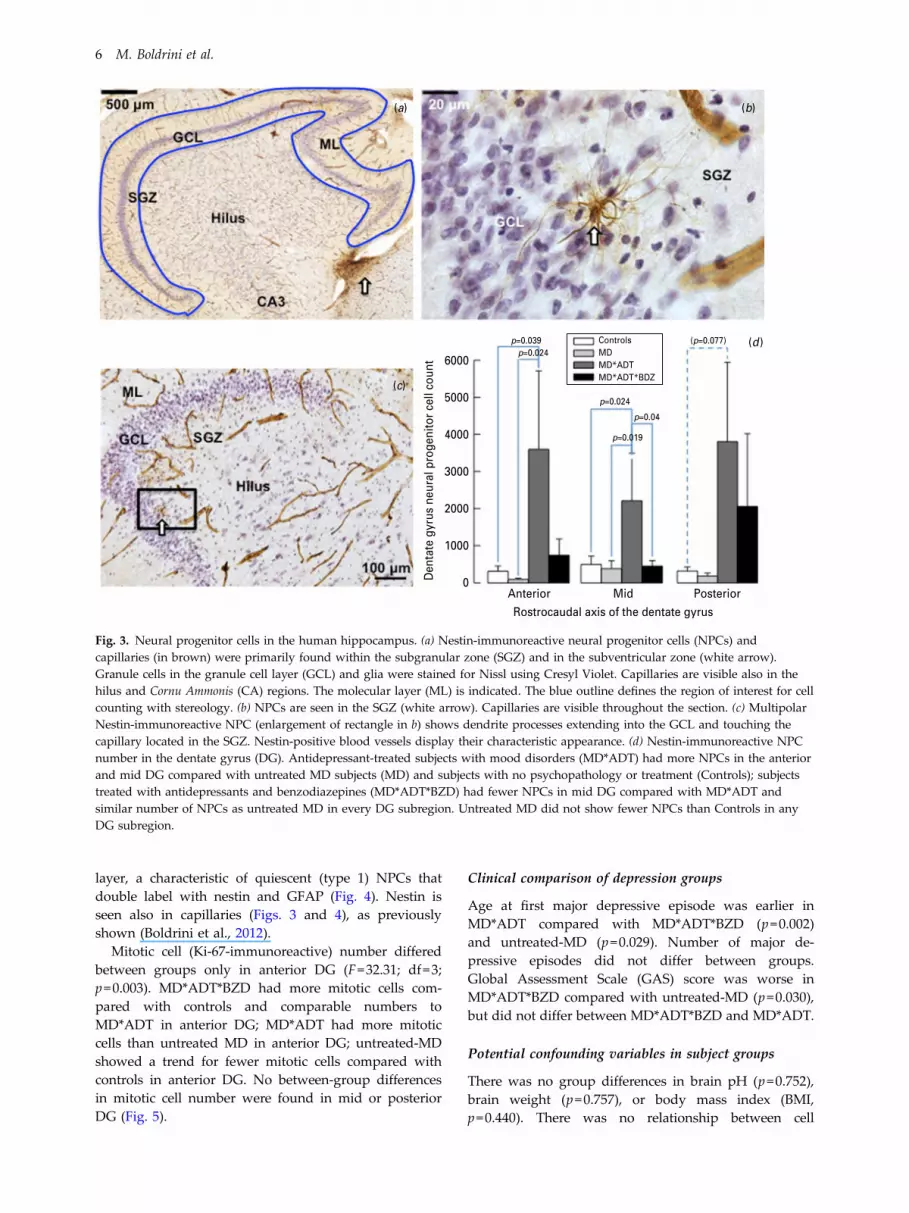
Mature granule neuron (NeuN-immunoreactive) num-ber differed between groups in the anterior (F=6.846;df=3; p=0.001) and mid (F=4.475; df=3,40; p=0.008)DG. MD*ADT*BZD had fewer granule neurons com-pared with MD*ADT in anterior DG, and with controlsin mid DG; MD*ADT had more granule neurons thanuntreated-MD in anterior and mid DG, and comparablegranule neuron number to controls in all DG regions.Untreated-MD had fewer granule neurons than controlsin anterior and mid DG, but did not differ from anyother group in posterior DG; no between-group differ-ences in granule neuron number were found in posteriorDG (Fig. 2).
NPC (nestin-immunoreactive) number differed be-tween groups in anterior (F=4.949; df=3; p=0.014) andmid DG (F=12.965; df=3; p<0.001), but not in posteriorDG. MD*ADT*BZD had fewer NPCs compared withMD*ADT in mid DG. MD*ADT had more NPCs com-pared with controls and untreated MD in anterior andmid DG. Post-hoc test for multiple comparisons did notshow significance differences in NPC number betweenuntreated-MD (U-MD) and controls (C) in anterior(U-MD=84±24; C=583±180), mid (U-MD=323±196;C=649±221) or posterior (U-MD=171±65; C=1253±60)DG. No between-group differences in NPC numberwere found in posterior DG (Fig. 3). The nestin-immunoreactive cells counted in the present study werefound in the SGZ of the human DG and displayed themorphology of amplifying (type 2) NPCs, as previouslyshown (Boldrini et al., 2012). Nestin-immunoreactivecells did not have vertical processes crossing the granulecell layer and ending in elaborate arbors in the molecular
4 M. Boldrini et al.
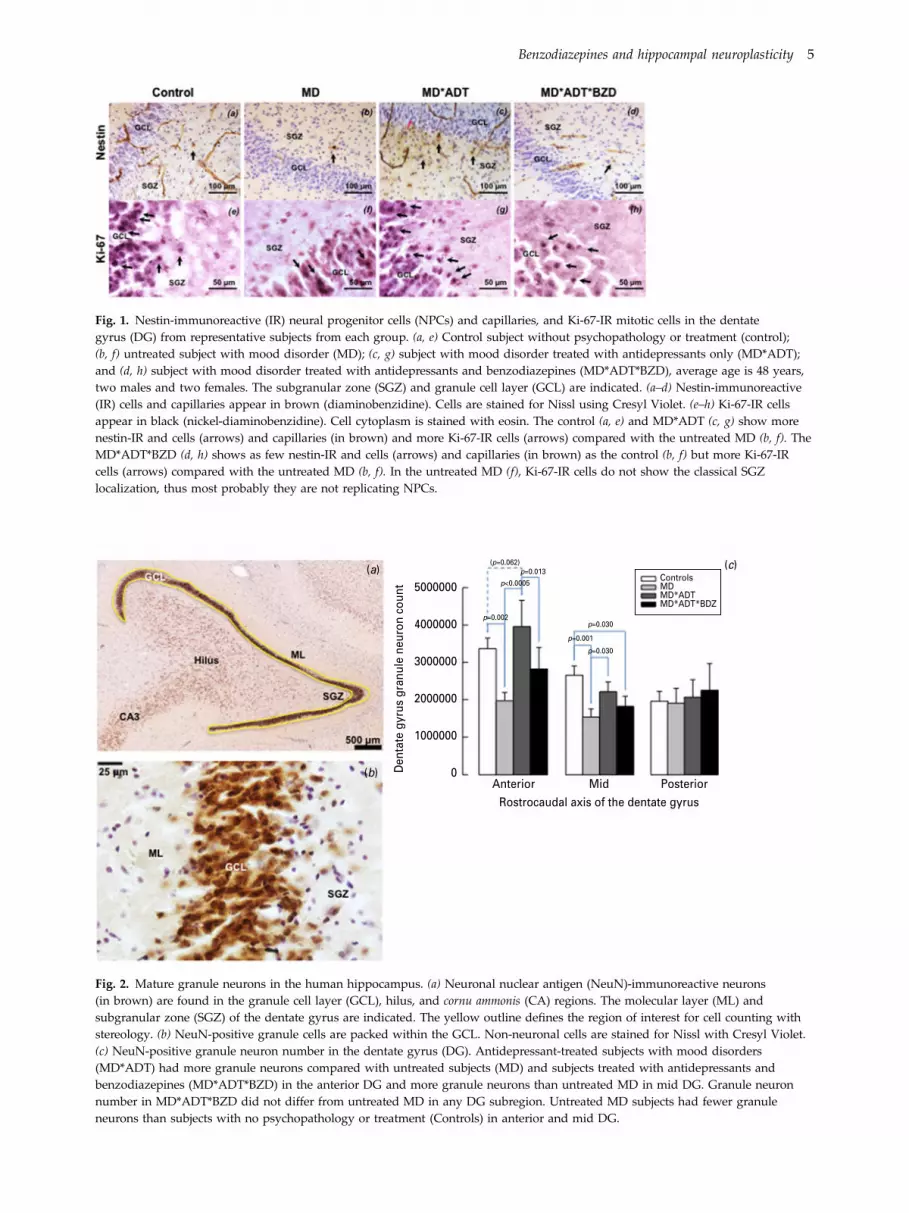
Fig. 1. Nestin-immunoreactive (IR) neural progenitor cells (NPCs) and capillaries, and Ki-67-IR mitotic cells in the dentategyrus (DG) from representative subjects from each group. (a, e) Control subject without psychopathology or treatment (control);(b, f) untreated subject with mood disorder (MD); (c, g) subject with mood disorder treated with antidepressants only (MD*ADT);and (d, h) subject with mood disorder treated with antidepressants and benzodiazepines (MD*ADT*BZD), average age is 48 years,two males and two females. The subgranular zone (SGZ) and granule cell layer (GCL) are indicated. (a–d) Nestin-immunoreactive(IR) cells and capillaries appear in brown (diaminobenzidine). Cells are stained for Nissl using Cresyl Violet. (e–h) Ki-67-IR cellsappear in black (nickel-diaminobenzidine). Cell cytoplasm is stained with eosin. The control (a, e) and MD*ADT (c, g) show morenestin-IR and cells (arrows) and capillaries (in brown) and more Ki-67-IR cells (arrows) compared with the untreated MD (b, f). TheMD*ADT*BZD (d, h) shows as few nestin-IR and cells (arrows) and capillaries (in brown) as the control (b, f) but more Ki-67-IRcells (arrows) compared with the untreated MD (b, f). In the untreated MD (f), Ki-67-IR cells do not show the classical SGZlocalization, thus most probably they are not replicating NPCs.
Den
tate
gyr
us g
ranu
le n
euro
n co
unt 5000000
(p=0.062)p=0.013
p=0.002p=0.030
p=0.001
p=0.030
p<0.0005ControlsMDMD*ADTMD*ADT*BDZ
4000000
3000000
2000000
1000000
0Anterior Mid
Rostrocaudal axis of the dentate gyrus
Posterior
(a)
(b)
(c)
Fig. 2. Mature granule neurons in the human hippocampus. (a) Neuronal nuclear antigen (NeuN)-immunoreactive neurons(in brown) are found in the granule cell layer (GCL), hilus, and cornu ammonis (CA) regions. The molecular layer (ML) andsubgranular zone (SGZ) of the dentate gyrus are indicated. The yellow outline defines the region of interest for cell counting withstereology. (b) NeuN-positive granule cells are packed within the GCL. Non-neuronal cells are stained for Nissl with Cresyl Violet.(c) NeuN-positive granule neuron number in the dentate gyrus (DG). Antidepressant-treated subjects with mood disorders(MD*ADT) had more granule neurons compared with untreated subjects (MD) and subjects treated with antidepressants andbenzodiazepines (MD*ADT*BZD) in the anterior DG and more granule neurons than untreated MD in mid DG. Granule neuronnumber in MD*ADT*BZD did not differ from untreated MD in any DG subregion. Untreated MD subjects had fewer granuleneurons than subjects with no psychopathology or treatment (Controls) in anterior and mid DG.
Benzodiazepines and hippocampal neuroplasticity 5
layer, a characteristic of quiescent (type 1) NPCs thatdouble label with nestin and GFAP (Fig. 4). Nestin isseen also in capillaries (Figs. 3 and 4), as previouslyshown (Boldrini et al., 2012).
Mitotic cell (Ki-67-immunoreactive) number differedbetween groups only in anterior DG (F=32.31; df=3;p=0.003). MD*ADT*BZD had more mitotic cells com-pared with controls and comparable numbers toMD*ADT in anterior DG; MD*ADT had more mitoticcells than untreated MD in anterior DG; untreated-MDshowed a trend for fewer mitotic cells compared withcontrols in anterior DG. No between-group differencesin mitotic cell number were found in mid or posteriorDG (Fig. 5).
Clinical comparison of depression groups
Age at first major depressive episode was earlier inMD*ADT compared with MD*ADT*BZD (p=0.002)and untreated-MD (p=0.029). Number of major de-pressive episodes did not differ between groups.Global Assessment Scale (GAS) score was worse inMD*ADT*BZD compared with untreated-MD (p=0.030),but did not differ between MD*ADT*BZD and MD*ADT.
Potential confounding variables in subject groups
There was no group differences in brain pH (p=0.752),brain weight (p=0.757), or body mass index (BMI,p=0.440). There was no relationship between cell
6000
p=0.039p=0.024
p=0.024
p=0.04
(p=0.077)
p=0.019
5000
4000
3000
2000
1000
0Anterior Mid Posterior
Den
tate
gyr
us n
eura
l pro
geni
tor
cell
coun
t
Rostrocaudal axis of the dentate gyrus
ControlsMDMD*ADTMD*ADT*BDZ
(a) (b)
(d )
(c)
Fig. 3. Neural progenitor cells in the human hippocampus. (a) Nestin-immunoreactive neural progenitor cells (NPCs) andcapillaries (in brown) were primarily found within the subgranular zone (SGZ) and in the subventricular zone (white arrow).Granule cells in the granule cell layer (GCL) and glia were stained for Nissl using Cresyl Violet. Capillaries are visible also in thehilus and Cornu Ammonis (CA) regions. The molecular layer (ML) is indicated. The blue outline defines the region of interest for cellcounting with stereology. (b) NPCs are seen in the SGZ (white arrow). Capillaries are visible throughout the section. (c) MultipolarNestin-immunoreactive NPC (enlargement of rectangle in b) shows dendrite processes extending into the GCL and touching thecapillary located in the SGZ. Nestin-positive blood vessels display their characteristic appearance. (d) Nestin-immunoreactive NPCnumber in the dentate gyrus (DG). Antidepressant-treated subjects with mood disorders (MD*ADT) had more NPCs in the anteriorand mid DG compared with untreated MD subjects (MD) and subjects with no psychopathology or treatment (Controls); subjectstreated with antidepressants and benzodiazepines (MD*ADT*BZD) had fewer NPCs in mid DG compared with MD*ADT andsimilar number of NPCs as untreated MD in every DG subregion. Untreated MD did not show fewer NPCs than Controls in anyDG subregion.
6 M. Boldrini et al.

numbers and PMI, brain pH, brain weight, or BMI. Malesand females were equally distributed in the differentgroups (see methods). There was no sex difference innumber of NPC, mitotic cells, and granule neurons inany group. Age differed between groups and was usedas a covariate in the statistical analysis to testbetween-group differences (see methods).
Discussion
This first human brain study of the effect of augmentingantidepressant treatment with adjunctive BZDs on neuro-genesis in the dentate gyrus in mood disorders finds evi-dence consistent with mouse studies showing BZDsinhibit the antidepressant enhancement of neurogenesis.MD treated with BZDs and antidepressants had fewerNPCs and mature granule neurons in DG than MD trea-ted with antidepressant alone. Both MD groups had com-parable mitotic cell number.
Mature granule neurons
In the anterior DG, MD co-treated with antidepressantsand BZDs had granule neuron number comparable tountreated-MD and fewer granule neurons than MD trea-ted with antidepressants. Long-term BZD use has beenlinked to sustained cognitive and psychomotor
impairment (Lader, 2011; Dell’osso and Lader, 2013).Our results suggest longer-term impairment may be re-lated to fewer mature granule neurons in subjectsco-treated with BZD.
Untreated MD had fewer granule neurons comparedwith non-psychiatric controls in anterior and mid DG.We had previously reported fewer granule neurons in an-terior and mid DG in untreated major depressive dis-order, in a partially overlapping, smaller sample(Boldrini et al., 2013). This study includes subjects withbipolar disorder. The percentage of bipolar and major de-pressive disorder (MDD) subjects did not differ betweentreatment groups. Our subsample sizes were too smallto determine whether MD subtype influences cellnumbers.
MD treated with antidepressants alone had more ma-ture granule neurons in anterior and mid DG comparedwith untreated MD. In a previous study (Boldrini et al.,2013), we reported that SSRI-treated subjects with MDDhad more granule neurons than untreated MDD subjectsonly in mid DG. Our previous study sample differed fromthis sample because it did not exclude from theSSRI-treated MDD group subjects that had receivedBZD treatment. The present study also included in theantidepressant-treated group subjects treated with tri-cyclic antidepressants and subjects treated with lithium,but without BDZ treatment. The number of subjectsinvestigated was insufficient to allow analysis of poten-tially different effects of SSRIs, tricyclics, and lithium.The absence of subjects with BZD treatment likelystrengthened the results in terms of detecting the overalleffect of antidepressants.
Neural progenitor cells
Based on morphology and antigenicity, thenestin-immunoreactive cells we counted are amplifyingNPCs. MD co-treated with antidepressants and BZDdid not differ from untreated-MD in NPC number andhad fewer NPCs in mid DG compared with MDs treatedwith antidepressants alone. This picture is consistent withBZDs inhibiting early stage neuronal development.Another possibility is that subjects treated with BZDand antidepressants may have had more severe de-pression such as higher levels of anxiety compared tosubjects treated with antidepressants alone. Such clinicalcharacteristics may be a consequence or a cause of dimin-ished NPC and mature neuron number. However, age ofonset of major depression, number of episodes of de-pression, and GAS score, do not suggest that the BZD-antidepressant co-treated MD group was more severelyill than those treated with antidepressants only. Thereforeseverity of illness is unlikely to explain the findings.
Untreated MD did not have fewer NPCs comparedwith non-psychiatric controls in anterior and mid DG.This is in agreement with our findings in smaller samplesof untreated major depression (Boldrini et al., 2009, 2012).
50µm
Fig. 4. Stem cells and neural progenitor cells in the humandentate gyrus. Stem cells, labeled with antibody against glialfibrillary acid protein (GFAP, in red) were seen in the humandentate gyrus (yellow arrow). They show the typical apicaldendrite crossing the granule cell layer (GCL) typical of stemcells, or type 1 progenitors. Neural progenitor cells, or type 2progenitors, double labeled with nestin (green) and GFAP (inorange, cyan arrow) and were found in the subgranular zone(SGZ); they are the cells that we find proliferating in thehuman brain in adulthood. GFAP labeling (in red) was alsofound in astrocytes (white arrow), as expected. Cell nuclei werestained with DAPI (in blue).
Benzodiazepines and hippocampal neuroplasticity 7

Ablation of neurogenesis in mice precipitates behavioraldespair and anhedonia without exposing mice to anybehavioral stress (Snyder et al., 2011). Of note, wefound the mean NPC number in anterior DG of MDwas one seventh that of controls, but the difference wasnot statistically significant owing to the low number ofNPCs in both groups and the variance in counts.Alternatively, as stress does induce a form of depressionin mice (Santarelli et al., 2003; Surget et al., 2008), anNPC deficit in mood disorders may be present only in aphenotype that involves stress exposure.
Antidepressant-alone treated MD had higher NPCnumber in the mid DG compared with controls anduntreated MD. Our results are consistent with rodent(Malberg et al., 2000), non-human primate (Perera et al.,2011), and our previous human studies (Boldrini et al.,2009, 2012), which suggest that antidepressants increaseamplifying or type 2 NPCs (Encinas et al., 2006).
Mitotic cells
MD subjects co-treated with antidepressants and BZD didnot differ from antidepressants-alone treated MD in
mitotic cell number, and both treatment groups hadmore mitotic cells compared with untreated MD and con-trols in anterior DG. Thus we do not observe an effect ofBZD co-treatment on mitotic cells stained with Ki-67, butnote that these cells include glial and neuronal lineages.Moreover, the effects of BZDs on mitotic cells in rodentsare complex, depending on duration of exposure. Short-term exposure increases proliferation, whereas long-termexposure reduces overall neurogenesis (Zhao et al., 2012).
Consistent with our previous findings (Boldrini et al.,2009), in this larger sample, although the mean numberof mitotic cells in untreated MD subjects is about a halfthat in controls, the difference did not reach statisticalsignificance.
Effects of GABA-mediated signaling on hippocampalneurogenesis
A BZD effect on neurogenesis may be mediated by theirallosteric effect on the GABA receptor. Type-2 progenitorsin adult hippocampus express GABAA receptors that canbe activated by synaptic stimuli and stimulate differen-tiation (Deisseroth and Malenka, 2005; Wang et al., 2005).
Den
tate
gyr
us m
itotic
cel
l cou
nt 160000 ControlsMDMD*ADTMD*ADT*BDZ
140000
120000
100000
80000
60000
40000
20000
0Head Mid Tail
Rostrocaudal axis of the dentate gyrus
p=0.032
p=0.046
(p=0.056)
(p=0.063)
(a) (b)
(c)
Fig. 5. Mitotic cells in the human hippocampus. (a) Mitotic cells visualization with monoclonal antibody against the nuclearproliferation antigen Ki-67. This antigen is present in the nuclei of cells in the G1, S, and G2 phases of the cell division cycle as wellas in mitosis, whereas quiescent or resting cells in the G0 phase do not express Ki-67 (Scholzen and Gerdes, 2000). The greenoutline defines the region of interest for cell counting with stereology. The granule cell layer (GCL), subgranular zone (SGZ),molecular layer (ML) and hilus are indicated. (b) Ki-67 immunoreactive cells with stained nuclei (in black) are found in the innerportion of the GCL of the dentate gyrus and in the SGZ. (c) Ki-67-immunoreactive mitotic cell number in the dentate gyrus (DG).Non-parametric statistical analysis shows that antidepressant-treated subjects with mood disorders (MD*ADT) had more mitoticcells compared with untreated subjects (MD) in the anterior DG. Subjects treated with antidepressants and benzodiazepines(MD*ADT*BZD) had more mitotic cells compared with subjects with no psychopathology or treatment (Controls) in the anteriorDG. Mitotic cell number in MD*ADT*BZD did not differ from untreated MD*ADT. Untreated MD showed a trend to fewer mitoticcells than Controls in anterior DG. No between-groups differences in mitotic cell number were found in mid and posterior DG.
8 M. Boldrini et al.

Antiepileptic drugs, such as the barbiturate phenobar-bital, which also act on the GABAergic system by bindingto GABAA receptors (Rudolph et al., 1999), decreaseneurogenesis by about 60% in the DG (Chen et al.,2009) and have cognitive effects that persist after drugcessation (Chen et al., 2009). Alcohol, which, likeBZDs, modulates GABAA receptors to induce sedation(Laukkanen et al., 2013), decreases DG neurogenesis byup to 50% in adult rats after long-term exposure (Janget al., 2002; Nixon and Crews, 2002; Rice et al., 2004).
The reduction in hippocampal neurogenesis and thedepression-like responses induced by chronic alcoholself-administration in mice are reversible by fluoxetine(Stevenson et al., 2009). Whether treatment with BDZalone would be associated with an apparent detrimentaleffect on hippocampal neurogenesis or granule cell num-ber relative to untreated depression should be investi-gated in future studies.
Increased GABA activation via GABAA receptor ago-nists, such as BZD, appears to cause neuronal apoptosisin the developing brain of rodents (Chen et al., 2009).We found that mature granule neurons were fewer insubjects co-treated with BZD compared with subjectstreated with antidepressant alone, and in BZD co-treatedMD were comparable to untreated MD. Based on rodentstudies, our findings may be partly the result of increasedapoptosis.
Clinical relevance of hippocampal neuroplasticity
We found that cell number changes associated with MDand with treatment occur selectively in the anterior andmid DG and are not detected in posterior DG. Theposterior primate hippocampus, and its homolog in therodent, the dorsal hippocampus, are mainly implicatedin memory (Risold and Swanson, 1996; Bannermanet al., 2004), whereas the anterior hippocampus inprimates projects to prefrontal cortex and amygdala(Thierry et al., 2000), and participates in emotional regu-lation (Nettles et al., 2000; Sahay and Hen, 2007) and pat-tern discrimination (Clelland et al., 2009; Aimone et al.,2011). Our results are consistent with a functional re-lationship between anterior and mid DG NPC and gran-ule neuron number and indicate that mood disorders ortreatment effects may impact emotional responses andpattern discrimination via the anterior hippocampus.
We found fewer mature DG neurons correlated withworse global clinical functioning, earlier onset of majordepressive disorder, and more lifetime major depressiveepisodes, indicating the potential clinical relevance ofDG cell viability in major depression (Boldrini et al.,2013). Adult neurogenesis, like developmental neurogen-esis, in non-human primates is about five times less thanin mice (Kohler et al., 2011). Human levels of neurogen-esis decline modestly during aging (Spalding et al.,2013), but our MD treated and untreated groups didnot differ in age, thus age does not explain our findings.
The proportion of subjects who died by suicide was notdifferent between the MD groups, and so suicide as acause of death did not explain our treatment groupfindings. Of note, clinician-rated severity of depressionis weakly related to risk for suicidal behavior (Mannet al., 1999) and therefore, suicide is not considered a use-ful measure of depression severity. The pathophysiologyof suicide, as opposed to depression, is related to the di-athesis for suicidal behavior (Mann et al., 1999). What ispuzzling is why patients with robust increase in neuro-genesis still die by suicide. Perhaps neurogenesis aloneis insufficient protection from suicide, and other mechan-isms and pathways lead to suicide. Alternatively, an in-termediate level of improvement in depression andneurogenesis, while suicidal ideation persists, may pro-vide an increase in drive and effectiveness to overcomethe helplessness and inertia of depression and act onsuicidal thoughts.
Study limitations
The sample included major depressive disorder andbipolar disorder. It is not known whether the two dis-orders differ in hippocampal DG morphometry. Themood disorder-treated group included subjects treatedwith lithium, SSRIs, and tricyclic antidepressants.Larger sample sizes would allow for testing the effect ofBZD co-treatment with different classes of antidepres-sants and lithium. Subjects co-treated with BZD may beanxious compared with MD not prescribed BZD. The re-lationship between anxiety and neurogenesis remainsunclear, and excessive anxiety might inhibit neurogenesisinstead of the BZD used to treat it and thereby could ex-plain our results.
Animal studies and an expanded human study maybegin to further address these questions which are of criti-cal importance to doctors and patients trying to decidewhether to use a benzodiazepine for treatment of anxietysymptoms in major depression.
Acknowledgments
Supported by MH83862, MH94888, MH64168, MH40210,the American Foundation for Suicide Prevention and theDiane Goldberg Foundation.
Statement of Interest
Maura Boldrini, Tanya H Butt, Adrienne N. Santiago,Andrew Dwork, Gorazd Rosoklija, Hadassah Tamir,and Victoria Arango declare that, except for incomereceived from their primary employer, no financial sup-port or compensation has been received from any individ-ual or corporate entity over the past three years forresearch or professional services and there are no per-sonal financial holdings that could be perceived as consti-tuting a potential conflict of interest. Dr Andrew J. Dwork
Benzodiazepines and hippocampal neuroplasticity 9

received loans and gifts of equipment and software fromOlympus and Visiopharm for research unrelated to thisstudy. Dr Rene´ Hen receives compensation as a consult-ant for Roche and Lundbeck. Dr J. John Mann receivedpast unrelated grants from GlaxoSmithKline andNovartis.
References
Aimone JB, Deng W, Gage FH (2011) Resolving new memories: acritical look at the dentate gyrus, adult neurogenesis, andpattern separation. Neuron 70:589–596.
Bandelow B, Zohar J, Hollander E, Kasper S, Möller HJ; WFSBPTask Force on Treatment Guidelines for Anxiety, Obsessive-Compulsive and Post-Traumatic Stress Disoders, Zohar J,Hollander E, Kasper S, Moller HJ, Allgulander C, Ayuso-Gutierrez J, Baldwin DS, Buenvicius R, Cassano G, Fineberg N,Gabriels, Hindmarch I, Kaiya H, Klein DF, Lader M,Lecrubier Y, Lépine JP, Liebowitz MR, Lopez-Ibor JJ,Marazziti D, Miguel EC, Oh KS, Preter M, Rupprecht R,Sato M, Starcevic V, Stein DJ, van Ameringen M, Vega J (2008)World Federation of Societies of Biological Psychiatry (WFSBP)guidelines for the pharmacological treatment of anxiety,obsessive-compulsive and post-traumatic stress disorders -first revision. World J Biol Psychiatry 9:248–312.
Bannerman DM, Rawlins JN, McHugh SB, Deacon RM, Yee BK,Bast T, Zhang WN, Pothuizen HH, Feldon J (2004) Regionaldissociations within the hippocampus–memory and anxiety.Neurosci Biobehav Rev 28:273–283.
Boldrini M, Underwood MD, Hen R, Rosoklija GB, Dwork AJ,John MJ, Arango V (2009) Antidepressants increase neuralprogenitor cells in the human hippocampus.Neuropsychopharmacology 34:2376–2389.
Boldrini M, Hen R, Underwood M, Rosoklija G, Dwork A,Mann JVA (2012) Hippocampal Angiogenesis and progenitorcell proliferation are increased with antidepressant use inmajor depression. Biol Psychiatry 75:562–571.
Boldrini M, Santiago AN, Hen R, Dwork AJ, Rosoklija GB,Tamir H, Arango V, John Mann J (2013) Hippocampal granuleneuron number and dentate gyrus volume inantidepressant-treated and untreated major depression.Neuropsychopharmacology 38:1068–1077.
Brook GA, Perez-Bouza A, Noth J, Nacimiento W (1999)Astrocytes re-express nestin in deafferented target territories ofthe adult rat hippocampus. Neuroreport 10:1007–1011.
Chen J, Cai F, Cao J, Zhang X, Li S (2009) Long-term antiepilepticdrug administration during early life inhibits hippocampalneurogenesis in the developing brain. J Neurosci Res87:2898–2907.
Clelland CD, Choi M, Romberg C, Clemenson GD Jr.,Fragniere A, Tyers P, Jessberger S, Saksida LM, Barker RA,Gage FH, Bussey TJ (2009) A functional role for adulthippocampal neurogenesis in spatial pattern separation.Science 325:210–213.
David DJ, Samuels BA, Rainer Q, Wang JW, Marsteller D,Mendez I, Drew M, Craig DA, Guiard BP, Guilloux JP,Artymyshyn RP, Gardier AM, Gerald C, Antonijevic IA,Leonardo ED, Hen R (2009) Neurogenesis-dependent and-independent effects of fluoxetine in an animal model ofanxiety/depression. Neuron 62:479–493.
Deisseroth K, Malenka RC (2005) GABA excitation in the adultbrain: a mechanism for excitation- neurogenesis coupling.Neuron 47:775–777.
Dell’osso B, Lader M (2013) Do benzodiazepines still deserve amajor role in the treatment of psychiatric disorders? A criticalreappraisal. Eur Psychiatry 28:7–20.
Duggal N, Schmidt-Kastner R, Hakim AM (1997) Nestinexpression in reactive astrocytes following focal cerebralischemia in rats. Brain Res 768:1–9.
Edwards JG, Anderson I (1999) Systematic review and guide toselection of selective serotonin reuptake inhibitors. Drugs57:507–533.
Encinas JM, Vaahtokari A, Enikolopov G (2006) Fluoxetinetargets early progenitor cells in the adult brain. Proc Natl AcadSci USA 103:8233–8238.
Endicott J, Spitzer RL, Fleiss JL, Cohen J (1976) The globalassessment scale. A procedure for measuring overall severityof psychiatric disturbance. Arch Gen Psychiatry 33:766–771.
Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM,Nordborg C, Peterson DA, Gage FH (1998) Neurogenesis inthe adult human hippocampus. Nat Med 4:1313–1317.
Ge S, Goh EL, Sailor KA, Kitabatake Y, Ming GL, Song H (2006)GABA regulates synaptic integration of newly generatedneurons in the adult brain. Nature 439:589–593.
Itoh T, Satou T, Nishida S, Hashimoto S, Ito H (2006) Cultured ratastrocytes give rise to neural stem cells. Neurochem Res31:1381–1387.
Jang MH, Shin MC, Kim EH, Kim CJ (2002) Acute alcoholintoxication decreases cell proliferation and nitric oxidesynthase expression in dentate gyrus of rats. Toxicol Lett133:255–262.
Kalman M, Ajtai BM (2001) A comparison of intermediatefilament markers for presumptive astroglia in the developingrat neocortex: immunostaining against nestin reveals moredetail, than GFAP or vimentin. Int J Dev Neurosci 19:101–108.
Kelly TM, Mann JJ (1996) Validity of DSM-III-R diagnosis bypsychological autopsy: a comparison with clinicianante-mortem diagnosis. Acta Psychiatr Scand 94:337–343.
Kempermann G, Kronenberg G (2003) Depressed new neurons–adult hippocampal neurogenesis and a cellular plasticityhypothesis of major depression. Biol Psychiatry 54:499–503.
Klein DN, Arnow BA, Barkin JL, Dowling F, Kocsis JH, Leon AC,Manber R, Rothbaum BO, Trivedi MH, Wisniewski SR (2009)Early adversity in chronic depression: clinical correlates andresponse to pharmacotherapy. Depress Anxiety 26:701–710.
Kohler SJ, Williams NI, Stanton GB, Cameron JL, Greenough WT(2011) Maturation time of new granule cells in the dentategyrus of adult macaque monkeys exceeds six months. ProcNatl Acad Sci USA 108:10326–10331.
Lader M (2011) Benzodiazepines revisited–will we ever learn?Addiction 106:2086–2109.
Laukkanen V, Storvik M, Hakkinen M, Akamine Y, Tupala E,Virkkunen M, Tiihonen J (2013) Decreased GABA(A)benzodiazepine binding site densities in postmortem brains ofCloninger type 1 and 2 alcoholics. Alcohol 47:103–108.
Lobbestael J, Leurgans M, Arntz A (2011) Inter-Rater reliability ofthe structured clinical interview for DSM-IV axis I disorders(SCID I) and axis II disorders (SCID II). Clin Psychol Psychot18:75–79.
Malberg JE, Eisch AJ, Nestler EJ, Duman RS (2000) Chronicantidepressant treatment increases neurogenesis in adult rathippocampus. J Neurosci 20:9104–9110.
10 M. Boldrini et al.

Mann JJ, Waternaux C, Haas GL, Malone KM (1999) Towarda clinical model of suicidal behavior in psychiatric patients.Am J Psychiatry 156:181–189.
McKinnon MC, Yucel K, Nazarov A, MacQueen GM (2009)A meta-analysis examining clinical predictors of hippocampalvolume in patients with major depressive disorder.J Psychiatry Neurosci 34:41–54.
Mokry J, Ehrmann J, Karbanova J, Cizkova D, Soukup T,Suchanek J, Filip S, Kolar Z (2008) Expression of intermediatefilament nestin in blood vessels of neural and non-neuraltissues. Acta Medica (Hradec Kralove) 51:173–179.
Nettles KW, Pesold C, Goldman MB (2000) Influence of theventral hippocampal formation on plasma vasopressin,hypothalamic-pituitary-adrenal axis, and behavioral responsesto novel acoustic stress. Brain Res 858:181–190.
Nixon K, Crews FT (2002) Binge ethanol exposure decreasesneurogenesis in adult rat hippocampus. J Neurochem83:1087–1093.
Perera TD, Dwork AJ, Keegan KA, Thirumangalakudi L,Lipira CM, Joyce N, Lange C, Higley JD, Rosoklija G, Hen R,Sackeim HA, Coplan JD (2011) Necessity of hippocampalneurogenesis for the therapeutic action of antidepressants inadult nonhuman primates. PLoS ONE 6:e17600.
Rice AC, Bullock MR, Shelton KL (2004) Chronic ethanolconsumption transiently reduces adult neural progenitor cellproliferation. Brain Res 1011:94–98.
Risold PY, Swanson LW (1996) Structural evidence for functionaldomains in the rat hippocampus. Science 272:1484–1486.
Rudolph U, Crestani F, Benke D, Brünig I, Benson JA,Fritschy J-M, Martin JR, Bluethmann H, Möhler H (1999)Benzodiazepine actions mediated by specific γ-aminobutyricacidA receptor subtypes. Nature 401:796–800.
Sahay A, Hen R (2007) Adult hippocampal neurogenesis indepression. Nat Neurosci 10:1110–1115.
Sahay A, Scobie KN, Hill AS, O’Carroll CM, Kheirbek MA,Burghardt NS, Fenton AA, Dranovsky A, Hen R (2011)Increasing adult hippocampal neurogenesis is sufficient toimprove pattern separation. Nature 472:466–470.
Sahin KS, Mahmood A, Li Y, Yavuz E, Chopp M (1999)Expression of nestin after traumatic brain injury in rat brain.Brain Res 840:153–157.
Salehi F, Kovacs K, Cusimano MD, Horvath E, Bell CD,Rotondo F, Scheithauer BW (2008) Immunohistochemicalexpression of nestin in adenohypophysial vessels duringdevelopment of pituitary infarction. J Neurosurg 108:118–123.
Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S,Weisstaub N, Lee J, Duman R, Arancio O, Belzung C, Hen R(2003) Requirement of hippocampal neurogenesis for thebehavioral effects of antidepressants. Science 301:805–809.
Saravia F, Beauquis J, Pietranera L, De Nicola AF (2007)Neuroprotective effects of estradiol in hippocampal neuronsand glia of middle age mice. Psychoneuroendocrinology32:480–492.
Saxe MD, Battaglia F, Wang JW, Malleret G, David DJ,Monckton JE, Garcia AD, Sofroniew MV, Kandel ER,Santarelli L, Hen R, Drew MR (2006) Ablation of hippocampalneurogenesis impairs contextual fear conditioning andsynaptic plasticity in the dentate gyrus. Proc Natl Acad SciUSA 103:17501–17506.
Schloesser RJ, Lehmann M, Martinowich K, Manji HK,Herkenham M (2010) Environmental enrichment requires
adult neurogenesis to facilitate the recovery from psychosocialstress. Mol Psychiatry 15:1152–1163.
Scholzen T, Gerdes J (2000) The Ki-67 protein: from the knownand the unknown. J Cell Physiol 182:311–322.
Sheline YI, Gado MH, Kraemer HC (2003) Untreated depressionand hippocampal volume loss. Am J Psychiatry 160:1516–1518.
Snyder JS, Soumier A, Brewer M, Pickel J, Cameron HA (2011)Adult hippocampal neurogenesis buffers stress responses anddepressive behaviour. Nature 476:458–461.
Spalding KL, Bergmann O, Alkass K, Bernard S, Salehpour M,Huttner HB, Bostrom E, Westerlund I, Vial C, Buchholz BA,Possnert G, Mash DC, Druid H, Frisen J (2013) Dynamicsof hippocampal neurogenesis in adult humans. Cell153:1219–1227.
Stevenson JR, Schroeder JP, Nixon K, Besheer J, Crews FT,Hodge CW (2009) Abstinence following alcohol drinkingproduces depression-like behavior and reduced hippocampalneurogenesis in mice. Neuropsychopharmacology34:1209–1222.
Sun B, Halabisky B, Zhou Y, Palop JJ, Yu G, Mucke L, Gan L(2009) Imbalance between GABAergic and Glutamatergictransmission impairs adult neurogenesis in an animal model ofAlzheimer’s disease. Cell Stem Cell 5:624–633.
Sun Y, Evans J, Russell B, Kydd R, Connor B (2013) Abenzodiazepine impairs the neurogenic and behaviouraleffects of fluoxetine in a rodent model of chronic stress.Neuropharmacology 72:20–28.
Surget A, Saxe M, Leman S, Ibarguen-Vargas Y, Chalon S,Griebel G, Hen R, Belzung C (2008) Drug-dependentrequirement of hippocampal neurogenesis in a model ofdepression and of antidepressant reversal. Biol Psychiatry64:293–301.
Thierry AM, Gioanni Y, Degenetais E, Glowinski J (2000)Hippocampo-prefrontal cortex pathway: anatomical andelectrophysiological characteristics. Hippocampus 10:411–419.
Tronel S, Belnoue L, Grosjean N, Revest JM, Piazza PV,Koehl M, Abrous DN (2012) Adult-born neurons are necessaryfor extended contextual discrimination. Hippocampus22:292–298.
Wang LP, Kempermann G, Kettenmann H (2005) Asubpopulation of precursor cells in the mouse dentate gyrusreceives synaptic GABAergic input. Mol Cell Neurosci29:181–189.
Wu CS, Wang SC, Chang IS, Lin KM (2009) The associationbetween dementia and long-term use of benzodiazepine in theelderly: nested case-control study using claims data. Am JGeriatr Psychiatry 17:614–620.
Wu X, Castren E (2009) Co-treatment with diazepam preventsthe effects of fluoxetine on the proliferation and survival ofhippocampal dentate granule cells. Biol Psychiatry 66:5–8.
Yamaguchi M, Saito H, Suzuki M, Mori K (2000) Visualization ofneurogenesis in the central nervous system using nestinpromoter-GFP transgenic mice. Neuroreport 11:1991–1996.
Yoo YM, Lee U, Kim YJ (2005) Apoptosis and nestin expressionin the cortex and cultured astrocytes following 6-OHDAadministration. Neurosci Lett 382:88–92.
Zhao Y, Wang Z, Dai J, Chen L, Huang Y, Zhan Z (2012)Beneficial effects of benzodiazepine diazepam on chronicstress-induced impairment of hippocampal structuralplasticity and depression-like behavior in mice. Behav BrainRes 228:339–350.
Benzodiazepines and hippocampal neuroplasticity 11