babesia microti: prevalence in wild rodents and ixodes ricinus ticks from the mazury lakes district...
TRANSCRIPT

ARTICLE IN PRESS
1438-4221/$ - se
doi:10.1016/j.ijm
�CorrespondE-mail addr
International Journal of Medical Microbiology 296 (2006) S1, 137–143
www.elsevier.de/ijmm
Babesia microti: Prevalence in wild rodents and Ixodes ricinus ticks from
the Mazury Lakes District of north-eastern Poland
Edward Sinskia,�, Anna Bajera, Renata Welca, Agnieszka Pawe"czykb,Maria Ogrzewalskaa, Jerzy M. Behnkec
aDepartment of Parasitology, Institute of Zoology, Warsaw University, Miecznikowa 1 Street, PL-02096 Warsaw, PolandbDepartment of Immunopathology, Institute of Infection Diseases, Medical University of Warsaw, Warsaw, PolandcSchool of Biology, University Park, University of Nottingham, Nottingham, UK
Abstract
Infections of Babesia microti (Apicomplexa, Piroplasmida), a common erythroparasitic protozoon of Holarcticrodents, are not widely acknowledged in Poland. The presence of this parasite in various species of wild rodents hasbeen well documented throughout the northern temperate zone of North America, Europe, and Eurasia. However,human babesiosis attributable to infection with B. microti has been reported only from the north-eastern and uppermidwestern United States and Japan. We recently carried out an epizootiological survey investigating the prevalence ofB. microti both in the tick Ixodes ricinus and in wild rodents in north-eastern Poland. Blood samples were collectedfrom a total of 483 animals comprising three species: Apodemus flavicollis, Microtus arvalis, and Microtus oeconomus
trapped at Urwita"t near Miko"ajki in the Mazury Lakes District. Questing adult I. ricinus ticks were collected in thestudy sites by blanket dragging of vegetation in heterogeneous, deciduous woodland, and, in addition, rodents werecarefully examined for feeding larvae and nymphs. Altogether, B. microti was detected in 9 out of 1513 I. ricinus ticks(0.6%) examined by PCR. This included 163 adults (92 females and 71 males), 50 nymphs, and 1300 larvae 3%, 8%,and 0% of which were PCR-positive, respectively. Of 85 A. flavicollis, 374 M. arvalis and 24 M. oeconomus, 1%,12.8%, and 42% were parasitaemic, respectively, as determined by microscopic examination of blood smears stainedwith Giemsa. B. microti DNA, extracted from 53 M. arvalis and 5 M. oeconomus and examined by nested PCR,targeting a piroplasm-specific portion of the 18S ribosomal DNA, revealed 72% and 40%, respectively, to be PCRpositive. Sequence analysis showed that all PCR-positive samples had rDNA sequences identical (100% homology) tothat of the Munich B. microti strain (GenBank Accession no. AB071177) isolated from Mus musculus. The results ofthis study indicate that the B. microti commonly encountered among Microtus spp. rodents is probably not a zoonoticstrain and, therefore, that it is most unlikely to represent a risk to public health in the Mazury Lakes District of north-eastern Poland.r 2006 Elsevier GmbH. All rights reserved.
Keywords: Babesia microti; Ixodes ricinus; Rodents; Poland
e front matter r 2006 Elsevier GmbH. All rights reserved.
m.2006.01.015
ing author. Tel.: +4822 554 1113.
ess: [email protected] (E. Sinski).
Introduction
Babesia microti (Apicomplexa, Piroplasmida), anintra-erythrocytic, tick-transmitted haemoprotozoan, is

ARTICLE IN PRESSE. Sinski et al. / International Journal of Medical Microbiology 296 (2006) S1, 137–143138
a common parasite of Holarctic small wild rodents. Thepresence of this parasite in various rodent species hasbeen documented throughout the northern temperatezones of North America (Steketee et al., 1985; Watkinset al., 1991; Burkot et al., 2000), Europe (Sebek et al.,1977; Healing, 1981; Karbowiak and Sinski, 1996; Bajeret al., 2001; Pawe"czyk et al., 2004), and Eurasia (Shihet al., 1997; Tsuji et al., 2001). Human babesiosiscaused by B. microti is gaining increasing interest as anemerging zoonosis (Homer et al., 2000). Particularly, inNorth America most human cases of babesiosis are dueto the rodent parasite B. microti; however, a newlyrecognized species called the WA1 piroplasm may alsobe responsible for this disease (Dammin et al., 1981;Persing et al., 1995). However, in Europe, the 31confirmed cases of human babesiosis were mainlycaused by Babesia divergens (Kjemtrup and Conrad,2000). Moreover, quite recently a new pathogenicB. microti-like species has been described from dogs inGermany (Zahler et al., 2000) and a non-B. divergens
piroplasm (EU1), causing two human cases of babesio-sis, has been reported in Italy and Austria (Herwaldtet al., 2003).
In Europe, B. microti is transmitted by ticks of the‘Ixodes persulcatus complex’, predominantly by nymphsof I. ricinus, which appear to be vectors of B. microti
(Gray et al., 2002). Contrary to the earlier belief thatI. ricinus is infected with B. microti relatively infre-quently (Walter, 1981), under the natural conditions ofthe middle-European biotopes the carriers and mostlikely vectors of B. microti are Ixodes trianguliceps
(Randolph, 1995) and I. ricinus (Duh et al., 2001). Ourprevious studies showed that in the study area subadultI. ricinus abundantly feed on Clethrionomys glareolus,Apodemus flavicollis, and Microtus arvalis, some ofwhich are parasitaemic, with chronic infections thatevidently persist for several weeks at a time (Sinski andKarbowiak, 1995; Sinski, 1999; Bajer et al., 2001;Pawe"czyk et al., 2004). However, recent studies indicatethat B. microti DNA may be detected within questingI. ricinus ticks (Duh et al., 2001; Skotarczak andCichocka, 2001; Foppa et al., 2002; Stanczak et al.,2004) but molecular evidence also points to B. microti
comprising a diverse species complex in wild rodentpopulations. There appear to be at least two clades, onerepresenting zoonotic isolates that are transmissible tohumans, and the other apparently maintained only inMicrotus spp. (Goethert and Telford, 2003).
In the Mazury Lakes District of north-eastern Po-land, sympatric populations of wild rodents andI. ricinus ticks are extremely abundant but the zoonoticpotential of B. microti in Poland is still unresolved.Therefore, the present study was undertaken to assessthe prevalence of B. microti in both rodents and ticks toascertain whether the genotypes involved are thoseassociated with infections in humans.
Materials and methods
Field studies
Small wild rodents were studied mostly in heterogeneous,
deciduous woodland and fallow land at Urwita"t near
Miko"ajki in the Mazury Lakes District, north-eastern Poland,
as part of long-term epizootiologic surveys in the region.
Rodents were live trapped in wooden traps which were
inspected twice daily and processed according to the proce-
dures described in detail by Bajer et al. (2001). In the
laboratory, at the field station in Urwita"t, each trapped
rodent was marked, identified to species level and sexed,
weighed, and examined for ectoparasites, mostly for feeding
tick larvae and nymphs. In the same study sites, questing ticks
were collected by blanket dragging of vegetation in hetero-
geneous, deciduous woodland.
Prevalence and morphology of Babesia
Thin blood smears were prepared using blood from the tip
of the tail from a total of 483 A. flavicollis, M. arvalis, and
M. oeconomus. Some animals were anaesthetized with ether
and their blood was immediately collected by heart puncture,
using a heparinized syringe. Blood smears were air-dried, fixed
in absolute methanol, and stained with Giemsa at pH 7.1.
Smears were viewed at 1000� magnification under oil
immersion using an Olympus AX70 microscope. Approxi-
mately 200 fields of vision were inspected and the number of
red blood cells (RBC) infected with Babesia-like organisms
were recorded and expressed as number of infected cells/100
RBC.
Analysis of rDNA sequences
Genomic DNA was prepared from frozen blood samples of
53 M. arvalis and 5 M. oeconomus as well as from EDTA-
stabilized whole blood from BALB/c mice experimentally
infected with the reference strain (King’s 67 BALB/c mice
strain) of B. microti using a whole-blood DNA extraction kit,
Genomic Mini AX Blood (A&A Biotechnology, Gdansk,
Poland). To determine whether B. microti-specific DNA was
present, the extracted DNA was subjected to a nested PCR
reaction with outer primers Bab1 and Bab4 and inner primers
Bab2 and Bab3, targeting a specific fragment of the gene
encoding the nuclear small-subunit ribosomal RNA (18S
rDNA). The primer sets amplify 238 base pairs (bp) and
154 bp fragments, respectively (Persing et al., 1992). After
denaturation for 2min at 94 1C, 40 cycles were performed, with
45 s at 94 1C, 45 s at 55 1C, and 45 s at 72 1C, followed by a
7min final extension. PCR products were subjected to
electrophoresis on a 2% agarose gel, stained with ethidium
bromide, and photographed. PCR products were purified by
using the QIAquicks PCR Purification Kit (Qiagen). Sequenc-
ing reactions were conducted with the ABI-PRISM 377
automatic DNA sequencer (Applied Biosystems). The result-
ing sequences were assembled by using the program ABI
PrismTM BigDyeTM and compared with specimens deposited
in GenBank using the Blast-NCBI program (Table 1).
ARTICLE IN PRESS
Table 1. Babesia microti 18S rRNA gene homology (Blast-NCBI program) of selected isolates estimated from GenBank sequences
with reference to the isolates from Microtus arvalis (GenBank Accession no. AY918165) and M. oeconomus (GenBank Accession no.
AY918164)
Species (strain)a Accession no. Base pairs compared % homology Source
B. microti (Munich) AB071177 131 100 Mus musculusb
B. microti (GI) AF231348 131 98.19 Human
B. microti (unknown) AF373331 131 98.16 Ixodes ricinus
B. microti (Gray) AY693840 131 98.05 Human
B. microti (Xinjiang 1647) AB083375 131 97.97 Lagurus luteus
B. microti (Otsu) AB119446 131 97.91 Apodemus sp.
B. microti (Hobetsu) AB050732 131 97.91 Apodemus sp.
B. microti (HK) AB085191 131 97.83 Clethrionomys glareolus
B. microti (Ruebosh Peabody) U09833 131 97.74 Mus musculusb
B. microti (Kobe524) AB032434 131 97.71 Human
B. microti (RcMS201, Meisham) AB112050 131 97.70 Rattus coxinga
B. microti (Berlin) AF231349 131 97.61 Ixodes ricinus
B. divergens (unknown) AY572456 71 88.73 Rangifer tarandus
aThe strains cited refer to the GenBank database, but not to any publications.bLaboratory host.
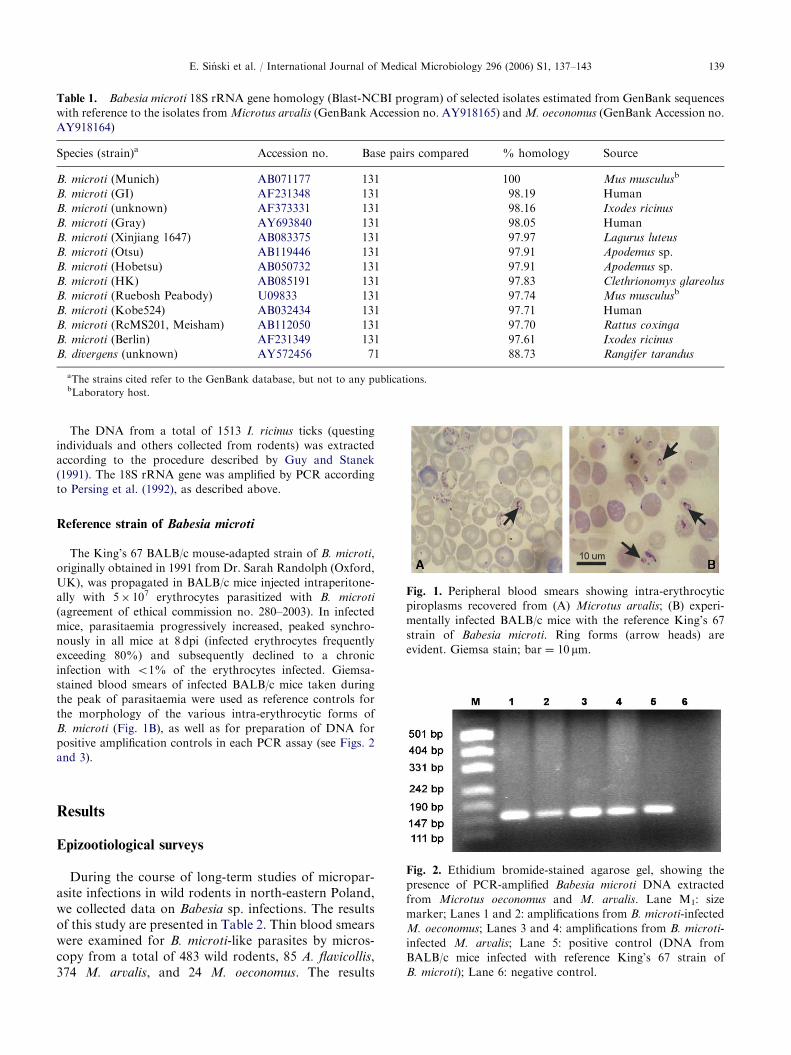
Fig. 1. Peripheral blood smears showing intra-erythrocytic
piroplasms recovered from (A) Microtus arvalis; (B) experi-
mentally infected BALB/c mice with the reference King’s 67
strain of Babesia microti. Ring forms (arrow heads) are
evident. Giemsa stain; bar ¼ 10 mm.
E. Sinski et al. / International Journal of Medical Microbiology 296 (2006) S1, 137–143 139
The DNA from a total of 1513 I. ricinus ticks (questing
individuals and others collected from rodents) was extracted
according to the procedure described by Guy and Stanek
(1991). The 18S rRNA gene was amplified by PCR according
to Persing et al. (1992), as described above.
Reference strain of Babesia microti
The King’s 67 BALB/c mouse-adapted strain of B. microti,
originally obtained in 1991 from Dr. Sarah Randolph (Oxford,
UK), was propagated in BALB/c mice injected intraperitone-
ally with 5� 107 erythrocytes parasitized with B. microti
(agreement of ethical commission no. 280–2003). In infected
mice, parasitaemia progressively increased, peaked synchro-
nously in all mice at 8 dpi (infected erythrocytes frequently
exceeding 80%) and subsequently declined to a chronic
infection with o1% of the erythrocytes infected. Giemsa-
stained blood smears of infected BALB/c mice taken during
the peak of parasitaemia were used as reference controls for
the morphology of the various intra-erythrocytic forms of
B. microti (Fig. 1B), as well as for preparation of DNA for
positive amplification controls in each PCR assay (see Figs. 2
and 3).
Fig. 2. Ethidium bromide-stained agarose gel, showing the
presence of PCR-amplified Babesia microti DNA extracted
from Microtus oeconomus and M. arvalis. Lane M1: size
marker; Lanes 1 and 2: amplifications from B. microti-infected
M. oeconomus; Lanes 3 and 4: amplifications from B. microti-
infected M. arvalis; Lane 5: positive control (DNA from
BALB/c mice infected with reference King’s 67 strain of
B. microti); Lane 6: negative control.
Results
Epizootiological surveys
During the course of long-term studies of micropar-asite infections in wild rodents in north-eastern Poland,we collected data on Babesia sp. infections. The resultsof this study are presented in Table 2. Thin blood smearswere examined for B. microti-like parasites by micros-copy from a total of 483 wild rodents, 85 A. flavicollis,374 M. arvalis, and 24 M. oeconomus. The results

ARTICLE IN PRESSE. Sinski et al. / International Journal of Medical Microbiology 296 (2006) S1, 137–143140
indicate that prevalence of infection in A. flavicollis wasrather low with 1% of the sampled population. Incontrast, prevalence of infection in populations of thetwo microtine rodents was much higher with 12.8% and42% for M. arvalis and M. oeconomus, respectively. Theparasites examined in field surveys were mostly ring-and pear-shaped forms, 1.5–3.0 mm in diameter andmorphologically similar to the King’s 67 BALB/cmouse-adapted strain of B. microti (see Fig. 1A andB). A tetrad (Maltese cross form) was observed onlyvery rarely.
Fig. 3. Ethidium bromide-stained agarose gel, showing the
presence of nested PCR-amplified Babesia microti DNA
extracted from nymphs of Ixodes ricinus. Lane M1: size
marker; Lane 1: negative control; Lane 2: positive control
(DNA from BALB/c mice infected with reference King’s 67
strain of B. microti); Lane 3: amplification from B. microti-
infected nymph of I. ricinus.
Table 2. Levels of Babesia microti infection among wild rodents in
Host species Study period No. of positive ticks
(microscopy, x/n)
Apodemus flavicollis 2001 1/85
Microtus arvalis 2000 48/374
Microtus oeconomus 2000–2004 10/24
Total 59/483
Molecular analysis of Babesia isolates from
microtine rodents
B. microti DNA was extracted from 53 M. arvalis and5 M. oeconomus and examined by nested PCR, targetinga piroplasm-specific portion of the 18S ribosomal DNA(Persing et al., 1992). This analysis revealed that 72%and 40% of the two vole species, respectively, werePCR-positive. Sequencing analysis showed that allPCR-positive samples of isolates from M. arvalis
(GenBank Accession no. AY918165) and fromM. oeconomus (GenBank Accession no. AY918164)had rDNA sequences identical (100% homology) to thatof the Munich B. microti strain (GenBank Accession no.AB071177) isolated originally from Mus musculus. Gelelectrophoresis of nested PCR-amplified B. microti
DNA is presented in Fig. 2. Significant differences inthe rDNA sequences to other B. microti isolates orBabesia species are shown in Table 1.
Tick survey
The prevalence of B. microti infection in I. ricinus
ticks from the study sites was assessed by nested PCR.Altogether, B. microti was detected in 9 (0.6%) out of1513 samples of DNA from I. ricinus ticks. Thisincluded 163 adults, 50 nymphs and 1300 larvae 3%,8%, and 0% of which, respectively, were PCR-positive(Table 3). An ethidium bromide-stained agarose gel,showing the presence of PCR-amplified B. microti
DNA extracted from nymphs of I. ricinus is presentedin Fig. 3.
Discussion
B. microti-like piroplasms were first isolated morethan a century ago from a Portuguese vole Microtus
incertus (after Sebek et al., 1977). This parasite has beendetected in small mammals and ticks throughoutEurasia since then (Foppa et al., 2002). Humaninfections were first recognized in the late 1950s, thefirst confirmed case of human babesiosis being docu-mented in 1957 in an asplenic man in former Yugoslavia(Skrabalo and Deanovic, 1957). From that time on,
the Mazury Lakes District of north-eastern Poland
% positive No. of positive
ticks (PCR, x/n)
% positive
1 — —
12.8 38/53 72
42 2/5 40
12.2 40/58 69

ARTICLE IN PRESS
Table 3. Levels of Babesia microti infection in Ixodes ricinus collected from rodents and vegetation in the Mazury Lakes District of
north-eastern Poland
Tick developmental stage No. examined No. PCR positive % PCR positive
Larvae feeding on rodentsa 1300 0 0
Nymphs feeding on rodentsa 50 4 8
Questing femalesb 92 4 4
Questing malesb 71 1 1
Total 1513 9 0.6
aCollected from Apodemus flavicollis in 2001.bCollected from the vegetation in the same areas at the same time as feeding larvae and nymphs.
E. Sinski et al. / International Journal of Medical Microbiology 296 (2006) S1, 137–143 141
babesiosis has been considered a human disease, and inEurope, 31 cases of human babesiosis were diagnosed todate attributed mainly to infection with B. divergens
(Zintl et al., 2003). However, hundreds of cases ofhuman babesiosis caused by B. microti or the WA1-typebabesial parasite have been reported in the UnitedStates (Homer et al., 2000).
The present study comprises a comprehensive surveyof B. microti natural infections in A. flavicollis occurringin woodland and two other rodent species (M. arvalis,M. oeconomus) occurring in meadowland. The resultsbased on morphological studies indicate that theprevalence of infection in A. flavicollis was low, mostlyabout 1% of the sampled population. In markedcontrast, prevalence was very high in populations ofthe two microtine rodents, amounting to 12.8% and42% for M. arvalis and M. oeconomus, respectively. Inthis study, Giemsa-stained peripheral blood smears wereused to determine the presence of intra-erythrocyticparasites, but it is important to be aware that thistechnique is not sensitive enough to detect the very lowparasitaemias that often occur in nature, and hence theprevalence data must be treated with some caution,probably representing an underestimation of the realprevalence in any sampled population. All of theparasites detected at the microscopical level weremorphologically similar to the King’s 67 BALB/cmouse-adopted strain of B. microti. With the develop-ment of a more sensitive PCR method described for thedetection of B. microti infection in humans (Persinget al., 1992), molecular studies and more reliablemonitoring of babesial infection became possible.
In the present study, perhaps not surprisingly,evaluation of the prevalence of B. microti in naturallyinfected microtine rodents (M. arvalis) by PCR provedto be more sensitive and equally specific when comparedwith direct examination of blood smears. The twospecies of voles clearly play an important role asreservoirs of B. microti, especially M. arvalis, the speciesthat is most abundant and most widely distributedthroughout the country. Based on morphology and on
the host spectrum, B. microti was previously regarded asjust a single species. Recent work, however, indicatesthat this is perhaps an over-simplistic view. Advances inphylogenetic analyses of molecular data have providednew insights, and it is increasingly clear that what waspreviously considered to be just one species actuallyconsists of a genetically diverse species complex, withsignificant sequence diversity between the constituentclades (Goethert and Telford, 2003). Within thiscomplex, based on analyses of the 18S rRNA gene,three distinct clades have now been recognized. Rodentbabesial parasites comprise two clades, one representingzoonotic isolates, and the other one apparently main-tained only in microtine rodents. In our study, thesequences amplified from microtine rodents and depos-ited in GenBank (accession nos. AY918164 andAY918165), have rDNA sequences identical (100%homology) to that of the Munich B. microti strain(GenBank accession no. AB071177) originally isolatedfrom Mus musculus. Moreover, our sequences wereclearly distinct from the GI isolate (AF231348) fromhumans and the unknown B. microti (AF373331)isolated from I. ricinus ticks (see Table 1).
Through the data reported in this paper, we haveclearly demonstrated that the carriers of B. microti innorth-eastern Poland, the nymphal and adult stages ofthe tick I. ricinus, which are sympatric with the rodentsin our study sites, most likely have the potential totransmit babesial parasites. Four of 50 nymphs (8%),4 of 92 females and 1 of 71 males were found to beinfected with B. microti. These data therefore providesupplementary evidence that the tick species responsiblefor maintaining B. microti in rodent populations inEurope is not only I. trianguliceps but also includesI. ricinus (Duh et al., 2001; Gray et al., 2002).
Regardless of the compelling evidence that zoonoticB. microti occurs elsewhere in Europe (Gray, 2004), thedata presented in this paper indicate that microtinerodents (M. oeconomus and M. arvalis) in north-easternPoland carry the enzootic strain of B. microti. Our datashow that the homology of the 18S rRNA gene, which is

ARTICLE IN PRESSE. Sinski et al. / International Journal of Medical Microbiology 296 (2006) S1, 137–143142
relatively conserved among Babesia species, to theknown zoonotic strain (GI) from Nantucket (USA) isonly 98.19%. Since adult ticks do not feed on rodentsand we failed to detect any infections among unfedI. ricinus larvae, Babesia circulation might be dependenton I. ricinus larvae feeding on infected rodents and inturn on the infection of new rodents by feeding infectednymphs. The conclusion that rodents in our study sitesdo not carry the zoonotic strain of B. microti isstrengthened by the total absence of any records ofhuman babesiosis in the region, although ticks feedon people and evidence of transmission of other tick-borne pathogens, e.g. Borrelia burgdorferi, Anaplasma
phagocytophilum, and tick-borne encephalitis virus(Hermanowska-Szpakowicz et al., 2004). However,although we cannot find any evidence of strains ofB. microti causing infection in humans and thereforebelieve that there is very little risk to public health, wecannot totally exclude the possibility that the zoonoticstrain is present at a very low prevalence. Since this is animportant issue for tourism and for local inhabitants ofthe Mazury Lakes District of Poland, we urge thecontinuation of genetic and biological characterizationon B. microti isolates from Poland and more extensivecoverage of the region than was feasible during ourproject.
Acknowledgments
This study was supported by a KBN Grant no. 2P04C09827. We are grateful to Ma"gorzata Bednarska andKinga Jozwicka for the maintenance of the laboratorystrain of Babesia.
References
Bajer, A., Pawe"czyk, A., Behnke, J.M., Gilbert, F.S., Sinski,
E., 2001. Factors affecting the component community
structure of haemoparasites in bank voles (Clethrionomys
glareolus) from the Mazury Lake District region of Poland.
Parasitology 122, 43–54.
Burkot, T.R., Schneider, B.S., Pieniazek, N.J., Happ, C.M.,
Rutherford, J.S., Slemenda, S.B., Hoffmeister, E., Maupin,
G.O., Zeidner, N.S., 2000. Babesia microti and Borrelia
bissetti transmission by Ixodes spinipalpis ticks among
prairie voles, Microtus ochrogaster in Colorado. Parasitol-
ogy 121, 595–599.
Dammin, G.J., Spielman, A., Benach, J.L., Piesman, J., 1981.
The rising incidence of clinical Babesia microti infection.
Human Pathol. 12, 398–400.
Duh, D., Petrovec, M., Avsic-Zupanc, T., 2001. Diversity
of Babesia infecting European sheep ticks (Ixodes ricinus).
J. Clin. Microbiol. 39, 3395–3397.
Foppa, I.M., Krause, P.J., Spielman, A., Goethert, H.K.,
Gern, L., Brand, B., Telford 3rd., S.R., 2002. Entomologic
and serologic evidence of zoonotic transmission of Babesia
microti in eastern Switzerland. Emerg. Infect. Dis. 8,
722–726.
Goethert, H.K., Telford III, S.R., 2003. What is Babesia
microti? Parasitology 127, 301–309.
Gray, J.S., 2004. Babesia sp.: emerging intracellular parasites
in Europe. Polish J. Microbiol. 53 (Suppl.), 55–60.
Gray, J.S., von Stedingk, L.V., Gurtelschmid, M., Granstrom,
M., 2002. Transmission studies on Babesia microti in
Ixodes ricinus ticks and gerbils. J. Clin. Microbiol. 40,
1258–1263.
Guy, E.C., Stanek, G., 1991. Detection of Borrelia burgdorferi
in patients with Lyme disease by the polymerase chain
reaction. J. Clin. Pathol. 44, 103–110.
Healing, T.D., 1981. Infections with blood parasites in the
small British rodents Apodemus sylvaticus, Clethrionomys
glareolus and Microtus agrestis. Parasitology 83, 179–189.
Hermanowska-Szpakowicz, T., Skotarczak, B., Kondrusik,
M., Rymaszewska, A., Sawczuk, M., Maciejewska, A.,
Adamska, M., Pancewicz, S., Zajkowska, J., 2004. Detect-
ing DNAs of Anaplasma phagocytophilum and Babesia in
the blood of patients suspected of Lyme disease. Ann.
Agric. Environ. Med. 11, 351–354.
Herwaldt, B.L., Caccio, S., Gherlinzoni, F., Aspock, H.,
Slemenda, S.B., Piccaluga, P., Martinelli, G., Edelhofer, R.,
Hollenstein, U., Poletti, G., Pampiglione, S., Loschenber-
ger, K., Tura, S., Pieniazek, N.J., 2003. Molecular
characterization of a non-Babesia divergens organism
causing zoonotic babesiosis in Europe. Emerg. Infect. Dis.
9, 942–948.
Homer, M.J., Aguilar-Delfin, I., Telford III, S.R., Krause,
P.J., Persing, D.H., 2000. Babesiosis. Clin. Microbiol. Rev.
13, 451–469.
Karbowiak, G., Sinski, E., 1996. The finding of Babesia
microti in bank vole Clethrionomys glareolus in the district
of Mazury Lakes (Poland). Acta Parasitol. 41, 50–51.
Kjemtrup, A.M., Conrad, P.A., 2000. Human babesiosis: an
emerging tick-borne disease. Int. J. Parasitol. 30, 1323–1337.
Pawe"czyk, A., Bajer, A., Behnke, J.M., Gilbert, F.S., Sinski,
E., 2004. Factors affecting the component community
structure of haemoparasites in common voles (Microtus
arvalis) from the Mazury Lake District region of Poland.
Parasitol. Res. 92, 270–284.
Persing, D.H., Mathiesen, D., Marshall, W.F., Telford, S.R.,
Spielman, A., Thomford, J.W., Conrad, P.A., 1992.
Detection of Babesia microti by polymerase chain reaction.
J. Clin. Microbiol. 30, 2097–2103.
Persing, D.H., Herwaldt, B.L., Glaser, C., Lane, R.S.,
Thomford, J.W., Mathiesen, D., Krause, P.J., Phillip,
D.F., Conrad, P.A., 1995. Infection with a Babesia-like
organism in northern California. N. Engl. J. Med. 332,
298–303.
Randolph, S.E., 1995. Quantifying parameters in the transmis-
sion of Babesia microti by the tick Ixodes trianguliceps
amongst voles (Clethrionomys glareolus). Parasitology 110,
287–295.
Sebek, Z., Rosicky, B., Sixl, W., 1977. The occurrence of
babesiasis affecting small terrestrial mammals and the
importance of this zoonosis in Europe. Folia Parasitol. 24,
211–220.

ARTICLE IN PRESSE. Sinski et al. / International Journal of Medical Microbiology 296 (2006) S1, 137–143 143
Shih, C.M., Liu, L.P., Chung, W.C., Ong, S.J., Wang, C.C.,
1997. Human babesiosis in Taiwan: asymptomatic infection
with a Babesia microti-like organism in a Taiwanese
woman. J. Clin Microbiol. 35, 450–454.
Sinski, E., 1999. Enzootic reservoir for new Ixodes ricinus-trans-
mitted infections. Wiad. Parazytol. 45, 135–142 (in Polish).
Sinski, E., Karbowiak, G., 1995. Ticks (Ixodidae) in zoonotic
transmission of Babesia microti and B. divergens. Wiad.
Parasitol. 41, 321–327 (in Polish).
Skotarczak, B., Cichocka, A., 2001. Isolation and amplifica-
tion by polymerase chain reaction DNA of Babesia microti
and Babesia divergens in ticks in Poland. Ann. Agric.
Environ. Med. 8, 187–189.
Skrabalo, Z., Deanovic, Z., 1957. Piroplasmosis in man: report
on a case. Doc. Med. Geogr. Trop. 9, 11–16.
Stanczak, J., Gabre, R.M., Kruminis-Lozowska, W., Race-
wicz, M., Kubica-Biernat, B., 2004. Ixodes ricinus as a
vector of Borrelia burgdorferi sensu lato, Anaplasma
phagocytophilum and Babesia microti in urban and sub-
urban forests. Ann. Agric. Environ. Med. 11, 109–114.
Steketee, R.W., Eckman, M.R., Burgess, E.C., Kuritsky, J.N.,
Dickerson, J., Schell, W.L., Godsey, M.S., Davis, J.P.,
1985. Babesiosis in Wisconsin: a new focus of disease
transmission. JAMA 53, 2675–2678.
Tsuji, M., Wei, Q., Zamoto, A., Morita, C., Arai, S., Shiota,
T., Fujimagari, M., Itagaki, A., Fujita, H., Ishihara, C.,
2001. Human babesiosis in Japan: epizootiologic survey
of rodent reservoir and isolation of new type of
Babesia microti-like parasite. J. Clin. Microbiol. 39,
4316–4322.
Walter, G., 1981. Isolierung von Babesia microti (Franca 1912)
aus freilebenden Nymphen von Ixodes ricinus (Linnaeus
1758). Acta Trop. 38, 187–188.
Watkins, R.A., Moshier, S.E., O’Dell, W.D., Pinter, A.J.,
1991. Splenomegaly and reticulocytosis caused by Babesia
microti infections in natural populations of the montane
vole, Microtus montanus. J. Protozool. 38, 573–576.
Zahler, M., Rinder, H., Schein, E., Gothe, R., 2000. Detection
of a new pathogenic Babesia microti-like species in dogs.
Vet. Parasitol. 89, 241–248.
Zintl, A., Mulcahy, G., Skerrett, H.E., Taylor, S.M., Gray,
J.S., 2003. Babesia divergens, a bovine blood parasite of
veterinary and zoonotic importance. Clin. Microbiol. Rev.
16, 622–636.