ixodes (ixodes) pararicinus keirans & clifford, 1985 (acari: ixodidae): description of the...
TRANSCRIPT

Ixodes (Ixodes) pararicinus Keirans & Clifford, 1985 (Acari: Ixodidae):description of the immature stages, distribution, hosts andmedical/veterinary importance
Jose Manuel Venzal1, Agustın Estrada-Pena2, Darci Moraes Barros-Battesti3,Valeria Castilho Onofrio3 & Pablo Martın Beldomenico41Departamento de Parasitologıa Veterinaria, Facultad de Veterinaria, Universidad de la Republica, Av.Alberto Lasplaces 1550, 11600 Montevideo, Uruguay2Departamento de Parasitologıa, Facultad de Veterinaria, Universidad de Zaragoza, Spain3Laboratorio de Parasitologia, Instituto Butantan, Sao Paulo, SP, Brazil4Facultad de Ciencias Veterinarias, Universidad Nacional del Litoral, Esperanza, Santa Fe, Argentina
Accepted for publication 2nd September, 2004
Abstract
The immature stages of the Neotropical tick Ixodes (Ixodes) pararicinus Keirans & Clifford, 1985 are de-scribed from specimens in a laboratory colony initiated from engorged females collected on cattle and larvaefed onmice and chickens. The larva and nymph of I. pararicinus are described using SEMmicrographs as wellas drawings for some features of the larva. Additionally, immature stages of I. pararicinus collected on wildmice and birds in Uruguay and Argentina were compared with specimens from the laboratory colony. Ataxonomic key to the nymphs of the species of the ricinus complex established in the Western Hemisphere ispresented. The distribution of I. pararicinus comprises Argentina, Colombia and Uruguay, but it is alsoprobably established in Bolivia, Brazil, Chile and Peru. Most adult ticks of this species have been found onintroduced domestic artiodactyles, although Neotropical deer species must have been the ancestral host.Larvae and nymphs of I. pararicinus have also been found on sigmodontine rodents and passeriformbirds. Although I. pararicinus is a member of the ricinus complex, which contains the main vectors ofBorrelia burgdorferi (sensu lato), there are few studies concerning its potential for pathogen transmission.
Introduction
According to Horak et al. (2002), Ixodes Latreille,1795 is the largest tick genus, with 241 describedspecies. This list has been recently increased bytwo new Neotropical representatives, I. dicei Kei-rans & Ahjoda, 2003 and I. paranaensis BarrosBattesti, Arzua, Pichorim & Keirans, 2003. Someof these species are vectors of prominent diseasesof medical and veterinary importance (Oliveret al., 1987).
Clifford et al. (1973) recognised 14 subgeneraof Ixodes, with 79 species in the subgenus Ix-odes. Later, Robbins & Keirans (1992) revali-dated the subgenus Ixodiopsis Filippova, 1957,and Durden & Keirans (1996) recognised the
subgenus Trichotoixodes Reznik, 1961, bringingthe total number of Ixodes subgenera to 16. Thisclassification is currently accepted by most ticktaxonomists, in contrast to the division of thegenus Ixodes into several genera as proposed byCamicas et al. (1998). According to Cliffordet al. (1973), some species were not included inany of the previously described subgenera be-cause they were insufficiently known and couldnot be readily identified with any of these speciesgroups (e.g. Ixodes amarali Fonseca, 1935).Nevertheless, Onofrio (2003) included I. amaraliin the subgenus Ixodes after evaluating its mor-phological characteristics, and also becauseI. amarali shared the same type of hosts with otherNeotropical representatives of this subgenus,
Systematic Parasitology (2005) 60: 225–234 � Springer 2005

such as I. loricatus Neumann, 1899, I. luciaeSenevet, 1940 and I. schulzei Aragao & Fonseca,1951. The Ixodes ricinus (L., 1758) complexhas been defined by Keirans et al. (1999) asconsisting of 14 species. Xu et al. (2003) addedthree species to this complex after a molecularanalysis of several species of Ixodes. Theseauthors also proposed the polyphyletic origin ofthat group, suggesting the existence of fourclades. However, I. aragaoi Fonseca, 1935 was notconsidered by Xu et al. (2003), although this is avalid species (Horak et al., 2002; Gugliel-mone et al., 2003) included in the ricinus com-plex (Onofrio, 2003). Therefore, the list of validspecies in the complex should be raised to 18.Of these, eight species are established in theWestern Hemisphere, namely the Nearctic speciesIxodes jellisoni Cooley & Kohls, 1938, I. murisBishopp & Smith, 1937, I. pacificus Cooley &Kohls, 1943 and I. scapularis Say 1821; theNeotropical I. aragaoi and I. pararicinus Kei-rans & Clifford, 1985; and the Neotropical-Nearctic I. affinis Neumann, 1899 and I. minorNeumann, 1902. This group of species has severaltaxonomic problems. For instance,I. aragaoi is considered a synonym of I. affinisby some authors (Cooley & Kohls, 1945;Keirans & Clifford, 1978), and Keirans (1992)does not mention I. aragoi in his list of validtick names. This is unjustified, as morphologicaldifferences between the females of both specieswere clearly defined by Aragao & Fonseca (1952)(Guglielmone et al., 2003). The type-specimensof the Uruguayan I. ricinus rochensis Calzada,1936 are lost, but Camicas et al. (1998) con-sidered it, and also I. pararicinus, as synonymsof I. aragaoi, although they failed to provideany comparative descriptions to support theiropinion. Indeed, the figures of the female ofI. aragaoi as presented by Fonseca (1935) andthose for adults of I. r. rochensis presented byCalzada (1936) show great similarities withI. pararicinus. However, molecular, biologicaland additional morphological studies are neededbefore confirming any synonymy. Currently, bothI. aragaoi and I. pararicinus were consideredvalid names by Horak et al. (2002) and wereincluded in a list of Neotropical tick species byGuglielmone et al. (2003).
The morphological features of immature tickstages are of importance in determining the
taxonomic status of this genus. There aredescriptions of the immature stages of the Nearcticand Neotropical-Nearctic species of the ricinuscomplex, but not of the Neotropical I. pararicinusand I. aragaoi (see Guglielmone et al., 2003).
Herein we describe the larva and the nymph ofI. pararicinus from a large array of specimens ob-tained under laboratory conditions. Furthermore,this material was compared with larvae andnymphs collected previously from rodents andbirds in Uruguay and Argentina (Beldomenicoet al., 2003; Venzal et al., 2003). A key to thenymphs of the species of the ricinus complexpresent in the Western Hemisphere is alsoprovided.
Materials and methods
Engorged females of I. pararicinus were collectedfrom cattle at Laguna Negra, Department ofRocha, Uruguay (34�03¢ S, 53�40¢ W). They werekept in the dark under controlled conditions at27 �C and 90% RH. Larvae were mounted inHoyer’s medium to make semi-permanent slides.A subset of larvae fed on chickens and laboratorymice. Engorged larvae were kept under the sameconditions as the females until moulting. Othermaterial examined was collected from wild ani-mals, as described below. All measurements are inmillimetres; the average is followed by the stan-dard deviation and by the range in parentheses.The larval chaetotaxy of the idiosoma and tarsus Ifollows the terminology by Clifford & Anastos(1960), and that of the palpi follows that proposedby Hess & Vlimant (1983). Material for SEM wasprepared according to the methods presented byKeirans et al. (1976). The micrographs were ob-tained using a JEOL JMS-5900 scanning electronmicroscope. The taxonomy of hosts followsWilson & Reeder (1993) and the nomenclatureproposed by Gonzalez (2001) for mammals andAzpiroz (2001) for birds.
Ixodes (I.) pararicinus Keirans & Clifford, 1985
Material examinedAdditional Uruguayan and Argentine materialexamined was as follows: 4 larvae, 2 nymphs,
226

Syndactyla rufosuperciliata (Lafresnaye) (Passeri-formes: Furnariidae), coll.S. Claramunt, Gruta delos Helechos (31�37¢S, 56�02¢W), Tacuarembo,April 10, 1995; 6 nymphs,Phacellodomus straticollis(d’Orbigny & Lafresnaye) (Passeriformes: Fur-nariidae), coll. J. Venzal and C. de Souza, LagunaNegra (34�03¢S, 53�40¢W), Rocha, March 27, 2000;1 nymph, S. rufosuperciliata, coll. S. Claramunt,Paso Centurion (32�10¢S, 53�45¢W), Cerro Largo,March 27, 2000; 6 nymphs,Akodon azarae (Fischer)(Rodentia: Muridae), coll. E.M. Gonzalez, PosadaLa Laguna (34�17¢S, 54�40¢W), Maldonado, Janu-ary 28, 2001; 4 nymphs, Oligoryzomys delticola(Thomas) (Rodentia: Muridae), coll. E.M. Gon-zalez, Posada La Laguna (34�17¢S, 54�40¢W),Maldonado, January 28, 2001; 2 larvae, 2 nymphs,Calomys sp. cf. C. fecundus (Thomas) (Rodentia:Muridae), coll. P. Beldomenico, Parque NacionalEl Rey (24�42¢S, 64�38¢W), Salta, June 1, 2001; 1larva, 1 nymph, Calomys sp. cf. C. fecundus (Tho-mas) (Rodentia: Muridae), coll. P. Beldomenico,Parque Nacional El Rey (24�42¢S, 64�38¢W), Salta,November 11, 2000.
Diagnosis
Larva: scutum rounded, clearly broader thanlong; basis capituli long, triangular; cornua ab-sent; ventral basis with transverse suture, auric-ulae short; hypostome presenting apex clearlypointed with dental formula 3/3 to midlength,then 2/2 to base, with all denticles sharplypointed; palpi with inconspicuous suture betweenarticles II and III; anal groove slightly divergent.Nymph: characterised by the presence of abroadly pyriform scutum; lateral carinae slightlyapparent; cornua robust, posterolaterally pro-duced; palpi flattened on the internal surface;auriculae long; hypostome with sharply pointedapex, presenting dental formula 3/3 at tip, then2/2 to base, all denticles sharply pointed; analgroove slightly diverging posteriorly.
Description (Figures 1–17)
Larva (Figures 1–11) (based on 10 measuredspecimens)Idiosoma.Dorsal surface oval (Figures 1,6). Lengthfrom apex of scapulae to posterior margin of id-iosoma 0.594 ± 0.025 (0.550–0.630), width0.472 ± 0.018 (0.450–0.500). Sensilla sagittiformia
(large wax gland) absent. Setae (Figure 6): 11pairs-3 pairs of central dorsal setae (Cd1–Cd3), 7pairs of marginal dorsal setae (Md1–Md7) and 1pair of supplementary setae (S). Scutum broadlyrounded, length along median line 0.333 ± 0.118(0.320–0.350), width 0.383 ± 0.014 (0.360–0.410);tegument with irregular hexagonal ornamenta-tion, punctations rare; eyes absent; cervicalgrooves (Figure 1) converging at level of posteriorpart of coxa I then diverging until level of coxaII. Scutal setae: 5 pairs (Sc1–Sc5). Ventral surface(Figure 2) with 13 pairs of setae plus 1 pair on theanal valves, i.e. 3 sternal setae (St1–St3), with St1located below level of coxae I, St2 below of coxaeII and St3 between coxae III, 2 pre-anal pairs(Pa1–Pa2), 4 premarginal pairs (Pm1–Pm4), withPm5 pair observed in 2 specimens, and 4 marginalventral pairs (Mv1–Mv4). Anal groove slightlydivergent (Figure 7).
Gnathosoma. Basis capituli (Figures 3,9) triangu-lar, with posterior margin slightly convex andcornua absent, length 0.219 ± 0.011 (0.212–0.237), width 0.150 ± 0.005 (0.140–0.155). Palpilong (Figure 9) excavated close to femur, withsuture between article II (femur) and III (genu)inconspicuous, length 0.176 ± 0.005 (0.170–0.180), width 0.044 ± 0.024 (0.040–0.047); palpaltrochanter length 0.282 ± 0.025 (0.263–0.315),width 0.368 ± 0.026 (0.342–0.394); palpal femurand genu combined length 0.143 ± 0.01 (0.1367–0.157). Ventral basis (Figures 4,10) with 2 pairs ofposthypostomal setae (Ph1–Ph2), distance be-tween Ph1 0.045 ± 0.015 (0.044–0.047) and Ph20.031 ± 0.015 (0.029–0.031); posterior margin ofventral part of capitulum clearly sinuous; auric-ulae (Figure 10) short, widely separated fromeach other, 0.129 ± 0.005 (0.125–0.137), roun-ded, projected laterally; hypostome (Figure 10)total length from apex to Ph1 setae0.144 ± 0.002 (0.142–0.147), width 0.040 ±0.002 (0.037–0.045), with clearly pointed apex;dental formula 3/3 to mid-length, then 2/2 tobase, with all denticles sharply pointed, 9/10denticles in row I, 9 in row 2 and 6 in row 3.Palpal setae (Figures 3,4): 7 on genu, i.e. 1 par-axial (Gp1), 2 antiaxial (Ga1–Ga2), 3 dorsal(Gd1–Gd3) and 1 ventral (Gv1); 6 on femur, i.e. 1paraxial (Fp1), 2 antiaxial (Fa1–Fa2), 1 dorsal(Fd1) and 2 ventral (Fv1–Fv2); 10 on tibiotarsus(Ttt1–Ttt10); trocanter 0.
227

Legs. Coxa I with 2 triangular spurs; internallonger (Figure 8). Coxa II with single externalspur, shorter than homologous spur of coxa I.Coxa III without spur. Coxa I with 3 setae, 1anterior (CIa), 1 posterior (CIp) and 1 paraxial(CIpa); coxae II and III each with 1 anterior seta
(CIIa, CIIIa) and 1 posterior seta (CIIp, CIIIp);trochanters without spurs. Tarsus I (Figure 5)length 0.211 ± 0.002 (0.200–0.234), width0.071 ± 0.001 (0.070–0.073). Setae on tarsus I:group I absent; 4 setae in group II–2 dorsal (dII1,dII2), 1 lateral antiaxial (laII1) and 1 lateral
0,1 mm
Sc1
Sc2
Sc3
Sc4
Sc5Md1
Md2
Md3
Md4
Md5
Md6Md7
S Cd1
Cd2
Cd3
1
A
0,1 mm 2
CIa
CIpCIpa
CIIa
CIIpSt1
St2
St3
CIIIa
CIIIp
Pm1
Pm2
Pm3
Pm4
Mv1
Mv2
Mv3
Mv4
Pa1
Pa2
0,1 mm
Gp1 Gd1
Gd2
Ga1
Ga2
Gd3
Fa1Fd1
Fp1
Fa2
3
0,1
mm
Ttt
Gv1
Fv1
Fv2
Ph1
Ph2
4
VvII1-2
CHO
Pv
_
0,1
mm
5VvIII1-2
lpII3
laII3
lpII2
laII2
laII1
lpII1
laI1
lpI1
VvI1-2
dVI1-2
dIII1-4dII1-2
Figures 1–5. Ixodes pararicinus larva. 1. Idiosoma dorsal. 2 Idiosoma ventral. 3. Gnathosoma dorsal. 4. Gnathosoma ventral. 5.Tarsus I, lateral view. Abbreviations: CHO, capsule of Hallers organ; pv, pulvillus; Ttt, tibiotarsus; Setae, A, anal; Cd, central dorsal;CIa, coxa I anterior; CIp, coxa I posterior; CIpa, coxa I paraxial; CIIa, coxa II anterior; CIIp, coxa II posterior; CIIIa, coxa IIIanterior; CIIIp, coxa III posterior; dII, dorsal group II; dIII, dorsal group III; dVI, dorsal group VI; Fa, femoral antiaxial; Fd, femoraldorsal; Fp, femoral paraxial; Fv, femoral ventral; Ga, genual antiaxial; Gd, genual dorsal; Gp, genual paraxial; Gv, genual ventral; laI,lateral anterior group I; laII, lateral anterior group II; lpI, lateral posterior group I; lpII, lateral posterior group II; Md, marginaldorsal; Mv, marginal ventral; Pa, pre-anal; Ph, post hypostomal; Pm, premarginal; S, supplementary; Sc, scutal; St, sternal; VvI,ventral group I; VvII, ventral group II; VvIII, ventral group III.
228

paraxial (lpII1); 4 dorsal in group III (dIII1–4);groups IV and V dorsal setae absent; 2 dorsal inthe group VI (dVI1–2). Ventral and lateral setae:
4 setae in group I – 2 ventral (VvI1–2); 2 lateralantiaxial (laI1) and lateral paraxial (lpI1); 4 setaein group II – 2 ventral (VvII1–2); 1 lateral antiaxial
Figures 6–11. Ixodes pararicinus larva. 6. Idiosoma dorsal, bar 200 lm. 7. Idiosoma ventral. 8. Coxae I–III. 9. Gnathosoma dorsal. 10.Gnathosoma ventral. 11. Haller’s organ. Abbreviations: a, auriculae; ag, anal groove; bc, basis capituli; cg, cervical groove; CHO,capsule of Haller’s organ; Fe, femur; Ge, genu; Ph, post hypostomal setae; phs, prehalleral setae; Tr, trochanter; Ttt, tibiotarsus.Scale-bars: 6,7, 200 lm; 8-10, 100 lm; 11, 40 lm.
229
(laII2) and 1 lateral paraxial (lpII2); 4 setae ingroup III – 2 ventral (VvIII1–2); 2 lateral antiaxial(laIII1) and lateral paraxial (lpIII1). Haller’s organ(Figure 11) with 5 prehalleral setae (phs) plus3 apparent setae in the capsule.
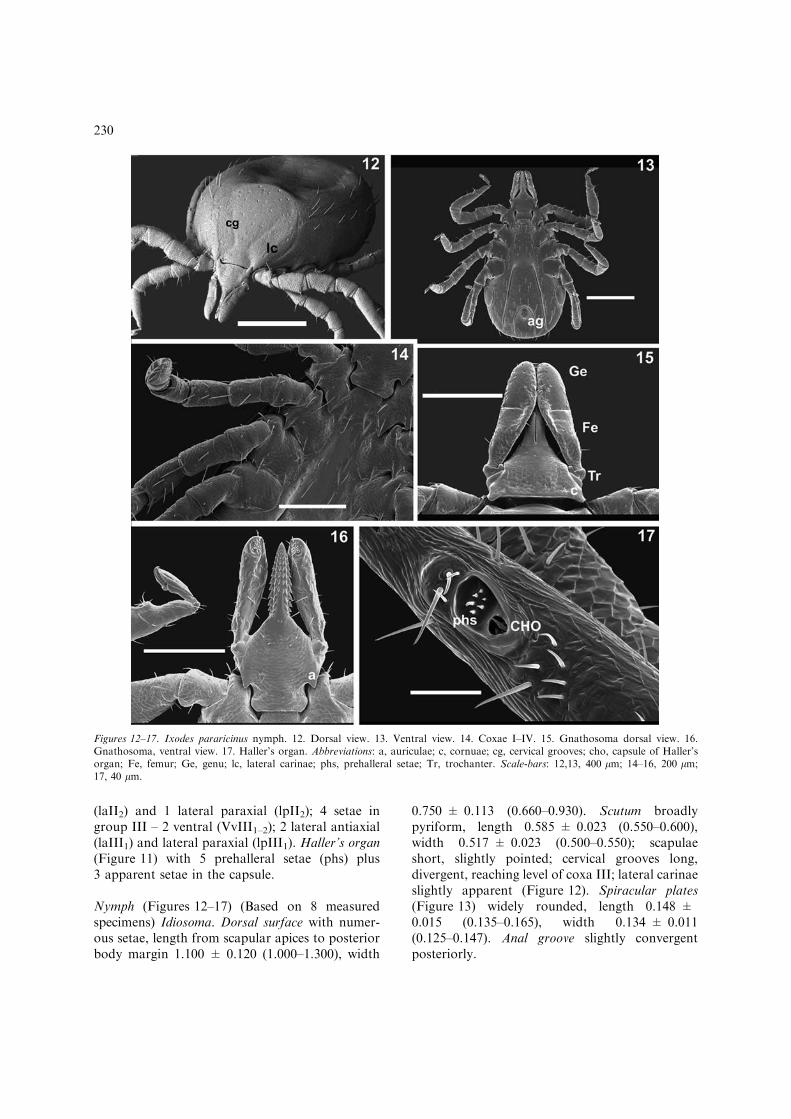
Nymph (Figures 12–17) (Based on 8 measuredspecimens) Idiosoma. Dorsal surface with numer-ous setae, length from scapular apices to posteriorbody margin 1.100 ± 0.120 (1.000–1.300), width
0.750 ± 0.113 (0.660–0.930). Scutum broadlypyriform, length 0.585 ± 0.023 (0.550–0.600),width 0.517 ± 0.023 (0.500–0.550); scapulaeshort, slightly pointed; cervical grooves long,divergent, reaching level of coxa III; lateral carinaeslightly apparent (Figure 12). Spiracular plates(Figure 13) widely rounded, length 0.148 ±0.015 (0.135–0.165), width 0.134 ± 0.011(0.125–0.147). Anal groove slightly convergentposteriorly.
Figures 12–17. Ixodes pararicinus nymph. 12. Dorsal view. 13. Ventral view. 14. Coxae I–IV. 15. Gnathosoma dorsal view. 16.Gnathosoma, ventral view. 17. Haller’s organ. Abbreviations: a, auriculae; c, cornuae; cg, cervical grooves; cho, capsule of Haller’sorgan; Fe, femur; Ge, genu; lc, lateral carinae; phs, prehalleral setae; Tr, trochanter. Scale-bars: 12,13, 400 lm; 14–16, 200 lm;17, 40 lm.
230

Gnathosoma. Total length from palpal apices tobase of gnathosoma 0.345 ± 0.005 (0.340–0.350).Basis capituli (Figure 15) triangular, width0.228 ± 0.007 (0.215–0.235); posterior marginslightly convex; cornua robust, wide, posterolat-erally produced, not sharply pointed. Ventral basis(Figure 16) length from base of hypostome toposterior margin 0.223 ± 0.007 (0.215–0.230),width 0.193 ± 0.005 (0.190–0.197) between au-riculae; lateral margins essentially straight frompalpal insertions to auriculae, then concave, thendiverging to widely convex at posterior margin;auriculae long, essentially straight, pointed. Palpilong and internally excavated, combined length0.301 ± 0.008 (0.290–0.310), width 0.066 ± 0.004(0.062–0.072); palpal article I (trochanter) length0.054 ± 0.003 (0.052–0.057), width 0.055 ± 0.003(0.052–0.057); palpal article II (femur) length0.146 ± 0.008 (0.139–0.158); palpal article III(genu) length 0.136 ± 0.005 (0.131–0.144). ArticleII slightly larger than III; both are 3 times longerthan article I; length of articles in descending order2, 3, 1, 4. Hypostome length 0.272 ± 0.008 (0.263–0.279), width 0.064 ± 0.007 (0.060–0.075), withvery pointed apex; dental formula essentially 3/3 attip, then 2/2 to base, with all denticles sharplypointed.
Legs. Moderately long; coxa I with an externalspur reaching level of coxa II and with prominent,sharply pointed internal spur; external spurs equalin size on coxae II–III, smaller on coxa IV (Fig-ure 14). Spurs on trochanters absent. Haller’sorgan (Figure 17) with 6 prehalleral setae (phs)plus 6 apparent setae in capsule.
Discussion
Species relationships
As expected, the immature stages of I. pararici-nus are morphologically close to correspondingstages of species belonging to the ricinus com-plex. Among the Nearctic and Neotropical rep-resentatives of this group, I. pararicinus hasmany morphological similarities with the larvaand nymph of I. scapularis. However, accordingto the molecular analysis performed by Xu et al.(2003), I. pararicinus should be placed in the
clade shared with I. affinis, I. muris and I. minor.The larvae of the seven Western Hemisphererepresentatives of the ricinus complex (the larvaof I. aragaoi is unknown) appear to be quitesimilar. Moreover, the descriptions of the larvaof I. pacificus, I. minor and I. muris are notadequate for comparison and we have thereforedecided to defer construction of larval keys,pending further chaetotactic studies and SEM-based redescriptions of the larva of these species.Durden & Keirans (1996) produced an SEM-based key for the nymphs of all Ixodes speciesfound in the USA; six of the eight Westernspecies of the ricinus complex are included inthat study. Our key includes one Neotropicalspecies, since the nymph of I. aragaoi is stillundescribed.
Key to nymphs
1. Hypostome apically rounded..…………………2– Hypostome apically pointed.…………………..42. Apex of hypostome with 4/4 dentition; denticles
sharply pointed. Dorsal posterior margin ofbasis capituli straight……….…………..I. affinis
– Apex of hypostome with 3/3 dentition…….…33. Dorsal posterior margin of basis capituli
straight; hypostomal teeth in rows 2 and 3rounded, not pointed...…..................I. pacificus
– Dorsal posterior margin of basis capituli con-cave…….…......…...................…........I. jellisoni
4. Palpi sinuous……………………………I. minor– Palpi straight……………………………………55. Auriculae salient posterolaterally; posterior
margin of the basis capituli straight. Nearcticspecies……………………………………I. muris
– Auriculae not salient posterolaterally.....………...............…...…............................... 6
6. Basis capituli dorsally with posterior marginslightly convex; Neotropical species...................................................…….......I. pararicinus
– Basis capituli dorsally with posterior margindistinctly or strongly convex; Nearctic spe-cies…………………………………..I. scapularis
Distribution and hosts
I. pararicinus has a Neotropical distribution. It hasbeen recorded in Argentina and Uruguay (Keirans
231

et al., 1985; Guglielmone et al., 1992; Venzalet al., 2003) as well as Colombia (Mattar & Lopez-Valencia, 1998), and its presence in both Braziland Peru is probable (Keirans et al., 1999). SomeIxodes specimens collected on horses in Bolivia(Torregiani, 1914), on cattle in Chile (Gervais,1849) and on sheep and cattle in Peru (Escomel,1929; Escalante & Mollinedo, 1989) may also beI. pararicinus.
There are no records of human infestations withI. pararicinus. We consider that most Neotropicalrecords of ‘I. ricinus’ represent of I. pararicinus.Adults of this tick species parasitise domesticanimals, such as cattle, horses, sheep and goats(Gervais, 1849; Torregiani, 1914; Escomel, 1929;Boero, 1957; Keirans et al., 1985; Escalante &Mollinedo, 1989; Guglielmone et al., 1992; Mattar& Lopez-Valencia, 1998; Venzal et al., 2003). Allthese hosts were introduced into the Neotropicalregion and, therefore, surely do not represent theoriginal host of I. pararicinus. There are few recordsof I. pararicinus or ‘I. ricinus’ parasitising Neo-tropical artiodactyles, such as the grey brocket deerMazama gouazoupira (Fischer) (Calzada, 1938;Ivancovich, 1973; Venzal et al., 2003). This, orother species of Neotropical artiodactyles, mayrepresent the natural host(s) of I. pararicinus,although this tick species has the ability to surviveon introduced hosts. The preference for large un-gulates is typical of several species of the ricinuscomplex in the Western Hemisphere. The black-tailed deer Odocoileus hemionus columbianus(Richardson) has been recorded as the main hostfor I. pacificus, while the white-tailed deer Odocoi-leus virginianus (Zimmermann) is considered themain host for adults of both I. scapularis andI. affinis (see Durden & Keirans, 1996).
The larvae and nymphs of I. pararicinus arefound mostly on sigmodontine rodents and pass-eriform birds, but they have never been nevercollected on reptiles, although these hosts arecommon for both I. pacificus and I. scapularis insome parts of their range (Durden & Keirans,1996). The most important hosts for larvae andnymphs of I. pararicinus in Argentina andUruguay appear to be the rodents Akodon spp.,A. azarae, Oryzomys spp., Oligoryzomys spp.,O. delticola and Calomys spp., and also small birds,such as Phacellodomus striaticollis, Syndactylarufosuperciliata and Cyanocorax chrysops (Vieillot)(Beldomenico et al., 2003; Venzal et al., 2003).
There is also a record on Thylamys venusta(Thomas) (Didelphimorphia: Didelphidae) inArgentina (Beldomenico et al., 2003). Rodentsand birds are also common hosts for other speciesof the ricinus complex established in the WesternHemisphere. All stages of I. jellisoni are parasites onheteromyid rodents (Durden & Keirans, 1996;Keirans et al., 1996). Both I. minor and I. murisutilise rodents as their principal hosts, while theimmature stages of both species have been alsorecorded on birds (Durden & Keirans, 1996).
Medical and veterinary importance
The ricinus complex of species contains themain vectors of the Lyme disease spirochaeteBorrelia burgdorferi (sensu lato); these are I. scap-ularis and I. pacificus in North America, I. ricinusin the Western Palaeartic and I. persulcatus in Asiaand parts of the former Soviet Union (Oliver,1996). I. jellisoni, I. minor and I. muris have beensuggested as potential vectors of B. burgdorferi(s. l.) or may be involved in the epidemiology ofthe disease (Durden & Keirans, 1996 Keiranset al., 1996; Banks et al., 1998; Lane et al., 1999;Dolan et al., 2000).
The importance in human health of bothI. affinis and I. aragaoi is currently unknown, asLyme disease has not been adequately defined noraccurately diagnosed for the Neotropical region,where I. pararicinus prevails. Search for the agentof Lyme disease in I. pararicinus in Colombia(Mattar & Lopez-Valencia, 1998) and Ixodes spp.in Chile (Osorio, 2001) were negative. The veteri-nary importance of I. pararicinus cannot be dis-counted, as other species in the group are mainvectors of viruses, bacteria and protozoa, and havealso been also in clinical cases of tick paralysis(Durden & Keirans, 1996). Boero (1957) men-tioned that ‘I. ricinus’ was involved in a severereaction of cattle against the adult stages at theirattachment site. We have noted the same skininflammatory reaction in cattle in the RochaDepartment of Uruguay.
Acknowledgements
This study was supported in part by theFundacao de Amparo a Pesquisa do Estado deSao Paulo through project Biota-Fapesp
232

99/05446-8 to DMBB. We thank Alberto Gugl-ielmone for a critical reading of the manuscript,Vanessa Maria Martins Rebello for technicalassistance in the laboratory and Ana Vieira daCosta for the larval drawings. We are alsograteful to Alejandro Marquez and Jorge Troc-coli from the Unidad de Microscopıa Electro-nica, Facultad de Ciencias, Montevideo(Uruguay) for their assistance in providingscanning electron micrographs, and to EnriqueM. Gonzalez and Santiago Claramunt for field-collected specimens.
References
Aragao, H.B. & Fonseca, F. (1952) Notas de ixodologia.III. Confirmacao de Ixodes aragaoi Fonseca, 1935,de Ixodes amarali Fonseca, 1935, e lista das especies dogenero Ixodes que ocorrem no Brasil (Acari, Ixodidae).Memorias do Instituto Butantan, 50, 727–728.
Azpiroz, A.B. (2001) Aves del Uruguay. Lista e introduccion a subiologıa y conservacion. Montevideo: Aves Uruguay-GUPE-CA, 104 pp.
Banks, C.W., Oliver, J.H. Jr., Phillips, J.B. & Clark, K.L.(1998) Life cycle of Ixodes minor (Acari: Ixodidae) in thelaboratory. Journal of Medical Entomology, 35, 496–499.
Beldomenico, P.M., Baldi, C.J., Antoniazzi, L.R., Orduna,G.M., Mastropaolo, M., Macedo, A.C., Ruiz, M.F.,Orcellet, V.M., Peralta, J.L., Venzal, J.M., Mangold, A.J.& Guglielmone, A.A. (2003) Ixodid Ticks (Acari: Ixodidae)present at Parque Nacional El Rey, Argentina. NeotropicalEntomology, 32, 273–277.
Boero, J.J. (1957) Las garrapatas de la Republica Argentina(Acarina: Ixodoidea). Buenos Aires: Universidad de BuenosAires, Departamento Editorial, 113 pp.
Calzada, V. (1936) Comprobacion de ‘‘Ixodes ricinus’’ (nuevasubespecie) en el Uruguay. Boletın Mensual, Direccion deGanaderıa, Uruguay, 20, 103–109.
Calzada, V. (1938) Nueva comprobacion de Ixodes ricinus en elpaıs. Boletın Mensual, Direccion de Ganaderıa, Uruguay, 3,238–241.
Camicas, J.L., Hervy, J.P., Adam, F. & Morel, P.C. (1998)Les tiques du monde. Paris: Editions de l’Orstom, InstitutFrancais de Recherche Scientifique pour le Developpementen Cooperation, 233 pp.
Clifford, C.M. & Anastos, G. (1960) The use of chaetotaxy forthe identification of larval ticks (Acarina: Ixodidae). Journalof Parasitology, 46, 567–578.
Clifford, C.M., Sonenshine, D.E., Keirans, J.E. & Kohls, G.M.(1973) Systematics of the subfamily Ixodinae (Acarina:Ixodidae). I. The subgenera of Ixodes. Annals of theEntomological Society of America, 66, 489–500.
Cooley, R.A. & Kohls, G.M. (1945) The genus Ixodes in NorthAmerica. National Institute of Health Bulletin, 184, 1–246.
Dolan, M.C., Lacombe, E.H. & Piesman, J. (2000) Vectorcompetence of Ixodes muris (Acari: Ixodidae) for Borreliaburgdorferi. Journal of Medical Entomology, 37, 766–768.
Durden, L.A. & Keirans, J.E. (1996) Nymphs of the genusIxodes (Acari: Ixodidae) of the United States: taxonomy,
identification key, distribution, hosts, and medical/veterinaryimportance. Lanham, Maryland: Entomological Society ofAmerica, 95 pp.
Escalante, J.A. & Mollinedo, L. (1989) Ocho artropodos deinteres veterinario en el Cusco. Revista Peruana de Entomo-logıa, 31, 46–47.
Escomel, E. (1929) Fauna de Arequipa. Obras CientıficasEscomel, 1, 367–418.
Fonseca, F. (1935) Notas de acareologia. XV. Occorrencia desub-especie de Ixodes ricinus (L., 1758) no Estado de S. Paulo(Acarina, Ixodidae). Memorias do Instituto Butantan, 9, 131–135.
Gervais, P. (1849) Aracnidos. In: Gay, C. (Ed.) Historia fısica ypolıtica de Chile, Volume 4. Parıs, Santiago, pp. 5–52.
Gonzalez, E.M. (2001) Guıa de campo de los mamıferos deUruguay. Introduccion al estudio de los mamıferos. Montevi-deo: Vida Silvestre, 339 p.
Guglielmone, A.A., Mangold, A.J. & Aufranc, C.R. (1992)Haemaphysalis juxtakochi, Ixodes pararicinus (Ixodidae) andOtobius megnini (Argasidae) in relation to the phytogeogra-phy of Argentina. Annales de Parasitologie Humaineet Comparee, 67, 91–93.
Guglielmone, A.A., Estrada-Pena, A., Keirans, J.E. & Robbins,R.G. (2003) Ticks (Acari: Ixodida) of the Neotropicalzoogeographic region. Atalanta, Houten, The Netherlands:Special Publication of the International Consortium onTicks and Tick-borne Diseases, 173 pp.
Hess, E. & Vlimant, M. (1983) The tarsal sensory system ofAmblyomma variegatum Fabricius (Ixodidae: Metastriata).II. No pore sensilla. Revue Suisse de Zoologie, 90, 157–167.
Horak, I.G., Camicas, J.L. & Keirans, J.E. (2002) TheArgasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida): aworld list of valid tick names. Experimental and AppliedAcarology, 28, 27–54.
Ivancovich, J.C. (1973) Las garrapatas del Chaco y Formosa.Revista de Investigaciones Agropecuarias. Serie 4. PatologiaAnimal, 10, 9–24.
Keirans, J.E. (1992) Systematics of the Ixodida (Argasidae,Ixodidae, Nuttaliellidae): An overview and some problems.In: Fivaz, B.H., Petney, T.N. & Horak, I.G. (Eds) TickVector Biology Medical and Veterinary Aspects. Berlin:Springer Verlag, pp. 1–21.
Keirans, J.E., Clifford, C.M. & Corwin, D. (1976) Ixodes sigelosn. sp. (Acarina: Ixodidae), a parasite of rodents in Chile,with a method for preparing ticks for examination byscanning electron microscopy. Acarologia, 18, 217–225.
Keirans, J.E. & Clifford, C.M. (1978) The genus Ixodes in theUnited States: a scanning electron microscope study and keyto adults. Journal of Medical Entomology, 15 (Suppl. 2), 149.
Keirans, J.E., Clifford, C.M., Guglielmone, A.A. & Mangold,A.J. (1985) Ixodes (Ixodes) pararicinus, n. sp. (Acari:Ixodoidea: Ixodidae), a South American cattle tick longconfused with Ixodes ricinus. Journal of Medical Entomology,22, 401–407.
Keirans, J.E., Brown, R.N. & Lane, R.S. (1996) Ixodes (Ixodes)jellisoni and I. (I.) neotomae (Acari: Ixodidae): descriptionsof the immature stages from California. Journal of MedicalEntomology, 33, 319–27.
Keirans, J.E., Needham, G.R. & Oliver, J.H. Jr. (1999) TheIxodes ricinus complex worldwide: diagnosis of the species inthe complex, hosts and distribution. In: Needham, G.R.et al. (Eds) Symposia, Acarology 9, Volume 2, pp. 341–347.
Lane, R.S., Peavey, C.A., Padgett, K.A. & Hendson, M. (1999)Life history of Ixodes (Ixodes) jellisoni (Acari: Ixodidae) and
233

its vector competence for Borrelia burgdorferi sensu lato.Journal of Medical Entomology, 36, 329–334.
Mattar, S. & Lopez-Valencia, G. (1998) Searching for Lymedisease in Colombia: a preliminary study on the vector.Journal of Medical Entomology, 35, 324–326.
Oliver, J.H. Jr. (1996) Lyme borreliosis in the Southern UnitedStates: A review. Journal of Parasitology, 82, 926–935.
Oliver, J.H. Jr.,Keirans, J.E., Lavender,D.R.&Hutcheson,H.J.(1987) Ixodes affinis Neumann (Acari: Ixodidae): new hostand distribution records, description of inmatures, seasonalactivities in Georgia, and laboratory rearing. Journal ofParasitology, 73, 646–652.
Onofrio, V.C. (2003) O genero Ixodes Latreille, 1795 (Acari:Ixodidae) no Brasil: distribuicao geografica, hospedeiros,taxonomia e chave de identificacao para as especies.Dissertacao de Mestrado. Universidade Federal Rural doRio de Janeiro, Seropedica, Rio de Janeiro, 108 pp.
Osorio, G. (2001) Busqueda de la espiroqueta Borreliaburgdorferi sensu lato mediante PCR en garrapatas ixoid-
eas chilenas silvestres. Revista Medica de Chile, 129, 270–276.
Robbins, R.G. & Keirans, J.E. (1992) Systematics and ecologyof the subgenus Ixodiopsis (Acari: Ixodidae: Ixodes).Volume XIV. Lanham, Maryland: Entomological Societyof America, 159 pp.
Torreggiani, G. (1914) I zooparassiti osservati nella faunaBoliviana. Nuovo Ercol, 19, 417–425.
Venzal, J.M., Castro, O., Cabrera, P.A., de Souza, C.G. &Guglielmone, A.A. (2003) Las garrapatas de Uruguay:especies, hospedadores, distribucion e importancia sanitaria.Veterinaria (Montevideo), 38, 17–28.
Wilson, D.E. & Reeder, D.M. (1993) Mammal species of theworld: a taxonomic and geographic reference. 2nd Edition.Washington: Smithsonian Institution Press, 1207 pp.
Xu, G., Quentin, Q.F., Keirans, J.E. & Durden, L.A. (2003)Molecular phylogenetic analyses indicate that the Ixodesricinus complex is a paraphyletic group. Journal of Parasi-tology, 89, 452–457.
234