at a glance: cellular biology for engineers
TRANSCRIPT

Computational Biology and Chemistry 32 (2008) 315–331
Contents lists available at ScienceDirect
Computational Biology and Chemistry
journa l homepage: www.e lsev ier .com/ locate /compbio lchem
Review
At a glance: Cellular biology for engineers
K. Khoshmanesha,∗, A.Z. Kouzania, S. Nahavandia, S. Baratchib, J.R. Kanwarb
a School of Engineering and IT, Deakin University, Geelong VIC 3217, Australiab Institute of Biotechnology (BioDeakin), Deakin University, Geelong VIC 3217, Australia
a r t i c l e i n f o
Article history:Received 14 March 2008Accepted 6 July 2008
Keywords:Cellular biologyCell organellesMicro-bio-factoryEnergy generationProtein synthesis
a b s t r a c t
Engineering contributions have played an important role in the rise and evolution of cellular biology.Engineering technologies have helped biologists to explore the living organisms at cellular and molecularlevels, and have created new opportunities to tackle the unsolved biological problems. There is now agrowing demand to further expand the role of engineering in cellular biology research. For an engineerto play an effective role in cellular biology, the first essential step is to understand the cells and theircomponents. However, the stumbling block of this step is to comprehend the information given in thecellular biology literature because it best suits the readers with a biological background. This paper aimsto overcome this bottleneck by describing the human cell components as micro-plants that form cells asmicro-bio-factories. This concept can accelerate the engineers’ comprehension of the subject. In this paper,first the structure and function of different cell components are described. In addition, the engineeringattempts to mimic various cell components through numerical modelling or physical implementationare highlighted. Next, the interaction of different cell components that facilitate complicated chemical
processes, such as energy generation and protein synthesis, are described. These complex interactionsare translated into simple flow diagrams, generally used by engineers to represent multi-componentprocesses.C
1d
© 2008 Elsevier Ltd. All rights reserved.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3162. The Classification of Microorganisms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3163. Contribution of Engineering to the Evolution of Cellular Biology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3174. Description of Human Cell Organelles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 318
4.1. Cell Membrane . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3194.2. Cytoplasm . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3204.3. Endoplasmic Reticulum (ER) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3214.4. Golgi Body . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3224.5. Lysosomes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3224.6. Centrosome . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3234.7. Mitochondria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3234.8. Nucleus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 324
4.8.1. Nuclear Envelope. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 324
4.8.2. Nucleoplasm . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4.8.3. Chromatin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4.8.4. Nucleolus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4.9. Ribosome . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
∗ Corresponding author. Tel.: +61 3 52272208; fax: +61 3 52272167.E-mail address: [email protected] (K. Khoshmanesh).
476-9271/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.oi:10.1016/j.compbiolchem.2008.07.010
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 324. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 324. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 325. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 325

316 K. Khoshmanesh et al. / Computational Biology and Chemistry 32 (2008) 315–331
5. Description of Cells’ Major Chemical Processes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3255.1. Energy Generation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3255.2. Protein Synthesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 327
6. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 329. . . . . .
1
ibWsenatmmtt
hpptcrtnacp
btuotbaoom
cfctmla
cctifemcp6
2
tbtbtaanil
uclmSwS(
cwctwooemaed(epe(
oftwostfb
e
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Introduction
In recent years, the fusion of engineering and biology has creatednnovative areas such as systems biology, synthetic biology, andiomedical engineering (Andrianantoandr et al., 2006; Brent, 2004;esterhoff and Palsson, 2004), and also promising technologies
uch as DNA microarrays, biosensors, and lab-on-a-chips (Alizadeht al., 2000; Buerk, 1993; Weigl et al., 2003). These areas and tech-ologies have enabled biologists to explore the living organismst cellular and molecular levels, and have opened up new oppor-unities to tackle the unsolved biological problems. Advances in
olecular and cell biology have revolutionised the healthcare, andore importantly initiated new exciting interdisciplinary direc-
ions in engineering. In this respect, there is an enormous demando expand the biology-related technologies and industries.
Thanks to engineering devices, considerable research progressas been made in different branches of healthcare. For exam-le, a variety of devices and technologies are being developed foroint-of-care detection, diagnostics, prognostics, and treatment ofhe most complicated diseases such as cancer. In contrast to theonventional methods of cancer diagnostics and prognostics thately on the invasive isolation of cancer tumors and visual inspec-ion of cell morphology, the emerging technologies facilitate theon-invasive scanning of tumor responses such as changes in thectivation or inactivation of specific proteins or expression of spe-ific genes. These approaches realise the profiling of tumors androduce their molecular signatures (Liotta and Petricoin, 2000).
Most of the new emerging technologies deal with cells as theasic structural components of living organisms. For an engineero play an effective role in these fields, the first essential step is tonderstand the basic elements and complexities of cellular biol-gy. However, the main bottleneck in accomplishing this step iso understand the detailed information given in academic cellulariology literature. The issue is that the material covered in the liter-ture best suits the readers with a prior exposure to biology. On thether hand, there exist some resources that describe cellular biol-gy in layman’s terms. But those resources deal with the subject in aanner that does not provide the reader with the necessary details.The objective of this paper is to bring the basic principles of
ellular biology under one umbrella, and explain the structure,unction, and interaction of human cell organelles in a way thatan benefit engineers interested in biology-related researches. Inhis paper, the cell is presented from engineering perspective as a
icro-bio-factory with several process plants. The inlets and out-ets of each plant as well as the plant’s role are discussed throughnumber of flow diagrams.
The paper is organised as follows. Section 2 presents the classifi-ation of the microorganisms to identify the position of the humanell among other cells. Section 3 gives the discovery timeline ofhe human cell organelles emphasising on the role of engineeringnstruments and interdisciplinary research. Section 4 describes theollowing organelles of the human cell: cell membrane, cytoplasm,
ndoplasmic reticulum, Golgi body, Lysosomes, centrosome,itochondria, nucleus and ribosome. Section 5 explains two majorhemical processes occurring in the cell: energy generation androtein synthesis. Finally, concluding remarks are given in Section.
(tmuc
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 329
. The Classification of Microorganisms
Most engineers have heard about the terms such as virus, bac-eria, human cell and plant cell but do not know about theiriological classification and main specifications. In this section,he basic elements of the biological organisms are classified andriefly described. Those elements are known as microorganisms ashey are too small to be seen by human eye. The microorganismsre classified into three groups: non-living agents, prokaryotic cell,nd Eukaryotic cells as shown in Fig. 1. Some references exclude theon-living agents from the microorganisms; however, it is assumed
n this paper that the microorganisms include both living and non-iving groups.
The non-living group refers to the viruses because they arenable to live independently. A virus penetrates a cell and takesontrol of its biological system to flourish and reproduce itself andater attack the other cells. The virus structure consists of a genetic
aterial (DNA or RNA) enclosed by a protein coat called capsid.ome viruses have a more complex structure and may be equippedith an envelope around the capsid or a tail attached to the capsid.
ome diseases caused by viruses include, HIV, Hepatitis-B and fluBecker et al., 2006; Clark, 2005).
The prokaryotic cell group refers to bacteria that live as single-ell microorganisms. The structure of the bacteria is rather simpleith a genetic material (DNA) suspended in a jelly-like material
alled cytoplasm. The cytoplasm is equipped with some tiny par-icles known as ribosomes that produce the needed proteins. Thehole structure is enclosed by the cell envelope that is made up
f two to three layers: a cytoplasmic membrane, a cell wall, and anuter capsule in some species of bacteria. Some bacteria are alsoquipped with hair-like structures called flagella that provide loco-otion. The bacteria exist almost everywhere, in soil, water, air
nd even our bodies. They can adapt themselves with the harshestnvironments of the world such as the acidic hot springs, or in theeepest locations of the earth’s crust. Bacteria could be harmlessnon-pathogenic) or harmful (pathogenic). The E. coli bacteria thatxist in the large intestine of our bodies are normally harmless androduce vitamin K. On the other hand, the Bacillus bacteria thatxist in soil or water are harmful and cause the fatal tuberculosisBecker et al., 2006; Clark, 2005).
The eukaryotic cell group refers to the cells that make up thergans of humans, animals, plants, and fungi. The structure andunction of eukaryotic cells are more complex in comparison tohose of prokaryotic cells. The genetic material (DNA) is enclosedithin a membrane called cell nucleus. Eukaryotic cells also have
ther specialised structures suspended in the cytoplasm. The maintructures are endoplasmic reticulum, Golgi body, Lysosomes, cen-rosome, mitochondria, and ribosome. These are discussed in theollowing sections of this paper. The whole structure is protectedy the cell membrane (Becker et al., 2006; Clark, 2005).
The cells of humans and animals are basically the same. How-ver, the plant cells differ from the human/animal cells because:
i) the membrane of the plant cells is covered with a cellulose wallhat stiffens the cell’s structure and allows the plant to grow tall, (ii)ost of the space within the plant’s cell is occupied by a large vac-ole that stores water, ions and toxins, and (iii) plant cells containhloroplast that is a pigment which determines the colour of the

K. Khoshmanesh et al. / Computational Biology and Chemistry 32 (2008) 315–331 317
on of
pgC
ftwca
t0ic2
3B
totthopdmF
wiWoiirr
orptbpIo11
goTmatcy
Esdlep1cao1
RBlXeUd
toUsing this technique, Claude discovered the ribosome particles in1938 (Claude, 1946). The technique was further utilised by Belgiancytologist and biochemist C.R. Duve to discover the Lysosomes in1956.
Fig. 1. Classificati
lant, and eases the conversion of carbon dioxide and water intolucose and oxygen by absorbing the sunlight (Becker et al., 2006;lark, 2005).
Fungi are more likely to be plants however the fungi cells differrom the plant cells because: (i) the fungi cells lack chloroplast andherefore absorbing food from their surrounding, and (ii) the cellall of the fungi cells is made of a nitrogenous substance known as
hitin. Yeast and mold are among the most famous fungi (Becker etl., 2006; Clark, 2005).
As the microorganisms become more complex, their size andhe number of their ingredient genes increase. A virus is typically.01–0.1 �m across and has around 200 genes, a bacterium is typ-
cally 1–10 �m across and has around 4000 genes, and a humanell is typically 10–100 �m across and poses 25,000 genes (Clark,005).
. Contribution of Engineering to the Evolution of Cellulariology
Cellular biology deals with the cells as the basic elements ofhe living organisms. Cellular biology could be called modern biol-gy as it is younger that the traditional biology which focuses onhe tissues as the structural elements of living organisms. Dueo its characteristics, the rise and evolution of cellular biologyas been more interwoven with two factors: (i) the inventionf engineering instruments, and (ii) the application of interdisci-linary approaches. Those two factors have led the biologists toiscover the cell organelles, and also to develop cell models for-ulating the complicated interactions between them as shown in
ig. 2.In this section, the evolution of cellular biology is reviewed
ith more emphasis on the role of the invention of engineeringnstruments and the application of interdisciplinary approaches.
ith regard to the engineering instruments, the contributionf compound microscopes, electron microscopes, X-ray imag-ng, and differential centrifuge in the growth of cellular biologys reviewed. With respect to interdisciplinary approaches, theole of biochemistry in the advancement of cellular biology iseviewed.
The birth of cellular biology was facilitated by the developmentf microscopes. This is due to the fact that cells are below the limit ofesolution of the human eye and can only be visualised by means ofroper instruments such as microscopes. The microscope has con-ributed more than any other instrument to the development ofiology as a science. In 1590, the first compound microscope was
roduced by the Dutch spectacle-maker Z. Janssen (Croft, 2006).n 1663, the English scientist R. Hooke, discovered cells in a piecef cork, which he examined under his primitive microscope (Ash,998). The value of this discovery was not appreciated until the839, when the cell theory was introduced by German physiolo-
Fa
microorganisms.
ists T. Schwann and M. Schleiden. The cell theory states that allrganisms are composed of similar units of elements called cells.he idea looks simple today, but it established the foundation ofodern biology (Mazzarello, 1999). The emergence of cell theory
nd the invention of more advanced compound microscopes ledhe biologists to discover five cell organelles: cell nucleus, mito-hondria, centrosome, cell membrane, and Golgi body during theears 1833–1897.
The invention of electron microscopes by German engineers. Ruska and M. Knoll in 1931 pushed forward the role of micro-copes in cellular biology. Electron microscopes were developedue to the fact that the magnification of light microscopes was
imited to 1000×. The electron microscope uses a beam of highlynergetic electrons to illuminate and create an image of a sam-le that can provide magnifications of up to 2,000,000× (Ruska,986). Taking advantage of electron microscopes, biologists dis-overed two more cell organelles in 1945: endoplasmic reticulumnd microtubules. They could also identify the traces of ribosomesn the rough endoplasmic reticulum and inside the cytoplasm by958.
The introduction of X-ray imaging by German physicist W.öntgen in 1895 advanced the boundaries of the modern biology.asically X-rays are a type of electromagnetic radiation with wave-
engths of around 10−10 m (Gray and Orton, 2000). Although the-ray imaging was vastly used to detect the bone defects, the sci-ntists applied this method to take the images of the DNA in 1952.sing those images, Watson and Crick put forward the currentouble-helix model of DNA by 1953 (Watson and Crick, 1953).
The development of differential centrifugation technique byhe Belgian biologist A. Claude in the late 1930s helped the biol-gists to sort the cell organelles and discover new components.
ig. 2. The evolution of cellular biology was facilitated by engineering instrumentsnd taking interdisciplinary approaches.

318 K. Khoshmanesh et al. / Computational Biology and Chemistry 32 (2008) 315–331
Table 1The timeline for the first wave of cellular biology evolution with respect to the invention of tools and techniques and the development of cell models
Invention of tools andtechniques
Discovery of cell organelles Development of cell models
Compound microscopeinvented by Janssen (1590)
Cell discovered by Hooke (1633)The biochemistry waspioneered by Wöhler (1828)
Cell nucleus reported by Brown (1833)Cell theory proposed by Schwann and Schleiden (1839)
Mitochondria discovered by Kölliker (1857)DNA isolated by Miescher (1869)The term ‘centrosome’ used by Boveri (1888)
X-ray imaging invented byRöntgen (1895)
Cell membrane distinguished by Overton(1895)Golgi body discovered by Golgi (1897)
Electron microscope built byRuska and Knoll (1931)Differential centrifugationtechnique was developed byClaude (1930s)
Krebs cycle of mitochondria identified by Krebs (1937)
Cytoplasm discussed by Just (1938)Ribosome discovered by Claude (1938)
The role of RNA in protein synthesis discovered by Caspersson, Brachet andSchultz (1939)
Endoplasmic reticulum and microtubulesidentified by Porter, Claude and Fullam (1945)
The role of ribosome in protein synthesis put forwarded by Claude (1946)The current model of DNA structure suggested by Watson and Crick, based onthe X-ray images taken by Franklin (1953)
Lysosomes discovered by Duvé (1956)The location of ribosomes in endoplasmic reticulum and cytoplasm noticed byPalade (1958)Mitochondria ATP synthesis proposed by Mitchell (1961)
anow1ghpottcs
im
mlat
fowflsime
((
eWfwr
titpatpbic
4
cmn
The biochemistry was one of the first interdisciplinarypproaches to discover the world of cells. The biochemistry was pio-eered by the German chemist F. Wöhler, who published a papern the synthesis of urea in 1828 (Morrison and Weiss, 2006). Itas further developed with the discovery of the first enzyme in
833. The evolution of biochemistry led the biologists to isolate theenetic material (DNA) in 1869 (Dahm, 2008). More experimentselped the biologists to uncover two enigmatic processes that takelace in the cell: protein synthesis and energy production. The rolef genetic information carriers (RNA) and ribosomes in protein syn-hesis was uncovered in a series of experiments by 1946. In abouthe same period, the mitochondrial Krebs cycle and the electronhain transport were discovered to reveal the energy productionecret of mitochondria by 1961 (Scheffler, 2000).
Table 1 summarises the evolution of cellular biology from thenvention of microscope through the introduction of the current
odel of cell membrane.The table does not cover the advancements beyond 1972. The
ain reason is that the first wave of the innovations in the cellu-ar biology that tended to the discovery of major cell organellesnd the development of cell models was completed by thatime.
After that milestone, most of the conducted research wasocused on exploring the behaviour of DNA. This second wavef innovations is known as molecular genetics because it dealsith genes as the basic elements of DNA. Molecular genetics was
ourished by developing DNA sequencing (determination of geneequence along DNA polymer) and DNA amplification (produc-ng multiple copies of specific genes) methods. Invention of DNAicroarrays that analyse several thousand genes at the same timenabled the scientists to accomplish the human genome project
feate
The fluid mosaic model of cell membrane proposed by Singer and Nicolson(1972)
identification of all human genes and their sequences) by 2003Liotta and Petricoin, 2000).
Mapping the complete human genome led the biologists tovolve the third wave of innovations that is known as genomics.hilst the genetics deals with the single gene to understand its
unction and response to external factors, the genomics dealsith the all genes to understand their interaction and combined
esponse to external factors (Liotta and Petricoin, 2000).The fourth wave of innovations targeted the proteins and
herefore is called proteomics. The DNA can be regarded as thenformation and instruction database of the cells while the pro-eins can be considered as the task forces or agents of the cell. Theroteins are produced from the genetic instructions stored in DNAnd then sent throughout the cell performing specific functionshrough biochemical reactions. Formulating the behaviour of theroteins is much more complicated than that of the genes. This isecause the genes are stable and common amongst all cells of liv-
ng organisms, while the proteins are variable in response to theonditions influencing the cell (Liotta and Petricoin, 2000).
. Description of Human Cell Organelles
This section presents the structure and function of humanell organelles including: cell membrane, cytoplasm, endoplas-ic reticulum, Golgi body, Lysosomes, centrosome, mitochondria,
ucleus and ribosome. These organelles act as the structural and
unctional elements of the cell, facilitating the activities of differ-nt human tissues and organs. Fig. 3 depicts the physical shapend location of the cell organelles as perceived by biologists. Onhe other hand, Fig. 4 depicts the cell organelles as perceived byngineers. Using appropriate symbols, the cell organelles are repre-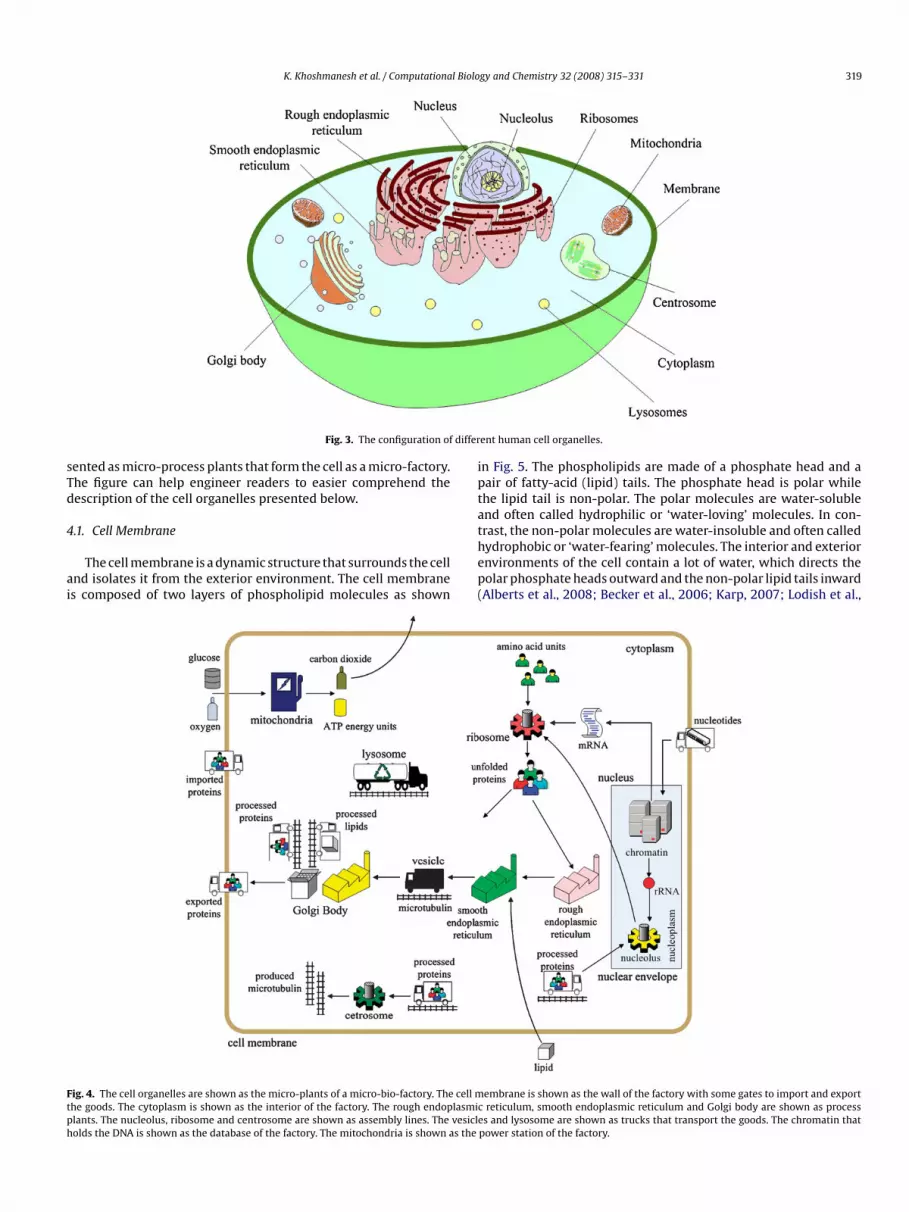
K. Khoshmanesh et al. / Computational Biology and Chemistry 32 (2008) 315–331 319
differ
sTd
4
ai
ipta
Ftph
Fig. 3. The configuration of
ented as micro-process plants that form the cell as a micro-factory.he figure can help engineer readers to easier comprehend theescription of the cell organelles presented below.
.1. Cell Membrane
The cell membrane is a dynamic structure that surrounds the cellnd isolates it from the exterior environment. The cell membranes composed of two layers of phospholipid molecules as shown
thep(
ig. 4. The cell organelles are shown as the micro-plants of a micro-bio-factory. The cell mhe goods. The cytoplasm is shown as the interior of the factory. The rough endoplasmiclants. The nucleolus, ribosome and centrosome are shown as assembly lines. The vesicleolds the DNA is shown as the database of the factory. The mitochondria is shown as the
ent human cell organelles.
n Fig. 5. The phospholipids are made of a phosphate head and aair of fatty-acid (lipid) tails. The phosphate head is polar whilehe lipid tail is non-polar. The polar molecules are water-solublend often called hydrophilic or ‘water-loving’ molecules. In con-
rast, the non-polar molecules are water-insoluble and often calledydrophobic or ‘water-fearing’ molecules. The interior and exteriornvironments of the cell contain a lot of water, which directs theolar phosphate heads outward and the non-polar lipid tails inwardAlberts et al., 2008; Becker et al., 2006; Karp, 2007; Lodish et al.,embrane is shown as the wall of the factory with some gates to import and exportreticulum, smooth endoplasmic reticulum and Golgi body are shown as processs and lysosome are shown as trucks that transport the goods. The chromatin that
power station of the factory.

320 K. Khoshmanesh et al. / Computational Biolo
22low(b
i
v
iicsbatoHbaod2e
4
TcoalaVt
Fig. 5. Structure and different components of the cell membrane.
008; Tobin and Morel, 1997; Voet et al., 2006; Walker and Rapley,000). Several models have been proposed to explain and formu-
ate the cell membrane structure. The basic fluid mosaic modelf the cell membrane is discussed in Singer and Nicolson (1972)hile the evolution of the proposed models is reviewed by Edidin
2003). The main functions and features of the cell membrane cane summarised as follows:
i. The membrane separates the cell organelles from the exteriorenvironment and also acts as a selectively preamble membrane.Small molecules of water, oxygen, and carbon dioxide can passthrough the membrane, while the big molecules of glucose andamino acids cannot do so. For the big molecules, some othermechanisms have been predicted according to their polarities.These mechanisms are described in ii and iii (Alberts et al., 2008;Becker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobin andMorel, 1997; Voet et al., 2006; Walker and Rapley, 2000).
ii. For the big non-polar molecules such as the nutrition particlesor cell waste products, the membrane applies the endocyto-sis and exocytosis mechanisms. In the endocytosis mechanismthat is applied to incoming nutrition particles, the particles arewrapped within a lipid cover called the vesicle and then pushedthrough it. On the other hand, in the exocytosis mechanism thatis applied to outgoing waste products, the vesicle surroundingthe particles is fused with membrane and then pushed throughit (Alberts et al., 2008; Becker et al., 2006; Karp, 2007; Lodish etal., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walker andRapley, 2000).
ii. For the big polar molecules such as glucose, the membrane isequipped with some transport proteins interspersed within thephospholipid layers. The transport proteins have a flexible struc-ture and are reshaped in response to the particle’s shape to graband swallow it (Alberts et al., 2008; Becker et al., 2006; Karp,2007; Lodish et al., 2008; Tobin and Morel, 1997; Voet et al.,2006; Walker and Rapley, 2000).
iv. The movement of the particles through the membrane is drivenby the concentration gradient over the two sides of the mem-brane that is known as diffusion mechanism. However, if theparticles should travel against the concentration gradient, someenergy is consumed to pump them in or out (Alberts et al., 2008;Becker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobin andMorel, 1997; Voet et al., 2006; Walker and Rapley, 2000).
v. Apart from the transport proteins, the membrane also takesadvantage of marker proteins and receptor proteins. The markerproteins or glycoproteins are made of protein and carbohydrate
elements. The protein end is anchored in the membrane whilethe carbohydrate end lays on the external surface of the mem-brane to help the immune system identifying the cell as a friend.Similarly, the receptor proteins lay on the external surface ofthe membrane. The receptor protein is responsible to receivegy and Chemistry 32 (2008) 315–331
the hormones circulating within the blood. The hormones actas information signals within the body and trigger correspond-ing reactions in the cell. For example, the metabolic hormonesthat control the digestion and storage of food may trigger thecorresponding cells to secret digestive enzymes (Alberts et al.,2008; Becker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobinand Morel, 1997; Voet et al., 2006; Walker and Rapley, 2000).
i. The membrane is equipped with cholesterol molecules. Thecholesterol stabilises the fluidity of membrane against tempera-ture variations. At high temperatures, cholesterol decreases themovement of the lipids, making the membrane less fluid andreducing its permeability. At low temperatures, on the otherhand, cholesterol increases the movement of the lipids, main-tains its fluidity preventing membrane from freezing (Albertset al., 2008; Becker et al., 2006; Karp, 2007; Lodish et al., 2008;Tobin and Morel, 1997; Voet et al., 2006; Walker and Rapley,2000). More research is conducted to address the remainedquestions about the structure, dynamic behaviour and functionsof the cell membrane as discussed in Dopico and Tigyi (2007),Fielding (2007), Heimburg (2007), Kusumi et al. (2005), Marguetet al. (2006), Ritchie et al. (2005), Weber et al. (2006) and Wu etal. (2004).
The engineering aspects of the cell membrane have been studiedn different works. The elastic deformation of the cell membranes modelled in Du et al. (2005). The selective permeability of theell membrane is taken as a template to synthesise ion-selectiveupramolecular polymers (Percec and Bera, 2002). Based on theilayer structure of the cell membrane, cylindrical nanofibersre synthesised to facilitate the self-assembly of proteins withinhe damaged tissues. This innovation leads to the developmentf regenerative medicines as discussed in Guler et al. (2006),artgerink et al. (2002) and Stupp et al. (1997). Nanoparticles withiodegradable shells are developed to penetrate the cell membraneccording to the endocytosis mechanism and deliver drugs to therganelles. This technique can revolutionise the therapeutic andiagnostic phases of enigmatic diseases such as cancer (Maysinger,007; Vasir and Labhasetwar, 2007; L. Zhang et al., 2007; B. Zhangt al., 2007).
.2. Cytoplasm
The nucleus of all eukaryotic cells is surrounded by cytoplasm.he cytoplasm is a jelly-like material that holds together theell organelles. The area of the cytoplasm outside the individualrganelles is called cytosol. The cytosol is made of mostly water. Itlso contains some proteins, enzymes, carbohydrates, salts, sugars,ipids, nucleotides, and amino acids (Alberts et al., 2008; Becker etl., 2006; Karp, 2007; Lodish et al., 2008; Tobin and Morel, 1997;oet et al., 2006; Walker and Rapley, 2000). The main functions of
he cytoplasm can be summarised as follows:
i. The molecules that enter the cell membrane should pass throughthe cytosol before entering their final destinations. Therefore,the cytosol acts as a medium between the cell membrane anddifferent cell organelles. In this sense, it may act as a primaryreactor to undertake some chemical processes on the passingmolecules. For example, the cytosol takes the incoming 6-carbonglucose molecules and split them into two 3-carbon moleculesin a process called the glycolysis (Alberts et al., 2008; Becker et
al., 2006; Karp, 2007; Lodish et al., 2008; Tobin and Morel, 1997;Voet et al., 2006; Walker and Rapley, 2000). The reaction of thecytoplasm after the invasion of infectious viruses or bacteria isdiscussed in Yoneyama and Fujita (2007). The cytosol acts as abig reservoir for the materials needed for different processes.
l Biology and Chemistry 32 (2008) 315–331 321
i
bciaaag
4
nwruLa
efcciR
i
ftDt
K. Khoshmanesh et al. / Computationa
For example, in order to produce a protein molecule, the aminoacids that are submerged in the cytosol are collected and laidin a special arrangement in a process called the protein synthe-sis (Alberts et al., 2008; Becker et al., 2006; Karp, 2007; Lodishet al., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walkerand Rapley, 2000). The behaviour and associated mechanismsof cytoplasm during the cancer development can be found inGermain and Frank (2007), Hanahan and Weinberg (2000) andIfergan et al. (2005).
i. The cytosol contains a skeletal structure, called the cytoskele-ton. The cytoskeleton is a dynamic three-dimensional structureof protein fibres. The cytoskeleton has three major elements:microfilaments, microtubules, and intermediate filaments. Themicrofilaments are thin filaments of protein accumulated underthe cell membrane. They stiffen the cell structure and facilitatetiny cellular movements including gliding and contraction. Onthe other hand, the microtubules are hollow tubes consisting ofseveral protein filaments distributed under the microfilaments.The microtubules act as a scaffold to determine the cell shapeand divide the organelles. They also provide a set of tracks forthe movement of organelles and vesicles within the cell. Finally,the intermediate filaments that are made of several proteinfilaments coil around each other to form a rope. The name ‘inter-mediate’ comes from the fact that the diameter of these elementsis bigger than the diameter of microfilaments and smaller thanthe diameter of microtubules. The intermediate filaments pro-vide tensile strength for the cell and help it withstand againstthe mechanical stresses (Alberts et al., 2008; Becker et al., 2006;Karp, 2007; Lodish et al., 2008; Tobin and Morel, 1997; Voet etal., 2006; Walker and Rapley, 2000).
The composite structure of the cytoskeletons and cytosol haseen of great interest to engineering researchers. The mechani-al characteristics of the cytoplasm are comprehensively reviewedn Alt et al. (1997) and Zhu et al. (2000). The elastic deformationsnd the propagation of mechanical stresses through the cytoplasmre analysed in Gladilin et al. (2007), Hu et al. (2005) and Tracquind Ohayon (2004). The hydrodynamic analysis of the cytoplasm isiven in Alt and Dembo (1999) and Yoneda and Nagai (1988).
.3. Endoplasmic Reticulum (ER)
The term ‘endo’ means within and the term ‘reticulum’ meansetwork, thus the endoplasmic reticulum (ER) means a networkithin the cytoplasm. There are two types of ER within the cell:
ough endoplasmic reticulum (RER) and smooth endoplasmic retic-lum (SER) (Alberts et al., 2008; Becker et al., 2006; Karp, 2007;odish et al., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walkernd Rapley, 2000).
With regard to the RER, the membrane of the cell nucleus isxtended at some spots. At those points, the extended membraneorms a series of flattened sacs or sheet-like structures that arealled RER. The sacs are interconnected and spread throughout theell as shown in Fig. 6. They provide enough extended surface tonteract with the cytoplasm. Due to the specific characteristics ofER, its main functions can be summarised as follows:
i. The RER plays a vital role in the process of protein production.Some of the ribosomes employ the extended surface of RERas their unloading docks to discharge their produced proteins.Thousands of ribosomes might bind to the RER at the same time,
therefore its surface looks bumpy or so called ‘rough’ under themicroscope. The other key players of protein synthesis processare introduced in Section 5.2 (Alberts et al., 2008; Becker et al.,2006; Karp, 2007; Lodish et al., 2008; Tobin and Morel, 1997;Voet et al., 2006; Walker and Rapley, 2000).Fig. 6. The structure of the endoplasmic reticulum.
ii. The endoplasmic reticulum (ER) is equipped with chaperonesand folding enzymes that facilitate the folding of protein chains.The chaperones control the quality of the folded proteins toensure that only the correctly folded proteins are exported fromthe ER. The misfolded proteins are separated and later degraded.The amount of proteins that are processed by the ER is setby specific signalling pathways and effector mechanisms. Theupstream signal that activates these pathways is called ER stressand indicates the difference between the amount of the pro-duced protein chains and the ER capacity (Alberts et al., 2008;Becker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobin andMorel, 1997; Voet et al., 2006; Walker and Rapley, 2000). Moredetails about the ER functions including signalling pathways,folding and quality control of proteins are discussed in Hollienand Weissman (2006), Kleizen and Braakman (2004), Marciniakand Ron (2006) and Ron (2002). The disruption of the ER duringthe cancer development that leads to the misfolding of proteinsis discussed in Carracedo et al. (2006), Teiten et al. (2003) andZong et al. (2003).
ii. The space enclosed by the RER membrane is called lumen. Theproteins travel through the lumen to enter the SER. The RERoffers energy-efficient pathways for the transportation and dif-fusion of proteins (Alberts et al., 2008; Becker et al., 2006; Karp,2007; Lodish et al., 2008; Tobin and Morel, 1997; Voet et al.,2006; Walker and Rapley, 2000). Recent studies introduce theER network of neurons as the conveyor and integrator of signalsthat activate the dendrites (Park et al., 2008).
The SER is the extension of RER network. The main structuraleatures of the SER include its network of tubular-like sacs, and alsohe lack of ribosomes on its membrane surfaces as shown in Fig. 6.ue to those specifications the functionality of the SER differs from
he RER as described below:
i. Once the protein leaves the RER, it is sent to the SER to be pack-aged via vesicle and released within the cytoplasm (Alberts etal., 2008; Karp, 2007; Lodish et al., 2008; Tobin and Morel, 1997;Voet et al., 2006; Walker and Rapley, 2000).
ii. The SER is not involved in protein production, but its largeextended surface facilitates other chemical reactions in thecytoplasm. Some of the major chemical reactions that arerealised by the SER include: (a) the synthesis of phosolipidsthat is the essential part of cell membrane, (b) the metabolism
of cholesterol that is a part of cell membrane and also plays akey role in the regulation of some hormones and digestion offats, and (c) the breakdown of glycogen, that is the stored formof glucose, to release energy during the peak times (Lodish et
322 K. Khoshmanesh et al. / Computational Biolo
i
steR
4
ioaaTipTaa1ad(aaetfp(f
i
a
i
4
phtsdpwaLa
Fig. 7. The structure and different components of Golgi body.
al., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walker andRapley, 2000).
ii. The SER detoxifies the drugs and chemical agents by adding OHgroups to them so that they can be dissolved and flushed outof the body easier (Becker et al., 2006; Karp, 2007; Lodish etal., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walker andRapley, 2000).
The SER provides necessary space for the storage of the synthe-ised materials and minerals such as calcium ions that are needed torigger muscle contractions (Alberts et al., 2008; Karp, 2007; Lodisht al., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walker andapley, 2000).
.4. Golgi Body
The Golgi body, also known as Golgi apparatus or Golgi complex,s an arc-shaped organelle that is found nearby the SER. It consistsf five to eight flattened sacs that are covered by lipid membraness shown in Fig. 7. The Golgi sacs can be divided into three regionsccording to their roles: cis-Golgi, medial-Golgi, and trans-Golgi.he cis-Golgi is oriented toward the ER and is responsible for receiv-ng the vesicles that are released by the ER. These vesicles carry theroteins and lipids that are synthesised within the RER and SER.he vesicles fuse with the cis-Golgi membrane, and their contentsre consequently released in the sacs (Becker et al., 2006; Farquharnd Palade, 1981; Karp, 2007; Lodish et al., 2008; Tobin and Morel,997; Voet et al., 2006; Walker and Rapley, 2000). The formationnd assembly of the Golgi body as well as the hypothesis around theynamic behaviour of its structural proteins are discussed in Barr2002), Jiang et al. (2006) and Puri and Linstedt (2003). The proteinsnd lipids then enter the medial-Golgi, where they are processednd modified. To do so, the medial-Golgi is equipped with differ-nt enzymes that add carbohydrates or phosphate supplementarieso incoming molecules. Some recent works on the structural andunctional characteristics of the Golgi body as well as the trans-ortation of proteins are given in Berger and Roth (1997), Cao et al.2000), Cosson et al. (2005) and Hermo and Smith (1998). The mainunctions of the Golgi body can be summarised as follows:
i. The glycoprotein that serves as the marker protein of the cellmembrane is modified while passing through the Golgi body.
The glycoprotein is produced within the ER by adding a car-bohydrate link to the protein. The Golgi enzymes modify thecarbohydrate link by adding and removing some sugar com-pounds (Karp, 2007; Lodish et al., 2008; Tobin and Morel, 1997;Voet et al., 2006; Walker and Rapley, 2000).l
gy and Chemistry 32 (2008) 315–331
ii. The lysosomal protein that serves as the degrading enzymewithin the cell is modified by adding phosphate links to theprotein. Its function is described in Section 4.5 (Alberts et al.,2008; Becker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobinand Morel, 1997; Voet et al., 2006; Walker and Rapley, 2000).
ii. The ceramide that belongs to the lipid family is synthesisedby the Golgi body. The ceramides, phospholipids, and choles-terols are the three lipids that are produced within the ER.The Golgi enzymes add carbohydrate links to the cearmides tomake glycolipids. Alternatively, they add phosphate links to thecearmides to make sphingomyelin. Both of the glycolipids andsphingomyelins are further incorporated in the cell membrane(Becker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobin andMorel, 1997; Voet et al., 2006; Walker and Rapley, 2000).
The proteins and lipids then enter the trans-Golgi, where theyre sorted and packaged within different vesicles as follows:
i. The lysosome vesicles carry the lysosomal proteins and remainwithin the cytosol. Their function is described in Section 4.5(Alberts et al., 2008; Karp, 2007; Lodish et al., 2008; Tobin andMorel, 1997; Voet et al., 2006; Walker and Rapley, 2000).
ii. The membrane vesicles carry the proteins and lipids that areapplied in the structure of the cell membrane. The proteins suchas transport, marker, and receptor proteins as well as the lipidssuch as cholesterol, glycolipid, and sphingomyelin use this path-way to reach the cell membrane. The phospholipid that is themain structural element of the cell membrane is provided by thevesicles as they fuse with the membrane (Becker et al., 2006;Karp, 2007; Lodish et al., 2008; Tobin and Morel, 1997; Voet etal., 2006; Walker and Rapley, 2000).
ii. The secretory vesicles carry the proteins and lipids that aresecreted to the extracellular environment. These vesicles aredivided into constitutive secretion and regulated secretiongroups. The constitutive secretion vesicles move toward thecell membrane and secret their contents as soon as they leavethe trans-Golgi. For example, the liver vesicles that contain theserum albumin proteins continuously secret their proteins intothe blood vessels. On the other hand, the regulated vesicles arestored in the cytosol and are activated when are required. Forexample, the pancreas vesicles that contain digestive enzymesare secreted only after the stomach is filled with food (Alberts etal., 2008; Karp, 2007; Tobin and Morel, 1997; Voet et al., 2006;Walker and Rapley, 2000). The role of the Golgi body in thedevelopment of the Alzheimer disease is explained in Dal Canto(1996) and Hu et al. (2007).
.5. Lysosomes
The lysosomes are small spherical vesicles immersed in the cyto-lasm. The vesicles contain some digestive enzymes known as acidydrolysis. The enzymes have a normal pH level of 4.8, which makeshem effective for selectively dissolving the macromolecules intoeveral parts. In this sense, the lysosomes act as the cell garbageisposal system. The acidic enzymes are produced in the SER andackaged within the vesicles in the Golgi body. As a safety matter,hen the membrane of lysosome is damaged, the acidic enzymes
re disabled (Alberts et al., 2008; Becker et al., 2006; Karp, 2007;odish et al., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walkernd Rapley, 2000). The functions of the lysosomes include the fol-
owing:i. They digest the defective organelles such as mitochondria in aprocess called autophagy. For this, a section of the RER wrapsaround the old organelle as a big vesicle. The lysosomes fuse
K. Khoshmanesh et al. / Computational Biology and Chemistry 32 (2008) 315–331 323
i
i
4
ocnUc
i
4
cplitcicfbdbBSaIm
Fig. 8. The structure of the centrosome.
with the formed vesicle, releasing their digestive enzymes todegrade the worn out organelle. Thus, the nutrients are recy-cled to the cytoplasm and later used within the cell (Albertset al., 2008; Karp, 2007; Kroemer and Jäättelä, 2005; Lodishet al., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walkerand Rapley, 2000). The malfunction or absence of specific diges-tive enzymes leads to the accumulation of specific substanceswithin the cytoplasm and impairing of the cell. The group of dis-eases originated from this problem are called lysosomal storagediseases (Futerman and van Meer, 2004; Rohrbach and Clarke,2007; Settembre et al., 2008).
ii. They degrade the enemies such as bacteria that enter the cell.The enemy particle is trapped within a vesicle. The lysosomesattack the vesicle, releasing their acidic enzymes to degrade theparticle (Alberts et al., 2008; Becker et al., 2006; Karp, 2007;Lodish et al., 2008; Tobin and Morel, 1997; Voet et al., 2006;Walker and Rapley, 2000).
ii. They are involved in the programmed cell death that is essen-tial for the development of different structures and is known asautolysis. To do so, the contents of the lysosomes are deliberatelyreleased to the cytoplasm and the cell digests itself (Becker etal., 2006; Holt et al., 2006; Luzio et al., 2007).
v. They provide a membrane patch to repair the damaged parts ofthe cell membrane (McNeil and Kirchhausen, 2005). Role of thelysosomes in the development and therapy of cancer are dis-cussed in Fehrenbacher and Jättelä (2005), Glunde et al. (2003)and Maclean et al. (2008).
.6. Centrosome
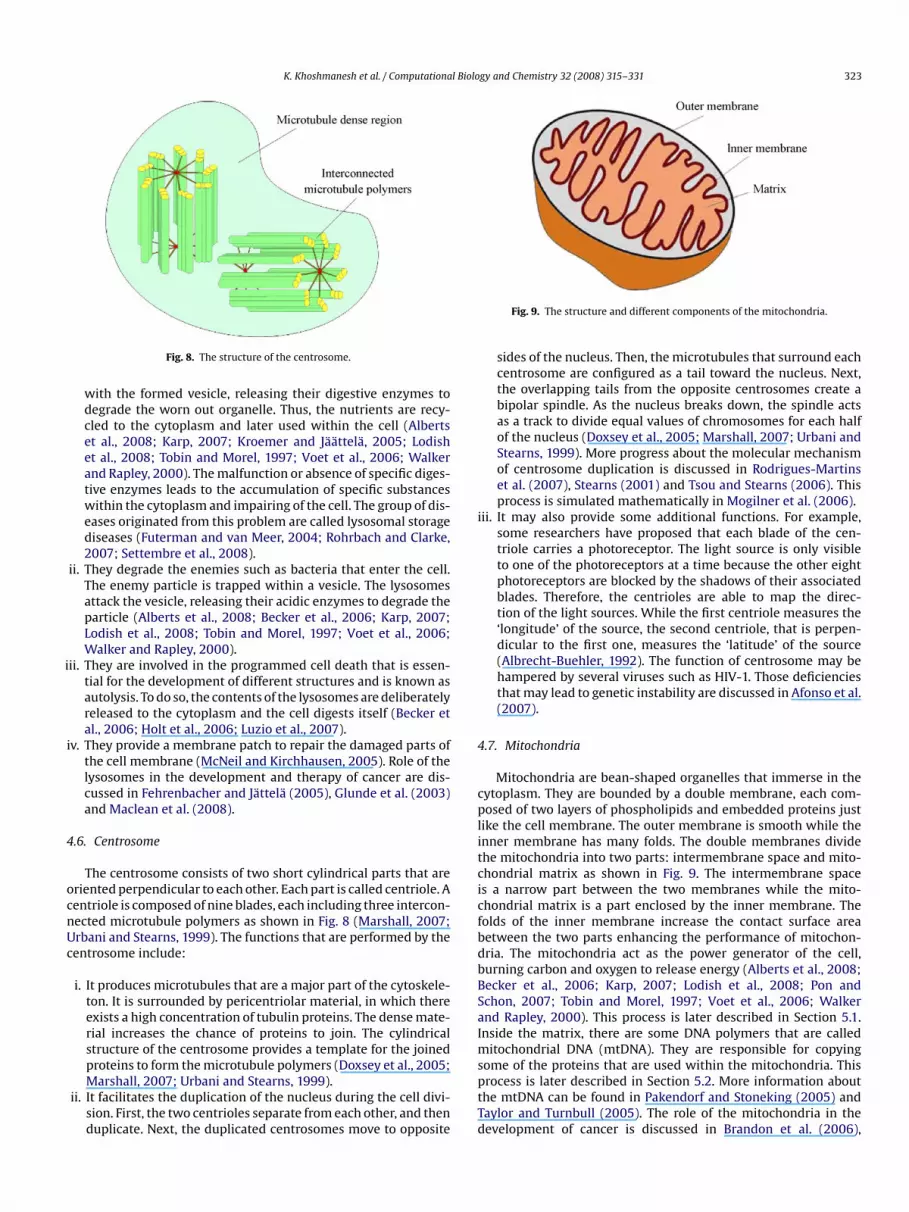
The centrosome consists of two short cylindrical parts that areriented perpendicular to each other. Each part is called centriole. Aentriole is composed of nine blades, each including three intercon-ected microtubule polymers as shown in Fig. 8 (Marshall, 2007;rbani and Stearns, 1999). The functions that are performed by theentrosome include:
i. It produces microtubules that are a major part of the cytoskele-ton. It is surrounded by pericentriolar material, in which thereexists a high concentration of tubulin proteins. The dense mate-rial increases the chance of proteins to join. The cylindricalstructure of the centrosome provides a template for the joined
proteins to form the microtubule polymers (Doxsey et al., 2005;Marshall, 2007; Urbani and Stearns, 1999).ii. It facilitates the duplication of the nucleus during the cell divi-sion. First, the two centrioles separate from each other, and thenduplicate. Next, the duplicated centrosomes move to opposite
sptTd
Fig. 9. The structure and different components of the mitochondria.
sides of the nucleus. Then, the microtubules that surround eachcentrosome are configured as a tail toward the nucleus. Next,the overlapping tails from the opposite centrosomes create abipolar spindle. As the nucleus breaks down, the spindle actsas a track to divide equal values of chromosomes for each halfof the nucleus (Doxsey et al., 2005; Marshall, 2007; Urbani andStearns, 1999). More progress about the molecular mechanismof centrosome duplication is discussed in Rodrigues-Martinset al. (2007), Stearns (2001) and Tsou and Stearns (2006). Thisprocess is simulated mathematically in Mogilner et al. (2006).
ii. It may also provide some additional functions. For example,some researchers have proposed that each blade of the cen-triole carries a photoreceptor. The light source is only visibleto one of the photoreceptors at a time because the other eightphotoreceptors are blocked by the shadows of their associatedblades. Therefore, the centrioles are able to map the direc-tion of the light sources. While the first centriole measures the‘longitude’ of the source, the second centriole, that is perpen-dicular to the first one, measures the ‘latitude’ of the source(Albrecht-Buehler, 1992). The function of centrosome may behampered by several viruses such as HIV-1. Those deficienciesthat may lead to genetic instability are discussed in Afonso et al.(2007).
.7. Mitochondria
Mitochondria are bean-shaped organelles that immerse in theytoplasm. They are bounded by a double membrane, each com-osed of two layers of phospholipids and embedded proteins just
ike the cell membrane. The outer membrane is smooth while thenner membrane has many folds. The double membranes dividehe mitochondria into two parts: intermembrane space and mito-hondrial matrix as shown in Fig. 9. The intermembrane spaces a narrow part between the two membranes while the mito-hondrial matrix is a part enclosed by the inner membrane. Theolds of the inner membrane increase the contact surface areaetween the two parts enhancing the performance of mitochon-ria. The mitochondria act as the power generator of the cell,urning carbon and oxygen to release energy (Alberts et al., 2008;ecker et al., 2006; Karp, 2007; Lodish et al., 2008; Pon andchon, 2007; Tobin and Morel, 1997; Voet et al., 2006; Walkernd Rapley, 2000). This process is later described in Section 5.1.nside the matrix, there are some DNA polymers that are called
itochondrial DNA (mtDNA). They are responsible for copyingome of the proteins that are used within the mitochondria. This
rocess is later described in Section 5.2. More information abouthe mtDNA can be found in Pakendorf and Stoneking (2005) andaylor and Turnbull (2005). The role of the mitochondria in theevelopment of cancer is discussed in Brandon et al. (2006),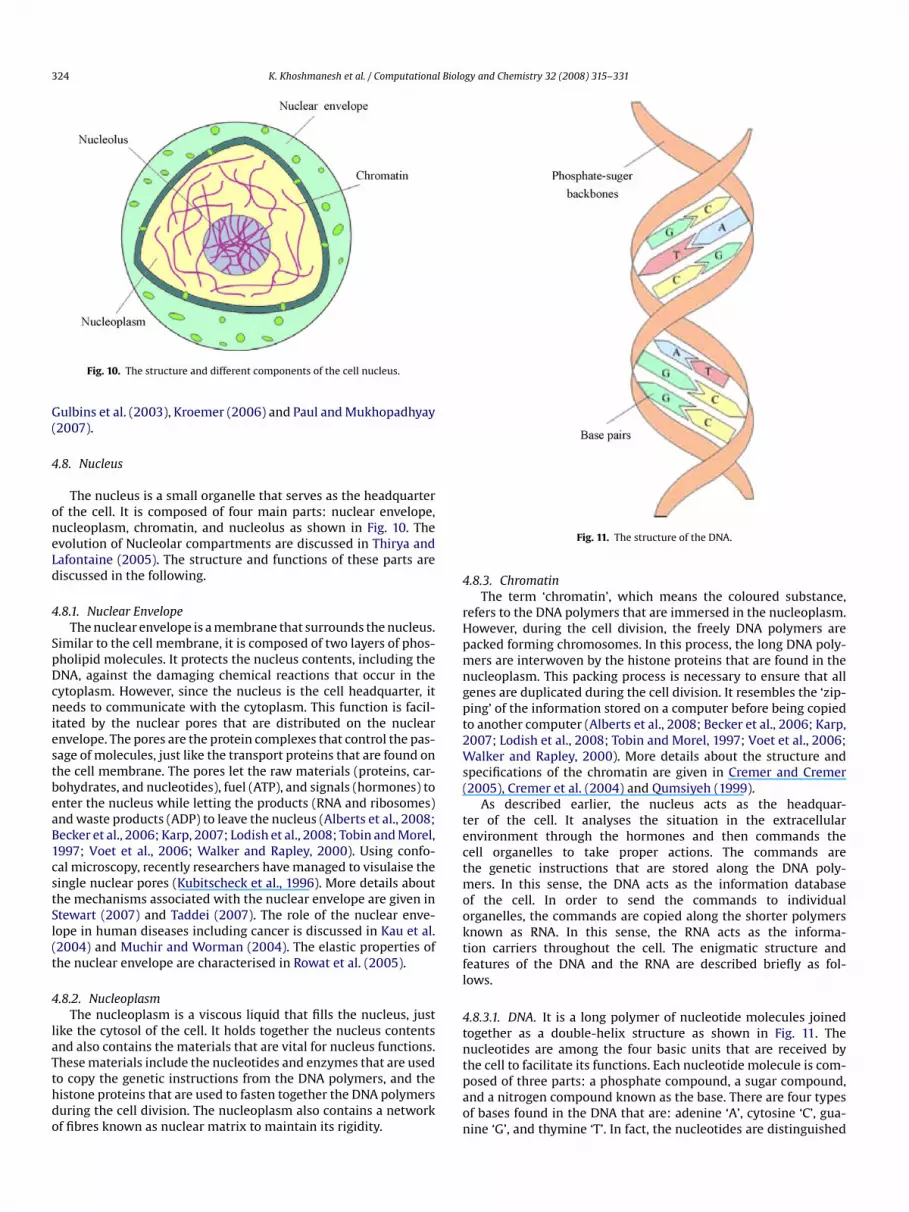
324 K. Khoshmanesh et al. / Computational Biology and Chemistry 32 (2008) 315–331
G(
4
oneLd
4
SpDcniestbeaB1cstSl(t
4
laTthdo
4
rHpmngpt2Ws(
tectmooktfl
4tn
Fig. 10. The structure and different components of the cell nucleus.
ulbins et al. (2003), Kroemer (2006) and Paul and Mukhopadhyay2007).
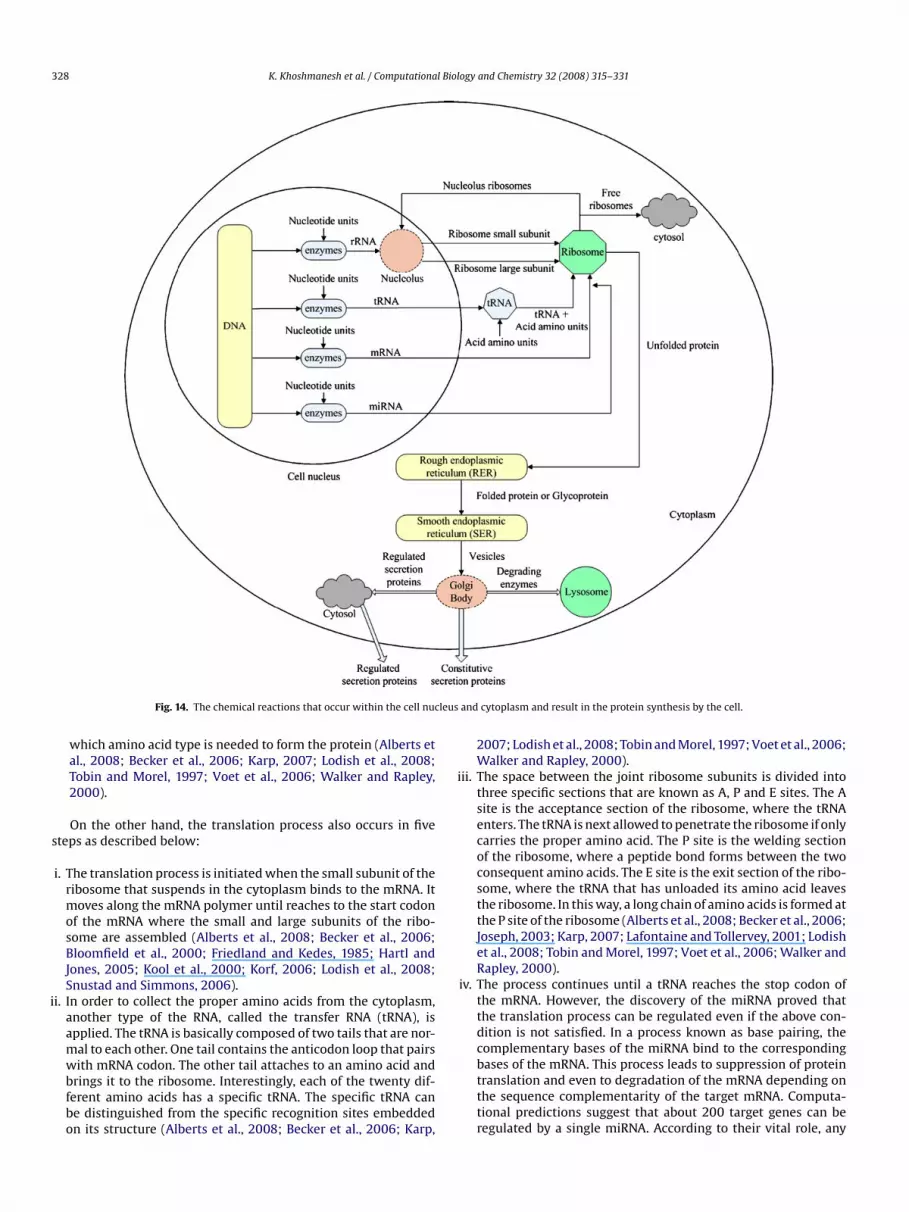
.8. Nucleus
The nucleus is a small organelle that serves as the headquarterf the cell. It is composed of four main parts: nuclear envelope,ucleoplasm, chromatin, and nucleolus as shown in Fig. 10. Thevolution of Nucleolar compartments are discussed in Thirya andafontaine (2005). The structure and functions of these parts areiscussed in the following.
.8.1. Nuclear EnvelopeThe nuclear envelope is a membrane that surrounds the nucleus.
imilar to the cell membrane, it is composed of two layers of phos-holipid molecules. It protects the nucleus contents, including theNA, against the damaging chemical reactions that occur in theytoplasm. However, since the nucleus is the cell headquarter, iteeds to communicate with the cytoplasm. This function is facil-
tated by the nuclear pores that are distributed on the nuclearnvelope. The pores are the protein complexes that control the pas-age of molecules, just like the transport proteins that are found onhe cell membrane. The pores let the raw materials (proteins, car-ohydrates, and nucleotides), fuel (ATP), and signals (hormones) tonter the nucleus while letting the products (RNA and ribosomes)nd waste products (ADP) to leave the nucleus (Alberts et al., 2008;ecker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobin and Morel,997; Voet et al., 2006; Walker and Rapley, 2000). Using confo-al microscopy, recently researchers have managed to visulaise theingle nuclear pores (Kubitscheck et al., 1996). More details abouthe mechanisms associated with the nuclear envelope are given intewart (2007) and Taddei (2007). The role of the nuclear enve-ope in human diseases including cancer is discussed in Kau et al.2004) and Muchir and Worman (2004). The elastic properties ofhe nuclear envelope are characterised in Rowat et al. (2005).
.8.2. NucleoplasmThe nucleoplasm is a viscous liquid that fills the nucleus, just
ike the cytosol of the cell. It holds together the nucleus contentsnd also contains the materials that are vital for nucleus functions.
hese materials include the nucleotides and enzymes that are usedo copy the genetic instructions from the DNA polymers, and theistone proteins that are used to fasten together the DNA polymersuring the cell division. The nucleoplasm also contains a networkf fibres known as nuclear matrix to maintain its rigidity.tpaon
Fig. 11. The structure of the DNA.
.8.3. ChromatinThe term ‘chromatin’, which means the coloured substance,
efers to the DNA polymers that are immersed in the nucleoplasm.owever, during the cell division, the freely DNA polymers areacked forming chromosomes. In this process, the long DNA poly-ers are interwoven by the histone proteins that are found in the
ucleoplasm. This packing process is necessary to ensure that allenes are duplicated during the cell division. It resembles the ‘zip-ing’ of the information stored on a computer before being copiedo another computer (Alberts et al., 2008; Becker et al., 2006; Karp,007; Lodish et al., 2008; Tobin and Morel, 1997; Voet et al., 2006;alker and Rapley, 2000). More details about the structure and
pecifications of the chromatin are given in Cremer and Cremer2005), Cremer et al. (2004) and Qumsiyeh (1999).
As described earlier, the nucleus acts as the headquar-er of the cell. It analyses the situation in the extracellularnvironment through the hormones and then commands theell organelles to take proper actions. The commands arehe genetic instructions that are stored along the DNA poly-
ers. In this sense, the DNA acts as the information databasef the cell. In order to send the commands to individualrganelles, the commands are copied along the shorter polymersnown as RNA. In this sense, the RNA acts as the informa-ion carriers throughout the cell. The enigmatic structure andeatures of the DNA and the RNA are described briefly as fol-ows.
.8.3.1. DNA. It is a long polymer of nucleotide molecules joinedogether as a double-helix structure as shown in Fig. 11. Theucleotides are among the four basic units that are received byhe cell to facilitate its functions. Each nucleotide molecule is com-
osed of three parts: a phosphate compound, a sugar compound,nd a nitrogen compound known as the base. There are four typesf bases found in the DNA that are: adenine ‘A’, cytosine ‘C’, gua-ine ‘G’, and thymine ‘T’. In fact, the nucleotides are distinguished
l Biolo
tptsptdawobgcee2hCUht(
io(h(gtZ
4oasmtwbtRrttpa2tKg1tpt2eRi
4
lna
DitsscaZpc
4
ctrtis(22
dasarwtf‘edittbSBe2
5
wtdqpcim
5
e
K. Khoshmanesh et al. / Computationa
hrough their bases. In order to create a DNA polymer, the sugar andhosphate compounds of nearby nucleotides attach to each othero form a single strand. The bases that are arranged along the singletrand then recall their pairs to form the second strand. The ‘A’ basesair with the ‘T’ bases while the ‘C’ bases pair with the ‘G’ bases. Thewo attached strands then are twisted around each other to form aouble-helix structure known as the DNA. If the bases are regardeds genetic letters, then the three consequent bases make a geneticord, and finally the neighbouring words make a genetic sentence
r the so called gene. For example, the sequence of ATG TTA CTC GTAases can represent the gene that specifies the colour of the hair. Theenes contain the genetic instructions that are used to control theell activities (Alberts et al., 2008; Becker et al., 2006; Bloomfieldt al., 2000; Friedland and Kedes, 1985; Hartl and Jones, 2005; Koolt al., 2000; Korf, 2006; Lodish et al., 2008; Snustad and Simmons,006). Most of the human genes have been identified during theuman genome project (International Human Genome Sequencingonsortium, 2001; The International HapMap Consortium, 2005).sing DNA microarrays, the parallel examination of several genesas become possible (Jordan, 2001; Rinaldis and Lahm, 2007) andherefore the molecular profiling of cancer cells has been realisedEsteller, 2007).
The unique helical shape of the DNA has inspired some engineer-ng studies as follows. The mechanical and structural characteristicsf the helical DNA are analysed in Bao and Suresh (2003), Marko2005) and Travers and Thompson (2004). A water turbine withelical blades is reported to harness the energy of gulf streamsGorlov, 1995, 1998). The unique capability of the DNA to store theenetic information has inspired the idea of molecular informa-ion storage and processing systems (Ezziane, 2006; Shapiro, 2006;auner, 2005).
.8.3.2. RNA. It is copied from the DNA to facilitate the cellularperations. RNA is similar to the DNA in many ways. However, therere some differences between them as follows: (i) The RNA is muchhorter than the DNA as it carries only a small portion of the infor-ation stored on the DNA. The DNA is as big as the hard disk while
he RNA is as big as the flash memory. (ii) The RNA is single-strandedhile the DNA is double-stranded. (iii) The RNA includes uracil ‘U’
ase while the DNA contains the thymine ‘T’ base. Conventionally,he RNA polymers are classified into three major groups: ribosomalNA (rRNA), transfer RNA (tRNA), and messenger RNA (mRNA). TheRNA travels to the nucleolus to produce the ribosome particles, theRNA travels to the cytoplasm to gather the pair amino acids andransfer them to the ribosomes, and the mRNA travels to the cyto-lasm to produce the proteins (Bloomfield et al., 2000; Friedlandnd Kedes, 1985; Hartl and Jones, 2005; Kool et al., 2000; Korf,006; Snustad and Simmons, 2006). Recent studies about the func-ions of these RNAs can be found in Borukhov and Nudler (2008),öhler and Hurt (2007) and Mcnamara (2006). However, anotherroup of RNAs that is called microRNA (miRNA) was discovered5 years ago. The miRNA regulates protein production by bindingo the mRNA inside the cytoplasm. After binding, the process ofrotein production by the mRNA is inhibited and in some caseshe mRNA is degraded (Bushati and Cohen, 2007; Lee and Ambros,001; Pillai, 2005; Kent and Mendell, 2006; Kosik, 2006; L. Zhangt al., 2007; B. Zhang et al., 2007). The function of the describedNA groups is described in Section 5.2, where the protein synthesis
s discussed.
.8.4. NucleolusNucleolus is a spherical body within the nucleus. It is membrane
ess and surrounded by the chromatins. The main function of theucleolus is to produce the ribosome particles. This function is initi-ted by be copying of the ribosomal RNA (rRNA) polymers from the
ictdo
gy and Chemistry 32 (2008) 315–331 325
NA. The nucleolus takes the rRNA as well as the proteins that aremmersed in the nucleoplasm to make ribosomes. The ribosomeshen leave the nucleus through the nuclear pores (recent researchesuggest that the nucleolus has some non-conventional functionsuch as the assembly of various kinds of ribonucleoprotein parti-les, the modification of small RNAs, the control of the cell cyclend the nuclear export processes; Boisvert et al., 2007; Schmidt-achmann, 2005). Moreover, the recent investigations indicate aossible connection between the malfunction of the nucleolus andell aging (Guarente, 2007; Scaffi di et al., 2005).
.9. Ribosome
The ribosomes are small organelles that immerse within theytoplasm. They are composed of two parts: the small subunit andhe large subunit. The two parts are suspended in the cytosol sepa-ately; however they join and behave as a single entity facilitatinghe protein synthesis. Each subunit consists of rRNA and protein. Its believed that the rRNA component handles the protein synthe-is while the protein component stabilises the ribosome structureAlberts et al., 2008; Becker et al., 2006; Karp, 2007; Lodish et al.,008; Tobin and Morel, 1997; Voet et al., 2006; Walker and Rapley,000).
The ribosomes have a major role in the protein synthesis asescribed in Section 5.2. They hold the single amino acid moleculesnd weld them together to form a chain of amino acids, or theo called protein. The ribosomes can be divided into two groupsccording to their produced proteins: free ribosomes and boundibosomes. The free ribosomes produce the ‘domestic’ proteins,hich are consumed by the cell organelles to facilitate their func-
ions. These proteins are released within the cytoplasm withouturther processing. In contrast, the bound ribosomes produce theforeign export’ proteins, which are secreted to the extracellularnvironment via the cell membrane. These proteins need moreelicate packaging and therefore should undergo further process-
ng. Once the bound ribosomes produce these proteins, they attacho the RER membrane. The proteins penetrate the RER interspacehrough the translocation proteins embedded on the RER mem-rane. Then the proteins experience a long journey through RER,ER, and Golgi body before being secretion (Alberts et al., 2008;ecker et al., 2006; Berk et al., 2006; Karp, 2007; Liljas, 2004; Lodisht al., 2008; Stephens, 2005; Tobin and Morel, 1997; Voet et al.,006; Walker and Rapley, 2000).
. Description of Cells’ Major Chemical Processes
This section presents two major chemical processes that occurithin the cell. They are the energy generation process and the pro-
ein synthesis process. Despite the comprehensive research workevoted to exploring these enigmatic processes, there are manyuestions yet to be answered. Different cell organelles, or differentarts of an individual cell organelle, interact to realise each pro-ess. In the following, these complex interactions are translatednto simple flow diagrams, generally used by engineers to deal with
ulti-component processes.
.1. Energy Generation
Energy generation is performed by mitochondria. Like any otherngine, it burns carbon and oxygen to release energy. The carbon
s mainly carried to the cell as glucose, a 6-carbon sugar with thehemical formula of C6H12O2 while the air is carried to the cell viahe blood vessels. The products of the burning process are carbonioxide and water molecules. These are consumed within the cell,r leave the cell via blood vessels. The energy that is released during
3 l Biolo
thimp(M2
C
A
aka
i
aGa
26 K. Khoshmanesh et al. / Computationa
his reaction is stored as adenosine triphosphate (ATP) molecule, aeavy compound of adenine, ribose and phosphate with the chem-
cal formula of C10H16N5O13P3. When energy is needed, each ATPolecule loses one of its phosphates to form adenosine diphos-
hate (ADP) molecule and release 7.3 kcal of energy as followsBerg et al., 2007; Champe et al., 2005; Garrett and Grisham, 2007;
athews et al., 2000; Nelson and Cox, 2005; Pratt and Cornely,004):
6H12O2 + 6O2 → 6CO2 + 6H2O + 8ATP (1)
TP + H2O → ADP + PO43− + 7.3 kcal (2)
The process of energy generation in the cell is not as simples described above. It occurs in a series of biochemical reactionsnown as glycolysis, Krebs cycle, and electron transport chain thatre explained below:
i. The glycolysis reaction occurs within the cytosol. During thisreaction, the 6-carbon glucose molecule is broken into twohalves and converted to two 3-carbon molecules known as pyru-vate (C3H4O3). To facilitate this process, the cytoplasm employstwo special chemical compounds known as cytoplasm enzymesand cytoplasm coenzymes. The cytoplasm enzymes are the largeprotein molecules that act as catalysers and speed up the reac-tions. On the other hand, the cytoplasm coenzymes are thenon-protein molecules that assist the enzymes. The vitaminsare among the most well known coenzymes. They often serveas intermediate carriers of electrons, specific atoms or func-tional groups that are exchanged during the overall reaction. Thecontributing coenzyme involved in the process is nicotinamideadenine dinucleotide (NAD+), which resumes its role in Krebscycle, as well as electron transport chain. The overall reactionof glycolysis can be summarised as follows (Berg et al., 2007;Champe et al., 2005; Garrett and Grisham, 2007; Mathews etal., 2000; Nelson and Cox, 2005; Pratt and Cornely, 2004):
(3)
ii. The Krebs cycle reactions occur in the matrix of the mitochon-dria. The pyruvate molecules that were produced during theglycolysis enter the matrix to feed the cycle. The cycle involvesthe periodic composition and decomposition of 6-, 5- and 4-carbon molecules, which results in the gradual releasing ofpyruvate energy content. The cycle reactions can be divided intofour steps as follows (Berg et al., 2007; Champe et al., 2005;Garrett and Grisham, 2007; Mathews et al., 2000; Nelson andCox, 2005; Pratt and Cornely, 2004):(a) To initiate the cycle, the 3-carbon pyruvate molecule loses
a carbon dioxide and is converted into a 2-carbon moleculeof acetyl group. The acetyl reacts with a matrix coenzymethat is called coenzyme-A to form acetyl coenzyme-A.
(b) The acetyl coenzyme-A reacts with a 4-carbon molecule ofoxaloacetate that exists in the matrix to form a 6-carbonmolecule of citrate.
(c) The citrate loses a carbon dioxide to form a 5-carbon
molecule of �-ketoglutarate.(d) The �-ketoglutarate loses the last carbon dioxide to repro-duce the 4-carbon molecule of oxaloacetate and restart thecycle. The contributing coenzymes involved in the processare NAD+ and flavin adenine dinucleotide (FAD).
gy and Chemistry 32 (2008) 315–331
The overall reaction of Krebs cycle can be summarised as fol-lows:
(4)
ii. The electron transport chain reactions occur along the innermembrane of the mitochondria. The route of the inner mem-brane is windy to provide a large surface area, and moreover isequipped with protein complexes and electron shuttles to facil-itate its performance. The reactions can be divided into six stepsas follows (Berg et al., 2007; Champe et al., 2005; Garrett andGrisham, 2007; Mathews et al., 2000; Nelson and Cox, 2005;Pratt and Cornely, 2004):(a) The reactions are initiated by the coenzymes such as NADH
and FADH2 that were produced through the glycolysis andKrebs cycle reactions. To do so, the coenzymes donate theirelectrons to the protein complexes of the inner membrane.
(b) The electrons pass through the protein complexes andrelease some of their internal energy. This energy triggersthe protein complexes to pump out some H+ ions from thematrix to the intermembrane space of the mitochondria.
(c) The electrons travel between the protein complexes via theelectron shuttles. The process of pumping out H+ ions con-tinues until the energy level of the electrons drops below acertain value. At that point the electrons return to the matrix.The electrons donated by NADH can pass through three pro-tein complexes while the electrons donated by FADH2 canonly pass through two protein complexes.
(d) Once the electrons return to the matrix, they are absorbedby the oxygen molecules. The oxygen molecules becomecharged and the H+ ions of the matrix gather around them toreceive the electrons. A chemical reaction occurs between H+
and oxygen that results in the formation of H2O. The reactionis associated with releasing some energy that is consumedto pump out more H+ ions from the matrix.
(e) The consequent pumping out of H+ ions increases the con-centration gradient of positive ions over the two sides of theinner membrane. The gradient then pumps in the H+ ionsto the matrix through some specific proteins know as ATPsynthesis complexes.
(f) The ATP complexes act as hydroelectric turbines. They har-ness the energy of the flowing H+ ions and consume it toweld the ADP molecules and organic phosphates that existwithin the matrix. The result is the ATP molecule that acts asthe energy carrier throughout the cell.
The overall reaction of electron transport chain be summariseds follows (Berg et al., 2007; Champe et al., 2005; Garrett andrisham, 2007; Mathews et al., 2000; Nelson and Cox, 2005; Prattnd Cornely, 2004):
(5)

K. Khoshmanesh et al. / Computational Biology and Chemistry 32 (2008) 315–331 327
ria th
Kg
i(MefDdGemin
5
sbnstmstt
i
i
stranded, and contains the uracil ‘U’ bases instead of the thymine‘T’ bases. The sequence of each three neighbouring bases of themRNA is called the codon. There are twenty different types ofamino acids available within the cytoplasm. The codon indicates
Fig. 12. Chemical reactions of the mitochond
Fig. 12 integrates the chemical reactions that occur in glycolysis,rebs cycle, and electron transport chain and result in the energyeneration by the cell.
Several models have been developed to simulate the biochem-cal reactions happening within the cytoplasm and mitochondriaBanaji, 2006; Bertrama et al., 2006; Loebl and Matthai, 2004;
oser et al., 2006; Peterson et al., 2007; Wu et al., 2007). Fromngineering perspective, the mitochondria is an enzymatic bio-uel cell that generates electric power from the burning of glucose.esign and fabrication of biofuel cells that mimic the mitochon-ria is reported in Barrière et al. (2006), Furukawa et al. (2006),ao et al. (2007), Moore et al. (2005), Togo et al. (2007) and Zhangt al. (2006). The simplicity and biocompatibility of these systemsake them good candidates to serve as the power source of the
n vivo bioelectronic devices such as micropumps, pacemakers andeuromorphic circuits.
.2. Protein Synthesis
The process of protein production by the cell is called proteinynthesis. A protein is a long chain of amino acids that are attachedy peptide bonds. In order to produce a protein, three elements areeeded: (i) a genetic template to recall the appropriate amino acidubunits from the cytoplasm, (ii) the amino acid subunits that formhe chain, and (iii) a machine to join and weld the amino acids to
ake a peptide bond. The first element is provided trough the tran-cription process, while the second and third elements are providedhrough the translation process. The transcription process occurs inhe following four steps:
i. The transcription process is initiated when an enzyme, calledRNA polymerase-II, binds to a specific region of the DNA calledthe promoter region. The enzyme then unwinds the joint strandsof DNA at that region (Alberts et al., 2008; Becker et al., 2006;
Bloomfield et al., 2000; Friedland and Kedes, 1985; Hartl andJones, 2005; Kool et al., 2000; Korf, 2006; Lodish et al., 2008;Snustad and Simmons, 2006).ii. The enzyme reads the genetic sequence of the single-strandedDNA, recalls the proper nucleotide subunits from the nucleo-
Fl
at tend to energy generation within the cell.
plasm, and put them in a row. The ‘C’ bases pair with the ‘G’ baseswhile the ‘A’ bases prefer to pair with the ‘U’ bases (Alberts et al.,2008; Becker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobinand Morel, 1997; Voet et al., 2006; Walker and Rapley, 2000).
ii. The process continues until the enzyme reaches the terminationsequence of the DNA, where it leaves the DNA (Alberts et al.,2008; Becker et al., 2006; Karp, 2007; Lodish et al., 2008; Tobinand Morel, 1997; Voet et al., 2006; Walker and Rapley, 2000).
v. The outcome of this process is the formation of a messen-ger RNA (mRNA) polymer that serves as an information carrierthroughout the cell. The mRNA migrates from the nucleus tothe cytoplasm. The mRNA is similar to DNA but is shorter, single
ig. 13. The merging of small and large subunits of ribosome to facilitate the trans-ation process.
328 K. Khoshmanesh et al. / Computational Biology and Chemistry 32 (2008) 315–331
leus a
s
i
ii
iv
Fig. 14. The chemical reactions that occur within the cell nuc
which amino acid type is needed to form the protein (Alberts etal., 2008; Becker et al., 2006; Karp, 2007; Lodish et al., 2008;Tobin and Morel, 1997; Voet et al., 2006; Walker and Rapley,2000).
On the other hand, the translation process also occurs in fiveteps as described below:
i. The translation process is initiated when the small subunit of theribosome that suspends in the cytoplasm binds to the mRNA. Itmoves along the mRNA polymer until reaches to the start codonof the mRNA where the small and large subunits of the ribo-some are assembled (Alberts et al., 2008; Becker et al., 2006;Bloomfield et al., 2000; Friedland and Kedes, 1985; Hartl andJones, 2005; Kool et al., 2000; Korf, 2006; Lodish et al., 2008;Snustad and Simmons, 2006).
i. In order to collect the proper amino acids from the cytoplasm,another type of the RNA, called the transfer RNA (tRNA), isapplied. The tRNA is basically composed of two tails that are nor-mal to each other. One tail contains the anticodon loop that pairs
with mRNA codon. The other tail attaches to an amino acid andbrings it to the ribosome. Interestingly, each of the twenty dif-ferent amino acids has a specific tRNA. The specific tRNA canbe distinguished from the specific recognition sites embeddedon its structure (Alberts et al., 2008; Becker et al., 2006; Karp,nd cytoplasm and result in the protein synthesis by the cell.
2007; Lodish et al., 2008; Tobin and Morel, 1997; Voet et al., 2006;Walker and Rapley, 2000).
i. The space between the joint ribosome subunits is divided intothree specific sections that are known as A, P and E sites. The Asite is the acceptance section of the ribosome, where the tRNAenters. The tRNA is next allowed to penetrate the ribosome if onlycarries the proper amino acid. The P site is the welding sectionof the ribosome, where a peptide bond forms between the twoconsequent amino acids. The E site is the exit section of the ribo-some, where the tRNA that has unloaded its amino acid leavesthe ribosome. In this way, a long chain of amino acids is formed atthe P site of the ribosome (Alberts et al., 2008; Becker et al., 2006;Joseph, 2003; Karp, 2007; Lafontaine and Tollervey, 2001; Lodishet al., 2008; Tobin and Morel, 1997; Voet et al., 2006; Walker andRapley, 2000).
. The process continues until a tRNA reaches the stop codon ofthe mRNA. However, the discovery of the miRNA proved thatthe translation process can be regulated even if the above con-dition is not satisfied. In a process known as base pairing, thecomplementary bases of the miRNA bind to the corresponding
bases of the mRNA. This process leads to suppression of proteintranslation and even to degradation of the mRNA depending onthe sequence complementarity of the target mRNA. Computa-tional predictions suggest that about 200 target genes can beregulated by a single miRNA. According to their vital role, any
l Biolo
v
rFnc
RnsD2O
6
oaieafbcFiptswb
R
A
A
A
A
A
AA
AB
B
B
B
B
B
BB
B
B
B
B
B
B
B
B
B
C
C
C
CC
C
C
C
C
D
D
D
D
D
E
E
E
F
F
F
F
F
K. Khoshmanesh et al. / Computationa
misregulation of the miRNA could lead to great regulatory dis-orders in the cell and may cause cancer or neuronal diseases(Bushati and Cohen, 2007; Lee and Ambros, 2001; Pillai, 2005;Kent and Mendell, 2006; Kosik, 2006; L. Zhang et al., 2007; B.Zhang et al., 2007).
. After stopping the mRNA, two different circumstances are pos-sible for the ribosome: (a) if the produced protein is consumablewithin the cell the chain of amino acids leaves the ribosome, (b)if the product protein is consumable out of the cell the chainof amino acids is unloaded on the RER membrane for furtherprocess. In either case, the ribosome subunits disassemble inthe end (Joseph, 2003; Lafontaine and Tollervey, 2001). Usingcryo-electron microscopy, the dynamic structure of the ribosomeduring the different steps of translation has been studied in Mitraand Frank (2006).
Fig. 13 shows the merging of the small and large subunits of theibosome to facilitate the translation process. On the other hand,ig. 14 integrates the chemical reactions that occur within the cellucleus and cytoplasm and result in the protein synthesis by theell.
From engineering perspective, the enigmatic interaction of DNA,NA, proteins and other organelles is a sophisticated informationetwork. Several models been developed to simulate the proteinynthesis and map the genetic regulatory networks (Bolouri andavidson, 2002; Fisher and Henzinger, 2007; Schlitt and Brazma,006; Skjøndal-Bar and Morris, 2007; Walhout, 2006; Weitzke andrtoleva, 2003; Yeo et al., 2005).
. Conclusion
The paper aimed to facilitate the comprehension of cellular biol-gy for engineers. In this respect, first the position of human cellmong microorganisms was stated and its main features were clar-fied. Then, the evolution of cellular biology was reviewed with anmphasis on the contributions of engineering instruments and thepplication of interdisciplinary approaches. Next, the structure andunction of different human cell organelles, including cell mem-rane, cytoplasm, endoplasmic reticulum, Golgi body, Lysosomes,entrosome, mitochondria, nucleus and ribosome, were described.inally, the interaction of different cell organelles or parts of anndividual cell organelle, realising the two complicated chemicalrocesses within the cell, were reviewed. Those processes includehe energy generation and protein synthesis that are crucial forurvival and routine activities of the cell. The involved interactionsere translated into simple flow diagrams that are generally used
y engineers to represent multi-component processes.
eferences
fonso, P.V., Zamborlini, A., Saïb, A., Mahieux, R., 2007. Centrosome and retroviruses:the dangerous liaisons. Retrovirology 4.
lberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., Walter, P., 2008. MolecularBiology of the Cell, fifth ed. Garland Science, New York.
lbrecht-Buehler, G., 1992. Speculation about the function and formation of centri-oles and basal bodies. In: The Centrosome. Academic Press, pp. 69–102.
lizadeh, A.A., et al., 2000. Distinct types of diffuse large B-cell lymphoma identifiedby gene expression profiling. Nature 403, 503–511.
lt, W., Dembo, M., 1999. Cytoplasm dynamics and cell motion: two-phase flowmodels. Mathematical Biosciences 156, 207–228.
lt, W., Deutsch, A., Dunn, G., 1997. Dynamics of Cell and Tissue Motion. Birkhäuser.ndrianantoandr, E., Basu, S., Karig, D.K., Weiss, R., 2006. Synthetic biology: new
engineering rules for an emerging discipline. Molecular Systems Biology 2, 1–14.sh, C., 1998. Hooke’s microscope. Trends in Microbiology 6, 391.anaji, M., 2006. A generic model of electron transport in mitochondria. Journal of
Theoretical Biology 243, 501–516.ao, G., Suresh, S., 2003. Cell and molecular mechanics of biological materials. Nature
Materials 2, 715–725.
F
F
gy and Chemistry 32 (2008) 315–331 329
arr, F.A., 2002. The Golgi apparatus: going round in circles? Trends in Cell Biology12, 101–104.
arrière, F., Kavanagh, P., Leech, D., 2006. A laccase–glucose oxidase biofuelcell prototype operating in a physiological buffer. Electrochimica Acta 51,5187–5192.
ecker, W.M., Kleinsmith, L.J., Hardin, J., 2006. The World of the Cell, 6th ed. Pear-son/Benjamin Cummings, San Francisco.
erg, J.M., Tymoczko, J.L., Stryer, L., 2007. Biochemistry, sixth ed. W. H. Freeman, NewYork.
erger, E.G., Roth, J., 1997. The Golgi Apparatus. Birkhäuser Verlag, Basel.erk, V., Zhang, W., Pai, R.D., Cate, J.H.D., 2006. Structural basis for mRNA and tRNA
positioning on the ribosome. PNAS 103, 15830–15834.ertrama, R., Pedersenb, M.G., Lucianic, D.S., Sherman, A., 2006. A simplified
model for mitochondrial ATP production. Journal of Theoretical Biology 243,575–586.
loomfield, V.A., Crothers, D.M., Tinoco, I., 2000. Nucleic Acids: Structures, Proper-ties, and Functions. University Science Books, Sausalito.
oisvert, F.M., Koningsbruggen, S.V., Navascués, J., Lamond, A.I., 2007. The multifunc-tional nucleolus. Nature Reviews Molecular Cell Biology 8, 574–585.
olouri, H., Davidson, E.H., 2002. Modeling transcriptional regulatory networks.BioEssays 24, 1118–1129.
orukhov, S., Nudler, E., 2008. RNA polymerase: the vehicle of transcription. Trendsin Microbiology 16, 126–134.
randon, M., Baldi, P., Wallace, D.C., 2006. Mitochondrial mutations in cancer. Onco-gene 25, 4647–4662.
rent, R., 2004. A partnership between biology and engineering. Nature Biotechnol-ogy 22, 1211–1214.
uerk, D.G., 1993. Biosensors: Theory and Applications. Technomic Publishing Com-pany, Lancaster.
ushati, N., Cohen, S.M., 2007. microRNA Functions. The Annual Review of Cell andDevelopmental 23, 175–205.
ao, H., Thompson, H.M., Krueger, E.W., McNiven, M.A., 2000. Disruption of Golgistructure and function in mammalian cells expressing a mutant dynamin. Jour-nal of Cell Science 113, 1993–2002.
arracedo, A., Gironella, M., Lorente, M., Garcia, S., Guzmán, M., Velasco, G., Juan,L., Iovanna, 2006. Cannabinoids induce apoptosis of pancreatic tumor cells viaendoplasmic reticulum stress-related genes. Cancer Research 66, 6748–6755.
hampe, P.C., Harvey, R.A., Ferrier, D.R., 2005. Biochemistry, third ed. LippincottWilliams & Wilkins, Philadelphia.
lark, D.P., 2005. Molecular Biology, first ed. Elsevier Academic Press.laude, A., 1946. Fractionation of mammalian liver cells by differential centrifu-
gation: experimental procedures and results. The Journal of ExperimentalMedicine 84, 61–89.
osson, P., et al., 2005. Dynamic transport of SNARE proteins in the Golgi apparatus.PNAS 102, 14647–14652.
remer, T., Cremer, C.G., 2005. Chromatin in the Cell Nucleus: Higher-order Organi-zation, Encyclopaedia of Life Sciences. John Wiley & Sons.
remer, T., Küpper, K., Dietzel, S., Fakan, Stanislav, 2004. Higher order chromatinarchitecture in the cell nucleus: on the way from structure to function. Biologyof the Cell 96, 555–567.
roft, W.J., 2006. Under the Microscope: A Brief History of Microscopy. World Sci-entific, Singapore.
ahm, R., 2008. Discovering DNA: Friedrich Miescher and the early years of nucleicacid research. Human Genetics 122, 565–581.
al Canto, M.C., 1996. The Golgi apparatus and the pathogenesis of Alzheimer’sdisease. American Journal of Pathology 148, 355–360.
opico, A.M., Tigyi, G.J., 2007. A Glance at the Structural and Functional Diversity ofMembrane Lipids, Methods in Molecular Biology, Methods in Membrane Lipids.Humana Press Inc, Totowa, pp. 1–13.
oxsey, S., McCollum, D., Theurkauf, W., 2005. Centrosomes in Cellular Regulation.Annual Review of Cell and Developmental Biology 21, 411–434.
u, Q., Liu, C., Ryham, R., Wang, X., 2005. Modeling the spontaneous curvature effectsin static cell membrane deformations by a phase field formulation. Communi-cations on Pure and Applied Analysis 4, 537–548.
didin, M., 2003. Lipids on the frontier: a century of cell-membrane bilayers. NatureReviews Molecular Cell Biology 4, 414–418.
steller, M., 2007. Cancer epigenomics: DNA methylomes and histone-modificationmaps. Nature Reviews Genetics 8, 287–298.
zziane, Z., 2006. DNA computing: applications and challenges. Nanotechnology 17,R27–R39.
arquhar, M.G., Palade, G.E., 1981. The Golgi apparatus (Complex)—(1954–1981) fromartifact to center stage. The Journal of Cell Biology 91, 77s–103s.
ehrenbacher, N., Jättelä, M., 2005. Lysosomes as targets for cancer therapy. CancerResearch 65, 2993–2995.
ielding, C.J., 2007. Lipid Rafts and Caveolae: From Membrane Biophysics to CellBiology. Wiley-VCH, Weinheim.
isher, J., Henzinger, T.A., 2007. Executable cell biology. Nature Biotechnology 25,1239–1249.
riedland, P., Kedes, L.H., 1985. Discovering the secrets of DNA. Communications of
the ACM 28, 1164–1186.urukawa, Y., et al., 2006. Design principle and prototyping of a direct pho-tosynthetic/metabolic biofuel cell (DPMFC). Journal of Micromechanics andMicroengineering 16, S220–S225.
uterman, A.H., van Meer, G., 2004. The cell biology of lysosomal storage disorders.Nature Reviews Cell Molecular Biology 5, 554–565.

3 l Biolo
G
GG
G
G
G
G
G
G
G
G
HH
H
HH
H
H
H
H
I
I
J
J
J
K
K
K
K
K
K
KK
KK
K
K
L
L
LL
L
L
L
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
N
P
P
P
P
P
P
PPP
Q
R
R
R
R
R
R
30 K. Khoshmanesh et al. / Computationa
ao, F., Yan, Y., Su, L., Wang, L., Mao, L., 2007. An enzymatic glucose/O2 biofuelcell: Preparation, characterization and performance in serum. ElectrochemistryCommunications 9, 989–996.
arrett, R.H., Grisham, C.M., 2007. Biochemistry, third ed. Thomson Brooks/Cole.ermain, D., Frank, D.A., 2007. Targeting the cytoplasmic and nuclear functions of
signal transducers and activators of transcription 3 for cancer therapy. ClinicalCancer Research 13, 5665–5669.
ladilin, E., Micoulet, A., Hosseini, B., Rohr, K., Spatz, J., Eils, R., 2007. 3D finite elementanalysis of uniaxial cell stretching: from image to insight. Physical Biology 4,104–113.
lunde, K., et al., 2003. Extracellular acidification alters lysosomal trafficking inhuman breast cancer cells. Neoplasia 5, 533–545.
orlov, A.M., 1995. The helical turbine: a new idea for low-head hydropower. HydroReview 14, 44–50.
orlov, A.M., 1998. Helical turbines for the Gulf Stream. Marine Technology 35,175–182.
ray, E., Orton, C.G., 2000. Medical physics: some recollections in diagnostic X-rayimaging and therapeutic radiology. Radiology 217, 619–625.
uarente, L., 2007. Link between aging and the nucleolus, genes & development.Journal of Cellular Biochemistry 100, 2449–2455.
ulbins, E., Dreschers, S., Bock, J., 2003. Role of mitochondria in apoptosis. Experi-mental Physiology 88, 85–90.
uler, M.O., Hsu, L., Soukasene, S., Harrington, D.A., Hulvat, J.F., Stupp, S.I., 2006. Pre-sentation of RGDS epitopes on self-assembled nanofibers of branched peptideamphiphiles. Biomacromolecules 7, 1855–1863.
anahan, D., Weinberg, R.A., 2000. The hallmarks of cancer. Cell 100, 57–70.artgerink, J.D., Beniash, E., Stupp, S.I., 2002. Peptide-amphiphile nanofibers: a
versatile scaffold for the preparation of self-assembling materials. PNAS 99,5133–5138.
artl, D.L., Jones, E.W., 2005. Genetics: Analysis of Genes and Genomes, sixth ed.Jones & Bartlett.
eimburg, T., 2007. Thermal Biophysics of Membranes. Wiley-VCH, Weinheim.ermo, L., Smith, C.E., 1998. The structure of the Golgi apparatus: a sperm’s eye
view in principal epithelial cells of the rat epididymis. Histochemistry and CellBiology 109, 431–447.
ollien, J., Weissman, J.S., 2006. Decay of endoplasmic reticulum-localized mRNAsduring the unfolded protein response. Science 313, 104–107.
olt, O.J., Gallo, F., Griffiths, G.M., 2006. Regulating secretory lysosomes. Journal ofBiochemistry 140, 7–12.
u, S., Chen, J., Butler, J.P., Wang, N., 2005. Prestress mediates force propagationinto the nucleus. Biochemical and Biophysical Research Communications 329,423–428.
u, Z., Zeng, L., Huang, Z., Zhang, J., Li, T., 2007. The study of Golgi apparatus inAlzheimer’s disease. Neurochemistry Research 32, 1265–1277.
fergan, I., Jansen, G., Assaraf, Y.G., 2005. Cytoplasmic confinement of breastcancer resistance protein (BCRP/ABCG2) as a novel mechanism of adapta-tion to short-term folate deprivation. Molecular Pharmacology 67, 1349–1359.
nternational Human Genome Sequencing Consortium, 2001. Initial sequencing andanalysis of the human genome. Nature 409, 861–921.
iang, S., Rhee, S.W., Gleeson, P.A., Storrie, B., 2006. Capacity of the Golgi apparatusfor cargo transport prior to complete assembly. Molecular Biology of the Cell 17,4105–4117.
ordan, B., 2001. DNA Microarrays: Gene Expression Applications. Springer-Verlag,Heidelberg.
oseph, S., 2003. After the ribosome structure: how does translocation work? RNA9, 160–164.
arp, G., 2007. Cell and Molecular Biology: Concepts and Experiments, fifth ed. WileyDesktop Edition E-Book.
au, T.R., Way, J.C., Silver, P.A., 2004. Nuclear transport and cancer: from mechanismto intervention. Nature Reviews Cancer 4, 106–117.
ent, O.A., Mendell, J.T., 2006. A small piece in the cancer puzzle: microRNAs astumor suppressors and oncogenes. Oncogene 25, 6188–6196.
leizen, B., Braakman, I., 2004. Protein folding and quality control in the endoplasmicreticulum. Current Opinion in Cell Biology 16, 343–349.
öhler, A., Hurt, E., 2007. Exporting RNA from the nucleus to the cytoplasm. NatureReviews Molecular Cell Biology 8, 761–773.
ool, E.T., Morales, J.C., Guckian, K.M., 2000. Mimicking the structure and func-tion of DNA: insights into DNA stability and replication. Angewandte ChemieInternational Edition 39, 990–1009.
orf, B.R., 2006. Human genetics and genomics, third ed. Wiley-Blackwell.osik, K.S., 2006. The neuronal microRNA system. Nature Reviews Neuroscience 7,
911–920.roemer, G., 2006. Mitochondria in cancer. Oncogene 25, 4630–4632.roemer, G., Jäättelä, M., Lysosomes, 2005. Autophagy in cell death control. Nature
Reviews Cancer 5, 886–897.ubitscheck, U., Wedekind, P., Zeidler, O., Grote, M., Peters, R., 1996. Single nuclear
pores visualized by confocal microscopy and image processing. Biophysical Jour-nal 70, 2067–2077.
usumi, A., et al., 2005. Paradigm shift of the plasma membrane concept from thetwo-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annual Review of Biophysics andBiomolecular Structure 34, 351–378.
afontaine, D.L.J., Tollervey, D., 2001. The function and synthesis of ribosomes. Molec-ular Cell Biology 2, 515–520.
R
S
gy and Chemistry 32 (2008) 315–331
ee, R.C., Ambros, V., 2001. An extensive class of small RNAs in Caenorhabditis ele-gans. Science 294, 862–864.
iljas, A., 2004. Structural Aspects of Protein Synthesis. World Scientific, Singapore.iotta, L., Petricoin, E., 2000. Molecular profiling of human cancer. Nature Reviews
Genetics 1, 48–56.odish, H., Berk, A., Zipursky, S.L., Matsudaira, P., Baltimore, D., Darnell, J., 2008.
Molecular Cell Biology, sixth ed. W. H. Freeman, New York.oebl, H.C., Matthai, C.C., 2004. Simulation studies of protein translocation in Mito-
chondria. Physica A 342, 612–622.uzio, J.P., Pryor, P.R., Bright, N.A., 2007. Lysosomes: fusion and function. Nature
Reviews Molecular Cell Biology 8, 622–632.aclean, K.H., Dorsey, F.C., Cleveland, J.L., Kastan, M.B., 2008. Targeting lysosomal
degradation induces p53-dependent cell death and prevents cancer in mousemodels of lymphomagenesis. Journal of Clinical Investigation 118, 79–88.
arciniak, S.J., Ron, D., 2006. Endoplasmic reticulum stress signaling in disease.Physiological Reviews 86, 1133–1149.
arguet, D., Lenne, P.F., Rigneault, H., He, H.T., 2006. Dynamics in the plasma mem-brane: how to combine fluidity and order. The EMBO Journal 25, 3446–3457.
arko, J.F., 2005. Introduction to Single-DNA Micromechanics, Department ofPhysics, University of Illinois Reports, 2005.
arshall, W.F., 2007. What is the function of centrioles? Journal of Cellular Biochem-istry 100, 916–922.
athews, C.K., Van Holde, K.E., Ahern, K., 2000. Biochemistry, third ed. Ben-jamin/Cummings, San Francisco.
aysinger, D., 2007. Nanoparticles and cells: good companions and doomed part-nerships. Organic and Biomolecular Chemistry 5, 2335–2342.
azzarello, P., 1999. A unifying concept: the history of cell theory. Nature Cell Biology1, E13–E15.
cnamara, P.A., 2006. Trends in RNA Research, first ed. Nova Science Publishers,Hauppauge.
cNeil, P.L., Kirchhausen, T., 2005. An emergency response team for membranerepair. Nature Reviews Molecular Cell Biology 6, 499–505.
itra, K., Frank, J., 2006. Ribosome dynamics: insights from atomic structure mod-eling into cryo-electron microscopy maps. Annual Review of Biophysics andBiomolecular Structure 35, 299–317.
ogilner, A., Wollman, R., Civelekoglu-Scholey, G., Scholey, J., 2006. Modeling mito-sis. Trends in Cell Biology 16.
oore, C.M., Minteer, S.D., Martin, R.S., 2005. Microchip-based ethanol/oxygen bio-fuel cell. Lab Chip 5, 218–225.
orrison, K.L., Weiss, G.A., 2006. The origins of chemical biology. Nature ChemicalBiology 2, 3–6.
oser, C.C., Farid, T.A., Chobot, S.E., Dutton, P.L., 2006. Electron tunneling chains ofmitochondria. Biochimica et Biophysica Acta 1757, 1096–1109.
uchir, A., Worman, H.J., 2004. The nuclear envelope and human disease. Physiology19, 309–314.
elson, D.L., Cox, M.M., 2005. Lehninger Principles of Biochemistry, forth ed. W. H.Freeman, New York.
akendorf, B., Stoneking, M., 2005. Mitochondrial DNA and human evolution. AnnualReview of Genomics and Human Genetics 6, 165–183.
ark, M.K., Choi, Y.M., Kang, Y.K., Petersen, O.H., 2008. The endoplasmic reticulumas an integrator of multiple dendritic events. The Neuroscientist 14, 68–77.
aul, M.K., Mukhopadhyay, A.K., 2007. Cancer—the mitochondrial connection. Biolo-gia Bratislava 62, 371–380.
ercec, V., Bera, T.K., 2002. Cell membrane as a model for the design of semifluo-rinated ion-selective nanostructured supramolecular systems. Tetrahedron 58,4031–4040.
eterson, J.R., Smith, T.A., Thordarson, P., 2007. Photo-induced reduction of catalyt-ically and biologically active Ru(II)bisterpyridine-cytochrome c bioconjugates.Chemical Communications (Cambridge) 19, 1899–1901.
illai, R.S., 2005. MicroRNA function: multiple mechanisms for a tiny RNA? RNA 11,1753–1761.
on, L.A., Schon, E.A., 2007. Mitochondria. Elsevier/Academic Press.ratt, C.W., Cornely, K., 2004. Essential Biochemistry. J. Wiley, Hoboken.uri, S., Linstedt, A.D., 2003. Capacity of the Golgi apparatus for biogenesis from the
endoplasmic reticulum. Molecular Biology of the Cell 14, 5011–5018.umsiyeh, M.B., 1999. Structure and function of the nucleus: anatomy and physiol-
ogy of chromatin, CMLS. Cellular and Molecular Life Sciences 55, 1129–1140.inaldis, E.D., Lahm, Armin, 2007. DNA Microarrays: Current Applications. Horizon
Scientific Press, Wymondham.itchie, K., Shan, X.Y., Kondo, J., Iwasawa, K., Fujiwara, T., Kusumi, A., 2005. Detection
of non-brownian diffusion in the cell membrane in single molecule tracking.Biophysical Journal 88, 2266–2277.
odrigues-Martins, A., et al., 2007. Revisiting the role of the mother centriole incentriole biogenesis. Science 316, 1046–1050.
ohrbach, M., Clarke, J.T.R., 2007. Treatment of lysosomal storage disorders: progresswith enzyme replacement therapy. Drugs 67, 2697–2716.
on, D., 2002. Translational control in the endoplasmic reticulum stress response.Journal of Clinical Investigation 110, 1383–1388.
owat, A.C., et al., 2005. Characterization of the elastic properties of the nuclear
envelope. Journal of the Royal Society Interface 2, 63–69.uska, E., 1986. The development of the electron microscopeand of electron microscopy, Nobel Lecture, December 8,http://nobelprize.org/nobel prizes/physics/laureates/1986/ruska-lecture.html.
caffi di, P., Gordon, L., Misteli, T., 2005. The cell nucleus and aging: tantalizing cluesand hopeful promises. PLoS Biology 3, 1855–1859.

l Biolo
S
S
S
S
S
S
S
SS
S
S
S
T
T
T
T
T
TT
T
T
T
U
V
V
W
W
W
W
W
W
W
W
W
Y
Y
Y
Z
Z
Z
Z
K. Khoshmanesh et al. / Computationa
cheffler, I.E., 2000. A century of mitochondrial research: achievements and per-spectives. Mitochondrion 1, 3–31.
chlitt, T., Brazma, A., 2006. Modelling in molecular biology: describing transcriptionregulatory networks at different scales. Philosophical Transactions of the RoyalSociety B 361, 483–494.
chmidt-Zachmann, M.S., 2005. The nucleolus—a multifunctional nuclear body. Zell-biologie aktuell 31, 22–25.
ettembre, C., et al., 2008. A block of autophagy in lysosomal storage disorders.Human Molecular Genetics 17, 119–129.
hapiro, J.A., 2006. Genome informatics: the role of DNA in cellular computations.Biological Theory 1, 288–301.
inger, S.J., Nicolson, G.L., 1972. The fluid mosaic model of the structure of cellmembranes. Science 175, 720–731.
kjøndal-Bar, N., Morris, D.R., 2007. Dynamic model of the process of protein syn-thesis in eukaryotic cells. Bulletin of Mathematical Biology 69, 361–393.
nustad, D.P., Simmons, M.J., 2006. Principles of Genetics, fourth ed. Wiley.tearns, T., 2001. Centrosome duplication: a centriolar pas de deux. Cell 105, 417–
420.tephens, S.B., 2005. Stable ribosome binding to the endoplasmic reticulum enables
compartment-specific regulation of mRNA translation. Molecular Biology of theCell 16, 5819–5831.
tewart, M., 2007. Molecular mechanism of the nuclear protein import cycle. NatureReviews Molecular Cell Biology 8, 195–208.
tupp, S.I., et al., 1997. Supramolecular materials: self-organized nanostructures.Science 276, 384.
addei, A., 2007. Active genes at the nuclear pore complex. Current Opinion in CellBiology 19, 305–310.
aylor, R.W., Turnbull, D.M., 2005. Mitochondrial DNA mutations in human disease.Nature Reviews Genetics 6, 389–402.
eiten, M.H., Bezdetnaya, L., Morlière, P., Santus, R., Guillemin, F., 2003. Endoplasmicreticulum and Golgi apparatus are the preferential sites of Foscan localisation incultured tumour cells. British Journal of Cancer 88, 146–152.
he International HapMap Consortium, 2005. A haplotype map of the humangenome. Nature 437, 1299–1320.
hirya, M., Lafontaine, D.L.J., 2005. Birth of a nucleolus: the evolution of nucleolarcompartments. Trends in Cell Biology 15, 194–199.
obin, A.J., Morel, R.E., 1997. Asking About Cells, first ed. Saunders College Division.ogo, M., Takamura, A., Asai, T., Kaji, H., Nishizawa, M., 2007. An enzyme-based
microfluidic biofuel cell using vitamin K3-mediated glucose oxidation. Elec-trochimica Acta 52, 4669–4674.
racqui, P., Ohayon, J., 2004. Transmission of mechanical stresses within thecytoskeleton of adherent cells: a theoretical analysis based on a multi-component cell model. Acta Biotheoretica 52, 323–341.
ravers, A.A., Thompson, J.M.T., 2004. An introduction to the mechanics of DNA.Philosophical Transactions of the Royal Society of London Series A- Mathemat-ical Physical and Engineering Sciences 362, 1265–1279.
sou, M.B., Stearns, T., 2006. Controlling centrosome number: licenses and blocks.Current Opinion in Cell Biology 18, 74–78.
rbani, L., Stearns, T., 1999. The centrosome: a primer. Current Biology 9, R315–R317.
Z
Z
gy and Chemistry 32 (2008) 315–331 331
asir, J.K., Labhasetwar, V., 2007. Biodegradable nanoparticles for cytosolic deliveryof therapeutics. Advanced Drug Delivery Reviews 59, 718–728.
oet, D., Voet, J.G., Pratt, C.W., 2006. Fundamentals of Biochemistry: Life at theMolecular Level, Second ed. Wiley, New York.
alhout, A.J.M., 2006. Unraveling transcription regulatory networks byprotein–DNA and protein–protein interaction mapping. Genome Research16, 1445–1454.
alker, J.M., Rapley, R., 2000. Molecular Biology and Biotechnology, fourth ed. RoyalSociety of Chemistry.
atson, J.D., Crick, F.H.C., 1953. A structure for deoxyribose nucleic acid. Nature 171,737–738.
eber, P., Wagner, M., Schneckenburger, H., 2006. Microfluorometry of cell mem-brane dynamics. Cytometry Part A 69A, 185–188.
eigl, B.H., Bardell, R.L., Cabrera, C.R., 2003. Lab-on-a-chip for drug development.Advanced Drug Delivery Reviews 55, 349–377.
eitzke, E.L., Ortoleva, P.J., 2003. Simulating cellular dynamics through a coupledtranscription, translation, metabolic model. Computational Biology and Chem-istry 27, 469–480.
esterhoff, H.V., Palsson, B.O., 2004. The evolution of molecular biology into systemsbiology. Nature Biotechnology 22, 1249–1252.
u, M., Holowka, D., Craighead, H.G., Baird, B., 2004. Visualization of plasmamembrane compartmentalization with patterned lipid bilayers. PNAS 101,13798–13803.
u, F., Yang, F., Vinnakota, K.C., Beard, D.A., 2007. Computer modeling of mitochon-drial tricarboxylic acid cycle, oxidative phosphorylation, metabolite transportand electrophysiology. The Journal of Biological Chemistry 282, 24525–24537.
eo, J.W., Samarrai, W., Barjis, I., Ryu, Y.S., 2005. Data-flow diagram representationof biological process: DNA, RNA, and protein, bioengineering conference. In:Proceedings of the IEEE 31st Annual Northeast.
oneda, M., Nagai, R., 1988. Structural basis of cytoplasmic streaming in Characeaninternodal cells, a hydrodynamic analysis. Protoplasma 147, 64–76.
oneyama, M., Fujita, T., 2007. Cytoplasmic double-stranded DNA sensor. NatureImmunology 8, 907–908.
auner, K.P., 2005. Molecular Information Technology. Solid State and Material Sci-ences 30, 3–69.
hang, X., Ranta, A., Halme, A., 2006. Direct methanol biocatalytic fuelcell—Considerations of restraints on electron transfer. Biosensors and Bioelec-tronics 21, 2052–2057.
hang, L., Gu, F.X., Chan, J.M., Wang, A.Z., Langer, R.S., Farokhzad, O.C., 2007.Nanoparticles in medicine: therapeutic applications and developments. ClinicalPharmacology and Therapeutics.
hang, B., Pan, X., Cobb, G.P., Anderson, T.A., 2007. microRNAs as oncogenes andtumor suppressors. Developmental Biology 302, 1–12.
hu, C., Bao, G., Wang, N., 2000. Cell mechanics: mechanical response, cell adhe-sion, and molecular deformation. Annual Review of Biomedical Engineering 2,189–226.
ong, W.X., Li, C., Hatzivassiliou, G., Lindsten, T., Yu, Q.C., Yuan, J., Thompson, C.B.,2003. Bax and Bak can localize to the endoplasmic reticulum to initiate apopto-sis. The Journal of Cell Biology 162, 59–69.