assessing lake typologies and indicator fish species for italian natural lakes using past fish...
TRANSCRIPT

PRIMARY RESEARCH PAPER
Assessing lake typologies and indicator fish speciesfor Italian natural lakes using past fish richnessand assemblages
Pietro Volta • Alessandro Oggioni •
Roberta Bettinetti • Erik Jeppesen
Received: 6 January 2011 / Revised: 12 April 2011 / Accepted: 16 April 2011 / Published online: 29 April 2011
� Springer Science+Business Media B.V. 2011
Abstract In order to establish a fish-based typology
of Italian lakes and identify possible reference and
indicator fish species for each lake type, we analysed
historical data on fish assemblages of all Italian
natural lakes [0.5 km2 from the period prior to the
major decline in water quality in the 1950s. General
linear regression models showed the ecoregion and
lake altitude being the best predictors of fish species
richness. The number of species was significantly
higher in the Alpine than in the Mediterranean
ecoregion. Among Alpine lakes, the number of fish
species increased significantly with lake volume
whilst decreased with altitude. In the Mediterranean
lakes, none of the selected parameters was significant.
Cluster analysis of fish assemblages (presence/
absence) divided the lakes of the Alpine and Med-
iterranean ecoregions into four and two types,
respectively. Pike (Esox lucius), rudd (Scardinius
erythrophthalmus) and tench (Tinca tinca) were the
main indicator species for the small and mostly
shallow lakes in both the Alpine (Type 1) and
Mediterranean (Type 6) ecoregions, minnow (Phox-
inus phoxinus) for the alpine high altitude lakes
(Type 2) and landlocked shad (Alosa fallax lacustris),
European whitefish (Coregonus lavaretus) and burbot
(Lota lota) for the large and very deep alpine lakes
(Type 4). The European whitefish was the only
indicator species for the deep Mediterranean lakes
(Type 5). These species and associated fish assem-
blages may be useful indicators in future assessments
of the ecological status of Italian lakes according to
the European Directives (2000/60/EC and 2008/105/
EC).
Keywords Fish fauna �Biodiversity �Bioindicators �Reference conditions � Biomonitoring � Pollutants
Handling editor: P. Noges
Electronic supplementary material The online version ofthis article (doi:10.1007/s10750-011-0720-6) containssupplementary material, which is available to authorized users.
P. Volta (&) � A. Oggioni
CNR-Institute of Ecosystem Study, L.go Tonolli 50,
28922 Verbania Pallanza, Italy
e-mail: [email protected]
R. Bettinetti
Department of Chemical and Environmental Sciences,
University of Insubria, Via Valleggio 11, 22100 Como,
Italy
E. Jeppesen
National Environmental Research Institute, Aarhus
University, Vejlsøvej 25, 8600 Silkeborg, Denmark
E. Jeppesen
Greenland Climate Research Centre (GCRC), Greenland
Institute of Natural Resources, Kivioq 2, P.O. Box 570,
3900 Nuuk, Greenland
E. Jeppesen
Sino-Danish Centre for Education and Research (SDC),
Beijing, China
123
Hydrobiologia (2011) 671:227–240
DOI 10.1007/s10750-011-0720-6

Introduction
During the past 50 years, fish assemblages in lakes
and reservoirs have been widely investigated at a
regional and ecoregional scale in many North and
Central European countries. Such studies have linked
the characteristics of the fish community with phys-
ico-chemical and morphological variables of the
lakes, as well as with geographical and climatic
variables and anthropogenic pressures (e.g. Eckmann,
1995; Holmgren & Appelberg, 2000; Jeppesen et al.,
2000; Tammi et al., 2003; Penczak et al., 2004;
Diekmann et al., 2005; Mehner et al., 2005; Garcia
et al., 2006; Eros et al., 2009). In contrast, only few
studies have been conducted in countries around the
Mediterranean Sea (Godinho et al., 1998; Argillier
et al., 2002; Irz et al., 2002, 2004; Carol et al., 2006).
In Italy, most studies of freshwater fishes have
considered taxonomic issues (Bianco, 1998; Bianco
& Ketmaier, 2001; Ketmaier et al., 2003; Marchetto
et al., 2010), and most studies focusing on ecological
and biogeographical investigations have been spa-
tially scattered (Galli et al., 2007; Orban et al., 2007;
Pedicillo et al., 2008; Volta & Jepsen, 2008;
Lorenzoni et al., 2009; Volta et al., 2009; Volta &
Giussani, 2010) and not encompassed wide geo-
graphical or temporal scales. At present, Italian lakes
are only divided into three very general categories:
those suitable for salmonids, percids and cyprinids,
respectively. The environmental and fishing author-
ities do not consider lake types or classify lakes
according to type of fish assemblages. In conse-
quence, environmental assessments and conservation
measures are based on limited information of refer-
ence conditions. However, with the establishment of
the Water Framework Directive (WFD; EU, 2000),
there is an urgent need for expanding our knowledge
of biological communities (including fish) in Italian
lakes. The WFD requires assessment of the ecolog-
ical status of lakes using four biological elements
(phytoplankton, macrophytes, macrobenthos and fish)
supported by physical–chemical and hydromorpho-
logical information. The present status of each
biological quality element (BQE) has to be compared
with the community in a ‘near-natural state’, i.e. a
situation of minimum or no anthropogenic distur-
bance, the so-called ‘reference state’. To determine
the reference state, the WFD proposes a comparison
of the statuses of the BQEs of a particular waterbody
with those of another waterbody of the same type, but
in a ‘near-natural’ (least disturbed) state. If this is not
feasible, use of historical data, modelling or expert
judgement is required.
Although the use of historical data involves a
degree of uncertainty due to the different quality and
quantity of data (often only qualitative data are
available), it may provide important information on
the composition and structure of the fish assemblages
in a near-natural state (Steedman et al., 1996).
Therefore, identifying indicator fish species for
different lake types using historical data and thereby
defining the minimum required fish assemblages for
each lake type in a near-natural state might be an
effective approach to overcome the present lack of
reference sites. Furthermore, the Directive 2008/105/
EC (EU, 2008), amending the WFD 2000/60/EC,
includes fish for the monitoring of the levels of
pollutants that are hardly measurable in water
(Galassi & Cassi, 2001; Belpaire & Goemans,
2007), such as mercury, hexachlorobenzene and
hexachlorobutadiene. However, identification of ref-
erence species that can realistically be used as
bioindicators in freshwaters is still subject to debate
(Bettinetti et al., 2010). Therefore, the elaboration of
a list of lake type specific fish species present prior to
pollution is also important for the risk assessment of
pollutants.
The present study describes the fish assemblages in
all Italian natural lakes with a surface area [0.5 km2
prior to major anthropogenic disturbance. The year
1950 was a significant time threshold between low
and high level of anthropogenic pressures on lakes, as
it corresponds to the period between low and high
human development in Italy (before and after the
Second World War). This is particularly true for
eutrophication which, although it began to increase
already in the 18th century, did not seriously affect
the Italian lakes until the 1950s (Guilizzoni et al.,
1982; Margaritora, 1992; Marchetto et al., 2004;
Salmaso et al., 2007). We therefore explore the fish
assemblages prior to 1950 and relate our findings to
limnological characteristics and the geographical
location of the lakes in order to establish a fish-based
typology of Italian lakes and to identify possible
reference and indicator fish species for each lake
type.
228 Hydrobiologia (2011) 671:227–240
123

Materials and methods
Study area
This study considers all natural lakes[0.5 km2 located
in the Italian peninsula. The 46 lakes covered a
latitudinal and longitudinal range between 46�450N–
37�300N and 7�220E–14�180E, respectively. From a
zoogeographical point of view, Italy is divided into
two different ichthyogeographical districts (Bianco,
1995; Abell et al., 2008): the Padano-Venetian district
(from the Alps to the eastern edge of the Apennine
mountains facing the Adriatic Sea) and the Tuscano-
Latium district (facing the Tyrrhenean Sea and the
insular regions), being identical with the two separated
ecoregions identified by the WFD guidelines: the
Alpine and the Mediterranean. Hence, we a priori
separated the lakes as belonging to either the Alpine or
the Mediterranean ecoregion, corresponding more or
less to north and south of the River Po.
Data collection and elaboration
To determine the reference state, we analysed the
available literature data extracted from the historical
library of the Italian Institute of Hydrobiology
(presently CNR-Institute of Ecosystem Study) which
holds documents covering a period from 1700 to
1950 (see Appendix I—Electronic supplementary
materials). Most of the documents are from the period
1850–1930 and include scientific papers and reports
from universities and research institutions and notes
in technical reports from the Ministry of Agriculture
and fisheries authorities on regional economies and
commercial trades. The data achieved from this
historical analysis were checked against those from
recent publications (Tortonese, 1971; Tortonese,
1975; Gandolfi et al., 1991) functioning as the official
(Ministry of the Environment) reference publications
on the Italian fish fauna: scientific names of the fish
species in this study are those included in Gandolfi
et al. (1991).
Thirty-five fish species were included in the
database, belonging to 9 orders and 14 families
(Table 1). Barbel Barbus plebejus, lamprey Lampetra
planeri, grayling Thymallus thymallus, sturgeon
Acipenser spp. and eel were excluded from the
analyses since they are not usually present in lakes or
typical sea migratory species without a predictable
life span in freshwaters. The fish assemblages
included in the database consisted of a mixture
between native and a few non-native species, the
latter being introduced or translocated at different
times from Eastern Europe, Austria, Switzerland and
USA to the Alpine ecoregion and later from north to
south (Gandolfi et al., 1991). This holds true for 7
species: carp, char, European whitefish, ‘bondella’
whitefish, black bullhead, largemouth bass and
pumpkinseed. Also, perch was translocated from
north to the south during the first decades of the
1900s, and big-scale smelt was introduced to lakes in
central Italy from coastal waters.
Data on available morphometric, geographical and
limnological characteristics of lakes including lake
area, maximum depth, volume, altitude and catch-
ment area, alkalinity and retention time were
obtained from LIMNO, the national database of
Italian lakes (Tartari et al., 2004). Data were log-
transformed before statistical analyses and checked
for multicollinearity. Hence, lake area and maximum
depth were excluded since they were highly corre-
lated (r [ 0.80, P \ 0.05) with lake volume both in
the alpine and in the Mediterranean lakes.
General linear regression models (STATISTICA
software) were used to assess the relationship
between the number of species (dependent variable),
the lake characteristics (independent variable) and the
ecoregion (categorical variable) within the whole
dataset. Multiple linear regression was used to assess
the same relationships within each single ecoregion.
Differences between the number of fish species
between ecoregions were tested by Student t test.
Additionally, we calculated the average ratio between
the number of indicator species and the total number
of fish species in each lake (called later as dispersal
ratio) and we used it as a measure of the dispersal
effectiveness. The higher is the ratio, the higher is the
number of species shared within a lake type, indicat-
ing higher dispersal effectiveness. The relationship of
dispersal ratio with the other lake parameters was
tested by general linear regression models, using
ecoregion as categorical variable and other parame-
ters as independent continuous variables.
Lake typology
We divided the lakes into types with similar fish
assemblages, based on the fish data collected, and for
Hydrobiologia (2011) 671:227–240 229
123
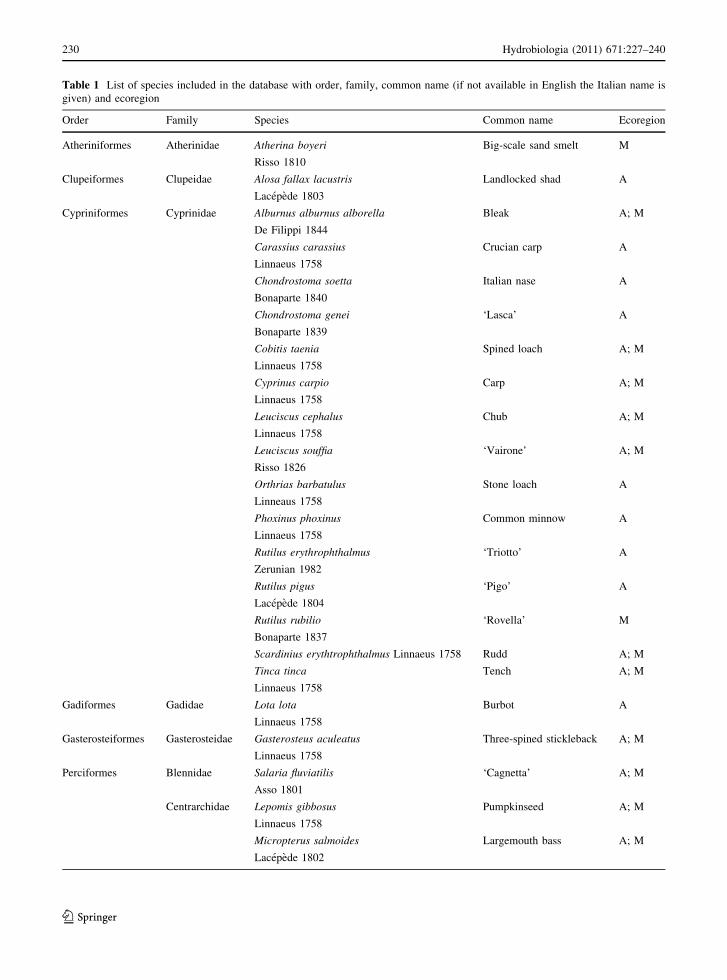
Table 1 List of species included in the database with order, family, common name (if not available in English the Italian name is
given) and ecoregion
Order Family Species Common name Ecoregion
Atheriniformes Atherinidae Atherina boyeri Big-scale sand smelt M
Risso 1810
Clupeiformes Clupeidae Alosa fallax lacustris Landlocked shad A
Lacepede 1803
Cypriniformes Cyprinidae Alburnus alburnus alborella Bleak A; M
De Filippi 1844
Carassius carassius Crucian carp A
Linnaeus 1758
Chondrostoma soetta Italian nase A
Bonaparte 1840
Chondrostoma genei ‘Lasca’ A
Bonaparte 1839
Cobitis taenia Spined loach A; M
Linnaeus 1758
Cyprinus carpio Carp A; M
Linnaeus 1758
Leuciscus cephalus Chub A; M
Linnaeus 1758
Leuciscus souffia ‘Vairone’ A; M
Risso 1826
Orthrias barbatulus Stone loach A
Linneaus 1758
Phoxinus phoxinus Common minnow A
Linnaeus 1758
Rutilus erythrophthalmus ‘Triotto’ A
Zerunian 1982
Rutilus pigus ‘Pigo’ A
Lacepede 1804
Rutilus rubilio ‘Rovella’ M
Bonaparte 1837
Scardinius erythtrophthalmus Linnaeus 1758 Rudd A; M
Tinca tinca Tench A; M
Linnaeus 1758
Gadiformes Gadidae Lota lota Burbot A
Linnaeus 1758
Gasterosteiformes Gasterosteidae Gasterosteus aculeatus Three-spined stickleback A; M
Linnaeus 1758
Perciformes Blennidae Salaria fluviatilis ‘Cagnetta’ A; M
Asso 1801
Centrarchidae Lepomis gibbosus Pumpkinseed A; M
Linnaeus 1758
Micropterus salmoides Largemouth bass A; M
Lacepede 1802
230 Hydrobiologia (2011) 671:227–240
123

each lake type we identified indicator fish species for
the reference state. The lake types were identified by
means of cluster analysis using a dissimilarity matrix
calculated from the fish communities occurring in the
lakes prior to 1950. We used the Jaccard index as it is
considered a useful index for identifying ecological
gradients (Faith et al., 1987) and a presence/absence
approach (Heino et al., 2010). For identifying lake
types, we first conducted an assessment of the best
linkage method using the ‘cophenetic correlation
coefficient’ function in R software (VEGAN pack-
age). The cophenetic correlation for a cluster tree is
defined as the linear correlation coefficient between
the distances obtained from the tree and the original
distances (or dissimilarities) used to construct the
tree. Thus, it is a measure of how well the tree
represents the dissimilarities among observations and
it has a statistical meaning indicating the statistical
differences between the different clusters and there-
fore the robustness of the results (Becker et al., 1988).
We chose the Average Linkage Method as it reached
the highest cophenetic value (0.97). The numerical
analyses were carried out by means of R Software
version 2.11.0 (R Development Core Team, 2010)
and Vegan Community Ecology Package version
1.17-2-8 (Oksanen et al., 2010).
Table 1 continued
Order Family Species Common name Ecoregion
Percidae Perca fluviatilis Perch A, M
Linnaeus 1758
Stizostedion lucioperca Pikeperch A
Linnaeus 1758
Gobidae Orsinogobius punctatissimus ‘Panzarolo’ A
Canestrini 1864
Padogobius martensi Padanian goby A
Gunther 1861
Salmoniformes Coregonidae Coregonus lavaretus European whitefish A; M
Linnaeus 1758
Coregonus macrophthalmus ‘Bondella’ A
Nusslin 1882
Esocidae Esox lucius Pike A; M
Linnaeus 1758
Salmonidae Salmo carpio ‘Carpione del Garda’ A
Linnaeus 1758
Salmo trutta lacustris Trout A
Linnaeus 1758
Salvelinus alpinus Char A
Linnaeus 1758
Salmo trutta macrostigma Mediterranean trout M
Dumeril 1858
Salmo trutta marmoratus Marble trout A
Linnaeus 1758
Salmo trutta trutta Trout A
Linnaeus 1758
Scorpaeniformes Cottidae Cottus gobio Bullhead A
Linnaeus 1756
Siluriformes Ictaluridae Ictalurus melas Black bullhead A
Rafinesque 1820
Hydrobiologia (2011) 671:227–240 231
123

Differences in morphometric and chemical charac-
teristics of lake types were tested by Mann–Whitney
statistical test (SYSTAT Software). Significant levels
were set at P \ 0.05.
Indicator fish species identification
Indicator fish species, i.e. guiding and accompanying
fish species, were determined according to Gassner
et al. (2005) and Volta and Oggioni (2010). Guiding
fish species were those that discriminate well between
lake types, i.e. ideally are present in one lake type and
absent in the others. Guiding species were defined by
an occurrence of 100% in one lake type and by a
value of factor a B 7, where a is:
a ¼ ðRpiÞ � range,
‘pi’ is the occurrence (between 0 = absent and
1 = ubiquitous) of a fish species in one lake type
and ‘range’ is the number of lake types where the
species is present. Factor ‘a’ enables selection of
species indicative of one lake type, i.e. species which
are present in all lakes of a particular type, but rare in
the other types.
Accompanying fish species were defined by an
occurrence C80% in one lake type.
Results
General linear regression models (R2 = 0.694,
P \ 0.001) show ecoregion and lake altitude being
the best predictor of fish species richness in Italian
lakes (P = 0.028 and P = 0.027, respectively). The
dispersal ratio (R2 = 0.479, P \ 0.001) was explained
significantly only by the altitude (P = 0.017). The
number of species was significantly higher (t test,
P \ 0.001) in the lakes in the Alpine ecoregion
(n = 34) than in the Mediterranean ecoregion
(n = 13), the average being 12.5 (±3.1) and 7.6
(±2.8) species, respectively. Limited to Alpine lakes
(R2 = 0.724, P \ 0.001), the number of fish species
increases with lake volume (b = 0.569), whilst
decrease significantly with altitude (b = -0.22). In
the Mediterranean lakes, instead none of the selected
parameters contributed to explain significantly the fish
species richness (R2 = 0.487, P = 0.314).
Cluster analysis of the presence/absence of fish
revealed different lake types (Fig. 1a, b). By
maximizing the homogeneity within clusters and the
heterogeneity between clusters, four lake types in the
Alpine ecoregion and two lake types in the Mediter-
ranean were identified. Main lake type characteristics
are shown in Table 2. The lake types were charac-
terized by significant differences in morphometric
and physico-chemical variables as well as in fish
species richness (Table 3).
Sixteen relatively shallow (20.0 ± 15.2 m) and
small (2.9 ± 3.4 km2) lakes were included in Type 1
of the Alpine ecoregion. They are located mostly in
the lowlands (altitude 245.0 ± 74.4 m a.s.l.), just
beyond the moraine edge of the old glaciers, follow-
ing a wide longitude range. Phytophilic species such
as pike, tench and rudd were the guiding species
accompanied by bleak, perch, carp and Padanian
goby (Table 4; Fig. 2).
The second group comprised three high altitude
(1198 ± 242.6 m a.s.l.) lakes located in North-East
Italy. They had low species richness (3.7 ± 1.2) and
were characterized by cold stenothermal and sensitive
species such as the small cyprinid, common minnow,
which is the guiding species, bullhead and salmonids.
Where streams were absent, char was the only
salmonid present, while trout, requiring streams for
reproduction, was only present in lakes associated with
streams.
Type 3 comprised the deep (75.9 ± 50.4 m) but
small (3.5 ± 3.8 km2) lakes located in the north-east.
They constitute an ‘intermediate environment’
between the very deep (Type 4) and shallow (Type
1) lakes. Compared to Type 1, these lakes differed
significantly in depth, but not in area. Therefore,
phytophilic species such as pike, rudd and tench had
the highest indicator value, although trout was still
included in the accompanying species group. The
Italian nase was also accompanying species in these
lakes together with perch and carp.
Type 4 comprised seven large (115.6 ±
136.6 km2) and very deep (238.0 ± 144.1 m) subal-
pine lakes in north-west Italy. The number of species
present was generally high (14.0 ± 4.8) due to the
availability of several different habitats. A well
oxygenated and cold hypolimnion is a suitable habitat
for the sensitive cold stenothermal burbot and Euro-
pean whitefish, which were guiding species, and for
trout. Gregarious euritherm zooplanktivorous species
such as the landlocked shad were also guiding species
in this type of lakes. The number of accompanying
232 Hydrobiologia (2011) 671:227–240
123
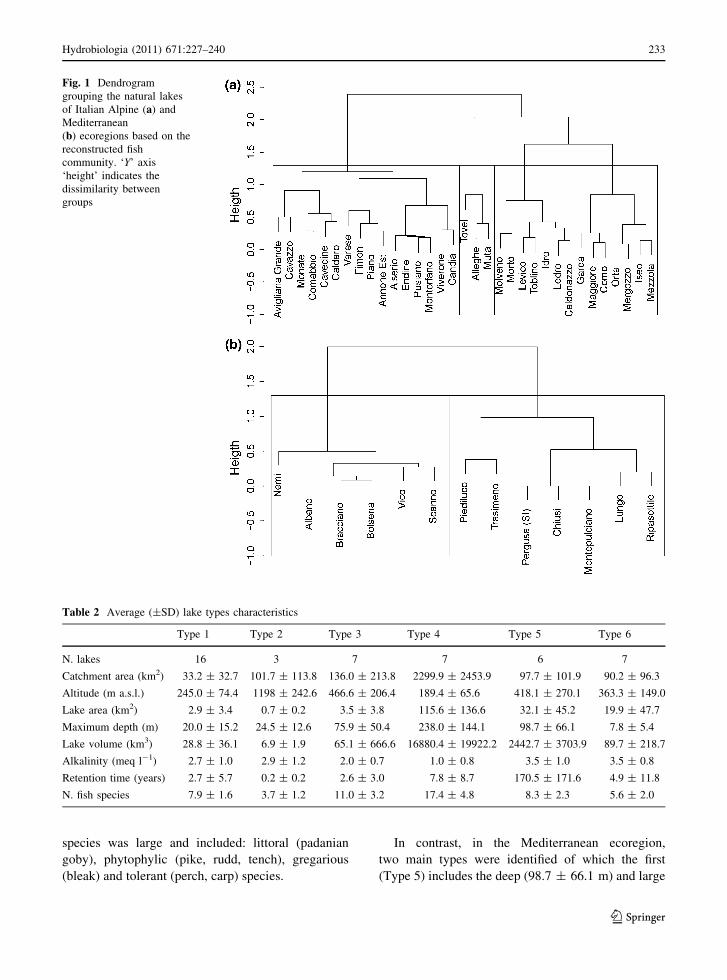
species was large and included: littoral (padanian
goby), phytophylic (pike, rudd, tench), gregarious
(bleak) and tolerant (perch, carp) species.
In contrast, in the Mediterranean ecoregion,
two main types were identified of which the first
(Type 5) includes the deep (98.7 ± 66.1 m) and large
Fig. 1 Dendrogram
grouping the natural lakes
of Italian Alpine (a) and
Mediterranean
(b) ecoregions based on the
reconstructed fish
community. ‘Y’ axis
‘height’ indicates the
dissimilarity between
groups
Table 2 Average (±SD) lake types characteristics
Type 1 Type 2 Type 3 Type 4 Type 5 Type 6
N. lakes 16 3 7 7 6 7
Catchment area (km2) 33.2 ± 32.7 101.7 ± 113.8 136.0 ± 213.8 2299.9 ± 2453.9 97.7 ± 101.9 90.2 ± 96.3
Altitude (m a.s.l.) 245.0 ± 74.4 1198 ± 242.6 466.6 ± 206.4 189.4 ± 65.6 418.1 ± 270.1 363.3 ± 149.0
Lake area (km2) 2.9 ± 3.4 0.7 ± 0.2 3.5 ± 3.8 115.6 ± 136.6 32.1 ± 45.2 19.9 ± 47.7
Maximum depth (m) 20.0 ± 15.2 24.5 ± 12.6 75.9 ± 50.4 238.0 ± 144.1 98.7 ± 66.1 7.8 ± 5.4
Lake volume (km3) 28.8 ± 36.1 6.9 ± 1.9 65.1 ± 666.6 16880.4 ± 19922.2 2442.7 ± 3703.9 89.7 ± 218.7
Alkalinity (meq l-1) 2.7 ± 1.0 2.9 ± 1.2 2.0 ± 0.7 1.0 ± 0.8 3.5 ± 1.0 3.5 ± 0.8
Retention time (years) 2.7 ± 5.7 0.2 ± 0.2 2.6 ± 3.0 7.8 ± 8.7 170.5 ± 171.6 4.9 ± 11.8
N. fish species 7.9 ± 1.6 3.7 ± 1.2 11.0 ± 3.2 17.4 ± 4.8 8.3 ± 2.3 5.6 ± 2.0
Hydrobiologia (2011) 671:227–240 233
123

Ta
ble
3L
ake
var
iab
les
dif
feri
ng
sig
nifi
can
tly
bet
wee
nth
ela
ke
typ
eso
fea
chec
ore
gio
n(M
ann
–W
hit
ney
stat
isti
cal
test
:P
lev
els
are
rep
ort
ed).
Fis
has
sem
bla
ge
com
par
iso
ns
amo
ng
lak
ety
pes
are
bas
edo
nav
erag
e(±
SD
)d
issi
mil
arit
ysh
ow
nin
bo
ld.
Th
eav
erag
e(±
SD
)d
isp
ersa
lra
tio
(%)
isre
po
rted
init
alic
s
Ty
pe
1T
yp
e2
Ty
pe
3T
yp
e4
Ty
pe
5T
yp
e6
Ty
pe
18
8.4 (±
24
.3)
Lak
ear
ea(P
=0
.04
4).
Alt
itu
de
(P=
0.0
08
),
N.
fish
spec
ies
(P=
0.0
09
)
Cat
chm
ent
area
(P=
0.0
42
),A
ltit
ud
e
(P=
0.0
02
),M
ax
dep
th(P
=0
.00
4),
N.
fish
spec
ies
(P=
0.0
23
)
Cat
chm
ent
area
(P=
0.0
03
),L
ake
area
(P=
0.0
01
),A
ltit
ud
e(P
=0
.01
9),
Max
dep
th(P
\0
.00
1),
Lak
ev
olu
me
(P\
0.0
01
),A
lkal
init
y(P
=0
.00
3),
Ret
enti
on
tim
e(P
P\
0.0
01
),N
.fi
sh
spec
ies
(P\
0.0
01
)
Max
dep
th(P
=0
.00
3),
Lak
e
Vo
lum
e(P
=0
.00
6),
Ret
enti
on
tim
e(P
=0
.00
9)
Alt
itu
de
(P=
0.0
14
),
Are
a(P
=0
.00
8)
Ty
pe
28
8.1
1(–
15
.9)
66
.7(±
14
.4)
Alt
itu
de
(P=
0.0
17
),
Alk
alin
ity
(P=
0.0
17
),n
.fi
sh
spec
ies
(P=
0.0
17
)
Lak
ear
ea(P
=0
.01
7),
Alt
itu
de
(P=
0.0
17
),M
axd
epth
(P=
0.0
17
),
Lak
ev
olu
me
(P=
0.0
17
),N
.fi
sh
spec
ies
(P=
0.0
17
)
Lak
ear
ea(P
=0
.03
3),
Lak
e
Alt
itu
de
(P=
0.0
22
)
Ty
pe
35
7.0
0(–
10
.48
)7
6.9
8(–
15
.99
)6
8.5
(±2
4.1
)C
atch
men
tar
ea(P
=0
.03
8),
Lak
ear
ea
(P=
0.0
11
),A
ltit
ud
e(P
=0
.00
2),
Max
dep
th(P
=0
.00
7),
Alk
alin
ity
(P=
0.0
26
),N
.fi
shsp
ecie
s
(P=
0.0
13
)
Alk
anli
nit
y(P
=0
.00
8),
Ret
enti
on
tim
e(P
=0
.03
5)
Max
dep
th(P
=0
.00
1),
Ret
enti
on
tim
e
(P=
0.0
02
),N
.fi
sh
spec
ies
(P=
0.0
02
)
Ty
pe
45
9.1
9(–
10
.19
)8
6.6
7(–
11
.71
)5
0.8
4(–
11
.70
)7
3.9
(±1
7.8
)A
ltit
ud
e(P
=0
.03
5),
Alk
alin
ity
(P=
0.0
01
),
Ret
enti
on
tim
e(P
=0
.03
5),
N.
fish
spec
ies
(P=
0.0
01
)
Cat
chm
ent
area
(P=
0.0
38
),L
ake
area
(P=
0.0
26
),
Alt
itu
de
(P=
0.0
04
),
Max
dep
th
(P\
0.0
01
),L
ake
vo
lum
e(P
=0
.00
2),
Alk
anli
nit
y
(P\
0.0
01
),N
.fi
sh
spec
ies
(P\
0.0
01
),
Ty
pe
57
6.1
(±1
8.4
)M
axd
epth
(P=
0.0
01
),
Lak
ev
olu
me
(P=
0.0
14
),
Ret
enti
on
tim
e
(P=
0.0
08
),N
.fi
sh
spec
ies
(P=
0.0
14
)
Ty
pe
65
6.7
6(–
6.7
8)
86
.4(±
22
.5)
234 Hydrobiologia (2011) 671:227–240
123
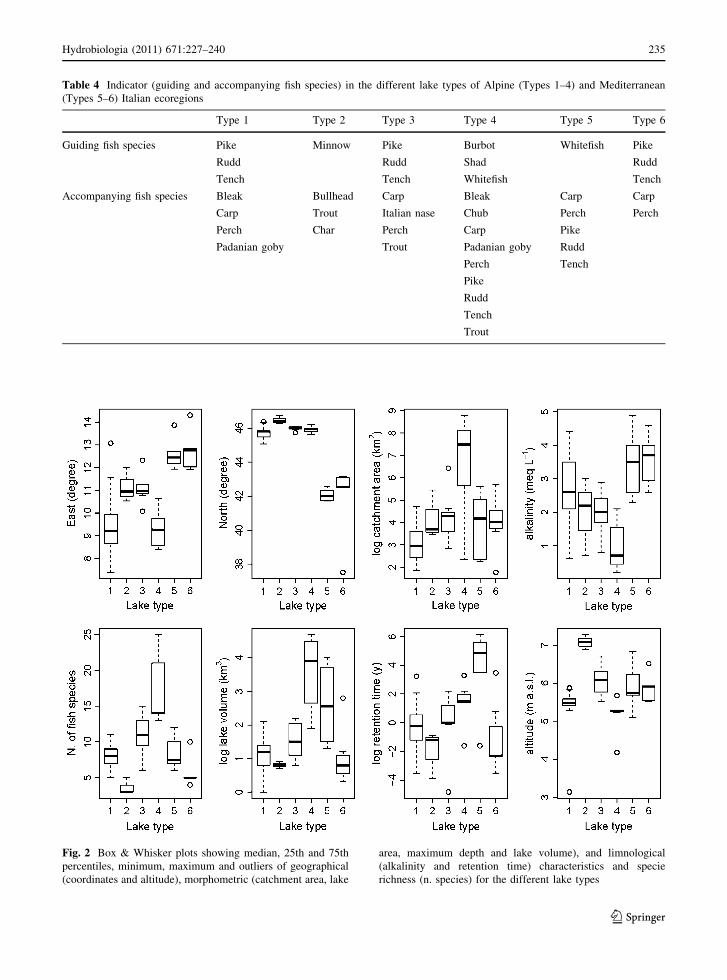
Table 4 Indicator (guiding and accompanying fish species) in the different lake types of Alpine (Types 1–4) and Mediterranean
(Types 5–6) Italian ecoregions
Type 1 Type 2 Type 3 Type 4 Type 5 Type 6
Guiding fish species Pike Minnow Pike Burbot Whitefish Pike
Rudd Rudd Shad Rudd
Tench Tench Whitefish Tench
Accompanying fish species Bleak Bullhead Carp Bleak Carp Carp
Carp Trout Italian nase Chub Perch Perch
Perch Char Perch Carp Pike
Padanian goby Trout Padanian goby Rudd
Perch Tench
Pike
Rudd
Tench
Trout
Fig. 2 Box & Whisker plots showing median, 25th and 75th
percentiles, minimum, maximum and outliers of geographical
(coordinates and altitude), morphometric (catchment area, lake
area, maximum depth and lake volume), and limnological
(alkalinity and retention time) characteristics and specie
richness (n. species) for the different lake types
Hydrobiologia (2011) 671:227–240 235
123

(32.1 ± 45.2 km2) lakes of Central Italy, mostly of
volcanic origin and located in the Latium region. The
number of species was relatively high (8.3 ± 2.3), and
European whitefish was the guiding species. The
second (Type 6) comprised all the shallow (7.8 ±
5.4 m) lakes of Central and South Italy; these were
characterized by a few typically phytophilic and highly
tolerant species (5.6 ± 2.0) such as tench, rudd
and carp.
Discussion
The implementation of the WFD requires as a first
step identification of waterbody types including sites
with reference conditions for each biological quality
element (WFD CIS, 2003). Typology of waterbodies
is usually based on physico-chemical variables and
hydromorphological characteristics. However, if ref-
erence sites at national scale are scarce or missing,
use of alternative methods to establish reference
benchmarks is required.
Fish are the only group of freshwater organisms for
which adequate historical information may be avail-
able, which makes them useful indicators of the
reference state of freshwaters (Schmutz et al., 2000).
The quality of historical data is variable and the
information on fish composition of lakes is often
biased towards commercial interesting species, non-
commercial species often being excluded from fishery
statistics. Also, historical data may include synonyms
and unclear taxonomy that are not easy to update to the
current nomenclature. However, even though use of
historical data is somewhat uncertain they may provide
valuable information on the composition and structure
of the fish assemblages in a near-natural state (Steed-
man et al., 1996). In our study, we checked for
synonyms, and all fish species were validated with the
current official Italian taxonomy (Gandolfi et al.,
1991). Moreover, our data have been gathered from
multiple sources coupling scientific observations,
paintings and photos from universities, research and
fisheries institutions, commercial and fisheries statis-
tics and local information from leasing contracts and
fish market trades. Also, due to the different sources
considered, the data cover the whole geographical
range and lake sizes enabling a satisfactory recon-
struction of the fish assemblages in the first decades of
the twentieth century in Italian lakes.
Using data for all Italian natural lakes [0.5 km2,
we obtained six lake types, four in the Alpine
ecoregion and two in the Mediterranean ecoregion
showing differences both in fish richness and com-
position. The fish assemblages were significantly
influenced by a complex set of geological, climato-
logical and limnological variables. According to
statistical analyses, the ecoregion is an important
factor explaining the fish species richness in Italian
lakes. The Alps and Apennines have acted as barriers
modulating the fish distribution by slowing down
migrations from Alpine areas to Mediterranean ones
(Bianco, 1995; Hewitt 1999; Griffiths, 2006; Reyjol
et al., 2007). For instance, the number of fish species
was lower in the deep of Type 5 situated mainly in
volcanic areas of central Italy and therefore isolated
than in those of Type 4, belonging to the river Po
basin and therefore linked to Adriatic Sea. In
addition, species with high mobility and migratory
behaviour, such as trout, chub and nase, are common
species in deep lakes in the North, while they are less
frequently occurring in the more isolated deep lakes
in the Mediterranean ecoregion.
As in other studies (see Zhao et al., 2006), we
found species richness to decrease with lake altitude.
We also found that the dispersal effectiveness
decreased with altitude, which is not surprisingly as
high altitude watersheds are less assessable for fish
migrations than those at low altitudes (e.g. Albert &
Crampton, 2010).
However, although barriers and glaciation events
likely determine species richness, species that over-
come such factors face various abiotic and biotic
constraints and opportunities modulating the fish
assemblages (Griffiths, 2006).
Also, fish species richness was highest in large and
deep lakes, while small and shallow lakes had
relatively simple fish communities. This holds true
for both high and low altitude lakes and confirms
previous findings covering a wider (Jackson &
Harvey, 1989; Tonn et al., 1990; Amarasinghe &
Welcomme, 2002; Zhao et al., 2006) and a smaller
geographical area (Eckmann, 1995; Irz et al., 2002).
Lake morphology (depth, surface and catchment
area) and altitude affects physical and chemical
characteristics such as retention time, nutrient con-
centrations and primary productivity (Wetzel, 1975).
Deep and large lakes and high altitude lakes are often
oligo-mesotrophic in the natural state, while shallow
236 Hydrobiologia (2011) 671:227–240
123

lakes are typically meso-eutrophic (Noges, 2009). We
found that high altitude or deep lakes sustain
populations of sensitive/intolerant fish species such
as salmonids (trout, char), bullhead and coregonids,
while shallow and relatively small lakes were char-
acterized mainly by phytophilic and benthivorous-
tolerant species. This is in line with numerous studies
in the northern hemisphere showing that enhanced
primary productivity reinforced by specific morpho-
metric characteristics of shallow lakes leads to a shift
in fish communities from Salmoniformes (mainly
coregonids) to phytophilic or tolerant species (Mar-
shall & Ryan, 1987; Persson et al., 1991; Holmgren
& Appelberg, 2000; Mehner et al., 2005; Garcia
et al., 2006). Furthermore, at species level, Heino
et al. (2010) found that pike and perch were among
the most common species in meso-eutrophic boreal
lakes, while brown trout was common in deep
oligotrophic lakes in the same region. Mehner et al.
(2007) found vendace Coregonus albula and burbot
to be the most indicative fish species of deep and
oligotrophic lakes in the European ‘Central Plains’
ecoregion.
We identified indicator fish species for each lake
type in the near-natural state, which may be used in the
assessment of ecological status according to the
relevant EU directives in combination with appropri-
ate metrics. Noble et al. (2007) suggested development
of metrics for young-of-the-year indicator (sentinel)
species supplemented by an examination of their
population age structure. Zick et al. (2007) and
Gassner & Wanzenbock (2007) applied size structure
indices (Gablehouse, 1984) to char and whitefish
populations in Austrian lakes, and Volta (2010) did the
same to three deep Italian lakes exhibiting differences
in trophic status and fishing pressure. Size structure
indices may be useful proxies for the determination of
age structure, a labour-demanding process that is
currently a demand of the WFD.
The indicator fish species may potentially also be
used as biomonitors for aquatic chemicals when
concentrations are at or below the detection level
(Barron et al., 1996; Volta et al., 2009) and may
provide information allowing assessment of the threats
posed by water contamination to aquatic communities
(Koponen et al., 2001; Volta et al., 2009).
The indicator species identified in our study
include pelagic and littoral feeders, benthivores and
zooplanktivores, prey and top predators, representing
different lake environments and common fish food
webs. The species are moreover easy to capture by
standardized techniques such as gillnetting and
electrofishing, enabling the development of routine
surveys and effective monitoring programs.
In conclusion, we have developed a fish-based
typology of Italian natural lakes [0.5 km2 supple-
mented with a list of indicator reference fish species for
each lake type to be used in the assessment of the
ecological and chemical quality of natural Mediterra-
nean lakes. As most of the species are of commercial
and recreational value, they are relevant as a reference
for managers of both fisheries and the environment.
The reference species and assemblages can further be
used as a basis when changes in the ichthyofauna of
lakes at local and ecoregional scale are studied and for
implementation of lake type specific management and
restoration measures, when necessary.
Acknowledgments P. Volta (PV) and E. Jeppesen (EJ) were
partially funded by the EU-FP7 WISER project; PV also by
LIFE? INHABIT and CIPAIS (Commissione Internazionale
per la Protezione delle Acque Italo-Svizzere), and EJ also by
EU-FP7 REFRESH, CRES and The Danish Council for
Independent Research: Natural Sciences (272-08-0406). We
thank Anne Mette Poulsen for most valuable editorial
comments and the two anonymous referees for the valuable
comments which improved the manuscript importantly.
References
Abell, M., M. L. Thieme, C. Revenga, M. Bryer, M. Kottelat,
N. Bugutskaya, B. Coad, N. Mandrak, S. Contreras Bal-
deras, W. Bussing, M. L. J. Stiassny, P. Skelton, G.
R. Allen, P. Unmack, A. Naseka, R. Ng, N. Sindorf, J.
Robestson, E. Armijo, J. V. Higgins, T. J. Heibel, E.
Wikramanayake, D. Olson, H. L. Lopez, R. E. Reis, J.
G. Lundberg, M. H. Sabaj Perez & P. Petry, 2008.
Freshwater ecoregions of the world: a new map of bio-
geographic units for freshwater biodiversity conservation.
BioScience 403: 403–414.
Albert, J. S. & W. G. Crampton, 2010. Geography and ecology
of diversification in Neotropical freshwaters. Nature
Education Knowledge 1: 13.
Amarasinghe, U. S. & R. L. Welcomme, 2002. An analysis of
fish species richness in natural lakes. Environmental
Biology of Fishes 65: 327–339.
Argillier, C., O. Pronier & P. Irz, 2002. A typology of fish
communities in French lakes. II. Fish assemblages in lakes
below 1500 m in altitude. Bulletin Francaise Peche
Pisciculture 365–366: 365–366.
Barron, M. G., C. Nardo & J. J. Yurk, 1996. Bioavailability of
PCBs in channel catfish exposed to a treated municipal
effluent. Environmental Technology 17: 557–560.
Hydrobiologia (2011) 671:227–240 237
123

Becker, R. A., J. M. Chambers & A. R. Wilks, 1988. The New
S Language: A Programming Environment for Data
Analysis and Graphics. Wadsworth & Brooks/Cole,
Pacific Grove, CA.
Belpaire, C. & G. Goemans, 2007. The European eel Anguillaanguilla, a rapporteur of the chemical status for the water
framework directive? Vie et Milieu – Life and Environ-
ment 57: 235–252.
Bettinetti, R., S. Galassi, S. Quadroni, P. Volta, E. Ciccotti, F.
Capoccioni & G. A. De Leo, 2010. Use of Anguillaanguilla for biomonitoring persistent organic pollutants
(POPs) in brackish and riverine waters in Central and
Southern Italy. Water Air & Soil Pollution. doi:
10.1007/s11270-010-0590-y.
Bianco, P. G., 1995. Mediterranean endemic freshwater fishes
of Italy. Biological Conservation 72: 159–170.
Bianco, P. G., 1998. Diversity of Barbinae fishes in southern
Europe with description of a new genus and a new species
(Cyprinidae). Italian Journal of Zoology 65(Suppl.):
125–136.
Bianco, P. G. & V. Ketmaier, 2001. Anthropogenic changes in
the freshwater fish fauna of Italy, with reference to the
central region and Barbus graellsii, a newly established
alien species of Iberian origin. Journal of Fish Biology 59:
190–208.
Carol, J., L. Benejam, C. Alcaraz, A. Vila-Gispert, L. Zamora,
E. Navarro, J. Armengol & E. Garcia-Berthou, 2006. The
effects of limnological features on fish assemblages of 14
Spanish reservoirs. Ecology of Freshwater Fish 15: 66–77.
Diekmann, M., U. Bramick, R. Lemcke & T. Mehner, 2005.
Habitat-specific fishing revealed distinct indicator species
in German lowland lake fish communities. Journal of
Applied Ecology 42: 901–909.
Eckmann, R., 1995. Fish richness in lakes of the northeastern
lowlands in Germany. Ecology of Freshwater Fish 4:
62–69.
Eros, T., J. Heino, D. Schmera & M. Rask, 2009. Character-
ising functional trait diversity and trait-environment
relationships in fish assemblages of boreal lakes. Fresh-
water Biology 54: 1788–1803.
EU, 2000. Directive 2000/60/EC of the European Parliament
and the Council of 23 October 2000 establishing a
framework for community action in the field of water
policy. The European Parliament and the Council of the
European Union, Brussels: 72 pp.
EU, 2008. Directive 2008/105/EC of the European Parliament
and of the Council of 16 December 2008 on environ-
mental quality standards in the field of water policy,
amending and subsequently repealing Council Directives
82/176/EEC, 83/513/EEC, 84/156/EEC, 84/491/EEC,
86/280/EEC and amending Directive 2000/60/EC of the
European Parliament and of the Council. The European
Parliament and the Council of the European Union,
Brussels: 14 pp.
Faith, D. P., P. R. Minchin & L. Belbin, 1987. Compositional
dissimilarity as a robust measure of ecological distance.
Vegetation 69: 57–68.
Gablehouse, D. W. Jr., 1984. A length categorization system to
assess fish stocks. North American Journal of Fisheries
Management 4: 273–285.
Galassi, S. & R. Cassi, 2001. Key species for monitoring
persistent and bioaccumulable pesticides. Fresenius
Environmental Bulletin 10: 451–454.
Galli, P., G. Strona, F. Benzoni, G. Crosa & F. Stefani, 2007.
Monogenoids from freshwater fish in Italy, with com-
ments on alien species. Comparative Parasitology 74:
264–272.
Gandolfi, G., S. Zerunian, P. Torricelli & A. Marconato, 1991.
I Pesci delle acque interne italiane. Istituto Poligrafico e
Zecca dello Stato (Eds), Rome.
Garcia, X. F., M. Diekmann, U. Bramick, R. Lemcke & T.
Mehner, 2006. Correlations between type-indicator fish
species and lake productivity in German lowland lakes.
Journal of Fish Biology 68: 1144–1157.
Gassner, H. & J. Wanzenbock, 2007. Application of population
size structure indices to Austrian whitefish (Coregonussp.) stocks. Fundamental and Applied Limnology. Special
Issues: Advances in Limnology 60: 377–384.
Gassner, H., J. Wanzenbock, D. Zick, G. Tischler & B.
Pamminger-Lahnsteiner, 2005. Development of a fish
based lake typology for natural Austrian lakes [50 ha
based on the reconstructed historical fish communities.
International Review of Hydrobiology 90: 422–432.
Godinho, F. N., M. T. Ferreira & M. I. Portugal e Castro, 1998.
Fish assemblage composition in relation to environmental
gradients in Portuguese reservoirs. Aquatic Living
Resources 11: 325–334.
Griffiths, A., 2006. Pattern and process in the ecological bio-
geography of European freshwater fish. Journal of Animal
Ecology 75: 734–751.
Guilizzoni, P., G. Bonomi, G. Galanti & D. Ruggiu, 1982.
Basic trophic status and recent development of some
Italian Lakes as revealed by plant pigments and other
chemical components in sediment cores. Memorie
dell’Istituto italiano di Idrobiologia 40: 79–98.
Heino, J., T. Eros, J. Kotanen & M. Rask, 2010. Describing
lake fish communities: do presence-absence and biomass
data show similar spatial and environmental relationships?
Boreal Environmental Research 15: 69–80.
Hewitt, G. M., 1999. Post-glacial recolonization of European
biota. Biological Journal of the Linnean Society 68:
87–112.
Holmgren, K. & M. Appelberg, 2000. Size structure of benthic
freshwater fish communities in relation to environmental
gradients. Journal of Fish biology 57: 1312–1330.
Irz, P., A. Laurent, S. Messad, O. Pronier & C. Argillier, 2002.
Influence of site characteristics on fish community pat-
terns in French reservoirs. Ecology of Freshwater Fishes
11: 123–136.
Irz, P., C. Argillier & T. Oberdorff, 2004. Native and intro-
duced fish species richness in French lakes: local and
regional influences. Global Ecology and Biogeography
13: 335–344.
Jackson, D. A. & H. H. Harvey, 1989. Biogeographic associ-
ation in fish assemblages: local vs regional processes.
Ecology 70: 1472–1484.
Jeppesen, E., J. P. Jensen, M. Søndergaard, T. L. Lauridsen &
F. Landkildehus, 2000. Trophic structure, species richness
and biodiversity in Danish lakes: changes along a phos-
phorus gradient. Freshwater Biology 45: 201–218.
238 Hydrobiologia (2011) 671:227–240
123

Ketmaier, V., P. G. Bianco, M. E. Cobollo & E. De Matthaeis,
2003. Genetic differentiation and biogeography in south-
ern European populations of the genus Scardinius (Pisces,
Cyprinidae) based on allozyme data. Zoologica Scripta
32: 13–22.
Koponen, K., M. S. Myers, O. Ritola, S. E. Huuskonen & P.
Lindstrom-Seppa, 2001. Histopathology of feral fish from
a PCB-contaminated freshwater lake. Ambio 30:
122–126.
Lorenzoni, M., R. Pace, G. Pedicillo, P. Viali & A. Carosi,
2009. Growth, catches and reproductive biology of ruffe
Gymnocephalus cernuus in Lake Piediluco (Umbria,
Italy). Folia Zoologica 58: 420–435.
Marchetto, A., A. Lami, S. Musazzi, J. Masaferro, L. Langone
& P. Guilizzoni, 2004. Lake Maggiore (N. Italy) trophic
history: fossil diatoms, plant pigments, chironomids and
comparison with long-term limnological data. Quaternary
International 113: 97–110.
Marchetto, F., S. Zaccara, F. M. Muenzel & W. Salzburger,
2010. Phylogeography of the Italian vairone (Telestesmuticellus, Bonaparte 1837) inferred by microsatellite
markers: evolutionary history of a freshwater fish species
with a restricted and fragmented distribution. BMC Evo-
lutionary Biology 10: 111.
Margaritora, F. G., 1992. Limnology in Latium: the volcanic
lakes. Memorie dell’Istituto Italiano di Idrobiologia 50:
319–336.
Marshall, T. R. & P. A. Ryan, 1987. Abundance patterns and
community attributes of fishes relative to environmental
gradients. Canadian Journal of Fisheries and Aquatic
Sciences 44: 198–215.
Mehner, T., M. Diekmann, U. Bramick & R. Lemcke, 2005.
Composition of fish communities in German lakes as
related to lake morphology, trophic state, shore structure
and human use intensity. Freshwater Biology 50: 70–85.
Mehner, T., K. Holmgren, T. L. Lauridsen, E. Jeppesen & M.
Diekmann, 2007. Lake depth and geographical position
modify lake fish assemblages of the European ‘Central
Plains’ ecoregion. Freshwater Biology 52: 2285–2297.
Noble, R. A. A., I. G. Cowx, D. Goffaux & P. Kestemont, 2007.
Assessing the health of European rivers using functional
ecological guilds of fish communities: standardising spe-
cies classification and approaches to metric selection.
Fisheries Management and Ecology 14: 453–463.
Noges, T., 2009. Relationships between morphometry, geo-
graphic location and water quality parameters of Euro-
pean lakes. Hydrobiologia 633: 33–43.
Oksanen, J., R. Kindt, P. Legendre, B. O’Hara & M. H. H.
Stevens, 2010. VEGAN: Community Ecology Package.
R Package. Version 1.8-8. http://cran.r-project.org/, http://
r-forge.r-project.org/projects/vegan/.
Orban, E., T. Nevigato, M. Masci, G. Di Lena, I. Casini, R.
Caproni, L. Gambelli, P. De Angelis & M. Rampacci,
2007. Nutritional quality and safety of European perch
(Perca fluviatilis) from three lakes of Central Italy. Food
Chemistry 100: 482–490.
Pedicillo, G., A. Bicchi, V. Angeli, A. Carosi, P. Viali & M.
Lorenzoni, 2008. Growth of black bullhead Ameiurusmelas (Rafinesque, 1820) in Corbara Reservoir (Umbria,
Italy). Knowledge and Management of Aquatic Ecosys-
tems 389: 05.
Penczak, T., W. Galicka, Ł. Głowacki, H. Koszalinski, A.
Kruk, G. Zieba, J. Kostrzewa & L. Marszał, 2004. Fish
assemblage changes relative to environmental factors and
time in the Warta River, Poland, and its oxbow lakes.
Journal of Fish Biology 64: 483–501.
Persson, L., S. Diehl, L. Johansson, G. Anderson & S.
F. Hamrin, 1991. Shifts in fish communities along the
productivity gradient of temperate lakes – patterns and the
importance of size structured interactions. Journal of Fish
Biology 38: 281–293.
R Development Core Team, 2010. R: A Language and Envi-
ronment for Statistical Computing. R Foundation for
Statistical Computing, Vienna, Austria. ISBN 3-900051-
07-0, URL:http://www.R-project.org.
Reyjol, Y., B. Hugueny, D. Pont, P. G. Bianco, U. Beier, N.
Caiola, F. Casals, I. Cowx, A. Economou, T. Ferreira, G.
Haidvogl, R. Noble, A. de Sostoa, T. Vigneron & T.
Virbickas, 2007. Patterns in species richness and ende-
mism of European freshwater fish. Global Ecology and
Biogeography 16: 65–75.
Salmaso, N., G. Morabito, L. Garibaldi & R. Mosello, 2007.
Trophic development of the deep lakes south of the Alps:
a comparative analysis. Fundamental and Applied Lim-
nology – Archiv fur Hydrobiologie 170: 177–196.
Schmutz, S., M. Kaufmann, B. Vogel, M. Jungwirth & S.
Muhar, 2000. A multilevel concept for fish-based, river-
type-specific assessment of ecological integrity. Hydro-
biologia 422(423): 279–289.
Steedman, R. J., T. H. Whillans, A. P. Behm, K. E. Bray, K.
I. Cullis, M. M. Holland, S. J. Stoddart & R. J. White,
1996. Use of historical information for conservation and
restoration of Great Lakes aquatic habitat. Canadian
Journal of Fisheries and Aquatic Sciences 53: 415–423.
Tammi, J., M. Appelberg, U. Beier, T. Hesthagen, A. Lappa-
lainen & M. Rask, 2003. Fish status survey of Nordic
lakes: effects of acidification, eutrophication and stocking
activity on present fish species composition. Ambio 32:
98–105.
Tartari, G., E. Buraschi, C. Monguzzi, A. Marchetto, D. Co-
petti, L. Previtali, S. Taiti, G. Barbiero & R. Pagnotta,
2004. Progetto Limno: Qualita delle Acque Lacustri ital-
iane, Vol. 1. Sintesi dei risultati. Quaderni IRSA 120:
1–333.
Tonn, W. M., J. J. Magnusson, M. Rask & J. Toivonen, 1990.
Incontinental comparison of small-lake fish assemblages:
the balance between local and regional processes. Amer-
ican Naturalist 136: 345–375.
Tortonese, E., 1971. Osteichthyes, parte I. Fauna d’Italia, Vol.
X. Calderini, Bologna.
Tortonese, E., 1975. Osteichthyes, parte II. Fauna d’Italia, Vol.
XI. Calderini, Bologna.
Volta, P., 2010. Analysis of the population structure of
Coregonus lavaretus (Linneus 1758) in three deep Italian
lakes using stock density indexes. Studi Trentini Scienze
Naturali 87: 257–260.
Volta, P. & G. Giussani, 2010. Weight–length relationships
(WLRs) and von Bertalanffy growth function (VBGF)
parameters of Alosa agone, Rutilus pigus and the anad-
romous Salmo trutta from a deep south-alpine lake: Lago
Maggiore (northern Italy). Journal of Applied Ichthyology
26: 474–476.
Hydrobiologia (2011) 671:227–240 239
123

Volta, P. & N. Jepsen, 2008. The recent invasion of Rutilusrutilus (L.) (Pisces: Cyprinidae) in a large South-Alpine
lake: Lago Maggiore. Journal of Limnology 67: 163–170.
Volta, P. & A. Oggioni, 2010. Key- and type-specific fish
species in natural lakes of Italian Alpine ecoregion
reconstructed from historical data: a preliminary index to
assess the quality status of fish fauna according to WFD
2000/60/CE. Studi Trentini Scienze Naturali 87: 97–104.
in Italian.
Volta, P., P. Tremolada, M. C. Neri, G. Giussani & S. Galassi,
2009. Age-dependent bioaccumulation of organochlorine
compounds in fish and their selective biotransformation in
top predators from Lake Maggiore (Italy). Water Air &
Soil Pollution 197: 193–209.
Wetzel, R. G., 1975. Limnology. W. B. Saunders Co., Phila-
delphia/London/Toronto.
WFD CIS, 2003. Rivers and lakes – typology, reference con-
ditions and classification systems. In: Guidance Document
No. 10. Published by the Directorate General Environ-
ment the European Commission, Brussels.
Zhao, S. Q., J. Y. Fang, C. H. Peng, Z. Y. S. L. Tang & Z. Y. S.
L. Piao, 2006. Patterns of fish species richness in China’s
lakes. Global Ecology and Biogeography 4: 386–394.
Zick, D., H. Gassner, M. Rinnerthaler, P. Jager & R. Patzner,
2007. Application of population size structure indices to
Arctic charr Salvelinus alpinus (L.) in Alpine lakes in
Austria. Ecology of Freshwater Fish 16: 54–63.
240 Hydrobiologia (2011) 671:227–240
123