arabidopsis rab geranylgeranyl transferase β-subunit mutant is constitutively photomorphogenic, and...
TRANSCRIPT

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/41561223
ArabidopsisRABgeranylgeranyltransferaseβ‐subunitmutantisconstitutivelyphotomorphogenic,andhasshootgrowthandgravitropicdefects
ARTICLEinTHEPLANTJOURNAL·FEBRUARY2010
ImpactFactor:5.97·DOI:10.1111/j.1365-313X.2010.04172.x·Source:PubMed
CITATIONS
15
READS
28
4AUTHORS,INCLUDING:
HanaSoukupova
AcademyofSciencesoftheCzechRepublic
4PUBLICATIONS285CITATIONS
SEEPROFILE
LukasSynek
AcademyofSciencesoftheCzechRepublic
13PUBLICATIONS503CITATIONS
SEEPROFILE
Availablefrom:LukasSynek
Retrievedon:04February2016

Arabidopsis RAB geranylgeranyl transferase b-subunitmutant is constitutively photomorphogenic, and hasshoot growth and gravitropic defects
Michal Hala1, Hana Soukupova1, Lukas Synek1 and Viktor Zarsky1,2*
1Institute of Experimental Botany, Academy of Sciences of the Czech Republic, Rozvojova 263, 165 02 Prague 6, Czech Republic,
and2Department of Plant Physiology, Faculty of Science, Charles University, Vinicna 5, 128 44 Prague 2, Czech Republic
Received 18 September 2009; revised 26 January 2010; accepted 4 February 2010; published online 23 March 2010.*For correspondence (fax +420 225106461; e-mail [email protected]).
SUMMARY
RAB GTPases are important directional regulators of intracellular vesicle transport. Membrane localization of
RAB GTPases is mediated by C-terminal double geranylgeranylation. This post-translational modification is
catalyzed by the a-b-heterodimer catalytic core of RAB geranylgeranyl transferase (RAB-GGT), which
cooperates with the RAB escort protein (REP) that presents a nascent RAB. Here, we show that RAB-
geranylgeranylation activity is significantly reduced in two homozygous mutants of the major Arabidopsis
b-subunit of RAB-GGT (AtRGTB1), resulting in unprenylated RAB GTPases accumulation in the cytoplasm.
Both endocytosis and exocytosis are downregulated in rgtb1 homozygotes defective in shoot growth and
morphogenesis. Root gravitropism is normal in rgtb1 roots, but is significantly compromised in shoots.
Mutants are defective in etiolation and show constitutive photomorphogenic phenotypes that cannot be
rescued by brassinosteroid treatment, similarly to the det3 mutant that is also defective in the secretory
pathway. Transcriptomic analysis revealed an upregulation of specific RAB GTPases in etiolated wild-type
plants. Taken together, these data suggest that the downregulation of the secretory pathway is interpreted as
a photomorphogenic signal in Arabidopsis.
Keywords: RAB GTPases, cell expansion, photomorphogenesis, vesicle trafficking, Arabidopsis thaliana,
prenylation.
INTRODUCTION
RAB proteins represent the largest subset of the Ras
superfamily of small GTPases in yeast and animals, as well
as in plants (Pereira-Leal and Seabra, 2001; Rutherford and
Moore, 2002). Although only some members of the RAB
family were characterized on the functional level, it seems
that most RABs are involved in the regulation of various
steps of vesicular transport within the secretion and
endocytotic pathways, including vesicle budding, transport
along the cytoskeleton and vesicle fusion with target
membranes (Deneka et al., 2003; Novick et al., 2006;
Fukuda, 2008; Woollard and Moore, 2008). The number of
distinct RABs varies considerably among species: 11 in
Saccharomyces cerevisiae, 29 in Caenorhabditis elegans,
40–60 in angiosperm plants and more than 60 in mammals
(Pereira-Leal and Seabra, 2001; Woollard and Moore,
2008).
Membrane localization of hydrophilic RAB proteins is
achieved by their post-translational hydrophobic modifica-
tion catalyzed by the RAB geranylgeranyl transferase (RAB-
GGT), one of three protein prenyl transferases present in
eukaryotic cells (Maurer-Stroh et al., 2003). Most RABs
contain a C-terminal motif with two cysteine residues
(usually -XXCC or -XCXC), to which geranylgeranyl moieties
are covalently attached via a thioether bond. Although RAB-
GGT prenylates exclusively RAB GTPases, it remains unclear
whether it is the only enzyme geranylgeranylating RABs
in vivo, as several members of the RAB family (e.g. the
mammalian Rab8 containing the -CXXX C-terminal motif)
have also been shown to serve as a substrate for geranyl-
geranyl transferase I (GGTI) in vitro (Wilson et al., 1998). In
plants, a novel type of RABs lacking the C-terminal preny-
lation motif was reported, and was shown to associate with
ª 2010 The Authors 615Journal compilation ª 2010 Blackwell Publishing Ltd
The Plant Journal (2010) 62, 615–627 doi: 10.1111/j.1365-313X.2010.04172.x

membranes via N-terminal myristoylation and palmitoyla-
tion (Bolte et al., 2000; Ueda et al., 2001). The functional
cycle of these RABs seems to be RAB-GDI-independent
(Ueda et al., 2001).
Similarly to other protein prenyl transferases, RAB-GGT is
a heterodimer of tightly associated a- and b-subunits
(Maurer-Stroh et al., 2003). However, in contrast to farnesyl
transferase and GGTI, it does not recognize its protein
substrate directly, but only in a complex with an accessory
RAB escort protein (REP). The mechanism of the reaction
has been well described in mammals and yeast (Anant et al.,
1998; Thoma et al., 2001a,b). Nascent RAB GTPase and REP
form a cytoplasmic complex which then binds to RAB-GGT
in a prenyl-substrate-dependent manner. Recently, Baron
and Seabra (2008) proposed that an alternative scenario
might be prevalent in vivo: first, REP binds to a catalytic
a-b heterodimer, and then nascent RAB is loaded into this
pre-assembled ternary complex. Geranylgeranylation pro-
ceeds step by step so that mono-geranylgeranylated inter-
mediates can be isolated (Shen and Seabra, 1996; Thoma
et al., 2001a). The use of mutant RAB GTPases partially or
completely lacking a prenylation motif inhibits the activity of
RAB-GGT, as shown for the plant RAB GTPase LeRab1 by
Loraine et al. (1996).
The interface of the REP-RAB-GGT complex is limited to a
relatively small area, and involves a combination of hydro-
phobic and charge interactions. It was shown that the
a-subunit of RAB-GGT undergoes a series of conformational
changes preceded by the formation of the a-b dimer in the
presence of a phosphoisoprenoid substrate. These changes
also force the REP-binding domains into the right confor-
mation for binding the REP-RAB complex (Pylypenko et al.,
2003).
Loss-of-function mutations in RAB-GGT subunits in yeast
are lethal (http://www.yeastgenome.org). In animals, the
gunmetal mouse, carrying a mutant allele of the RAB-GGT
a-subunit with mutated splicing site, displays reduced RAB
geranylgeranylation activity, resulting in a weak phenotype
characterized by platelets malformation (Detter et al., 2000).
Mutation in human REP1 causes choroideremia disease, an
X-chromosome-linked retinal dystrophy (Cremers et al.,
1992; Merry et al., 1992; Seabra et al., 1992). Surprisingly,
it seems that the mutation of the single C. elegans REP
selectively affects only specific RABs, whereas others are
correctly geranylated and localized in the supposed absence
of wild-type REP (Tanaka et al., 2008).
Protein prenylation was first documented in plants by
Randall et al. (1993). The Arabidopsis farnesyl transferase
has been found to be involved in abscisic acid-dependent
plant developmental processes, as demonstrated by the
phenotype of era1, a mutant in the farnesyl transferase
b-subunit (Cutler et al., 1996), or in the a-subunit mutant plp
(Running et al., 2004). Also, the ggb mutant in the Arabid-
opsis geranylgeranyl transferase I b-subunit is compro-
mised in some aspects of ABA and auxin responses;
interestingly, the phenotype of a double mutant combining
ggb and era1 lesions is identical to that of the plp mutant
(Johnson et al., 2005). Functional analysis of the Arabidop-
sis CaaX processing enzymes also using, among others,
RNAi suppression of isoprenyl cysteine methyltransferase,
showed that phenotypic deviations of these RNAi plants are
somewhat similar to those in the era1 mutant phenotype
(Bracha-Drori et al., 2008).
Biermann et al. (1996) characterized several prenylated
GTPases in plants, and also described the RAB-GGT activity
prenylating recombinant Rab1 and Rab2 in tobacco extracts.
Yalovsky et al. (1996) reported a wide substrate specificity of
different plant RAB-GGTs in total extracts, a preference for
geranylgeranyl pyrophosphate (GGPP) and an inhibitory
effect of RAB GTPases lacking C-terminal cysteine motifs.
Loraine et al. (1996) observed an ability of tomato Rab1 to
undergo geranylgeranylation in the yeast extract in vitro,
and to complement the yeast ypt1 mutant. We have previ-
ously characterized a single Arabidopsis RAB escort protein,
and demonstrated its plant-specific structural feature in the
RAB-GGT interaction with REP: site-directed amino acid
substitution into the Opisthokontian consensus sequence
allowed plant REP to complement mrs6, the yeast REP
mutant (Hala et al., 2005). Recently, the plant-specific plas-
tidial pathway of prenylation precursors biosynthesis was
shown to be crucial for protein geranylgeranylation in plant
cells (Gerber et al., 2009).
In this paper, we report that a mutation in the major
b-subunit of Arabidopsis RAB-GGT results in RAB geranyl-
geranylation deficiency, causes distinct phenotypic changes
and affects many physiological processes, such as shoot
gravitropism and cell elongation. Surprisingly, rgtb1
mutants also exhibit a constitutive photomorphogenesis
phenotype.
RESULTS
RAB geranylgeranyl transferase in the Arabidopsis genome
A search of the Arabidopsis genome revealed the presence
of two genes encoding RAB-GGT a-subunits (AtRGTA1, lo-
cus At4g24490; AtRGTA2, locus At5g41820), as well as two
genes encoding RAB-GGT b-subunits (AtRGTB1, locus
At5g12210; AtRGTB2, locus At3g12070).
The AtRGTA1 protein shares 27% sequence identity and
41% similarity with the human RAB-GGT a-subunit;
AtRGTB1 shares 56% identity and 71% similarity with the
human RAB-GGT b-subunit. Although AtRGTA2 and
AtRGTA1 subunits share 66% identity and 74% similarity
with each other, AtRGTB1 and AtRGTB2 are more similar to
each other (83% identity and 89% similarity).
The presence of two RAB-GGT subunits in the Arabidop-
sis genome is unusual when compared with other non-plant
eukaryotes. To test whether this is a common plant feature,
616 Michal Hala et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

we constructed a phylogenetic tree of protein prenyl trans-
ferase b-subunits (Figure S1). Both b-subunits of Arabidop-
sis thaliana, RGTB1 and RGTB2, branch together, but
separately from orthologous couples in Vitis vinifera, Physc-
omitrella patens, and Selaginella moellendorffii. This obser-
vation implies independent recent duplication events.
Comparison of expression profiles for all RAB-GGT genes
and REP using Genevestigator (Zimmermann et al., 2004)
revealed the relatively low level of AtREP expression in all
tissues examined (also in Hala et al., 2005; Wojtas et al.,
2007). AtRGTB1 mRNA is generally 10 times more abundant
than AtRGTB2 mRNA in sporophytes, whereas levels of both
mRNAs are comparable in male gametophytes. AtRGTA1 is
expressed in all tissues, in contrast to AtRGTA2, which is
only significantly expressed in pollen (Figure S2).
Disruption of Arabidopsis RGTB1 results in pleiotropic
growth and development defects
Two independent T-DNA insertional mutants in the
AtRGTB1 gene, rgtb1-1 and rgtb1-2, were obtained from the
SALK collection (Appendix S1). After germination the pop-
ulation started to segregate, with one-quarter producing
dwarf plants, both in vitro and in soil growth conditions.
These plants were confirmed by PCR genotyping to be
homozygous mutants.
Both mutant lines exhibited an identical phenotype: root-
tip organization, cell differentiation in the elongation zone,
total length of the primary root, as well as root-hair
formation were unaffected, and so were similar to that of
the wild-type (WT) plants, although a weak tendency
towards longer primary roots was observed in 1-week-old
etiolated mutants (see below). However, detailed study
revealed that differences between rgtb1 homozygotes and
WT plants are prominent in the shoot. The leaves of the
homozygotes became epinastic and smaller than wild-type
leaves (Figure 1a). The number of rgtb1-1 and rgtb1-2
rosette leaves is statistically indistinguishable from the
number in WT plants, but the diameter of rosettes is much
lower in homozygous mutants (Figure 1a,b).
Homozygous rgtb1-1 and rgtb1-2 plants produce multiple
inflorescence stems, indicating a loss of apical dominance
(Figure 1c). Although the number of floral organs remains
constant, mutant flowers never open fully, featuring distinct
pistil protuberance (Figure 1d). Flowering in homozygous
rgtb1-1 and rgtb1-2 plants started at the same time as in the
WT, but the siliques remained mostly sterile. Reciprocal
crossing with WT plants, however, resulted in fertilization,
regardless of whether rgtb1-1 or rgtb1-2 were pollen donors
or recipients. In all cases, the progeny was comprised of a
(a)
(b)
(c)
(d)
Figure 1. Phenotype of homozygous rgtb1
mutants.
(a) Two-week-old plants. Scale bars: 5 mm.
(b) Three-week-old plants.
(c) Four-week-old plants.
(d) Flowers of 4-week-old plants.
RAB prenylation and photomorphogenesis 617
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

uniform population of heterozygotes, suggesting that
mutant pollen is functional.
Although wild-type plants are typically more than 24 cm
high 30 days after germination, rgtb1-1 and rgtb1-2 homo-
zygotes reach only approximately 6 cm in height (Figure 1c).
In addition, the onset of senescence in rgtb1-1 and rgtb1-2
plants is delayed by 2 months, compared with wild-type
plants, in our growth conditions.
Biochemical characterization of rgtb1 mutants
Using RT-PCR, we analyzed homozygous rgtb1-1 and rgtb1-
2 mutants as well as WT plants for the expression of mRNA
encoding the RGTB1 subunit. It turned out that the full-
length mRNA of the RGTB1 subunit was absent in the
homozygous mutants tested, whereas actin (ACT2) and
RGTB2 were expressed at the normal level (see Figure S3).
Next, we tested the possible presence of unprenylated
RAB GTPases, substrates of the RAB-GGT, in rgtb1-1 and
rgtb1-2 homozygotes. Western blot analysis using the anti-
AtRAB-A2a antibody (provided by I. Moore, University of
Oxford) revealed a band of AtRAB-A2a in the WT. However,
in the case of both rgtb1-1 and rgtb1-2 homozygotes, this
band was accompanied by other weaker bands of higher
molecular weight, probably representing unprenylated
AtRAB-A2a (Figure 2a). This shift in mobility might be
caused by the well characterized more effective SDS binding
to the prenyl-modified proteins under conditions of SDS-
PAGE (Sanford et al., 1995). No accumulation of such
possibly unprenylated RAB GTPases was observed in the
WT. To further address the subcellular localization of these
putatively unprenylated RAB GTPases, we prepared micro-
somal and cytosolic fractions from etiolated hypocotyls of
WT and rgtb1-1 homozygotes. Although a large pool of
AtRAB-A2a was still attached to the membranes, a signifi-
cant accumulation of a higher molecular weight second
band of possibly unprenylated RAB occured in the cytosolic
fraction of rgtb1-1 homozygote hypocotyls (Figure S4).
In order to compare the activity of RAB-GGT in vitro,
extracts from 3-week-old light-grown plantlets were pre-
pared, and the specific RAB-GGT activity was tested by the
addition of recombinant AtRAB-A2a as a substrate.
Figure 2(b) shows that the RAB geranylgeranylation activity
in both rgtb1-1 and rgtb1-2 homozygous mutants was
reduced to �25% of the WT level (similar results were seen
using a few other recombinant RABs as substrates; data not
shown). As both rgtb1-1 and rgtb1-2 exhibit identical
phenotypes and biochemical characteristics mentioned
above, we used only rgtb1-1 for further experiments.
Arabidopsis rgtb1 mutants show gravitropic defect in the
shoot
To test for a possible defect in gravitropic response, often
associated with mutations in vesicle transport organizers
(e.g. SNARE proteins; Kato et al., 2002; Yano et al., 2003; for
a review see Bassham and Blatt, 2008), we set up a gravi-
tropic assay on roots in a light-grown segregating popula-
tion of the rbtg1-1/RGTB1 progeny. Ten days after
germination, the vertical agar plates were rotated by 90�.Roots of all plants responded to the change in gravity vector
equally (Figure 3a). Next, pots containing 3-week-old WT
plants or rgtb1-1 mutants were rotated by 90�, and shoot
gravitropism was tested. Plants were kept in the dark for
12 h before images were taken. Shoots of all WT plants re-
sponded to the new gravity direction, whereas no response
was observed in rgtb1-1 homozygotes (Figure 3b).
Arabidopsis rgtb1 mutants exhibit secretory defects
RAB GTPases are important regulators of vesicle trafficking
in the cell. Deficiency in RAB geranylgeranylation activity in
the rgtb1-1 mutants, as documented above, should result in
the defective functioning of the secretory machinery, there-
by leading to impaired cell elongation. To test this hypoth-
esis, we crossed rgtb1-1 mutants with plants transformed
with the secretory GFP construct (secGFP, provided by I.
Moore). Under normal conditions, secGFP is secreted out of
the cell, and the environment outside the cell (low pH and
(a)
(b)
Figure 2. Biochemical characteristics of homozygous rgtb1 mutants.
(a) Western blot analysis in extracts from rgtb1-1, rgtb1-2 and wild-type (WT)
plants using rabbit polyclonal anti-AtRAB-A2a antibody (1:1000). Two differ-
ent forms of AtRAB-A2a are labeled: unprenylated AtRAB-A2a and prenylated
AtRAB-A2a-GG.
(b) RAB geranylgeranylation activities detected after the addition of 4 lg of
recombinant AtRAB-A4a in cell extracts from rgtb1-1 and rgtb1-2 homozy-
gous mutants. Each column represents three independent measurements;
bars correspond to standard deviations.
618 Michal Hala et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

presence of proteases) quenches the fluorescence. When the
secretion is perturbed secGFP accumulates inside the cell,
and a fluorescent signal becomes visible (Batoko et al.,
2000).
We tested 1-week old dark-grown progeny of an rgtb1-1/
RGTB1 heterozygote expressing secGFP. We observed that
secGFP expression did not alter the rgtb1 phenotype.
Microscopic analysis revealed significantly higher accumu-
lation of secGFP inside the cells of rgtb1-1 hypocotyls
(Figure 4a,c), in contrast to the WT (Figure 4b,d). In rgtb1-1
cells, confocal microscopy showed that secGFP is largely
accumulated in diverse internal structures [such as the
endoplasmic reticulum (ER) and ER-like bodies], rather
than in the cytoplasm (Figure 4c). However, the strongly
increased overall fluorescence of rgtb1 mutant hypocotyls is
partly the result of increased non-specific autofluorescence.
Endocytosis is an essential process of the secretory
pathway, and is as important as exocytosis. Using confocal
microscopy, we visualized endocytic compartments by
vacuum infiltration of the styryl dye FM4-64 in etiolated
hypocotyls of 1-week-old seedlings. Whereas visible
FM-labeled moving particles (endocytic compartments)
soon appeared, reproducibly, inside the hypocotyl cells in
WT plants (Figure 4h–j), only a few of them were observed in
rgtb1-1 homozygotes up to 90 min (Figure 4e–g). Using
image analysis, we measured the fluorescence inside hypo-
cotyl cells cytoplasm. The average fluorescence was
17.5 � 2.0 relative units in WT cells and 7.3 � 4.3 relative
units in rgtb1 cells (n = 20; Student’s t-test P < 0.001).
Moreover, between 30–90 min after treatment with brefel-
din A (BFA), BFA compartments emerged in WT hypocotyl
cells, but not in rgtb1-1 hypocotyl cells. Taken together, we
conclude that the rgtb1-1 mutant has defects in the secretory
pathways.
We also observed roots of rgtb1 mutants in both exper-
iments. As expected, based on the WT-like root phenotypes
in both rgtb1 mutants, root cells secrete secGFP and perform
FM4-64 endocytosis like the WT (data not shown).
Arabidopsis rgtb1 mutant homozygotes show de-etiolated
phenotype in the dark
The dwarf phenotype of light-grown rgtb1 mutants led us to
test the ability of rgtb1 cells to elongate in the dark during
etiolation (skotomorphogenesis). We germinated a segre-
gating population of rgtb1-1/RGTB1 heterozygote progeny
in the dark. After 7 days we found a segregating subpopu-
lation of plantlets with short hypocotyls (Figures 5a and 6)
and opened upright cotyledons (Figure 6a). Mutant
hypocotyls consisted of irregularly patterned short cells
instead of the bundles of highly elongated cells typical for
the WT (Figure 5b), and possessed a portion of dead cells.
This subpopulation represented 26% of all plantlets
(n = 120), and following genotyping confirmed they were
rgtb1-1 homozygotes. At this stage, the rgtb1-1 phenotype is
very similar to the de-etiolated 3 (det3) phenotype, a repre-
sentative of a group of mutant plants showing de-etiolation
and constitutive photomorphogenesis obtained under the
same conditions (Figure 6).
To test for possible brassinosteroid dependence of the
rgtb1-1 phenotype, we grew etiolated seedlings on vertical
agar supplemented with 10)6, 10)7, 10)8 or 10)9M 24-
epibrassinolide. We saw no rescue of the rgtb1-1 phenotype
by external brassinosteroid addition for either concentration
(data not shown), as is the case for det3 (Szekeres et al.,
1996).
As the phenotype of dark-grown rgtb1-1 homozygotes
was similar to known det/cop mutants, we prolonged the
dark cultivation of plants for up to 3 weeks. Similarly to the
previous experiment, the population (n = 120) segregated
out 27% of plants exhibiting a constitutively photomorpho-
genic phenotype, which were proven by genotyping to be
rgtb1-1 homozygotes. These plants have fully opened
cotyledons with long petioles, the apical meristem protrud-
(a)
(b)
Figure 3. Gravitropic response of rgtb1-1 plants.
(a) Root gravitropism of the segregating population of rgtb1-1/RGTB1
progeny. Ten days after germination the plate was rotated by 90�. Plants are
labeled according to the later genotyping.
(b) Shoot gravitropism assay. Pots with either wild-type (WT) or rgtb1-1 plants
(3-weeks old) were reoriented and cultivated for 12 more hours. Arrows show
the gravity vector before (1.) and after (2.) reorientation.
RAB prenylation and photomorphogenesis 619
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

ing and are developing the first true leaves (Figure 6b).
Obviously, the phenotypic deviations of the 3-week-old det3
mutant grown under the same conditions are more pro-
nounced as compared with rgtb1 (e.g. higher number of
leaves and shorter petioles). The growth direction of dark-
grown rgtb1 plants was variable on vertical agar plates,
pointing again to the defect in shoot gravitropism.
Many RAB GTPases are upregulated during the etiolation
growth of Arabidopsis
The rapid elongation growth of etiolated Arabidopsis seed-
lings in the dark is supported by the activated exocytotic
machinery. We were interested whether expression levels of
RAB GTPases in dark-grown plantlets are generally
(a) (e) (f) (g)
(h) (i) (j)
(b)
(c)
(k) (l)
(d)
Figure 4. Vesicle trafficking defects in rgtb1-1 plants.
(a) Hypocotyls of the 1-week-old dark-grown rgtb1-1 mutant expressing secGFP. Scale bar: 100 lm.
(b) Hypocotyls of a 1-week-old dark-grown wild-type (WT) plant expressing secGFP. Scale bar: 100 lm.
(c) Confocal laser scanning microscope (CLSM) projection through an rgtb1-1 hypocotyl expressing secGFP. Scale bar: 50 lm.
(d) CLSM projection through a WT hypocotyl expressing secGFP. Scale bar: 50 lm.
(e, f, g) Endocytosis in hypocotyls of 1-week-old dark-grown rgtb1-1 plants visualized by FM4-64 dye: (e) immediately after vacuum infiltration, (f) 30 min later and (g)
90 min later. Scale bar: 20 lm.
(h, i, j) Endocytosis in hypocotyls of 1-week-old dark-grown WT plants visualized by FM4-64 dye: (e) immediately after vacuum infiltration, (f) 30 min later and (g)
90 min later. Scale bar: 20 lm.
(k) CLSM projection through a 1-week-old dark-grown rgtb1-1 hypocotyl treated with 50 lM brefeldin A (BFA) for 90 min. Diverse stable patches along the cell
periphery emitting autofluorescence were common in rgtb1-1 hypocotyls, in contrast to those of the WT. Scale bar: 50 lm.
(l) CLSM projection through a 1-week-old dark-grown WT hypocotyl treated with 50 lM BFA for 90 min. Scale bar: 50 lm.
620 Michal Hala et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

increased, as predicted for two pea RAB-A proteins (Yoshida
et al., 1993; Nagano et al., 1995). Therefore, we performed
microarray analysis of gene expression on both light- and
dark-grown 7-day-old WT seedlings using the Arabidopsis
ATH1 chip (Table 1, Table S1 and Table S2).
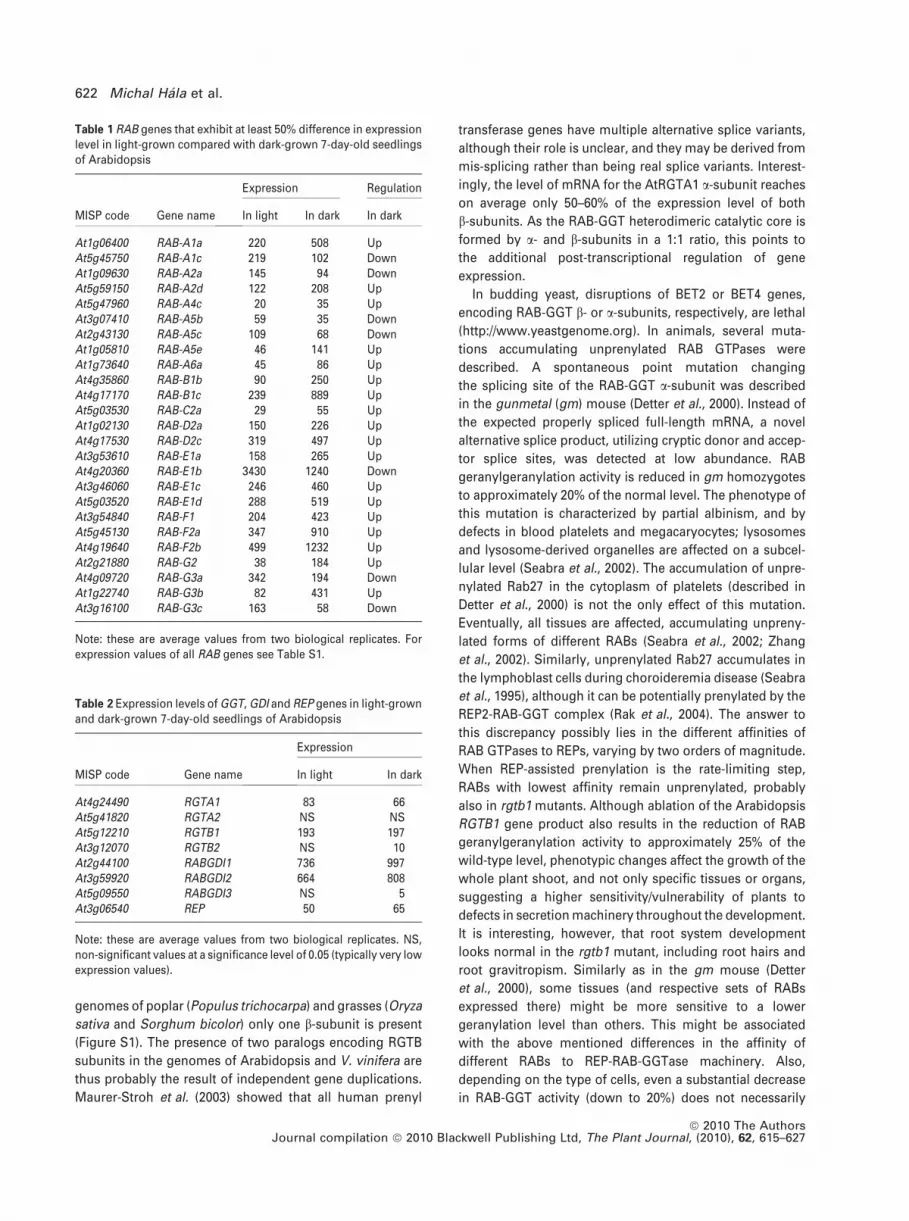
We found that one-third of Arabidopsis RAB GTPase
paralogs (especially in the B, D, E and F subfamilies) were at
least 50% upregulated in WT dark-grown plants when
compared with light-grown plants. Of these, RAB-G3b, -G2,
-B1c and -A5e showed at least a threefold increase in their
mRNA expression and RABGDI1 and 2 are also upregulated
(Tables 1 and 2). However, there was also one-tenth of
downregulated RAB GTPases, namely RAB-G3a, -G3c, -E1b
and -A1c in the dark-grown Arabidopsis plantlets. In any
case, these results are in accord with the data mentioned
above on the dark-induced upregulation of pea RABs, and
show the importance of the upregulation of RAB GTPases
for the implementation of the skotomorphogenic develop-
mental program.
DISCUSSION
Using phylogenetic analyses, we have found that different
angiosperm species acquired independently duplicated
loci encoding RAB-GGT subunits. As in Arabidopsis, two
paralogs of the b-subunit can also be found in the genomes
of V. vinifera, P. patens and S. moellendorffii, whereas in
(a)
(b)
Figure 5. Length of hypocotyls and roots and hypocotyl cell morphology of
dark-grown rgtb1-1 mutants.
(a) Length of hypocotyls (in white) and roots (in black) of rgtb1-1 and wild-type
(WT) plants. Scale bars correspond to standard deviations (n > 132 for each
column). For hypocotyl length, Student’s t-test P < 0.001; for root length,
Student’s t-test P < 0.001.
(b) Morphology of hypocotyl cells in dark-grown 1-week-old seedlings labeled
by 20 mM propidium iodide for 10 min (CLSM projections). Scale
bars: 50 lm.
(a)
(b)
Figure 6. Morphology of dark-grown rgtb1-1 seedlings.
(a) One-week-old rgtb1-1, det3 and wild-type (WT) seedlings grown in the
dark. Only the apical part of the WT is shown (the total hypocotyl length was
18 mm). Scale bars: 2 mm.
(b) Three-week-old rgtb1-1, det3 and WT seedlings grown in the dark. Only the
apical part of the WT is shown (the total hypocotyl length was 19 mm). Scale
bar: 2 mm.
RAB prenylation and photomorphogenesis 621
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627
genomes of poplar (Populus trichocarpa) and grasses (Oryza
sativa and Sorghum bicolor) only one b-subunit is present
(Figure S1). The presence of two paralogs encoding RGTB
subunits in the genomes of Arabidopsis and V. vinifera are
thus probably the result of independent gene duplications.
Maurer-Stroh et al. (2003) showed that all human prenyl
transferase genes have multiple alternative splice variants,
although their role is unclear, and they may be derived from
mis-splicing rather than being real splice variants. Interest-
ingly, the level of mRNA for the AtRGTA1 a-subunit reaches
on average only 50–60% of the expression level of both
b-subunits. As the RAB-GGT heterodimeric catalytic core is
formed by a- and b-subunits in a 1:1 ratio, this points to
the additional post-transcriptional regulation of gene
expression.
In budding yeast, disruptions of BET2 or BET4 genes,
encoding RAB-GGT b- or a-subunits, respectively, are lethal
(http://www.yeastgenome.org). In animals, several muta-
tions accumulating unprenylated RAB GTPases were
described. A spontaneous point mutation changing
the splicing site of the RAB-GGT a-subunit was described
in the gunmetal (gm) mouse (Detter et al., 2000). Instead of
the expected properly spliced full-length mRNA, a novel
alternative splice product, utilizing cryptic donor and accep-
tor splice sites, was detected at low abundance. RAB
geranylgeranylation activity is reduced in gm homozygotes
to approximately 20% of the normal level. The phenotype of
this mutation is characterized by partial albinism, and by
defects in blood platelets and megacaryocytes; lysosomes
and lysosome-derived organelles are affected on a subcel-
lular level (Seabra et al., 2002). The accumulation of unpre-
nylated Rab27 in the cytoplasm of platelets (described in
Detter et al., 2000) is not the only effect of this mutation.
Eventually, all tissues are affected, accumulating unpreny-
lated forms of different RABs (Seabra et al., 2002; Zhang
et al., 2002). Similarly, unprenylated Rab27 accumulates in
the lymphoblast cells during choroideremia disease (Seabra
et al., 1995), although it can be potentially prenylated by the
REP2-RAB-GGT complex (Rak et al., 2004). The answer to
this discrepancy possibly lies in the different affinities of
RAB GTPases to REPs, varying by two orders of magnitude.
When REP-assisted prenylation is the rate-limiting step,
RABs with lowest affinity remain unprenylated, probably
also in rgtb1 mutants. Although ablation of the Arabidopsis
RGTB1 gene product also results in the reduction of RAB
geranylgeranylation activity to approximately 25% of the
wild-type level, phenotypic changes affect the growth of the
whole plant shoot, and not only specific tissues or organs,
suggesting a higher sensitivity/vulnerability of plants to
defects in secretion machinery throughout the development.
It is interesting, however, that root system development
looks normal in the rgtb1 mutant, including root hairs and
root gravitropism. Similarly as in the gm mouse (Detter
et al., 2000), some tissues (and respective sets of RABs
expressed there) might be more sensitive to a lower
geranylation level than others. This might be associated
with the above mentioned differences in the affinity of
different RABs to REP-RAB-GGTase machinery. Also,
depending on the type of cells, even a substantial decrease
in RAB-GGT activity (down to 20%) does not necessarily
Table 2 Expression levels of GGT, GDI and REP genes in light-grownand dark-grown 7-day-old seedlings of Arabidopsis
MISP code Gene name
Expression
In light In dark
At4g24490 RGTA1 83 66At5g41820 RGTA2 NS NSAt5g12210 RGTB1 193 197At3g12070 RGTB2 NS 10At2g44100 RABGDI1 736 997At3g59920 RABGDI2 664 808At5g09550 RABGDI3 NS 5At3g06540 REP 50 65
Note: these are average values from two biological replicates. NS,non-significant values at a significance level of 0.05 (typically very lowexpression values).
Table 1 RAB genes that exhibit at least 50% difference in expressionlevel in light-grown compared with dark-grown 7-day-old seedlingsof Arabidopsis
MISP code Gene name
Expression Regulation
In light In dark In dark
At1g06400 RAB-A1a 220 508 UpAt5g45750 RAB-A1c 219 102 DownAt1g09630 RAB-A2a 145 94 DownAt5g59150 RAB-A2d 122 208 UpAt5g47960 RAB-A4c 20 35 UpAt3g07410 RAB-A5b 59 35 DownAt2g43130 RAB-A5c 109 68 DownAt1g05810 RAB-A5e 46 141 UpAt1g73640 RAB-A6a 45 86 UpAt4g35860 RAB-B1b 90 250 UpAt4g17170 RAB-B1c 239 889 UpAt5g03530 RAB-C2a 29 55 UpAt1g02130 RAB-D2a 150 226 UpAt4g17530 RAB-D2c 319 497 UpAt3g53610 RAB-E1a 158 265 UpAt4g20360 RAB-E1b 3430 1240 DownAt3g46060 RAB-E1c 246 460 UpAt5g03520 RAB-E1d 288 519 UpAt3g54840 RAB-F1 204 423 UpAt5g45130 RAB-F2a 347 910 UpAt4g19640 RAB-F2b 499 1232 UpAt2g21880 RAB-G2 38 184 UpAt4g09720 RAB-G3a 342 194 DownAt1g22740 RAB-G3b 82 431 UpAt3g16100 RAB-G3c 163 58 Down
Note: these are average values from two biological replicates. Forexpression values of all RAB genes see Table S1.
622 Michal Hala et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

cause a significant accumulation of unprenylated RABs (see
e.g. Seabra et al., 2002; Zhang et al., 2002). Genevestigator
data show that the expression level of RGTB2, the second
paralog, varies around 20% of RGTB1 expression on the
mRNA level, showing the potential to balance the lack of
RGTB1 activity.
Two major developmental abnormalities, apart from loss
of apical dominance and small stature, are typical for rgtb1
mutants: defective shoot gravitropic response (Figure 3) and
constitutive photomorphogenesis (Figure 6). Gravity sens-
ing is localized to the inflorescence stem endodermis in
Arabidopsis. At the tissue level, two opposite gradients of
auxin and jasmonate are established upon gravitropic
signaling, and these gradients are sufficient to start the
gravitropic response (Gutjahr et al., 2005). At the subcellular
level, it was shown that the normal gravitropic response
depends partially on vesicle transport from the trans-Golgi
network (TGN) to the vacuole – a process that might be
affected by the RGTB1 mutation along with the secretion.
The vacuolar t-SNARE protein AtVAM3 and the soluble
N-ethylmaleimide-sensitive factor AtVNI11 forming a
complex were shown to be necessary for shoot gravitro-
pism, directly affecting tonoplast dynamics and amyloplast
localization (Yano et al., 2003). Similarly, the GRAVITRO-
PISM DEFECTIVE 2 (GRV2) protein was shown to be local-
ized in the prevacuolar compartment, and to cooperate in
the final trafficking from this compartment to the vacuole
(Silady et al., 2008).
The observation of dark-grown mutant plants uncovered
the most interesting aspect of the rgtb1 mutant phenotype:
the de-etiolated (det) or constitutive photomorphogenesis
(cop) phenotypes. Many cop/det mutants were shown to be
brassinosteroid-deficient (reviewed in Schumacher and
Chory, 2000; Bishop and Koncz, 2002). Brassinosteroid-
deficient plants typically exhibit the development of primary
leaves in the dark, apical hook and cotyledons opening, and
thick short hypocotyls – phenotypes complemented by the
external addition of brassinosteroids (BRs) (Szekeres et al.,
1996). However, rgtb1 mutants seem to be most similar to
the det3 mutant. Szekeres et al. (1996) realized that in
contrast to most other cop/det mutants, det3 is insensitive
to complementation by BR treatment.
Arabidopsis det3 mutants grown in light lose apical
dominance, and are smaller than the WT (Cabrera y Poch
et al., 1993). When Schumacher et al. (1999) identified DET3
as the C-subunit of the vacuolar-type H+-ATPase (VHA), they
also found that its mutation leads to a defect in the execution
of the actual dark growth response, rather than in the
signaling pathways initiating it. Dettmer et al. (2006) and
Brux et al. (2008) proved that VHA function in plants is
essential for the operation of the secretory pathway – both
endocytosis and exocytosis via TGN (which serves as an
endosome in plants; Dettmer et al., 2006). Arabidopsis
mutants have shown that the VHA is essential for Golgi
function in pollen development, and during embryogenesis
(Dettmer et al., 2005; Strompen et al., 2005). All these results
imply that the growth inhibition observed in plants with
reduced VHA activity is caused by a defect in vesicle
trafficking, rather than by reduced turgor pressure attribut-
able to a lack of osmolyte transport into the vacuole (Dettmer
et al., 2005). Mutants in DET3, COP4 and SHY2/IAA3 also
exhibit a defective gravitropic response: shy2 and det3
exhibit a weaker defect in hypocotyls, whereas cop4 exhibits
the defect in both roots and hypocotyls (Hou et al., 1993; Kim
et al., 1998; Schumacher et al., 1999).
In all these aspects, we can see a remarkable similarity to
the rgtb1 mutant, which also exhibits a block in the secretory
pathway (both exocytosis and endocytosis), obviously via
insufficient prenylation of RAB GTPases. Already in the first
reports on plant RAB GTPases, the negative effect of light
(mediated by phytochrome) on RAB expression in etiolated
pea plants was recognized (Yoshida et al., 1993; Nagano
et al., 1995); likewise, the downregulation of a specific
RAB-A homolog in tobacco seems to induce hypocotyl
shortening in the dark (Kang et al., 2001). We have fully
corroborated these early observations in our Arabidopsis
transcriptome analysis, as we have found that group of RAB
GTPases, and RABGDI1 and RABGDI2 are upregulated in
dark-grown seedlings. Publicly available expression data
reveal that at least three RAB genes are significantly
deregulated in the det3 mutant under dark conditions
(Newman et al., 2004). Two of them also show the same
regulation in our expression analysis of the photomorpho-
genic WT: RAB-B1c is downregulated, whereas RAB-E1b is
upregulated. At least five RAB genes (RAB-A1f, -A2b, -A4a,
-A6a and -G3c) are also significantly deregulated in the cop1
mutant. Interestingly, the expression of RGTB1 is signifi-
cantly downregulated compared with the WT.
Mutant det3 and rgtb1 phenotypes fit very well into the
notion that re-programming and a boost of the secretory
pathway is necessary to support remarkable elongation
growth of etiolated plantlets. However, why do lesions in
secretory pathways result in the full-blown development of
the cop/det phenotype: i.e. why does compromised secre-
tion induce photomorphogenesis in the dark, and not just a
cessation of hypocotyl elongation? In this respect we can
only speculate. In the case of the conditional det3 mutant
phenotype, the involvement of oxylipin and ethylene in
mediating changes in gene expression in response to
cellulose deficiency were implicated in the development of
the cop/det phenotype (Brux et al., 2008). It might be
expected that the deficient secretory pathway of the Arabid-
opsis rgtb1 mutant could also result in a defective cell wall
composition, accompanied by a similar stress response as
described for the det3 mutant ultimately leading to the
cop/det phenotype. However, it is equally possible that a
direct signaling feedback relay from the secretory pathway
to plant morphogenic programs also exists, switching on
RAB prenylation and photomorphogenesis 623
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

photomorphogenesis when vesicle trafficking is downregu-
lated. This might obviously be the case in plants treated by
endosidin1 (ES1), a new anti-endocytosis drug, which
releases the ICR1 adaptor protein from ROP GTPases at the
tip of growing pollen tubes. ES1 inhibits secretion specifi-
cally via TGN-dependent endocytosis, and induces the cop/
det phenotype when applied to the whole dark-grown
seedlings (this cannot be relieved by the addition of BRs;
Robert et al., 2008). The report suggests that a compromised
secretory pathway might influence BR signaling by interfer-
ing with the BRI1 signaling endosome (Robert et al., 2008).
So there might be dark/light and BR signaling upstream of
the secretory pathway execution of etiolated growth, but
also the secretory pathway might reciprocally influence
signal transduction pathways regulating photomorphogen-
esis (e.g. BR signal transduction via a signalling endosome).
In any case, cell elongation growth supported by de novo
translated RAB GTPases is an important part for the imple-
mentation of the dark-induced morphogenetic programme
in Angiosperms, and the efficient geranylation of nascent
RAB GTPases is necessary for their function.
EXPERIMENTAL PROCEDURES
Plant material and growth conditions
Mutant lines of the Columbia-0 ecotype of Arabidopsis thaliana L.Heynh with T-DNA insertions were obtained from the SALK Institute(Alonso et al., 2003): rgtb1-1, SALK_015871; rgtb1-2, SALK_125416.The det3-1 mutant (At1g12840; Cabrera y Poch et al., 1993) waskindly provided by K. Schumacher (University of Heidelberg) andM. Campbell (University of Toronto). The location of each T-DNAinsertion within the RGTB1 gene (At5g12210) was verified bysequencing from each end of the insert (Appendix S1).
Arabidopsis seeds were surface-sterilized, stratified at 4�C for3–5 days and planted on vertical agar plates with growth media[half-strength MS salts, 2% (w/v) sucrose, vitamins and 1.6% agar]or soil. Light-grown plants were cultivated at 22�C under long-dayconditions (16 h of light per day). Plates with dark-grown plantswere wrapped into the aluminium foil after a light treatment for2–4 h to stimulate germination, and were cultivated at 22�C.
Preparation of plant extracts
For western blot analysis, either 100 mg of 7-day-old etiolatedplants or 100 mg of hypocotyls of these plants were ground inbuffer A [100 mM Tris, pH 7.8, 5 mM EGTA, 5 mM EDTA, 10 mM
beta-mercaptoethanol and 10% (w/v) glycerol] containing 1x plantprotease inhibitor cocktail and 1 mM phenylmethylsulfonyl fluoride(PMSF) (all from Sigma-Aldrich, http://www.sigmaaldrich.com) andcentrifuged for at 10 000 g for 5 min at 4�C. The supernatant waseither subjected to western analysis or further centrifuged at100 000 g for 1 h, at 4�C. The supernatant and pellet fractions wereprecipitated with trichloroacetic acid (TCA), the pellets were washedwith acetone and then dissolved in SDS/PAGE sample buffer.
For the geranylgeranylation assay, plant extracts were preparedaccording to the method described by Loraine et al. (1996). Freshplant material was frozen in liquid nitrogen and then ground on icein a mortar with acid-washed sand in buffer A (2 ml g)1 of cells).Lysates were centrifuged at 30 000 g for 30 min at 4�C, dialyzedagainst buffer B [50 mM Tris, pH 7.8, 10% (v/v) glycerol and 1 mM
DTT], aliquoted, frozen in liquid nitrogen and stored at )80�C untiluse.
The total protein concentration was measured using the Bio-RadDc Protein Assay according to the manufacturer’s protocol (Bio-RadLaboratories, http://www.bio-rad.com).
Preparation of recombinant fusion proteins
AtRAB-A2a was amplified from Arabidopsis pollen cDNA andcloned into the pET30a vector (Novagen, now part of Merck, http://www.merck-chemicals.com). Expression in Escherichia coli strainBL21 and purification of the recombinant N-terminal 6His-fusionprotein on Ni-NTA agarose (Qiagen, http://www.qiagen.com) wasperformed according to the Qiagen expression handbook.
Geranylgeranylation assay
The geranylgeranylation assay was performed as described byBenito-Moreno et al. (1994). Briefly, 200 lg of yeast or plant extractswere used in a total volume of 60 ll 1x geranylgeranylation buffer(50 mM phosphate buffer, pH 7.6, 10 mM MgCl2 and 5 mM DTT)with the addition of 0.5 lM [1-3H] all trans-geranylgeranyl pyro-phosphate (15–30 Ci mmol)1) (Amersham, now part of GE Health-care, http://www.gelifesciences.com) and 4 lg of recombinantAtRAB-A2a protein. The reaction mixture was then incubated at32�C for 40 min, and the reaction was stopped by the addition of12 ll of 6x loading buffer and boiled, and finally separated on 15%SDS-PAGE. Gels were boiled in 5% TCA solution for 5 min in thewater bath, and were briefly washed with distilled water. Acidictreatment was neutralized by 5 min of incubation with 100 mM TRISsolution at 25�C. Fluorographic intensification of the autoradio-graphic signal was achieved by incubation of the gel with 1 M
sodium salicylate. Gel was then dried on the gel drier and exposedto the X-Omat AR Kodak film or FOMA X-Ray film for 1 week at)80�C. The intensity of the bands occurring after the film develop-ment was measured by the IMAGEJ freeware program (http://rsb.info.nih.gov/ij/download.html).
Western blot analysis
An appropriate volume of the sample was loaded on 13.5% SDS-PAGE. Proteins were transferred on polyvinylidene fluoride (PVDF)membrane, blocked overnight at 4�C with 5% non-fat dry milk inTris-buffered saline (TBS), and incubated with the rabbit polyclonalanti-RAB-A2a antibody (diluted 1:1000 in 5% non-fat dry milk in TBSsupplemented with 0.5% Tween 20; kindly provided by I. Moore) for1 h. Secondary anti-rabbit alkaline-phosphatase-conjugated anti-body (1:5000; Sigma-Aldrich) was applied for 30 min, followed bycolour detection (Western Blue Stabilized Substrate; Promega,http://www.promega.com).
Phylogenetic analysis
Using the BLAST server and databases held at the National Centerfor Biotechnology Information (http://www.ncbi.nlm.nih.gov) andDOE Joint Genome Institute (http://genome.jgi-psf.org; http://www.phytozome.net), we searched for genes encoding b-subunitsof protein prenyl transferases in genomes of the following organ-isms: A. thaliana, Candida albicans, Caenorhabditis elegans,Drosophila melanogaster, Entomoeba histolytica, Giardia lamblia,Homo sapiens, Mus musculus, Neurospora crassa, O. sativa, Plas-modium falciparum, Physcomitrella patens, Populus trichocarpa,Saccharomyces cerevisiae, Schizosaccharomyces pombe,Selaginella moellendorffii, Sorghum bicolor, Volvox carteri andVitis vinifera.
624 Michal Hala et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

Multiple alignments of protein sequences were constructed usingthe CLUSTALX program with the default settings (Thompson et al.,1997) and manually adjusted. For phylogenetic analysis onlyreliably aligned regions were included: poorly conserved and gap-containing regions were removed (the alignment is available uponrequest). The phylogenetic tree was constructed using the maxi-mum-likelihood (ML) method, as implemented in the PHYML pro-gram (Guindon and Gascuel, 2003) with the JTT amino acidsubstitution matrix, discrete approximation to a C distribution (fourrate categories) and taking into account invariable sites (JTT+C+Imodel). Bacterial prenyl transferase from Mycobacterium tubercu-losis (MtPT) (Maurer-Stroh et al., 2003) was used as a root. Allmodel parameters were estimated from the data. ML bootstrapanalysis was performed on 100 bootstrap replicates (the JTT+C+Imodel with the same parameters as for the original data set), usingthe PHYML online execution server (http://atgc.lirmm.fr/phyml).
Microscopy
Styryl dye FM4-64 in a 15 lM concentration (diluted in 0.5x MSliquid medium) was vacuum infiltrated for 8 min into etiolated7-day-old plantlets. Quantification of endocytic compartments wasperformed as a measuring of the fluorescence inside hypocotylcells. The analyzed images were normalized by the signal of thecytoplasmic membranes, and the background was subtracted.Seedlings treated with 50 lM BFA were labeled with FM4-64 shortlybefore observation. Imaging of FM4-64 as well as secGFP was per-formed at different times using a confocal laser scanning micro-scope (LSM 510; Zeiss, http://www.zeiss.com) and a fluorescencemicroscope (BX51; Olympus, http://www.olympus-global.com)(Figure 4a,b).
Expression analysis
Seven-day-old seedlings grown in both continuous light and darkwere harvested from vertical agar plates to liquid nitrogen (200 mgof tissue per sample) and then ground. RNA was extracted in twoparallels for each set of conditions using an RNeasy� Mini kit(Qiagen) following the manufacturer’s instructions for plant mate-rial. Samples were sent on dry ice to NASC’s InternationalAffymetrix Service for standard processing on Affymetrix ATH1chips (http://affymetrix.arabidopsis.info). Values at a significancelevel of 0.05 were selected and average values for correspondingreplicas were calculated.
ACKNOWLEDGEMENTS
We thank I. Moore (University of Oxford) for help with the analysisof RAB prenylation, M. Potocky (IEB ASCR) for help with phyloge-netic analysis and for comments on the manuscript, andK. Schumacher (University of Heidelberg) and M. Campbell (Uni-versity of Toronto) for seeds of the det3-1 mutant. The work wassupported by the Grant Agency of the Czech Republic (204/06/P0457), the Ministry of Education, Youth and Sports of the CzechRepublic (MSMT LC06034) and part of VZ’s income is covered byMSM0021620858. Affymetrix data were supported by the GrantAgency of the Academy of Sciences of the Czech Republic (projectKJB600380802).
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Phylogenetic analysis of prenyl transferases b-subunits.Figure S2. Expression profiles of RAB-GGT subunits and AtREP.Figure S3. Semi-quantitative RT-PCR screening of AtRGTB1 andAtRGTB2 expression in the wild type and in rgtb1 mutants.
Figure S4. Biochemical characteristics of homozygous rgtb1mutants.Table S1. Expression levels of RAB, RAB-GDI, RAB-GGT and REPgenes in 7-day-old Arabidopsis seedlings (Col-0) in light or darkconditions.Appendix S1. Verification of T-DNA positions in rgtb1-1 and rgtb1-2insertion lines.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Alonso, J.M., Stepanova, A.N., Leisse, T.J. et al. (2003) Genome-wide inser-
tional mutagenesis of Arabidopsis thaliana. Science, 301, 653–657.
Anant, J.S., Desnoyers, L., Machius, M., Demeler, B., Hansen, J.C., Westover,
K.D., Deisenhofer, J. and Seabra, M.C. (1998) Mechanism of Rab geranyl-
geranylation: formation of the catalytic ternary complex. Biochemistry, 37,
12559–12568.
Baron, R.A. and Seabra, M.C. (2008) Rab geranylgeranylation occurs prefer-
entially via the pre-formed REP-RGGT complex and is regulated by gera-
nylgeranyl pyrophosphate. Biochem. J. 415, 67–75.
Bassham, D.C. and Blatt, M.R. (2008) SNAREs: cogs and coordinators in sig-
naling and development. Plant Physiol. 147, 1504–1515.
Batoko, H., Zheng, H.-Q., Hawes, C. and Moore, I. (2000) A Rab1 GTPase is
required for transport between the endoplasmic reticulum and Golgi
apparatus and for normal Golgi movement in plants. Plant Cell, 12, 2201–
2217.
Benito-Moreno, R.M., Miaczynska, M., Bauer, B.E., Schweyen, R.J. and Rag-
nini, A. (1994) Mrs6, the yeast homologue of the mammalian choroidere-
mia protein: immunological evidence for its function as the Ypt1p Rab
escort protein. Curr. Genet. 27, 23–25.
Biermann, B., Randall, S.K. and Crowell, D.N. (1996) Identification and
isoprenylation of plant GTP-binding proteins. Plant Mol. Biol. 31, 1021–
1028.
Bishop, G.J. and Koncz, C. (2002) Brassinosteroids and plant steroid hormone
signaling. Plant Cell, 14(Suppl.), S97–110.
Bolte, S., Schiene, K. and Dietz, K.J. (2000) Characterization of a small GTP-
binding protein of the rab 5 family in Mesembryanthemum crystallinum
with increased level of expression during early salt stress. Plant Mol. Biol.
42, 923–936.
Bracha-Drori, K., Shichrur, K., Lubetzky, T.C. and Yalovsky, S. (2008) Func-
tional analysis of Arabidopsis postprenylation CaaX processing enzymes
and their function in subcellular protein targeting. Plant Physiol. 148, 119–
31.
Brux, A., Liu, T.Y., Krebs, M., Stierhof, Y.D., Lohmann, J.U., Miersch, O.,
Wasternack, C. and Schumacher, K. (2008) Reduced V-ATPase activity in
the trans-Golgi network causes oxylipin-dependent hypocotyl growth
Inhibition in Arabidopsis. Plant Cell, 20, 1088–1100.
Cabrera y Poch, H.L., Peto, C.A. and Chory, J. (1993) A mutation in the
Arabidopsis DET3 gene uncouples photoregulated leaf development
from gene expression and chloroplast biogenesis. Plant J. 4, 671–
682.
Craig, S. (1992) The GUS reporter gene: application to light and transmission
electron microscopy. In GUS Protocols: Using the GUS Gene as a Reporter
of Gene Expression (Gallagher, S., ed.). San Diego, USA: Academic Press,
pp. 115–124.
Cremers, F.P., Molloy, C.M., van de Pol, D.J., van den Hurk, J.A., Bach, I.,
Geurts van Kessel, A.H. and Ropers, H.H. (1992) An autosomal homo-
logue of the choroideremia gene colocalizes with the Usher syndrome
type II locus on the distal part of chromosome 1q. Hum. Mol. Genet. 1,
71–75.
Cutler, S., Ghassemian, M., Bonetta, D., Cooney, S. and McCourt, P. (1996) A
protein farnesyl transferase involved in abscisic acid signal transduction in
Arabidopsis. Science, 273, 1239–1241.
Deneka, M., Neeft, M. and van der Sluis, P. (2003) Regulation of membrane
transport by rab GTPases. Crit. Rev. Biochem. Mol. Biol. 38, 121–142.
RAB prenylation and photomorphogenesis 625
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

Detter, J.C., Zhang, Q., Mules, E.H. et al. (2000) Rab geranylgeranyl transfer-
ase alpha mutation in the gunmetal mouse reduces Rab prenylation and
platelet synthesis. Proc. Natl Acad. Sci. USA, 97, 4144–4149.
Dettmer, J., Schubert, D., Calvo-Weimar, O., Stierhof, Y.D., Schmidt, R. and
Schumacher, K. (2005) Essential role of the V-ATPase in male gametophyte
development. Plant J. 41, 117–124.
Dettmer, J., Hong-Hermesdorf, A., Stierhof, Y.D. and Schumacher, K. (2006)
Vacuolar H+-ATPase activity is required for endocytic and secretory traf-
ficking in Arabidopsis. Plant Cell, 18, 715–730.
Dhonukshe, P., Tanaka, H., Goh, T. et al. (2008) Generation of cell polarity in
plants links endocytosis, auxin distribution and cell fate decisions. Nature,
456, 962–976.
Friml, J., Benfey, P., Benkova, E. et al. (2006) Apical-basal polarity: why plant
cells don’t stand on their heads. Trends Plant Sci. 11, 12–14.
Fukuda, M. (2008) Regulation of secretory vesicle traffic by Rab small GTP-
ases. Cell. Mol. Life Sci. 65, 2801–2813.
Geldner, N., Anders, N., Wolters, H., Keicher, J., Kornberger, W., Muller, P.,
Delbarre, A., Ueda, T., Nakano, A. and Jurgens, G. (2003) The Arabidopsis
GNOM ARF-GEF mediates endosomal recycling, auxin transport, and
auxin-dependent plant growth. Cell, 112, 219–230.
Geldner, N., Hyman, D.L., Wang, X., Schumacher, K. and Chory, J. (2007)
Endosomal signaling of plant steroid receptor kinase BRI1. Genes Dev. 21,
1598–1602.
Gerber, E., Hemmerlin, A., Hartmann, M. et al. (2009) The plastidial 2-C-me-
thyl-D-erythritol 4-phosphate pathway provides the isoprenyl moiety for
protein geranylgeranylation in tobacco BY-2 cells. Plant Cell, 21, 285–300.
Guindon, S. and Gascuel, O. (2003) A simple, fast, and accurate algorithm to
estimate large phylogenies by maximum likelihood. Syst. Biol. 52, 696–704.
Gutjahr, C., Riemann, M., Muller, A., Duchting, P., Weiler, E.W. and Nick, P.
(2005) Cholodny-Went revisited: a role for jasmonate in gravitropism of rice
coleoptiles. Planta, 222, 575–585.
Hala, M., Elias, M. and Zarsky, V. (2005) A specific feature of the angiosperm
Rab escort protein (REP) and evolution of the REP/GDI superfamily. J. Mol.
Biol. 348, 1299–1313.
Hedden, P. and Phillips, A.L. (2000) Gibberellin metabolism: new insights
revealed by the genes. Trends Plant Sci. 5, 523–530.
Hou, Y., von Arnim, A.G. and Deng, X.W. (1993) A new class of Arabidopsis
constitutive photomorphogenic genes involved in regulating cotyledon
development. Plant Cell, 5, 329–339.
Johnson, C.D., Chary, S.N., Chernoff, E.A., Zeng, Q., Running, M.P. and
Crowell, D.N. (2005) Protein geranylgeranyltransferase I is involved in
specific aspects of abscisic acid and auxin signaling in Arabidopsis. Plant
Physiol. 139, 722–733.
Kang, J.G., Yun, J., Kim, D.H. et al. (2001) Light and brassinosteroid signals
are integrated via a dark-induced small G protein in etiolated seedling
growth. Cell, 105, 625–636.
Kato, T., Morita, M.T., Fukaki, H., Yamauchi, Y., Uehara, M., Niihama, M. and
Tasaka, M. (2002) SGR2, a phospholipase-like protein, and ZIG/SGR4, a
SNARE, are involved in the shoot gravitropism of Arabidopsis. Plant Cell,
14, 33–46.
Kim, B.C., Soh, M.S., Hong, S.H., Furuya, M. and Nam, H.G. (1998) Photom-
orphogenic development of the Arabidopsis shy2-1D mutation and its
interaction with phytochromes in darkness. Plant J. 15, 61–68.
Loraine, A.E., Yalovsky, S., Fabry, S. and Gruissem, W. (1996) Tomato Rab1A
homologs as molecular tools for studying Rab geranylgeranyl transferase
in plant cells. Plant Physiol. 110, 1337–1347.
Maurer-Stroh, S., Washietl, S. and Eisenhaber, F. (2003) Protein pren-
yltransferases. Genome Biol. 4, 212.
Merry, D.E., Janne, P.A., Landers, J.E., Lewis, R.A. and Nussbaum, R.L. (1992)
Isolation of a candidate gene for choroideremia. Proc. Natl. Acad. Sci. USA,
89, 2135–2139.
Nagano, Y., Okada, Y., Narita, H., Asaka, Y. and Sasaki, Y. (1995) Location of
light-repressible, small GTP-binding protein of the YPT/rab family in the
growing zone of etiolated pea stems. Proc. Natl. Acad. Sci. USA, 92, 6314–
6318.
Newman, L.J., Perazza, D.E., Juda, L. and Campbell, M.M. (2004) Involvement
of the R2R3-MYB, AtMYB61, in the ectopic lignification and dark-photom-
orphogenic components of the det3 mutant phenotype. Plant J. 37, 239–250.
Novick, P., Medkova, M., Dong, G., Hutagalung, A., Reinisch, K. and
Grosshans, B. (2006) Interactions between Rabs, tethers, SNAREs and their
regulators in exocytosis. Biochem. Soc. Trans. 34, 683–686.
Pereira-Leal, J.B. and Seabra, M.C. (2001) Evolution of the Rab family of small
GTP-binding proteins. J. Mol. Biol. 313, 889–901.
Pylypenko, O., Rak, A., Reents, R. et al. (2003) Structure of Rab escort
protein-1 in complex with Rab geranylgeranyltransferase. Mol. Cell, 11,
483–494.
Rak, A., Pylypenko, O., Niculae, A., Pyatkov, K., Goody, R.S. and Alexan-
drov, K. (2004) Structure of the Rab7:REP-1 complex: insights into the
mechanism of Rab prenylation and choroideremia disease. Cell, 117,
749–760.
Randall, S.K., Marshall, M.S. and Crowell, D.N. (1993) Protein isoprenylation
in suspension-cultured tobacco cells. Plant Cell, 5, 433–442.
Robert, S., Chary, S.N., Drakakaki, G., Li, S., Yang, Z., Raikhel, N.V. and Hicks,
G.R. (2008) Endosidin1 defines a compartment involved in endocytosis of
the brassinosteroids receptor BRI1 and the auxin transporters PIN2 and
AUX1. Proc. Natl. Acad. Sci. USA, 105, 8464–8469.
Running, M.P., Lavy, M., Sternberg, H., Galichet, A., Gruissem, W., Hake, S.,
Ori, N. and Yalovsky, S. (2004) Enlarged meristems and delayed growth in
plp mutants result from lack of CaaX prenyltransferases. Proc. Natl Acad.
Sci. USA, 101, 7815–7820.
Rutherford, S. and Moore, I. (2002) The Arabidopsis Rab GTPase family: an-
other enigma variation. Curr. Opin. Plant Biol. 5, 518–528.
Sanford, J.C., Foster, L., Kapadia, Z. and Wessling-Resnick, M. (1995) Analysis
of the stoichiometry of Rab protein prenylation. Anal. Biochem. 224, 547–
556.
Schumacher, K. and Chory, J. (2000) Brassinosteroid signal transduction: still
casting the actors. Curr. Opin. Plant Biol. 3, 79–84.
Schumacher, K., Vafeados, D., McCarthy, M., Sze, H., Wilkins, T. and Chory,
J. (1999) The Arabidopsis det3 mutant reveals a central role for the
vacuolar H(+)-ATPase in plant growth and development. Genes Dev. 13,
3259–3270.
Seabra, M.C. (1996) Nucleotide dependence of Rab geranylgeranylation.
J. Biol. Chem. 271, 14398–14404.
Seabra, M.C., Brown, M.S., Slaughter, C.A., Sudhof, T.C. and Goldstein, J.L.
(1992) Purification of component A of Rab geranylgeranyl transferase:
possible identity with the choroideremia gene product. Cell, 70, 1049–1057.
Seabra, M.C., Ho, Y.K. and Anant, J.S. (1995) Deficient geranylgeranylation of
Ram/Rab27 in choroideremia. J. Biol. Chem. 270, 24420–24427.
Seabra, M.C., Mules, E.H. and Hume, A.N. (2002) Rab GTPases, intracellular
traffic and disease. Trends Mol. Med. 8, 23–30.
Shen, F. and Seabra, M.C. (1996) Mechanism of digeranylgeranylation of Rab
proteins. J. Biol. Chem. 271, 3692–3698.
Silady, R.A., Ehrhardt, D.W., Jackson, K., Faulkner, C., Oparka, K. and Som-
erville, C.R. (2008) The GRV2/RME-8 protein of Arabidopsis functions in the
late endocytic pathway and is required for vacuolar membrane flow. Plant
J. 53, 29–41.
Strompen, G., Dettmer, J., Stierhof, Y.D., Schumacher, K., Jurgens, G. and
Mayer, U. (2005) Arabidopsis vacuolar H-ATPase subunit E isoform 1 is
required for Golgi organization and vacuole function in embryogenesis.
Plant J. 41, 125–32.
Szekeres, M., Nemeth, K., Koncz-Kalman, Z., Mathur, J., Kauschmann, A.,
Altmann, T., Redei, G.P., Nagy, F., Schell, J. and Koncz, C. (1996) Brassi-
nosteroids rescue the deficiency of CYP90, a cytochrome P450, controlling
cell elongation and de-etiolation in Arabidopsis. Cell, 19, 171–182.
Tanaka, D., Kameyama, K., Okamoto, H. and Doi, M. (2008) Caenorhabditis
elegans Rab escort protein (REP-1) differently regulates each Rab protein
function and localization in a tissue-dependent manner. Genes to Cells, 13,
1141–1157.
Thoma, N.H., Niculae, A., Goody, R.S. and Alexandrov, K. (2001a) Double
prenylation by RabGGTase can proceed without dissociation of the mono-
prenylated intermediate. J. Biol. Chem. 276, 48631–48636.
Thoma, N.H., Iakovenko, A., Kalinin, A., Waldmann, H., Goody, R.S. and
Alexandrov, K. (2001b) Allosteric regulation of substrate binding and
product release in geranylgeranyltransferase type II. Biochemistry, 40, 268–
274.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F. and Higgins, D.G.
(1997) The CLUSTAL_X windows interface: flexible strategies for multiple
sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25,
4876–4882.
Ueda, T., Yamaguchi, M., Uchimiya, H. and Nakano, A. (2001) Ara6, a plant-
unique novel type Rab GTPase, functions in the endocytic pathway of
Arabidopsis thaliana. EMBO J. 20, 4730–4741.
626 Michal Hala et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627

Ulmasov, T., Murfett, J., Hagen, G. and Guilfoyle, T.J. (1997) Aux/IAA
proteins repress expression of reporter genes containing natural and
highly active synthetic auxin response elements. Plant Cell, 9, 1963–
1971.
Wilson, R.N., Heckman, J.W. and Somerville, C.R. (1992) Gibberellin Is
Required for Flowering in Arabidopsis thaliana under Short Days. Plant
Physiol. 100, 403–408.
Wilson, A.L., Erdman, R.A., Castellano, F. and Maltese, W.A. (1998) Prenyla-
tion of Rab8 GTPase by type I and type II geranylgeranyl transferases.
Biochem. J. 333, 497–504.
Wojtas, M., Swiezewski, S., Sarnowski, T.J., Plochocka, D., Chelstowska, A.,
Tolmachova, T. and Swiezewska, E. (2007) Cloning and characterization of
Rab Escort Protein (REP) from Arabidopsis thaliana. Cell Biol. Int. 31, 246–
251.
Woollard, A.A. and Moore, I. (2008) The functions of Rab GTPases in plant
membrane traffic. Curr. Opin. Plant Biol. 11, 610–619.
Yalovsky, S., Loraine, A.E. and Gruissem, W. (1996) Specific prenylation
of tomato Rab proteins by geranylgeranyl type-II transferase requires
a conserved cysteine–cysteine motif. Plant Physiol. 110, 1349–
1359.
Yalovsky, S., Kulukian, A., Rodriguez-Concepcion, M., Young, C.A. and
Gruissem, W. (2000) Functional requirement of plant farnesyltransferase
during development in Arabidopsis. Plant Cell, 12, 1267–1278.
Yano, D., Sato, M., Saito, C., Sato, M.H., Morita, M.T. and Tasaka, M. (2003) A
SNARE complex containing SGR3/AtVAM3 and ZIG/VTI11 in gravity-
sensing cells is important for Arabidopsis shoot gravitropism. Proc. Natl
Acad. Sci. USA, 100, 8589–8594.
Yoshida, K., Nagano, Y., Murai, N. and Sasaki, Y. (1993) Phytochrome-
regulated expression of the genes encoding the small GTP-binding
proteins in peas. Proc. Natl Acad. Sci. USA, 90, 6636–6640.
Zhang, H., Seabra, M.C. and Deisenhofer, J. (2000) Crystal structure of
Rab geranylgeranyltransferase at 2.0 A resolution. Structure Fold Des. 8,
241–251.
Zhang, Q., Zhen, L., Li, W., Novak, E.K., Collinson, L.M., Jang, E.K., Haslam,
R.J., Elliott, R.W. and Swank, R.T. (2002) Cell-specific abnormal prenylation
of Rab proteins in platelets and melanocytes of the gunmetal mouse. Br. J.
Haematol. 117, 414–423.
Zimmermann, P., Hirsch-Hoffmann, M., Hennig, L. and Gruissem, W. (2004)
GENEVESTIGATOR. Arabidopsis microarray database and analysis
toolbox. Plant Physiol. 136, 2621–2632.
Accession numbers: Sequence data from this article can be found in the EMBL/GenBank data libraries under the following accession numbers(AGI codes): AtRGTB1, At5g12210; AtRGTB2, At3g12070; AtRGTA1, At4g24490; AtRGTA2, At5g41820. Seed stocks from the SALK Institute:rgtb1-1, SALK_015871; rgtb1-2, SALK_125416.
RAB prenylation and photomorphogenesis 627
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 62, 615–627