abiotic stresses and induced bvocs
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Special Issue: Induced biogenic volatile organic compounds from plants
Abiotic stresses and induced BVOCsFrancesco Loreto1 and Jorg-Peter Schnitzler2
1 Consiglio Nazionale delle Ricerche (CNR), Istituto per la Protezione delle Piante (IPP), Via Madonna del Piano 10, 50019 Sesto
Fiorentino, Firenze, Italy2 Karlsruhe Institute of Technology (KIT), Institute of Meteorology and Climate Research (IMK-IFU), Kreuzeckbahnstraße 19,
82467 Garmisch-Partenkirchen, Germany
Plants produce a wide spectrum of biogenic volatileorganic compounds (BVOCs) in various tissues aboveand below ground to communicate with other plantsand organisms. However, BVOCs also have various func-tions in biotic and abiotic stresses. For example abioticstresses enhance BVOCs emission rates and patterns,altering the communication with other organisms andthe photochemical cycles. Recent new insights on bio-synthesis and eco-physiological control of constitutiveor induced BVOCs have led to formulation of hypotheseson their functions which are presented in this review.Specifically, oxidative and thermal stresses are relievedin the presence of volatile terpenes. Terpenes, C6 com-pounds, and methyl salicylate are thought to promotedirect and indirect defence by modulating the signallingthat biochemically activate defence pathways.
The emission of BVOCs: few biochemical pathways butmany compounds emittedBiosynthesis of the main BVOCs
Plants produce a wide spectrum of BVOCs in varioustissues above and below ground. Most BVOCs are largelylipophilic and have enough vapour pressure to be releasedinto the atmosphere in significant amounts. The availabil-ity of new methods of head-space sampling (such as solidphase micro-extraction) in combination with gas chroma-tography–mass spectroscopy and new techniques for on-line analysis (proton transfer reaction-mass spectrometry)[1] has led, in the last 15 years, to a significant expansion ofour knowledge on the occurrence and temporal and spatialdistribution of BVOCs emissions. At present, about 1700substances have been found to be emitted from plants [2].Nearly all organs from vegetative parts, as well as flowers[3] and roots [4] emit these compounds. Many BVOCs areemitted constitutively and the emissions can be observedthroughout the life cycle of the plant or, more often, atspecific developmental stages (e.g. leaf and needle matu-ration, senescence, flowering, and fruit ripening). Theemission is biosynthetically controlled by abiotic factorssuch as light and/or temperature, atmospheric CO2 con-centration, or nutrition. Other BVOCs are induced afterwounding and herbivore feeding or after environmentalstresses. Stresses may induce change of constitutiveBVOCs, either stimulating or quenching the emissions(e.g. [5]) or may induce de novo synthesis and emission
of BVOCs. Induced emissions may occur in a systemic way,i.e. away from the site of damage [6].
The biosynthesis of most BVOCs can be assigned to thefollowing three major pathways: terpenes (= isoprenoids),oxylipins, and shikimate and benzoic acid [7,8]. Low mol-ecular weight, C1 and C2, compounds, such as methanol,ethanol, formaldehyde, and acetaldehyde can be synthes-ized via other biosynthetic routes [9]. Two other BVOCs aremethane and ethylene. Emission of non-microbialmethaneby vegetation has been discovered recently [10] andresearch on this important topic is still in its infancy.Recent isotope labelling studies provided evidence thatmethane can be generated from methoxyl groups derivingfrom breakdown of plant pectins [11].
In this review of the impact of abiotic factors on theinduction of BVOCs emissions, we will mostly concentrateon volatile terpenes which are the most important com-pounds for plant biology [12] and atmospheric chemistry[13] because of their role in plant protection (e.g. in theprotection of photosynthesis against thermal and oxidativestresses, and in direct and indirect defence against herbi-vores), as well as in the chemical properties of the atmos-phere (e.g. entering the cycle of photochemical production/destruction of ozone, aerosols, and particles). The emissionof volatile terpenes is estimated to account for more thanhalf of the total emission of BVOCs [14] and is constitu-tively ten times higher than other emissions, as heavyemitters can release isoprene at rates of 50–100 nmolm�2 s�1, representing up to 2–5% of the photosyntheticnet carbon uptake in tree species. Volatile terpene emis-sions are far more sustained than the emission of otherinduced volatiles, for which emissions are transient bynature and limited to specific periods after stress, depend-ing on the damage experienced and on the activation of thebiosynthetic pathways producing the volatiles.
Biosynthesis of volatile terpenesTerpenes are constitutively formed in some plant familiesthat store them in massive amounts in internal or externalstructures (e.g. the resin ducts of conifers or the glandularcells of Lamiaceae leaves). They may also be induced inresponse to wounding or herbivory attack. The emission ofterpenes from storage structures is generally uncoupledfrom photosynthesis as it may occur, for example, at night[15].
Direct emission of isoprene or monoterpenes from themesophyll is common in some tree species (Figure 1), in
Review
Corresponding author: Loreto, F. ([email protected]).
154 1360-1385/$ – see front matter � 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.tplants.2009.12.006 Available online 4 February 2010

Author's personal copy
particular from the Fagaceae and Salicaceae (oaks andpoplars) [16,17]. These emissions are light-dependent[18,19] and are closely linked to the availability of photo-synthetic intermediates [19,20]. The investigation ofvolatile terpene biosynthesis is a very active area of plantresearch, especially since the discovery and complete elu-cidation of the methylerythritol (MEP) pathway, respon-sible for the formation of the basic C5 units isopentenyldiphosphate (IDP) and dimethylallyl diphosphate(DMADP) in many bacteria and in the plastids of allorganisms from phototrophic phyla [21].
In plants, IDP and DMADP are formed via two alterna-tive pathways: (i) in the cytosolic mevalonic acid (MVA)pathway from acetyl-CoA, and (ii) in the plastidic MEPpathway from pyruvate and glyceraldehyde-3-phosphate[22]. Generally, the MEP pathway provides IDP andDMADP for hemiterpene and monoterpene biosynthesis,while the MVA pathway provides the C5 units for sesqui-terpene formation. However, a very recent report hasrevealed that the MEP pathway also contributes to sesqui-terpene formation [23]. In addition, some metabolic
crosstalk between both biosynthetic routes is possible[24] particularly (via IDP) in the direction from chloro-plasts to the cytosol [25].
Prenyl transferases catalyze the condensation of IDPand DMADP to form geranyl diphosphate (GDP) andfarnesyl diphosphate (FDP) [26]. Finally, the conversionof DMADP, GDP, and FDP, into volatile hemi-, mono-, andsesquiterpenes, respectively, is catalyzed by terpenesynthases (TPS), a large family of enzymes, encoded byclosely related genes [27]. Isolation and characterization ofprenyl transferase [28] and terpene synthase genes [29] isnow giving new insights into the evolutionary origin[30,31] and genetic and biochemical regulation of terpenebiosynthesis.
Isoprene and 2-methyl-3-buten-2-ol (MBO), a hemiter-pene common only in American western pines, [32] arebiochemically synthesized in chloroplasts by isoprene [33]and MBO synthase [34] from DMADP. Due to the import-ance of isoprene for atmospheric processes and plant func-tions, isoprene synthase (ISPS) became one of the beststudied TPS [35]. A positive correlation between ISPSactivity and basal standard emission capacity was foundin different isoprene-emitting species [36,37]. Up to now,five ISPS genes from different poplar (Populus) species orpoplarhybridshavebeendescribed [35]. The only ISPSgenecloned so far fromanother genus,Puerariamontana (kudzu)[38], shows only 52% identity with the poplar proteinsequences,although thestructureofpoplarandkudzugenesare similar (six intronsandsevenexons) [38].All ISPSgenesbelong to the subgroup b of the class 1 plant TPS-familywhich includes monomeric mono-, sesqui-, and diterpenesynthases, grouped insix subgroups (Tspa–TspfTpsa–Tpsf?)[27]. All known ISPS enzymes have a 10–100-fold higherMichaelis constant (kM) for its substrate DMADP (in themillimolar range) thanmonoterpene synthases forGDP [39]or prenyltransferases for DMADP [40]. The low kM of pre-nyltransferases may control the metabolic flux within theMEP pathway because downstream reactions leading tomonoterpene and non-volatile terpene biosynthesis arefavoured over isoprene biosynthesis. Based on this finding,it was suggested that isoprene emission occurs only whenplants’ need for ‘essential’, higher terpenes (hormones, e.g.ABA and gibberellins; tocopherol; phytosterols; and photo-synthetic pigments) are satisfied [41].
Interest in genetic regulation and biochemical proper-ties of TPS other than ISPS mostly focused on those TPSinvolved in the formation in storage structures (e.g. ingymnosperms and Lamiaceae) of those terpenes withdefensive or attractive functions [42] (see Dicke and Bald-win, this issue).
In light-dependent monoterpene emitters, the infor-mation about TPS regulation is scant. Differences inmono-terpene emission pattern of chemotypes and oak hybrids[43], the seasonal development of monoterpene emission[44,45], and the dependence of monoterpene emission onatmospheric CO2 [46] all appear to be controlled by TPSactivities. However, our knowledge about the underlyinggenes is scarce; in oak, only two mono-TPS genes, a b-myrcene synthase [47] and a multiproduct a-pinenesynthase [48] have been isolated and functionally charac-terized up to now.
Figure 1. Origin of volatile terpene emissions from different leaf types. In
deciduous leaves of many tree species (e.g. oaks and poplars) with no specific
storage structures for terpenes, isoprene and monoterpene, BVOCs emissions
originate from mesophyll cells in a light- and temperature-dependent manner. In
conifer needles (e.g. Picea abies - Norway spruce) light-dependent terpene
emission stems from photosynthetic tissue and is superposed by a temperature-
dependent volatilization of terpenes from resin ducts. Lamiaceae (e.g. Ocimum
basilicum basil) leaves release temperature-dependent volatile terpenes from
external glandular cells. Confocal laser scanning microscopic images were taken
from cross-sections of Quercus robur (top), Picea abies (middle) and leaf surface of
Ocimum basilicum (bottom).
Review Trends in Plant Science Vol.15 No.3
155

Author's personal copy
The commonly used model plant Arabidopsis thaliana(Arabidopsis) is a good example of how modern technol-ogies can improve knowledge of BVOCs biosynthesis andfunctions. In the past classified as ‘non-emitting’ species,analysis of the Arabidopsis genome revealed the existenceof over 30 putative genes belonging to themultigene familyof TPS [49,50]. Most of these genes are almost exclusivelyexpressed in flowers [50,51], but low constitutive terpeneemissions from leaves and siliques [50,52] and even emis-sions from roots (namely 1,8-cineole, [4]) could be detected.Isoprene synthase overexpression in Arabidopsis allowedverification of a ‘thermo-protective’ activity of isoprene[53,54], see below, and indicated an ecological functionof this hemiterpene in plant–insect interactions as a repel-ling cue [55]. The expression of b-caryophyllene was alsosuccessfully engineered in this plant (J. Gershenzon,personal communication) and may help assess functionsand fate of sesquiterpenes. Sesquiterpenes are not emittedin large amounts constitutively, but their biosynthesis canbe induced by biotic stresses, and may be important as anindirect defence mechanism (see Dicke and Baldwin, thisissue). Even at low concentrations sesquiterpenes are alsoimportant as nucleation factors, eventually leading toparticle formation in the atmosphere [56].
Poplar is another seminal model system to study regu-lation and function of plant volatiles. Besides ISPS [35],only one TPS gene [(�)-germacrene-D synthase] [57] ischaracterized. However, full length cDNAs [58], cDNAmicroarrays [59,60], and a largely sequenced genome ofPopulus trichocarpa [61] are now available, and these toolswill certainly allow for quick progress in the understandingof BVOCs formation in a near future.
Biosynthesis of C1 and C2 oxygenated compoundsVolatile terpenes are certainly the family of compoundsthat contributes the majority of BVOCs [14]. However,short-chained oxygenated compounds (especiallymethanoland acetaldehyde) are important components of constitu-tive and induced emissions of many plants [5] with a largepresence globally [62].
Methanol is predominantly emitted because of degra-dation and formation of cell wall pectins, e.g. (i) during cellexpansion in all types of plant tissue and seeds, and (ii)during leaf abscission, senescence, and seed maturation(Figure 2). The formation of methanol is catalyzed bypectin methylesterases (PME) which, among the otherfunctions, demethoxylates pectin [63,64]. To a minorextent, methanol emissions originate from protein meth-yltransferase and protein repair reactions [65], or fromtetrahydrofolate metabolism [66]. Emission of methanolcan be induced by mechanical wounding [67] or herbivorefeeding, due to an upregulation of PME expression. Thisfinding has stimulated a discussion about whether metha-nol might act as a signal in plant–plant communication[68].
The metabolic origin of acetaldehyde emitted by foresttrees is still a matter of debate [69,70]. This compoundseems to be predominantly induced by stresses. It is knownthat acetaldehyde emission correlates with root flooding[71,72] and with xylem sap ethanol concentrations [73,74].Ethanol formed under anoxic conditions in roots is trans-
ported to leaves by the transpiration stream, where it isoxidized to acetaldehyde by alcohol dehydrogenase (ADH).However, only a small portion of acetaldehyde is emittedwhile the bulk is furthermetabolized by aldehyde dehydro-genase (ALDH) to acetate and acetyl-CoA.
In some tree species, strong transient acetaldehydebursts during light–dark transitions have been reported[70,72,75]. These acetaldehyde bursts are thought to be theresult of a ‘pyruvate overflow mechanism’ [75]. In theproposed mechanism pyruvate decarboxylase (PDC) actsas a metabolic regulator converting excess cytosolic pyr-uvate into acetaldehyde, which is subsequently oxidized toacetate. Such an excess of cytosolic pyruvate may be theresult of transiently decreased transport rates of pyruvateequivalents [i.e. phosphoenolpyruvate (PEP)] into orga-nelles, or reduced pyruvate utilization in leaf cells immedi-ately after darkening [75]. However, emission ofacetaldehyde may also derive from cleavage of moietiesof C6 aldehydes, which emissions are also transientlystimulated during light–dark transitions [70]. In this case,acetaldehyde emission is independent of cytosolic pyruvateand is part of the leaf response to wounding that alsoincludes biosynthesis of C6 compounds (see below).
Biosynthesis of C6 aldehydes and alcoholsWounding induces the release of ‘green leaf volatiles’(GLV) as can be easily sensed in the odour of fresh hay.In most wounded plants GLV are C6 aldehydes, C6 alco-hols, and their derivatives, often collectively called C6- orLOX-products [76]. Physiologically these compounds have
Figure 2. Simplified scheme of the subcellular origin and biosynthesis of volatile
organic compounds upon abiotic stress. Abbreviations: CH4, methane; DMADP,
dimethylallyl diphosphate; DXS, 1-deoxy-D-xylulose 5-phosphate synthase (EC
4.1.3.37); FDP, farnesyl diphosphate; GDP, geranyl diphosphate; HMGR, 3-hydroxy-
3-methylglutaryl-CoA reductase (EC 1.1.1.34); 13-HPOT, 13S-hydroperoxy-
9(Z),11(E),15(Z)-octadecatrienoic acid; IDP, isopentenyl diphosphate; ISPS,
isoprene synthase (EC 4.2.3.27); a-LeA, a-linolenic acid; 13-LOX, 13-lipoxygenase
(EC 1.13.11.12); MeOH, methanol; MTS, monoterpene synthase (e.g. myrcene EC
4.2.3.14); PDC, pyruvate decarboxylase (EC 4.1.1.1); PEP, phosphoenolpyruvate;
PME, pectine methylesterase (EC 3.1.1.11); PYR, pyruvate; STS, sesquiterpene
synthase (e.g. epi-aristolochene EC 4.2.3.9); 13-HPL, 13-hydroxyperoxide lyase (not
listed in enzyme classification); TP, triose phosphate. The broken arrow indicates a
proposed, yet unidentified acetaldehyde emission path from chloroplasts.
Review Trends in Plant Science Vol.15 No.3
156

Author's personal copy
antibiotic properties inhibiting the invasion of damagedtissue [77], and can serve a signalling function withinplants to induce or prime defence [78].
GLV are derived from oxidized polyenoic fatty acids(PUFA), collectively called oxylipins. The initial formationis catalyzed by lipoxygenases (LOX), a large gene family ofnon-haeme iron containing fatty acid dioxygenases [8](Figure 2). LOXs (13-LOX, classified with respect to theirpositional specificity of oxidation at carbon-13 of the fattyacid hydrocarbon backbone) initiate the octadecanoicpathway by adding O2 stereospecifically to unsaturatedfatty acids [e.g. linoleic acid (18:2) and a-linolenic acid(18:3)] generating 13-(S)-hydroperoxides. Subsequently,13-(S)-hydroperoxide lyase (HPL) catalyzes the cleavagebetween C12 and C13 releasing n-hexanal (from linoleicacid) and (Z)-3-hexenal (from a-linolenic acid) which aretheparent compounds for all other aldehydes andalcohols,and enzymatically acetylated compounds such as hexylacetate and (Z)-3-hexenyl acetate [76]. Analysis of C6compounds is complicated by their chemical instabilityand the transient nature of their formation after wound-ing. Thanks to modern on-line techniques like protontransfer reaction mass spectrometry (PTR-MS) that avoidpre-concentration of the samples on adsorbent phases, therapid and transient emission of C6 upon various stimuli,such as physical damage [79], light–dark transitions, her-bivory, or senescence processes, can be easily monitored[70,79].
Plants and abiotic stresses: how stresses affect BVOCsbiosynthesis and emissionsAbiotic stresses affect primary and secondary metabolismin different ways. Stresses generally inhibit photosyn-thesis by reducing CO2 uptake and diffusion inside leavesto the site of fixation into carbohydrates or by altering thephotochemical or biochemical reactions of the photosyn-thetic cycle [80]. The impact of stresses on the secondarymetabolisms that produces BVOCs is more controversial,as some of the pathways may be elicited by stresses. In thecase of volatile terpenes, the uncoupling of photosynthesisand BVOCs emission is surprising, because of the well-known high requirement of photosynthetic carbon for ter-pene biosynthesis [81]. The stimulation of terpene pro-duction is made possible by the activation of sources ofcarbon which are alternatives to freshly fixed photo-synthates and not yet fully identified. Some labellingstudies have provided evidence that isoprene may alsobe formed from xylem-transported glucose and chloroplas-tic starch [82]. However, under stress conditions, other 13Clabelling studies point out that, because of starchdepletion, extra-chloroplastic sources of carbon may beactivated and feed carbon to volatile terpenes [83]. Stressesmay also induce damage that elicits or induces the syn-thesis of other volatiles with important consequences forplant protection, directly against the environmental con-straint [12] or indirectly, against possible, associatedbursts of pathogens and herbivores (see Dicke and Bald-win, this issue). The impact of the main environmentalfactors of stress on the biosynthesis and emission of vola-tiles will be reviewed in the following sections (see alsoFigure 3).
Temperature
BVOCs partition between the gas and liquid phase in theplants according to their Henry’s law constant (kH), whichis generally very high (e.g. kH �7500 Pa m3 mol�1 at 25 8Cfor isoprene) [84]. The equilibrium between gas and aqu-eous phases is indeed determined by the temperature andtherefore it is expected that more BVOCs enter the gasphase and are emitted at rising temperatures. This directeffect of temperature on BVOCs emission is, however,modulated by diffusion resistances encountered from thesites of synthesis inside the leaf to the atmosphere. BVOCsthat are stored in specialized structures (ducts or glands)reach very high concentrations and are tightly separatedby the surrounding cells by an impermeable cell layer,generally made by sub-cuticular cells [85]. This is becausehigh concentrations of BVOCs, which have probablyimportant ecological functions as deterrents of herbivoresand as anti-bacterial and anti-mycotic substances, could beauto-toxic for plant cells [86]. Temperature affects theevaporation and release of a minimal part of the pools ofBVOCs that leaks out the impermeable cell layer. Indeedplants with large storage pools are moderate emitters[15,17] unless the pools are opened up, e.g. by herbivory[87], strong winds, or forest fires [88]. High humidity mayalso lead to a swelling and to the consequent explosion ofthe structures containing the pools. In this case strongemissions of terpenesmay even be sensed, e.g. walking on apine forest at sunrise.
On the other hand, plants that do not store BVOCs intospecialized structures have small temporary pools in theleaf mesophyll that freely diffuse out of the leaf driven by aconcentration gradient. The only important constraint tothis diffusion is at the stomatal conductance to gas
Figure 3. Schematic overview of long-term and short-term regulation of terpene
emissions upon various abiotic stresses. Long-term adaptation of volatile terpene
emission (predisposition and/or priming) relates to differential gene expression
resulting in upregulation (green circles) and/or downregulation (red circles) of
specific enzymes and changed emission pattern and emission rates. Short-term
regulation reflects rapid changes in metabolic fluxes and enzyme kinetic properties
(e.g. temperature, pH and ion shifts, and redox/energy status) due to abiotic factors
like atmospheric CO2 concentration, light and temperature. Orientation of arrows
in the green and red circles indicate strength of upregulation and downregulation.
Abbreviations: h, hours; min, minutes; s, seconds.
Review Trends in Plant Science Vol.15 No.3
157

Author's personal copy
exchange. High temperatures often affect stomatal beha-viour, either per se or because this stress is generallyassociated with a drought stress. Stomatal opening undertransient heat stress is an important mechanism to dis-sipate latent heat through transpiration of water and touncouple the leaf temperature from air temperature. Onthe other hand, stomatal closure improves instantaneouswater use efficiency (the ratio between net CO2 assimila-tion and transpiration) and avoids excessive loss of waterdriven by increasing transpiration. Stomatal movementsdo not affect the steady-state diffusion rates of gases withhigh kH, such as isoprene and monoterpenes [84]. In thecase of stomatal closure, these compounds build up tran-siently significant partial pressures inside leaves thatcompensate for the increasing resistance at stomatal level[89,90]. However, diffusion of gases that are mainly parti-tioned into liquid phase, such as oxygenated VOCs (metha-nol, C6 aldehydes, and alcohols) might be stronglyrestrained by stomatal closure [87].
Temperature has a strong and immediate influence onthe activity of the enzymes that catalyze the synthesis ofmany BVOCs. Emissions of volatile terpenes typicallyhave a Q10 = 2–4, at temperatures variable between 20and 40 8C [91]. Thus the main effect of rising temperatureis a direct increment of the terpenes formed throughenzymatic reactions. However, when a heat stress occurswith temperature above the optimal enzyme temperature(around 40–45 8C for enzymes in the MEP pathway andmost TPS), then a very rapid inhibition of terpene emissionis observed [5]. This is probably due to downregulation orimpairment of primary metabolism and to the con-sequently insufficient supply of photosyntheticmetabolitesinto the MEP pathway [92,93]. However, there are cases inwhich the emission is not rapidly re-established upon heatstress removal, and in these cases a heat-induced dena-turation of the TPS is also likely to occur [5].
Interestingly, when the heat-induced inhibition ofvolatile terpenes occurs, then the emission of other BVOCsis highly enhanced [5]. This is particularly evident in thecase of methanol and C6 compounds, pinpointing that theinhibition of volatile terpene emission coincides with theoccurrence of damage to cell walls and membranes,respectively [5]. The emission of C6 compounds is sus-tained for the whole period of heat stress andmay continuefor a long time after temperatures go back to physiologicallevels [5]. Significant fluxes of methyl-butenol, ethanol,and acetaldehyde were found in North American conifersexposed to high temperature [94], and methanol andacetone emissions were also observed from bare agricul-tural soils following a heat wave episode [95]. Thus soilmicroorganisms may also contribute to the temperature-dependent emission of oxygenated BVOCs.
Finally, temperature seems to have an important role indetermining emissions of methyl salicylate (MeSA), a sig-nallingmolecule whose induction is frequent in response tobiotic stresses [74,96]. One study [97] reported induction ofMeSA only after spraying plants with jasmonic acid,another important signalling molecule activating theinduction of genes involved in hypersensitive responses,both after biotic and abiotic stresses. In another study asignificant induction of MeSA, with fluxes comparable to
those of monoterpenes was observed in plants exposed tonight chilling temperatures [98] and the induction of thiscompound correlated to the difference in temperaturebetween day and night that was experienced by walnut(Juglans californica � Juglans regia) plants.
Drought and salt
The impact of drought and salt on BVOCs emission issurprising. These abiotic stresses directly affect stomatalconductance and produce diffusive and biochemical limita-tions of photosynthesis [79]. Both the reduction of photo-synthesis and the stomatal closure are expected tonegatively impact on BVOCs emission by altering thecarbon supply into the MEP pathway and by increasingresistance to their emission.
In fact, the emission of volatile terpenes is resistant tothese stresses and is often elicited by stress occurrence.The original observation that isoprene is not reduced byincreasing drought stress until the stress becomes heavyand almost completely inhibits photosynthesis [99] hasbeen repeatedly made, also with experiments in whichthe drought stress has been controlled differently, e.g.[90,100]. The sustained emission of isoprene to salt anddrought [101,102] is made possible by the induction ofcarbon sources alternative to photosynthesis, possiblyrelated to respiration [103] or starch breakdown [82].Labelling experiments with 13CO2 [35,83] demonstrateclearly the preference of ‘old’ unlabeled (12C) carbonskeletons over recently fixed, 13C enriched photosyntheticintermediates when photosynthesis (which is heavilyreduced by drought and salt stress) [83,101] and isoprenebiosynthesis become uncoupled. This may also explain whyin some instances re-establishment of photosyntheticmetabolism, by re-watering plants, results in a burst ofisoprene emission [99]. However, it has been shown [83]that the alternative carbon sources rapidly cease to feedcarbon into the MEP pathway once the water status ofplants is reintegrated. A recent observation indicates thatin plants exposed to severe drought stress and in thoserecovering from drought stress the temperature depen-dency of isoprene emission is lost for at least several weeks,if not permanently [100]. The mechanism behind thisobservation is still unknown, but it is likely that the ISPSprotein is slowly or incompletely re-synthesized after thestress [100]. Climate-change impact on isoprene emissionhas been mainly attributed to positive long-term (enzy-matic) and short-term (substrate) feedback of risingtemperature [104,105], implying that future emissions ofisoprene will also increase [106]. However, as drought isoften associated with heat waves and summer climate, thefinding that drought suppresses temperature-dependencyof isoprene emission may have consequences for trees’thermal and ozone tolerance in regions (e.g. the Mediter-ranean basin) which will be plagued by climate-change-induced droughts associated with rising temperatures.
Laboratory measurements of the impact of drought onmonoterpene emissions are missing, but in field exper-iments monoterpene emissions are inhibited by droughtstress [46,107,108]. Interestingly, the inhibition is particu-larly evident only when drought stress is severe (e.g. thewater potential is less than –2 MPa) again suggesting that
Review Trends in Plant Science Vol.15 No.3
158

Author's personal copy
biosynthesis of volatile terpenes is resistant to milddrought stress [109]. It is unclear whether alternativecarbon sources can be used to generate monoterpenesunder these conditions. As for temperature, heavilydrought-stressed oak plants lose the capacity to respondto other environmental factors that are known to modulatethe emission, such as CO2 (see below) [46].
Drought seems to have a different effect on pools ofmonoterpenes stored in specialized structures or non-stored in the mesophyll. In conifers, if the drought eventoccurs in winter, when the biosynthesis of terpenes isrestrained by temperature, the pools appear to be nega-tively affected. However, summer droughts can furtherenrich the monoterpene pools. In non-storing species,summer drought depletes the pools of terpenes in themesophyll which are under direct control of photosyntheticcarbon via the MEP pathway [110].
As described above, the emission of oxygenated BVOCsdepends on stomatal opening [84]. Stomatal closure inresponse to drought and salt stress is therefore expectedto reduce particularly the emission of these compounds.However, morning peaks of acetone and methanol emis-sions may be very high in drought-stressed leaves, becausethe temporary opening of stomata during times of higherhumidity allows the release of large pools of oxygenatedBVOCs that were built inside the leaf mesophyll [111,112].Thus, drought stress does not inhibit per se the biosyn-thesis of oxygenated BVOCs. Moreover, if the stressreaches levels that are able to damage membranes andcell walls, further increments of the emission of C6 com-pounds and methanol must be expected. However, emis-sion of these compounds has not been reported generally inresponse to drought and salt [101]. Indeed, C6 emissionsoccur in bursts, immediately after the damage to cellularstructures has occurred [5,113]. A recent experiment hasestablished a correlation between the emission of C6 vola-tiles and the damage to membranes, as assessed by the ionleakage, under a developing drought [114].
Ozone and other oxidants
In the atmosphere, volatile terpenes perform a dual action,depending on the presence of anthropogenic pollutants.When these compounds are absent, volatile terpenescleanse the atmosphere of ozone. In the presence of NOx,however, these BVOCs initiate reactions leading toincreased ozone formation [13]. The chemical reactivityof terpenes in the atmosphere led to the idea that volatileterpenes play a similar dual action inside the leaves, beforethey are released into the atmosphere [115,116].
In general, terpenes have been demonstrated to reduceozone damage and to quench ozone and reactive oxygenspecies (ROS) [115–118]. The mechanism(s) by which thisprotective effect occurs is still under investigation and theprevalent theories will be discussed below. Here we con-centrate on the impact of oxidative stresses on the emissionof BVOCs. If isoprene reacts with ozone and other oxidativespecies then it is expected that it disappears, concurrentlywith the appearance of its reaction products. In the atmos-phere, these products are mainly methyl-vinyl-ketone andmethacrolein, two compounds that are indeed found inchamber studies in which isoprene-emitting trees were
fumigated with ozone [119]. In plants exposed to ozoneeither a reduction [113,120] or a stimulation of isopreneandmonoterpene emissions is observed. The stimulation ofthe emission is more evident in response to acute andheavy doses of ozone [121–123] (e.g. 150–300 ppb) whereasit is often absent when plants are exposed to low doses ofozone above background [122,124]. Ozone-enhanced emis-sion of isoprene is due to a higher expression of the ISPSmRNA which probably upregulates the protein and theactivity of the enzyme [123]. Interestingly, such an upre-gulation is more evident in leaves that develop underenriched ozone and that build up a better resistance topollutants, as well as in new leaves that develop abovethose that have been ozonated [123]. Evidently, impair-ment of photosynthetic activities prevents older ozonatedleaves from enhancing the secondary metabolism, leadingto volatile terpene formation.
When plants are exposed to low ormoderate and chronicdoses of ozone, such an induction of the volatile terpenepathway is absent, and the expression of ISPS mRNA andthe level of ISPS protein may even be reduced [120,124].Clearly, the signals that activate the biochemistry of ter-pene formation, andwhich are unknown at present, are notreleased under these conditions. It has been hypothesized[125] that the induction of volatile terpenes in response toozone follows a hormetic dose–response relationship, i.e.that the volatile terpenes are increasingly induced at lowbut growing doses of ozone until an ozone threshold isreached after which the biosynthesis of terpenes isrepressed. The experiments, however, reveal a more com-plex picture, with biosynthesis of volatile terpenes beinginduced only when ozone dose overcomes a threshold thatmarks cellular damage but at which photosynthesis is notyet so heavily suppressed to be unable to supply enoughsubstrate for volatile terpenes. On the basis of these find-ings a long-term induction of isoprene biosynthesis, andthe consequent evolutionary hypothesis that high terpeneemitters will be favoured in a future more oxidative atmos-phere [126] may not hold true.
Ozone is a very damaging pollutant for plant cells, andone of the first recognized ozone effects is the denaturationof the lipids in cellular membranes [127]. It is thereforeexpected that volatiles that are associated with lipid per-oxidation are also more emitted in ozone-stressed leaves.Accordingly, bursts of C6 compound emissions wereobserved in ozone-stressed leaves and the lag time withwhich these compounds were emitted was proportional tothe ozone dose absorbed by the leaves [128] and to theozone-induced injuries [113]. These experiments clearlyshow that C6 compounds are in vivo indicators of mem-brane denaturation and damage as already indicated alsoin response to drought [114]. In addition, these studies[113,128] also revealed transient pulses of methanol andMeSA. While methanol emission has been mainly attrib-uted to demethylation of cell wall pectins, and is thereforelikely to be another indicator of ozone damage, the induc-tion of MeSA is more intriguing. MeSA emissions showed aweak association with high levels of ozone recorded in non-manipulative field experiments [98], but acute exposuresto ozone and UV light are apparently also able to inducebursts of this compound (Velikova et al., personal com-
Review Trends in Plant Science Vol.15 No.3
159

Author's personal copy
munication). Emissions of MeSA induced by oxidative andother environmental stresses may further contribute tobiogenic secondary organic aerosols [129,130] and mayalter the network of plant communication with otherorganisms which is mediated by chemical messengers [98].
Other climate change factors: UV-B radiation
Studies of the effect of UV-B radiation (wavebands of 290–
320 nm) onBVOCs emissions are rare, despite the fact thatUV-B radiation is known (i) in moderate doses (environ-mentally relevant) as a priming abiotic agent, triggeringthe formation of UV screening pigments and modulatingplant growth; and (ii) in higher doses (e.g. as a result of thestratospheric ‘ozone hole’ and indeed transiently present incertain regions surrounding the South Pole) as an agentcausing severe damage to the photosynthetic machinery,nucleic acids, and proteins [127].
The impact of UV-B radiation on terpenes seems toindicate a stimulation of their biosynthesis and emission.In one study with European oak [131] the higher emissionof isoprene under UV-B radiation was attributed to ahigher biomass density rather than to a higher instan-taneous photosynthetic rate. A more recent study [132]again showed increased isoprene emission rates of sub-arctic peatlands when irradiated with increasing levels ofUV-B radiation, and explained the rising emission as aconsequence of oxidative damage to membranes and to theinduction of the terpene defensive antioxidant pathway[133].
UV damage to cellular structure may also induce emis-sion of other BVOCs. Emission of methane under aerobicconditions originating from plant material [10] is a con-troversial topic in recent plant and atmospheric research,e.g. [134,135]. Plant-mediated transport of methaneoriginating from methanogenic soil microorganismsthrough the aerenchyma and out of the leaves of wetlandplants, e.g. rice [136] has been known from many years.However, there is growing evidence that UV radiation canmediate non-microbial methane release from cell wallmaterial, in particular pectin. In the initial work [10] itwas suggested that the methoxyl groups of pectin can beone, albeit not the only, source of aerobic methane, e.g.[137,138]. It was recently demonstrated [137–139] that therelease of methane from structural cell wall components offresh and dried leaf tissue was UV-B dependent. In linewith this observation, the authors [139] also showed thatthe removal ofmethoxyl groups interruptedmethane emis-sion from UV-irradiated pectin. Very recently it wasdemonstrated [140] that the emission of methane from cellwall material is mediated by UV-generated ROS [hydroxylradicals (�OH) and singlet oxygen; but not hydrogen per-oxide or superoxide radicals].
Environmental stresses as well as cellular signallingprocesses involve the formation of ROS. Therefore it mightbe speculated that aerobic methane formation is a com-mon, yet overlooked part of plant stress responses com-plementing the transient burst of BVOCs, i.e. methanol,acetaldehyde, and C6 alcohols and aldehydes [67,79], andthe long-lasting stimulation of terpene biosynthesis. Sup-port for this assumption is given by recent work demon-strating increased emissions of methane upon bacterial
infection, chemical generation of ROS [139] or physicalinjury [141].
Other climate change factors: atmospheric CO2
Rising CO2 concentration at global level is dramaticallyaffecting plant life. The primary effect is the increasedavailability of substrate for Rubisco, and the consequentenhancement of photosynthesis [142]. However, CO2 is onemore factor that uncouples terpene metabolism and emis-sions from photosynthesis.
As early as 1964, G. Sanadze [143] demonstrated that atlow CO2 concentration photosynthesis is reduced, but iso-prene emission increases. In a more recent laboratorystudy the highest emission of isoprene was detected atan intercellular CO2 concentration (Ci) of around 150–200ppmv, not far from the Ci experienced in nature by leaves oftree species [18], and then progressively decreased athigher Ci. Since then, many studies on different plantspecies have shown an inhibition of isoprene biosynthesisunder higher than ambient CO2 concentrations [144–148].For example elevated CO2 in a natural CO2 spring reducesISPS activity and isoprene emission from common reed(Phragmites australis) [37]. Compared to isoprene, less isknown about the influence of CO2 on light-dependentmonoterpene emission. A study on holm oak (Quercus ilex)growing under CO2 concentration double than ambient inopen top chambers [46] showed a parallel downregulationof mono-TPS activities and monoterpene emission. How-ever, studies conducted in holm oak plants growing fortheir entire life in natural CO2 springs under very high butnot steady CO2 levels did not reveal substantial inhibitionof the emitted monoterpenes [149], probably because thetreatment was also associated with recurrent droughtstress episodes that could also affect the emission (seeabove).
While a downregulation of ISPS andmono-TPS is gener-ally associated with lower emission of isoprene and mono-terpenes grown under elevated CO2 [145], the enzymeproperties are not necessarily the factors controlling theemissions. This control may be exerted primarily byreduced substrate availability (DMADP and GDP) result-ing from a downregulation of the MEP pathway. Althoughthis assumption has no direct support from the literature, asimultaneous decrease in isoprene emission and DMADPcontent was observed [145]. This reduction in isoprene(and also monoterpene) emission under elevated CO2
might result from enforced higher consumption rates ofcytosolic PEP (phosphoenolpyruvate) through PEPcarboxylase activity, thus lowering the rates of PEP trans-port into the chloroplast, where PEP, in its dephosphory-lated form (pyruvate) feeds into the MEP pathway. Basedon this work [145] and earlier suggestions by G. Sanadze[143,150] a double carboxylation hypothesis was proposed[148,151] for C3 plants (to which all of isoprene emittersbelong) with cytosolic PEP carboxylase and plastidic RuBPcarboxylase as antithetic precursors controlling the flow ofcarbon to plastidic terpene metabolism (as well as to theshikimic acid pathway and fatty acid biosynthesis) inresponse to changes in CO2 concentration. This idea getssupport from a study [152] showing an inverse relationshipof isoprene biosynthesis and PEP carboxylase activity
Review Trends in Plant Science Vol.15 No.3
160

Author's personal copy
when cottonwoods (Populus deltoides) were grown on highnitrate concentration, a condition favouring cytosolicorganic acids synthesis. In line with this scheme, mito-chondrial respiration can constitute a growing sink forcytosolic PEP under rising CO2, thus competing with theimport of this substrate in the chloroplast for terpenebiosynthesis [145]. Recent experiments with Free AirCO2 Enhancement (FACE) facilities, however, indicatedthat the inverse relationship between isoprene and respir-ation was not straightforward under elevated CO2, andthat competition occurs only when oxaloacetate productionfromPEP for anabolic support of respiration is strong, suchas in young, expanding leaves [153]. A positive CO2 effecton isoprene [144] andmonoterpene [154] emissions may beoccasionally observed. Moreover, no effect on isoprene andmonoterpenes was reported on poplar [125,153] or on Scotspine (Pinus sylvestris) [155].
Currently we do not know whether the CO2 impact onlight-dependent isoprene and monoterpene emission is ageneral effect and which fundamental biochemical mech-anisms are responsible. If the biosynthesis of volatileterpenes is ubiquitously enhanced at CO2 lower thanambient, then stress conditions in which a lower intercel-lular CO2 concentration is set by increasing resistances toCO2 diffusion can generically lead to higher emissions.
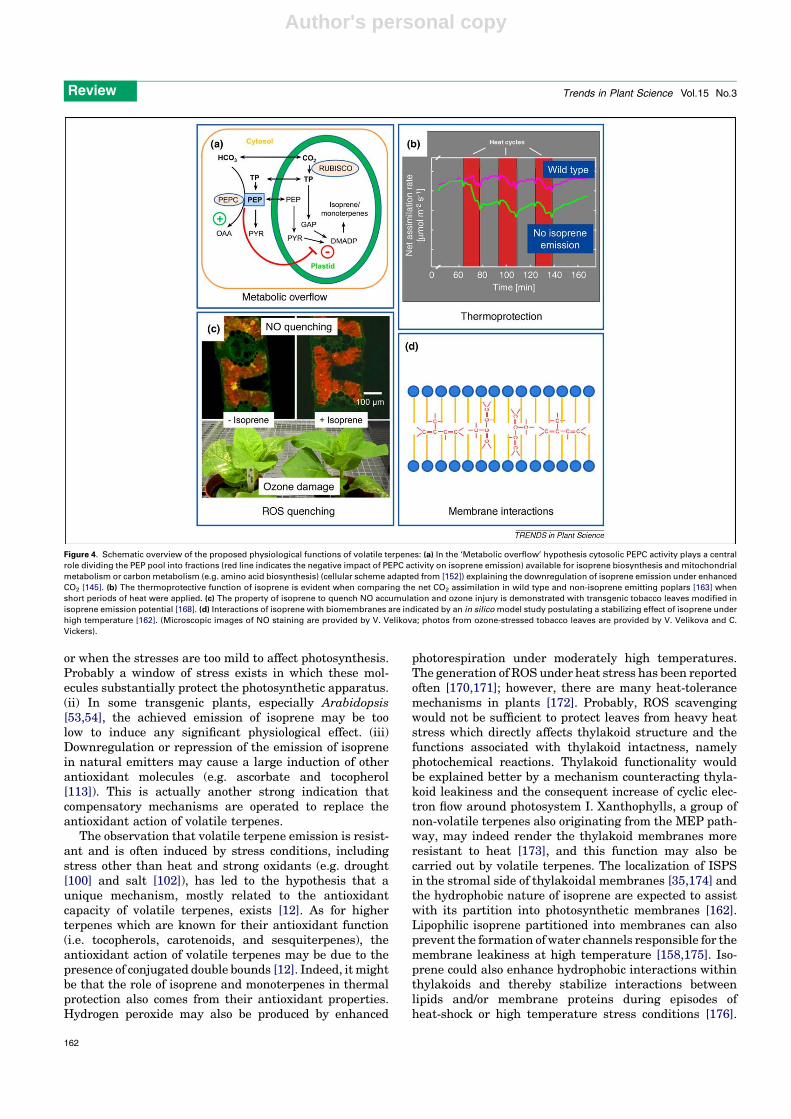
Why is emission of volatile terpenes induced byabiotic stress?Whereas oxygenated BVOCs aremainly catabolic productsof the denaturation of cellular walls and membranes, iso-prene andmonoterpenes are produced through a dedicatedmetabolic pathway that is stimulated by several abioticstresses (see above). This tightly regulated terpene biosyn-thesis and the observation that emission of volatile ter-penes represents a significant loss of photosyntheticcarbon, led to the proposition that these compounds playimportant physiological and ecological roles in the protec-tion of plants from environmental constraints. However,the debate is still ongoing whether one or more ecologicalactions should be attributed to volatile terpenes, andwhichare the physiological mechanisms that allow terpenes toexert their protective action. Two ‘metabolic’ hypothesessuggest that isoprene acts as a kind of ‘safety valve’ whichallows quenching energy or metabolites (Figure 4). Inparticular, one study [150] considers isoprene biosynthesisas a pathway for dissipation of excess photosyntheticenergy, whereas a more recent one [145] postulates thatisoprene biosynthesis prevents the overflow of chloroplas-tic DMADP. We consider as a ‘metabolic’ hypothesis alsothe ‘opportunistic hypothesis’ [41] which suggests thatvolatile terpenes are somehow alternative to ‘essential’terpenes (such as carotenoids, also formed through theMEP pathway, and for which an important antioxidantrole is clearly established). The same pool of carbon maygenerate volatile and/or essential terpenes according to theneed to face different constraints. This may be true alsowithin the class of volatile terpenes, as it was recentlydemonstrated that isoprene decreases whenmonoterpenesare synthesized in poplar leaves attacked by beetles [156].
A second group of hypotheses establishes a moredistinctive functional role for volatile terpenes in plant
protection against abiotic stressors (Figure 4). The mainhypothesis is that isoprene, as well as monoterpenes, arethermoprotective molecules, able to stabilize chloroplastmembranes during high temperature events [157] there-fore protecting the photosynthetic apparatus. There isample experimental support for this idea. In brief: (i)experiments in which plants have been fumigated withterpenes or in which the biosynthesis of these compoundshave been chemically blocked have shown that the photo-synthetic apparatus of isoprene-emitting plants is betterprotected against heat stress [158–160] and in particularagainst rapid temperature changes [161]. The interestingidea behind the latter observation is that isoprene acts as arapid mechanism of protection before plants can synthes-ize more complex molecules (including non-volatile ter-penes) that improve thermal stabilization. (ii) An insilico experiment has demonstrated that isoprene mayindeed partition into the phospho-lipid bilayer of mem-branes and maintain their stability, in particular duringexposure to high temperature [162]. (iii) Transgenic plantsthat have been engineered to emit isoprene or monoter-penes, or in which isoprene biosynthesis has beenrepressed are now available. Isoprene-emitting transgenicplants are more resistant to heat stress than wild types[53,54,163].
Volatile terpenes also appear to have a relevant anti-oxidant action. Again several lines of evidence support thishypothesis: (i) isoprene and monoterpenes have beenshown to reduce the damage caused by ozone [115,122]and ROS [117,133,164]. (ii) Evidence of the reaction be-tween isoprene and monoterpenes and ozone or other ROShas been produced by observing the appearance of thereaction products and the disappearance of the reagents[120,165]. In plants exposed to oxidative stresses isoprenehas also been shown to quench nitrogen reactive species(namely NO) that can also have an important role asmessenger molecules of the hypersensitive response tostress [166,167]. (iii) Transgenic tobacco (Nicotiana taba-cum) plants that have been engineered to emit isoprene aremore resistant to ozone toxicity than non-emitting wildtypes [168], and the ozone sensitivity of poplar clones isinversely related to their capacity to emit isoprene [169].
However, both functions of isoprene have been chal-lenged by studies in which the authors were not able toreproduce improved resistance to the stresses, especiallywhen using artificial systems or transgenic plants. Forinstance, photosynthesis analysis showed that transgenicArabidopsis plants do not benefit from isoprene when atransient heat stress occurs [53]. In transgenic, isoprene-emitting tobacco plants, tolerance of photosynthesisagainst transient high-temperature episodes could onlybe observed in lines emitting high levels of isoprene. More-over, this effect was very mild and could only be identifiedover repetitive stress events [168]. Finally, grey poplar(Populus � canescens) plants in which isoprene emissionwas efficiently repressed were not more sensible to oxi-dative stresses [113]. There may be good reasons thatexplain why the protection offered by volatile terpenes isnot observed in specific cases: (i) the protection offered bythese molecules may not be achieved when photosynthesisis already heavily impaired by prolonged or acute stresses,
Review Trends in Plant Science Vol.15 No.3
161
Author's personal copy
or when the stresses are too mild to affect photosynthesis.Probably a window of stress exists in which these mol-ecules substantially protect the photosynthetic apparatus.(ii) In some transgenic plants, especially Arabidopsis[53,54], the achieved emission of isoprene may be toolow to induce any significant physiological effect. (iii)Downregulation or repression of the emission of isoprenein natural emitters may cause a large induction of otherantioxidant molecules (e.g. ascorbate and tocopherol[113]). This is actually another strong indication thatcompensatory mechanisms are operated to replace theantioxidant action of volatile terpenes.
The observation that volatile terpene emission is resist-ant and is often induced by stress conditions, includingstress other than heat and strong oxidants (e.g. drought[100] and salt [102]), has led to the hypothesis that aunique mechanism, mostly related to the antioxidantcapacity of volatile terpenes, exists [12]. As for higherterpenes which are known for their antioxidant function(i.e. tocopherols, carotenoids, and sesquiterpenes), theantioxidant action of volatile terpenes may be due to thepresence of conjugated double bounds [12]. Indeed, it mightbe that the role of isoprene and monoterpenes in thermalprotection also comes from their antioxidant properties.Hydrogen peroxide may also be produced by enhanced
photorespiration under moderately high temperatures.The generation of ROS under heat stress has been reportedoften [170,171]; however, there are many heat-tolerancemechanisms in plants [172]. Probably, ROS scavengingwould not be sufficient to protect leaves from heavy heatstress which directly affects thylakoid structure and thefunctions associated with thylakoid intactness, namelyphotochemical reactions. Thylakoid functionality wouldbe explained better by a mechanism counteracting thyla-koid leakiness and the consequent increase of cyclic elec-tron flow around photosystem I. Xanthophylls, a group ofnon-volatile terpenes also originating from the MEP path-way, may indeed render the thylakoid membranes moreresistant to heat [173], and this function may also becarried out by volatile terpenes. The localization of ISPSin the stromal side of thylakoidal membranes [35,174] andthe hydrophobic nature of isoprene are expected to assistwith its partition into photosynthetic membranes [162].Lipophilic isoprene partitioned into membranes can alsoprevent the formation of water channels responsible for themembrane leakiness at high temperature [158,175]. Iso-prene could also enhance hydrophobic interactions withinthylakoids and thereby stabilize interactions betweenlipids and/or membrane proteins during episodes ofheat-shock or high temperature stress conditions [176].
Figure 4. Schematic overview of the proposed physiological functions of volatile terpenes: (a) In the ‘Metabolic overflow’ hypothesis cytosolic PEPC activity plays a central
role dividing the PEP pool into fractions (red line indicates the negative impact of PEPC activity on isoprene emission) available for isoprene biosynthesis and mitochondrial
metabolism or carbon metabolism (e.g. amino acid biosynthesis) (cellular scheme adapted from [152]) explaining the downregulation of isoprene emission under enhanced
CO2 [145]. (b) The thermoprotective function of isoprene is evident when comparing the net CO2 assimilation in wild type and non-isoprene emitting poplars [163] when
short periods of heat were applied. (c) The property of isoprene to quench NO accumulation and ozone injury is demonstrated with transgenic tobacco leaves modified in
isoprene emission potential [168]. (d) Interactions of isoprene with biomembranes are indicated by an in silico model study postulating a stabilizing effect of isoprene under
high temperature [162]. (Microscopic images of NO staining are provided by V. Velikova; photos from ozone-stressed tobacco leaves are provided by V. Velikova and C.
Vickers).
Review Trends in Plant Science Vol.15 No.3
162

Author's personal copy
Preliminary research using circular dichroism spec-troscopy, a valuable tool for probing the molecular archi-tecture of the complexes and supercomplexes and theirmacro-organization in the membrane system [177]confirms that a higher thermal stability of thylakoid mem-branes is induced in transgenic plants that are able to emitisoprene (Fortunati et al., personal communication).
Conclusions and future directionsWehave seen that abiotic stressesmay induce the emissionof multiple BVOCs. Many of the emitted compounds aresynthesized from the degradation of cellular structuresand may be used as reliable indicators of cell wall degra-dation (methanol and methane) or membrane denatura-tion (C6 volatiles). In the case of volatile terpenes theinduced emissions reflect the elicitation of the MEP path-way, revealing important function(s) of these compounds inthe protection against stresses. The physiological andecological functions of volatile terpenes are well estab-lished; however, more studies are needed to reveal themolecular and biochemical mechanisms that oversee theprotective role of volatile terpenes. Induction or alterationof these BVOCs emissions by abiotic stresses and otherclimate change factors may also contribute to modify thecommunication of plants with other organisms, namelyherbivores and carnivores that use BVOCs emissions asolfactory cues to retrieve hosts suitable both as food andshelter. Technical advances have made it possible to detectinduced emissions of MeSA not only in response to bioticstresses but also in plants subjected to abiotic stresses.This volatile is therefore emerging as a centralmolecule forthe signalling of stresses and for the consequent activationof systemic acquired resistance and hypersensitiveresponses.
AcknowledgmentsWe thank I. Zimmer for critical reading of the manuscript and C. Vickersand V. Velikova for providing us with tobacco images. The work wassupported by the ESF Project Volatile Organic Compounds in theBiosphere-Atmosphere System (VOCBAS). The German ResearchFoundation (DFG; SCHN653/4 to J.-P.S.) supported the research withinthe German joint research group ‘Poplar – a model to address tree-specificquestions’.
References1 Tholl, D. et al. (2006) Practical approaches to plant volatile analysis.
Plant J. 45, 540–5602 Knudsen, J.T. and Gershenzon, J. (2006) The chemical diversity of
floral scent. In Biology of Floral Scent (Dudareva, N. and Pichersky,E., eds), pp. 27–52, Taylor & Francis
3 Knudsen, J.T. et al. (1993) Floral scents: a check-list of volatilecompounds isolated by head-space techniques. Phytochemistry 33,253–280
4 Steeghs, M. et al. (2004) Proton-transfer-reaction mass spectrometryas a new tool for real time analysis of root-secreted volatile organiccompounds in Arabidopsis. Plant Physiol. 135, 47–58
5 Loreto, F. et al. (2006) On the induction of volatile organic compoundemissions by plants as consequence of wounding or fluctuations oflight and temperature. Plant Cell Environ. 29, 1820–1828
6 Pare, P.W. and Tumlinson, J.H. (1999) Plant volatiles as a defenseagainst insect herbivores. Plant Physiol. 121, 325–332
7 Dudareva, N. et al. (2004) Biochemistry of plant volatiles. PlantPhysiol. 135, 1993–2011
8 Feussner, I. and Wasternack, C. (2002) The lipoxygenase pathway.Annu. Rev. Plant Biol. 53, 275–297
9 Kreuzwieser, J. et al. (1999) Metabolic origin of acetaldehyde emittedby poplar (Populus tremula � P. alba) trees. J. Exp. Bot. 50, 757–
76510 Keppler, F. et al. (2006) Methane emission from terrestrial plants
under aerobic conditions. Nature 439, 187–19111 Keppler, F. et al. (2008)Methoxyl groups of plant pectin as a precursor
of atmospheric methane: evidence from deuterium labelling studies.New Phytol. 178, 808–814
12 Vickers, C.E. et al. (2009) A unified mechanism of action for volatileisoprenoids in plant abiotic stress. Nat. Chem. Biol. 5, 283–291
13 Fehsenfeld, F. et al. (1992) Emissions of volatile organic compoundsfrom vegetation and the implications for atmospheric chemistry.Global Biogeochem. Cyc. 6, 389–430
14 Guenther, A.B. et al. (1995) A global model of natural volatile organiccompound emissions. J. Geophys. Res. 100, 8873–8892
15 Ghirardo, A. et al. (2010) Determination of de novo and poolemissions of terpenes from four common boreal/alpine trees by13CO2 labeling and PTR-MS analysis. Plant Cell Environ DOI:10.1111/j.1365-3040.2009.02104.x (http://www3.interscience.wiley.com/journal/123224612/abstract)
16 Loreto, F. et al. (2009) One species, many terpenes: matching chemicaland biological diversity. Trends Plant Sci. 14, 416–420
17 Kesselmeier, J. and Staudt, M. (1999) Biogenic volatile organiccompounds (VOC): an overview on emission, physiology andecology. J. Atmos. Chem. 33, 23–88
18 Loreto, F. and Sharkey, T.D. (1990) A gas exchange study ofphotosynthesis and isoprene emission in red oak (Quercus rubraL.). Planta 182, 523–531
19 Loreto, F. et al. (1996) Influence of environmental factors and aircomposition on the emission of a-pinene from Quercus ilex leaves.Plant Physiol. 110, 267–275
20 Delwiche, C.F. and Sharkey, T.D. (1993) Rapid appearance of 13C inbiogenic isoprene when 13CO2 is fed to intact leaves. Plant CellEnviron. 16, 587–591
21 Rohmer, M. (2008) From molecular fossils of bacterial hopanoids tothe formation of isoprene units: Discovery and elucidation of themethylerythritol phosphate pathway. Lipids 43, 1095–1107
22 Lichtenthaler, H.K. (1999) The 1-deoxy-D-xylulose 5-phosphatepathway of isoprenoid biosynthesis in plants. Annu. Rev. PlantPhysiol. Plant Mol. Biol. 50, 47–65
23 Sallaud, C. et al. (2009) A novel pathway for sesquiterpenebiosynthesis from Z,Z-farnesyl pyrophosphate in the wild tomatoSolanum habrochaites. Plant Cell 21, 301–317
24 Schuhr, C.A. et al. (2003) Quantitative assessment of crosstalkbetween the two isoprenoid biosynthesis pathways in plants byNMR spectroscopy. Phytochem. Rev. 2, 3–16
25 Laule, O. et al. (2003) Crosstalk between cytosolic and plastidialpathways of isoprenoid biosynthesis in Arabidopsis thaliana. Proc.Natl. Acad. Sci. U. S. A. 100, 6866–6871
26 Schmidt, A. and Gershenzon, J. (2008) Cloning and characterizationof two different types of geranyl diphosphate synthases from Norwayspruce (Picea abies). Phytochemistry 69, 49–57
27 Bohlmann, J. et al. (1998) Plant terpenoid synthases: molecularbiology and phylogenetic analysis. Proc. Natl. Acad. Sci. U. S. A.95, 4126–4133
28 Vandermoten, S. et al. (2009) New insights into short-chainprenyltransferases: structural features, evolutionary history andpotential for selective inhibition. Cell. Mol. Life Sci. 66, 3685–
369529 Wise, M.L. and Croteau, R. (1999) Monoterpene biosynthesis. In
Comprehensive Natural Products Chemistry: Isoprenoids (Cane,D.E., ed.), pp. 97–153, Elsevier Science
30 Trapp, S.C. and Croteau, R.B. (2001) Genomic organization of plantterpene synthases and molecular evolutionary implications. Genetics158, 811–832
31 Degenhardt, J. et al. (2009) Monoterpene and sesquiterpenesynthases and the origin of terpene skeletal diversity in plants.Phytochemistry 70, 1621–1637
32 Gray, D. et al. (2006) Influences of temperature history, water stressand needle age on methylbutenol emissions. Ecology 84, 765–776
33 Silver, G.M. and Fall, R. (1995) Characterization of aspen isoprenesynthase, an enzyme responsible for leaf isoprene emission to theatmosphere. J. Biol. Chem. 270, 13010–13016
Review Trends in Plant Science Vol.15 No.3
163

Author's personal copy
34 Fisher, A.J. et al. (2000) Enzymatic synthesis of methylbutenol fromdimethylallyl diphosphate in needles of Pinus sabiniana. Arch.Biochem. Biophys. 383, 128–134
35 Schnitzler, J.P. et al. (2010) Poplar volatiles - biosynthesis, regulationand (eco)physiology of isoprene and stress-induced isoprenoids. PlantBiol. 12, 302–316
36 Lehning, A. et al. (1999) Isoprene synthase activity and its relation toisoprene emission in Quercus robur L. leaves. Plant Cell Environ. 22,495–504
37 Scholefield, P.A. et al. (2004) Impact of rising CO2 on VOC emissions:isoprene emission from Phragmites australis growing at elevated CO2
in a natural carbon dioxide spring. Plant Cell Environ. 27, 381–39238 Sharkey, T.D. et al. (2005) Evolution of the isoprene biosynthetic
pathway in kudzu. Plant Physiol. 137, 700–71239 Fischbach, R.J. et al. (2000) Monoterpene synthase activities in leaves
of Picea abies (L.) Karst. and Quercus ilex L. Phytochemistry 54,257–265
40 Tholl, D. et al. (2001) Partial purification and characterization of theshort-chain prenyltransferases, geranyl diphosphate synthase andfarnesyl diphosphate synthase, from Abies grandis (Grand Fir).Arch. Biochem. Biophys. 386, 233–242
41 Owen, S. and Penuelas, J. (2005) Opportunistic emissions of volatileisoprenoids. Trends Plant Sci. 10, 420–426
42 Dudareva, N. et al. (2006) Plant volatiles: Recent advances and futureperspectives. Crit. Rev. Plant Sci. 25, 417–440
43 Schnitzler, J.P. et al. (2004) Hybridisation of European oaks (Quercusilex x Q. robur) results in a mixed isoprenoid emitter type. Plant CellEnviron. 27, 585–593
44 Fischbach, R.J. et al. (2002) Seasonal pattern of monoterpenesynthase activities in leaves of the evergreen tree Quercus ilex L.Physiol. Plant. 114, 354–360
45 Grote, R. et al. (2009) Modelling the drought impact on monoterpenefluxes from an evergreenMediterranean forest canopy.Oecologia 160,213–223
46 Loreto, F. et al. (2001) Monoterpene emission and monoterpenesynthase activities in the Mediterranean evergreen oak Quercusilex L. grown at elevated CO2 concentrations. Global Change Biol.7, 709–717
47 Fischbach, R.J. et al. (2001) Isolation and functional analysis of acDNA encoding amyrcene synthase of holm oak (Quercus ilex L.).Eur.J. Biochem. 268, 5633–5638
48 Andres-Montaner, D. (2009) Study of gene regulation and function ofmonoterpene synthases in Holm oak (Quercus ilex L.) and transgenicbirch (Betula pendula Roth). PhD thesis, FZK-Bericht 7511
49 Aubourg, S. et al. (2002) Genomic analysis of the terpenoid synthase(AtTPS) gene family of Arabidopsis thaliana. Mol. Genet. Genomics267, 730–745
50 Chen, F. et al. (2003) Biosynthesis and emission of terpenoids volatilesfrom Arabidopsis flowers. Plant Cell 15, 481–494
51 Tholl, D. et al. (2005) Two sesquiterpene synthases are responsible forthe complex mixture of sesquiterpenes emitted from Arabidopsisflowers. Plant J. 42, 757–771
52 VanPoecke, R.M.P. et al. (2001)Herbivore induced volatile productionby Arabidopsis thaliana leads to attraction of the parasitoid Cotesiarubecula: Chemical, behavioural and gene-expression analysis. J.Chem. Ecol. 27, 1911–1928
53 Loivamaki, M. et al. (2007) Arabidopsis, a model to study biologicalfunctions of isoprene emission? Plant Physiol. 144, 1066–1078
54 Sasaki, K. et al. (2007) Plants utilize isoprene emission as athermotolerance mechanism. Plant Cell Physiol. 48, 1254–1262
55 Loivamaki, M. et al. (2008) Isoprene interferes with the attraction ofbodyguards by herbaceous plants. Proc. Natl. Acad. Sci. U. S. A. 105,17430–17435
56 Bonn, B. and Moortgart, G.K. (2003) Sesquiterpene ozonolysis: originof atmospheric new particle formation from biogenic hydrocarbons.Geophys. Res. Lett. 30, 1585–1589
57 Arimura, G. et al. (2004) Forest tent caterpillars (Malacosomadisstria) induce systemic and diurnal emissions of terpenoidvolatiles in hybrid poplar (Populus trichocarpa x deltoides): cDNAcloning, functional characterization, and patterns of gene expressionof (�)-germacrene D synthase, PtdTPS1. Plant J. 37, 603–616
58 Ralph, S.G. et al. (2008) Analysis of 4,664 high-quality sequence-finished poplar full-length cDNA clones and their utility for the
discovery of genes responding to insect feeding. BMC Genomics 9,57–75
59 Hertzberg, M. et al. (2001) A transcriptional roadmap to woodformation. Proc. Natl. Acad. Sci. U. S. A. 98, 14732–14737
60 Ralph, S.G. et al. (2006) Genomics of hybrid poplar (Populustrichocarpa x deltoides) interacting with forest tent caterpillars(Malacosoma disstria): normalized and full-length cDNA libraries,expressed sequence tags, and a cDNA microarray for the study ofinsect-induced defences in poplar. Mol. Ecol. 15, 1275–1297
61 Tuskan, G. et al. (2006) The genome of black cottonwood, Populustrichocarpa (Torr & Gray ex. Brayshaw). Science 313, 1596–1604
62 Guenther, A.B. (2002) The contribution of reactive carbon emissionsfrom vegetation to the carbon balance of terrestrial ecosystems.Chemosphere 49, 837–844
63 Ricard, J. andNoat, G. (1986) Electrostatic effects and the dynamics ofenzyme reactions at the surface of plant cells. 1. A theory of the ioniccontrol of a complex multi-enzyme system. Eur. J. Biochem. 155, 183–
19064 Gaffe, J. et al. (1994) Pectin methylesterase isoforms in tomato
(Lycopersicon esculentum) tissues. Effects of expression of a pectinmethylesterase antisense gene. Plant Physiol. 105, 199–203
65 Mudgett, M.B. and Clarke, S. (1993) Characterization of plant L-isoaspartyl methyltransferases that may be involved in seed survival.Purification, characterization and sequence analysis of the wheatgerm enzyme. Biochemistry 32, 11100–11111
66 Cossins, E.A. (1987) Folate biochemistry and the metabolism of one-carbon units. In The Biochemistry of Plants (Vol 11) (Davies, D.D.,ed.), In pp. 317–353, Academic Press
67 Davison, B. et al. (2008) Cut-induced VOC emissions from agriculturalgrasslands. Plant Biol. 10, 76–85
68 Baldwin, I.T. et al. (2006) Volatile signaling in plant-plantinteractions: ‘talking trees’ in the genomics era. Science 311, 812–
81569 Fall, R. (2003) Abundant oxygenates in the atmosphere: a biochemical
perspective. Chem. Rev. 103, 4941–495170 Graus, M. et al. (2004) Transient release of oxygenated VOC during
light–dark transitions. Plant Physiol. 135, 1967–197571 Kreuzwieser, J. et al. (2001) Acetaldehyde emission by the leaves of
Trees - correlation with physiological and environmental parameters.Physiol. Plant. 113, 41–49
72 Holzinger, R. et al. (2000) Emissions of volatile organic compoundsfrom Quercus ilex L. measured by Proton Transfer Reaction Massspectrometry (PTR-MS) under different environmental conditions. J.Geophys. Res. 105, 20573–20579
73 Kreuzwieser, J. et al. (2002) Xylem-transported glucose as additionalcarbon source for leaf isoprene formation in Quercus robur. NewPhytol. 156, 171–178
74 Cojocariu, C. et al. (2004) Correlation of short-chained carbonylsemitted from Picea abies with physiological and environmentalparameters. New Phytol. 162, 717–727
75 Karl, T. et al. (2002) Transient releases of acetaldehyde from treeleaves – products of a pyruvate overflow mechanism? Plant CellEnviron. 25, 1121–1131
76 Hatanaka, A. (1993) The biogeneration of green odour by green leaves.Phytochemistry 34, 1201–1218
77 Croft, K.P.C. et al. (1993) Volatile products of the lipoxygenasepathway evolved from Phaseolus vulgaris L. leaves inoculatedwith Pseudomonas syringae pv phaseolicola. Plant Physiol. 101,13–24
78 Frost, C.J. et al. (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primesresponses against herbivores. Ecol. Lett. 10, 490–498
79 Fall, R. et al. (1999) Volatile organic compounds emitted after leafwounding: on-line analysis by proton-transfer-reaction massspectrometry. J. Geophys. Res. 104, 15963–15974
80 Flexas, J. et al. (2006) Diffusive and metabolic limitations tophotosynthesis under drought and salinity in C3 plants. Plant Biol.6, 269–279
81 Loreto, F. et al. (1996) Different sources of acetyl CoA contribute toform three classes of terpenoid emitted byQuercus ilex L. leaves.Proc.Natl. Acad. Sci. U. S. A. 93, 9966–9969
82 Schnitzler, J.P. et al. (2004) Contribution of different carbon sources toisoprene biosynthesis in poplar leaves. Plant Physiol. 135, 152–160
Review Trends in Plant Science Vol.15 No.3
164

Author's personal copy
83 Brilli, F. et al. (2007) Response of isoprene emission and carbonmetabolism to drought in white poplar (Populus alba) saplings.New Phytol. 175, 244–254
84 Niinemets, U. et al. (2004) Physiological and physico-chemicalcontrols on foliar volatile organic compound emissions. TrendsPlant Sci. 9, 180–186
85 Gershenzon, J. et al. (2000) Regulation of monoterpene accumulationin leaves of peppermint. Plant Physiol. 122, 205–214
86 Pasqua, G. et al. (2002) The role of isoprenoid accumulation andoxidation in sealing wounded needles of Mediterranean pines.Plant Sci. 163, 355–359
87 Litvak, M.E. and Monson, R.K. (1998) Induced and constitutivemonoterpene defenses in conifer needles in relation to herbivorypatterns. Oecologia 114, 531–540
88 Alessio, G.A. et al. (2004) Direct and indirect impacts of fire on theisoprenoids emission fromMediterranean vegetation. Funct. Ecol. 18,357–364
89 Fall, R. and Monson, R.K. (1992) Isoprene emission rate andintercellular isoprene concentration as influenced by stomataldistribution and conductance. Plant Physiol. 100, 987–992
90 Bruggemann, N. and Schnitzler, J.P. (2002) Comparison of isopreneemission, intercellular isoprene concentration and photosyntheticperformance in water-limited oak (Quercus pubescens Willd. andQuercus robur L.) saplings. Plant Biol. 4, 456–463
91 Monson, R.K. et al. (1992) Relationship among isoprene emission rate,photosynthesis, and isoprene synthase activity as influenced bytemperature. Plant Physiol. 98, 1175–1180
92 Singsaas, E.L. and Sharkey, T.D. (2000) Regulation of isoprenesynthesis during high temperature stress. Plant Cell Environ. 23,751–757
93 Magel, E. et al. (2007) Determination of the role of products ofphotosynthesis in substrate supply of isoprenoid biosynthesis inpoplar leaves. Atmos. Environ. 40, S138–S151
94 Schade, G.W. andGoldstein, A.H. (2001) Fluxes of oxygenated volatileorganic compounds from a ponderosa pine plantation. J. Geophys. Res.106, 3111–3124
95 Schade, G.W. and Custer, T.G. (2004) OVOC emissions fromagricultural soil in northern Germany during the 2003 Europeanheat wave. Atmos. Environ. 38, 6105–6114
96 Dicke, M. et al. (1999) Jasmonic acid and herbivory differentiallyinduce carnivore-attracting plant volatiles in lima bean plants. J.Chem. Ecol. 25, 1907–1922
97 Filella, I. et al. (2006) Dynamics of the enhanced emissions ofmonoterpenes and methyl salicylate, and decreased uptake offormaldehyde, by Quercus ilex leaves after application of jasmonicacid. New Phytol. 169, 135–144
98 Karl, T. et al. (2008) Chemical sensing of plant stress at the ecosystemscale. Biogeosciences 5, 1287–1294
99 Sharkey, T.D. and Loreto, F. (1993) Water stress, temperature, andlight effects on the capacity for isoprene emission and photosynthesisof kudzu leaves. Oecologia 95, 328–333
100 Fortunati, A. et al. (2008) Isoprene emission is not temperature-dependent during and after severe drought-stress: a physiologicaland biochemical analysis. Plant J. 55, 687–697
101 Teuber, M. et al. (2008) VOC emission of Grey poplar leaves asaffected by salt stress and different N sources. Plant Biol. 10, 86–96
102 Loreto, F. and Delfine, S. (2000) Emission of isoprene fromsalt-stressed Eucalyptus globulus leaves. Plant Physiol. 123, 1605–
1610103 Loreto, F. et al. (2004) 13C labelling reveals chloroplastic and extra-
chloroplastic pools of dimethylallyl pyrophosphate and theircontribution to isoprene formation. Plant Physiol. 135, 1903–1907
104 Lehning, A. et al. (2001)Modeling of annual variations of oak (Quercusrobur L.) isoprene synthase activity to predict isoprene emissionrates. J. Geophys. Res. 106, 3157–3166
105 Rennenberg, H. et al. (2006) Physiological responses of forest trees toheat and drought. Plant Biol. 8, 556–571
106 Arneth, A. et al. (2008) Effects of species composition, land surfacecover, CO2 concentration and climate on isoprene emissions fromEuropean forests. Plant Biol. 10, 150–162
107 Bertin, N. and Staudt, M. (1996) Effect of water stress onmonoterpene emissions from young potted holm oak (Quercus ilexL.) trees. Oecologia 107, 456–462
108 Llusia, J. and Penuelas, J. (1998) Changes in terpene content andemission in potted Mediterranean woody plants under severedrought. Can. J. Bot. 76, 1366–1373
109 Lavoir, A.V. et al. (2009) Drought reduced monoterpene emissionsfrom Quercus ilex trees: Results from a throughfall displacementexperiment within a forest ecosystem. Biogeosciences 6, 863–893
110 Llusia, J. et al. (2006) Seasonal contrasting changes of foliarconcentrations of terpenes and other volatile organic compounds infour dominant species of a Mediterranean shrubland submitted to afield experimental drought and warming. Physiol. Plant 127, 632–649
111 Fares, S. et al. (2009) The ACCENT-VOCBAS field campaign onbiosphere-atmosphere interactions in a Mediterranean ecosystemof Castelporziano (Rome): site characteristics, climatic andmeteorological conditions, and eco-physiology of vegetation.Biogeosciences 6, 1043–1058
112 Filella, I. et al. (2009) Short-chained oxygenated VOC emissions inPinus halepensis in response to changes in water availability. ActaPhysiol. Plant. 31, 311–318
113 Behnke, K. et al. (2009) RNAi mediated suppression of isoprenebiosynthesis impacts ozone tolerance. Tree Physiol. 29, 725–736
114 Capitani, D. et al. (2009) In situ investigation of leaf water status byportable unilateral NMR. Plant Physiol. 149, 1638–1647
115 Loreto, F. et al. (2001) Ozone quenching properties of isoprene and itsantioxidant role in leaves. Plant Physiol. 26, 993–1000
116 Hewitt, N.C. et al. (1990) Hydroperoxides in plants exposed to ozonemediate air pollution damage to alkene emitters. Nature 344, 56–58
117 Affek, H.P. and Yakir, D. (2002) Protection by isoprene against singletoxygen in leaves. Plant Physiol. 129, 269–277
118 Loreto, F. and Fares, S. (2007) Is ozone flux inside leaves only adamage indicator? Clues from volatile isoprenoid studies. PlantPhysiol. 143, 1096–1100
119 Fares, S. et al. (2008) Stomatal uptake and stomatal deposition ofozone in isoprene and monoterpene emitting plants. Plant Biol. 10,44–54
120 Ryan, A. et al. (2009) Defining hybrid poplar (Populusdeltoides � Populus trichocarpa) tolerance to ozone: identifying keyparameters. Plant Cell Environ. 32, 31–45
121 Loreto, F. et al. (2004) Impact of ozone on monoterpene emissions andevidences for an isoprene-like antioxidant action of monoterpenesemitted by Quercus ilex (L.) leaves. Tree Physiol. 24, 361–367
122 Velikova, V. et al. (2005) Localized ozone fumigation system forstudying ozone effects on photosynthesis, respiration, electrontransport rate and isoprene emission in field-grown Mediterraneanoak species. Tree Physiol. 25, 1523–1532
123 Fares, S. et al. (2006) Impact of high ozone on isoprene emission,photosynthesis and histology of developing Populus alba leavesdirectly or indirectly exposed to the pollutant. Physiol. Plant. 128,456–465
124 Calfapietra, C. et al. (2007) Isoprene synthase expression and proteinlevels are reduced under elevated O3 but not under elevated CO2
(FACE) in field-grown aspen trees. Plant Cell Environ. 30, 654–
661125 Calfapietra, C. et al. (2009) Volatile organic compounds from Italian
vegetation and their interaction with ozone. Environ. Poll. 157, 1478–
1486126 Lerdau, M. (2007) A positive feedback with negative consequences.
Science 316, 212–213127 Pell, E.J. et al. (1997) Ozone induced oxidative stress: mechanism of
action and reaction. Physiol. Plant. 100, 264–273128 Beauchamp, J. et al. (2005) Ozone induced emissions of biogenic VOC
from tobacco: relationships between ozone uptake and emission ofLOX products. Plant Cell Environ. 28, 1334–1343
129 Clays, M. et al. (2004) Formation of secondary organic aerosolsthrough photooxidation of isoprene. Science 303, 1173–1175
130 Kanakidou, M. et al. (2005) Organic aerosol and global climatemodeling: a review. Atmos. Chem. Phys. 5, 1053–1123
131 Harley, P. et al. (1996) Effects of elevated levels of UV-B radiation onphotosynthesis and isoprene emission in Gambel’s oak and velvetbean. Global Change Biol. 2, 149–154
132 Tiiva, P. et al. (2007) Isoprene emission from a subarctic peatlandunder enhanced UV-B radiation. New Phytol. 176, 346–355
133 Loreto, F. and Velikova, V. (2001) Isoprene produced by leavesprotects the photosynthetic apparatus against ozone damage,
Review Trends in Plant Science Vol.15 No.3
165

Author's personal copy
quenches ozone products, and reduces lipid peroxidation of cellularmembranes. Plant Physiol. 127, 1781–1787
134 Dueck, T.A. et al. (2007) No evidence for substantial aerobic methaneemission by terrestrial plants: A 13C-labelling approach. New Phytol.175, 29–35
135 Nisbet, R.E.R. et al. (2009) Emission of methane from plants. Proc. R.Soc. B-Bio. Sci. 276, 1347–1354
136 Butterbach-Bahl, K. et al. (1997) Impact of gas transport through ricecultivars on methane emission from rice paddy fields. Plant CellEnviron. 20, 1175–1183
137 Vigano, I. et al. (2008) Effect of UV radiation and temperature on theemission of methane from plant biomass and structural components.Biogeosciences 5, 937–947
138 Bruggemann, N. et al. (2009) Nonmicrobial aerobic methane emissionfrom poplar shoot cultures under low-light conditions.NewPhytol. 23,912–918
139 McLeod, A.R. et al. (2008) Ultraviolet radiation drives methaneemissions from terrestrial plant pectins. New Phytol. 180, 124–132
140 Messenger, D.J. et al. (2009) The role of ultraviolet radiation,photosensitizers, reactive oxygen species and ester groups inmechanisms of methane formation from pectin. Plant Cell Environ.32, 1–9
141 Wang, Z.P. et al. (2009) Physical injury stimulates aerobic methaneemissions from terrestrial plants. Biogeosciences 6, 615–621
142 Long, S.P. et al. (2004) Rising atmospheric carbon dioxide: plants facethe future. Annu. Rev. Plant Biol. 55, 591–628
143 Sanadze, G.A. (1964) Light-dependent excretion of isoprene by plants.Photosyn. Res. 2, 701–707
144 Sharkey, T.D. et al. (1991) High carbon dioxide and sun/shade effectson isoprene emission from oak and aspen tree leaves. Plant CellEnviron. 14, 333–338
145 Rosenstiel, T.N. et al. (2003) Increased CO2 uncouples growth fromisoprene emission in an agriforest ecosystem. Nature 421, 256–259
146 Centritto, M. et al. (2004) Profiles of isoprene emission andphotosynthetic parameters in hybrid poplars exposed to free-airCO2 enrichment. Plant Cell Environ. 27, 403–412
147 Pegoraro, E. et al. (2004) Effect of elevated CO2 concentration andvapour pressure deficit on isoprene emission from leaves of Populusdeltoides during drought. Funct. Plant Biol. 31, 1137–1147
148 Wilkinson, M.J. et al. (2009) Leaf isoprene emission rate as a functionof atmospheric CO2 concentration.Global Change Biol. 15, 1189–1200
149 Rapparini, F. et al. (2004) Isoprenoid emission in trees of Quercuspubescens and Quercus ilex with lifetime exposure to naturally highCO2 environment. Plant Cell Environ. 27, 381–391
150 Sanadze, G.A. (2004) Biogenic isoprene (A review). Russ. J. PlantPhysiol. 51, 729–741
151 Monson, R.K. et al. (2009) Biochemical controls of the CO2 response ofleaf isoprene emission: an alternative view of Sanadze’s doublecarboxylation scheme. Ann. Agrar. Sci. 7, 21–29
152 Rosenstiel, T.N. et al. (2004) Induction of poplar leaf nitratereductase: a test of extrachloroplastic control of isoprene emissionrate. Plant Biol. 6, 12–21
153 Loreto, F. et al. (2007) The relationship between isoprene emissionrate and dark respiration rate in white poplar (Populus alba L.)leaves. Plant Cell Environ. 30, 662–669
154 Staudt, M. et al. (2001) Effect of elevated CO2 on monoterpeneemission of young Quercus ilex L. trees and its relation tostructural and ecophysiological parameters. Tree Physiol. 21, 437–
445155 Raisanen, T. et al. (2008) Effects of elevated CO2 and temperature on
monoterpene emission of Scots pine (Pinus sylvestris L.). Atmos.Environ. 42, 4160–4171
156 Brilli, F. et al. (2009) Constitutive and herbivore-inducedmonoterpenes emitted by Populus x euroamericana leaves are keyvolatiles that orient Chrysomela populi beetles. Plant Cell Environ.32, 542–552
157 Sharkey, T.D. and Singsaas, E.L. (1995) Why plants emit isoprene.Nature 374, 769–1769
158 Singsaas, E.L. et al. (1997) Isoprene increases thermotolerance ofisoprene-emitting species. Plant Physiol. 115, 1413–1420
159 Loreto, F. et al. (1998) On themonoterpene emission under heat stressand on the increased thermotolerance of leaves of Quercus ilexfumigated with selected monoterpenes. Plant Cell Environ. 21,101–107
160 Velikova, V. and Loreto, F. (2005) On the relationship betweenisoprene emission and thermotolerance in Phragmites australisleaves exposed to high temperatures and during the recovery froma heat stress. Plant Cell Environ. 28, 318–327
161 Sharkey, T.D. et al. (2008) Isoprene emission from plants: Why andhow. Ann. Bot. 101, 5–18
162 Siwko, M.E. et al. (2007) Does isoprene protect plant membranes fromthermal shock? Amolecular dynamics study. Biochim. Biophys. Acta -Biomembranes 1768, 198–206
163 Behnke, K. et al. (2007) Transgenic, non-isoprene emitting poplarsdon’t like it hot. Plant J. 51, 485–499
164 Velikova, V. et al. (2004) Endogenous isoprene protects Phragmitesaustralis leaves against singlet oxygen. Physiol. Plant. 122, 219–225
165 Pinto, D.M. et al. (2007) The effects of increasing atmospheric ozoneon biogenic monoterpene profiles and the formation of secondaryaerosols. Atmos. Environ. 41, 4877–4887
166 Velikova, V. et al. (2005) Isoprene decreases the concentration of nitricoxide in leaves exposed to elevated ozone. New Phytol. 166, 419–426
167 Velikova, V. et al. (2008) Isoprene and nitric oxide reduce damages inleaves exposed to oxidative stress. Plant Cell Environ. 31, 1882–1894
168 Vickers, C.E. et al. (2009) Isoprene synthesis protects tobacco plantsfrom oxidative stress. Plant Cell Environ. 32, 520–531
169 Calfapietra, C. et al. (2008) Isoprene emission rates under elevatedCO2 and O3 in two field-grown aspen clones differing for theirsensitivity to O3. New Phytol. 179, 55–61
170 Vallelian-Bindschedler, L. et al. (1998) Heat-induced resistance inbarley to powdery mildew (Blumeria graminis f.sp. hordei) isassociated with a burst of active oxygen species. Physiol. Mol.Plant Pathol. 52, 185–199
171 Vacca, R.A. et al. (2004) Production of reactive oxygen species,alteration of cytosolic ascorbate peroxidase, and impairment ofmitochondrial metabolism are early events in heat shock-inducedprogrammed cell death in tobacco bright-yellow 2 cells. Plant Physiol.134, 1100–1112
172 Sharkey, T.D. and Schrader, S.M. (2006) High temperature stress. InPhysiology and Molecular Biology of Stress Tolerance in Plants(Madhava Rao, K.V. et al., eds), pp. 101–129, Springer
173 Havaux,M. and Tardy, F. (1996) Temperature-dependent adjustmentof the thermal stability of photosystem II in vivo: possibleinvolvement of xanthophyll-cycle pigments. Planta 198, 324–333
174 Wildermuth, M.C. and Fall, R. (1998) Biochemical characterization ofstromal and thylakoid-bound isoforms of isoprene synthase in willowleaves. Plant Physiol. 116, 1111–1123
175 Sharkey, T.D. et al. (2001) Isoprene increases thermotolerance offosmidomycin-fed leaves. Plant Physiol. 125, 2001–2006
176 Sharkey, T.D. and Yeh, S. (2001) Isoprene emissions from plants.Annu. Rev. Plant Physiol. Plant Mol. Biol. 52, 407–436
177 Garab, G. and van Amerongen, H. (2009) Linear dichroism (LD) andcircular dichroism (CD) in photosynthesis research. Photosyn. Res.101, 135–146
Review Trends in Plant Science Vol.15 No.3
166