transcription factors and genes in abiotic stress
TRANSCRIPT

Transcription Factors and Environmental Stresses in Plants
Loredana F. Ciarmiello1*, Pasqualina Woodrow2, Pasquale Piccirillo1, Antonio De Luca1
and Petronia Carillo2
1)Consiglio per la Ricerca e la Sperimentazione in Agricoltura – Fruit Tree Research Unit,
Via Torrino, 3 – 81100 Caserta, Italy 2)II University of Naples, Dipartimento di Scienze e Tecnologie Ambientali, Biologiche e
Farmaceutiche, via Vivaldi 43, 81100 Caserta, Italy
Abstract
Plants are constantly exposed to changes in environmental conditions that, when are
rapid and extreme, can be perceive as stresses. Abiotic stresses such as salt, drought and
extreme temperatures, whose effects have been exacerbated by climate changes, are the
most serious factors limiting the productivity of agricultural crops, with adverse effects
on germination, plant vigour and crop yield. Worldwide, it has been estimated that
approximately 70% of yield reduction is the direct result of abiotic stresses. Moreover
the ever increasing pressure put on agricultural land by burgeoning human populations
has resulted in land degradation, a cultivation shift to more marginal areas and soil
types, and heavier requirements for agricultural productivity per unit area.
A large number of abiotic stress responsive genes have been reported in a variety of
plants including Arabidopsis but also the major crops such as rice, wheat, maize and
barley. Transcriptional control of the expression of these genes is a crucial part of the
plant response to abiotic stresses. Therefore in the last years the transcriptional
regulatory networks in response to abiotic stresses have been the subject of intense
research, that have been productive in identifying transcription factors (TFs) as
important 'key or master regulators' of gene expression under stress. An increasing
number of TFs have been recently described and essential transcription factor binding
regions have been identified for many genes. In fact, these regulatory systems are
achieved through specific cis-elements in the promoter regions of target genes, which
are called regulons. At least four different regulons that are active in response to abiotic
stresses have been identified. Dehydration-responsive element binding protein 1
(DREB1)/C-repeat binding factor (CBF) that function in the cold stress response, and
DREB2 regulons acts in ABA-independent gene expression for response to heat and
1

osmotic stress, whereas the ABA-responsive element (ABRE) binding protein
(AREB)/ABRE binding factor (ABF) regulon functions in ABA-dependent gene
expression under osmotic stress conditions. In addition to these major pathways, other
regulons, including the NAC and MYB/MYC regulons, are involved in abiotic stress-
responsive gene expression.
Recent studies evidence that TFs are powerful tools to engineer enhanced stress
tolerance in plants. In this chapter, we will outline the major yield-limiting abiotic
stresses faced by crop plants and their adaptive mechanisms at the molecular level. A
major knowledge on molecular mechanisms that occur in stress condition are the one
way pass for the improvement of stress tolerance in crop plants.
Key words: transcriptional factors, abiotic stress, cold, drought, salt, AP2/ERF,
MYB/MYC, NAC
2

1 Transcription Factors Activate Stress Responsive Genes
Organisms rely on regulatory networks to integrate and process signals from various
sources and to orchestrate the transcription of genes controlling a range of cellular
processes. Importantly, changes in the architecture of these regulatory networks play a
significant role in survival or adaptation of organisms to changing environments (Pérez
and Groisman, 2009).
Tolerance or susceptibility to abiotic stresses, for example, are a very complex
phenomenon, both because stress may occur at multiple stages of plant development
and more than one stress simultaneously affects the plant. Therefore, the perception of
abiotic stresses and signal transduction to switch on adaptive responses are critical steps
in determining the survival and reproduction of plants exposed to adverse environments
(Chinnusamy et al. 2004).
During the past few years, transcriptome analysis has indicated that distinct
environmental stresses induce similar responses. Overlap between stress responses can
explain the phenomenon known as cross-tolerance, a capability to limit collateral
damage inflicted by other stresses accompanying the primary stress.
The functions of stress-inducible genes involved in stress response and tolerance are
classified into two major groups, one involved in stress tolerance, such as chaperones,
late embryogenesis abundant (LEA) proteins, enzymes for osmolyte biosynthesis and
detoxification enzymes and the other in gene expression and signal transduction in
abiotic stress response, such as protein kinases, transcription factors (TFs) and enzymes
in phospholipids metabolism (Yamaguchi-Shinozaki and Shinozaki 2005). In addition,
new transcripts are synthesised and within a few hours a steady-state level of stress
adaptation has been reached. Gene induction occurs primarily at transcriptional level,
the temporal and spatial regulation of stress-induced expression patters is an important
part of the plant response (Rushton and Somssich 1998).
In the past two decades, important advances have been made in order to understand
transcriptional changes induced by environmental constraints and to identify signalling
proteins and TFs which regulate the stress-induced gene expression. In general, the
transcriptional regulation of genes is directly controlled by a network of TFs and
transcription factor binding sites (TFBS) (Chaves and Oliveira 2004).
The families of the plant-specific TFs are defined by their characteristic TFBS. After
completion of the Arabidopsis thaliana (Arabidopsis) genome sequence, approximately
3
1500 probable TFs belonging to approximately 30 TF families were identified.
Approximately half of the families were considered plant-specific, because no members
have been identified in other eukaryotic lineages. In particular, a recent plant TFBS
three-dimensional structures analysis done by Yamasaki and co-workers (2013) has
demonstrated that they are likely to have originated from endonucleases associated with
transposable elements. After the TFBSs have been established in unicellular eukaryotes,
they experienced extensive plant-specific expansion, by acquiring new functions and
complexity.
Indeed, responses to abiotic stresses are not linear pathways, but are complicated
integrated circuits involving the interaction of additional cofactors and/or signalling
molecules to coordinate a specified response to a given stimulus. The regulation of
these responses requires proteins operating in the signal transduction pathways, such as
TFs, which regulate gene expression by binding to specific cis-elements in the promoter
regions of target genes and forming a transcriptional initiation complex on the TATA
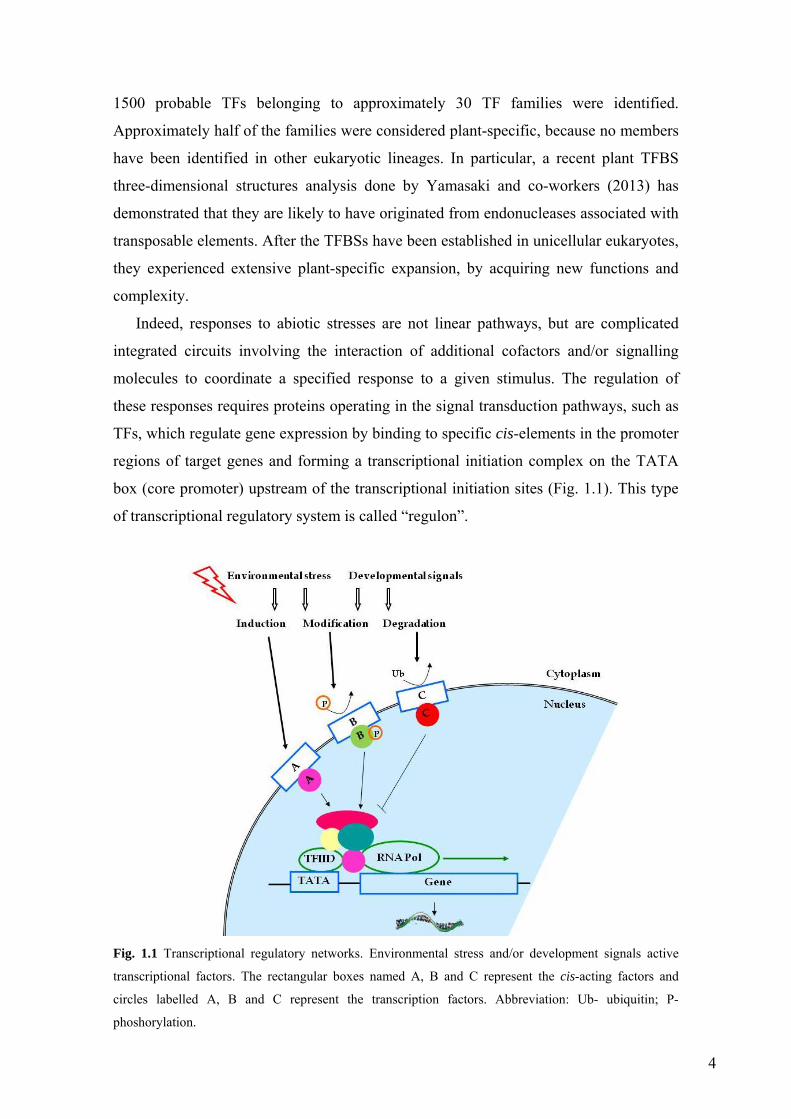
box (core promoter) upstream of the transcriptional initiation sites (Fig. 1.1). This type
of transcriptional regulatory system is called “regulon”.
Fig. 1.1 Transcriptional regulatory networks. Environmental stress and/or development signals active
transcriptional factors. The rectangular boxes named A, B and C represent the cis-acting factors and
circles labelled A, B and C represent the transcription factors. Abbreviation: Ub- ubiquitin; P-
phoshorylation.
4
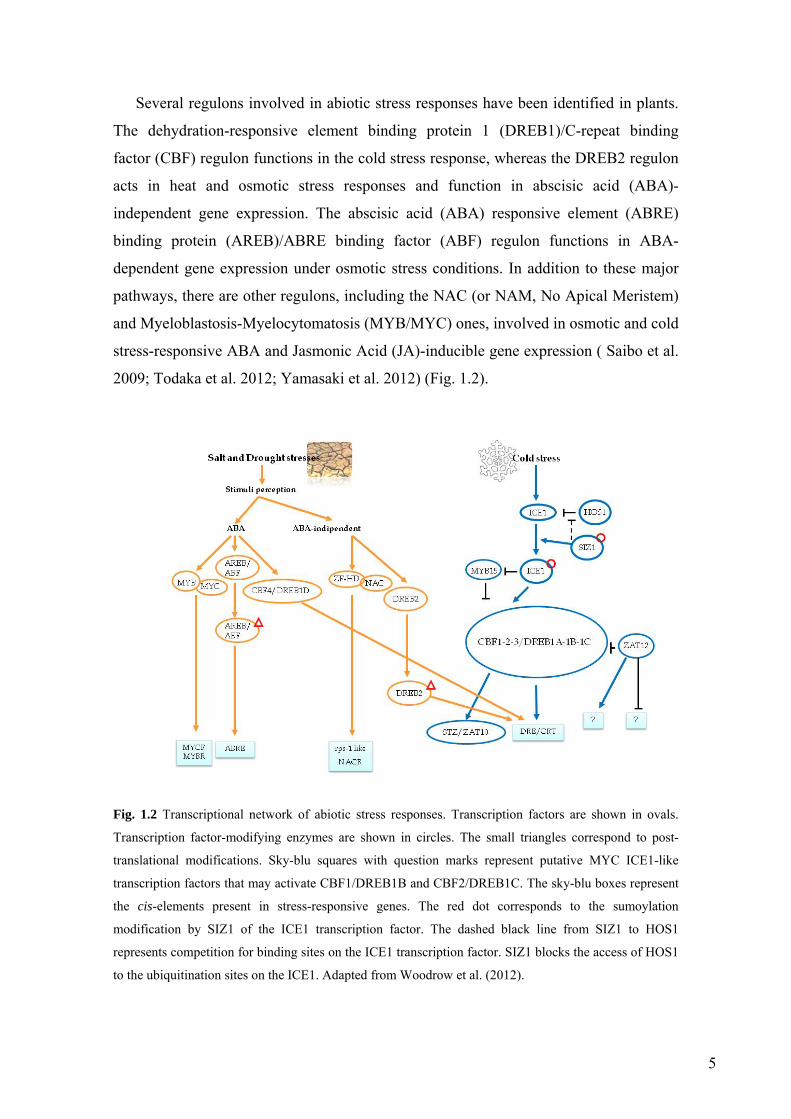
Several regulons involved in abiotic stress responses have been identified in plants.
The dehydration-responsive element binding protein 1 (DREB1)/C-repeat binding
factor (CBF) regulon functions in the cold stress response, whereas the DREB2 regulon
acts in heat and osmotic stress responses and function in abscisic acid (ABA)-
independent gene expression. The abscisic acid (ABA) responsive element (ABRE)
binding protein (AREB)/ABRE binding factor (ABF) regulon functions in ABA-
dependent gene expression under osmotic stress conditions. In addition to these major
pathways, there are other regulons, including the NAC (or NAM, No Apical Meristem)
and Myeloblastosis-Myelocytomatosis (MYB/MYC) ones, involved in osmotic and cold
stress-responsive ABA and Jasmonic Acid (JA)-inducible gene expression ( Saibo et al.
2009; Todaka et al. 2012; Yamasaki et al. 2012) (Fig. 1.2).
Fig. 1.2 Transcriptional network of abiotic stress responses. Transcription factors are shown in ovals.
Transcription factor-modifying enzymes are shown in circles. The small triangles correspond to post-
translational modifications. Sky-blu squares with question marks represent putative MYC ICE1-like
transcription factors that may activate CBF1/DREB1B and CBF2/DREB1C. The sky-blu boxes represent
the cis-elements present in stress-responsive genes. The red dot corresponds to the sumoylation
modification by SIZ1 of the ICE1 transcription factor. The dashed black line from SIZ1 to HOS1
represents competition for binding sites on the ICE1 transcription factor. SIZ1 blocks the access of HOS1
to the ubiquitination sites on the ICE1. Adapted from Woodrow et al. (2012).
5

We will discuss the different TFs and their transcriptional gene networks by
focusing on the main regulons and therefore on the set of their target genes in order to
understand the architecture of regulatory networks, and inferring how these networks
change upon exposures to abiotic stress.
2 APETALA 2/Ethylene-Responsive Element Binding Factor
One class of protein that is unique in plants and plays a vital role in biotic and abiotic
stress response is the APETALA 2/ethylene-responsive element binding factor
(AP2/ERF) (Agarwal et al. 2006). The AP2/ERF proteins family is a large group of TFs
containing AP2/ERF-type DNA-binding motif (DBDs). Four major subfamilies belong
to AP2/ERF family: the AP2, Related to AB13/VP1 (RAV), ERF and dehydration-
responsive element-binding protein (DREB) subfamilies (Sakuma et al. 2002).
All AP2/ERF family members share a conserved DBD of approximately 60 amino
acids that binds to two cis-elements, the GCC box, found in many PR (Pathogens
Related) gene promoters conferring ethylene responsiveness (Gu et al. 2000), and the C
repeat CRT/dehydration responsive element (DRE) motif involved in the expression of
cold and dehydration responsive genes (Agarwal et al. 2006).
DNA sequences recognized and bound by DBDs differ among the subfamilies and,
the amino acid sequences are conserved within a family. Members of a family encode
related proteins that share a homologous DNA binding domain. Furthermore, individual
members of the same family respond differently in response to different stress stimuli
(Akhtar et al. 2012).
Deletion and mutational analyses of promoter genes responsive to light, ultraviolet
radiation and JA have shown that mutations in G-box compromise the ability of each
promoter to respond to its respective stimulus. The expression of these genes require at
least one cis-acting element in addition to the G-box for appropriate transcriptional
activation within promoter regions.
An AP2/ERF binding domain was found for the first time in the Arabidopsis
homeotic gene APETALA 2 (Jofuku et al. 1994), and a similar domain was found in
tobacco (Nicotiana tabacum) ethylene-responsive element binding proteins (EREBPs)
(Ohme-Takagi et al. 1995). Sequences homologous to the AP2/ERF domain have been
found in bacterial and viral endonucleases, and thus, it has been hypothesised that the
6

AP2/ERF domain was transferred from a cyanobacterium by endosymbiosis or from a
bacterium or a virus by other lateral gene transfer events (Magnani and Sjölander 2004).
An NMR-based structural analysis of the Arabidopsis ERF1 protein in complex with
a target DNA molecule revealed that the AP2/ERF domain contains an N-terminal
three-strand antiparallel β-sheet that recognises a target sequence connected by loops, as
well as a C-terminal α-helix, packed approximately parallel to each other (Allen et al.
1998; Fig. 1.3).
Fig. 1.3 AP2; DNA-binding domain found in transcription regulators in plants such as APETALA2 and
ethylene responsive element binding protein (EREBP). In EREBPs the domain specifically binds to the
11bp GCC box of the ethylene response element (ERE), a promotor element essential for ethylene
responsiveness. EREBPs and the C-repeat binding factor CBF1, which is involved in stress response,
contain a single copy of the AP2 domain (PubMed 9736626; Modified from Allen et al. 1998).
DREBs, also referred as CBF (C-repeat binding factor) proteins, belong to ERF
subfamily and play a crucial role in plant response to abiotic stresses. DREBs are
important TFs which induce a set of abiotic stress-related genes and confer stress
resistance to plants. As previously mentioned, DREB TFs could be dichotomized as
DREB1 and DREB2, which are involved in two separate signal transduction pathways
under low temperature and dehydration, respectively.
The AP2 family genes possess two repeats of the AP2/ERF domain (14 members in
Arabidopsis) and the ERF family proteins contain a single AP2/ERF domain, whereas
RAV subfamily proteins (6 members) have an additional B3 DNA binding domain
(Kagaya et al. 1999; Rashid et al. 2012).
In the ERF/AP2 domain, two amino acids, valine (14) and glutamic acid (19) play a
crucial role in the determination of DNA-binding specificity of DRE cis-elements
7

(Sakuma et al. 2002). Other amino acids that facilitate direct contact with DNA for
DNA binding activity are arginine (6), arginine (8), trytophan (10), glutamic acid (16),
arginine (25) and tryptophan (27) (Allen et al. 1998). A conserved Ser/Thr-rich region
that is adjacent to the ERF/AP2 domain is considered to be responsible for
phosphorylation of DREB proteins, while a conserved alanine (37) is capable of binding
to the DRE element (Liu et al. 1998).
The entry of DREB proteins is mediated by one or two nuclear localization signal
(NLS) (Akhtar et al. 2012).
The DREB1/CBF1-type proteins have a basic-aminoacid-rich stretch having a
consensus sequence PKRPAGRTKFRETRHP. This motif distinguishes these proteins
from other ERF/AP2 proteins.
Ethylene-responsive TF RAV1 binds specifically to bipartite recognition sequences
composed of two unrelated motifs, 5’-CAACA-3’ and 5’-CACCTG-3’. This TF
contains two distinct DNA-binding domains (Kagaya et al. 1999): one is located in the
N-terminal region and binds the 5’-CAACA-3’ motif and the other one is located in the
C-terminal region and binds the 5’-CACCTG-3’ motif (Kagaya et al. 1999). RAV1, in
fact, interacts in a selective and non-covalent manner with the two previously
mentioned DNA sequences, in order to modulate transcription. RAV1 expression is
down-regulated by brassinosteroid and zeatin (Hu et al. 2004). RAV proteins have
important roles in several physiological process such as lateral root development and
leaf and flower development (Ikeda et al. 2009). Moreover they are involved in plant
response to both biotic and abiotic stresses. For example, RAV1 is expressed in pepper
and in Brassica napus where it has functional roles in bacterial disease resistance and
drought and salt stress tolerance (Shon et al. 2006; Zhuang et al. 2011). In Arabidopsis,
members of RAV family as RAV1 and RAV2 exhibit transcriptional repressive activity
when they bind a plant specific repression domain that contains the conserved amino
acid sequence LXLXL (Ikeda et al. 2009).
AP2 domain includes two regions: YRG and RAYD. The first region (YRG
element) is constituted of 20 amino acids rich in basic and hydrophobic amino acids in
the N-terminal stretch. Due to its basic nature, it is proposed to have a role in DNA-
binding activity. The second region, RAYD element is approximately 40 amino acids.
RAYD element also regulates the special binding activity of DREB proteins by
influencing the conformation of the YRG element (Okamuro et al. 1997).
8

The DSAW motif (A (A/V)xxA (A/V)xxF,) at the end of the ERF/AP2 domain and
LWSY motif with a consensus sequence of
(L/Y)(L/Y)x(N/S)(M/L)A(E/Q)G(M/L)(L/M)xxPP at the end of the C-terminal, are
conserved in most of the DREB1-type proteins (Fig. 1.4). A 18-amino-acid stretch in
the C-terminal region is able to form an amphipathic α-helix that could mediate protein-
protein interaction.
Eudicot plants have LLxNM at the beginning of the domain, while the monocot
CBF homologues have YYxSL (Akhtar et al. 2012).
Fig. 1.4 Schematic structure of typical DREB1/CBF domains.
2.1 Cis-Acting Regulatory Element
Various TFs interact with cis-acting elements in promoter regions and form a
transcriptional initiation complex on the TATA box (core promoter) upstream of
transcriptional initiation sites.
In Arabidopsis, promoter analysis of RD29A/COR78/LT178 genes induced by
drought, high-salinity and cold highlighted a 9 bp conserved sequence (TACCGA-CAT)
(Akhtar et al. 2012).
Similar cis-acting elements, named C-repeat (CRT) and low-temperature-responsive
element (LTRE), both containing an A/GCCGAC motif that forms the core of the DRE
sequence, regulate cold-inducible promoters (Stockinger et al. 1997; Thomashow 1999).
A conserved sequence, PyACGTGGC, was found in the promoter regions of several
ABA-inducible genes. This sequence, first identified as a cis-acting element named
ABA responsive element (ABRE) in the wheat Em gene (Guiltinan et al. 1990) and then
in the rice RAB16 gene, is expressed in both dehydrated vegetative tissues and
maturating seeds (Mundy et al. 1990).
Detailed analyses of the binding preferences of DREB1A and DREB2A demonstrate
that these TFs have the highest affinity for the DRE core sequence A/GCCGAC
9

(Sakuma et al. 2002). Furthermore, DREB1A binds preferentially A/GCCGACNT,
while DREB2A binds preferentially ACCGAC, as confirmed by the analysis of
upregulated genes promoters in transgenic plants overexpressing DREBs (Sakuma et al.
2006).
The expression of DREB2A and its homolog DREB2B are induced by dehydration
and high salt stress (Liu et al. 1998). DREB2A and DREB2B proteins require post-
translational modification for their activation by a mechanism that has not yet been
unravelled. It has been found in some studies that the expression of DREB genes is
induced by abiotic stresses, but at different time periods. AtDREB2A is induced within
10 min under drought or salt stress (250 mM NaCl ) (Liu et al. 1998). In rice, Os-
DREB2A is induced within 24 h after dehydration and salt stress (250 mM NaCl) and it
responds faintly to ABA and cold stress (Dubouzet et al. 2003).
Presently, there is not much information available on the tissue-specific expression
of DREBs. The transcription of GmDREBa and GmDREBb is induced by cold, drought
and salt in leaves of soybean seedlings. The expression of GmDREBc is not significant
in leaves but shows high expression levels in roots under drought, salt, low temperature
and ABA treatments (Li et al. 2005; Chen et al. 2007).
Five ERF subfamily proteins display the highest affinity for the AGCCGCC
sequence, which is the core sequence of the ERE (Fujimoto et al. 2000). A DREB2 (A-
2) subgroup protein from barley (HvDRF1) preferentially bounds TT/AACCGCCTT
sequence (Xue et al. 2004).
Based on the various studies summarized above, it is clear that the DREB and ERF
proteins are important TFs involved in the regulation of abiotic stress-related genes and
play a critical role in imparting stress endurance to plants.
3 The MYC /MYB Transcriptional Factors
MYC/MYB TFs participate in the ABA-dependent stress-signalling pathway up-
regulating the abiotic stress responsive genes.
The N-terminal region of the plant MYC-like basic helix–loop–helix TFs (bHLH
TFs) contains two domains, one around 190 amino acids is located at the N-terminus
that comprises a domain of interaction with MYB-like TFs. This interaction domain is
an activation domain responsible for transactivation. Indeed, the activation domain
forms a protein-binding surface that binds the RNA polymerase II machinery and
10

afterwards initiates transcription (Pattanaik et al. 2008). Another feature of MYC-like
bHLH TFs is the presence of multiple protein dimer interfaces. One of these interfaces
is an ACT-like domain located at the C-terminus.
The bHLH proteins belong to a large family of transcriptional regulators and are
characterized by the signature bHLH domain comprising approximately 60 amino acids.
The critical region of the domain binds to an E-box (CANNTG) sequence and the HLH
region is involved in homo- and heterodimerization (Li et al. 2006).
MYB domain is usually composed of one to three imperfect repeats, each with about
52 amino acid residues with a conformation helix-turn-helix that intercalates the DNA
in the major groove (Yanhui et al. 2006). Plant MYB proteins are divided into three
major groups: group I (R2R3-MYB) that shows two adjacent repeats; group II
(R1R2R3-MYB) with three adjacent repeats; and group III (MYB-related proteins)
usually containing a single MYB repeat (Stracke et al. 2001; Lata et al. 2011).
MYB TFs function as important components in the complex signalling pathways of
plant responses to abiotic stress (i.e. drought, cold, light, and nutrient deficiency) and
they also play important roles in several developmental and physiological processes (i.e.
cell morphogenesis, meristem formation and floral and seed development, cell cycle
control and hormone signalling) (Kirik et al., 1998; Higginson et al., 2003; Araki et al.,
2004; Newman et al., 2004).
MYC and MYB TFs accumulation occurs only after an ABA concentration increase.
MYB and MYC, together with a family of transcriptional repressors (Cys2/His2-type
zinc-finger proteins) are also involved in the ABA response to water deficit.
MYC, in cooperation with MYB is involved in the regulation of ABA-inducible
genes under drought stress conditions, in fact expression of the drought-inducible gene
Responsive to Dehydration 22 (RD22) from Arabidopsis was found to be induced by
ABA. The promoter region of RD22 contains MYC (CANNTG) and MYB
(C/TAACNA/G) cis-element recognition sites. Over-expression of these TFs results in
enhanced sensitivity to ABA and drought tolerance (Abe et al. 2003).
According to Denekamp and Smeekens (2003), AtMYB102 integrates dehydration,
osmotic, or salinity stress, ABA and wound-signalling pathways. AtMYB60 and
AtMYB61 are, respectively, involved in stomata light-induced opening (Cominelli et
al., 2005) and stomata dark-induced closure (Liang et al., 2005). These TFs can operate
as negative regulator of blue light-mediated photomorphogenic growth and blue- and
far-red-light regulated gene expression (Yadav et al. 2005). Moreover, recent studies
11

have shown that AtMYB15 expression is up-regulated by cold and salt stresses
(Agarwal et al., 2006). In Arabidopsis, ICE1 (Inducer of CBF Expression1), a MYC-
type bHLH TF, can bind to MYC recognition elements in the CBF3 promoter, allowing
CBF3 expression during cold acclimation. ICE1 is constitutively expressed and
localized in the nucleus, but it induces expression of CBFs only under cold stress (Fig.
1.5). This suggests that a cold stress-induced post-translational modification is
necessary for ICE1 to activate downstream genes in plants (Chinnusamy et al. 2003).
Fig. 1.5 Cold-responsive transcriptional network in Arabidopsis. Constitutive expressed ICE1 is actived
by cold stress after sumoylation and phosphorylation post-transcriptional modifications. Active ICE1
induces the transcription of CBFs. Broken arrows indicate post-translational regulation; solid arrows
indicate activation.
4 NAC Transcriptional Factors
The plant-specific NAC [No Apical Meristem (NAM); Arabidopsis transcription
activation factor (ATAF); Cup-shaped cotyledon (CUC)] proteins constitute a major TF
superfamily renowned mostly for their important functions in plant development and
stress responses (Fujita et al. 2004; Nakashima et al. 2011; Puranik et al. 2011). NAC
TFs were originally identified due to their role in diverse developmental processes
12

(Souer et al. 1996); a growing body of evidence suggests that they also play a
significant role in both biotic and abiotic stress responses. These hypotheses have been
made mainly based on observations of rapid induction of NAC genes in response to
stress stimuli and enhanced stress tolerance in transgenic plants over-expressing these
genes (Hu et al. 2006; Wu et al. 2009; Zheng et al. 2009; Jeong et al. 2010; Takasaki et
al. 2010; Hao et al. 2011; Liu et al. 2011; Xue et al. 2011). Whole-genome expression
profiling studies in Arabidopsis have shown that most NAC genes are induced by at
least one type of abiotic stress signal, such as salinity, drought, cold, or ABA (Jensen et
al. 2010).
In addition to rapid gene expression in response to stress stimuli, NAC transcription
factors can be regulated at the mRNA or/and protein level.
NAC family members have been identified and characterized in model plants like
Arabidopsis (Ooka et al. 2003; Tran et al. 2004; Fujita et al. 2004), crops such as rice
(Oryza sativa) (Ooka et al. 2003; Hu et al. 2006; Hu et al. 2008), soybean (Glycine max)
(Le et al. 2011; Hao et al. 2011) and wheat (Triticum sp.) (Kawaura et al. 2008; Xia et
al. 2010), as well as in some model tree species like poplar (Populus trichocarpa) (Hu
et al. 2010), citrus (Citrus sp.) (Liu et al. 2009; de Oliveira et al. 2011) and grape (Vitis
vinifera) (Wang et al. 2013). The availability of several complete plant genomic
sequences has allowed the identification of 117 NAC genes in Arabidopsis (Jensen et al.
2010), 151 in rice (Nuruzzaman et al. 2010), 79 in grape (Vitis vinifera), 163 in poplar
and 152 each in soybean and tobacco (Nicotiana tabacum) (Le et al. 2011; Hu et al.
2010; Nuruzzaman et al. 2010; Rushton et al. 2008) which makes them one of the
largest family of TFs in plants (Puranik et al. 2012).
4.1 NAC TFs Role in Abiotic Stress Response
NAC TFs are implicated in abiotic stress responses such as salt stress and ABA (He et
al. 2005), and plays a role in salt-promoted senescence (Balazadeh et al. 2010).
Some of the well-characterized abiotic stress-associated NAC TFs belong to sub-
group III-3, also referred to as ATAF (Jensen et al. 2010) or SNAC (Stress-responsive
NAC) sub-group (Nuruzzaman et al. 2010). Members of this sub-group, ANAC019,
ANAC055, and RESPONSIVE TO DEHYDRATION 26 (RD26) (ANAC072), are
induced by drought, high salinity or cold stress, and are under the control of the central
ABA perception and signalling network (Tran et al. 2004; Jensen et al. 2010). The over-
13

expression of these genes induced up-regulation of several stress-inducible genes and
conferred drought tolerance to the transformed plants (Tran et al. 2004; Nakai et al.
2013). In fact, the TFs were shown to bind the promoter of EARLY RESPONSIVE TO
DEHYDRATION STRESS 1 (ERD1) (Tran et al. 2004; Nakashima et al. 2012) which
is not only induced by dehydration but is also up-regulated during senescence and dark-
induced etiolation.
Up-regulation of the ERD1 gene depends on co-overexpression of both NAC TFs
and the ZF homeodomain transcriptional activator ZFHD, suggesting the existence of
cooperative regulation between these two cis-acting elements (Lindemose et al. 2013).
Overexpression of ATAF1 also affects plant tolerance to drought, although
contradicting reports suggest positive and/or negative regulatory effects (Lu et al. 2007;
Wu et al. 2009). Therefore, subgroup III-3 NAC TFs play important roles in abiotic
stress perception and individual NAC members display functional overlap in stress
responses (Fig. 1.6).
Fig. 1.6 A model representing the regulation of NAC TFs at different levels during stress. Upstream TFs
like DREBs, WRKYs or ABREs may bind to stress-related cis-regulatory elements in the promoter of
regulated NAC genes and influence their transcription. Post-transcriptional control of pre-NAC mRNAs
by miRNA164 and alternate splicing may also regulate their expression. The translated protein may be
subjected to activation by phosphorylation or glycosylation. Upon their nuclear import, the activated
NAC proteins homo- or heterodimerize or bind to promoters of their target genes to control their
14

expression. Abbreviations: AREB, ABA-responsive element binding protein; DREB, Dehydration
responsive element binding protein; G, N-acetyl glucosamine; miRNA, microRNA; and P, phosphate.
Adapted from Puranik et al. (2012).
Expression of stress-responsive NACs may be tightly regulated by several stress-
responsive cis-acting elements contained in the promoter region. These include the
ABREs (ABA-responsive elements), DREs (Dehydration Responsive Element), LTREs
(Low-temperature Responsive Elements), MYB (Myeloblastosis) and MYC
(Myelocytomatosis) binding sites, W-Box, jasmonic acid responsive element and
salicylic acid responsive element (Nakashima et al. 2012).
4.2 Structural Features of NAC TFs
The DNA sequences that are recognized show weak differences among the family
members and possess palindromic properties with different distances, such as
AN5TCN7ACACGCATGT with a consecutive pseudo-palindromic core (underlined),
for abscisic-acid-responsive NAC (ANAC) proteins (Tran et al. 2004) and
(T/A)NN(C/T)(T/C/G)TN7A(A/C)GN(A/C/T)(A/T) with a pseudo-palindromic
sequence with a seven-base gap for SND1 and VND7 (Yamasaki et al. 2012).
NAC domain proteins share a conserved N-terminal DNA binding domain of
approximately 150 amino acids and originally found in four genes (NAM, ATAF1,
ATAF2 and CUC2) (Christianson et al. 2010). The N-terminal domain is further
divided into five conserved sub-domains: this region holds DNA-binding (DB) ability
and/or is responsible for protein-binding and dimerization (Fig. 1.7). The C-terminal
region is more divergent and serves as a potential transcriptional regulatory (TR)
domain which has either activator or repressor function and has sometimes protein
binding activity (Fig. 1.7). In some cases, the NAC domain may contain a negative
regulatory domain and the C-terminal may have a transmembrane motif (Olsen et al.
2005).
15

Fig. 1.7 Schematic structure of a typical NAC domain protein with a highly conserved NAC domain at
the N-terminal which is further divided into five conserved subdomains (indicated with the letters A-E).
This region holds DNA-binding (DB) ability and/or is responsible for protein-binding and dimerization.
Secondary elements are shown at the top of subdomains (A and B α-helices; C-E β-strands). The C-
terminal region is more diverged and serves as a potential transcriptional regulatory (TR) domain
(activator or repressor) may occasionally have protein binding activity.
Variations from typical NAC proteins include those encoding only the NAC
domains, two tandemly repeated NAC domains, a N-terminally extended (NTE) region
preceding the conserved NAC domain, and VOZ proteins, where the NAC domain lies
at the C-terminal and the TR domain lies at the N-terminal with a zinc finger (ZF) motif
functioning as a conserved DB domain (Puranik et al. 2012). These remarkable
diversification across plants reflects their numerous functions.
5 WRKY Transcriptional Factors
Members of plant WRKY TFs families are among the best-characterized classes of
plant TFs. They participate in various disease resistance (Pandey et al. 2009) and
several developmental and physiological processes (i.e. embryogenesis, seed coat and
trichome development, hormone signalling, leaf senescence) (Song et al. 2010; Zhou et
al. 2011). Furthermore, WRKY TFs function as important components in the complex
signalling networks in plant responses to abiotic stresses, such as drought, cold and
nutrient deficiency.
WRKY proteins belong to the WRKY-GCM1 superfamily of ZF TFs that evolved
from Mutator or Mutator-like (Mule) transposases (Marquez et al. 2010). WRKY
proteins are a class of DNA-binding proteins that recognize the TTGAC(C/T) W-box
elements found in the promoters of a large number of plant defense-related genes. The
DNA binding domain shows an almost invariant WRKY amino acid sequence at the N-
terminus (Eulgem et al. 2000). Only in a few WRKY proteins, the WRKY amino acid
16

sequences have been replaced by WRRY, WSKY, WKRY, WVKY or WKKY (Xie et
al. 2005).
The WRKY TFs are classified into three groups according to the number of WRKY
domains: group I has two domains, group II has one domain and group III has one
domain plus one ZF structure (Cys2-His/Cys or Cys2-His2 ZF motif) (Chen et al.
2012). Moreover, group II genes, being not monophyletic, can be further divided into
IIa, IIb, IIc, IId and IIe based on the primary amino acid sequence (Zhang et al. 2005;
Rushton et al. 2008).
5.1 The WRKY Domain and the W Box
The WRKY protein family shows a highly conserved 60 amino acid long peptide
region, termed WRKY domain (Maeo et al. 2001; Ciolkowski et al. 2008). In this
domain an invariant heptapeptide WRKYGQK at N-terminus that shows slight
variations in a few WRKY proteins (Yang et al. 2009), partly protrudes from one
surface of the protein enabling contacts with the DNA. In fact, it has been well
established that the b-strand containing the WRKYGQK motif makes contact with an
approximately 6-bp region, and this is consistent with the length of the W box (Rushton
et al. 2010).
At C-terminus of the WRKY domain there is also a novel ZF-like motif that
contributes to the largely strength of DNA binding and probably the N-terminal domain
might participate in the binding process, increasing the affinity or specificity of these
proteins for their target sites (Fig. 1.8). Therefore not the WRKYGQK residues as well
as the ZF motif are required for proper DNA binding of the protein (Rushton et al.
2010).
Fig. 1.8 Crystal structure of Arbidopsis C-Terminal WRKY domain (PubMed 118137307; adapted from
Eulgem et al. 2000).
17

The conservation of the WRKY domain is mirrored by a remarkable conservation of
its cognate binding site, the W box (TTGACC/T) (Eulgem et al. 2000). W box
represents the minimal consensus required for specific DNA binding as well established
by gel shift experiments, random binding site selection, one-hybrid system assays and
co-transfection assays performed with many different WRKY proteins (Ciolkowski et
al. 2008). There are a few reports of WRKY proteins binding to non-W box sequences
as the OsWRKY13 can also bind to the PRE4 element (TGCGCTT) (Cai et al. 2008) or
the SUSIBA2 (a sucrose-regulated barley WRKY transcription factor) that, in addition
to W boxes, can bind a sugar-responsive (SURE) element
(TAAAGATTACTAATAGGAA) (Sun et al. 2003).
5.2 Role of WRKY Genes Under Abiotic Stresses
The WRKY TFs are induced in response to different abiotic stresses and so they could
be an useful tool for conferring stress tolerance. The role of WRKY TFs under
environmental stimuli is not clearly unravelled. Microarray analysis on Arabidopsis
transcriptome under salt stress reveals up-regulation of 18 WRKY and down-regulation
of 8 WRKY genes (Jiang et al. 2006). The WRKY TFs can be induced by ABA, NaCl,
PEG, cold and heat treatment (Xie et al. 2005; Ramamoorthy et al. 2008) (Fig. 1.9).
Fig. 1.9 Role of Arabidopsis WRKY transcriptional factors in abiotic stress response. ABA signalling
pathway leads to the activation of ABL1 expression. This latter stimulates the expression of WRKY
18

genes, which differentially regulate the downstream W-box containing genes to regulate abiotic responses
and ABA signalling.
WRKY TFs regulate the plant stress response in two ways: by interacting with other
TFs or directly regulating some stress-responsive genes. WRKY TFs are also in turn
early response genes, and their target genes encoding enzymes such as chitinase,
glutathione S-transferase, and strictosidine synthase are late response genes (Cheong et
al. 2002) (Fig. 1.10).
Fig. 1.10 Role of Arabidopsis WRKY transcriptional factors in wounding response pathways. After
activation of MAPK phosphorylation cascade, WRKY3 regulate the expression of WRKY genes. The
WRKY6 actives the expression of late genes, such as chitinase and glutathione S-transferase which allow
plants to defence.
GmWRKY21 and GmWRKY54 transgenic Arabidopsis plants show a good
tolerance to environmental stimuli such as cold, salt and drought stresses. This ability is
probably due to WRKY TFs which regulate TFs like DREB2A and STZ/Zat10 (Wei et
al. 2008; Zhou et al. 2008). This example shows that WRKY factors are integrated part
of the signalling processes associated with TFs regulation of gene expression under
abiotic stresses.
6 Cys2His2 Zinc Finger (C2H2 ZF) TFs
19

Zinc Finger proteins (ZFPs) are one of the most important TFs found abundantly in
plants. They contain sequence motifs in which cysteines and/or histidines coordinate
zinc atom(s) forming local peptide structures required for their specific functions (Singh
et al. 2010). Cys2/His2 (C2H2)-type ZFPs containing the ERF-associated amphiphilic
repression (EAR) transcriptional repressor domain, play a key role in regulating the
defense responses of plants to biotic and abiotic stress conditions (Singh et al. 2010).
Several ZAT proteins, a family of putative ZF-containing TFs, are implicated in
abiotic stress responses. This is confirmed by several studies on plant response to
abiotic stresses. For example, in Arabidopsis, as reported by Lee et al. (2002),
Zat10/STZ can repress the RD29A gene, and constitutive expression of Zat10 in
Arabidopsis enhance plants tolerance to salinity, heat and osmotic stress. By contrast,
over-expression of Zat7 also results in increased tolerance to salinity stress (Ciftci-
Yilmaz et al. 2007). ZAT12 is recognized as a component in the oxidative stress
response signalling network of Arabidopsis (Rizhsky et al. 2004). During oxidative
stress, ZAT12 promotes expression of other TFs and the up-regulation of cytosolic
ascorbate peroxidase 1, a key enzyme in the removal of H2O2. Accumulation of ROS is
a common consequence of biotic and abiotic stresses, including oxygen deprivation.
ZAT12 transcript levels were significantly elevated in response to hypoxia and anoxia
in several independent analyses (Branco-Price et al. 2005), indicating involvement of
this potentially redox-regulated factor.
AZF1 and AZF2 are induced by osmotic stress and ABA, and over-expression of
these genes compromise plant growth and viability. AZF1 and AZF2 repressed a set of
genes that were down-regulated by osmotic stress and ABA treatment and many auxin
(IAA)-responsive genes, and AZF1 and AZF2 bound the promoter of two of these,
SAUR63 and SAUR20, in electrophoretic mobility shift assays (EMSAs). This
suggested that AZF1 and AZF2 function as transcriptional repressors to inhibit plant
growth (Kodaira et al. 2011).
Recently the C2H2 ZF TF DROUGHT AND SALT TOLERANCE (DST) is shown to
control stomatal aperture under drought and salt stress in rice by direct modulation of
genes related to H2O2 homeostasis (Huang et al. 2009).
6.1 C2H2 ZF DBD and Overall Structure of Zat7
20

The 168 amino acid residues C2H2-type ZF TF ZAT7 contain two C2H2 ZFs both with
a conserved plant-specific DNA-binding QALGGH motif that plays a critical role in
DNA binding activity (Ciftci-Yilmaz et al. 2008). The C2H2-motif, constituted by
approximately 30 amino acid residues, has two conserved Cys and His bound to one
zinc ion (Willemijn et al. 2005; Fig. 1.11). C2H2-type ZFs (Ciftci-Yilmaz et al. 2008)
structure shows two fingers, in ZAT7, each formed by two β strands and one α helix
that are separated by a long spacer predicted to be flexible. Moreover, ZAT7 protein
contains two EAR repression domains (Lindemose et al. 2013).
Fig. 1.11 Schematic structure of a Cys2-His2 type ZF. This Zinc Finger motif consists of a simple two β
strands and one α helix separated by a long spacer. Amino acid residues within the α-helix recognize the
target DNA sequence.
7 Conclusion and future perspectives
Extremes of environmental conditions, such as, drought, salinity, heat, cold and
wounding, induce stress and decrease plants growth and crops productivity. A large
number of genes have been reported as abiotic stress responsive genes and the
expression of these genes is crucial for providing stress tolerance to plants. Thanks to
the use of diverse technologies and approaches, including physiology, chemical,
genetic, molecular computational and informational biology, the field of plant signal
transduction and gene regulation has shown rapid progress (Chen et al. 2012), which
helps us to move forward to address and to unravel several important unresolved
questions on the complex mechanisms of plant responses to abiotic stresses and to
21

develop tolerant cultivars better able to cope with the increasing environmental
constraints.
This book chapter summarizes the structure and the role of important plant TFs such as
AP2/ERF, DREBs/CBF, MYC/MYB, and NAC that regulate various stress responsive
gene expression.
TFs play a crucial role in integrating the response among multiple stresses,
providing tolerance generally in both ABA dependent and -independent manner and
through respective cis-elements and DNA binding domains. Their further
characterization and in particular their functional analysis will thus provide more
information on the intricate regulatory networks involved in abiotic stress responses and
the cross-talk between different signalling pathways during stress adaptation. The
ability of TFs to control the expression of stress responsive genes may therefore prove
to be a practical way of engineering multiple stress tolerance into crops. Therefore,
considering TFs as candidate genes in breeding and other crop improvement programs
will improve the understanding of abiotic stress related signal transduction events and
eventually will improve and facilitate the development of crop varieties superior in
stress tolerance to drought, salinity, heat and cold.
References
Abe H, Urao T, Ito T, Seki M, Shinozaki K and Yamaguchi-Shinozaki K (2003)
Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional
activators in abscissic acid signalling. The Plant Cell 15:63-78
Agarwal M, Hao Y, Kapoor A, Dong CH, Fuji H, Zheng X and Zhu JK (2006) A R2R3
type MYB TF is involved in the cold regulation of CBF genes and in acquired
freezing tolerance. J Biol Chem 281:37636-37645
Agarwal PK, Agarwal P, Reddy MK and Sopory SK (2006) Role of DREB transcription
factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep 25:1263-1274
Akhtar M, Jaiswal A, Taj G, Jaiswal JP, Qureshi MI and Singh NK (2012) DREB1/CBF
transcription factors: their structure, function and role in abiotic stress tolerance in
plants. J Genet 91:385-395
Allen MD, Yamasaki K, Ohme-Takagi M, Tateno M and Suzuki M (1998) A novel
mode of DNA recognition by a β-sheet revealed by the solution structure of the
GCC-box binding domain in complex with DNA. EMBO J 17(18):5484-5496
22

Araki S, Ito M, Soyano T, Nishihama R and Machida Y (2004) Mitotic cyclins
stimulate the activity of c-Myb-like factors for transactivation of G2/M phase-
specific genes in tobacco. J Biol Chem 279:32979-88
Balazadeh S, Siddiqui H, Allu AD, Matallana-Ramirez LP, Caldana C, Mehrnia M,
Zanor MI, Kohler B and Mueller-Roeber B (2010) A gene regulatory network
controlled by the NAC transcription factor ANAC092/AtNAC2/ORE1 during salt-
promoted senescence. Plant J 62:250-264
Branco-Price C, Kawaguchi R, Ferreira R et al (2005) Genome-wide analysis of
transcript abundance and translation in Arabidopsis seedlings subjected to oxygen
deprivation. Annals of Botany 96:647-660
Cai M, Qiu D, Yuan T, Ding X, Li H, Duan L, Xu C, Li X and Wang S (2008)
Identification of novel pathogen-responsive cis-elements and their binding proteins
in the promoter of OsWRKY13, a gene regulating rice disease resistance. Plant Cell
Environ 31:86-96
Chen L, Song Y, Li S, Zhang L, Zou C and Yu D (2012) The role of WRKY
transcription factors in plant abiotic stresses. Biochim Biophys Acta 1819:120-128
Chen M, Wang QY, Cheng XG, Xu ZS, Li LC, Ye XG, Xia LQ, Ma YZ (2007)
GmDREB2, a soybean DRE-binding transcription factor, conferred drought and
high-salt tolerance in transgenic plants. Biochem Biophys Res Commun 353:299-305
Cheong YH, Chang H-S, Gupta R, Wang X, Zhu T and Luan S (2002) Transcriptional
profiling reveals novel interactions between wounding, pathogen, abiotic stress, and
hormonal responses in Arabidopsis. Plant Physiol 129:661-677
Chinnusamy V, Ohta M, Kanrar S, Lee B, Hong X, Agarwal M and Zhu J-K (2003)
ICE1: a regulator of cold-induced transcriptome and freezing tolerance in
Arabidopsis. Gene Dev 17:1043-1054
Christianson JA, Dennis ES, Llewellyn DJ and Wilson IW (2010) ATAF NAC
transcription factors: regulators of plant stress signalling. Plant Signal Behav 5:428-
432
Ciftci-Yilmaz S, Mittler R (2008) The zinc finger network of plants. Cell Mol Life Sci
65:1150-1160
Ciftci-Yilmaz S, Morsy MR, Song L, Coutu A, Krizek BA, Lewis MW, Warren D,
Ciolkowski I, Wanke D, Birkenbihl RP and Somssich IE (2008) Studies on DNA-
binding selectivity of WRKY transcription factors lend structural clues into WRKY
domain function. Plant Mol Biol 68:81-92
23

Cominelli E, Galbiati M, Vavasseur A, Conti L, Sala T, Vuylsteke M, Leonhardt N,
Dellaporta SL and Tonelli C (2005) A guard-cell-specific MYB transcription factor
regulates stomatal movements and plant drought tolerance. Curr Biol 15:1196-1200
Cushman J, Connolly EL, Mittler R (2007) The EAR-motif of the Cys2/His2-type zinc
fingerprotein Zat7 plays a key role in the defense response of Arabidopsis to salinity
stress. J Biol Chem 282:9260-9268
de Oliveira TM, Cidade LC, Gesteira AS, Coelho Filho MA, Soares Filho WS and
Costa MGC (2011) Analysis of the NAC transcription factor gene family in citrus
reveals a novel member involved in multiple abiotic stress responses. Tree Genet
Genomes 7:1123-1134
Denekamp M and Smeekens SC (2003) Integration of wounding and osmotic stress
signals determines the expression of the AtMYB102 transcription factor gene. Plant
Physiol 132:1415-1423
Dietz K-J, Vogel MO and Viehhauser A (2010) AP2/EREBP transcription factors are
part of gene regulatory networks and integrate metabolic, hormonal and
environmental signals in stress acclimation and retrograde signalling. Protoplasma
254:3-14
Dubouzet JG, Sakuma Y, Ito Y, Kasuga M, Dubouzet EG, Miura S, Seki M, Shinozaki
K, Yamaguchi-Shinozaki K (2003) OsDREB genes in rice, Oryza sativa L., encode
transcription activators that function in drought-, high-salt- and cold-responsive gene
expression. Plant J 33:751-763
Eulgem T, Rushton PJ, Robatzek S and Somssich IE (2000) The WRKY superfamily of
plant transcription factors. Trends Plant Sci 5:199-206
Fujimoto SY, Ohta M, Usui A, Shinshi H and Ohme-Takagi M (2000) Arabidopsis
ethylene-responsive element binding factors act as transcriptional activators or
repressors of GCC box-mediated gene expression. Plant Cell 12:393-404
Fujita M, Fujita Y, Maruyma K, Seki M, Hiratsu K, Ohmetakagi M, Trans LS,
Shinozaki KY and Shinozaki K (2004) A dehydration-induced NAC protein, RD26,
is involved in a novel ABA-dependent stress-signalling pathway. Plant J 39:863-876
Gommans WM, Haisma HJ, Rots MG (2005) Engineering zinc finger protein
transcription factors: The therapeutic relevance of switching endogenous gene
expression on or off at command. J Mol Biol 354(3):507-519
http://dx.doi.org/10.1016/j.jmb.2005.06.082
24

Gu YQ, Yang C, Thara VK, Zhou J and Martin GB (2000) Pti4 is induced by ethylene
and salicylic acid, and its product is phosphorylated by Pto kinase. Plant Cell 12:771-
786
Guiltinan MJ, Marcotte Jr WR and Quatrano RS (1990) A plant leucine zipper protein
that recognizes an abscisic acid response element. Science 250:267-271
Hao YJ, Song QX, Chen HW, Zou HF, Wei W and Kang XS et al. (2010) Plant NAC-
type transcription factor proteins contain a NARD domain for repression of
transcriptional activation. Planta 232:1033-1043
Hao YJ, Wei W, Song QX, Chen HW, Zhang YQ, Wang F, Zou HF, Lei G, Tian AG,
Zhang WK, Ma B, Zhang JS and Chen SY (2011) Soybean NAC transcription
factors promote abiotic stress tolerance and lateral root formation in transgenic
plants. Plant J 68:302–313
He XJ, Mu RL. Cao WH, Zhang ZG, Zhang JS and Chen SY (2005) AtNAC2, a
transcription factor downstream of ethylene and auxin signalling pathways, is
involved in salt stress response and lateral root development. Plant J 44:903-916
Higginson T, Li SF and Parish RW (2003) AtMYB103 regulates tapetum and trichome
development in Arabidopsis thaliana. Plant J 35:177-192
Hu H, Dai M, Yao J, Xiao B, Li X, Zhang Q and Xiong L (2006) Overexpressing a
NAM, ATAF, and CUC transcription factor enhances drought resistance and salt
tolerance in rice. PNAS 103:12987-12992
Hu H, You J, Fang Y, Zhu X, Qi Z and Xiong L (2008) Characterization of
transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol
Biol 67:169-181
Hu R, Qi G, Kong Y, Kong D, Gao Q and Zhou G (2010) Comprehensive analysis of
NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant
Biol 10:145
Hu YX, Wang YH, Liu XF, Li JY (2004) Arabidopsis RAV1 is down-regulated by
brassinosteroid and may act as a negative regulator during plant development. Cell
Res 14:8-15
Huang XY, Chao DY, Gao JP, Zhu MZ, Shi M, Lin HX (2009) A previously unknown
zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal
aperture control. Genes Dev 23:1805-1817
Ikeda M, Ohme-Takagi M (2009) A novel group of transcriptional repressors in
Arabidopsis. Plant Cell Physiol 50:970-975
25

Jensen MK, Kjaersgaard T, Nielsen MM, Galberg P, Petersen K, O'Shea C and Skriver
K (2010) The Arabidopsis thaliana NAC transcription factor family: structure–
function relationships and determinants of ANAC019 stress signalling. Biochem J
426:183-196
Jeong JS, Kim YS, Baek KH, Jung H, Ha S-H, Choi YD, Kim M, Reuzeau C and Kim
J-K (2010) Root-specific expression of OsNAC10 improves drought tolerance and
grain yield in rice under field drought conditions. Plant Physiol 153:185-197
Jiang Y and Deyholos MK (2006) Comprehensive transcriptional profiling of NaCl-
stressed Arabidopsis roots reveals novel classes of responsive genes. BMC Plant Biol
6:25
Jofuku KD, Boer B, Montagu MV and Okamuro JK (1994) Control of Arabidopsis
flower and seed development by the homeotic gene APETALA2. Plant Cell 6:1211-
1225
Kagaya Y, Ohmiya K and Hattori T (1999) RAV1, a novel DNA-binding protein, binds
to bipartite recognition sequence through two distinct DNA-binding domains
uniquely found in higher plants. Nucleic Acids Res 27:470-478
Kawaura K, Mochida K and Ogihara Y (2008) Genome-wide analysis for identification
of salt-responsive genes in common wheat. Funct Integr Genom 8:277-286
Kirik V, Kölle K, Miséra S and Baümlein H (1998) Two novel MYB homologues with
changed expression in late embryogenesis-defective Arabidopsis mutants. Plant Mol
Biol 37:819-827
Kodaira KS, Qin F, Tran LS, Maruyama K, Kidokoro S, Fujita Y, Shinozaki K,
Yamaguchi-Shinozaki K (2011) Arabidopsis Cys2/His2 zinc-finger proteins AZF1
and AZF2 negatively regulate abscisic acid-repressive and auxin-inducible genes
under abiotic stress conditions. Plant Physiol 157:742-756
Lata C, Yadav A and Prasad M (2011) Role of Plant Transcription Factors in Abiotic
Stress Tolerance DOI: 10.5772/23172. Book chapter within book titled “Abiotic
Stress Response in Plants- Physiological, Biochemical and Genetic Perspectives”
edited by Arun Shanker and B. Venkateswarlu, ISBN 978-953-307-672-0
Le DT, Nishiyama R, Watanabe Y, Mochida K, Yamaguchi-Shinozaki K, Shinozaki K
and Tran LS (2011) Genome-wide survey and expression analysis of the plant-
specific NAC transcription factor family in soybean during development and
dehydration stress. DNA Res 18:263–276
26

Lee H, Guo Y, Ohta M, Xiong L, Stevenson B, Zhu JK (2002) LOS2, a genetic locus
required for cold-responsive gene transcription encodes a bi-functional enolase.
EMBO J 21:2692-2702
Li X, Duan X, Jiang H, Sun Y, Tang Y, Yuan Z, Guo J, Liang W, Chen L, Yin J, Ma H,
Wang J and Zhang D (2006) Genome-wide analysis of basic/helix–loop–helix
transcription factor family in rice and Arabidopsis. Plant Physiol 141:1167-1184
Li XP, Tian AG, Luo GZ, Gong ZZ, Zhang JS, Chen SY (2005) Soybean DRE-binding
transcription factors that are responsive to abiotic stresses. Theor Appl Genet
110:1355–1362
Liang YK, Dubos C, Dodd IC, Holroyd GH, Hetherington AM and Campbell MM
(2005) AtMYB61, an R2R3-MYB transcription factor controlling stomatal aperture
in Arabidopsis thaliana. Curr Biol 15:1201-1206
Lindemose S, O’Shea C, Krogh Jensen M and Skriver K (2013) Structure, Function and
Networks of Transcription Factors Involved in Abiotic Stress Responses. Int J Mol
Sci 14:5842-5878 doi:10.3390/ijms14035842
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K and Shinozaki
K (1998) Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA
binding domain separate two cellular signal transduction pathways in drought- and
low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell
10:1391-1406
Liu QL, Xu KD, Zhao LJ, Pan YZ, Jiang BB, Zhang HQ and Liu GL (2011)
Overexpression of a novel chrysanthemum NAC transcription factor gene enhances
salt tolerance in tobacco. Biotechnol Lett 33:2073-2082
Liu YZ, Baig MNR, Fan R, Ye JL, Cao YC and Deng XX (2009) Identification and
expression pattern of a novel NAM, ATAF, and CUC-like gene from Citrus sinensis
Osbeck. Plant Mol. Biol. Rep. 27:292–297
Lu PL, Chen NZ, An R, Su Z, Qi BS, Ren F, Chen J and Wang XC (2007) A novel
drought-inducible gene, ATAF1, encodes a NAC family protein that negatively
regulates the expression of stress-responsive genes in Arabidopsis. Plant Mol Biol
63:289-305
Maeo K, Hayashi S, Kojima-Suzuki H, Morikami A and Nakamura D (2001) Role of
conserved residues of the WRKY domain in the DNA-binding of tobacco WRKY
family proteins. Biosci Biotechnol Biochem 65:2428–2436
27

Magnani E, Sjölander K and Hake S (2004) From endonucleases to transcription
factors: evolution of the AP2 DNA binding domain in plants. Plant Cell 16:2265-
2277
Marquez CP and Pritham EJ (2010) Phantom, a new subclass of Mutator DNA
transposons found in insect viruses and widely distributed in animals. Genetics
185:1507-1517
Mizoi J, Shinozaki K and Yamaguchi-Shinozak K (2012) AP2/ERF family transcription
factors in plant abiotic stress responses. Biochem Biophys Acta 1819:86-96
Mundy J, Yamaguchi-Shinozaki K and Chua N-H (1990) Nuclear proteins bind
conserved elements in the abscisic acid-responsive promoter of a rice rab gene.
PNAS 87:1406-1410
Nakashima K, Takasaki H, Mizoi J, Shinozaki K and Yamaguchi-Shinozaki K (2012)
NAC transcription factors in plant abiotic stress responses. Biochim Biophys Acta
1819(2):97-103
Newman LJ, Perazza DE, Juda L and Campbell MM (2004) Involvement of the R2R3-
MYB, AtMYB61, in the ectopic lignification and dark-photomorphogenic
components of the det3 mutant phenotype. Plant J 37:239-250
Nuruzzaman M, Manimekalai R, Sharoni AM, Satoh K, Kondoh H, Ooka H and
Kikuchi S (2010) Genome-wide analysis of NAC transcription factor family in rice.
Gene 465:30-44
Ohme-Takagi M and Shinshi H (1995) Ethylene-inducible DNA binding proteins that
interact with an ethylene-responsive element. Plant Cell 7:173-182
Okamuro JK, Caster B, Villarroel R, Van Montagu M and Jofuku KD (1997) The AP2
domain of APETALA2 defines a large new family of DNA binding proteins in
Arabidopsis. PNAS 94:7076-7081
Olsen AN, Ernst HA, Leggio LL and Skriver K (2005) NAC transcription factors:
Structurally distinct, functionally diverse. Trends Plant Sci 10:79-87
Ooka H, Satoh K, Doi K, Nagata T, Otomo Y, Murakami K, Matsubara K, Osato N,
Kawai J, Carninci P, Hayashizaki Y, Suzuki K, Kojima K, Takahara Y, Yamamoto K
and Kikuchi S (2003) Comprehensive analysis of NAC family genes in Oryza sativa
and Arabidopsis thaliana. DNA Res 10:239-47
Pandey SP and Somssich IE (2009) The role of WRKY transcription factors in plant
immunity. Plant Physiol 150:1648-1655
28

Pattanaik S, Xie CH and Yuan L (2008) The interaction domains of the plant Myc-like
bHLH transcription factors can regulate the transactivation strength. Planta 227:707-
715
Pérez JC, Groisman EA. (2009) Transcription factor function and promoter architecture
govern the evolution of bacterial regulons. PNAS 106:4319-4324
Puranik S, Bahadur RP, Srivastava PS and Prasad M (2011) Molecular cloning and
characterization of a membrane associated NAC family gene, SiNAC from foxtail
millet [Setaria italica (L.) P. Beauv.]. Mol Biotech 49:138–150
Puranik S, Sahu PP, Srivastava PS and Prasad M (2012) NAC proteins: regulation and
role in stress tolerance. Trends Plant Sci 17(6):369-381
Ramamoorthy R, Jiang SY, Kumar N, Venkatesh PN and Ramachandran S (2008) A
comprehensive transcriptional profiling of the WRKY gene family in rice under
various abiotic and phytohormone treatments. Plant Cell Physiol 49:865-879
Rashid M, Guangyuan H, Guangxiao Y, Hussain J and Xu Y (2011) AP2/ERF
transcription factor in Rice: genome-wide convas and syntenic relationships between
monocots and eucots. Evol Bioinformatics 8:321-355 DOI: 10.4137/EBO.59369
Rizhsky L, Davletova S, Liang H et al (2004) The zinc finger protein Zat12 is required
for cytosolic ascorbate peroxidase 1 expression during oxidative stress in
Arabidopsis. J Biol Chem 279:11736-11743
Rushton PJ, Bokowiec MT, Han S, Zhang H, Brannock JF, Chen X, Laudeman TW and
Timko MP (2008) Tobacco transcription factors: novel insights into transcriptional
regulation in the solanaceae. Plant Physiol 147:280-295
Rushton PJ, Somssich IE, Ringler P and Shen QJ (2010). WRKY transcription factors.
Trends Plant Sci 15(5):247-258
Saibo NJ, Lourenço T, Oliveira MM (2009) Transcription factors and regulation of
photosynthetic and related metabolism under environmental stresses. Ann Bot
103:609-623
Sakuma Y, Liu Q, Dubouzet JG, Abe H, Shinozaki K and Yamaguchi-Shinozaki K
(2002) DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs,
transcription factors involved in dehydration- and cold-inducible gene expression.
Biochem Biophys Res Commun 290:998-1009
Sakuma Y, Maruyama K, Osakabe Y, Qin F, Seki M, Shinozaki K and Yamaguchi-
Shinozaki K (2006) Functional analysis of an Arabidopsis transcription factor,
DREB2A, involved in drought-responsive gene expression. Plant Cell 18:1292-1309
29

Sohn KH, Lee SC, Jung HW, Hong JK, Hwang BK (2006) Expression and function
roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease
resistance, and drought and salt stress tolerance. Plant Mol Biol 61(6):897-915
Song Y, Ai CR, Jing SJ and Yu DQ (2010) Research progress on function analysis of
rice WRKY gene. Rice Sci 17:60-72
Souer E, van Houwelingen A, Kloos D, Mol J and Koes R (1996) The No Apical
Meristem gene of petunia is required for pattern formation in embryos and flowers
and is expressed as meristem and primordia boundaries. Cell 85:159-170
Stockinger EJ, Gilmour SJ and Thomashow MF (1997) Arabidopsis thaliana CBF1
encodes an AP2 domain-containing transcription activator that binds to the C-
repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in
response to low temperature and water deficit. PNAS 94:1035-1040
Stracke R, Werber M and Weisshaar B (2001) The R2R3-MYB gene family in
Arabidopsis thaliana. Curr Opin Plant Biol 4:447-456
Sun C, Palmqvist S, Olsson H, Borén M, Ahlandsberg S and Jansson C (2003) A novel
WRKY transcription factor, SUSIBA2, participates in sugar signalling in barley by
binding to the sugar responsive elements of the iso1 promoter. Plant Cell 15:2076-
2092
Takasaki H, Maruyama K, Kidokoro S, Ito Y, Fujita Y, Shinozaki K, Yamaguchi-
Shinozaki K and Nakashima K (2010) The abiotic stress-responsive NAC-type
transcription factor OsNAC5 regulates stress-inducible genes and stress tolerance in
rice. Mol Genet Genom 284:173-183
Thomashow MF (1999) Plant cold acclimation: freezing tolerance genes and regulatory
mechanisms. Annu Rev Plant Physiol Plant Mol Biol 50:571-599
Tran LS, Nakashima K, Sakuma Y, Simpson SD, Fujita Y, Maruyama K, Fujita M, Seki
M, Shinozaki K andYamaguchi-Shinozaki K (2004) Isolation and functional analysis
of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-
responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant
Cell 16:2481-2498
Wang N, Zheng Y, Xin H, Fang L and Li S (2013) Comprehensive analysis of NAC
domain transcription factor gene family in Vitis vinifera. Plant Cell Rep. 32:61-75
Wei W, Zhang Y, Han L, Guan Z and Chai T (2008) A novel WRKY transcriptional
factor from Thlaspi caerulescens negatively regulates the osmotic stress tolerance of
transgenic tobacco. Plant Cell Rep 27:795-803
30

Woodrow P, Pontecorvo G, Ciarmiello LF, Annunziata MG, Fuggi A and Carillo P
(2012) Transcription factors and Genes in Abiotic stress DOI 10.1007/978-94-007-
2220-0_9. Book chapter within book titled “Crop Stress and its Management:
Perspectives and Strategies” Springer, edited by Bandi V, Shanker AK, Shanker C,
Mandapaka M ISBN 978-94-007-2219-4
Wu Y, Deng Z, Lai J, Zhang Y, Yang C, Yin B, Zhao Q, Zhang L, Li Y, Yang C and
Xie Q (2009) Dual function of Arabidopsis ATAF1 in abiotic and biotic stress
responses. Cell Res 19:1279-1290
Xia N, Zhang G, Liu XY, Deng L, Cai GL, Zhang Y, Wang XJ, Zhao J, Huang LL and
Kang ZS (2010) Characterization of a novel wheat NAC transcription factor gene
involved in defense response against stripe rust pathogen infection and abiotic
stresses. Mol Biol Rep 37:3703-3712
Xie Z, Zhang ZL, Zou X, Huang J, Ruas P, Thompson D and Shen QJ (2005)
Annotations and functional analyses of the rice WRKY gene superfamily reveal
positive and negative regulators of abscisic acid signalling in aleurone cells. Plant
Physiol 137:176-189
Xue GP, Loveridge CW (2004) HvDRF1 is involved in abscisic acid-mediated gene
regulation in barley and produces two forms of AP2 transcriptional activators,
interacting preferably with a CT-rich element. Plant J 37:326-339
Xue GP, Way HM, Richardson T, Drenth J, Joyce PA and McIntyre CL (2011)
Overexpression of TaNAC69 leads to enhanced transcript levels of stress up-
regulated genes and dehydration tolerance in bread wheat. Mol Plant 4:697-712
Yadav V, Mallappa C, Gangappa SN, Bhatia S, Chattopadhyay S (2005) A basic helix-
loop-helix transcription factor in Arabidopsis, MYC2, acts as a repressor of blue
light-mediated photomorphogenic growth. Plant Cell 17:1953-1966
Yamaguchi-Shinozaki K (2012) Rice phytochrome-interacting factor-like protein
OsPIL1 functions as a key regulator of internode elongation and induces a
morphological response to drought stress. PNAS 109:15947-15952
Yamaguchi-Shinozaki K and Shinozaki K (2005) Organization of cis-acting regulatory
elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci 10:88-
94
Yamaguchi-Shinozaki K and Shinozaki K (2006) Transcriptional regulatory networks in
cellular responses and tolerance to dehydration and cold stresses. Annu Rev Plant
Biol 57:781-803
31

Yamasaki K, Kigawa T, Seki M, Shinozaki K, Yokoyama S (2012) DNA-binding
domains of plant-specific transcription factors: structure, function, and evolution.
Trends Plant Sci S1360-1385(12)00196-3
Yang B, Jiang Y, Rahman MH, Deyholos MK and Kav NN (2009) Identification and
expression analysis of WRKY transcription factor genes in canola (Brassica napus
L.) in response to fungal pathogens and hormone treatments. BMC Plant Biol 9:68
Yanhui C, Xiaoyuan Y, Kun H, Meihua L, Jigang L, Zhaofeng G, Zhiqiang L, Yunfei
Z, Xiaoxiao W, Xiaoming Q, Yunping S, Xiaohui D, Jingchu L, Xing-Wang D,
Zhangliang C, Hongya G and Li-Jia Q (2006) The MYB transcription factor
superfamily of Arabidopsis: expression analysis and phylogenetic comparison with
the rice MYB family. Plant Mol Biol 60:107-124
Zhang Y and Wang L (2005) The WRKY transcription factor superfamily: its origin in
eukaryotes and expansion in plants. BMC Evol Biol 5:1
Zheng X, Chen B, Lu G and Han B (2009) Overexpression of a NAC transcription
factor enhances rice drought and salt tolerance. Biochem Biophys Res Commun
379:985-989
Zhou QY, Tian AG, Zou HF, Xie ZM, Lei G, Huang J, Wang CM, Wang HW, Zhang
JS and Chen SY (2008) Soybean WRKY-type transcription factor genes,
GmWRKY13, GmWRKY21, and GmWRKY54, confer differential tolerance to
abiotic stresses in transgenic Arabidopsis plants. Plant Biotechnol J 6:486-503
Zhou X, Jiang YJ and Yu DQ (2011) WRKY22 transcription factor mediates dark-
induced leaf senescence in Arabidopsis. Mol Cells 31:303-313
Zhuang J, Sun CC, Zhou XR, Xiong AS, Zhang J (2011) Isolation and characterization
of an AP2/ERF-RAV transcription factor Bna RAV-1-HY15 in Brassica napus L.
HuYou15. Mol Biol Rep 38:3921-3928
32