a review of the glycine clandestina species complex (fabaceae: phaseolae) reveals two new species
TRANSCRIPT

P u b l i s h i n g
Volume 14, 2001© CSIRO 2001
Australian Systematic BotanyCSIRO PublishingPO Box 1139 (150 Oxford St)Collingwood, Vic. 3066, Australia
Telephone: +61 3 9662 7613Fax: +61 3 9662 7611Email: [email protected]
Published by CSIRO Publishing for CSIRO and the Australian Academy of Science
w w w . p u b l i s h . c s i r o . a u / j o u r n a l s / a s b
All enquiries and manuscripts should be directed to:
An international journal devoted to the taxonomy, b iogeographyand evolution of all plant groups
a u s t r a l i a ns y s t e m at i cb ota n y

Australian Systematic Botany 14, 891–900
© CSIRO 2001 1030-1887/01/0689110.1071/SB00041
A Review of the Glycine clandestina Species Complex (Fabaceae: Phaseolae) reveals Two New Species
B. E. PfeilAC, M. D. TindaleB and L. A. CravenA
ACentre for Plant Biodiversity Research, CSIRO Plant Industry, GPO Box 1600,Canberra, ACT 2601, Australia.BNational Herbarium of New South Wales, Royal Botanic Gardens, Mrs Macquaries Rd,Sydney, NSW 2000, Australia.CCorresponding author: [email protected]
Abstract
Digitate-leaved specimens of the genus Glycine from across southern Australia that have been determinedas G. clandestina J.C.Wendl. encompass a wide range of morphological variation. Re-evaluation of thismaterial has resulted in the conclusion that three species should be recognised. The original concept ofG. clandestina (digitate-leaved plants with long, c. nine-seeded pods) can be applied only to plants fromQueensland, New South Wales, Victoria and Tasmania. Two new species, G. peratosa B.E.Pfeil & Tindaleand G. rubiginosa Tindale & B.E.Pfeil, are described from Western Australia and South Australia,respectively. Variation in G. clandestina s.s. is discussed in relation to contact zones with three other closelyrelated species.Revi ew of Gl yci ne cl andes ti naB. E. Pf eilet alSB00041
Introduction
As part of our revision of the genus Glycine Willd. for the Flora of Australia, anexamination of various species complexes in the genus has been undertaken. This paperdiscusses the variation in the Glycine clandestina J.C.Wendl. species complex and includesdescriptions of two new species that have become apparent as a result of our investigations.
Glycina clandestina s.l.
The taxonomic and nomenclatural confusion regarding Glycine clandestina has beentreated by Tindale (1986) who, when choosing a neotype for the name, sought to embodythe traditional concept of G. clandestina (i.e. sensu Wendland 1798) while exposing thecommon confusion of G. microphylla (Benth.) Tindale with G. clandestina.
Tindale (1986) noted that Wendland’s description matched the taxon that occurs ineastern Australia with long legumes and c. nine black seeds per legume. This taxon isusually digitately tri-foliate (rarely pinnate), with straight or nearly straight legumes, rough-surfaced seeds (the perisperm usually adheres) and 7–12 seeds per legume. Other speciescited by Wendland as occurring in the ‘South Sea Islands’ (as G. clandestina is cited) areknown to be restricted to New South Wales (Tindale 1986), so it is most likely thatWendland’s concept was of a taxon occurring in eastern Australia.
However, Bentham’s (1864) treatment of the genus stated that G. clandestina occurredin both Western Australia (WA) and South Australia (SA), as well as the eastern states(Queensland, Qld; New South Wales, NSW; Victoria, Vic.; Tasmania, Tas.). Bentham’s(1864) key describes G. clandestina as possessing digitate leaves and nearly straight pods,but he did not mention the number of seeds per pod. Bentham also erected G. clandestinavar. sericea Benth. for plants with ‘very rusty-villous’ calyces and he considered all the WAand most of the SA material, but only a few specimens from other parts of Australia, tobelong to this variety.
18 December 2001

892 B. E. Pfeil et al.
Bentham’s G. clandestina var. sericea has been applied to eastern Australian plants withrusty calyx hairs by Stanley and Ross (1983) and by numerous botanists makingdeterminations of herbarium specimens. There seems to be confusion between Bentham’sconcept and part of Wendland’s G. clandestina s.s. There are quite a number of easternAustralian G. clandestina specimens with longer (7–12-seeded) legumes, leaflet secondaryvenation c. 90° from the primary vein, seeds with adherent perisperm and rusty calyx hairs.These would appear to be the plants to which Bentham was referring, when stating that thevariety was also present in the eastern states, but his concept included two other taxa asdetailed below.
A large number of specimens with predominantly rusty calyx hairs have recently beencollected from WA and SA by A. H. D. Brown, T. Hymowitz (University of Illinois) andJ. P. Grace (CSIRO Plant Industry). Included in these collections are a number of specimenswith digitate leaves that are unassignable to any known digitate Glycine. Although thesespecimens have previously been considered to belong to G. clandestina, they do not fit thetaxon embodied in Wendland’s description.
The SA material has short straight pods, containing five to nine mottled seeds with asmooth surface (the perisperm does not adhere). The leaflets of these plants are muchsmaller than in G. clandestina from the eastern states (length × width up to 50 × 5 mm inthe SA plants, but up to 105 × 14 mm in G. clandestina) and almost always glabrous on theadaxial surface (G. clandestina is sparsely to moderately hairy). Additionally, the terminalstipels in the SA material are usually absent, or extremely small (up to 0.25 mm long) ifpresent, whereas those in G. clandestina are often present and longer (up to 0.75 mm long).The leaflets have secondary venation from 20° to 50° from the primary vein, whereas thosein G. clandestina are from 40 to 90°. It should also be noted that the secondary venation ofthe SA material is very difficult to see on herbarium material, whereas that ofG. clandestina is readily visible.
Numerous accessions of G. clandestina from the eastern states and SA material havebeen grown under controlled conditions at CSIRO Plant Industry, Canberra, and both haveretained their distinctive features. Character comparisons between G. clandestina and theSA plants are based on extensive field-collected and glasshouse-grown herbarium sheets,as well as observations of several live glasshouse-grown plants. Therefore, we conclude thatthe co-variation of several characters, which appears to be genetically based, is bestexplained by the presence of a separate taxon. Bentham’s concept of G. clandestina var.sericea could, in part, incorporate the SA material, but this taxon warrants recognition atspecific rank due to consistent differences in numerous characters between it andG. clandestina s.s. As the epithet ‘sericea’ is unavailable at specific rank (see Hermann1962), the name Glycine rubiginosa is proposed (description below).
Some of the material from WA is of a different nature to that from SA. Many digitate (orvery shortly pinnate) specimens are from the central part of WA, south from the RobinsonRanges (c. 25°S) to near Merredin (c. 32°S). These plants have markedly curved legumes,containing from three to seven squarish seeds with adherent perisperms and secondaryleaflet venation from 20 to 40° from the primary vein. The latter characters clearly separatethese plants from Wendland’s G. clandestina. Glycine canescens F.J.Herm. also possessesmarkedly curved legumes, with three to seven squarish seeds and secondary leafletvenation from 20 to 50° from the primary vein. The plants from the central part of WAdiffer from other G. canescens plants only by their rusty calyx hairs and often more shortlypinnate leaves and are treated here as part of that species. Furthermore, these plants differfrom G. rubiginosa in that they have obvious secondary venation, again similar to
Review of Glycine clandestina 893
G. canescens, as well in other features already mentioned (curved legumes and roughsquarish seed).
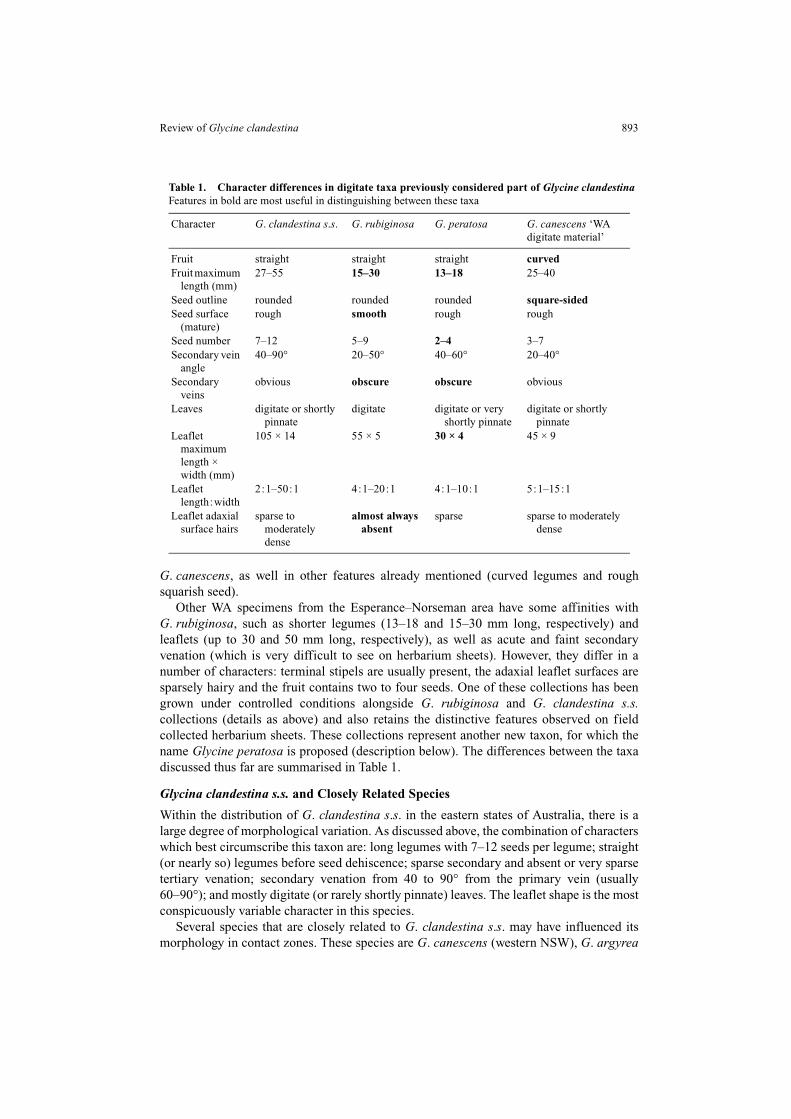
Other WA specimens from the Esperance–Norseman area have some affinities withG. rubiginosa, such as shorter legumes (13–18 and 15–30 mm long, respectively) andleaflets (up to 30 and 50 mm long, respectively), as well as acute and faint secondaryvenation (which is very difficult to see on herbarium sheets). However, they differ in anumber of characters: terminal stipels are usually present, the adaxial leaflet surfaces aresparsely hairy and the fruit contains two to four seeds. One of these collections has beengrown under controlled conditions alongside G. rubiginosa and G. clandestina s.s.collections (details as above) and also retains the distinctive features observed on fieldcollected herbarium sheets. These collections represent another new taxon, for which thename Glycine peratosa is proposed (description below). The differences between the taxadiscussed thus far are summarised in Table 1.
Glycina clandestina s.s. and Closely Related Species
Within the distribution of G. clandestina s.s. in the eastern states of Australia, there is alarge degree of morphological variation. As discussed above, the combination of characterswhich best circumscribe this taxon are: long legumes with 7–12 seeds per legume; straight(or nearly so) legumes before seed dehiscence; sparse secondary and absent or very sparsetertiary venation; secondary venation from 40 to 90° from the primary vein (usually60–90°); and mostly digitate (or rarely shortly pinnate) leaves. The leaflet shape is the mostconspicuously variable character in this species.
Several species that are closely related to G. clandestina s.s. may have influenced itsmorphology in contact zones. These species are G. canescens (western NSW), G. argyrea
Table 1. Character differences in digitate taxa previously considered part of Glycine clandestinaFeatures in bold are most useful in distinguishing between these taxa
Character G. clandestina s.s. G. rubiginosa G. peratosa G. canescens ‘WA digitate material’
Fruit straight straight straight curvedFruit maximum
length (mm)27–55 15–30 13–18 25–40
Seed outline rounded rounded rounded square-sidedSeed surface
(mature)rough smooth rough rough
Seed number 7–12 5–9 2–4 3–7Secondary vein
angle40–90° 20–50° 40–60° 20–40°
Secondary veins
obvious obscure obscure obvious
Leaves digitate or shortly pinnate
digitate digitate or very shortly pinnate
digitate or shortly pinnate
Leaflet maximum length × width (mm)
105 × 14 55 × 5 30 × 4 45 × 9
Leaflet length:width
2:1–50:1 4:1–20:1 4:1–10:1 5:1–15:1
Leaflet adaxial surface hairs
sparse to moderately dense
almost always absent
sparse sparse to moderately dense
894 B. E. Pfeil et al.
Tindale (south-eastern Qld and northern NSW) and G. latrobeana (Meissn.) Benth. (Vic.,Tas.). Features of the leaflets, seed and fruit appear to reflect this possible influence.
Glycina clandestina s.s.–G. canescens
The following information is largely based on Brown et al. (1990) but is included forcompleteness.
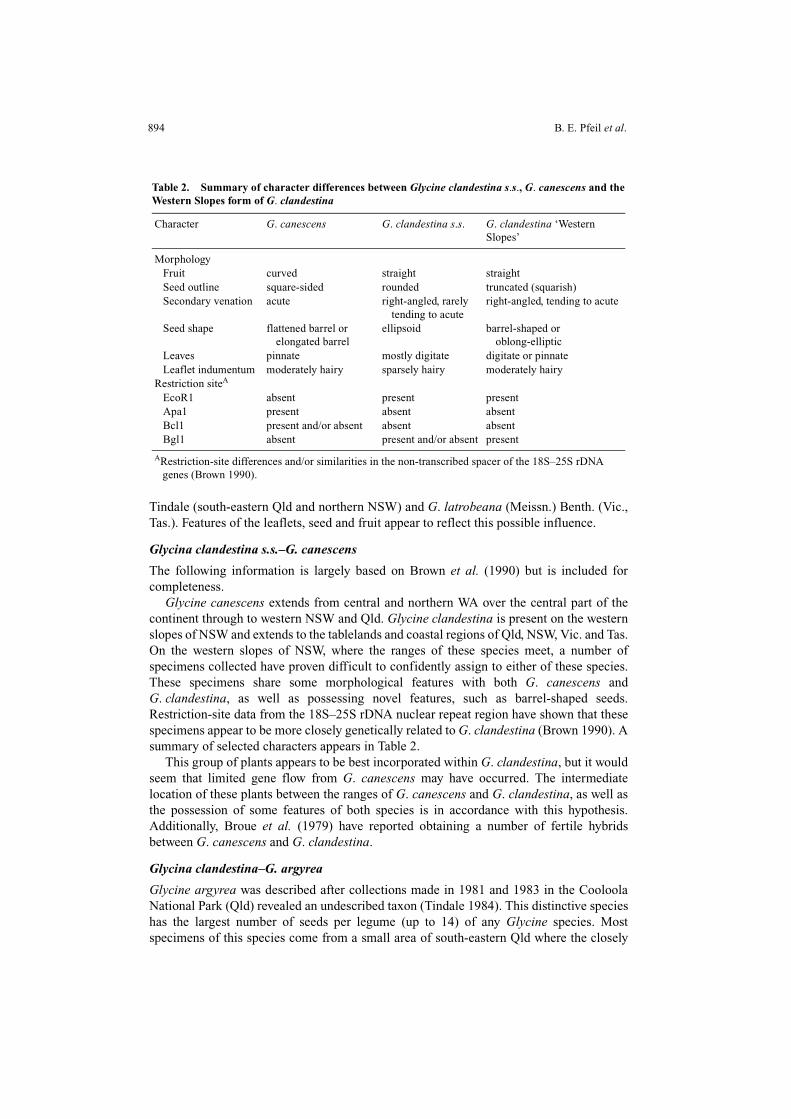
Glycine canescens extends from central and northern WA over the central part of thecontinent through to western NSW and Qld. Glycine clandestina is present on the westernslopes of NSW and extends to the tablelands and coastal regions of Qld, NSW, Vic. and Tas.On the western slopes of NSW, where the ranges of these species meet, a number ofspecimens collected have proven difficult to confidently assign to either of these species.These specimens share some morphological features with both G. canescens andG. clandestina, as well as possessing novel features, such as barrel-shaped seeds.Restriction-site data from the 18S–25S rDNA nuclear repeat region have shown that thesespecimens appear to be more closely genetically related to G. clandestina (Brown 1990). Asummary of selected characters appears in Table 2.
This group of plants appears to be best incorporated within G. clandestina, but it wouldseem that limited gene flow from G. canescens may have occurred. The intermediatelocation of these plants between the ranges of G. canescens and G. clandestina, as well asthe possession of some features of both species is in accordance with this hypothesis.Additionally, Broue et al. (1979) have reported obtaining a number of fertile hybridsbetween G. canescens and G. clandestina.
Glycina clandestina–G. argyrea
Glycine argyrea was described after collections made in 1981 and 1983 in the CooloolaNational Park (Qld) revealed an undescribed taxon (Tindale 1984). This distinctive specieshas the largest number of seeds per legume (up to 14) of any Glycine species. Mostspecimens of this species come from a small area of south-eastern Qld where the closely
Table 2. Summary of character differences between Glycine clandestina s.s., G. canescens and theWestern Slopes form of G. clandestina
Character G. canescens G. clandestina s.s. G. clandestina ‘Western Slopes’
Morphology Fruit curved straight straight Seed outline square-sided rounded truncated (squarish) Secondary venation acute right-angled, rarely
tending to acuteright-angled, tending to acute
Seed shape flattened barrel or elongated barrel
ellipsoid barrel-shaped or oblong-elliptic
Leaves pinnate mostly digitate digitate or pinnate Leaflet indumentum moderately hairy sparsely hairy moderately hairyRestriction siteA
EcoR1 absent present present Apa1 present absent absent Bcl1 present and/or absent absent absent Bgl1 absent present and/or absent present
ARestriction-site differences and/or similarities in the non-transcribed spacer of the 18S–25S rDNA genes (Brown 1990).

Review of Glycine clandestina 895
related G. clandestina also occurs; however, a more recent collection has been made nearPort Macquarie, NSW (J. P. Grace, pers. comm.).
Glycine clandestina specimens from northern NSW and Qld, in particular the areaclosest to the distribution of G. argyrea, show an increased similarity to G. argyrea, whencompared with specimens of G. clandestina from southern NSW and Vic. Some of thesefeatures are: an increase in the occurrence of the pinnate leaf condition; an increase inlegume length and number of seeds; and seeds occasionally with stellate (rather thancircular) tubercles.
As fertile hybrids have been easily obtained between G. argyrea and G. clandestina(Grant et al. 1986) and as these species occur in the same area, it is possible that gene flowbetween these species is responsible for the morphological intermediacy that has beenobserved. However, this must remain an unconfirmed hypothesis until appropriate data isavailable to test it. A putative F1 hybrid has also been found in nature (R. Pullen andJ. Grant 11094, 15.xii.1984; CSIRO accession G 736).
Glycina clandestina–G. latrobeana
Populations of G. clandestina in the higher parts of the Great Dividing Range in NSW andVic. often exhibit broader and shorter leaflets compared with plants from further north andcloser to the coast. Although G. latrobeana has a similar leaflet shape to theseG. clandestina populations, it is less likely that G. latrobeana has contributed to thisvariation for the following reasons. First, G. clandestina specimens do not show any signof intermediacy in the autapomorphy of G. latrobeana, the very broad, stem-sheathing andmostly fused (on one side) stipules. Second, the leaflet shape of the higher altitudeG. clandestina populations is more similar to the juvenile leaflet shape of this species thanit is to G. latrobeana leaflets. Third, no fertile hybrids between these species have beenreported. A simpler explanation may be that the juvenile foliage is retained longer in theseenvironments.
Juvenile leaflets of Glycine latrobeana are usually circular to obovate, as long as broad,with emarginate to obtuse apices. The adult leaflets are the same shape as the juveniles butlarger, or more rarely up to three times as long as broad with emarginate or obtuse apices.Juveniles of G. clandestina from Qld, NSW and Vic. only very rarely display emarginateapices, are usually from two to three times as long as broad and the adult leaflets are neveremarginate.
Variation in Leaflet Morphology within G. clandestina s.s.
A number of prominent leaf morphotypes exist within the range of G. clandestina, apartfrom those discussed above. Some of these morphotypes occupy restricted ranges withinthe overall distribution of the species, while others are widespread. However, the presenceof a high degree of intermediate forms precludes the recognition of any formal taxa withinthis species. Further work may reveal significant morphological and/or genetic structurethat may be accorded some taxonomic rank; however, it is beyond the scope of this review.A map of the main morphotypes and their distributions is presented to assist furtherinvestigation (Fig. 1).
Key to Glycine clandestina and Related Species
1. Leaflets with obscure secondary venation; leaflets up to 55 mm long and 5 mm wide . . . . . . . . . . . . . . . . . 2Leaflets with obvious secondary venation; leaflets up to 105 mm long and to 15 mm wide . . . . . . . . . . . . 3

896 B. E. Pfeil et al.
2. Seeds smooth at maturity; fruit 5–9-seeded; leaves always digitate; leaflets almost always glabrous onthe adaxial surface . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G. rubiginosa
Seeds rough at maturity; fruit 2–4-seeded; leaves digitate to very shortly pinnate; leaflets sparsely hairyon the adaxial surface . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G. peratosa
3. Apex of mature fruit curved upwards before dehiscence; leaves pinnate; 2° venation 20–50° from mid-vein . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G. canescens
Apex of mature fruit straight before dehiscence; leaves pinnate or digitate; 2° venation 30–90° from mid-vein . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
4. Stipules partly fused on one side (around the stem), broad-lanceolate and often blunt; fruit 4–6-seeded;2° venation 30–50° from mid-vein . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G. latrobeana
Stipules free, narrow-lanceolate and acute; fruit 7–14-seeded; 2° venation 40–90° from mid-vein (often60–90°) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Fig. 1. Distribution of leaflet variation in Glycine clandestina s.s. The morphotype (�) corresponds withG. clandestina ‘Western Slopes’ in Table 2 (G. canescens ‘group X’ in Brown et al. 1990). Othermorphotypes (�, �, �, �, �, �) represent examples of extremes of variation within Glycineclandestina s.s. There are many intermediates to be found between these forms.

Review of Glycine clandestina 897
5. Leaflet abaxial surfaces densely hairy, hairs white to sometimes pale rusty; leaflets discolorous (adaxialsurface dark green, abaxial surface made much paler by the indumentum); fruit 10–14-seeded;terminal leaflet stipels present and to 1.25 mm long; leaves usually pinnate . . . . . . . . . . . . . . . G. argyrea
Leaflet abaxial surfaces sparsely to moderately hairy, hairs white to rusty; leaflets often not noticablydiscolorous, but if so, then sparsely hairy; fruit 7–12-seeded; terminal stipels often present, if so, upto 0.75 mm long; leaves occasionally shortly pinnate . . . . . . . . . . . . . . . . . . . . . . . . . . . . G. clandestina s.s.
Descriptions of New Taxa
Glycine rubiginosa Tindale et B.E.Pfeil, sp. nov.
Glycine rubiginosa Tindale et B.E.Pfeil: a G. clandestina J.C.Wendl. foliolis plerumqueminoribus usque ad 55 mm longus et ad 5 mm latis, venis secondariis inconspicuis etacutioribus ab 20–50°, foliolis adaxialibus laminis et venis primariis pilis plerumquecarentibus, leguminibus minoribus usque ad 30 mm longis et ad 2.5 mm latis; seminibusmaculosis et laevibus differt.
Typus: SA, on hill slopes at South Blinman, D.E. Symon 6766, 5.ix.1969 (holo: CANB;iso: AAU, AD, AHUC, B, K, US—all iso n.v.).Glycine clandestina var. sericea Benth., Fl. Austral. 2: 244 (1864). p.p.
Climbing, non-stoloniferous, perennial herb. Stem hairs white to rusty (usually white),appressed to ascending-spreading, sparse to moderately dense; stipules up to 1.5 mm longand up to 0.5 mm wide, not fused, the hairs white to rusty, appressed to ascending, sparseto moderately dense. Petioles to 11 mm long, the hairs white to dark brown (usually white),appressed to ascending, sparse to moderately dense. Leaves digitately trifoliate. Lateralstipels up to 0.5 mm long; terminal stipels obscure if present (up to 0.25 mm long), butusually absent; stipel hairs white, appressed, moderately dense to dense. Petiolules to 1 mmlong, the hairs white to rusty; ascending to spreading; moderately dense. Terminal rachisabsent. Leaflets up to 55 mm long and up to 5 mm wide, usually linear but also narrowlyobovate, narrowly elliptic or elliptic; margin flat or slightly recurved; apex acuminate andmucronate; terminal leaflets may be slightly larger than lateral leaflets. Leafletlength:width ratio 4:1–20:1. Leaflet venation brochidodromous; secondary veins faint,typically 20–50° from mid-vein; reticulation obscure, sparse. Leaflet hairs: abaxial mid-vein and laminal hairs white to rusty, appressed, sparse to rarely moderately dense; adaxialmid-vein and laminal hairs usually absent, if present then white, appressed and very sparse.
Inflorescences loosely racemose; each bract subtending a single flower. Peduncles up to120 mm long, the hairs white to dark brown, appressed to ascending, sparse to dense; bractsup to 2 mm long and up to 0.5 mm wide, the hairs white to dark brown, ascending toascending-spreading, moderately dense to dense. Pedicels up to 5 mm long, the hairs whiteto dark brown, appressed to ascending, moderately dense to dense; bracteoles up to 1 mmlong, the hairs white to dark brown, appressed to ascending, moderately dense to dense;bracteoles inserted at the base of the calyx or a little below. Calyx up to 2.25 mm long and2 mm wide, the sinus between the two adaxial teeth 0.25–0.5 mm long; the calyx hairs darkbrown (sometimes with scattered white hairs intermixed), appressed to ascending,moderately dense to dense. Corolla purple to mauve; the standard 6–8 mm long, 4–8 mmwide, without lobes on the lower edge; wings 5 mm long; keel 4–5 mm long. Carpel hairs:sometimes forming a collar on the style.
Chasmogamous and cleistogamous legumes 15–30 mm long and up to 2.5 mm wide,brown when mature, initially straight but becoming twisted after dehiscence; the legumehairs white to rusty, appressed to ascending, usually sparse but sometimes absent.Cleistogamous legumes solitary in the leaf axils. Seeds 5–9 per chasmogamous legume

898 B. E. Pfeil et al.
(3–8 per cleistogamous legume), c. 3 mm long; mottled; barrel-shaped or ellipsoidal(somewhat irregular), usually with a rounded outline; surface smooth, lacking surfacetubercles and reticulation.
Distribution and Habitat (Fig.2)
This species occurs in and near the Flinders, Lofty and Gawler Ranges in SA, the BarrierRanges in NSW and as far west as Caiguna in WA (at the western end of the GreatAustralian Bight). The altitude range of this species is from 20 to 600 m and it is commonlyassociated with quartzite outcrops (A. H. D. Brown, pers. comm.).
Selected Specimens
WESTERN AUSTRALIA: ‘Iron Tanks’, 27 km S Caiguna via Baxter’s Memorial Track, M. FitzgeraldC-12, 24.viii.1983 (PERTH); near Point Dover, c. 380 km W of SA–WA border on edge of GreatAustralian Bight, P.G. Wilson 5925, 19.vii.1967 (PERTH). SOUTH AUSTRALIA: Mt ChambersGorge, near aboriginal site, J.P. Grace, A.H.D. Brown, L.A. Craven, H. Hurka and U.Hurka 801,13.ii.1993 (AD, CANB, NSW); track to Bungeroo Gorge, SA, M.E. Phillips 131, 21.viii.1964(CANB); Goolwa, old golf course, 2.5 km W from Goolwa towards Victor Harbour, J.P. Grace, A.H.D.Brown and J.E. Grant 271, 2.xi.1985 (AD, CANB, K, NSW); Cadbury Springs, 20 km W of WilpenaHeadquarters, J.P. Grace, A.H.D. Brown and J.E. Grant 261, 28.x.1985 (AD, CANB, K, NSW). NEWSOUTH WALES: Silverton, collector unknown, 18.viii.1958 (UNE); 12 miles from Broken Hill onMenindee Road, R.A. Bourne, 14.viii.1968 (UNE).
Notes
Flowers have been recorded from July to November. The latin rubiginosus means rusty-red,which alludes to the calyx hairs.
Glycine peratosa B.E.Pfeil et Tindale, sp. nov.
Glycine peratosa B.E.Pfeil et Tindale: a G. clandestina J.C. Wendl. venis secondariisinconspicuis et plerumque acutioribus ab 40–60°; a G. rubiginosa Tindale et B.E.Pfeil etG. clandestina foliolis minoribus usque ad 30 mm longis et ad 4 mm latis, 2–4–seminalis;a G. rubiginosa seminibus scabris differt.
Fig. 2. Distributions of Glycine peratosa (�) and G. rubiginosa (�).

Review of Glycine clandestina 899
Typus: WA, Pallinup River crossing, Hassell Highway, Albany side, A.H.D. Brown, J.J.Burdon and J.A. Thompson 1301 (=CSIRO Accession G2916), 12.xi.1991 (holo: CANB).
Twining and climbing, non-stoloniferous, perennial herb. Stem hairs white to rusty,appressed to spreading, sparse to dense; stipules up to 2 mm long and up to 0.5 mm wide, notfused, the hairs white to rusty, appressed to ascending, sparse to moderately dense. Petiolesup to 10 mm long, the hairs white to rusty, appressed to spreading, sparse to moderatelydense. Leaves digitately or rarely pinnately trifoliate. Lateral stipels up to 0.5 mm long;terminal stipels obscure if present (up to 0.25 mm long), but often absent; stipel hairs whiteto rusty, appressed to ascending, sparse to moderately dense. Petiolules up to 0.5 mm long,hairs white to rusty, appressed to spreading, sparse to moderately dense. Terminal rachisrarely up to 2 mm long (usually less than 0.5 mm long). Leaflets up to 30 mm long and up to4 mm wide (usually less than 20 mm long), linear or narrowly elliptic, margin flat or slightlyrecurved, apex acute or obtuse and often mucronate; terminal leaflets sometimes slightlylarger than lateral leaflets. Leaflet length:width ratio 4 : 1–10:1. Leaflet venationbrochidodromous; secondary veins faint, typically 40–60° from mid-vein; reticulationobscure. Leaflet hairs: leaflet abaxial mid-vein and lamina hairs white to pale rusty,appressed to ascending-spreading, sparse to moderately dense; leaflet adaxial mid-vein andlaminal hairs white, appressed to ascending-spreading, sparse to moderately dense.
Inflorescences racemose, but clustered towards the apex; each bract subtending a singleflower. Peduncles up to 40 mm long, the hairs white to rusty, ascending to ascending-spreading, moderate to dense; bracts up to 2 mm long and up to 0.5 mm wide, the hairswhite to rusty, ascending, moderately dense. Pedicels up to 2 mm long, the hairs white torusty, ascending, moderate to dense; bracteoles up to 1 mm long, the hairs white to darkbrown, appressed to ascending-spreading, moderately dense; bracteoles inserted at the baseof the calyx or partway along the pedicel. Calyx up to 3 mm long and to 1.75 mm wide, thesinus between the two adaxial teeth 0.5–1 mm long; calyx hairs rusty to dark brown withsome white hairs intermixed, appressed to ascending-spreading, moderately dense to dense.Corolla blue, violet, mauve or purple, with a yellow centre; standard 5–7 mm long and 5–6mm wide, with or without lobes on the lower edge; wings 4–6 mm long; keel 4–5 mm long.Carpel hairs: greatest density on the apex of the ovary, but not forming a distinct collar.
Chasmogamous legumes not seen. Cleistogamous legumes 13–18 mm long and up to 3mm wide, brown when mature, initially straight but becoming twisted after dehiscence;hairs white to dark brown, appressed to ascending, sparse to dense. Cleistogamous legumessolitary in the leaf axils. Seeds 2–4 per cleistogamous legume, c. 2.5 mm long; black orbrown, not mottled; irregularly barrel-shaped or ellipsoidal, with a rounded outline; rough,lacking surface tubercles; reticulation ill-defined.
Distribution and Habitat (Fig. 2)
This species occurs in the Norseman–Esperance area of WA, extending from Cundeelee inthe north-east to the Pallinup River in the south-west. Plants of this species are commonlyassociated with granite outcrops.
Selected Specimens
WESTERN AUSTRALIA: Fraser Range [as Frazers Range], A.W. Humphries P26, 9.viii.1951 (PERTH);Hassall Highway, Pallinup River crossing, J.W. Green 4848, 23.viii.1978 (PERTH); Cundeelee, P. BoswellF80, 1967 (PERTH); 23 km SE of Sinclair Soak, c. 70 km NE of Norseman, K.R. Newbey 6902, 5.viii.1980(PERTH); 10 km E of Norseman, R.J. Cranfield 743, 22.ix.1978 (PERTH); 120–121 mile peg, E ofNorseman, A.R. Main s.n., no date (PERTH); 3 km SE of Ravensthorpe, A. Sanders AS40, 3.ix.1998(PERTH).

900 B. E. Pfeil et al
http://www.publish.csiro.au/journals/asb
Notes
Flowers have been recorded in August and September. The species epithet is based on theGreek peratos meaning ‘west’ or ‘opposite’, thereby referring to the geographic location ofthese plants.
Acknowledgments
Our thanks go to our collegues Tony Brown and Jim Grace for discussions and assistancethroughout; to Brendan Lepschi and Anna Monro for reviewing the manuscript; to TimEntwisle, David Mabberley and Peter Wilson for useful comments; to the curators of thefollowing herbaria for the opportunity to study specimens in their care: BRI, CANB, DNA,HO, K, MBA, MEL, NSW, PERTH; and to Roy Pullen and ABRS for providing fundingsupport for the Flora of Australia treatment, from which this work stems.
References
Bentham, G. (1864). ‘Flora Australiensis.’ Leguminosae to Combretaceae, Vol. 2. (Lovell Reeve: London.)Broue, P., Marshall, D. R., and Grace, J. P. (1979). Hybridization among the Australian wild relatives of the
soybean. Journal of the Australian Institute of Agricultural Science 45, 256–257.Brown, A. H. D. (1990). The role of isozyme studies in molecular systematics. Australian Systematic
Botany 3, 39–46.Brown, A. H. D., Burdon, J. J., and Grace, J. P. (1990). Genetic structure of Glycine canescens, a perennial
relative of soybean. Theoretical and Applied Genetics 79, 729–736.Grant, J. E., Pullen, R., Brown, A. H. D., Grace, J. P., and Gresshoff, P. M. (1986). Cytogenetic affinity
between the new species Glycine argyrea and its congeners. Journal of Heredity 77, 423–426.Hermann, F. J. (1962). A revision of the genus Glycine and its immediate allies. USDA Technical Bulletin
1268, 1–79.Stanley, T. D., and Ross E. M. (1983). ‘Flora of South-eastern Queensland.’ Vol. 1. (Queensland
Department of Primary Industries: Brisbane.)Tindale, M. D. (1984). Two new eastern Australian species of Glycine Willd. (Fabaceae). Brunonia 7,
207–213.Tindale, M. D. (1986). Taxonomic notes on three Australian and Norfolk Island species of Glycine Willd.
(Fabaceae: Phaseolae) including the choice of a neotype for G. clandestina Wendl. Brunonia 9,179–191.
Wendland, J. C. (1798). ‘Botanische Beobachtungen, nebst einigen neuen Gattungen und Arten.’ (GebrüberHahn: Hannover.) (m.f. at CANB.)
Manuscript received 22 December 2000, accepted 2 April 2001