cloning, expression and cross-linking analysis of the murine p55 tumor necrosis factor receptor
TRANSCRIPT

Eur. J. Immunol. 1991. 21: 1649-1656 Cloning and characterization of a murine p55 TNFR 1649
Cloning, expression and cross-linking analysis of the murine p55 tumor necrosis factor receptor*
Kathy Barrett, David A. Taylor-Fishwick, Andrew P. Cope, A. Maija Kissonerghis, Patrick W. Grayo, Marc Feldmann and Brian M. J. Foxwell
Charing Cross Sunley Research Centre, London
Tumor necrosis factor (TNF) mediates its pleiotropic effects via high-affinity cell surface receptors. In man, molecular cloning has identified two distinct, independent TNF receptors (TNFR) of 55 and 75 kDa. It is unclear, however, whether the multiple effects of TNFare segregated between the receptor types. In the mouse, previous studies had shown functional heterogeneity of TNFR, since the WEHI 164 fibroblast line is sensitive to the cytotoxic effects of both murine and human TNF, whereas the murine Tcell line, CT6, proliferates in response to murine but not human TNE In this study, the cloning of a cDNA encoding the murine homologue of the p55 TNFR is reported. This receptor binds murine and human TNF with equal affinity and is expressed on WEHI 164 and a number of other cell lines, but only low levels of mRNA and no protein is detectable on CT6 cells. CT6 cells, however, express a second TNFR of approximately 75 kDa, identified by cross-linking analysis, which is also found on WEHI 164 cells, and binds only murine TNF. These studies establish that there are also two TNFR in the mouse, and suggests that there may be segregation of the cytotoxic and proliferative responses between different receptors, at least in these cell lines.
1 Introduction
The diverse actions of TNF-a (reviewed [ l , 21) are mediated by interaction with high-affinity receptors, which also bind the related cytokine, lymphotoxin (TNF-p) [3]. In man, TNF receptors (TNFR) have been characterized biochemically as proteins with high affinity for TNF (Kd - M) [4], and recent cDNA cloning studies have led to the definition of two discrete receptors, of 55 kDa, termed p55 [5-71 and approximately 75 kDa, termed p75 [8, 91. The similarity between these receptors is restricted to the extracellular domains, which have four homologous repeats of -40 amino acids, each containing 6 cysteine residues. This structure is characteristic of a family of cell surface proteins, including the nerve growth factor receptor and surface antigens CD40 and OX40 [5 ] . Immunological studies have shown that expression of the individual TNFR is not restricted to cell type, with most cells studied expressing both [4,10,11], but in varying proportions. I t is not yet known whether each receptor mediates all the actions of TNF, or whether cytolytic, proinflammatory or proliferative actions are selectively mediated by one or another receptor. The difference between the cytoplasmic domains of the two receptors suggests differences in signal transduction mechanisms.
[I 93991
* This work was supported by the Arthritis and Rheumatism Council, Wellcome Trust, U.S. Navy Medical Research and Development Command, National Kidney Foundation and Xenova plc.
O Present address: ICOS Corporation, 22021 20th Avenue SE, Bothell, WA 98021, USA.
Correspondence: Kathy Barrett, Charing Cross Sunley Research Centre, 1 Lurgan Avenue, Hammersmith, London W68LW, GB
Abbreviation: PVP: Polyvinylpyrrolidone
Compared with the human TNFR, there is currently little information describing the structure of the murine TNFR. Functional studies have shown that while TNF is indeed cytotoxic for many cell lines, it causes the proliferation of other cell lines [12]. In particular, the murineTcell line CT6 [13] and murine thymocytes [14] bind and proliferate in response to murine TNF but not human TNF. In contrast, the murine fibroblast lines WEHI 164 and L929 [15] show little species specificity in their sensitivity to the cytotoxic action of TNF, and are used as routine assays for human TNF. These results suggest a heterogeneity in the structure and function of murine TNFR.
Characterization of the murine TNFR is essential for defining the basis of the heterogeneity observed above. Moreover, these studies are important, since TNF has been implicated in the pathogenesis of a variety of human diseases and also their corresponding murine models. These include multiple sclerosis [16] and its animal model, EAE [17], rheumatoid arthritis [18] and collagen-type I1 arthritis in mouse (Williams, Williams, Maini and Feldmann, sub- mitted). Understanding the interaction of TNF with its receptors will facilitate studies of the role of TNF in the pathogenesis of these diseases. The murine system will provide a useful model for testing therapeutic agents based on altering the TNF and TNFR interaction in viva.
This study reports the cloning and expression of the murine homologue of the human p55 TNFR. This receptor binds both human and murine TNF with similar affinity, and is expressed in several murine lines including WEHI 164, but is barely detectable on CT6 T cells. However, affinity cross-linking studies demonstrate that the CT6 line expresses at least one other TNF-binding protein, with an estimated molecular mass of 75 kDa, which appears to bind only murine TNF. Therefore, this study of the murineTNFR provides biochemical evidence for their heterogeneity, and suggests that there may be segregation of the diverse responses of cytotoxicity and proliferation between differ- ent receptors.
0 VCH Verlagsgesellschaft mbH, D-6940 Weinheim, 1991 0014-2980/91/0707-1649$3.50 + .25/0

1650
2 Materials and methods
K. Barrett, D. A. Taylor-Fishwick, A. P. Cope et al. Eur. J. Immunol. 1991.21: 1649-1656
2.1 Reagents
Recombinant murine and human TNF were a generous gift of Genentech Inc. (San Francisco, CA) and Upjohn (Kalamazoo, MI). The specific activities of these prepara- tions were approximately lo7 units/mg, as measured in the L929 cytotoxicity assay. 1251-labeled TNF (1251-TNF) was prepared using Iodogen (Pierce, Rockford, IL), as pre- viously described [5,27]. Both murine and human TNF were labeled to a specific activity of - 100 pCi (= 3.7 MBq)/pg.
2.2 Cell lines
2.2.1 Murine cell lines
CT6 (T cytotoxic) cells, a gift of Dr. M. Palladino, Genentech, and HT2 (Th) cells were maintained in RPMI 1640 supplemented with L-glutamine, 25 mM Hepes (Gibco, Paisley, Scotland), 5% heat-inactivated FCS (Bio- logical Industries, Glasgow, Scotland), 100 U/ml penicil- lin/streptomycin (Gibco). All T cell lines were cultured in the presence of 20 ng/ml human rIL2 (Takeda and Ajino- moto, Osaka, Japan) and 2-ME at 0.3 pM. WEHI164 clone13 fibrosarcoma cell line, a gift of Dr. A. Meager, N.I.B.S. (South Mimms, GB) was maintained in Dh4EM (Gibco) plus 5% heat-inactivated FCS, and antibiotics as above.
2.2.2 Other cell lines
The human histiocytic cell line U-937 (ICRF, London, GB) was maintained in RF'MI 1640 plus 10% FCS, and the monkey kidney fibroblast line COS 7 (ATCC, Rockville, MD) in DMEM plus 10% FCS.
2.3 Isolation of TNFR cDNA clones
Approximately 3 x lo5 phage from a murine T cell library [19] in hgtlO were transferred to nitrocellulose filters.These were screened at low stringency using a 32P-labeled Eco RI fragment containing 1272 bp of the 5' coding region of the human 55-kDaTNFR cDNA [5]. Filters were incubated at 42°C in 0.05 M sodium phosphate, pH 6.5; 20% formam- ide; 5 X SSC (1 x SSC is 0.15 M sodium chloride; 0.015 M sodium citrate); 0.01% polyvinylpyrrolidone (PVP; Sigma, St. Louis, MO); 0.1% Ficoll; 0.1% BSA (Sigma) and sonicated salmon sperm DNA (Sigma) at 50 pg/ml. Radio- labeled probe was added to the filters at 1 x 106cpm/ml and hybridized at 42°C for 16 h. Filters were washed at low stringency in 0.5 x SSC, 0.1% SDS at 37°C and positive clones identified by autoradiography. Four hybridizing clones were isolated and plaque purified. The inserts of the longest two of these clones were sequenced by the dideoxy chain-termination technique [20].
2.4 Southern and Northern blot analysis
A 950-bp EcoRI fragment comprising the 5' end of the murine TNFR cDNA was labeled with 32P to high specific
activity (2 x lo8 cpdpg) . High-molecular weight DNA was isolated [21] from WEHI 164 clone 13 cells, and digested with restriction endonucleases (New England Biolabs, Beverly, MA), run on 1% agarose and transferred to nitrocellulose by the method of Southern [22]. The filter was incubated at 42°C for 16 h in 30% formamide; 5 x SSC; 0.1% PVP; 0.1% ficoll; 0.1% BSA; 1% glycine; 50 mM sodium phosphate and 250 pg/ml sonicated salmon sperm DNA. The buffer was then exchanged for hybridi- zation buffer containing 30% formamide; 5 x SSC; 0.02% PVP; 0.02% ficoll; 0.02% BSA; 20 mM sodium phosphate; 100 pg/ml sonicated salmon sperm DNA; 10% dextran sulfate (Sigma) and probe at 1 x lo6 cpdml . Incubation was continued at 42°C for 16 h, followed by washing in 0.1 x SSC, 0.1% SDS at 65°C. The blot was then autora- diographed.
RNA was extracted from cell lines (WEHI, CTLL, CT6, HT2, EL4, 70213 and B9) using RNAzol B (Cinna Bio- tec X Labs International Inc., Friendswood,%) following the manufacturer's instructions.Tota1 RNA (20 pg) was run on a 6% formaldehyde/l% agarose gel and blotted to nitrocellulose [23]. Prehybridization was carried out at 42°C for 16 h in 50% formamide; 5 x SSC; 0.1% PVP; 0.1% Ficoll; 0.1% BSA; 50mM sodium phosphate and 250 pg/ml sonicated salmon sperm DNA.The blot was then hybridized in the same solution containing labeled probe at 1 x lo6 cpdml , at 42°C for 16 h, followed by washing in 0.2 x SSC, 0.1% SDS at 50°C and autoradiography for 2days. This blot was also hybridized under the same conditions, except for washing at 42°C in 0.5 x SSC, 0.1% SDS, with a human control probe, 7B6, the expression of which is constant throughout the cell cycle [24].
2.5 Mammalian cell expression and analysis of receptor
Clone K was isolated as a 950-bp EcoRI fragment and subcloned into a mammalian cell expression vector contain- ing the cytomegalovirus promoter and SV40 termination sequences [5]. This was transfected into the monkey fibroblast cell line COS 7 using lipofectin (Gibco) according to the manufacturers' instructions. Mock-transfected cells were treated similarly, omitting DNA. Transfected COS 7 cells were incubated for 24 h, seeded into 6-well culture plates (Nunc, Roskilde, Denmark) at 2 x lo5 cells/well and incubated for a further 48 h. Cells were washed on the plates once in cold DMEM, 10% FCS. Increasing concen- trations of 1251-TNF in 0.5 ml DMEM, 10% FCS were bound for 1.5 h at 4°C. Unbound ligand was removed by washing extensively with DMEM and 10% FCS. Cells were lysedin 1% Triton X-100 (BDH, Poole, GB), 10% glycerol, 25 mM Hepes, pH 7.4, at 37°C for 30 min and counted. Nonspecific binding was determined in the presence of 500 X molar excess of unlabeled ligand. Binding data was analyzed using the method of Scatchard [25].
2.6 Affinity c r o s s - l i g of radiolabeled TNF to its receptor
Cross-linking of 4 nM human or murine 1251-TNF to trans- fected COS7 cells or cell lines was performed using disuccinimidyl suberate (DSS; Pierce) on 4 X lo6 cells as described previously [26]. Cross-linked cells were analyzed

Eur. J. Immunol. 1991. 21: 1649-1656 Cloning and characterization of a murine p55 TNFR 1651
by SDS-PAGE [27], using 7.5% w/v polyacrylamide gels under reducing conditions, followed by autoradiography of dried gels at -70 "C using Hyperfilm M p (Amersham Int., Amersham, GB).
3 Results
3.1 Isolation and characterization of murine TNFR cDNA
The human p55 TNFR cDNAwas used to screen a murineT cell cDNA library in hgtl0. The library was screened at low stringency and four positive clones were identified. The longest two of these clones,K and 4, were sequenced and found to be overlapping, together comprising a single long open reading frame of 454 amino acids (Fig. la). The complete cDNA sequence and deduced amino acid sequence are depicted in Fig. lb.
Two start codons are observed at nucleotides 163 and 194 (Fig. lb). The first is followed 10 nucleotides later by an in-frame termination codon. Several tripeptide-coding sequences have been identified prior to the initiation codon of other receptors, including the human p55 TNFR [5].The second occurs in the context of a nucleotide sequence favorable to initiation of translation [28], followed by a termination codon at nucleotide 1557 and a potential polyadenylation signal at 2030. A hydropathy plot of the amino acid sequence reveals two hydrophobic regions consistent with a simple transmembrane protein (results not shown).The first hydrophobic region of 29 amino acids constitutes a potential signal peptide [29] and the second, a putative 23 amino acid transmembrane region, divides the extracellular and cytoplasmic domains of 183 and 219 amino acids, respectively. The protein appears to be encoded by a single gene as Southern blot analysis (Fig. 2), in which clone K was hybridized to murine genomic DNA, revealed discrete bands on Hind III and Pst I restriction digests .
3.2 Comparison of the murine and human p55 TNFR
Comparison of the amino acid sequence with that of the human homologue (Fig. 3) reveals considerable similarity between the two species, with a greater degree of homology in the extracellular domain than the cytoplasmic, 71% vs. 58% . Twenty-four highly conserved cysteine residues delin- eate four repeated sequences of approximately 40 amino acids in the extracellular domain, in common with related receptors [5]. Potential phosphorylation sites for PKC at residue 411 and for tyrosine kinase at residue 401 [7] are also conserved between the human and murine TNFR. Minor differences occur between the three potential N- linked glycosylation sites, with only two of the three sites between human and murine receptors being conserved.
This truncated receptor encodes the first 250 amino acids of the murine TNFR, including the extracellular and trans- membrane domains, and 16 amino acids of the intracellular domain. After 3 days, the ability of the expressed protein to bind human and murine 1251-TNF was assessed. The trans- fected cells specifically bound labeled TNF of both species with high affinity, as shown in Fig. 4.The binding affinity of this receptor for murine and human ligands was comparable at Kd - 3 nM (Fig. 4). No specific binding of 1251-TNF of either species (data not shown) to mock-transfected COS 7 cells was detected.
3.4 Affinity cross-linking of recombinant receptor
The truncated recombinant receptor contains 55% of the coding region of the cDNA and three potential N-linked glycosylation sites. Therefore, the predicted size is likely to be in the range 30-40 kDa. This recombinant receptor construct was expressed on COS 7 cells and affinity cross- linked to both murine and human 1251-TNF (Fig. 5). Both ligands gave labeled products of approximately 54, 70 and 85 kDa on SDS-PAGE analysis, compatible with linking of a truncated receptor of -35 kDa to TNF monomers, dimers and trimers, of unit molecular mass 17 kDa. The additional 34-kDa product observed is consistent with a homodimer of TNF. Cross-linking of labeled murine TNF was effectively inhibited by excess cold ligand of both species (Fig. 5, lanes b and c), and excess unlabeled human TNF blocked cross-linking of radiolabeled human TNF (Fig. 5, lane e). No cross-linking was observed between either species of TNF and mock-transfected COS cells (Fig. 5, lanes f-j), or untreated COS cells (data not shown).
3.5 Analysis of mRNA of various murine cell lines
To study the distribution of this receptor in different cell types, we carried out Northern blot analysis using RNA isolated from various murine cell lines (Fig. 6). These included the fibroblast line, WEHI 164, which is used for TNF cytotoxicity assays [15], several T cell lines (CT6, CTLL, EL4, HT2) and lines of B cell lineage (B9 and 702/3). A 2.4-kb transcript was observed in the fibroblast line, and three of the four Tcell lines. However, mRNA for this receptor was only faintly visible in the T cell line CT6 and the B cell line B9 on four times-longer exposure of the autoradiograph.The pre-B cell line, 702/3, did not produce detectable message for this receptor. Hybridization of the blot with a control probe, 7B6, indicated that equal amounts of RNA were loaded in each track (data not shown). The cDNA clones isolated comprise 2063 nucleo- tides in total, which is slightly shorter than the message size estimated from the Nothern blot. These clones may, therefore, lack some of the untranslated region or poly(A) tail.
3.3 Expression of recombinant receptor cDNA in COS7 3.6 Affiity cross-linking of cell lines
Radiolabeled TNF was cross-linked to the natural receptor expressed on several cell lines, and multiple labeled products were observed by SDS-PAGE analysis (Fig. 7). Affinity cross-linking of murine lZ5I-TNF to the WEHI 164
cells
Clone K was subcloned into a mammalian cell expression vector containing a cytomegalovirus promoter and SV40 termination sequences and transfected into COS 7 cells.

1652
A SIG 1 2 3 4 TM CYT
K. Barrett, D. A. Taylor-Fishwick, A. P. Cope et al. Eur. J. Immunol. 1991. 21: 1649-1656
c In+xisssK\\ An
m
v) E
z x Q X m I 0 m o - c > c m -
A
I I I 1 I I Lt
A, 500 bp
B 1 TGAGGGGAGAGAGGTCCTGGAGGACCGTACCCTGATCTCTATCTGCCTCTGACTTTCAGCTTCTCGAA~CGAGG
7 6 CCCAGGCTGCCATCGCCCGGGCCACCTGGTCCGATCATCTTACTTCATTCACGAGCGTTGTCMTTGCTGCCCTGTCCCCAGCCCCMTGGGGGAGTGAGAGGC~CTGCCGGCCGGAC
1 L 1 9 4 ATG GGT CTC CCC ACC GTG CCT GGC CTG CTG CTG TCA CTC GTG CTC CTG GCT CTG CTG ATG CGG ATA CAT CCA TCA GGG GTC ACT GGA CTA
3 1 V P S L G D R E K R D S L P Q G K Y V H S K N N S I C C T 2 8 4 GTC CCT TCT CTT GGT GAC CGG GAG M G AGG GAT AGC TTG TGT CCC CAA GGA AAG TAT GTC CAT TCT M G M C AAT TCC ATC TGC TGC ACC
1 1
6 1 K C H K G T Y L V S D C P S P G R D T V C R E m m C E K G T F T , 3 7 4 AAG TGC CAC AAA GGA ACC TAC TTG GTG AGT GAC TGT CCG AGC CCA GGG CGG GAT ACA GTC TGC AGO GAG TGT GAA AAG GGC ACC TTT ACG
9 1 A S Q N Y L R Q C L S C K T C R K E H S Q V E I S P C Q A D 464 GCT TCC CAG AAT TAC CTC AGG CAG TGT CTC ACT TGC AAG ACA TGT CGG M A GAA ATG TCC CAG GTG GAG ATC TCT CCT TGC CAA GCT GAC
1 2 1 m ' C K E N Q F Q R Y L S E T H F Q C V D C S P C F 5 5 4 AAG GAC ACG GTG TGT GGC TGT AAG GAG AAC CAG TTC CAA CGC TAC CTG AGT GAG ACA CAC TTC CAG TGC GTG GAC TGC AGC CCC TGC TTC
1.
1 5 1 N G T V T I P C K E T Q N T V C N . 5 H A G F F L R E S E C V 6 4 4 M C GGC ACC GTG ACA ATC CCC TGT M G GAG ACT CAG AAC ACC GTG TGT M C TGC CAT GCA GGG TTC TTT CTG AGA GAA AGT GAG TGC GTC
181 P C S H C K K N E E C M K L C L P P P L A N 'V T N P Q D S G 7 3 4 CCT TGC AGC CAC TGC AAG AAA AAT GAG GAG TGT ATG M G TTG TGC CTA CCT CCT CCG CTT GCA AAT GTC ACA AAC CCC CAG GAC TCA GGT
211 T A R Y P R W 824 ACT GCG GTG CTG TTG CCC CTG GTT ATC TTG CTA GGT CTT TGC CTT CTA TCC TTT ATC TTC ATC AGT TTA ATG TGC CGA TAT CCC CGG TGG
2 4 1 R P E V Y S I I C R D P V P V K E E K A G K P L T P A P S P 9 1 4 AGG CCC GAA GTC TAC TCC ATC ATT TGT AGG GAT CCC GTG CCT GTC M A GAG GAG AAG GCT GGA AAG CCC CTA ACT CCA GCC CCC TCC CCA
2 7 1 A F S P T S G F N P T L G F S T P G F S S P V S S T P I S P 1 0 0 4 GCC TTC AGC CCC ACC TCC GGC TTC AAC CCC ACT CTG GGC TTC AGC ACC CCA GGC TTT AGT TCT CCT GTC TCC AGT ACC CCC ATC AGC CCC
3 0 1 1 F G P S N W H F H P P V S E V V P T Q G A D P L L Y E S L 1 0 9 4 ATC TTC GGT CCT ACT M C TGG CAC TTC ATG CCA CCT GTC AGT GAG GTA GTC CCA ACC CAG GGA GCT GAC CCT CTG CTC TAC GAA TCA CTC
3 3 1 C S V P A P T S V Q K W E D S A H P Q R P D N A D L A I L Y 1 1 8 4 TGC TCC GTG CCA GCC CCC ACC TCT GTT CAG M A TGG GAA GAC TCC GCC CAC CCG CAA CGT CCT GAC AAT GCA GAC CTT GCG ATT CTG TAT
3 6 1 A V V D G V P P A R W K E F M R F H G L S E H E I E R L E H 1 2 7 4 GCT GTG GTG GAT GGC GTG CCT CCA GCG CGC TGG AAG GAG TTC ATG CGT TTC ATG GGG CTG AGC GAG CAC GAG ATC GAG AGG CTG GAG ATG
3 9 1 0 N G R C L R E A 0 Y S M L E A W R R R T P R H E D T L E V 1 3 6 4 CAG AAC GGG CGC TGC CTG CGC GAG GCT CAG TAC AGC ATG CTG GAA GCC TGG CGG CGC CGC ACG CCG CGC CAC GAG GAC ACG CTG GAA GTA
4 2 1 V G L V L S K M N L A G C L E N I L E A L R N P A P S S T T 1 4 5 4 GTG GGC CTC GTG CTT TCC AAG ATG M C CTG GCT GGG TGC CTG GAG AAT ATC CTC GAG GCT CTG AGA AAT CCC GCC CCC TCG TCC ACG ACC
4 5 1 R L P R 1 5 4 4 CGC CTC CCG CGA TAA A G C ~ ~ C C C A C M C C T T A ~ G A ~ ~ f f T C ~ G A ~ T C ~ G C T A G A T G C C C T A C T C C C ~ ~ G T ~ G T G ~ T C ~ M 1658 G G G G G A A G G C T C G A G C T G G T A C C A C T T C C T T G G T G C T A C C G A C T T G G T ~ A C A T A G ~ ~ T ~ C A G ~ G C C G A ~ C T G C C T G A ~ C A G C ~ C ~ G T ~ G T ~ ~ ~ T G T ~ C A 1 7 7 7 T C A G C T C C T G G C C A G C T G A C G T G C ~ G A C A G G A T T G T A G A G ~ G ~ ~ T G T A T C T G G T G C C C A C T T G ~ T G C A C A G ~ C C C A A G C C A A G C T T C T C A G G G C C ~ ~ A ~ G G
2015 G A C T C T T C T A M T h C A C T A M C G T ~ ~ T ~ 1896 GTTTCTGGGCCTTTTTCACTTTTGATMG~TCTTTGTATCAATTA~ATCACACTAATGGATGAACTGTGTAAGGTAAGGACAAGCATA~GGC~GGTCTCCAGCTGGAGCCCTC

Eur. J. Immunol. 1991. 21: 1649-1656
a b
23,130 - 9,419 - 6,557 - 4.317 - 3,054 - 2,036 - 1,636 -
1.018 -
510 -
MURINE
HUMAN
MURINE
HUMAN
MURINE
HUMAN
MURINE
HUMAN
MURINE
HUMAN
MURINE
HUMAN
MURINE
HUMAN
MURINE
HUMAN
MURINE
n w
Figure 2. Southern blotting of murine genomic DNA probed with murine p55 receptor cDNA. Genomic DNA ( 5 pg) from the WEHI164 cell line was digested with restriction enzymes, (a) Hind111 and (b) PstI, run on a 1% agarose gel and blotted to nitrocellulose. Hybridization was carried out at high stringency using a 32P- labeled cDNA probe for h u - man p55 receptor.
Cloning and characterization of a murine p55 TNFR 1653
line identified complexes of 75, 95 and 130 kDa (Fig. 7, lane a). On the HT2 line, similar products were also seen, but with slightly higher apparent molecular mass of 80,100 and 135 kDa, respectively (Fig. 7, lane e).The CT6cell line only displayed a 100- and 135-kDa complex, even after overexposure of the autoradiograph. Specificity of cross- linking was verified by complete inhibition using excess unlabeled murine TNF (Fig. 7, lanes b , d and f). Cross- linking of human lZ5I-TNF to receptors from WEHI 164 cells with DSS resulted in a faint but detectable labeled 75-kDa complex (data not shown). These results are consistent with the cross-linking of monomeric murineTNF to a p55 (- 75 kDa) and p75 (-95 kDa) receptor and monomeric human TNF to a p55 (- 75 kDa) receptor only. Furthermore, the failure to detect a p55 cross-linked product from CT6 cells correlates with the low mRNAleve1 observed by Northern analysis (Fig. 6).
Figure3. Comparison of the amino acid sequences of murine and human p55 TNFR by the method of Lipman and Pearson [31]. Matching amino acids are printed between the two aligned sequences. Amino acids with a positive score on the PAM matrix are shown with a colon (:), those that are negatively related show a blank and those with a zero-value relationship are shown with a period (.). Transmembrane domain and signal peptide are in shaded boxes, conserved cysteine residues in open boxes and potential N-Linked glycosylation sites are overlined.
4 Figure 1 . Nucleotide sequence, predicted amino acid sequence and localization of cDNA clones of murine p55 TNFR. (A) Schematic representation of murine p55 TNFR cDNA. The 5’ untranslated region (left) is followed by the signal sequence (SIG), the extracellular domain containing four cysteine-rich repeats [l-41, the transmembrane domain (TM) and cytoplasmic domain (CYT). The two cDNA clones (K and 4) are aligned with respect to the schematic map. (B) Nucleotide and deduced amino acidsequence of the cDNA for murine p55 TNFR. Consensus signal sequence and transmembrane domain are boxed, the four cysteine-rich repeats are overlined and the presumed poly-adenylation signal underlined.
1654 K. Barrett, D. A. Taylor-Fishwick, A. P. Cope et al.
5-
Eur. J. Immunol. 1991.21: 1649-1656
0
A B
2 0 1 2 3 4 5 6 7 8
Mu TNF bound (fmoles)
K D a a b c d e f g h i j
200-
97.5-
69-
46-
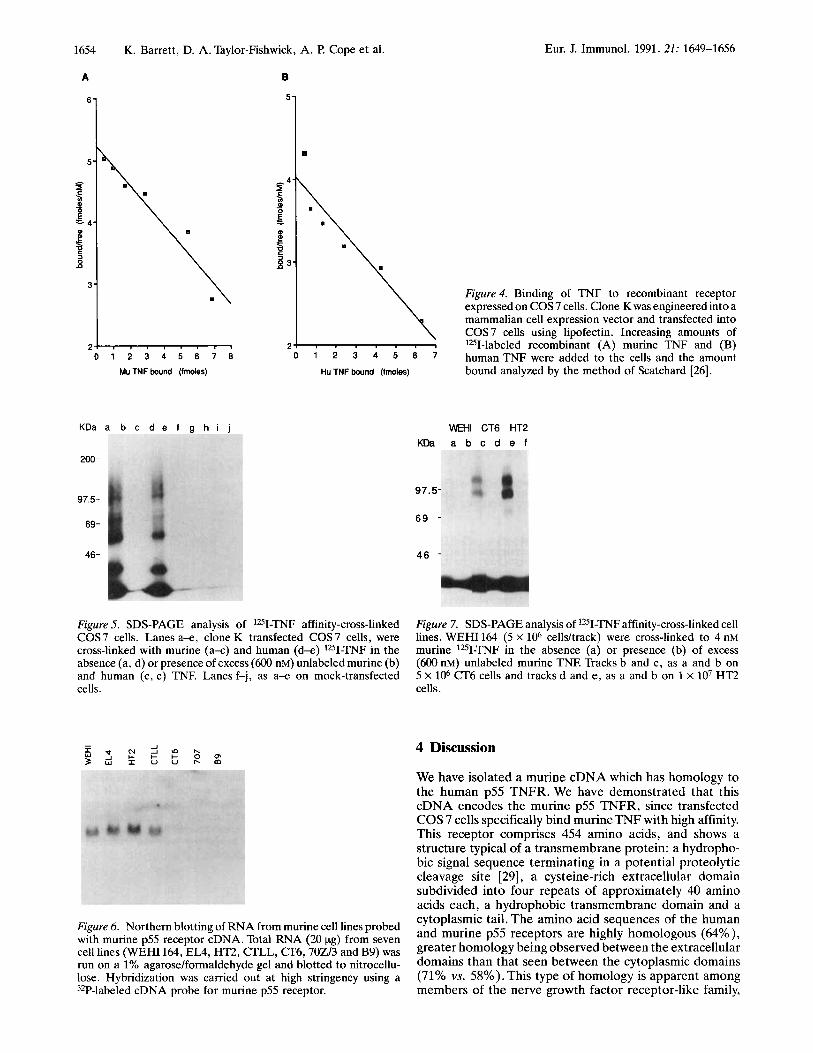
Figure 4. Binding of TNF to recombinant receptor expressed on COS 7 cells. Clone K was engineered into a mammalian cell expression vector and transfected into COS 7 cells using lipofectin. Increasing amounts of lZ5I-labeled recombinant (A) murine TNF and (B)
0 * 3 5 8 7 humanTNF were added to the cells and the amount bound analyzed by the method of Scatchard [26]. Hu TNF bound (fmoles)
Figure 5. SDS-PAGE analysis of lZ5I-TNF affinity-cross-linked COS7 cells. Lanes a x , clone K transfected COS7 cells, were cross-linked with murine (a-c) and human (d-e) 1251-TNF in the absence (a, d) or presence of excess (600 nM) unlabeled murine (b) and human (c, e) TNF. Lanes f-j, as a-e on mock-transfected cells.
Figure 6. Northern blotting of RNA from murine cell lines probed with murine p55 receptor cDNA. Total RNA (20 pg) from seven cell lines (WEHI 164, EL4, HT2, CTLL, CT6,702/3 and B9) was run on a 1% agarose/formaldehyde gel and blotted to nitrocellu- lose. Hybridization was carried out at high stringency using a 32P-labeled cDNA probe for murine p55 receptor.
WEHl CT6 HT2 KDa a b c d e f
97.5-
69 -
4 6 -
Figure 7. SDS-PAGE analysis of lZ5I-TNF affinity-cross-linked cell lines. WEHI 164 (5 X lo6 cellsltrack) were cross-linked to 4 nM murine 1251-TNJ? in the absence (a) or presence (b) of excess (600 nM) unlabeled murine TNE Tracks b and c, as a and b on 5 X lo6 CT6 cells and tracks d and e , as a and b on 1 x lo7 HT2 cells.
4 Discussion
We have isolated a murine cDNA which has homology to the human p55 TNFR. We have demonstrated that this cDNA encodes the murine p55 TNFR, since transfected COS 7 cells specifically bind murineTNF with high affinity. This receptor comprises 454 amino acids, and shows a structure typical of a transmembrane protein: a hydropho- bic signal sequence terminating in a potential proteolytic cleavage site [29], a cysteine-rich extracellular domain subdivided into four repeats of approximately 40 amino acids each, a hydrophobic transmembrane domain and a cytoplasmic tail. The amino acid sequences of the human and murine p55 receptors are highly homologous (64%), greater homology being observed between the extracellular domains than that seen between the cytoplasmic domains (71% vs. 58%). This type of homology is apparent among members of the nerve growth factor receptor-like family,

Eur. J. Immunol. 1991.21: 1649-1656
which are characterized by the well-conserved cysteine-rich repeat sequences described above (discussed in [5]).
Cloning and characterization of a murine p55 TNFR 1655
The mechanisms by which TNFexerts its pleiotropic effects are still not clear.Various mechanisms for cytotoxicity have been proposed in man (reviewed in [34]). InTNF-mediated killing, antibody studies have suggested a requirement for p55 receptor [ l l , 351 or other cellular components, for features Of the TNFR are 'Onserved between
species, potential phosphorylation sites in the cytoplasmic domain. A potential pKc site is located On
example the Fas protein [36]. Antibodies recognizing the human p55 receptor are able to induce cytotoxicity of the
residue 411 of both receptors, and its conservation would suggest that this site is involved in receptor function. PKC has been shown to modulate the affinity of the human TNFR on PBL [30]. Initial studies on recombinant human receptor demonstrated that, while a truncated version lacking the C-terminal 44 amino acids, and therefore the
u937 fibroblast line, and proliferation of Fs4 fibroblasts ~351. The involvement of the p55 receptor in human TNF-mediated cytotoxicity is supported by our study, in which the murine p55 mFR, which does not discriminate between human and mufine TNF binding, appears to mediate the cytotoxic effects to WEHI 164 cells. Moreover,
phosphorylation site, has a high affinity for the ligand L 5 3 6 1 9 a full-length recombinant receptor exhibited
using the truncated murine
cT6 cells, which proliferate only to murine TW, do not express detectable p55 TNFR on the cell surface. Thus, it Seems likely that the p75 T W R on c T 6 cells transduces the affinity I71. Our
receptor cDNA, clone K (Fig. 4), also identified a single affinity for a truncated receptor. Further analysis is in progress to define the contribution of PKC phosphorylation and truncation to receptor function.
The difference between the species specificity of TNF binding and activity on C T ~ and W E ~ 164 cells implies the existence of more than one murine TNFR. T~~ human TNFR have been identified, and we have also isolated cDNA clones corresponding to the murine homologue of the human P75 TNFR (manuscript in Preparation) - Since the recombinant murine p55 receptor bound both murine and human TNF with equal affinity, and a previous study indicated that CT6 cells did not respond to human TNF [13], it was considered unlikely that this receptor would be expressed on CT6 cells. Consistent with this, mRNA levels for the p55 receptor were considerably lower in CT6 cells than in other murine T cell lines tested and in WEHI 164 cells. Furthermore, no complex corresponding to a putative p55 receptor was detectable on these cells after affinity cross-linking. In contrast, such a complex was present on W E ~ 164 and H= cells. (36 cells did display a 1O()-kDa complex, which would be compatible with the expression of p75 TNFR. Again, this band was detectable on WEHI 164 and HT2 The is in agreement with the affinity cross-linking of two receptors similar in size to the human p55 and p75 receptors [lo]. As expected, cross-linking with human 1251-TNF labeled the p55 protein on WEHI 164 cells, but a similar band was not detectable on c T 6 cells. The other proteins which labeled
These results suggest that the murine p75 TNFR binds only murine TNF.
Cross-linking studies also detected a 130-135-kDa complex on all cell lines investigated. Complexes larger in size than a single receptor bound to a TNF mommer have also been identified on HeLa cells [32].These could be resolved into monomeric receptor and ligand, although the presence of other accessory molecules was not excluded. A complex consisting of the p75 receptor and a trimer of TNF (51 kDa)
give a molecular close to the 130-kDa product seen here, but it is surprising that no - 110-kDa p75/TNF dimer was Observed' the 130-kDa product could be the result of cross-linking of an accessory molecule of - 110-115 kDa associated with the P75 recep- tor, analogous to the 130-kDa molecule associated with the
proliferative signal.
The cloning, characterization and expression of the mufine p55 TNFR described here, and of the p75 TNFR will facilitate studies of their differential function, and of TNFR signal transduction. It will also facilitate studies of the role of TNF in animal models of disease, and the generation and testing of therapeutics based on this cytokine and its receptors.
We thank Genentech Inc. and The Upjohn Company for supplying recombinant T N e Anne Hales for growing cell lines, Y Chernajovs- ky and A . Corcoran for helpful comments on the manuscript. cDNA sequence submitted to EMBL Database, Accession number x59238.
Received March 22, 1991.
References
1 Beutler, B. and Cerami, A. , N . Engl. J. Med. 1987. 316: 379.
2 Tracey, K. J., Vlassara, H. and Cerami, A . , Lancet 1989. i: 1122.
3 Agganval, B. B., Eessalu, T. E. and Hass, €? E. , Nature 1985. 318: 665.
4 Hohmann, H. , Remy, R., Brockhaus, M. and van Loon, A. F! G. M., J. Biol. Chem. 1989. 264: 14927.
5 Gray, P. W., Banett, K., Chantry, D. H. . Turner, M. and Feldmann, M., Proc. Natl. Acad. Sci. USA 1990. 87: 7380.
6 Loetscher, H. , Pan, Y-C. E. , Lahm, H.-W., G e n e R.3
Brockhaus, M., Tabuchi, H. and Lesslauer, W., Cell 1990. 61: 351.
7 Schall, T. J., Lewis, M., Koller, K. J., Lee, A. , Rice, G. C., Wong, G. H. W., Gatanaga, T., Granger, G. A. , Lentz, R., Raab, H. , Kohr, W. K. and Goeddel, D. V., Cell 1990. 61: 361.
8 Smith, c. A. , ~ ~ ~ i ~ , T., Anderson, D., solam, L., ~ ~ ~ k ~ ~ ~ ~ , M. F!, Jerzy, R. , Dower, S. K., Cosman, D. and Goodwin, R. G., Science 1990. 248: 1019.
9 DembiC, Z. Loetscher, H., Gubler, U., Pan,Y.-C. E. , Lahm, H.-W., Gentz, R. , Brockhaus, M. and Lesslauer,W., Cytokine 1990.2: 2 3 .
10 Brockhaus, M., Schoenfeld, H. J., Schlaeger, E. J. , Hunziker, W., Lesslauer,W. and Loetscher, H. , Proc. Natl. Acad. Sci. USA 1990. 87,. 3127.
11 Thoma, B., Grell, M., Pfizenmaier, K. and Scheurich, P., J. E ~ ~ . Med. 1990. 172: 1019.
12 Sugarman, B. J., Aggarwal, B. B., Hass, P. E., Figari, I. S., Palladino, M. A. , Jr. and Shepard, H. M. , Science 1985.230:
data On these
with murine TNF, were not cross-linked to human TNE
IL 6R [33]. 943.

1656 K. Barrett, D. A. Taylor-Fishwick, A. P. Cope et al. Eur. J. Immunol. 1991.21: 1649-1656
13 Ranges, G. E., Bombara, M. €?, Aiyer, R. A., Rice, G. G. and Palladino, M. A., Jr., J. Irnmunol. 1989. 142: 1203.
14 Ranges, G. E., Zlotnik, A., Espevik,T., Dinarello, C., Cerami, A. and Palladino, M. A., Jr., J. Exp. Med. 1988. 167: 1472.
15 Espevik, T. and Nissen-Meyer, J., J. lmrnunol. Methods 1986. 95: 99.
16 Hofman, F. M., Hinton, D. R., Johnson, K. and Merrill, J. E., J. Exp. Med. 1989. 170: 607.
17 Ruddle, N. H., Bergman, C. M., McGrath, K. M., Lingenheld, E. G., Grunnet, M. L., Padula, S. J. and Clark, R. B., J. Exp. Med. 1990. 172: 1193.
18 Brennan, F. M., Chanty, D., Jackson, A., Maini, R. N. and Feldmann, M., Lancet 1989. ii: 244.
19 Gray, P.W., Leong, S., Fennie, E. H., Farrar, M. A., Pingel, J. T., Fernandez-Luna, J. and Schreiber, R. D., Proc. Natl. Acad. Sci. USA 1989. 86: 8497.
20 Smith, A. J. H., Methods Enzyrnol. 1980. 65: 560. 21 Sambrook, J., Fritsch, E. F. and Maniatis, T., Molecular
Cloning, a Laboratory Manual, 2nd Edn., Cold Spring Harbor Laboratory Press, New York 1989, p. 9.16.
22 Southern, E. M., J. Mol. Biol. 1975. 98: 503. 23 Dobner, P. R., Kawasaki, E. S.,Yu, L.Y. and Bancroft, F. C.,
Proc. Natl. Acad. Sci. USA 1981. 78: 2230.
24 Kaczmarek, L., Calabretta, B. and Baserga, R., Proc. Natl.
25 Scatchard, G., Ann. N.Y Acad. Sci. 1949. 51: 660. 26 Foxwell, B. M. J.,Woerly, G. and Ryffel, B., Eur. J. Irnrnunol.
27 Laemmli, U. K., Nature 1970. 227: 680. 28 Kozak, M., J. Cell Biol. 1989. 108: 229. 29 Von Heijne, G., Nucleic Acids Res. 1986. 14: 4683. 30 Unglaub, R. B., Maxeiner, B.,Thoma, K., Pfizenmaier, K. and
31 Lipman, D. J. and Pearson, W. R., Science 1985. 227: 1435. 32 Smith, R. A. and Baglioni, C., J. Biol. Chern. 1989. 264:
14 646. 33 Taga, T., Hibi, M., Hirata, Y., Yamasaki, K., Yasukawa, K.,
Matsuda, T., Hirano, T. and Kishimoto, T., Cell 1989. 58: 573.
Acad. Sci. USA 1985. 82: 5375.
1989. 19: 1637.
Scheurich, P., J. Exp. Med. 1987. 167: 1778.
34 Lamck, J. W. and Wright, S. C., FASEB J. 1990. 4: 3215. 35 Espevik, T., Brockhaus, M., Loetscher, H., Nonstad, U. and
36 Yonehara, S., Ishii, A. and Yonehara, M., J. Exp. Med. 1989. Shalaby, R., J. Exp. Med. 1990. 171: 415.
169: 1747.