cilia and ciliopathies in congenital heart disease
TRANSCRIPT

Cilia and Ciliopathies in Congenital HeartDisease
Nikolai T. Klena, Brian C. Gibbs, and Cecilia W. Lo
Department of Developmental Biology, University of Pittsburgh School of Medicine, Pittsburgh,Pennsylvania 15201
Correspondence: [email protected]
A central role for cilia in congenital heart disease (CHD) was recently identified in a large-scale mouse mutagenesis screen. Although the screen was phenotype-driven, the majority ofgenes recovered were cilia-related, suggesting that cilia play a central role in CHD patho-genesis. This partly reflects the role of cilia as a hub for cell signaling pathways regulatingcardiovascular development. Consistentwith this, manycilia-transducedcell signalinggeneswere also recovered, and genes regulating vesicular trafficking, a pathway essential for cilio-genesis and cell signaling. Interestingly, among CHD-cilia genes recovered, some regulateleft–right patterning, indicating cardiac left–right asymmetry disturbance may play signifi-cant roles in CHD pathogenesis. Clinically, CHD patients show a high prevalence of ciliarydysfunction and showenrichment for de novo mutations in cilia-related pathways. Combinedwith the mouse findings, this would suggest CHD may be a new class of ciliopathy.
Congenital heart disease (CHD) is one of themost common birth defects, found in an
estimated 1% of live births (Hoffman andKaplan 2002). With advances in surgical pallia-tion, most patients with CHD now survive theircritical heart disease such that currently thereare more adults with CHD than infants bornwith CHD each year (van der Bom et al.2012). However, CHD patient prognosis is var-iable, with long-term outcome shown to be de-pendent on patient intrinsic factors rather thansurgical parameters (Newburger et al. 2012;Marelli et al. 2016). This is likely driven by ge-netic factors, given CHD is highly associatedwith chromosomal anomalies (Fahed et al.2013), and with copy number variants (Gless-ner et al. 2014). In addition, CHD has been
shown to have a high recurrence risk, with fa-milial clustering indicating a genetic contribu-tion (Gill et al. 2003; Oyen et al. 2009). Theidentification of the genetic causes of CHDmay provide mechanistic insights that canhelp stratify patients for guiding the therapeuticmanagement of their clinical care.
Investigations into the genetic causes ofCHD in human clinical studies have been chal-lenging given the high degree of genetic diver-sity in the human population. This has made acompelling case for pursuing the use of a sys-tems genetic approach with large-scale forwardgenetic screens in animal models to investigatethe genetic etiology of CHD. Although manyanimal models have provided invaluable in-sights into the developmental regulation of car-
Editors: Wallace Marshall and Renata Basto
Additional Perspectives on Cilia available at www.cshperspectives.org
Copyright # 2017 Cold Spring Harbor Laboratory Press; all rights reserved
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
1
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

diovascular development, investigations intothe genetic etiology of CHD must be conductedin a model system with the same four-chambercardiac anatomy that is the substrate of humanCHD. The mouse is one such model system,advantageous not only given its similar four-chamber cardiac anatomy, but also inbredmouse strains are readily available with ge-nomes that are fully sequenced and annotatedthat would facilitate mutation recovery. More-over, cardiovascular development in the mouseembryo is well studied, providing a strong foun-dation to interrogate the developmental and ge-netic etiology of CHD.
DEVELOPMENT OF THE CARDIOVASCULARSYSTEM
Congenital heart defect is a structural birth de-fect arising from disruption of cardiovasculardevelopment. Formation of the four-chamberheart in mammals is orchestrated by the highlycoordinated specification and migration of dif-ferent cell populations in the embryo that to-gether form the complex left–right asymmetricanatomy of the cardiovascular system. In themouse embryo, ingression of cells through theprimitive streak at E7.5 generates the anteriormesoderm forming the cardiac crescent–con-taining cells of the first heart field (FHF) andadjacent to it, the second heart field (SHF) (Fig.1) (Buckingham 2016). Cells of the FHF mi-grate toward the midline, fusing to form thelinear heart tube at E8.0 (Fig. 1). Pharyngealmesoderm located anterior and medially con-tinues to be added to the expanding heart tube,as the heart tube undergoes rightward loopingat E8.5, delineating the primitive anlage of theleft ventricle (LV) (Fig. 1). This is followed byaddition of SHF cells to the anterior and poste-rior poles of the heart tube, giving rise to theoutflow tract (OFT), right ventricle (RV), andmost of the left and right atria (LA, RA) (Fig. 1).
Normal development of the heart also re-quires the contribution and activity of severalother extracardiac cell lineages, including thecardiac neural crest cells derived from the dorsalhindbrain neural fold. The cardiac neural crestcells migrate into the cardiac OFT in two spiral
streams, helping to remodel the pharyngeal archarteries and orchestrating OFT septation toform the two great arteries—the aorta and pul-monary artery (Kirby and Waldo 1990). Thepharyngeal endoderm and ectoderm also playan important regulatory function in develop-mental patterning of the aortic arch arteriesand the OFT. Dynamic processes mediating en-docardial epithelial–mesenchyme transforma-tion (EMT) lead to formation of the cushionmesenchyme that provides early valve functionin the embryonic heart. These endocardial cush-ion tissues later remodel to form the matureleaflets of the outflow semilunar and atrioven-tricular valves (Fig. 1). Another extracardiac cellpopulation required for heart development arethe pro-epicardial cells that originate near theseptum transversum. These cells migrate to theheart via the sinus venosus, delaminating ontothe surface of the heart, and forming the epicar-dium that plays an essential role in developmentof the coronary arteries. Together, these diversecell populations are recruited to orchestrate for-mation of the mammalian heart, an organ thatis an unexpected mosaic of distinct cell lineages.
FOUR-CHAMBER HEART—THEANATOMICAL SUBSTRATE FORCONGENITAL HEART DISEASE
The cardiovascular system in mouse and humanis adapted for breathing air, being comprised offour chambers organized into functionally dis-tinct left versus right sides. This allows the for-mation of a separate pulmonary circuit thatpumps deoxygenated blood from the body tothe lungs via the RV and a systemic circuitpumping oxygenated blood from the lung tothe body via the LV. This left–right asymmetricorganization is critically dependent on appro-priate patterning of the left–right body axis andentails formation of an atrial and ventricularseptum separating the right versus left sides ofthe heart. This allows for compartmentalizationof the heart into four chambers, LA versus RAand LV versus RV. This is coupled with septationof the OFT into two great arteries, the aorta,which is inserted into the LV and pulmonaryartery into the RV, and formation of the atrio-
N.T. Klena et al.
2 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Sec
ond
hear
t fie
ld
Ven
ous
pole
Ven
tric
le
Atr
ial
pole
Firs
the
art f
ield
A DE
F
BC
AV
cana
l
Atr
ium
Ven
tric
le
E8.
5 H
eart
loop
ing
E9.
5 A
V c
anal
form
atio
nE
8.0
Line
ar h
eart
tube
E7.
5 C
ardi
ac c
rese
nt
EC
cus
hion
sC
utflo
wtr
act
E10
.5 O
FT
rem
odel
ing
E13
.5 M
atur
e he
art
Left
vent
ricle
Left
atriu
m
Aor
taP
ulm
onar
yar
tery
Rig
htat
rium
Rig
htve
ntric
le
E9.
5 E
C fo
rmat
ion
E10
.5 T
rabe
cula
tion
E10
.5 S
epta
tionTra
becu
latio
nV
entr
icul
arse
ptat
ion
Figu
re1.
Dia
gram
ofm
ou
seca
rdio
vasc
ula
rd
evel
op
men
t.(A
)C
ard
iac
cres
cen
tfo
rmat
ion
con
tain
ing
firs
th
eart
fiel
d(F
HF
)an
dse
con
dh
eart
fiel
d(S
HF
)ce
lls.
(B)
Car
dia
ccr
esce
nt
cell
sm
igra
teto
war
dth
em
idli
ne
crea
tin
gth
eli
nea
rh
eart
tub
ew
ith
its
arte
rial
and
ven
ou
sp
ole
san
da
pri
mit
ive
ven
tric
le.(
C)
AtE
8.5,
dex
tral
loo
pin
go
fth
eh
eart
tub
ele
adst
ofo
rmat
ion
oft
he
pri
mit
ive
atri
alan
dve
ntr
icu
lar
cham
ber
sin
the
mo
rph
olo
gica
lly
corr
ect
po
siti
on
.(D
)A
tE
9.5,
the
end
oca
rdia
lcu
shio
nce
lls
pin
chin
war
d,
crea
tin
gth
eat
riov
entr
icu
lar
can
al.
At
E9.
5,th
een
do
card
ial
cush
ion
sfo
rmat
the
do
rsal
and
ven
tral
lum
eno
fth
eat
rial
can
alas
the
end
oca
rdia
lce
lls
un
der
goep
ith
elia
lto
mes
ench
ymal
tran
siti
on
.C
ard
iac
trab
ecu
lati
on
init
iate
sat
E10
.5,c
reat
ing
bu
nd
les
ofc
ard
iom
yocy
tes
that
exte
nd
into
the
pri
mit
ive
card
iac
cham
ber
s.Se
pta
tio
nin
itia
tes
atE
10.5
,sta
rtin
gd
ivis
ion
oft
he
cham
ber
sin
toth
efo
ur-
cham
ber
anat
om
y.(E
)A
tE10
.5,t
he
ou
tflow
trac
t(O
FT
)is
rem
od
eled
lead
ing
toth
ep
rim
itiv
eco
nn
ecti
on
oft
he
aort
aan
dp
ulm
on
ary
arte
ryfr
om
the
pri
mit
ive
ven
tric
le.(
F)
By
E13
.5,t
he
hea
rtis
full
yd
evel
op
edin
tofo
ur
dis
tin
ctch
amb
ers
wit
hap
pro
pri
ate
aort
aan
dp
ulm
on
ary
arte
ryco
nn
ecti
on
sto
the
mo
rph
o-
logi
call
efta
nd
righ
tve
ntr
icle
s(R
Vs)
,res
pec
tive
ly.D
ark
pin
k,F
HF
;lig
ht
pin
k,SH
F;l
igh
tgre
en,a
trio
ven
tric
ula
r(A
V)
can
al;
dar
kb
lue,
end
oca
rdia
l(E
C)
cush
ion
s;ye
llow
gree
n,
sep
tati
on
;p
urp
le,
trab
ecu
lati
on
s;ye
llow
,O
FT.
Cilia and Ciliopathies in Congenital Heart Disease
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266 3
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

ventricular and outflow valves that allow unidi-rectional blood flow. It is this complex left–right asymmetric developmental patterning ofthe cardiovascular anatomy that ensures effi-cient oxygenation of blood with air exchangevia the lungs. The perturbation of this distinctfour-chamber cardiac anatomy in CHD invari-ably results in neonatal mortality unless surgicalintervention is provided to palliate the struc-tural heart defects. Identifying the genetic caus-es of CHD may help elucidate the developmen-tal processes contributing to CHD and suggestsnew avenues for prevention or intervention.
CENTRAL ROLE OF CILIA INCARDIOVASCULAR DEVELOPMENT ANDCONGENITAL HEART DISEASE
To elucidate the genetic etiology of CHD, alarge-scale, near-saturation level forward genet-ic screen with ethylnitrosourea (ENU) chemicalmutagenesis was conducted (Li et al. 2015b).This phenotype-driven cardiovascular screenused fetal echocardiography, a noninvasive im-aging modality routinely used clinically forCHD diagnosis (Fig. 2). This allowed high de-tection sensitivity and specificity for CHD di-agnosis and allowed the recovery of a wide spec-trum of CHD in the mouse screen similar tothose observed clinically (Figs. 2 and 3) (Liuet al. 2014). From ultrasound screening of�100,000 mouse fetuses, we recovered .200mutant mouse lines with a wide variety of CHD.
Using exome-sequencing analysis, �100CHD-causing mutations were recovered in 61genes, with more than half being cilia-related(Fig. 2) (Li et al. 2015). The cilia genes recoveredincluded proteins localized in the cilia transitionzone, basal body/centrosome, ciliary axoneme,and also multiprotein complexes in the cyto-plasm required for cilia assembly (Fig. 2). Mostof the proteins recovered are expressed in bothmotile (9þ 2) and primary cilia (9þ 0), such ascomponents of the cilia transition zone. How-ever, some genes encode proteins unique to mo-tile cilia, such as the motor dyneins Dnah5 andDnah11 localized in the outer dynein arm re-quired for motile cilia function (Fig. 2). Manyof these cilia protein components are known to
cause various human ciliopathies, mostly in-volving nonmotile, primary cilia defects, suchas in Joubert syndrome (JBTS), Jeune syndrome,nephronophthisis, Meckel–Gruber syndrome,and others. The motile cilia mutations recoveredin the screen are linked to the sinopulmonarydisease primary ciliary dyskinesia (PCD). Al-though CHD is not an essential feature of cilio-pathies, it is notable that the mutants we recov-ered were all based on having CHD phenotypes.
Further indicating the important role of cil-ia in CHD pathogenesis, we also recovered mu-tations in 12 CHD genes that are in cilia-trans-duced cell signaling pathways, including genesmediating sonic hedgehog (Shh), transforminggrowth factor b and bone morphogenetic pro-teins (TGF-b/BMPs), and Wnt signaling (Fig.2). This enrichment of genes mediating cell sig-naling reflects the central role of cilia as a hubfor signal transduction pathways essential to theregulation of key cardiovascular developmentalprocesses. Also unexpected was the recovery of10 CHD genes involved in vesicular trafficking.This included Dynamin 2 and Ap2b1 requiredfor clathrin-mediated endocytosis, adaptin pro-teins Ap1b1 and Ap2b1, and Lrp1, Lrp2, andSnx17 mediating endocytic receptor recycling(Li et al. 2015b). Significantly, vesicular traffick-ing plays an essential role in cilia biology, withciliogenesis initiated with capture of a ciliaryvesicle by the mother centriole followed bydocking of the basal body to the cell membraneand fusion of additional secondary vesicles thatallow lengthening of the ciliary axoneme (So-rokin 1962; Kobayashi and Dynlacht 2011; Reiteret al. 2012). Vesicular trafficking and receptorrecycling also play important roles in the regu-lation of cell signaling. Although the endocyticpathway was not previously known to play a rolein CHD, its importance can be easily appreciat-ed in the context of its role in regulating cilio-genesis and cilia-transduced cell signaling.
CILIA AND CILIA-TRANSDUCED CELLSIGNALING IN HEART DEVELOPMENT
The overall finding that the large majority of theCHD genes recovered were cilia or cilia-relatedwas unexpected, given the screen was entirely
N.T. Klena et al.
4 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

phenotype-driven. Hence, they point to a cen-tral role for cilia biology in regulating cardio-vascular development and the pathogenesisof CHD. Primary cilia in the developing heartwere first identified via electron microscopy inthe chick, rabbit, mouse, and lizard embryos(Rash et al. 1969). These were observed onlyin nonmitotic cardiomyocytes or myoblasts,
whereas in the adult heart tissue, cilia wereonly observed in fibroblasts. A more recentstudy of the mouse embryo showed that ciliacan be found throughout the early E9.5 hearttube (Slough et al. 2008). As developmentprogresses to E12.5, cilia continue to be ex-pressed in the atria and in the trabeculated myo-cardium (Fig. 3J). Cilia are also found in the
Outerdynein
arm
Ciliogenesis Cell signaling
Inversincompartment
andtransition
zone
Ciliaassembly
Ciliary pocket
Cep290Cc2d2aMks1*
Celsr
Rab8
DvlFuz
Ca++
Tbc1d32 Nucleus MegF8
Smad6
Smad4Smad2,3
MegF8
IntegrinCfc1
Ltbp1
Pcsk5
Rsg1
FuzBr2
Br1
TGF-β
TGF-β/BMP/nodalsignalingGli
Gli
Kif7
Sufu
Sufu
Zbtb14
Smarca4
Prdm1
Tab1Fuz
Rsg1Prickle1
Vangl
ShhPtch1
Smo
Ptk7
Basalbody
Armc4Ccdc151Drc1Ccdc39
Ift74Ift140Dync2h1
Wntsignaling Calcium
signaling
Hedgehogsignaling
Pkd2Pkd1
Pkd2
Pkd1l1
Lrp2
IFTtransport
Dnah5Dnah11Dnal1Daw1
Anks6Cc2d2aCep290Jbts17Lox
Recyclingendosome Recycling
endosome
Rab8Rabin8Rab11
Bicc1Anks6
Anks6
Nek8
Pskh1
Cep110
Centrosome
Nucleus
Foxj1
Dyx1c1Dnaaf3
TGN
Cilia cytoplasmicpreassembly
Lrp2Myh10
Nek8Pde2a
Tmem67Wdpcp*
1 mm 0.5 mm
RVVSD LV
Cd
Ao
RVLV
PA
LA
RAVSD
Cd
AVSD
AVSD
VSDRV
LV
RL
Cr
LV
RV
AVSD
DORV
PA
R L
Cr
LV
Cd
Cr
RV LV
VSD
R
Cd
Cr
LPA
Aorticatresia
Ao
PA
BAVPA
Ao
R L
1 mm 0.5 mm 1 mm
1 mm 0.5 mm 0.2 mm
0.2 mm
0.2 mm
RV
AoPA
Ao
b2b2
025
Con
trol
E F
A B C
G H
K
D I
J
Ao
RVLV
PAAo
RVLV
PA
RVVSD LV
RVVSD LV
0.5 mm0.5 mm
Ao IAA
PA
Ao IAA
PA
Common AV valveCommon AV valve
Figure 2. Congenital heart disease (CHD) mutants recovered from mouse mutagenesis screen by fetal echocar-diography show preponderance of cilia-related mutations. Vevo 2100 color flow Doppler imaging showed criss-cross pattern of blood flow indicating normal aorta (Ao) and pulmonary artery (PA) alignment (A) confirmedby histopathology (B). E16.5 mutant (line b2b327) showed blood flow pattern indicating single great artery (PA)and ventricular septal defect (VSD) (C), suggesting aortic atresia with VSD, confirmed by histopathology (D).Color flow imaging of E15.5 mutant (line b2b2025) with heterotaxy (stomach on right) showed Ao/PA side-by-side with Ao emerging from right ventricle (RV) (E), indicating double outlet right ventricle (DORV)/VSD (F)and presence of atrioventricular septal defect (AVSD) (G,H ). Histopathology also showed bicuspid aortic valve(BAV) (I), interrupted aortic arch (IAA) (J ), and common atrioventricular (AV) valve (K). (Bottom) Diagramssummarize genes recovered causing CHD that are related to cilia or cell signaling, providing biological context ofCHD gene function. Color highlighting indicates CHD genes recovered; asterisks denote CHD genes recoveredfrom previous screen (Shen et al. 2005). R, Receptor; TGN, trans-Golgi network (adapted from data in Li et al.2015b).
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266 5
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from
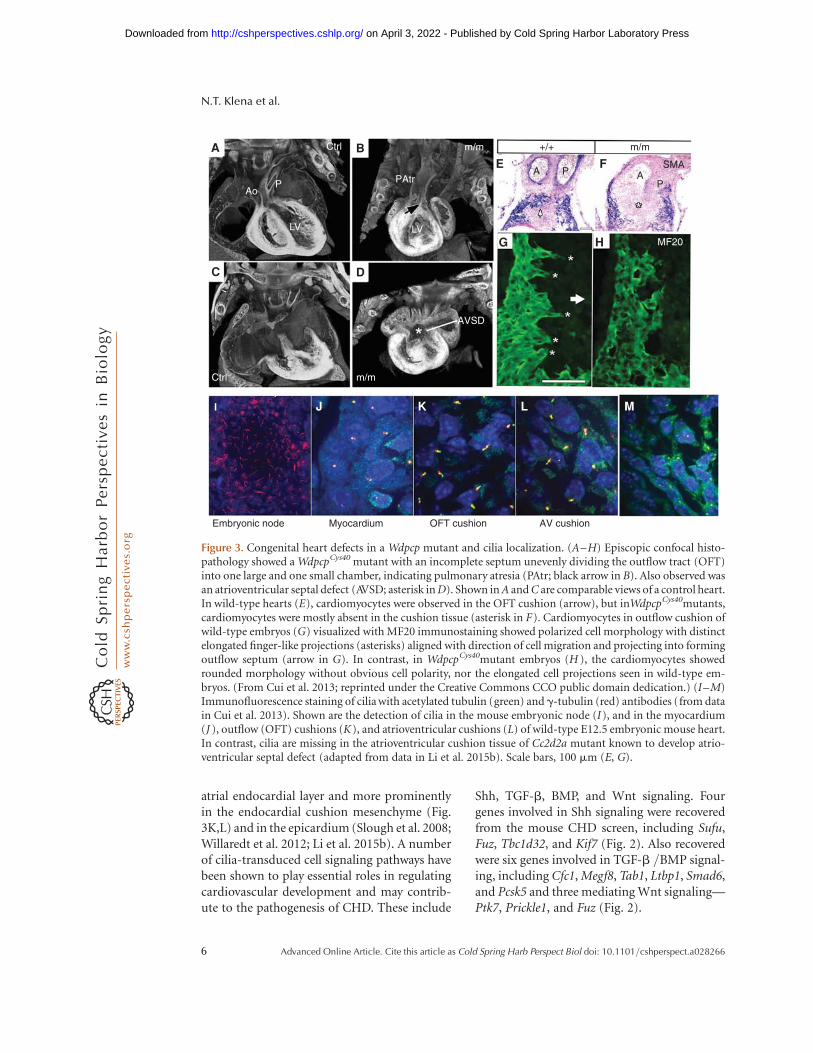
atrial endocardial layer and more prominentlyin the endocardial cushion mesenchyme (Fig.3K,L) and in the epicardium (Slough et al. 2008;Willaredt et al. 2012; Li et al. 2015b). A numberof cilia-transduced cell signaling pathways havebeen shown to play essential roles in regulatingcardiovascular development and may contrib-ute to the pathogenesis of CHD. These include
Shh, TGF-b, BMP, and Wnt signaling. Fourgenes involved in Shh signaling were recoveredfrom the mouse CHD screen, including Sufu,Fuz, Tbc1d32, and Kif7 (Fig. 2). Also recoveredwere six genes involved in TGF-b /BMP signal-ing, including Cfc1, Megf8, Tab1, Ltbp1, Smad6,and Pcsk5 and three mediating Wnt signaling—Ptk7, Prickle1, and Fuz (Fig. 2).
P
LV
PAtr
LV
AVSD
Ao
Ctrl
Ctrl m/m
m/m m/m
A AP
MF20
P
SMA
+/+A
C
I J K
Embryonic node Myocardium OFT cushion AV cushion
L M
B
D
G H
E F
Figure 3. Congenital heart defects in a Wdpcp mutant and cilia localization. (A–H) Episcopic confocal histo-pathology showed a WdpcpCys40 mutant with an incomplete septum unevenly dividing the outflow tract (OFT)into one large and one small chamber, indicating pulmonary atresia (PAtr; black arrow in B). Also observed wasan atrioventricular septal defect (AVSD; asterisk in D). Shown in A and C are comparable views of a control heart.In wild-type hearts (E), cardiomyocytes were observed in the OFT cushion (arrow), but inWdpcpCys40mutants,cardiomyocytes were mostly absent in the cushion tissue (asterisk in F). Cardiomyocytes in outflow cushion ofwild-type embryos (G) visualized with MF20 immunostaining showed polarized cell morphology with distinctelongated finger-like projections (asterisks) aligned with direction of cell migration and projecting into formingoutflow septum (arrow in G). In contrast, in WdpcpCys40mutant embryos (H ), the cardiomyocytes showedrounded morphology without obvious cell polarity, nor the elongated cell projections seen in wild-type em-bryos. (From Cui et al. 2013; reprinted under the Creative Commons CCO public domain dedication.) (I–M)Immunofluorescence staining of cilia with acetylated tubulin (green) and g-tubulin (red) antibodies (from datain Cui et al. 2013). Shown are the detection of cilia in the mouse embryonic node (I), and in the myocardium(J ), outflow (OFT) cushions (K), and atrioventricular cushions (L) of wild-type E12.5 embryonic mouse heart.In contrast, cilia are missing in the atrioventricular cushion tissue of Cc2d2a mutant known to develop atrio-ventricular septal defect (adapted from data in Li et al. 2015b). Scale bars, 100 mm (E, G).
N.T. Klena et al.
6 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Role of Shh Signaling in Cardiac Developmentand CHD
Shh signaling is the best-described cilia-trans-duced cell signaling pathway. Numerous studieshave shown that ablation of cilia can result in adrastic reduction of Shh signaling (Huangfuet al. 2003; Han et al. 2008; Goetz and Anderson2010). During heart development, Shh is ex-pressed in the pharyngeal endoderm and inthe foregut endoderm adjacent to incomingSHF derivatives in the dorsal mesenchyme pro-trusion (Dyer and Kirby 2009). Shh knockoutmice show atrial and atrioventricular septationdefects, defects in OFTseptation, and abnormalpharyngeal arch artery patterning (WashingtonSmoak et al. 2005). The outflow septation de-fects are characterized by the aorta shifted right-ward overriding the septum, and with eitherpulmonary atresia or a hypoplastic pulmonaryartery observed in conjunction with a variabledegree of ventricular hypertrophy. This constel-lation of defects is reminiscent of tetralogy ofFallot (TOF), one of the most common complexCHD observed clinically (Washington Smoaket al. 2005). Using Cre targeted deletion analy-sis, it was shown that these outflow defects re-flect a dual requirement for pharyngeal endo-dermal-derived Shh in the cardiac neural crestcells and the SHF derivatives (Goddeeris et al.2007). These studies showed Shh signaling tothe SHF and cardiac neural crest cells are re-quired for OFT septation, but not for eitherOFT lengthening, or cushion formation, respec-tively. As the Shh knockout embryos showed areduction in the number of SHF derivatives, thissuggested a requirement for Shh in the specifi-cation of the SHF (Hildreth et al. 2009).
Role of Wnt Signaling in CardiacDevelopment and CHD
Primary cilia also play a role in the transductionof canonical and noncanonical Wnt signaling(Clevers 2006; MacDonald et al. 2009; Walling-ford and Mitchell 2011; May-Simera and Kelley2012) pathways that are also essential for nor-mal heart development. One early evidencelinking cilia with b-catenin-dependent canoni-
cal Wnt signaling was the observation thatknockdown of basal body components bbs1,bbs4, and bss6 resulted in several-fold increasein Wnt activity in zebrafish (Gerdes et al. 2007).The functional link between Wnt signaling andcilia was also shown by the observed localiza-tion of noncanonical Wnt/planar cell polarity(PCP) components, such as Inversin, Dishev-elled, Vangl2, and Wdpcp in the basal bodyand/or ciliary axoneme (Fig. 4A,B) (Montcou-quiol et al. 2003; May-Simera and Kelley 2012;Cui et al. 2013). Other studies also showed a rolefor cilia as a switch that can constrain canonicalversus noncanonical Wnt signaling (Ross et al.2005; Simons et al. 2005; Barrow et al. 2007;Gerdes et al. 2007; Corbit et al. 2008; Huangand Schier 2009; Stottmann et al. 2009; Lien-kamp et al. 2012; Oh and Katsanis 2013). How-ever, the precise mechanism by which ciliaregulate Wnt signaling is not well understood.
In mice, the noncanonical Wnt/PCP genessuch as Celsr, Frizzled3 (Fzd3), Fzd6, Vangl1-2,and Dvl1-3 are highly expressed in the OFT(Etheridge et al. 2008; Paudyal et al. 2010).Mice with mutations in the PCP genes Vangl2,Scrib (Phillips et al. 2007), Dvl 1, 2, and 3(Hamblet et al. 2002; Etheridge et al. 2008;Sinha et al. 2012), Wdpcp (Cui et al. 2013),and Pk1 (Gibbs et al. 2016) show a spectrumof CHD phenotypes involving OFT malalign-ment and septation defects, such as double out-let RV (Fig. 2E,F), overriding aorta, pulmonaryatresia (Fig. 3B), and persistent truncus arteri-osus (Henderson et al. 2006; Cui et al. 2013;Boczonadi et al. 2014; Gibbs et al. 2016). Thesecardiac defects likely reflect a role for nonca-nonical Wnt/PCP pathway in regulating thepolarized migration of cardiac neural crest andSHF derivatives (Tada and Smith 2000; Mont-couquiol et al. 2003; Simons et al. 2005; Verziet al. 2005; Cohen et al. 2007; Simons and Mlod-zik 2008; Schlessinger et al. 2009; Gibbs et al.2016). Consistent with this, examination ofmouse embryonic fibroblasts derived from theWdpcp or Pk1 mutant embryos showed inabilityof the cells to polarize and engage in directionalcell migration (Figs. 4C–G, 5K–N). In contrastto Shh deficiency, Wnt/PCP disruption causedfailure of the OFT to appropriately lengthen
Cilia and Ciliopathies in Congenital Heart Disease
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266 7
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

A B
C D
H
I J
+/+ m/m
E G70
+/+ 94/117 in (0–60)m/m 64/161 in (0–60)±[
± p < 0.000160504030
Perce
ntag
e
Golgi orientation
2010
0 0–30°
30–60°
60–90°
90–120°
120–150°
150–180°
K
L M
N
Merged
Merged
Merged
F+/+ m/mm/m
Figure 4. Wdpcp is a cilia protein regulating cell polarity, directional cell migration, and the actin cytoskeleton.(A,B) IMCD3 cells immunostained with Wdpcp (green) and acetylateda-tubulin (red) antibodies show Wdpcplocalization in the axoneme and ring-like structure (arrowhead) at the ciliary base. Localization of Wdpcp (red)in this ring-like structure is better seen with a 3D isosurface reconstruction, which also shows some colocaliza-tion of Septin-2 (green) with the Wdpcp ring. (C–G) In a wound-healing assay, control mouse embryonicfibroblasts (MEFs) (A) show good alignment with the direction of wound closure (indicated by white arrow). Incontrast, WdpcpCys40 mutant MEFs (B) showed a disorganized distribution. These differences in cell polaritywere also reflected in the Golgi orientation (white line drawn through the center of the Golgi stained green)(E,F). In wild-type MEFs, the Golgi (green) was mostly situated at the cell’s leading edge (E,G), aligned with thedirection of wound closure (white arrow), whereas the WdpcpCys40 mutant MEFs show randomized Golgiorientation (F,G). Scale bars, 20 mm (A, B, C, D, F). (H–N) Confocal imaging of Sept2 (red) and Wdpcp(green) showed they are colocalized in actin stress fiber (phalloidin stained, blue) in wild-type MEFs (H–J),but in the WdpcpCys40 mutant MEFs, Wdpcp expression was lost (blue, L), whereas Sept2 immunostaining (red,K,M) showed the loss of colocalization with actin (blue) (K,L,M). (L) Wdpcp (green) is enriched at the cellcortex where actin filaments (phalloidin) insert into vinculin (red)-containing focal adhesions (N) in wild-typeMEFs. (Adapted from Cui et al. 2013 under the Creative Commons CC0 public domain dedication.)
N.T. Klena et al.
8 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

A B C +/–
D
I
J
FE
HG
K L
m/m
TZ
+/– m/m
+/– m/m
+/–
+/+
M N+/+
m/m
m/m
V+/+ = 1.367±V–/– = 1.657±
p = 0.0017
D+/+ = 0.513±0
0 20 40 60 80 0 20
Migration orientation (degrees)
40 60 80
Per
cent
age
1
2
3
4
5 +/+p = 0.0261
m/m
D–/– = 0.523±p = 0.7864
m/m
m/m
Apical
+/+
Apical
TZ
Figure 5. Shortened outflow tract (OFT) and defects in cell polarity and directional cell migration in the Pk1Bj
mutant. (A–D) E10.5 Pk1Bj mutant embryo. B and D show shortened OFT as compared with that of hetero-zygous embryo (C,D). (E–H) Islet1 immunostaining show distribution of SHF cells in the dorsal pericardialwall of the OFTof control (E,G) and Pk1Bj mutant embryos (F,H). Magnified views of region denoted by arrow-heads in E and F revealed a cuboidal (H ) rather than flat squamous (G) epithelial morphology in the homo-zygous mutant versus heterozygous embryo. (I,J ) b-Catenin (green) and laminin (red) antibody staining ofwild-type (I) and Bj mutant embryos (J ) shown in the E10.5 Bj mutant embryo, marked disorganization of theepithelium in the transition zone (TZ) of the pericardial wall where SHF derivatives are found. Confocal imagingshowed laminin (red) is localized basally (arrowhead I) in the TZ of the control embryo, but in the mutantembryo, it is localized apically (arrow) and basally (arrowhead J ), indicating a loss of normal epithelial polarity.The distribution of b-catenin (green) remains at the cell surface in both the control and Bj mutant embryos.(K,L) Myocardiolization defect in the OFTof Pk1Bj mutants. Examination of the striated banding pattern fromMF20 immunostain showed the developing myofilaments in the heart are closely aligned and oriented towardthe direction of myocardialization in the wild-type E14.5 embryo (K), but in the Bj mutant, the myofilamentsare sparse and are largely oriented perpendicular to the direction of myocardialization and septum formation(L). (M,N) Wound closure assay shows a defect in directional cell migration in Pk1Bj mutant mouse embryonicfibroblasts (MEFs). The migration path of MEFs 8 h after wound scratch were well aligned with the direction ofwound closure, but tortuous paths were observed with increased velocity for the Pk1Bj mutant MEFs (M,N)(adapted from data in Gibbs et al. 2016). Scale bars, 0.5 mm (A, B); 50 mm (E); 20 mm (K).
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266 9
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

(Fig. 5A–D). In the Pk1 mutant, the epithelialorganization and apical-basal polarity of theSHF derivatives in the OFT are disrupted. Thiswould suggest a defect in convergent-extensioncell movement required for delamination of acohesive epithelial sheet mediating OFT length-ening (Fig. 5E–J). This is followed later by amyocardialization defect of the OFT (Figs.3E–H, 5K–L), that together with the shortenedOFT likely account for the great artery mala-lignment defect in the Pk1 mutant.
Role of TGF-b Signaling in CardiacDevelopment and CHD
A role for cilia in mediating TGF-b signalingwas recently shown with the finding that ligandbinding causes accumulation of TGF-b recep-tors at the base of the cilium, in a region knownas the ciliary pocket (Clement et al. 2013). Thistriggers receptor-mediated endocytosis involv-ing clathrin-coated vesicles, leading to down-stream activation of SMAD phosphorylation(Clement et al. 2013). The essential role ofTGF-b/BMP signaling in CHD is well de-scribed via in vitro and in vivo analyses of chickand mouse embryos, and also with the exami-nation of knockout mouse models (Combs andYutzey 2009; de Vlaming et al. 2012; Kruithofet al. 2012; von Gise and Pu 2012). These studiesshow TGF-b/BMP signaling has multiple rolesin cardiovascular development that include theregulation of both endocardial EMT and endo-cardial cushion development (Potts and Run-yan 1989; Camenisch et al. 2002). For example,early endocardial cushion development to ac-quire critical valve-like function requires BMPsignaling in cardiac neural crest cells via theBMPRIA receptors (Nomura-Kitabayashi et al.2009). A role for Tgfb2 in OFT and aortic archremodeling is indicated by the finding thatTgfb2 knockout mice die perinatally with dou-ble outlet RV and interrupted aortic arch (San-ford et al. 1997).
The disturbance of TGF-b/BMP signalingis likely to play a major role in the valvular de-fects seen in mice harboring mutations disrupt-ing clathrin-mediated endocytosis and endo-cytic receptor recycling (Ap2b1, Dnm2, Ap1b1,
Snx17, LRP1, LRP2). These endocytic mutantsall show OFT malalignment and endocardialcushion defects, phenotypes reminiscent ofthose observed in mutants with disruption ofTGF-b/BMP signaling (Li et al. 2015b). Simi-larly, mutations affecting cilia integrity in theendocardial cushions may cause disruption ofcilia-transduced TGF-b/BMP signaling re-quired for normal valve development. Thus,mutation in Cc2d2a, a cilia transition zonecomponent, causes selective loss of cilia in theatrioventricular (AV) but not outflow cushions,and as might be expected, such mutants showedAV valve defects, while the outflow valves werespared (Fig. 3K–M).
ROLE OF CILIA IN SPECIFICATIONOF CELL POLARITY AND POLARIZEDCELL MIGRATION
Some cilia proteins may help regulate cardio-vascular development through cross talk, di-rectly or indirectly, with the cytoskeleton tospecify cell polarity and directional cell migra-tion, morphogenetic cell movements, and epi-thelial–mesenchyme cell transformation. Giv-en the basal body is a microtubule organizingcenter that can regulate nucleation and organi-zation of microtubule outgrowth, one conceptthat has emerged is that cilia may regulate thecytoskeleton through dynamic interactionswith PCP components and, in this manner,specify cell polarity and polarized cell migration(Figs. 4 and 5) (Wallingford and Mitchell 2011;May-Simera and Kelley 2012). These dynamiccell processes may help to direct the long-dis-tance migration of multiple extracardiac cellpopulations to the embryonic heart that are re-quired for normal heart development. This in-cludes cells from the SHF, neural crest cells, andthe pro-epicardial cells. In addition, cilia direct-ed reorganization of the actin cytoskeleton mayalso contribute to the regulation of EMT, suchas required for the emergence of cardiac neuralcrest cells from the dorsal neural fold, endocar-dial EMT mediating formation of the cardiaccushions and valves, or epicardial EMT thatgenerate the epicardially derived cells formingthe coronary vessels. These developmental pro-
N.T. Klena et al.
10 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

cesses involving dynamic reorganization of thecytoskeleton is impacted by cilia and, in con-junction with cilia-transduced cell signaling,may help orchestrate development of the car-diovascular system.
Although the role of cilia in the regulation ofcell polarity and directional cell migration in thecardiovascular development is well described inthe context of OFT morphogenesis (see above),the precise mechanism and role of the cilia inmodulating cell polarity is less understood. Inthis regard, it is worth pointing out that Wdpcp,a PCP component also known as Fritz, is local-ized not only in the cilia, but it is also colocal-ized with septins in the cilia (Kim et al. 2010;Cui et al. 2013) and in the actin cytoskeleton(Kim et al. 2010; Cui et al. 2013). In mouseembryonic fibroblast (MEF) cells deficient inWdpcp, a marked reorganization of the actincytoskeleton is observed (Fig. 4H–L), and thisis associated with altered focal contacts (Fig.4N) inability to establish cell polarity and en-gage in directional cell migration (Fig. 4C–G).Similar studies of MEFs harboring a mutationin the PCP component Pk1 also showed a sim-ilar loss of cell polarity and defect in directionalcell migration (Fig. 5M,N) (Gibbs et al. 2016).Together, these findings suggest that cilia muta-tions may cause CHD not only via the disrup-tion of cilia-transduced cell signaling, but ciliamutations also may disrupt the cytoarchitectureand perturb the establishment of cell polarity,polarized cell migration, and/or EMT.
CILIA IN LEFT–RIGHT PATTERNINGAND CONGENITAL HEART DISEASE
The enrichment of cilia genes was also notablein that it included a subset of genes that causedCHD in conjunction with left–right patterningdefects. This likely reflects the known require-ment for cilia in left–right patterning, with pre-vious studies indicating that motile cilia at theembryonic node is required to break symmetry(Fig. 3I) (Hirokawa et al. 2009; Nakamura andHamada 2012). Analysis of motile cilia mutantmice revealed CHD is typically observed in con-junction with heterotaxy, the randomization ofleft–right patterning (Tan et al. 2007). This is
consistent with the well-described clinical asso-ciation of complex CHD with heterotaxy (Linet al. 2014). As the heart is the most left–rightasymmetric organ, and this asymmetry is essen-tial for efficient oxygenation of blood, it is per-haps not surprising that left–right patterningdefects may play a major role in CHD patho-genesis.
Among 34 cilia mutations recovered causinglaterality defects, 22 genes perturbed the prima-ry cilia (Cc2d2a, Anks6, Nek8, Mks1, Cep290,Bicc1) versus 12 genes that disrupted motile cil-ia (Dnah5, Dnah11, Dnai1, Daw1, Armc4,Ccdc151, Drc1, Ccdc39, Dyxc1x1, Dnaaf3) (Liet al. 2015b). The latter genes are known tocause PCD, a ciliopathy that is autosomal reces-sive (Collins et al. 2014; Horani et al. 2014; Loboet al. 2015). In PCD, immotile/dyskinetic ciliain the airway cause mucociliary clearance de-fects that can lead to severe sinopulmonary dis-ease. Approximately half of PCD patients showsitus solitus, half situs inversus totalis, and vary-ing numbers up to 8% may show CHD withheterotaxy (Kennedy et al. 2007; Shapiro et al.2014). The disturbance of laterality with PCDreflects the essential role of motile cilia in left–right patterning. Studies in the PCD mutantmouse models showed each PCD mutationcan give rise to three phenotypes—approxi-mately half with situs solitus or situs inversusand half with heterotaxy, with complex CHDobserved only with heterotaxy (Tan et al.2007). Although the heterotaxy mutants mostlydie prenatally or neonatally from the CHD, mu-tants with situs solitus or inversus are largelyviable postnatally without CHD. Videomicros-copy showed most of these PCD mutants haveimmotile cilia in the embryonic node, even ashalf of the mutants show normal or invertedconcordant situs that indicate the breaking ofsymmetry.
These striking observations suggest thatmotile cilia are not absolutely required forbreaking symmetry, nor for left–right axis spec-ification, although motile cilia are clearly re-quired for high-fidelity situs solitus specifica-tion. As CHD is only seen with heterotaxy,this provides a clue that patterning of the car-diovascular system may occur very early in de-
Cilia and Ciliopathies in Congenital Heart Disease
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266 11
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

velopment, at the time the left–right body axisis specified. Even as these findings show thatmotile cilia play an important role in left–rightpatterning, the recovery of 24 mutations affect-ing primary cilia suggests nonmotile cilia alsoplay an essential role in laterality specification(Li et al. 2015b). Previous studies suggested atwo-cilia hypothesis in which motile cilia at thenode generated right to left flow (for additionalinformation, see Shinohara and Hamada 2016).This is proposed to trigger mechanosensorytransduction of primary cilia in the perinodalcrown cells, causing left-sided calcium releasethat is propagated into the surrounding lateralplate mesoderm, causing the breaking of sym-metry (Nonaka et al. 2002; McGrath et al. 2003;Bruekner 2007; Yoshiba et al. 2012). However,this model has been called into question recent-ly given the failure to detect cilia-mediated me-chanosensation and calcium release (Dellinget al. 2016).
A role for primary cilia in left–right pattern-ing could be easily understood neverthelesswithout invoking mechanosensation, becauseShh and TGF-b signaling, both cilia-transducedpathways, play important roles in left–right pat-terning. Although Shh knockout mice do notshow overt laterality defects, they show LAisomerism (Hildreth et al. 2009). Furthermore,the single outflow vessel seen in the Shh knock-out mouse is said to represent pulmonary atre-sia, as the single great artery shows Pitx2c, indi-cating a left-sided identity (Washington Smoaket al. 2005). It is interesting to note in chickembryos where Shh plays a much more primaryrole in left–right patterning, the experimentalmanipulation of left–right expression of Shhcan cause CHD, confirming its importance ofleft–right patterning in the pathogenesis ofCHD (Levin et al. 1995). Signaling mediatedby the TGF-b family of growth factors, includ-ing nodal, lefty1, and lefty2, are well describedto specify the left–right axis. This nodal signal-ing cascade is believed to propagate left–rightspecification initiated at the node. How muta-tions affecting primary cilia may contribute tothe disruption of left–right patterning is notknown, but it is thought to cause disturbance inthe propagation of this nodal signaling cascade.
CILIARY DYSFUNCTION AND CILIOMEMUTATIONS IN CHD PATIENTS
The unexpected enrichment for mutations incilia-related (ciliome) genes and genes involvedin endocytic trafficking and in cilia-transducedcell signaling (Shh, WNT/Pcp, TGF-b) in themouse mutagenesis screen point to a centralrole for cilia in CHD pathogenesis. To assessthe relevance of these findings to human CHD,we investigated the findings from exome-se-quencing analysis of CHD patients by the Pedi-atric Cardiac Genomics Consortium (PCGC)(Zaidi et al. 2013). In this analysis, the focuswas on examining de novo predicted pathogeniccoding variants. Although the PCGC publica-tion focused on the recovery of de novo variantsin a number of chromatin-modifying genes,interestingly, we noted among the 28 de novodamaging mutations identified in the PCGCCHD patient cohort, 13 or nearly half were ingenes associated with pathways identified in themouse forward genetic screen—that is, ciliogen-esis, endocytic trafficking, and cilia-transducedcell signaling (SHH, WNT, TGF-b) (Table 1),with LRP2 being a gene recovered in both thePCGC CHD patients and the mouse CHD mu-tants recovered in our screen. We also noted therecovery in the PCGC cohort of a de novo vari-ant in Pitx2, a gene known to play an essentialrole in left–right patterning, supporting an im-portant role for left–right patterning distur-bance in CHD pathogenesis.
Further supporting a central role for cilia inthe pathogenesis of CHD are clinical studiesshowing a high prevalence of ciliary dysfunctionin CHD patients (Nakhleh et al. 2012; Garrodet al. 2014). Given that respiratory compli-cations are among the biggest postsurgicalcomplications for CHD patients, we previouslyhypothesized that some CHD patients with res-piratory complications may have undiagnosedPCD. These studies were initiated with an exam-ination of CHD patients with heterotaxy. Nasalscrapes were conducted and video microscopywas used to examine cilia motility in the nasalepithelium. This analysis showed a high preva-lence of ciliary dysfunction in CHD patientswith heterotaxy. The ciliary motion defects
N.T. Klena et al.
12 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

observed span a spectrum that included someshowing dyskinetic ciliary motion to slow oreven immotile cilia. Overall, .40% of the pa-tients showed ciliary dysfunction (Nakhleh et al.2012). Although this was associated with an en-richment for coding variants in PCD genes, nopatient was either homozygous or compoundheterozygous for any PCD gene mutations.Thus, although CHD patients with heterotaxyare at high risk for ciliary dysfunction, these pa-tients largely do not have PCD. Since this initialstudy, a large study has been conducted compris-ing .200 patients with CHD of a broad spec-trum, mostly without heterotaxy. This analysisshowed a similar high prevalence of ciliary dys-function and this was correlated with increasedrisk of having PCD-related respiratory symp-toms (Garrod et al. 2014). Together, these find-ings suggest ciliary dysfunction is commonly as-sociated with CHD in the human population.
Although these studies focused on assessingmotile cilia function, we note many cilia genesare expressed in both motile and primary cilia.Hence, the high prevalence of ciliary dysfunc-tion in CHD patients may reflect not only theperturbation of motile cilia genes, but also genesrequired for primary cilia function. Indeed, werecently showed a patient harboring compoundheterozygous mutations in WDR35 causing
Sensenbrenner syndrome, a ciliopathy thoughtto affect only the primary cilia, showed motilecilia dysfunction. Pulmonary function assess-ments indicated obstructive airway disease thatsuggested possible mucociliary clearance de-fects in the airway (Li et al. 2015a). Indeed, sev-eral clinical studies have shown an increase inrespiratory symptoms and disease in patientswith other ciliopathies thought to affect onlythe primary cilia, indicating the distinction be-tween ciliopathies involving motile versus pri-mary cilia may not be so clear cut (Tobin andBeales 2009). These findings suggest furtherstudies are warranted to assess ciliopathy pa-tients of a wide spectrum for potential pulmo-nary complications, especially for those whowill undergo high-risk surgeries, such as thoseinvolving cardiopulmonary bypass.
CONGENITAL HEART DISEASE ANDCILIOPATHIES
It is notable that many cilia genes recovered inthe mouse forward genetic screen for CHD-causing mutations are genes clinically knownto cause various human ciliopathies. This in-cludes not only motile cilia genes associatedwith PCD, but also cilia genes linked to variousciliopathies thought to affect the primary cilia,
Table 1. Functional annotation for 13 PCGC patients with de novo mutations
Patient ID CHD typea Gene Mutation Gene function annotation
1-00638 CTD FBN2 p.D2191N TGF-b signaling1-02020 HTX SMAD2 p.IVS12 þ 1G . A TGF-b signaling1-02621 HTX SMAD2 p.W244C TGF-b signaling1-00197 LVO BCL9 p.M1395K Wnt signaling1-01828 CTD DAPK3 p.P193L Wnt signaling1-01138 LVO USP34 p.L432P Wnt signaling1-00802 LVO PTCH1 p.R831Q Shh signaling/ciliome1-02598 HTX LRP2b p.E4372K Shh signaling/endocytic trafficking1-01913 Other RAB10 p.N112S Endocytic trafficking1-00750 LVO HUWE1 p.R3219C Ciliome1-01151 CTD SUV420H1 p.R143C Ciliome1-00853 CTD WDR5 p.K7Q Ciliome1-02952 LVO PITX2 p.A47V Laterality-related
Based on exome-sequencing analysis of congenital heart disease (CHD) patients by Pediatric Cardiac Genomics
Consortium (Data from Zaidi et al. 2013).aCTD, Conotruncal defect; HTX, heterotaxy; LVO, left ventricular obstruction.bLRP2 is an endocytic gene also recovered from our mouse screen.
Cilia and Ciliopathies in Congenital Heart Disease
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266 13
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

such as in JBTS, polycystic kidney disease, acro-callosal syndrome, hydroelethalus, Leber con-gential amaurosis, Meckel–Gruber syndrome,Bardet–Biedl syndrome, etc. (Li et al. 2015b).While in our mouse screen, ciliopathy geneswere recovered based on mutations causingCHD phenotypes, clinically these ciliopathiesare not commonly associated with CHD. Thismay reflect ascertainment bias given that thepatient population represent only human fetus-es that can survive to term and, hence, are lesslikely to have severe cardiac anomalies. Indeed,clinical studies of aborted or stillborn fetuseshave shown that the human fetal populationhas more than ten times higher incidence ofCHD as compared with those in the clinical pa-tient population (Hoffman and Kaplan 2002).Consistent with this, most of the CHD ciliop-athy mutants recovered from our screen wereinviable to term and were harvested pretermafter in utero phenotyping by fetal echocardiog-raphy. On the flip side, there is undoubtedlyascertainment bias in our screen in the recoveryof mutations in ciliopathy genes that specificallycan cause CHD. That different ciliopathy mu-tations may have varying levels of penetrancefor CHD phenotypes is suggested by observa-tions of our mutant Hug (Damerla et al. 2015).This mutant has a mutation in Jbts17, a geneencoding a cilia transition zone protein knownto cause JBTS (Srour et al. 2012). Hug mutantsshow cerebellar defects expected for JBTS andthey also can show CHD comprising of pulmo-nary atresia. However, the CHD phenotype isincomplete in penetrance, as some Hug mutantsshow no heart defects (Damerla et al. 2015).These observations suggest that different muta-tions in the same ciliopathy gene may generatedifferent phenotypic outcome and this perhapscan be further modified by the genetic back-ground of the individual.
In light of these observations, we suggestthat, clinically, CHD may be considered a struc-tural birth defect related to ciliopathies. Howev-er, unlike other ciliopathies, which are relativelyrare (,1 in 10,000) and with a Mendelian re-cessive inheritance, the much higher prevalenceof CHD (up to 1%) and its sporadic occurrencewould suggest the contribution of cilia-related
or ciliome genes in CHD will be multigenic andhighly genetically heterogeneous. Such complexgenetics is expected to reflect the complexity ofcilia biology in which sequence variants foundamong different “ciliome” genes may affect thefunction of large multiprotein complexes thatregulate ciliogenesis and cilia structure andfunction. Given that there are hundreds of cil-iome genes that contribute to cilia assembly andcilia structure and function, it is perhaps notsurprising that CHD patients are observed tohave a high prevalence of ciliary dysfunction.While the CHD genes recovered from the mousescreen were by design recessive mutations, weexpect mutations in these same genes can con-tribute to more complex genetic models ofdisease. Such complex genetics may also con-tribute to classic ciliopathies, as there are clinicalreports of PCD patients and patients with otherciliopathies that have no homozygous or com-pound heterozygous ciliopathy mutations, butinstead show multiple heterozygous mutationsin known PCD or other ciliopathy genes (dePontual et al. 2009; Li et al. 2016). A future chal-lenge is to develop an effective bioinformaticspipeline for modeling and interrogating suchcomplex genetics and assess the contributionof ciliome mutations in the pathogenesis ofCHD and other structural birth defects.
CONCLUSIONS
CHDs are the most common structural birthdefects, and despite its prevalence, the geneticetiology of CHD remains poorly understood.Interrogations into the genetic landscape forCHD using a large-scale forward genetic screenin mice unveiled a central role for ciliome genesin the pathogenesis of CHD. These studies sug-gest the perturbation of cilia and cilia-trans-duced cell signaling pathways may play a centralrole in the pathogenesis of CHD. The futurechallenge is to clinically translate these findingsin mice to patients with CHD. The finding ofa high prevalence of ciliary dysfunction in CHDpatients and the enrichment of de novo patho-genic variants in cilia and cilia-related pathwaysin CHD patients would suggests such studieswill be fruitful and may provide the basis for
N.T. Klena et al.
14 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

stratifying patients to optimize the clinicalmanagement of patient care. The recent findingof primary cilia in the endothelial cells of theaorta regulating anti-atherosclerotic responsesalso point to a potential role for cilia in adultcardiac disease (Dinsmore and Reiter 2016).Further work in the future will be needed toclarify the role of cilia biology in human CHDand perhaps other cardiovascular diseases, andwith such insights may come new avenues oftherapeutic intervention to improve the out-come for patients with critical heart disease.
REFERENCES�Reference is also in this collection.
Barrow JR, Howell WD, Rule M, Hayashi S, Thomas KR,Capecchi MR, McMahon AP. 2007. Wnt3 signaling in theepiblast is required for proper orientation of the antero-posterior axis. Dev Biol 312: 312–320.
Boczonadi V, Gillespie R, Keenan I, Ramsbottom SA,Donald-Wilson C, Al Nazer M, Humbert P, SchwarzRJ, Chaudhry B, Henderson DJ. 2014. Scrib:Rac1 in-teractions are required for the morphogenesis of theventricular myocardium. Cardiovasc Res 104: 103–115.
Brueckner M. 2007. Heterotaxia, congenital heart disease,and primary ciliary dyskinesia. Circulation 115: 2793–2795.
Buckingham M. 2016. First and second heart field. In Con-genital heart diseases: The broken heart: Clinical features,human genetics and molecular pathways (ed. Rickert-Sperling S, Kelly GR, Driscoll JD), pp. 25–40. Springer,Vienna.
Camenisch TD, Molin DG, Person A, Runyan RB, Gitten-berger-de Groot AC, McDonald JA, Klewer SE. 2002.Temporal and distinct TGF-b ligand requirements dur-ing mouse and avian endocardial cushion morphogene-sis. Dev Biol 248: 170–181.
Clement CA, Ajbro KD, Koefoed K, Vestergaard ML, VelandIR, Henriques de Jesus MP, Pedersen LB, Benmerah A,Andersen CY, Larsen LA, et al. 2013. TGF-b signaling isassociated with endocytosis at the pocket region of theprimary cilium. Cell Rep 3: 1806–1814.
Clevers H. 2006. Wnt/b-catenin signaling in developmentand disease. Cell 127: 469–480.
Cohen ED, Wang Z, Lepore JJ, Lu MM, Taketo MM, EpsteinDJ, Morrisey EE. 2007. Wnt/b-catenin signaling pro-motes expansion of Isl-1-positive cardiac progenitor cellsthrough regulation of FGF signaling. J Clin Invest 117:1794–1804.
Collins SA, Walker WT, Lucas JS. 2014. Genetic testing in thediagnosis of primary ciliary dyskinesia: State-of-the-artand future perspectives. J Clin Med 3: 491–503.
Combs MD, Yutzey KE. 2009. Heart valve development:Regulatory networks in development and disease. CircRes 105: 408–421.
Corbit KC, Shyer AE, Dowdle WE, Gaulden J, Singla V, ChenMH, Chuang PT, Reiter JF. 2008. Kif3a constrains b-cat-enin-dependent Wnt signalling through dual ciliary andnon-ciliary mechanisms. Nat Cell Biol 10: 70–76.
Cui C, Chatterjee B, Lozito TP, Zhang Z, Francis RJ, YagiH, Swanhart LM, Sanker S, Francis D, Yu Q, et al. 2013.Wdpcp, a PCP protein required for ciliogenesis, reg-ulates directional cell migration and cell polarity by directmodulation of the actin cytoskeleton. PLoS Biol 11:e1001720.
Damerla RR, Cui C, Gabriel GC, Liu X, Craige B, Gibbs BC,Francis R, Li Y, Chatterjee B, San Agustin JT, et al. 2015.Novel Jbts17 mutant mouse model of Joubert syndromewith cilia transition zone defects and cerebellar and otherciliopathy related anomalies. Hum Mol Genet 24: 3994–4005.
Delling M, Indzhykulian AA, Liu X, Li Y, Xie T, Corey DP,Clapham DE. 2016. Primary cilia are not calcium-re-sponsive mechanosensors. Nature 531: 656–660.
de Pontual L, Zaghloul NA, Thomas S, Davis EE,Mcgaughey DM, Dollfus H, Baumann C, Bessling SL,Babarit C, Pelet A, et al. 2009. Epistasis between RETand BBS mutations modulates enteric innervation andcauses syndromic Hirschsprung disease. Proc Natl AcadSci 106: 13921–13926.
de Vlaming A, Sauls K, Hajdu Z, Visconti RP, Mehesz AN,Levine RA, Slaugenhaupt SA, Hagege A, Chester AH,Markwald RR, et al. 2012. Atrioventricular valve devel-opment: New perspectives on an old theme. Differentia-tion 84: 103–116.
Dinsmore C, Reiter JF. 2016. Endothelial primary cilia in-hibit atherosclerosis. EMBO Rep 17: 156–166.
Dyer LA, Kirby ML. 2009. Sonic Hedgehog maintains pro-liferation in secondary heart field progenitors and is re-quired for normal arterial pole formation. Dev Biol 330:305–317.
Etheridge SL, Ray S, Li S, Hamblet NS, Lijam N, Tsang M,Greer J, Kardos N, Wang J, Sussman DJ, et al. 2008. Mu-rine dishevelled 3 functions in redundant pathways withdishevelled 1 and 2 in normal cardiac outflow tract, co-chlea, and neural tube development. PLoS Genet 4:e1000259.
Fahed AC, Gelb BD, Seidman JG, Seidman CE. 2013. Ge-netics of congenital heart disease: The glass half empty.Circ Res 112: 707–720.
Garrod AS, Zahid M, Tian X, Francis RJ, Khalifa O, DevineW, Gabriel GC, Leatherbury L, Lo CW. 2014. Airwayciliary dysfunction and sinopulmonary symptoms in pa-tients with congenital heart disease. Ann Am Thorac Soc11: 1426–1432.
Gerdes JM, Liu Y, Zaghloul NA, Leitch CC, Lawson SS, KatoM, Beachy PA, Beales PL, DeMartino GN, Fisher S, et al.2007. Disruption of the basal body compromises protea-somal function and perturbs intracellular Wnt response.Nat Genet 39: 1350–1360.
Gibbs BC, Damerla RR, Vladar EK, Chatterjee B, Wan Y, LiuX, Cui C, Gabriel GC, Zahid M, Yagi H, et al. 2016.Prickle1 mutation causes planar cell polarity and direc-tional cell migration defects associated with cardiac out-flow tract anomalies and other structural birth defects.Biol Open 5: 323–335.
Cilia and Ciliopathies in Congenital Heart Disease
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266 15
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Gill HK, Splitt M, Sharland GK, Simpson JM. 2003. Patternsof recurrence of congenital heart disease: An analysis of6,640 consecutive pregnancies evaluated by detailed fetalechocardiography. J Am Coll Cardiol 42: 923–929.
Glessner JT, Bick AG, Ito K, Homsy JG, Rodriguez-MurilloL, Fromer M, Mazaika E, Vardarajan B, Italia M, Leipzig J,et al. 2014. Increased frequency of de novo copy numbervariants in congenital heart disease by integrative analysisof single nucleotide polymorphism array and exome se-quence data. Circ Res 115: 884–896.
Goddeeris MM, Schwartz R, Klingensmith J, Meyers EN.2007. Independent requirements for Hedgehog signalingby both the anterior heart field and neural crest cells foroutflow tract development. Development 134: 1593–1604.
Goetz SC, Anderson KV. 2010. The primary cilium: A sig-nalling centre during vertebrate development. Nat RevGenet 11: 331–344.
Hamblet NS, Lijam N, Ruiz-Lozano P, Wang J, Yang Y, LuoZ, Mei L, Chien KR, Sussman DJ, Wynshaw-Boris A.2002. Dishevelled 2 is essential for cardiac outflow tractdevelopment, somite segmentation and neural tube clo-sure. Development 129: 5827–5838.
Han YG, Spassky N, Romaguera-Ros M, Garcia-VerdugoJM, Aguilar A, Schneider-Maunoury S, Alvarez-BuyllaA. 2008. Hedgehog signaling and primary cilia are re-quired for the formation of adult neural stem cells. NatNeurosci 11: 277–284.
Henderson DJ, Phillips HM, Chaudhry B. 2006. Vang-like 2and noncanonical Wnt signaling in outflow tract devel-opment. Trends Cardiovasc Med 16: 38–45.
Hildreth V, Webb S, Chaudhry B, Peat JD, Phillips HM,Brown N, Anderson RH, Henderson DJ. 2009. Left car-diac isomerism in the Sonic Hedgehog null mouse. J Anat214: 894–904.
Hirokawa N, Tanaka Y, Okada Y. 2009. Left–right determi-nation: Involvement of molecular motor KIF3, cilia, andnodal flow. Cold Spring Harb Perspect Biol 1: a000802.
Hoffman JI, Kaplan S. 2002. The incidence of congenitalheart disease. J Am Coll Cardiol 39: 1890–1900.
Horani A, Brody SL, Ferkol TW. 2014. Picking up speed:Advances in the genetics of primary ciliary dyskinesia.Pediatr Res 75: 158–164.
Huang P, Schier AF. 2009. Dampened Hedgehog signalingbut normal Wnt signaling in zebrafish without cilia. De-velopment 136: 3089–3098.
Huangfu D, Liu A, Rakeman AS, Murcia NS, Niswander L,Anderson KV. 2003. Hedgehog signalling in the mouserequires intraflagellar transport proteins. Nature 426:83–87.
Kennedy MP, Omran H, Leigh MW, Dell S, Morgan L, Mo-lina PL, Robinson BV, Minnix SL, Olbrich H, Severin T,et al. 2007. Congenital heart disease and other heterotaxicdefects in a large cohort of patients with primary ciliarydyskinesia. Circulation 115: 2814–2821.
Kim SK, Shindo A, Park TJ, Oh EC, Ghosh S, Gray RS, LewisRA, Johnson CA, Attie-Bittach T, Katsanis N, et al. 2010.Planar cell polarity acts through septins to control collec-tive cell movement and ciliogenesis. Science 329: 1337–1340.
Kirby ML, Waldo KL. 1990. Role of neural crest in congenitalheart disease. Circulation 82: 332–340.
Kobayashi T, Dynlacht BD. 2011. Regulating the transitionfrom centriole to basal body. J Cell Biol 193: 435–444.
Kruithof BP, Duim SN, Moerkamp AT, Goumans MJ. 2012.TGF-b and BMP signaling in cardiac cushion formation:Lessons from mice and chicken. Differentiation 84: 89–102.
Levin M, Johnson RL, Stern CD, Kuehn M, Tabin C. 1995. Amolecular pathway determining left–right asymmetry inchick embryogenesis. Cell 82: 803–814.
Li Y, Garrod AS, Madan-Khetarpal S, Sreedher G, McGuireM, Yagi H, Klena NT, Gabriel GC, Khalifa O, Zahid M,et al. 2015a. Respiratory motile cilia dysfunction in apatient with cranioectodermal dysplasia. Am J Med GenetA 167A: 2188–2196.
Li Y, Klena NT, Gabriel GC, Liu X, Kim AJ, Lemke K, Chen Y,Chatterjee B, Devine W, Damerla RR, et al. 2015b. Globalgenetic analysis in mice unveils central role for cilia incongenital heart disease. Nature 521: 520–524.
Li Y, Yagi H, Onuoha EO, Damerla RR, Francis R, Furutani Y,Tariq M, King SM, Hendricks G, Cui C, et al. 2016.DNAH6 and its interactions with PCD genes in hetero-taxy and primary ciliary dyskinesia. PLoS Genet 12:e1005821.
Lienkamp S, Ganner A, Walz G. 2012. Inversin, Wnt signal-ing and primary cilia. Differentiation 83: S49–S55.
Lin AE, Krikov S, Riehle-Colarusso T, Frias JL, Belmont J,Anderka M, Geva T, Getz KD, Botto LD; National BirthDefects Prevention S. 2014. Laterality defects in the na-tional birth defects prevention study (1998–2007): Birthprevalence and descriptive epidemiology. Am J Med Ge-net A 164: 2581–2591.
Liu X, Francis R, Kim AJ, Ramirez R, Chen G, SubramanianR, Anderton S, Kim Y, Wong L, Morgan J, et al. 2014.Interrogating congenital heart defects with noninvasivefetal echocardiography in a mouse forward geneticscreen. Circ Cardiovasc Imaging 7: 31–42.
Lobo J, Zariwala MA, Noone PG. 2015. Primary ciliary dys-kinesia. Semin Respir Crit Care Med 36: 169–179.
MacDonald BT, Tamai K, He X. 2009. Wnt/b-catenin sig-naling: Components, mechanisms, and diseases. Dev Cell17: 9–26.
Marelli A, Miller SP, Marino BS, Jefferson AL, NewburgerJW. 2016. Brain in congenital heart disease across thelifespan: The cumulative burden of injury. Circulation133: 1951–1962.
May-Simera HL, Kelley MW. 2012. Cilia, Wnt signaling, andthe cytoskeleton. Cilia 1: 7.
McGrath J, Somlo S, Makova S, Tian X, Brueckner M. 2003.Two populations of node monocilia initiate left–rightasymmetry in the mouse. Cell 114: 61–73.
Montcouquiol M, Rachel RA, Lanford PJ, Copeland NG,Jenkins NA, Kelley MW. 2003. Identification of Vangl2and Scrb1 as planar polarity genes in mammals. Nature423: 173–177.
Nakamura T, Hamada H. 2012. Left–right patterning: Con-served and divergent mechanisms. Development 139:3257–3262.
Nakhleh N, Francis R, Giese RA, Tian X, Li Y, Zariwala MA,Yagi H, Khalifa O, Kureshi S, Chatterjee B, et al. 2012.
N.T. Klena et al.
16 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

High prevalence of respiratory ciliary dysfunction in con-genital heart disease patients with heterotaxy. Circulation125: 2232–2242.
Newburger JW, Sleeper LA, Bellinger DC, Goldberg CS,Tabbutt S, Lu M, Mussatto KA, Williams IA, GustafsonKE, Mital S, et al. 2012. Early developmental outcome inchildren with hypoplastic left heart syndrome and relatedanomalies: The single ventricle reconstruction trial. Cir-culation 125: 2081–2091.
Nomura-Kitabayashi A, Phoon CK, Kishigami S, RosenthalJ, Yamauchi Y, Abe K, Yamamura K, Samtani R, LoCW, Mishina Y. 2009. Outflow tract cushions performa critical valve-like function in the early embryonicheart requiring BMPRIA-mediated signaling in cardiacneural crest. Am J Physiol Heart Circ Physiol 297: H1617–H1628.
Nonaka S, Shiratori H, Saijoh Y, Hamada H. 2002. Deter-mination of left–right patterning of the mouse embryoby artificial nodal flow. Nature 418: 96–99.
Oh EC, Katsanis N. 2013. Context-dependent regulation ofWnt signaling through the primary cilium. J Am SocNephrol 24: 10–18.
Oyen N, Poulsen G, Boyd HA, Wohlfahrt J, Jensen PK, Mel-bye M. 2009. Recurrence of congenital heart defects infamilies. Circulation 120: 295–301.
Paudyal A, Damrau C, Patterson VL, Ermakov A, FormstoneC, Lalanne Z, Wells S, Lu X, Norris DP, Dean CH, et al.2010. The novel mouse mutant, chuzhoi, has disruptionof Ptk7 protein and exhibits defects in neural tube, heartand lung development and abnormal planar cell polarityin the ear. BMC Dev Biol 10: 87.
Phillips HM, Rhee HJ, Murdoch JN, Hildreth V, Peat JD,Anderson RH, Copp AJ, Chaudhry B, Henderson DJ.2007. Disruption of planar cell polarity signaling resultsin congenital heart defects and cardiomyopathy attribut-able to early cardiomyocyte disorganization. Circ Res 101:137–145.
Potts JD, Runyan RB. 1989. Epithelial-mesenchymal celltransformation in the embryonic heart can be mediated,in part, by transforming growth factor b. Dev Biol 134:392–401.
Rash JE, Shay JW, Biesele JJ. 1969. Cilia in cardiac differen-tiation. J Ultrastruct Res 29: 470–484.
Reiter JF, Blacque OE, Leroux MR. 2012. The base of thecilium: Roles for transition fibres and the transition zonein ciliary formation, maintenance and compartmentali-zation. EMBO Rep 13: 608–618.
Ross AJ, May-Simera H, Eichers ER, Kai M, Hill J, Jagger DJ,Leitch CC, Chapple JP, Munro PM, Fisher S, et al. 2005.Disruption of Bardet–Biedl syndrome ciliary proteinsperturbs planar cell polarity in vertebrates. Nat Genet37: 1135–1140.
Sanford LP, Ormsby I, Gittenberger-de Groot AC, SariolaH, Friedman R, Boivin GP, Cardell EL, Doetschman T.1997. TGF-b2 knockout mice have multiple develop-mental defects that are non-overlapping with otherTGF-b knockout phenotypes. Development 124: 2659–2670.
Schlessinger K, Hall A, Tolwinski N. 2009. Wnt signalingpathways meet Rho GTPases. Genes Dev 23: 265–277.
Shapiro AJ, Davis SD, Ferkol T, Dell SD, Rosenfeld M, Oli-vier KN, Sagel SD, Milla C, Zariwala MA, Wolf W, et al.
2014. Laterality defects other than situs inversus totalis inprimary ciliary dyskinesia: Insights into situs ambiguusand heterotaxy. Chest 146: 1176–1186.
Shen Y, Leatherbury L, Rosenthal J, Yu Q, Pappas MA,Wessels A, Lucas J, Siegfried B, Chatterjee B, Svenson K,et al. 2005. Cardiovascular phenotyping of fetal miceby noninvasive high-frequency ultrasound facilitates re-covery of ENU-induced mutations causing congenitalcardiac and extracardiac defects. Physiol Genomics 24:23–36.
� Shinohara K, Hamada H. 2016. Cilia in left–right symmetrybreaking. Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028282.
Simons M, Mlodzik M. 2008. Planar cell polarity signaling:From fly development to human disease. Annu Rev Genet42: 517–540.
Simons M, Gloy J, Ganner A, Bullerkotte A, Bashkurov M,Kronig C, Schermer B, Benzing T, Cabello OA, Jenny A,et al. 2005. Inversin, the gene product mutated in neph-ronophthisis type II, functions as a molecular switchbetween Wnt signaling pathways. Nat Genet 37: 537–543.
Sinha T, Wang B, Evans S, Wynshaw-Boris A, Wang J. 2012.Disheveled mediated planar cell polarity signaling is re-quired in the second heart field lineage for outflow tractmorphogenesis. Dev Biol 370: 135–144.
Slough J, Cooney L, Brueckner M. 2008. Monocilia in theembryonic mouse heart suggest a direct role for cilia incardiac morphogenesis. Dev Dyn 237: 2304–2314.
Sorokin S. 1962. Centrioles and the formation of rudimen-tary cilia by fibroblasts and smooth muscle cells. J CellBiol 15: 363–377.
Srour M, Schwartzentruber J, Hamdan FF, Ospina LH,Patry L, Labuda D, Massicotte C, Dobrzeniecka S,Capo-Chichi JM, Papillon-Cavanagh S, et al. 2012.Mutations in C5ORF42 cause Joubert syndrome inthe French Canadian population. Am J Hum Genet90: 693–700.
Stottmann RW, Tran PV, Turbe-Doan A, Beier DR. 2009.Ttc21b is required to restrict Sonic Hedgehog activityin the developing mouse forebrain. Dev Biol 335: 166–178.
Tada M, Smith JC. 2000. Xwnt11 is a target of XenopusBrachyury: Regulation of gastrulation movements via Di-shevelled, but not through the canonical Wnt pathway.Development 127: 2227–2238.
Tan SY, Rosenthal J, Zhao XQ, Francis RJ, Chatterjee B, SabolSL, Linask KL, Bracero L, Connelly PS, Daniels MP, et al.2007. Heterotaxy and complex structural heart defects ina mutant mouse model of primary ciliary dyskinesia. JClin Invest 117: 3742–3752.
Tobin JL, Beales PL. 2009. The nonmotile ciliopathies. GenetMed 11: 386–402.
van der Bom T, Bouma BJ, Meijboom FJ, ZwindermanAH, Mulder BJ. 2012. The prevalence of adult con-genital heart disease, results from a systematic reviewand evidence based calculation. Am Heart J 164: 568–575.
Verzi MP, McCulley DJ, De Val S, Dodou E, Black BL.2005. The right ventricle, outflow tract, and ventricularseptum comprise a restricted expression domain within
Cilia and Ciliopathies in Congenital Heart Disease
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266 17
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

the secondary/anterior heart field. Dev Biol 287: 134–145.
von Gise A, Pu WT. 2012. Endocardial and epicardial epi-thelial to mesenchymal transitions in heart developmentand disease. Circ Res 110: 1628–1645.
Wallingford JB, Mitchell B. 2011. Strange as it may seem:The many links between Wnt signaling, planar cell po-larity, and cilia. Genes Dev 25: 201–213.
Washington Smoak I, Byrd NA, Abu-Issa R, Goddeeris MM,Anderson R, Morris J, Yamamura K, Klingensmith J,Meyers EN. 2005. Sonic hedgehog is required for cardiacoutflow tract and neural crest cell development. Dev Biol283: 357–372.
Willaredt MA, Gorgas K, Gardner HA, Tucker KL. 2012.Multiple essential roles for primary cilia in heart devel-opment. Cilia 1: 23.
Yoshiba S, Shiratori H, Kuo IY, Kawasumi A, Shinohara K,Nonaka S, Asai Y, Sasaki G, Belo JA, Sasaki H, et al.2012. Cilia at the node of mouse embryos sense fluidflow for left–right determination via Pkd2. Science 338:226–231.
Zaidi S, Choi M, Wakimoto H, Ma L, Jiang J, Overton JD,Romano-Adesman A, Bjornson RD, Breitbart RE,Brown KK, et al. 2013. De novo mutations in histone-modifying genes in congenital heart disease. Nature 498:220–223.
N.T. Klena et al.
18 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a028266
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

published online February 3, 2017Cold Spring Harb Perspect Biol Nikolai T. Klena, Brian C. Gibbs and Cecilia W. Lo Cilia and Ciliopathies in Congenital Heart Disease
Subject Collection Cilia
Polycystic Kidney DiseaseCiliary Mechanisms of Cyst Formation in
SomloMing Ma, Anna-Rachel Gallagher and Stefan
Right Symmetry Breaking−Cilia in LeftKyosuke Shinohara and Hiroshi Hamada
Photoreceptor Cilia and Retinal CiliopathiesKinga M. Bujakowska, Qin Liu and Eric A. Pierce Ciliopathies
Discovery, Diagnosis, and Etiology of Craniofacial
Elizabeth N. Schock and Samantha A. BrugmannG-Protein-Coupled Receptor Signaling in Cilia
Kirk Mykytyn and Candice AskwithAxoneme Structure from Motile Cilia
Takashi IshikawaEvolution of Cilia
David R. MitchellCilia and Ciliopathies in Congenital Heart Disease
Nikolai T. Klena, Brian C. Gibbs and Cecilia W. Lo
Centriole ElongationCytoplasmic Ciliogenesis and Postaxonemal Transition Zone Migration: A Mechanism for
BasiriTomer Avidor-Reiss, Andrew Ha and Marcus L.
Sperm Sensory Signaling
KauppDagmar Wachten, Jan F. Jikeli and U. Benjamin
Cilia and Obesity
BerbariChristian Vaisse, Jeremy F. Reiter and Nicolas F.
) Signalingβ (TGF-βFactor Tyrosine Kinase (RTK) and Transforming Growth Primary Cilia and Coordination of Receptor
B. Mogensen, et al.Søren T. Christensen, Stine K. Morthorst, Johanne
CiliaPosttranslational Modifications of Tubulin and
et al.Dorota Wloga, Ewa Joachimiak, Panagiota Louka,
Primary Cilia and Mammalian Hedgehog SignalingFiona Bangs and Kathryn V. Anderson
FlagellaRegulation, Assembly, and Evolution of Cilia and
A Snapshot of the Motility−−Radial Spokes
Xiaoyan Zhu, Yi Liu and Pinfen Yang
Cilia and Mucociliary Clearance
OstrowskiXimena M. Bustamante-Marin and Lawrence E.
http://cshperspectives.cshlp.org/cgi/collection/ For additional articles in this collection, see
Copyright © 2017 Cold Spring Harbor Laboratory Press; all rights reserved
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

http://cshperspectives.cshlp.org/cgi/collection/ For additional articles in this collection, see
Copyright © 2017 Cold Spring Harbor Laboratory Press; all rights reserved
on April 3, 2022 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from