生体膜の動的構造と生体膜の機能ohki/biophys/dec01.pdf生体膜の動的構造と生体膜の機能...
TRANSCRIPT

生体膜の動的構造と生体膜の機能
脂質膜の構造と物性が蛋白質との相互作用を介して生体膜の機能(活性)に反映
両親媒性の脂質分子は水中で自発的に膜構造に集合し、蛋白質が組み込まれて生体膜となる。
相転移、相分離などの統計的な現象を示す。
側方拡散、回転拡散、フリップ・フロップなどの膜流動性を示す。
7-10 nm
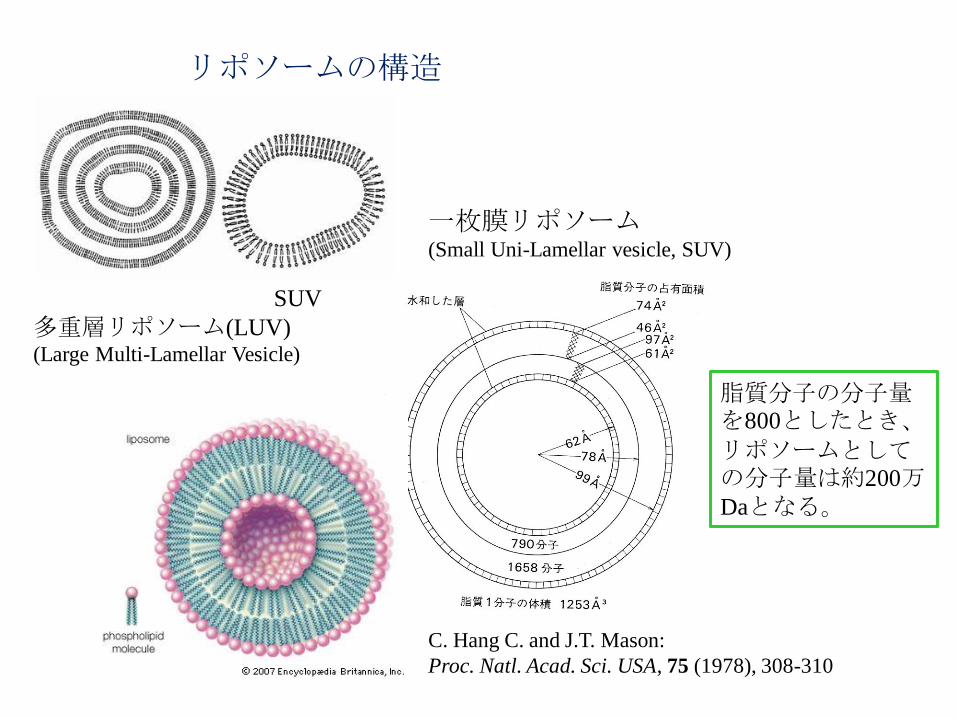
リポソームの構造
一枚膜リポソーム (Small Uni-Lamellar vesicle, SUV)
C. Hang C. and J.T. Mason:
Proc. Natl. Acad. Sci. USA, 75 (1978), 308-310
脂質分子の分子量を800としたとき、
リポソームとしての分子量は約200万Daとなる。
SUV
多重層リポソーム(LUV) (Large Multi-Lamellar Vesicle)

DPPC脂質膜の温度変化のX線回折測定

Temperature [℃]
脂肪酸側鎖長の異なるホスファチジルコリン脂質膜の相転移
DSC測定
脂肪酸側鎖の炭素数
DMPC:14
DPPC:16
DSPC:18
分子間の相互作用(Van del Waals力)が
大きい程相転移温度が高くなる。

DSPC
C18:0/C18:0-PC
C18:1Δn,cis/C18:1Δn,cis-PC
C18:0/C18:1Δn,cis-PC
脂肪酸側鎖へのシス二重結合の導入による相転移温度の変化
オレイン酸
C18:1
リノレイン酸
C18:2
飽和ホスファチジルコリン 1つのシス結合
2つのシス結合
ステアリン酸
C18:0

各種脂質組成をもつリン脂質の構造と相転移温度
相転移温度: 降下 上昇
脂肪酸鎖長: 短 長
Trans/cis結合: cis trans
極性頭部: 大 小
脂肪酸結合型: エーテル エステル

全ての生物は多細胞生物も単細胞生物も個体として、それを取り囲む環境の中で生息しており、常に変動する種々の環境因子の下で一定の状態を維持しようとする能力(ホメオスタシス=恒常性)を備えている。 多細胞生物の各細胞も直接的な周囲は水環境を介した細胞間相互作用の下にある。 個々の細胞は自身の遺伝情報をもっており、環境から受けた刺激に対して応答する能力を示すことができる。
細胞と外界の環境
Swiss3T3細胞
環境
刺激 (物理的・化学的)
応答

環境:温度因子
物性の温度依存性
生物機能への温度因子による影響
進化の過程での温度適応機構の獲得と恒温動物の誕生
恒温動物のラットやチャイニーズハムスターから株化した培養細胞で観察される温度適応は進化の過程で蓄積された遺伝子が発現していると考えられる。
環境の温度因子(物理的因子)
生体膜と温度適応:生体膜の基本構造である脂質二分子膜は相転移・相分離、側方拡散、回転拡散などの物性は生体膜の脂質構成に依存している。脂質の代謝変換と温度適応機構が相関している。脂質変換は細胞の情報伝達機構でも主要な伝達経路となっており、細胞の機能制御が環境からの刺激に対する応答であると理解できる。 つまり、細胞の環境適応機構と情報伝達機構は発生・分化の過程も含む細胞の機能制御の本質的な部分で共通の起源をもつものと考えられる。
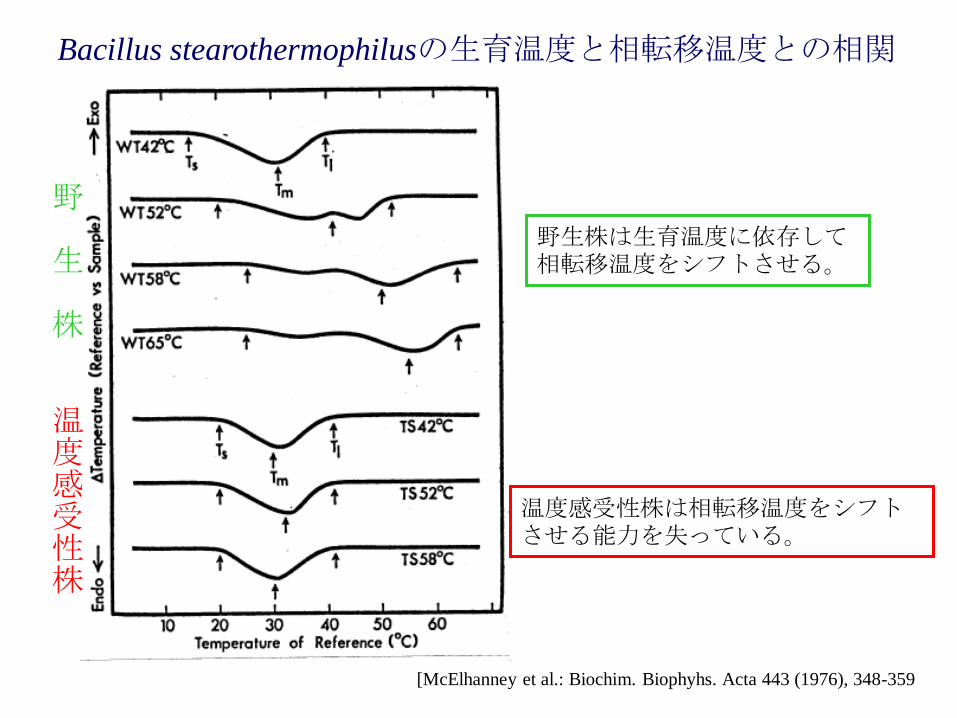
Bacillus stearothermophilusの生育温度と相転移温度との相関
[McElhanney et al.: Biochim. Biophyhs. Acta 443 (1976), 348-359
野
生
株
温度感受性株
野生株は生育温度に依存して相転移温度をシフトさせる。
温度感受性株は相転移温度をシフトさせる能力を失っている。
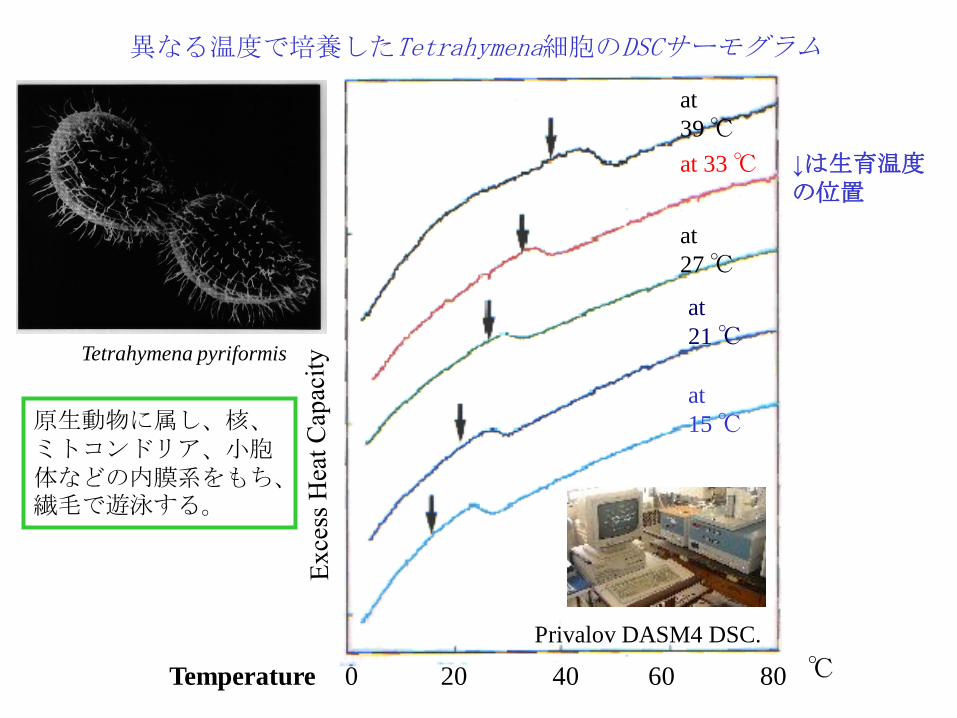
at
39 ℃
at 33 ℃
at
21 ℃
at
15 ℃
異なる温度で培養したTetrahymena細胞のDSCサーモグラム
at
27 ℃
Temperature ℃ 0 20 40 60 80
Privalov DASM4 DSC.
Tetrahymena pyriformis
原生動物に属し、核、ミトコンドリア、小胞体などの内膜系をもち、繊毛で遊泳する。
↓は生育温度の位置

desA:Δ12 (炭素数18の脂肪酸のカルボキシル基側から
12番目の位置に二重結合を導入)
desB:ω3 (炭素数18の脂肪酸のメチル基末端から
3番目の位置に二重結合を導入)
desC:Δ9 (炭素数18の脂肪酸のカルボキシル基側から
9番目の位置に二重結合を導入)
desD:Δ6 (炭素数18の脂肪酸のカルボキシル基側から
6番目の位置に二重結合を導入)
シアノバクテリア Cyanobacterium
(Synechocystis sp. PCC 6803) の脂肪酸不飽和化酵素をコードする遺伝子
特定の遺伝子を破壊して、その遺伝子がどのような作用をもつかを調べるノックアウト法を用い、培養温度変化時の不飽和化酵素の役割を調べた。
光合成細菌(藍藻)
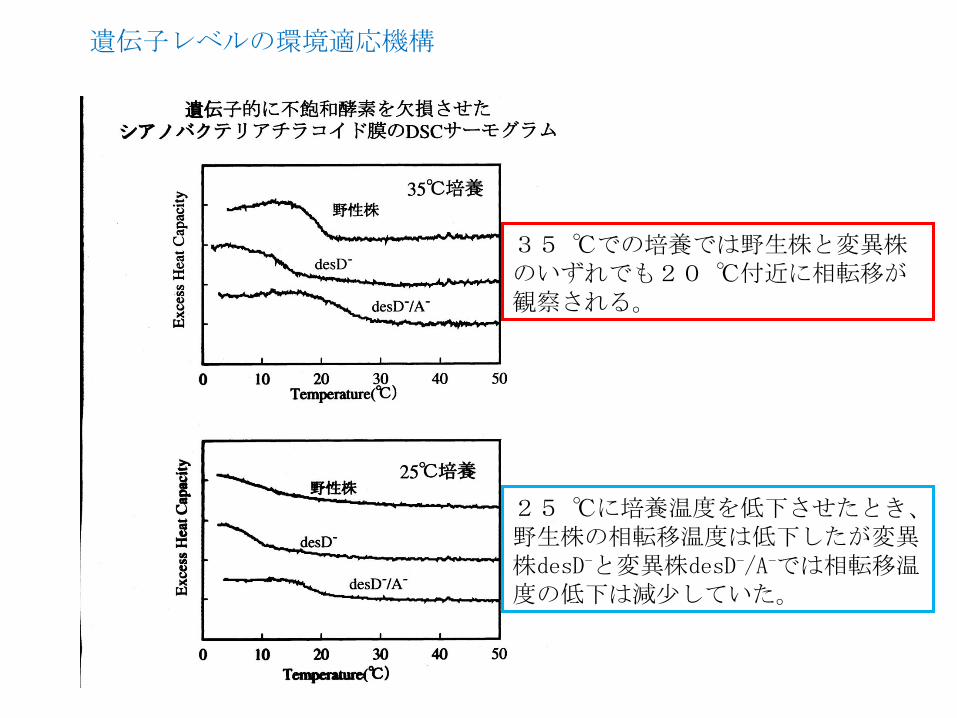
35 ℃での培養では野生株と変異株のいずれでも20 ℃付近に相転移が観察される。
25 ℃に培養温度を低下させたとき、野生株の相転移温度は低下したが変異株desD-と変異株desD-/A-では相転移温度の低下は減尐していた。
遺伝子レベルの環境適応機構

生育温度[℃] 測定温度[℃] 粘度[poise]
抽出脂質15 15 1.8
30 30 1.9
37 37 1.8 43 43 2.0 43 15 15 大腸菌膜 23 23 2.5 37 37 2.5
大腸菌のHomeoviscous adaptation
23 37 1.0
※脂肪酸エステルのスピンプローブを用いて、ESRスペクトルから粘度を求めている。

電子スピン共鳴とゼーマン分裂
磁場中にある磁気モーメントがもつエネルギーは古典的には角度に依存する内積となる。
量子力学では電子スピンとして量子化されて+1/2
と-1/2のスピン量子数となる
エネルギーの分離幅に等しいエネルギーに対して磁気共鳴が生じる
生体膜の粘度(1/流動性)測定

ゼーマン分裂と電子スピン共鳴
共鳴条件 ⊿E=gβH

スピンラベル法: 常磁性分子として、安定なニトロキシドラジカル類を導
入してスピン・プローブとし、そのESRスペクトルから生体分子系にダイナミックスについての知見を得る方法
[新生化学実験講座20 機器分析概論 電子スピン共鳴 pp.275-288
(大木和夫・大西俊一)(東京化学同人 1993)]
[現代物理学・展開シリーズ8 生物物理学 磁気共鳴 pp.43-51
(大木和夫・宮田英威)(朝倉書店 2010)]
その共鳴条件は9.2 GHz (Xバンドのマイクロ波)に対して 3280 G となり、Spin Hamiltonianは
Hamiltonian = βH・g・S + S・T・I - gN βN H・I であり、そのテンソルの主値は gxx = 2.0088 Txx = 5.9 gauss
gyy = 2.0058 Tyy = 5.4 gauss
gzz = 2.0022 Tzz = 32.9 gauss であり、
Order parameterはこの異方性を利用する。
用いる電磁波はマイクロ波の領域であり、可視光の領域で不透明な物質に対しても測定が可能である。
生体膜の粘度(1/流動性)測定

リン脂質スピンプローブによる膜流動性の測定
S=T -T a
Tzz-Txx a’・
回転軸
速い軸対称回転運動 θはNOラジカルの2pz軌道が回転軸となす角
このスペクトルから膜流動性の指標となるオーダーパラメーター(S)を定義する。
Txx = Tyy, <α2> + <β2> + <γ2> = 1 を用いて TZZ' = a + ・(Tzz - Txx)・S
a = Tr(T) = Tr(T')
S = (3<γ2> - 1) α, β, γはZ のx,y,z 主軸系に対する方向余弦
gZZ = < α2>gxx + < β2>gyy + < γ2>gzz
TZZ = < α2>Txx + < β2>Tyy + < γ2>Tzz
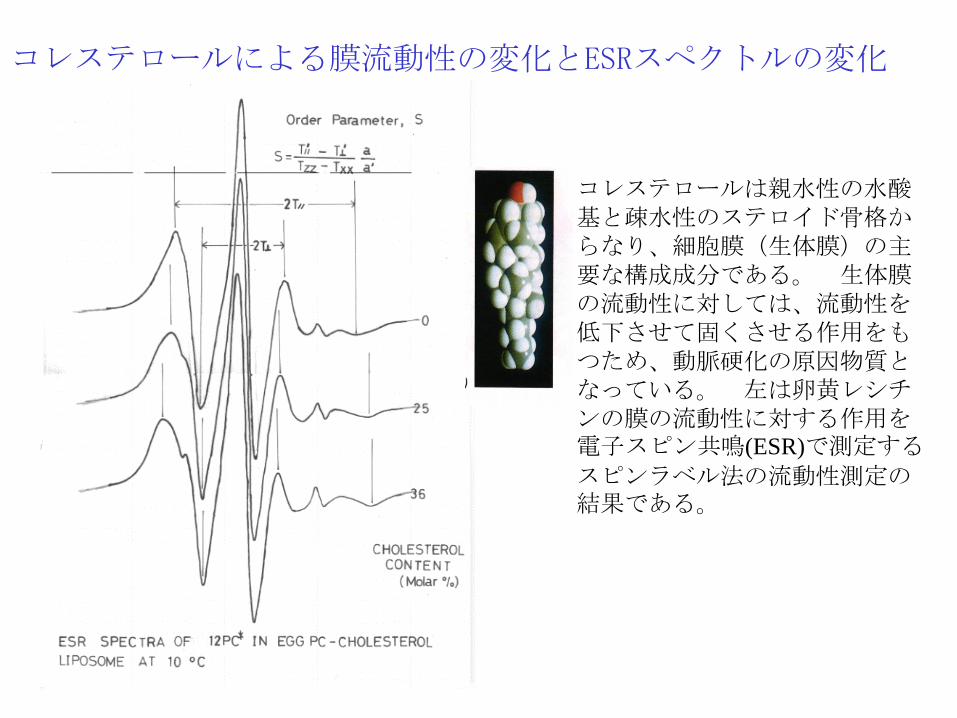
コレステロールによる膜流動性の変化とESRスペクトルの変化
コレステロールは親水性の水酸基と疎水性のステロイド骨格からなり、細胞膜(生体膜)の主要な構成成分である。 生体膜の流動性に対しては、流動性を低下させて固くさせる作用をもつため、動脈硬化の原因物質となっている。 左は卵黄レシチンの膜の流動性に対する作用を電子スピン共鳴(ESR)で測定する
スピンラベル法の流動性測定の結果である。

・Nitroxide radicalの運動が等方的なとき、球の運動で近似
・Spin probeを半径rの球と仮定
ブラウン運動についてのEinsteinの関係 τC=4πr3η/kT
「粘度ηと回転相関時間τC の関係を示している。」
・ESRスペクトルの線幅についての理論は線幅ΔHと回転相関時間τCの関係、
ΔH=τC(A+B・m+C・m2)
(mは核の量子数で、A,B,Cは定数)を与えるので、生体膜および脂質二重層膜の粘度測定を可能にする。
実際の表式は τC =(3πgβΔH(0)/h[c1m-c2m2])
(h(m):ピークの高さ、 ΔH(0):中央ピークの幅 ここで、
c1 =-(16π|β|H/45h)[Tzz-Txx][gzz-(gxx+gyy)/2]
Hamiltonian = βH・g・S + S・T・I - gN βN H・I
=(time-independent)-{(time-dependent) - (time-independent)} Averaged out (can be neglected) Effective Hamiltonian=βgZZSZHZ + hTZZ'SZIZ - gNβNIZHZ = β(<α2>gxx + < β
2>gyy + < γ2>gzz)SZHZ + h(<α
2>Txx + < β2>Tyy + <
γ2>Tzz)SZIZ - gN βNIZHZ α, β, γはZ のx,y,z 主軸系に対する方向余弦 gZZ = < α
2>gxx + < β2>gyy + < γ
2>gzz TZZ = < α2>Txx + < β2>Tyy + < γ
2>Tzzc2 =(2π2/9)[Tzz-Txx]2
脂質スピンプローブによる膜粘度の測定 生体膜の粘度(1/流動性)測定
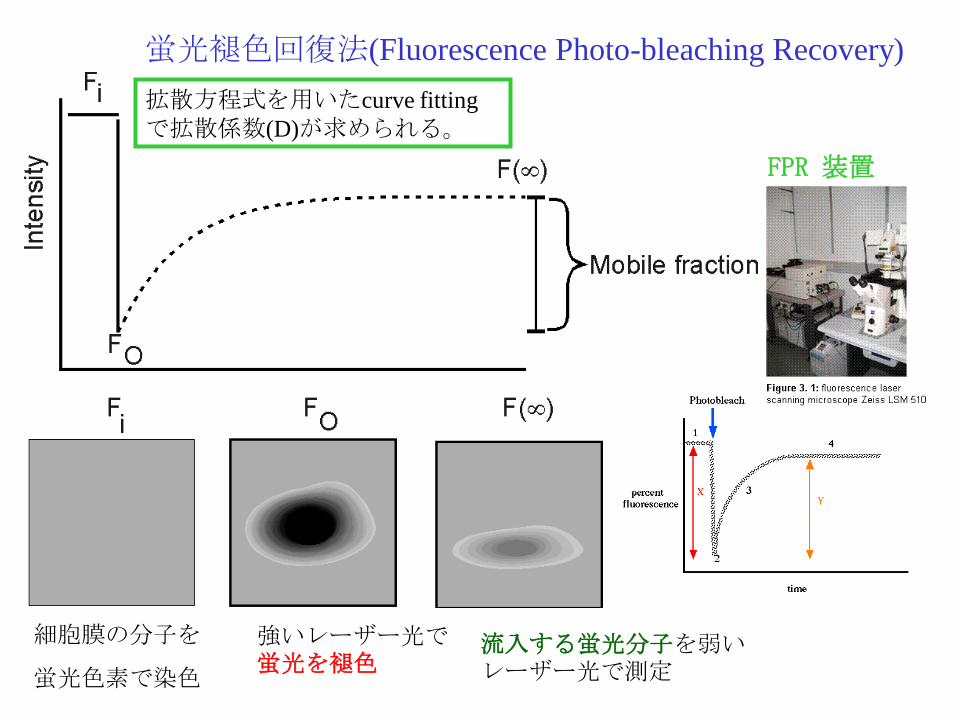
蛍光褪色回復法(Fluorescence Photo-bleaching Recovery)
細胞膜の分子を
蛍光色素で染色
強いレーザー光で蛍光を褪色
流入する蛍光分子を弱いレーザー光で測定
拡散方程式を用いたcurve fitting
で拡散係数(D)が求められる。
FPR 装置

蛍光褪色回復法(Fluorescence Photobleach Recovery)
[ D. Axelrod, D.E. Koppel, J. Schlessinger, E. Elson and W.W. Webb (1976) Biophys. J. 16, 1055-1069]
・蛍光色素が光褪色により非蛍光性になる過程は速度定数 αI(r)に従う 単純な非可逆一次反応と仮定する。 輸送がない条件下で未褪色の蛍光色素の濃度は位置 r、時間 tにおいて、 :褪色光の強度 拡散により1種の蛍光色素が側方に輸送される微分方程式は D :拡散係数 V0:x方向での一様な流れの速度 この微分方程式の解を 境界条件:C(∞,t)=C0・・・蛍光色素の一様な分布 初期条件:C(r,0)=C0 exp[-αTI(r)] で求める
t)(r,c(r)Itd
t)c(r,d (r)I
x
t)(r,cV-t)(r,cD
t
t)(r,c0
2
側方拡散測定法

相 (phase)
明確な物理的な境界により区別される物質系の均一な部分
気体→気相、 液体→液相、固体→固相
・相転移は異なる相構造の間を温度変化などで協同的に移行する現象であるが、異なる相構造が共存して存在することがある。 このとき、異なる相(phase)の間に相平衡が存在しており、相分離の状態は相図として表現で
きる。 リン脂質二重膜では脂質2成分系での相分離が広く研究されている。 生体膜は脂質の多成分系であるが、相分離の存在が観察されておりこれは生体膜内に不均一な領域を形成させることから、その物性の相違により生体膜の機能制御が行われていると推測されている。
・構造相転移(気相、液相、固相間の転移など)
・磁気相転移(常磁性、強磁性、反強磁性などの間での転移)
・金属-絶縁体転移(モット転移など)
・常伝導-超伝導転移(超伝導)
・常誘電体-強誘電体転移
・真空の相転移(宇宙論)

物質の相転移は自由エネルギーから考察できる。温度T、体積Vのとき、
ヘルムホルツの自由エネルギーが最小となる相が最も安定となる。
相1と相2のヘルムホルツの自由エネルギーをそれぞれ、F1(T,V)、F2(T,V)
とすればF1(T,V)<F2(T,V)のとき、相1が実現する。
TとVが変化するとき、F1(T,V)=F2(T,V)が大小関係の境界となり、転移点を
示す方程式となる。
体積Vを一定に保って、温度を低下させるとTに関しての解であるTc(転移温度)が両関数の交点となる。
この点で、高温相2から低温相1に転移することになる。転移点での値は等しいが、その微係数(傾き)も等しいとは限らない。ここで、内部エネルギーEとエントロピーSについて、S1(V,Tc)≠S2(V,Tc)、E1(V,Tc)≠E2(V,Tc)であれば、潜熱はL=Tc(S2-S1)=E2-E1となる。
温度
自由エネルギー
転移点で が不連続な場合を第1種または1次の相転移と呼ぶ。 T
F
相転移 (phase transition)

温度Tと圧力Pが与えられる場合はギブスの自由エネルギーから転移温度
TcはG1(T,P)=G2(T,P)の条件により求められる。1次相転移における潜熱
はエントロピーSとエンタルピーHについて、L=Tc(S2-S1)=H2-H1となる。
ここで、 , であるので、1次転移では体積(密度)、または、
エントロピーの不連続変化が生じている。
Vp
G
S
T
G
相平衡
水分子が氷(固体)、水(液体)、水蒸気(気体)の3つの相を取るように、同じ物質が複数の異なる相を取るとき、これらの相の間で平衡状態にあることを相平衡と呼ぶ。熱的、拡散的、機械的に接触している二つの相(相1と相2)が共存して平衡にある条件は温度、圧力、化学ポテンシャルについて、両相の値が等しいことであり、次のようになる。
T1 =T2;p1 =p2; μ1 =μ2
気液平衡/液固平衡
気体(gas)の相と液体(liquid)の相が平衡状態にある気液平衡では Tl=Tg ; pl=pg; μl =μg となる。 2相が共存するならば、そこに存在する同種の物質の化学ポテンシャルは両相で等しくなければならず、μl (p, T)=μg (p, T)である。