balancing efficacy and safety of an anti-dll4 antibody ... · once weekly for 3 weeks. vehicle was...
TRANSCRIPT

Cancer Therapy: Preclinical
Balancing Efficacy and Safety of an Anti-DLL4Antibody through Pharmacokinetic ModulationJessica A. Couch1, Gu Zhang2, Joseph C. Beyer1, Christina L. Zuch de Zafra1,Priyanka Gupta1, Amrita V. Kamath1, Nicholas Lewin-Koh3, Jacqueline Tarrant1,KrishnaP.Allamneni1,GaryCain1, SharonYee4, SarajaneRoss4,RyanCook5, SiaoPingTsai5,Jane Ruppel1, John Brady Ridgway2, Maciej Paluch6, Philip E. Hass6, Jayme Franklin7, andMinhong Yan2
Abstract
Purpose: Although agents targeting Delta-like ligand 4(DLL4) have shown great promise for angiogenesis-based can-cer therapy, findings in recent studies have raised serious safetyconcerns. To further evaluate the potential for therapeutic tar-geting of the DLL4 pathway, we pursued a novel strategy toreduce toxicities related to DLL4 inhibition by modulating thepharmacokinetic (PK) properties of an anti-DLL4 antibody.
Experimental Design: The F(ab0)2 fragment of anti-DLL4 anti-body (anti-DLL4 F(ab0)2) was generated and assessed in efficacyand toxicity studies.
Results: Anti-DLL4 F(ab0)2 enables greater control overthe extent and duration of DLL4 inhibition, such that
intermittent dosing of anti-DLL4 F(ab0)2 can maintainsignificant antitumor activity while markedly mitigatingknown toxicities associated with continuous pathwayinhibition.
Conclusions: PK modulation has potentially broad implica-tions for development of antibody-based therapeutics.Our safety studies with anti-DLL4 F(ab0)2 also provide newevidence reinforcing the notion that the DLL4 pathway isextremely sensitive to pharmacologic perturbation, furtherunderscoring the importance of exercising caution to safelyharness this potent pathway in humans. Clin Cancer Res; 22(6);1469–79. �2015 AACR.
IntroductionInhibition of tumor angiogenesis has become a prominent
strategy in cancer therapy because tumor growth is criticallydependent on neovascularization to support ever-increasingmet-abolic demands. In addition to vascular endothelial growth factor(VEGF; refs. 1–3), members of the Notch signaling pathway,including the NOTCH1 receptor and its Delta-like ligand 4(DLL4), have been recognized as attractive targets for tumorangiogenesis due to their essential roles in vascular development.DLL4 was initially identified as an endothelium-specific Notchligand (4–7), and plays a unique role in embryonic and postnatalvascular development, wound healing, as well as tumor angio-
genesis (8, 9). Specifically, inhibition of DLL4 signaling duringangiogenesis disrupts the dynamic balance between tip and stalkcells (10), resulting in a chaotic vascular network characterized byexcessive sprouting and branching, dramatically reduced vessellumen size, and increased vascular density (11–13). DLL4 neu-tralizing (anti-DLL4) antibodies have been associatedwith robustantitumor activity in a wide range of preclinical efficacy models(14–18), likely by promoting nonproductive angiogenesis thatimpairs tumor microcirculation and induces hypoxia (19, 20). Inaddition to the potent antitumor activity observed following anti-DLL4 treatment, additive antitumor activity has also beenobserved in combination with anti-VEGF therapy and/or chemo-therapy (14, 16–18, 21). Based on the important role of DLL4 invascular biology aswell as the broad preclinical antitumor activityof anti-DLL4 monoclonal antibodies, several anti-DLL4 mole-cules are currently being investigated in clinical trials as potentialcancer therapeutics (22).
Despite the promise, preclinical studies have raised safetyconcerns related to inhibiting DLL4. Specifically, treatment ofmice, rats, and cynomolgus monkeys with anti-DLL4 resulted inpathologic changes in the liver, including marked atrophy ofcentrilobular hepatic cords, dilation of centrilobular hepaticsinusoids (sinusoidal dilation), bile duct proliferation, and ele-vated liver function tests (23). In addition to liver findings,vascular neoplasms occurred in skin, heart, and lungs of malerats after 8 weeks of continuous anti-DLL4 exposure (23). Finally,although vascular neoplasms were not observed in monkeys,dose-dependent, moderate-to-severe acute hemolytic anemiaoccurred in some animals, leading to mortality in the most severecases. Clinical trials of the DLL4-targeting antibody OMP-21M18
1Development Sciences, Genentech, Inc., South San Francisco, Cali-fornia. 2Molecular Oncology, Genentech, Inc., South San Francisco,California. 3Biostatistics, Genentech, Inc., South San Francisco, Cali-fornia. 4In Vivo Pharmacology, Genentech, Inc., South San Francisco,California. 5Biochemical and Cellular Pharmacology, South San Fran-cisco, California. 6Protein Chemistry, Genentech, Inc., South San Fran-cisco, California. 7Purification Development, Genentech, Inc., SouthSan Francisco, California.
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
Current address for K.P. Allamneni: Jazz Pharmaceuticals, Palo Alto, California.
Corresponding Author: Minhong Yan, Genentech, Inc., 1 DNA Way, M/S 93B,South San Francisco, CA 94080. Phone: 650-225-5691; Fax: 650-225-5691;E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-15-1380
�2015 American Association for Cancer Research.
ClinicalCancerResearch
www.aacrjournals.org 1469
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

have also revealed safety concerns in humans, including grade IIIasymptomatic hypertension, that were not noted in publishedpreclinical studies (22).
As the outcome of Notch signaling is highly dependent on thecontext, duration, and strength of pathway activation/inhibition,directly manipulating any of these parameters could offer anopportunity to improve the therapeutic window associated withtargeting DLL4. We hypothesized that incomplete or intermittentblockade of DLL4 may have the potential to reduce dose-relatedliver and vascular toxicities while still maintaining efficacy. Weexplored a novel strategy of inhibiting DLL4 by reducing the half-life of an anti-DLL4 IgG1 antibody (16, 23), and found thataltering the pharmacokinetic (PK) profile by administering arapidly clearing F(ab0)2 antibody fragment allowed more flexiblecontrol over the extent and duration of DLL4 inhibition. Accord-ingly, we demonstrated that pulsatile DLL4 inhibition with ananti-DLL4 F(ab0)2 antibody fragmentmaintained significant anti-tumor efficacy, yet ameliorated the continuous pathway block-ade-associated toxicities of the full-length IgG1 molecule. How-ever, the observation of unexpected safety-related findings furtherunderscores the importance of understanding both the contextand duration of NOTCH1 inhibition associated with therapiestargeting the DLL4 pathway.
Materials and MethodsExperiments were designed to explore the hypothesis that the
extent and duration of DLL4 inhibition is related to bothantitumor efficacy and pathway-related toxicity, and that itmay be possible to maintain efficacy while reducing toxicityto enhance the therapeutic potential of an anti-DLL4 antibodyin humans. To test this hypothesis, we altered the PK profile(i.e., half-life) of an anti-DLL4 neutralizing IgG1 antibody.Efficacy was evaluated in human tumor xenograft mouse mod-els, whereas toxicity was evaluated in normal mice, rats, andmonkeys.
All animal studies were conducted in accordance with theAnimal Welfare Act and the Guide for the Care and Use ofLaboratory Animals (NIH Publication 85-23, revised 1985). An
Institutional AnimalCare andUseCommittee (IACUC) approvedall animal protocols.
Generation of anti-DLL4 IgG1 lesser affinity variantsThe anti-DLL4 IgG1 antibody (YW152F; ref. 16) was subjected
to site-directed mutagenesis via alanine replacement of thosesingle amino acid residues in the complementarity determiningregions (CDR) that are predicted to be involved in antigenbinding.
Production of anti-DLL4 F(ab0)2Anti-DLL4 F(ab0)2 was prepared by using pepsin-based solu-
tion digestion of the full-length anti-DLL4 parental antibody(YW152F; see Supplemental methods).
Antitumor efficacy studies with xenograft tumor modelsThe human HM7 colorectal xenograft tumor model was used
to assess antitumor efficacy of affinity variants of anti-DLL4(LM1 and HM6). When the average tumor size reached 150mm3 in beige nude female mice, animals were sorted intogroups consisting of 8 to 10 mice/group and administered LM1,HM6, or vehicle (phosphate buffered saline, PBS) via i.p.injection at a dose of 10 mg/kg twice per week. The antitumorefficacy of anti-DLL4 F(ab0)2 was assessed in the human SW620colon carcinoma xenograft model in athymic nude female miceand the human Calu-6 lung cancer xenograft model in beigenude female mice. When the average tumor size reached a targetsize of 100 mm3, animals were sorted into groups consisting of8 to 10 mice/group and treated intraperitoneally with anti-DLL4 F(ab0)2, anti-DLL4 IgG1, or vehicle (PBS). Anti-DLL4 F(ab0)2 was administered at various dose levels (cumulativedoses of 20, 40, or 80 mg/kg/week) using several dosing regi-mens (1 on/6 off, 2 on/5 off, or 3 on/4 off), over a duration of 3weeks. Anti-DLL4 IgG1 was administered at 10 mg/kg/week,once weekly for 3 weeks. Vehicle was also administered onceweekly for 3 weeks. In all xenograft studies, tumor volumes andbody weights were recorded at regular intervals, and tumorgrowth was quantitated by caliper measurements. Tumor vol-ume (mm3) was determined by measuring the length (l) andwidth (w) and calculating the volume (V ¼ lw2/2).
PK analysis of anti-DLL4 antibodies in mice, rats, andcynomolgus monkeys
Female athymic nude mice received a single i.v. dose of anti-DLL4 F(ab0)2 (10 or 50 mg/kg; n ¼ 12/group) or anti-DLL4 IgG1(20 mg/kg; n ¼ 15/group) via the tail vein. Blood samples fromthreemice per timepoint at various timepoints up to28dayswerecollected via retro-orbital bleeds (150 uL of blood collected onceper eye prior to terminal sac) and a terminal sample was collectedvia cardiac stick from each animal in each dose group. Compositeserum concentration–time profiles were constructed for PK anal-ysis. In multiple-dose toxicokinetic studies, Sprague Dawley ratsreceived anti-DLL4 F(ab0)2 (n¼ 6 per group) i.v. via the tail vein at3, 10, 30 or 100mg/kg (2 doses/week) for a total of 18 doses in 8weeks and anti-DLL4 IgG1 (n¼ 3per gender per group) at 1, 3, 10,or 30 mg/kg (1 dose/week) for a total of nine doses in 8 weeks.Blood samples were collected from each animal via the jugularvein. Composite serum concentration–time profiles were con-structed for PK analysis. Female cynomolgus monkeys (n¼ 5 pergroup) received multiple doses of anti-DLL4 F(ab0)2 i.v. via the
Translational Relevance
Several anti-DLL4 molecules are currently being investigat-ed in clinical trials for their potential as cancer therapeutics.Although selective inhibition of DLL4 apparently avoids someknown toxicities that have hampered the therapeutic applica-tion of Notch inhibition using gamma secretase inhibitors(GSI), safety concerns related to DLL4 inhibition have beenraised. Clinical trials using the DLL4-targeting antibody OMP-21M18 have also revealed safety concerns in humans, includ-ing grade III asymptomatic hypertension. This manuscriptexplores a novel strategy tomitigate toxicitywhilemaintainingtherapeutic activity in the context of a potent biologic pathwaywith a relatively narrow therapeutic index, which could haveimportant ramifications beyond developing therapeutics tar-geting DLL4. In addition, we report new safety findings thatfurther underscore the importance of exercising caution inclinical development of any therapeutics targeting this potentpathway.
Couch et al.
Clin Cancer Res; 22(6) March 15, 2016 Clinical Cancer Research1470
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

saphenous vein at 5, 15, or 50 mg/kg (1 dose/week) for a total ofninedoses in 8weeks andmultiple doses of anti-DLL4 IgG1 (n¼3per group) at 0.2, 0.8, 3, or 12 mg/kg (1 dose/week) for a total ofnine doses in 8 weeks. Blood samples were collected from eachanimal via the femoral vein and processed to collect serum. Meanserum concentration–time profiles were constructed for PKanalysis.
The following PK parameters were estimated using noncom-partmental analysis (WinNonlin, version 5.2.1; Pharsight Cor-poration) based on mouse serum concentration–time profiles:total drug exposure defined as area under the serum concen-tration–time curve extrapolated to infinity (AUCinf), total clear-ance (CL), and observed maximum serum concentration(Cmax). The following PK parameters were estimated usingnoncompartmental analysis (WinNonlin, version 5.2.1; Phar-sight Corporation) based on rat and monkey serum concen-tration–time profiles: weekly AUC defined as area underthe serum concentration–time curve extrapolated to Day 7(AUC0–7) and observed maximum serum concentration (Cmax).A na€�ve pooled approach was used in mice and rats to provideone estimate for each dose group; whereas in monkeys serumfrom each animal was analyzed separately and results for eachdose group were summarized as mean � SD. Anti-DLL4 F(ab0)2or anti-DLL4 IgG1 serum concentrations and antitherapeuticantibody (ATA) responses were assessed using standard ELISAs(see Supplemental methods).
Pilot assessment of anti-DLL4 F(ab0)2 toxicity in rat liverExperimentally na€�ve male Sprague Dawley Crl:CD(SD) rats
(n¼9/group)were given anti-DLL4 F(ab0)2 i.v. via the tail vein at atotal dose of 0 (PBS vehicle), 10, 30, or 100 mg/kg weekly for 8weeks (2 on/5 off dosing schedule). Males were chosen for pilotevaluations based on the higher incidence of neoplastic lesionsseen in males versus females following treatment with anti-DLL4IgG1 (23). Toxicity evaluationwas based on clinical observations,body weights, clinical pathology (clinical chemistry, hematology,and/or coagulation parameters), and macroscopic and micro-scopic pathology. Serum PK and ATA responses were assessed.Animals were euthanized onDay 59 and organswere collected formicroscopic evaluationby aboard-certified veterinary pathologist(J.C. Beyer).
Full evaluation of anti-DLL4 F(ab0)2 toxicity profile in rats andcynomolgus monkeys
Experimentally na€�ve SpragueDawleyCrl:CD(SD) rats (n¼ 15/sex/group) were given anti-DLL4 F(ab0)2 i.v. via the tail vein at 0(vehicle), 3, 10, 30, or 100 mg/kg/week (total dose level) on a 2on/5 off dosing schedule over 8 weeks (e.g., Days 1, 2, 8, 9, 15, 16,22, 23, 29, 30, 36, 37, 43, 44, 50, 51, 57, and 58; 18 total doses).Vehicle consisted of 20 mmol/L Na2 succinate, 240 mmol/Lsucrose, 0.02% polysorbate 20, at pH 5.5. Intermittent assess-ments of standard toxicologic parameters, including clinical con-dition, body weight and food consumption, clinical pathology,ophthalmic examinations (using an indirect ophthalmoscopeand a slit lamp biomicroscope), and neurologic examinations(24, 25) were conducted. Satellite animals (6/sex/group) wereincluded in the study for the purpose of assessing the kinetics ofanti-DLL4 F(ab0)2 exposure; these animals were observed clini-cally but no other toxicity assessments were conducted. Serum forassessment of ATAs was collected from all animals. Animals wereeuthanized on Day 59 (terminal necropsy) and 144 (recovery
necropsy) and organs were collected for microscopic evaluationby a board-certified veterinary pathologist (J.C. Beyer or G. Cain).
Experimentally na€�ve cynomolgus monkeys (3–5 years of age,Chinese origin; n¼ 5/sex/group)were given anti-DLL4 F(ab0)2 i.v.via the saphenous vein at dose levels of 0 (vehicle), 5, 15, or 50mg/kg once weekly (nine doses total). Vehicle consisted of 20mmol/L Na2 succinate, 240 mmol/L sucrose, 0.02% P20, at pH5.5. Periodic assessments of standard toxicologic parameters,including clinical condition, body weight, clinical pathology(clinical chemistry, hematology, and/or coagulation parameters),as well as physical, ophthalmic (using an indirect ophthalmo-scope and a slit lamp biomicroscope), and neurologic examina-tions (ref. 26; to assess general neurobehavioral condition andreflexes) were conducted. A subset of animals was implanted withtelemetry devices for cardiovascular monitoring; cardiovascularassessments in nontelemetered animals were performed usingexternal leads. Serum PK and ATA assays were performed toconfirm exposure to anti-DLL4 F(ab0)2 and assess the antibodyresponse. Animals were euthanized on Day 58 (terminal necrop-sy) andDay113 (recovery necropsy) andorganswere collected formicroscopic evaluationby aboard-certified veterinary pathologist(J.C. Beyer or G. Cain). Both the rat and monkey studies wereconducted at Covance Laboratories under Good Laboratory Prac-tices (GLP).
ResultsReducing DLL4 binding is not adequate to separate efficacyfrom toxicity in mice
We reasoned that tumor vasculature and normal liver vascu-lature might be differentially sensitive to DLL4 inhibition, suchthat partial DLL4 blockade could provide an opportunity toseparate antitumor efficacy from liver toxicity and improve thetherapeutic window. To this end, we explored a strategy ofreducing antibody binding affinity through alanine-directedmutagenesis in the antibodyCDRs to generate a panel of antibodyvariants with reduced binding to DLL4 on endothelial cellsrelative to parental antibody (Supplementary Fig. S1). Thereduced-binding antibodies were evaluated for antitumor efficacyin an HM7 colon cancer xenograft model and for induction ofliver toxicity in nude mice. The results of these in vivo studiesdefined two general classes of antibody variants, exemplified bycandidate molecules LM1 and HM6 (Fig. 1). Specifically, thereduced binding of LM1 relative to the parental antibody wasassociated with attenuated antitumor activity (Fig. 1A) withoutnotable attenuation of liver toxicity (Fig. 1B) after 3 weeks ofcontinued exposure. Compared with LM1, HM6 had even lowerbinding affinity and resulted in a complete loss of both antitumoractivity and liver toxicity. Reducing antibody binding thereforefailed to yield a molecule with the favorable properties of sus-tained antitumor activity combined with reduced liver toxicity.
Qualitative characterization of activity and pharmacokineticsof anti-DLL4 F(ab0)2
The anti-DLL4 F(ab0)2 antibody fragment retains bivalentCDRs identical to the parental antibody, and is expected to haveequivalent target binding capacity. To confirm that generation ofthe F(ab0)2 antibody fragment did not impact activity, we firsttested the in vitro potency of the F(ab0)2 relative to the parentalIgG1 in a three-dimensional angiogenic sprouting assay (27). Asanticipated, anti-DLL4 F(ab0)2 caused amarked increase in angio-genic sprouting that was comparable to that observed following
Effects of Pharmacokinetic Modulation of Anti-DLL4 Antibody
www.aacrjournals.org Clin Cancer Res; 22(6) March 15, 2016 1471
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

treatment with anti-DLL4 IgG1 (Supplementary Fig. S2A). Wealso tested the activity of anti-DLL4 F(ab0)2 in vivo by evaluatingearly postnatal mouse retina, which develops a stereotypic vas-cular pattern in a well-defined sequence (28–30). Consistent withthe in vitro results, anti-DLL4 F(ab0)2 caused excessive angiogenicsprouting in themouse retina thatwas qualitatively indistinguish-able in terms of vascular morphology from that observed afteranti-DLL4 IgG1 treatment (Supplementary Fig. S2B). Althoughthe in vivo activity of some antibodies may involve Fc-mediatedeffector function, these results, together with the subsequenttumor efficacy studies (detailed below), demonstrate that thebiologic activity of anti-DLL4 antibodies is independent of effec-tor function and therefore supports the potential utility of an F(ab0)2 antibody fragment to evaluate intermittent DLL4 pathway
blockade. The pharmacokinetics of the F(ab0)2 and IgG1 anti-bodies were evaluated in athymic nude mice following admin-istration of a single i.v. bolus dose. The exposure profile of bothmolecules was also evaluated in repeat-dose toxicity studies in ratand cynomolgus monkeys. In mice, at a dose of 10 mg/kg, anti-DLL4 F(ab0)2 had a clearance of 362 mL/day/kg compared with 8mL/day/kg for the full-length IgG1 (Supplementary Table S1;ref. 31). Similarly, in rats and cynomolgus monkeys, anti-DLL4F(ab0)2 was cleared much more rapidly relative to the full-lengthIgG1 (Fig. 2, Supplementary Fig. S3; see Supplementary Table S2for a full listing of PK parameters). These results confirmed thatthe anti-DLL4 F(ab0)2 molecule was an appropriate tool to assessthe impact of intermittent DLL4 pathway inhibition on bothefficacy and toxicity.
Figure 2.PK profiles of anti-DLL4 IgG1 andF(ab0)2 in mouse and monkey.A, anti-DLL4 IgG1 serumconcentration–time data in athymicnudemice; B, anti-DLL4 F(ab0)2 serumconcentration–time data in athymicnude mice; C, anti-DLL4 IgG1 averageserum concentration–time data incynomolgus monkeys; D, anti-DLL4 F(ab0)2 average serum concentration–time data in cynomolgus monkeys.
Figure 1.Reducing binding affinity of anti-DLL4 IgG1 does not separate antitumor efficacy and liver toxicity in mice. A, robust antitumor efficacy was observed in ahuman HM7 xenograft model with anti-DLL4 IgG1 (YW152F), whereas lower affinity variants showed attenuated (LM1) or no efficacy (HM6.1) when administeredtwice per week i.p at 10 mg/kg. Tumor volumes are presented as mean � SEM (n ¼ 8–10/group). Two-tailed unpaired t test was used to calculate P valuesat Day 11. B, mice given anti-DLL4 IgG1 for 3 weeks at 30 mg/kg/week i.p. had marked sinusoidal dilation of the liver, as shown by hematoxylin and eosin (H&E)staining. Lower affinity variants showed either a similar severity of liver pathology (LM1) relative to the IgG1 molecule, or appeared histologically normal andsimilar to control-treated animals (HM6) following an equivalent dosing regimen. Scale bar ¼ 500 mm.
Couch et al.
Clin Cancer Res; 22(6) March 15, 2016 Clinical Cancer Research1472
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380
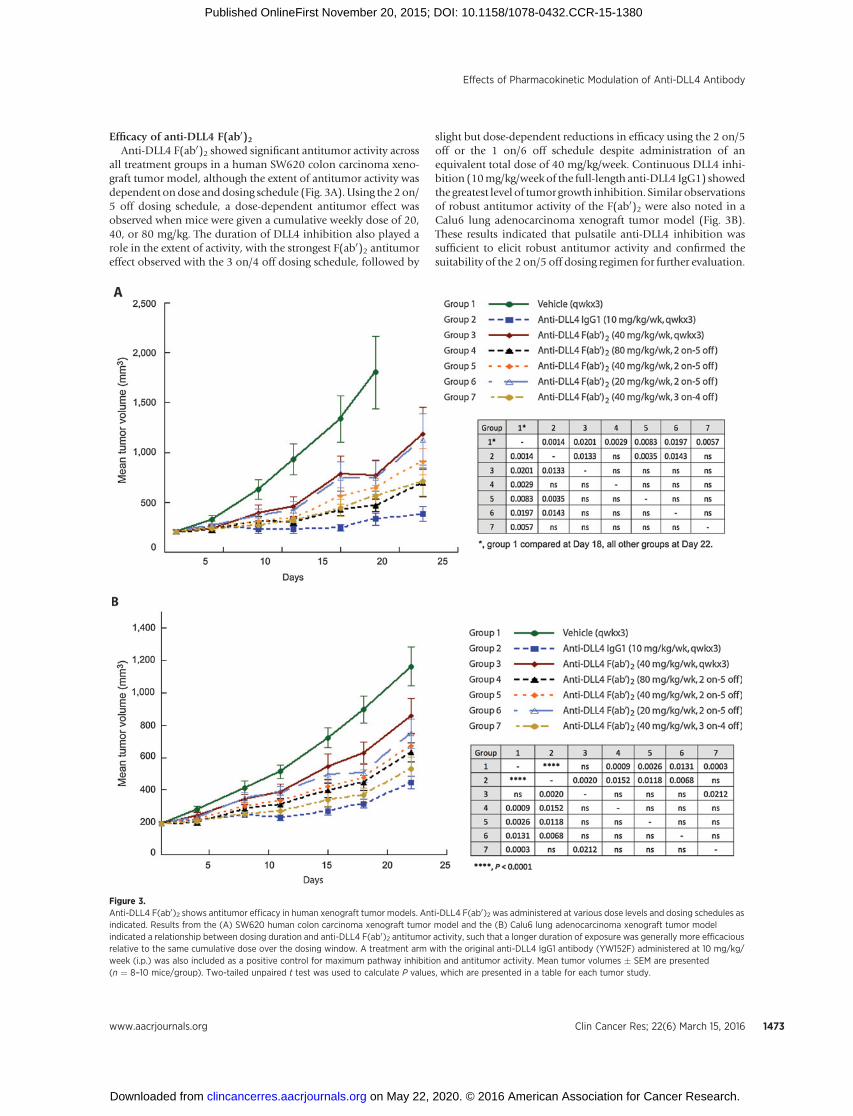
Efficacy of anti-DLL4 F(ab0)2Anti-DLL4 F(ab0)2 showed significant antitumor activity across
all treatment groups in a human SW620 colon carcinoma xeno-graft tumor model, although the extent of antitumor activity wasdependent on dose and dosing schedule (Fig. 3A). Using the 2 on/5 off dosing schedule, a dose-dependent antitumor effect wasobserved when mice were given a cumulative weekly dose of 20,40, or 80 mg/kg. The duration of DLL4 inhibition also played arole in the extent of activity, with the strongest F(ab0)2 antitumoreffect observed with the 3 on/4 off dosing schedule, followed by
slight but dose-dependent reductions in efficacy using the 2 on/5off or the 1 on/6 off schedule despite administration of anequivalent total dose of 40 mg/kg/week. Continuous DLL4 inhi-bition (10mg/kg/week of the full-length anti-DLL4 IgG1) showedthe greatest level of tumor growth inhibition. Similar observationsof robust antitumor activity of the F(ab0)2 were also noted in aCalu6 lung adenocarcinoma xenograft tumor model (Fig. 3B).These results indicated that pulsatile anti-DLL4 inhibition wassufficient to elicit robust antitumor activity and confirmed thesuitability of the 2 on/5 off dosing regimen for further evaluation.
Figure 3.Anti-DLL4 F(ab0)2 shows antitumor efficacy in human xenograft tumor models. Anti-DLL4 F(ab0)2 was administered at various dose levels and dosing schedules asindicated. Results from the (A) SW620 human colon carcinoma xenograft tumor model and the (B) Calu6 lung adenocarcinoma xenograft tumor modelindicated a relationship between dosing duration and anti-DLL4 F(ab0)2 antitumor activity, such that a longer duration of exposure was generally more efficaciousrelative to the same cumulative dose over the dosing window. A treatment arm with the original anti-DLL4 IgG1 antibody (YW152F) administered at 10 mg/kg/week (i.p.) was also included as a positive control for maximum pathway inhibition and antitumor activity. Mean tumor volumes � SEM are presented(n ¼ 8–10 mice/group). Two-tailed unpaired t test was used to calculate P values, which are presented in a table for each tumor study.
Effects of Pharmacokinetic Modulation of Anti-DLL4 Antibody
www.aacrjournals.org Clin Cancer Res; 22(6) March 15, 2016 1473
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

Mitigating toxicity with anti-DLL4 F(ab0)2 in mice and ratsTo address whether intermittent blockade of theDLL4 pathway
might ameliorate the toxicity associatedwith continuous pathwayinhibition, we first evaluated the effects of anti-DLL4 F(ab0)2 onDLL4 pathway-related liver gene expression and histopathologyin mice. Marked upregulation of DLL4 blockade-related geneexpression occurred in liver, as previously reported by Yan andcolleagues (ref. 23; Supplementary Fig. S4A and S4B). Despite asimilar cumulative weekly dose of anti-DLL4 F(ab0)2, significantattenuation of DLL4 pathway-related gene expression occurred inmice treatedon an intermittent dosing schedule (100mg/kg/weekgivenon the2on/5off schedule). Liver gene expression levels afteran 18-day recovery period were comparable between control andanti-DLL4 F(ab0)2-treated mice regardless of dosing regimen;however, gene expression increases were sustained in anti-DLL4IgG1-treated mice likely due to slower systemic clearance of theIgG1 antibody compared with the F(ab0)2 (Supplementary TableS1). The nature and severity of hepatic histopathological changescorresponded with altered gene expression in mouse livers for alltreatment groups at terminal and recovery time points (Supple-mentary Fig. S4C). Intermittent dosing of anti-DLL4 F(ab0)2mitigated the effect of DLL4 inhibition in the mouse liver.Furthermore, changes in liver gene expression and microscopicfindings were reversible after antibody clearance.
To extend and confirm findings indicating schedule-depen-dent amelioration of liver toxicity by anti-DLL4 F(ab0)2 admin-istration in the mouse, two follow-up pilot toxicity studies wereconducted in rats. Studies in rats were important for toxicityscreening because rats showed the highest sensitivity to anti-DLL4 IgG1-mediated liver histopathological changes among allnonclinical species evaluated (mouse, rat, and monkey), andalso demonstrated the potential for dose-dependent vascularneoplasia after treatment with anti-DLL4 IgG1 (23). In bothstudies, male Sprague Dawley rats were dosed i.v. with anti-DLL4 F(ab0)2 at 0, 10, 30, or 100 mg/kg weekly (2 on/5 offschedule) for 8 weeks (18 doses; n ¼ 9/group); the secondconfirmatory study also included an 8 week treatment-freerecovery period (n ¼ 9/group for both dosing and recoveryperiods) to evaluate the reversibility of any potential anti-DLL4F(ab0)2-induced lesions. These studies were conducted only inmales because vascular neoplasms following IgG1 treatmentwere observed only in males, although liver toxicity did notshow a difference between sexes. Representative results fromthe second F(ab0)2 pilot study are shown in Fig. 4B and D, as thetoxicity and exposure profiles of anti-DLL4 F(ab0)2-repeatedresults from the first pilot study. For comparison, the IgG1 ratstudy results (Fig. 4A and C) are shown with pooled data frommales and females, as there were no differences between sexesfor liver parameters. Similar to the mouse, the clearance of anti-DLL4 F(ab0)2 in rats was much faster than the full-length IgG1(Supplementary Table S2; Supplementary Fig. S3).
As anticipated based on previous experience with anti-DLL4IgG1, rats treated with anti-DLL4 F(ab0)2 at 30 and 100 mg/kg/week (i.e., 15 or 50 mg/kg/day on the 2 on/5 off schedule) hadchanges in serum chemistry parameters after 8 weeks of dosing,including minimal to mild increases in serum ALT (Fig. 4B),aspartate aminotransferase, alkaline phosphatase (ALP), totalbilirubin, total bile acids, and cholesterol. Corresponding histo-logic changes in the liver included sinusoidal dilation at all doselevels, as well as hepatocellular necrosis, mixed leukocyte inflam-mation, and bile duct hyperplasia (Fig. 4D). Notably, the severity
and incidence of hepatotoxicity (clinical and anatomic patholo-gy) was reduced relative to that observed with anti-DLL4 IgG1 atequivalent exposure levels (i.e., 3 mg/kg/week IgG1 vs. 10mg/kg/week F(ab0)2; or 30 mg/kg/week IgG1 vs. 100 mg/kg/week F(ab0)2), indicative of a shift in the overall toxicity dose–response(Fig. 4B–D; Supplementary Table S2). Evidence of reversibility ofF(ab0)2-associated clinical and anatomic pathology changes(sinusoidal dilation/hepatocyte loss) was present, with the excep-tion of nonreversible biliary hyperplasia and fibrosis at a dose of100 mg/kg/week.
Importantly, cutaneous vascular neoplasia was not present inrats at any F(ab0)2 dose level, further differentiating the F(ab0)2toxicity profile from that of the IgG1 molecule in which lesionswere present at doses as low as 3 mg/kg/week and increased inincidence at higher dose levels (Supplementary Table S3).Together, these results supported the potential for an improvedtherapeutic window and reduced toxicity profile with anti-DLL4 F(ab0)2 relative to anti-DLL4 IgG1, enabling a pathforward for additional characterization of the anti-DLL4 F(ab0)2molecule.
Novel toxicity findings associated with DLL4 pathwayinhibition
To extend the observations in mice and rats suggesting thepotential for an improved toxicity profile for anti-DLL4 F(ab0)2and to support future clinical studies, definitive (GLP) toxicologystudies were conducted in rats and cynomolgus monkeys. Ratswere dosed i.v. with anti-DLL4 F(ab0)2 at 0, 3, 10, 30, or 100mg/kg/week for 8 weeks, using the same 2 on/5 off dosingschedule as in the mouse efficacy and pilot rat toxicity studies.To account for distinct PK exposure profiles between species (Fig.2; Supplementary Table S2), monkeys were dosed once weeklyfor 8weekswith anti-DLL4F(ab0)2 i.v. at 0, 10, 30, and100mg/kg/week. Both studies included an 8-week treatment-free recoveryperiod to evaluate the reversibility of any drug-induced findings.Additionally, the dosing regimens were designed to producesystemic drug levels in both species that overlapped with expo-sures previously obtained from the anti-DLL4 IgG1 molecule,thereby enabling a comparison of the relative toxicity profilesbetween these two molecules (see Fig. 4).
Consistent with effects observed in the rat pilot toxicity studies,shifts in the dose–toxicity relationship of the F(ab0)2 moleculerelative to the IgG1 molecule were observed in both rats andcynomolgus monkeys. Higher dose levels of the F(ab0)2 werenecessary to elicit the same incidence and severity of IgG1-medi-ated changes, despite approximately equivalent AUC (Fig. 5A andB) and equivalent or higher Cmax (Supplementary Table S2).Together with the results of the rat pilot toxicity studies (Fig. 4;Supplementary Table S2), these observations suggest that main-tenance of anti-DLL4 concentrations over a threshold concentra-tion may be the driver of toxicity, because the faster clearance of F(ab0)2 allows the serum concentrations to drop to a much lowerlevel within the dosing period compared with the full-lengthIgG1. For example, the incidence and severity of anemia inmonkeys were decreased with the F(ab0)2 relative to the IgG1molecule (Fig. 5). Observations of an attenuated toxicity profilefor the F(ab0)2 relative to the full-length IgG1 were consistentacross species (mouse, rat, and cynomolgusmonkey) and indicatethat intermittent inhibition of DLL4 mitigates toxicities associat-ed with continuous DLL4 blockade, likely due to partial recoveryof pathway inhibition between dose administrations.
Couch et al.
Clin Cancer Res; 22(6) March 15, 2016 Clinical Cancer Research1474
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

Figure 4.Reduced toxicity of anti-DLL4 F(ab0)2 relative to IgG1 in rat liver. Comparison of liver enzyme changes and incidence of liver lesions revealed anamelioration of toxicity when dosing with anti-DLL4 F(ab0)2 relative to the IgG1 at comparable exposures (AUC0–7). A and B, liver enzyme (ALT) profiles ofindividual rats are shown over an 8-week dosing period with anti-DLL4 IgG1 (A) or anti-DLL4 F(ab0)2 (B); bold lines, the group mean with points at theactual measurement times. For both the IgG1 (n ¼ 15/sex/group) and F(ab0)2 (n ¼ 18 males/group) datasets, dosing and recovery animals werepooled across the dosing interval; for the IgG1, data were also pooled across sex as there was no apparent sex difference in liver toxicity. Only males wereused in the F(ab0)2 study. Scaling of the y-axis is on a log2 scale so that each axis tick represents a doubling of the ALT values, with labeling on the naturalscale to facilitate interpretation. C and D, the frequency and severity of major histologic findings in the liver was dose-dependent for both anti-DLL4 F(ab0)2and IgG1 following 8 weeks of IgG1 administration. However, relative to the IgG1 liver toxicity profile (n ¼ 10/sex/group) (C), dosing F(ab0)2 on a 2 on/5off dosing schedule (n ¼ 9 males/group) revealed amelioration of severe liver pathologies (BDH, fibrosis, necrosis) (D). SNDIL, sinusoidal dilation;BDH, bile duct hyperplasia.
Effects of Pharmacokinetic Modulation of Anti-DLL4 Antibody
www.aacrjournals.org Clin Cancer Res; 22(6) March 15, 2016 1475
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380
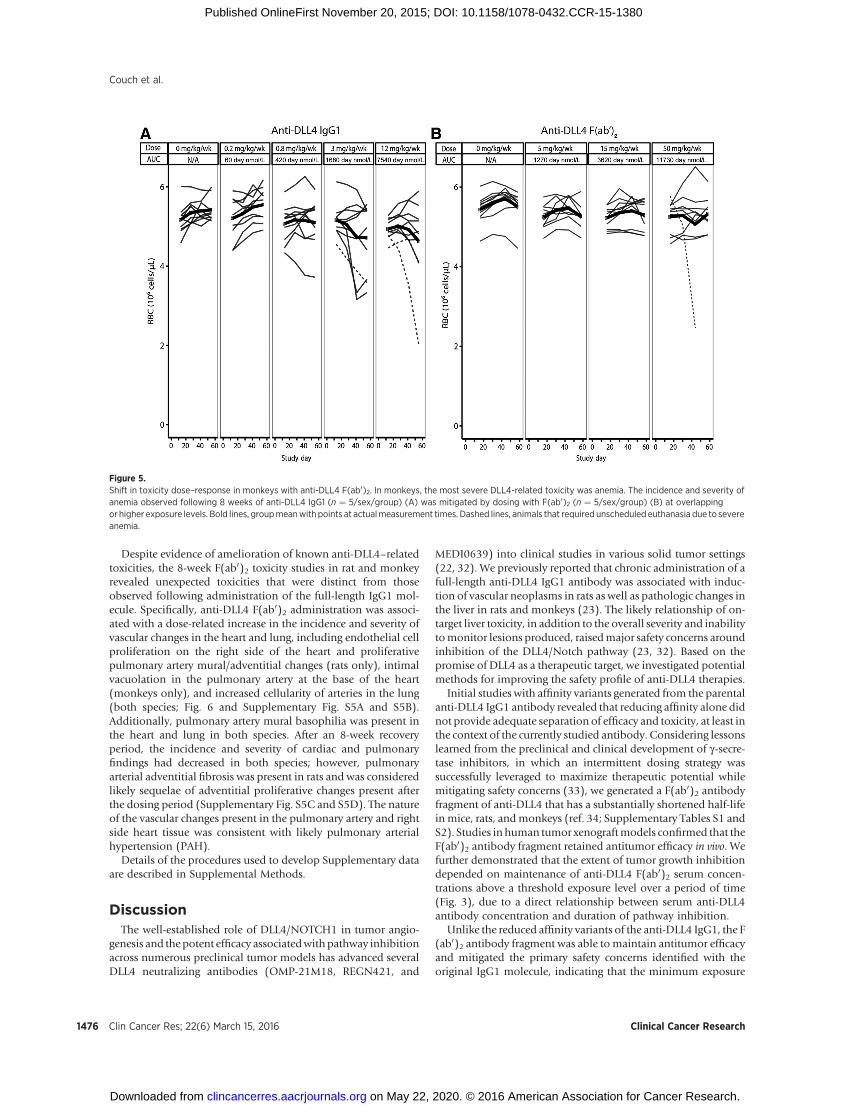
Despite evidence of amelioration of known anti-DLL4–relatedtoxicities, the 8-week F(ab0)2 toxicity studies in rat and monkeyrevealed unexpected toxicities that were distinct from thoseobserved following administration of the full-length IgG1 mol-ecule. Specifically, anti-DLL4 F(ab0)2 administration was associ-ated with a dose-related increase in the incidence and severity ofvascular changes in the heart and lung, including endothelial cellproliferation on the right side of the heart and proliferativepulmonary artery mural/adventitial changes (rats only), intimalvacuolation in the pulmonary artery at the base of the heart(monkeys only), and increased cellularity of arteries in the lung(both species; Fig. 6 and Supplementary Fig. S5A and S5B).Additionally, pulmonary artery mural basophilia was present inthe heart and lung in both species. After an 8-week recoveryperiod, the incidence and severity of cardiac and pulmonaryfindings had decreased in both species; however, pulmonaryarterial adventitial fibrosis was present in rats and was consideredlikely sequelae of adventitial proliferative changes present afterthe dosing period (Supplementary Fig. S5C and S5D). The natureof the vascular changes present in the pulmonary artery and rightside heart tissue was consistent with likely pulmonary arterialhypertension (PAH).
Details of the procedures used to develop Supplementary dataare described in Supplemental Methods.
DiscussionThe well-established role of DLL4/NOTCH1 in tumor angio-
genesis and the potent efficacy associatedwith pathway inhibitionacross numerous preclinical tumor models has advanced severalDLL4 neutralizing antibodies (OMP-21M18, REGN421, and
MEDI0639) into clinical studies in various solid tumor settings(22, 32). We previously reported that chronic administration of afull-length anti-DLL4 IgG1 antibody was associated with induc-tion of vascular neoplasms in rats as well as pathologic changes inthe liver in rats and monkeys (23). The likely relationship of on-target liver toxicity, in addition to the overall severity and inabilitytomonitor lesions produced, raisedmajor safety concerns aroundinhibition of the DLL4/Notch pathway (23, 32). Based on thepromise of DLL4 as a therapeutic target, we investigated potentialmethods for improving the safety profile of anti-DLL4 therapies.
Initial studies with affinity variants generated from the parentalanti-DLL4 IgG1 antibody revealed that reducing affinity alone didnot provide adequate separation of efficacy and toxicity, at least inthe context of the currently studied antibody. Considering lessonslearned from the preclinical and clinical development of g-secre-tase inhibitors, in which an intermittent dosing strategy wassuccessfully leveraged to maximize therapeutic potential whilemitigating safety concerns (33), we generated a F(ab0)2 antibodyfragment of anti-DLL4 that has a substantially shortened half-lifein mice, rats, andmonkeys (ref. 34; Supplementary Tables S1 andS2). Studies in human tumor xenograftmodels confirmed that theF(ab0)2 antibody fragment retained antitumor efficacy in vivo. Wefurther demonstrated that the extent of tumor growth inhibitiondepended on maintenance of anti-DLL4 F(ab0)2 serum concen-trations above a threshold exposure level over a period of time(Fig. 3), due to a direct relationship between serum anti-DLL4antibody concentration and duration of pathway inhibition.
Unlike the reduced affinity variants of the anti-DLL4 IgG1, the F(ab0)2 antibody fragment was able tomaintain antitumor efficacyand mitigated the primary safety concerns identified with theoriginal IgG1 molecule, indicating that the minimum exposure
Figure 5.Shift in toxicity dose–response in monkeys with anti-DLL4 F(ab0)2. In monkeys, the most severe DLL4-related toxicity was anemia. The incidence and severity ofanemia observed following 8 weeks of anti-DLL4 IgG1 (n ¼ 5/sex/group) (A) was mitigated by dosing with F(ab0)2 (n ¼ 5/sex/group) (B) at overlappingor higher exposure levels. Bold lines, groupmeanwith points at actualmeasurement times. Dashed lines, animals that required unscheduled euthanasia due to severeanemia.
Couch et al.
Clin Cancer Res; 22(6) March 15, 2016 Clinical Cancer Research1476
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

duration required for antitumor activity was less than thatrequired to elicit toxicity. Specifically, the F(ab0)2 moleculeappeared to result in a shift in the toxicity–exposure relationshiprelative to the IgG1 molecule, as evidenced by a reduction in theincidence and severity of serum liver enzyme elevations and liverhistopathology findings in rats and monkeys (Fig. 4), a reductionin the incidence and severity of anemia in monkeys (Fig. 5), aswell as a complete mitigation of vascular neoplasms in rats(Supplementary Table S3). These results indicate that alterationsin the exposure profile of an antibody (e.g., by using a F(ab0)2)may be a valuable strategy to mitigate toxicity while maintainingtherapeutic activity in the context of a potent biologic pathwaywith a relatively narrow therapeutic index.
Despite the mitigation of the well-known adverse effects, weobserved novel toxicities after repeated administration of anti-DLL4 F(ab0)2 over 8 weeks including right heart endothelial cellproliferative changes and proliferative pulmonary artery mural/
adventitial changes (rats only), intimal pulmonary artery vacuo-lation (monkeys only), and pulmonary artery basophilia in theheart and lung (both species). These right heart and pulmonaryarterial vascular changes are consistent with the histologic sequel-ae of drug-induced PAH (refs. 35, 36; Fig. 6); notably, thesefindings were not present after administration of the full-lengthanti-DLL4 IgG1. The cardiac andpulmonary vascular changes thatoccurred after administering the F(ab0)2 antibody fragment fur-ther underscore the importance of context-dependent outcomesassociated with perturbation of DLL4/NOTCH1 signaling withindifferent tissues.
Findings accumulated from our experience with anti-DLL4antibodies indicate that inhibition of the DLL4 pathway is broad-ly associated with adverse effects in at least two major organsystems: the liver and cardiopulmonary systems, specifically, theright heart and pulmonary arterial vasculature. Histopathologicalchanges in the heart and lung were not accompanied by overt
Figure 6.Novel toxicities identified with anti-DLL4 F(ab0)2 in rats and monkeys.A, representative H&E sections ofmonkey lung tissues show pulmonaryartery changes in an anti-DLL4 F(ab0)2-treated monkey (50 mg/kg/week); arrowhead, vacuolated arterialintima (endothelial) layer; arrow,basophilia of arterial muscular layer.B and C, representative H&E sectionsof rat tissues showing anti-DLL4 F(ab0)2-related changes in the lung (B)and heart (C). B, left panel, normalpulmonary artery in a vehicle-treatedrat; small arrow, normal arterial intima;large arrow, normal arterialmuscularis;arrowhead, normal arterial adventitiallayer. Right panel, pulmonary arterychanges in a rat administered anti-DLL4 F(ab0)2 at 50mg/kg/week, 2 on/5 off schedule. Small arrow, intimalayer (endothelium); large arrow,basophilia of arterial muscularis layer;arrowhead, basophilia of arterialadventitial layer. C, left panel, normalright atrium in a vehicle-treated rat.Right panel, interstitial hypercellularitydue to endothelial cell proliferation(arrows) in the right atrium of an anti-DLL4 F(ab0)2-treated rat (50 mg/kg/week; 2 on/5 off schedule).
Effects of Pharmacokinetic Modulation of Anti-DLL4 Antibody
www.aacrjournals.org Clin Cancer Res; 22(6) March 15, 2016 1477
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

changes in heart rate or blood pressure in telemetry-instrumentedmonkeys, suggesting that these changes are likely to be poorlymonitorable in the clinic. Independent preclinical and clinicalinvestigations using other anti-DLL4 antibodies indicate thatthe constellation of changes in the liver, skin, and cardiopul-monary systems may represent a class-effect of DLL4 inhibition.For instance, with respect to changes in the cardiopulmonarysystem, an anti-DLL4 antibody (MEDI0639) also evaluated inmonkeys was associated with pathologic changes in the heartand lung, as indicated by elevations in blood pressure, heartrate, and C-reactive protein, and observations of heart failure(37). In humans, another anti-DLL4 antibody (OMP-21M18)has been associated with grade III asymptomatic hypertensionin approximately 28% of phase I patients (22). In addition, ananti-DLL4 antibody (REGN421) reported hypertension as wellas grade II and III pulmonary hypertension in 14% of patientsin a phase I trial (38). Therefore, the totality of the effectsobserved over a range of anti-DLL4 neutralizing antibodiesindicates a liability of DLL4 inhibition in the cardiopulmonarysystem that may translate to a broad safety risk in humans. Ofparticular concern, our data indicate clear species differences insensitivity for particular DLL4-mediated toxicities (e.g., ratsappear most sensitive to effects on liver, whereas monkeys aremost sensitive to effects on RBC parameters). This species-specific heterogeneity of response to DLL4/NOTCH1 pathwayinhibition complicates prediction of the most likely or severesafety concerns in humans.
Taken together, by utilizing a F(ab0)2 antibody fragment againstDLL4 we have demonstrated a unique approach to antibody drugdevelopment that may prove advantageous for widening orimproving a narrow therapeutic index, particularly if toxicitiesappear related to chronic pathway inhibition. Importantly, ourdata provide further evidence that the DLL4 pathway is extremelysensitive topharmacologic perturbation and that toxicitiesmaybedifficult to accuratelymodel in preclinical species and/ormonitor
in the clinic. Therefore, cautionmust be exercised to safely harnessthis potent pathway to treat cancer.
Disclosure of Potential Conflicts of InterestA.V. Kamath has ownership interest (including patents) in Roche. No
potential conflicts of interest were disclosed by the other authors.
Authors' ContributionsConception and design: J.A. Couch, G. Zhang, J.C. Beyer, C.L.Z. de Zafra,P. Gupta, A.V. Kamath, J. Tarrant, K.P. Allamneni, G. Cain, M. YanDevelopment of methodology:G. Zhang, J.C. Beyer, P. Gupta, K.P. Allamneni,G. Cain, S.P. Tsai, J. Ruppel, J.B. Ridgway, P.E. HassAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): G. Zhang, J.C. Beyer, P. Gupta, K.P. Allamneni,G. Cain, S. Yee, J. Ruppel, J.B. Ridgway, P.E. HassAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): J.A. Couch, G. Zhang, J.C. Beyer, C.L.Z. de Zafra,P. Gupta, A.V. Kamath, N. Lewin-Koh, J. Tarrant, G. Cain, S. Yee, S. Ross,S.P. Tsai, J. Ruppel, M. YanWriting, review, and/or revision of the manuscript: J.A. Couch, J.C. Beyer,C.L.Z. de Zafra, P. Gupta, A.V. Kamath,N. Lewin-Koh, J. Tarrant, K.P. Allamneni,G. Cain, S. Ross, R. Cook, J. Ruppel, M. YanAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): J.A. Couch, G. Zhang, J.C. Beyer, C.L.Z. de Zafra,P. Gupta, M. Paluch, J. FranklinStudy supervision: J.C. Beyer, C.L.Z. de Zafra, K.P. AllamneniOther (assay support (development, data generation, raw data processing)):R. Cook
AcknowledgmentsThe authors thank Lisa Damico-Beyer, Cynthia Wong, Ina Rhee, Weilan Ye,
Lu Xu, Join Castro, Khiem Tran, Sock-Cheng Lewin-Koh, and Noel Dybdal fortheir critical contributions to this manuscript.
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received June15, 2015; revisedOctober 23, 2015; acceptedOctober 29, 2015;published OnlineFirst November 20, 2015.
References1. Ferrara N, Kerbel RS. Angiogenesis as a therapeutic target. Nature 2005;
438:967–74.2. Hurwitz H, Fehrenbacher L, NovotnyW, Cartwright T, Hainsworth J, Heim
W, et al. Bevacizumab plus irinotecan, fluorouracil, and leucovorin formetastatic colorectal cancer. N Engl J Med 2004;350:2335–42.
3. Laskin JJ, Sandler AB. First-line treatment for advanced non-small-cell lung cancer. Oncology (Williston Park) 2005;19:1671–6; discussion8–80.
4. Gale NW, Dominguez MG, Noguera I, Pan L, Hughes V, Valenzuela DM,et al. Haploinsufficiency of delta-like 4 ligand results in embryonic lethalitydue to major defects in arterial and vascular development. Proc Natl AcadSci U S A 2004;101:15949–54.
5. Mailhos C, Modlich U, Lewis J, Harris A, Bicknell R, Ish-Horowicz D.Delta4, an endothelial specific notch ligand expressed at sites of physio-logical and tumor angiogenesis. Differentiation 2001;69:135–44.
6. Rao PK, Dorsch M, Chickering T, Zheng G, Jiang C, Goodearl A, et al.Isolation and characterization of the notch ligand delta4. Exp Cell Res2000;260:379–86.
7. Shutter JR, Scully S, FanW, Richards WG, Kitajewski J, Deblandre GA, et al.Dll4, a novel Notch ligand expressed in arterial endothelium. Genes Dev2000;14:1313–8.
8. Duarte A, Hirashima M, Benedito R, Trindade A, Diniz P, Bekman E, et al.Dosage-sensitive requirement for mouse Dll4 in artery development.Genes Dev 2004;18:2474–8.
9. Krebs LT, Shutter JR, Tanigaki K, Honjo T, Stark KL, Gridley T. Haploin-sufficient lethality and formation of arteriovenousmalformations inNotchpathway mutants. Genes Dev 2004;18:2469–73.
10. Jakobsson L, Franco CA, Bentley K, Collins RT, Ponsioen B, Aspalter IM,et al. Endothelial cells dynamically compete for the tip cell position duringangiogenic sprouting. Nat Cell Biol 2010;12:943–53.
11. Thurston G, Noguera-Troise I, Lobov IB, Daly C, Rudge JS, Gale NW, et al.Delta-like Ligand 4/Notch pathway in tumor angiogenesis. In:Figg WD,Folkman J, editors. Angiogenesis: an integrative approach from science tomedicine. New York: Springer; 2008. p. 217–24.
12. Li JL, Harris AL. Crosstalk of VEGF and Notch pathways in tumourangiogenesis: therapeutic implications. Front Biosci (Landmark Ed)2009;14:3094–110.
13. Yan M, Plowman GD. Delta-like 4/Notch signaling and its therapeuticimplications. Clin Cancer Res 2007;13:7243–6.
14. Hoey T, YenWC,Axelrod F, Basi J, Donigian L,Dylla S, et al. DLL4 blockadeinhibits tumor growth and reduces tumor-initiating cell frequency. CellStem Cell 2009;5:168–77.
15. Noguera-Troise I,DalyC, PapadopoulosNJ, Coetzee S, BolandP,GaleNW,et al. Blockade of Dll4 inhibits tumour growth by promoting non-pro-ductive angiogenesis. Nature 2006;444:1032–7.
16. Ridgway J, Zhang G, Wu Y, Stawicki S, Liang WC, Chanthery Y, et al.Inhibition of Dll4 signalling inhibits tumour growth by deregulatingangiogenesis. Nature 2006;444:1083–7.
17. Scehnet JS, Jiang W, Kumar SR, Krasnoperov V, Trindade A, Benedito R,et al. Inhibition of Dll4-mediated signaling induces proliferationof immature vessels and results in poor tissue perfusion. Blood 2007;109:4753–60.
18. Yen WC, Fischer MM, Hynes M, Wu J, Kim E, Beviglia L, et al. Anti-DLL4has broad spectrum activity in pancreatic cancer dependent on targeting
Clin Cancer Res; 22(6) March 15, 2016 Clinical Cancer Research1478
Couch et al.
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

DLL4-Notch signaling in both tumor and vasculature cells. Clin Cancer Res2012;18:5374–86.
19. Pries AR, Hopfner M, le Noble F, Dewhirst MW, Secomb TW. The shuntproblem: control of functional shunting in normal and tumour vascula-ture. Nat Rev Cancer 2010;10:587–93.
20. Yan M. Therapeutic promise and challenges of targeting DLL4/NOTCH1.Vasc Cell 2011;3:17.
21. FischerM, YenWC, Kapoun AM,WangM,O'YoungG, Lewicki J, et al. Anti-DLL4 inhibits growth and reduces tumor-initiating cell frequency incolorectal tumors with oncogenic KRAS mutations. Cancer Res 2011;71:1520–5.
22. Smith DC, Eisenberg P, Stagg R, Manikhas G, Pavlovskiy A, Sikic B, et al. Afirst-in-human, phase I trial of the anti-DLL4 antibody (OMP-21M18)targeting cancer stem cells in patients with advanced solid tumors. 2ndEORTC-NCI-AACR symposium onmolecular targets and cancer therapeu-tics; 2010; Berlin, Germany; 2010.
23. Yan M, Callahan CA, Beyer JC, Allamneni KP, Zhang G, Ridgway JB,et al. Chronic DLL4 blockade induces vascular neoplasms. Nature2010;463:E6–7.
24. Moser VC. Approaches for assessing the validity of a functional observa-tional battery. Neurotoxicol Teratol 1990;12:483–8.
25. Tilson HA, Moser VC. Comparison of screening approaches. Neurotox-icology 1992;13:1–13.
26. Kallman MJ. Central nervous system assessment in nonhuman primates:behavioral assessment, neurological (electroencephalography, nerve con-duction), abuse liability, and cognitive assessment. Amsterdam, Nether-lands: Elsevier/Academic Press; 2015.
27. Nakatsu MN, Sainson RC, Aoto JN, Taylor KL, Aitkenhead M, Perez-del-Pulgar S, et al. Angiogenic sprouting and capillary lumen formationmodeled by human umbilical vein endothelial cells (HUVEC) in fibringels: the role of fibroblasts and Angiopoietin-1. Microvasc Res 2003;66:102–12.
28. Fruttiger M. Development of the mouse retinal vasculature: angio-genesis versus vasculogenesis. Invest Ophthalmol Vis Sci 2002;43:522–7.
29. Gerhardt H, Golding M, Fruttiger M, Ruhrberg C, Lundkvist A, AbramssonA, et al. VEGF guides angiogenic sprouting utilizing endothelial tip cellfilopodia. J Cell Biol 2003;161:1163–77.
30. Stone J, Dreher Z. Relationship between astrocytes, ganglion cells andvasculature of the retina. J Comp Neurol 1987;255:35–49.
31. Kamath AV, Yip V, Gupta P, Boswell CA, Bumbaca D, Haughney P, et al.Dose dependent pharmacokinetics, tissue distribution, and anti-tumorefficacy of a humanizedmonoclonal antibody against DLL4 inmice. MAbs2014;6:1631–7.
32. Tarrant JM, Lewin-Koh SC, Lewin-Koh N. Development of a safety bio-marker signature to detect hepatic sinusoidal dilation associated with ananti-DLL4 biotherapeutic. Biomarkers 2014;19:391–401.
33. Wei P, Walls M, QiuM, Ding R, Denlinger RH,Wong A, et al. Evaluation ofselective gamma-secretase inhibitor PF-03084014 for its antitumor efficacyand gastrointestinal safety to guide optimal clinical trial design. MolCancer Ther 2010;9:1618–28.
34. Covell DG, Barbet J, Holton OD, Black CD, Parker RJ, Weinstein JN.Pharmacokinetics of monoclonal immunoglobulin G1, F(ab0)2, and Fab0
in mice. Cancer Res 1986;46:3969–78.35. Cho YJ, Han JY, Lee SG, Jeon BT, Choi WS, Hwang YS, et al. Temporal
changes of angiopoietins and Tie2 expression in rat lungs after monocrota-line-induced pulmonary hypertension. Comp Med 2009;59:350–6.
36. StenmarkKR,Meyrick B,GalieN,MooiWJ,McMurtry IF. Animalmodels ofpulmonary arterial hypertension: the hope for etiological discovery andpharmacological cure. Am J Physiol Lung Cell Mol Physiol 2009;297:L1013–32.
37. Ryan P, Huang J, Bao H, Cho S, Brohawn P, Burke P. Nonclinical safetyevaluation of MEDI0639 (anti-DLL4Mab) to support first time In human:linking DLL4-notch signaling blockade to exaggerated pharmacologyeffects in cynomolgus monkeys. AACR 104th annual meeting; 2013;Washington, DC: AACR; 2013.
38. Chiorean EG, LoRusso P, Strother RM, Diamond JR, Younger A, Messer-smithWA, et al. A phase Ifirst-in-human study of enoticumab (REGN421),a fully human delta-like ligand 4 (Dll4) monoclonal antibody in patientswith advanced solid tumors. Clin Cancer Res 2015;21:2695–703.
www.aacrjournals.org Clin Cancer Res; 22(6) March 15, 2016 1479
Effects of Pharmacokinetic Modulation of Anti-DLL4 Antibody
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380

2016;22:1469-1479. Published OnlineFirst November 20, 2015.Clin Cancer Res Jessica A. Couch, Gu Zhang, Joseph C. Beyer, et al. Pharmacokinetic ModulationBalancing Efficacy and Safety of an Anti-DLL4 Antibody through
Updated version
10.1158/1078-0432.CCR-15-1380doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2015/11/20/1078-0432.CCR-15-1380.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/22/6/1469.full#ref-list-1
This article cites 34 articles, 13 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/22/6/1469To request permission to re-use all or part of this article, use this link
on May 22, 2020. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 20, 2015; DOI: 10.1158/1078-0432.CCR-15-1380