2. photosynthesis: the light reaction and …€¦ · 2. photosynthesis: the light reaction and...
TRANSCRIPT

2. PHOTOSYNTHESIS: THE LIGHT REACTION AND
CARBON METABOLISM
The photosynthesis is the sequence of reactions, performed by green plants, blue-green
algae and photosynthetic bacteria, in which light energy from the sun is converted into
chemical energy and used to produce carbohydrates and ultimately all the materials of the
plant (Figure 2.1.).
Fig. 2.1. The basic scheme of photosynthesis
in leaves of plants.
The photosynthetic reaction of green plants and blue-green algae can be summarized as:
6CO2 + 12H2O + light energy ──► C6H12O6 + 6O2 + 6H2O.
There are two distinct phases in photosynthesis, the light (or light-dependent) reactions
and the dark (or light-independent) reactions. In green plants and blue-green algae the light
reactions involve the photolysis of water, producing hydrogen atoms and molecular oxygen.
This oxygen, given off during photosynthesis, is the main source of atmospheric oxygen,
essential for aerobic organisms. The hydrogen atoms produced are used to reduce NADP+ to
NADPH+H+ and the energy released also forms of ATP from ADP and inorganic phosphate
(photophosphorylation). This ATP and NADPH+H+ are used up during the dark reactions in
which carbon dioxide is fixed into carbohydrates.

2.1. Leaf as photosynthetic organ The main photosynthetic organ of most green plants is a leaf, consisting of a lateral
outgrowth from a stem and comprising lamina, petiole, and leaf base. The leaf typically
consists of conducting tissues and photosynthetic cells (the mesophyll) often differentiated
into palisade and spongy mesophyll, surrounded by epidermis. The epidermis is perforated by
a leaf pores, called stomata, usually more numerous on the abaxial (lower) side of the leaf.
The epidermis is usually covered by a waxy cutinized layer termed the cuticle. This prevents
excessive water loss by transpiration.
2.1.1. Chloroplast
The photosynthetic cell includes the special cellular organelles called chloroplasts
(Figure 2.2.). Chloroplasts are one of the family organelles bounded by a double membrane
and known generally as plastids. As the name implies, chloroplasts are identified by the fact
that they contain the chlorophyll pigments responsible for the green colour of leaves. In
addition to chlorophyll, chloroplasts contain large amounts of carotenes and xanthophylls.
Fig. 2.2. Structural model of a
chloroplast from the assimilatory
tissue of a higher plant (from Mohr
et Schopfer 1995).
A typical higher plant
chloroplast is generally described
as discoid with a maximum
diameter of 5 to 10 µm.
Chloroplasts are located in the
cytosol of the cell and,
consequently, are normally seen
pressed between the cell wall and the prominent central vacuole. Chloroplasts are most often
limited to the inner, or mesophyll, leaf cells and stomatal
guard cells, although a species for which chloroplasts may be found in epidermal cells are
known. In those species that have epidermal chloroplasts and their number per cell are
generally smaller than in the mesophyll cells. The number of chloroplasts in the mesophyll

cell is typically in the range of 10 to 100, although values of several hundred have been
reported for some species. As well, the chloroplasts in palisade mesophyll cells are generally
larger and more numerous than in the spongy mesophyll cells.
We recognize four major structural regions or compartments in chloroplasts (see Figure
2.2.): (1) a pair of outer limiting membranes, collectively known as the envelope, (2) an
unstructured background matrix or stroma, (3) a highly structured internal system of
membranes, called thylakoids, and (4) the intrathylakoid space, or lumen. The envelope
defines the outer limits of the organelle. These membranes are 5.0 to 7.5 nm thick and are
separated by a 10.0 nm intermembrane space. Because the inner envelope membrane is
selectively permeable, the envelope also serves to isolate the chloroplast and regulate the
exchange of metabolites between the chloroplast and the cytosol that surrounds it. The
intermembrane space is freely accessible to metabolites in the cytoplasm. Thus it appears that
the outer envelope membrane offers little by way of a permeability barrier. It is left to the
inner envelope membrane to regulate the flow of molecular traffic between the chloroplast
and cytoplasm.
The envelope encloses the unstructured background matrix of the chloroplast or stroma.
The composition of the stroma is predominantly protein. The stroma contains all of the
enzymes responsible for photosynthetic carbon reduction, including ribulose-1,5-
bisphosphate carboxylase/oxygenase, generally referred to by the acronym rubisco. Rubisco,
which accounts for fully half of the total chloroplast protein, is no doubt the world’s single
abundant protein. In addition to rubisco and other enzymes involved to carbon reduction, the
stroma contains enzymes for a variety of other metabolic pathways as well as DNA, RNA,
and the necessary machinery for transcription and translation of protein.
The internal chloroplast membranes form a complex system of granal and intergranal
lamellae. The grana are formed from two or three up to approximately a hundred disclike
flattened vesicles (thylakoids) stacked on top of each other. They are oriented in a variety of
directions relative to the long axis of the chloroplast and their number and size varies with
different species. The thylakoids found within a region of membrane sacking are called grana
thylakoids. Some thylakoids, quite often every second one, extend beyond the grana stacks
into the stroma as single, nonappressed thylakoids. These stroma thylakoids are flexible
interconnecting channels, continuous with and linking together the channels of individual
grana. While the organization of thylakoids into stacked and unstacked regions is typical, it is
be no means universal. The chloroplasts in the bundle sheath cells of C4 photosynthetic plants
do not contain grana. The thylakoids membranes contain the chlorophyll and carotenoid

pigments and are the site of the light-dependent, energy-conserving reactions of
photosynthesis.
The interior space of the thylakoids is known as the lumen. The lumen is the site of
water oxidation and, consequently, the source of oxygen evolved in photosynthesis.
Otherwise it functions primarily as a reservoir for protons that are pumped across the
thylakoids membrane during electron transport and that are used to drive ATP synthesis.
2.1.2. Endosymbiotic theory about genesis of plastids
This theory says that plastids and mitochondria arose from symbiotic prokaryotic
organism living within a eukaryotic host cell. It is thought that plastids probably originated
from organism similar to present-day blue-green algae while mitochondria arose from aerobic
bacteria. Such conclusions are based on various similarities between plastids and
mitochondria and free-living prokaryotes. For example, such organelles are self replicating
and contain DNA, which in addition to having a different base composition from the nuclear
DNA, are circular rather than linear. The ribosomes of chloroplasts and mitochondria are
smaller than those in other parts of cytoplasm but similar in size to those of prokaryotes.
There is evidence moreover, that the origin of plastids could be polyphyletic, i.e. in different
groups of plants different types of symbionts have been incorporated. Thus in the higher
plants and Chlorophyta and Rhodophyta, which all have chloroplasts with a double
membrane, it is thought that the plastids are derived from a prokaryotic symbiont. However in
other groups of algae the chloroplasts may be surrounded by three or four membranes,
implying that these are derived from eukaryotic symbionts.
2.2. Photosynthetic pigments and light The light is a form of radiant energy. It is a visible electromagnetic radiation with
wavelengths ranging from 360 nm (violet) to 780 nm (far-red). Those regions of the spectrum,
which our eyes can detect, we perceive as violet, blue, green, yellow, orange, and red light.
Whereas ultraviolet (100-360 nm) and infrared (longer than 780nm) regions of the spectrum,
which our eyes cannot detected, are referred to as ultraviolet and infra-red radiation,
respectively (see Table 2.1.). The energy of light can be absorbed by molecules called
pigments.

Table 2.1. Radiation and relation between wavelength range and energy of photons.
Colour Wavelength range (nm) Average Energy (kJ.mol-1 photons)
Ultraviolet 100 - 360
UV-C 100 - 280 471
UV-B 280 - 320 399
UV-A 320 - 360 332
Visible 360 - 780
Violet 360 - 425 290
Blue 425 - 490 274
Green 490 - 550 230
Yellow 550 - 585 212
Orange 585 - 640 196
Red 640 - 700 181
Far-red 700 - 780 166
Infra-red longer than 780 85
The green plants use in photosynthesis the spectrum between 400 nm and 700 nm (see
Figure 2.3.). This range of light is broadly defined as photosynthetically active radiation
(PAR). The PAR intensity is measured as the photon fluence rates, expressed as mol photons
m-2 s-1 PAR, or energy fluence rates, expressed as W m-2 PAR.
Fig. 2.3. The photosynthetically active radiation.

The chlorophylls (Figure 2.4.) are the main class of photosynthetic pigments. They are
pigments primarily responsible for harvesting light energy used in photosynthesis.
Chlorophylls absorb red and blue-violet light and thus reflect green light, so giving plants
their characteristic green colour. Chlorophylls are involved in the light reactions of
photosynthesis and are located in the chloroplast in thylakoids membranes. The chlorophyll
molecule consists of two parts, a porphyrin head and a long hydrocarbon, or phytol tail. A
porphyrin is a cyclic tetrapyrrole, made up of four nitrogen-containing pyrrole rings arranged
in a cyclic fashion. Completing the chlorophyll molecule is a magnesium ion (Mg2+) chelated
to the four nitrogen atoms in the centre of the ring. Four species of chlorophyll, designated
chlorophyll a, b, c, and d, are known. The chemical structure of chlorophyll a, the primary
photosynthetic pigment in all higher plants, algae, and cyanobacteria, is shown in Figure. The
summary chemical formula is C55H72O5N4Mg. Chlorophyll b is similar except that a formyl
group (-CHO) substitutes for the methyl group on ring II. The summary chemical formula of
chlorophyll b is C55H70O6N4Mg. Chlorophyll b is found in virtually all higher plants and
green algae, although viable mutants deficient of chlorophyll b are known. The principal
difference between chlorophyll a and chlorophyll c lacks the phytol tail. Finally chlorophyll
d, found only in the red algae, is similar to chlorophyll a except that a (─O─OCHO) group
replaces the (─OCH═CH2) group on ring II.
When grown in the dark, angiosperm seedlings do not accumulate chlorophyll. Their
yellow colour is due primarily to the presence of carotenoids. Dark-grown seedlings do,
however accumulate significant amounts of protochlorophyll a, the immediate precursor to
chlorophyll a. The chemical structure of protochlorophyll differs from chlorophyll only by the
presence of a double bond between carbons 7 and 8 in ring IV. The reduction of this bond is
catalysed by the enzyme NADH: protochlorophyll oxidoreductase. In angiosperm this
reaction requires light, but in gymnosperm and most algae chlorophyll can synthesized in the
dark. There is a general consensus among investigators that chlorophyll b is synthesized from
chlorophyll a.
The carotenoids (Figure 2.4.) comprise a family of yellow, orange or red pigments
present in most photosynthetic organism. Found in large quantity in roots of carrot and tomato
fruit, carotenoid pigments are also prominent in green leaves, where are located in the
chloroplast in thylakoids membranes. The carotenoids absorb blue-violet, blue and blue-green
light. In the fall of the year, the chlorophyll pigments are degraded and the more stable
carotenoid pigments account for the brilliant orange and yellow colours so characteristics of
autumn foliage. The carotenoid pigments are C40 terpenoids biosynthetically derived from the
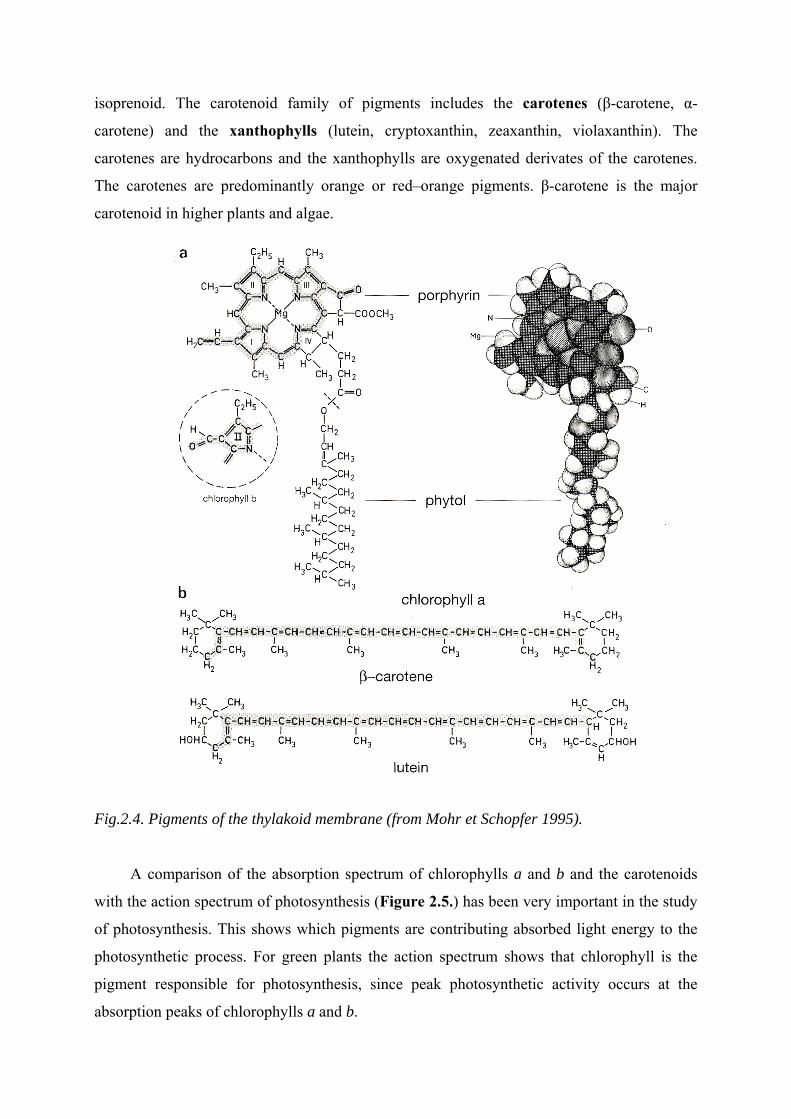
isoprenoid. The carotenoid family of pigments includes the carotenes (β-carotene, α-
carotene) and the xanthophylls (lutein, cryptoxanthin, zeaxanthin, violaxanthin). The
carotenes are hydrocarbons and the xanthophylls are oxygenated derivates of the carotenes.
The carotenes are predominantly orange or red–orange pigments. β-carotene is the major
carotenoid in higher plants and algae.
Fig.2.4. Pigments of the thylakoid membrane (from Mohr et Schopfer 1995).
A comparison of the absorption spectrum of chlorophylls a and b and the carotenoids
with the action spectrum of photosynthesis (Figure 2.5.) has been very important in the study
of photosynthesis. This shows which pigments are contributing absorbed light energy to the
photosynthetic process. For green plants the action spectrum shows that chlorophyll is the
pigment responsible for photosynthesis, since peak photosynthetic activity occurs at the
absorption peaks of chlorophylls a and b.

Fig. 2.5. A comparasion of the action spectrum of photosynthesis with the absorption spectra
of chlorophylls a and b and carotenoids (orig. Anonymous, modified by Hejnák 2005).
The phycobilins serve as accessory light-harvesting blue or red pigments in the blue-
green and red algae and cyanobacteria or as a critical regulatory system in green plants. Like
the carotenoids they are accessory pigments in photosynthesis, but unlike the chlorophylls and
carotenoids they are water soluble. Structurally they are very similar to the porphyrin part of
the chlorophyll molecule, except that they contain no magnesium. The three photosynthetic
phycobilins are phycoerythrin (also known as phycoerythrobilin), phycocyanin
(phycocyanobilin), and allophycocyanin (allophycocyanobilin). The phytochromobilin, fourth
of phycobilins, is an important photoreceptor that regulates various aspects of growth and
development of green plants. The phytochrome is a receptor that plays an important role in
many photomorphogenetic phenomena. Its chromophore structure and absorption spectrum
are similar to that allophycocyanin. The phytochrome (literally, plant pigment) is unique
because it exists in two forms that are photoreversible. The form P660 (or Pr) absorbs
maximally at 660 nm. However, absorption of 660 nm light converts the pigment to a second,
far-red absorption form P735 (or Pfr). Absorption of far-red light by Pfr converts it back to

the red-absorption form. Pfr is believed to be an active form of the pigment that is capable of
initiating a wide range of morphogenetic responses.
2.3. Photosynthetic electron transport and ATP synthesis In thylakoid membranes there are two photochemical systems containing photosynthetic
and accessory pigments and electron carriers. In this photosystems I and II (PSI and PSII)
there are groups of functionally cooperating pigment molecules consisting of photochemically
active chlorophyll a at the reaction centres and photochemically inactive chlorophylls a or b
and carotenoids as antenna pigments. The antenna pigments absorb light but do not
participate directly in photochemical reactions. However, antenna chlorophylls and
carotenoids lie very close together such that excitation energy can easily pass between
adjacent pigment molecules by a radiationless transfer process. The energy of absorbed
photons thus migrates through the antenna complex, passing from one chlorophyll molecule
to another until it eventually arrives at the reaction center.
Fig. 2.6. The model of a photosynthetic
pigment complex (photosystem). The energy
of the quanta absorbed by the antenna
pigments is transferred by radiationless
energy migration to a photochemically active
reaction centre of PSI with chlorophyll a
P700 or PSII with chlorophyll a P680. (after
Mohr et Schopfer 1995)
The reaction centre consists of one or
two molecules of chlorophyll a, called the
reaction centre chlorophyll, plus associated
proteins and cofactors. The reaction center chlorophyll is, in effect, an energy sink-it is the
longest wavelength, lowest energy-absorbing chlorophyll in the complex. Because the
reaction centre is the site of the primary photochemical redox reaction, it is here that light
energy is actually converted to chemical energy. The reaction centres of PSI and PSII are
designated as P700 and P680, respectively. These designations identify the reaction centre as
species of chlorophyll a, or pigment (P), with an absorbance maximum at either 700 nm (PSI)
or 680 nm (PSII). The efficiency of energy transfer through the antenna chlorophyll to the

reaction is very high-only about 10 percent of the energy is lost. The principal advantage of
associating a single reaction centre with a large number of antenna chlorophyll molecules is to
increase efficiency in the collection and utilization. The simplified model of the
photosynthetic pigment complex (photosystem) shows Figure 2.6. The photosystem I has a
chlorophyll a/b ratio of about 6-10/1, and the photosystem II has a chlorophyll a/b ratio of
about 1.2-2/1, most of the chlorophyll b. Third system in thylakoid membranes is the
cytochrome b/f complex, which transfer electrons from PSII to PSI. A schematic of the
photosynthetic electron transport chain depicting the arrangement of PSI, PSII, and the
cytochrome b/f complex in the thylakoid membrane is presented in Figure. A fourth complex-
the ATP synthase-is also shown. The ATP synthase use a proton gradient generated by
electron transport for ATP synthesis.
Fig. 2.7. The light reaction of photosynthesis. The Z-scheme of electron transport between
photosystems II and I (after Vodrážka 1993, modified).
P680 and P700 – reactions centre of photosystems II and I, with terminal pigments, Q and FeS – primary electron acceptors, PQ – plastoquinone, cyt b6/f – cytochromes, PC – plastocyanin, Fd – ferredoxin – translators of electrons, Z – donor of electrons, K – complex developing O2

2.3.1. Photophosphorylation
Light-driven production of ATP by chloroplasts is known as photophosphorylation
(photosynthetic phosphorylation). The light-induced photosynthetic electron transport is
utilized as a source of energy for the production of ATP from ADP and inorganic phosphate.
Photophosphorylation is very important because, in addition to using ATP (along with
NADPH+H+) for the reduction of CO2, a continual supply of ATP is required to support a
variety of other metabolic activities in the chloroplast. These activities include synthesis of
protein in the stroma and transport of proteins and metabolites across the envelope
membranes.
When the electron transport is operating according to the Z-scheme shown in Figures
2.7., electrons are continuously supplied from water and withdrawn as NADPH+H+ via
photosystems I and II. During the transfer of electrons from the primary electron acceptor of
photosystem II to P700 through plastoquinone, cytochrome b/f complex and plastocyanin, one
molecule of ATP is formed, because transfer of electrons between PSII and PSI is
energetically downhill. Some of that energy is used to move protons from the stroma side of
the membrane to the lumen side. These protons contribute to a proton gradient that can be
used to drive ATP synthesis (Figure 2.8.). This flow-through form of electron transport is
consequently known as noncyclic electron transport. Formation of ATP in association with
noncyclic electron transport is known as noncyclic photophosphorylation. The
photosynthetic splitting of water into gaseous oxygen and reducing equivalents is known as
photolysis of water (the Hill reaction).The two molecules of water are split to produce of one
molecule oxygen, four electrons, which go through the electron transport chain, and four
protons. The electrons and protons eventually reduce NADP+ to NADPH+H+. However PSI
units and PSII units in the membrane are not physically linked as implied by the Z-scheme,
but is even segregated into different regions of the thylakoid. One consequence of this
heterologous distribution in the membranes is that PSI units may transport electrons
independently of PSII, a process known as cyclic electron transport. In this case ferredoxin
transfers the electron back to PQ rather than to NADP+. The electron then returns to P700+,
passing through the cytochrome complex and plastocyanin. Since these electrons also pass
through PQ and the cytochrome complex, cyclic electron transport will also support ATP
synthesis, a process known as cyclic photophosphorylation. Cyclic photophosphorylation is a
source of ATP required for chloroplast activities over and above that required in the carbon
reduction cycle.

Fig. 2.8. The organization of the photosynthetic electron transport system in the thylakoid
membrane (from Hopkins 1995).
2.4. Photosynthetic carbon reduction (PCR) cycle In the chloroplast stroma there are the sequences of light-independent reactions (or dark
reactions) that utilize the energy (in the form of ATP) and reducing power (in the form of
NADPH) produced during light reactions of photosynthesis, to reduce carbon dioxide. This
process can take one of two forms, depending on whether the subject is a C3 or a C4 plant.
The details of the fixation of carbon dioxide differ in the two types of the plants but the end
result in both cases is the production of carbohydrates via the Calvin cycle.
The pathway by which all photosynthetic eukaryotic organisms ultimately incorporate
CO2 into carbohydrate is known as carbon fixation or the photosynthetic carbon reduction
cycle. Mapping the complex sequence of reactions involving the formation of organic carbon
and it is conversion to complex carbohydrates represented a major advance in plant
biochemistry. For his efforts and those of his associates, Calvin was awarded the Nobel Prize
for chemistry in 1961.
Calvin and his associates worked out this cycle of reactions by illuminating green algae
in the presence of radioactive carbon-14 (C14) dioxide for a couple of seconds and them
immersing the cell in boiling water to prevent further reaction. They then found which
metabolites first became radioactively labelled using chromatography.

Fig. 2.9. Metabolism and translocation of photosynthetic products formed by Calvin cycle
(from Mohr et Schopfer 1995).
2.4.1. Calvin cycle
The Calvin cycle (Figure 2.9.) begins with the carboxylation and cleavage of ribulose-
1,5-bisphosphate (RUBP) to form two molecules of three carbon acid, 3-phosphoglycerate
(3-PGA). Thus 3-PGA appeared to be the first stable product of photosynthesis. The
carboxylation reaction is catalyzed by the enzyme ribulose-1,5-bisphosphate
carboxylase/oxygenase, or Rubisco. Rubisco activity is light regulated. Its activity declines
rapidly to zero when the light is turned off and is regained only slowly when the light is once
again turned on. Rubisco is without doubt the most abundant protein in the world, accounting
for approximately 50 percent of the soluble protein in the most leaves. The enzyme also has a
high affinity for CO2 that, together with is high concentration in the chloroplast stroma,
ensures rapid carboxylation in the normally low atmospheric concentrations of CO2. Others
sugars that accumulated the label later in time were probably derived from 3-PGA. Because
Calvin’s group determined that the first product was a three-carbon molecule, the PCR cycle
is commonly referred to as the C3 cycle. Plants that incorporate carbon solely through the
PCR (or Calvin cycle) are generally known as C3 plants. The second step is the reduction of

3-PGA. The 3-PGA is removed by reduction to the triose phosphate, glyceraldehyd-3-
phosphate (G3P). The resulting triose sugar-phosphate, G3P, is available for export to the
cytoplasm, probably after conversion to dihydroxyacetone phosphate (DHAP). Once in the
cytoplasm, the triose molecules can easily be joined to synthesize hexose sugars, fructose-
phosphate and glucose-phosphate. These two hexose-phosphates then combine to form
sucrose phosphate. The glucose is subsequently converted to starch, cellulose, and other
polysaccharides. The acceptor molecule, RUBP is then regenerated by a complex series of
reactions involving 4-, 5- 6-, and 7-carbon sugar phosphates.
2.4.1.1. Energetic of the Calvin cycle
The reduction of each molecule of CO2 requires 2 molecules of NADPH and 3
molecules of ATP. This total presents energy input of 529 kJ mol-1. Oxidation of one mole of
hexose would yield like about 2817 kJ, or 469 kJ mol-1 of CO2. This represents an energy
storage efficiency of about 88 percent.
2.4.1.2. Photorespiration of C3 plants
The respiratory metabolism of green plants is not independent of light, but for most
autotrophic plants, respiration (CO2 release and O2 uptake) is much higher in the light than in
dark. This is substantiated by the observation that, for example, increase release of CO2 can
be measured for several minutes after illuminated leaves have been suddenly darkened. This
light-dependent CO2 evolution is called photorespiration. This process directly associated
with photosynthetic metabolism and involves the reoxidation of products just previously
assimilated in photosynthesis. It differs from dark respiration in that it does not occur in the
mitochondria and is not coupled to oxidative phosphorylation. The rate of CO2 release by
photorespiration in C3 plants can be three to five times greater than that released by dark
respiration. Since the process does not generate ATP it appears to be extremely wasteful. It
has been estimated that photosynthetic efficiency could be improved by 30-50 percent if
photorespiration were inhibited. Why photorespiration? It is because the Rubisco possesses a
dual function, carboxylation and oxygenation. As the concentration of O2 declines in air, the
relative level of carboxylation increases until, at zero O2, photorespiration is also zero. On the
other hand, an increase in the relative level of O2 (or decrease in CO2) shifts the balance in
favour of oxygenation. An increase in temperature will also favour oxygenation, since as the
temperature increases the solubility of gasses in water declines, but O2 solubility is less

affected than CO2. Thus O2 will inhibit photosynthesis, measured by net CO2 reduction, in
plants that photorespire.
Fig. 2.10. The photorespiration pathway
(from Hopkins 1995.)
The photorespiration pathway
(Figure 2.10.) involves the activities of
at least three different cellular organelles
(the chloroplast, the peroxisome, and the
mitochondrion) and, because CO2 is
evolved, results in a net loss of carbon
from the cell. The photorespiration
begins with oxidation of RUBP to 3-
PGA and phosphoglycolate (P-
glycolate). The 3-PGA is available for
further metabolism by the PCR cycle,
but the P-glycolate is rapidly
dephosphorylated to glycolate in the
chloroplast. The glycolate is exported
from the chloroplast and diffuses to the peroxisome. Taken up by the peroxisome, the
glycolate is oxidized to glycolate and hydrogen peroxide. The peroxide is broken down by
catalase and the glyoxylate undergoes a transamination reaction to form the amino acid
glycine. Glycine is then transferred to a mitochondrion where two molecules of glycine (4
carbons) are converted to one molecule of serine (3 carbons) plus one CO2. Glycine is thus
the immediate source of photorespired CO2. The serine then leaves the mitochondrion,
returning to the peroxisome where the amino group is given up in a transamination reaction
and the product, hydroxypyruvate, is reduced to glycerate. Finally, glycerate is returned to the
chloroplast where it is phosphorylated to 3-PGA.
2.4.2. C4 syndrome
Other groups of plants, known as C4 plants, posses the ability to reduce greatly
photorespiration by an additional extremely effective mechanism for CO2 fixation. In such
plants exists an alternative form of carbon dioxide fixation. The first product of CO2 fixation
is not the three-carbon phosphoglyceric acid but the four-carbon oxaloacetate. These C4

plants exhibit a number of specific anatomical, physiological and biochemical characteristics
that constitute C4 syndrome. One particular anatomical feature characteristics of most C4
leaves is the presence of two distinct photosynthetic tissues. In C4 leaves the vascular bundles
are quite close together and each bundle is surrounded by a tightly fitted layer of cells called
the bundle sheath. Between the vascular bundles and adjacent to the air spaces of the leaf are
more loosely arranged mesophyll cells. This distinction between mesophyll and bundle
sheath photosynthetic cells, called Kranz anatomy, plays a major role in the C4 syndrome. The
alternative C4 form of CO2 fixation was confirmed by M. D. Hatch and C. R. Slack in 1966
and is known as Hatch-Slack pathway (Figure 2.11.).
Fig. 2.11. The Hatch-Slack pathway (from Mohr et Schopfer).
2.4.2.1. Hatch-Slack pathway
The key to C4 cycle is the enzyme phosphoenol pyruvate carboxylase (PEPcase) which
catalyses the carboxylation of phosphoenol pyruvate (PEP) using the bicarbonate ion HCO3-
as the substrate (rather than CO2). The product of the PEPcase reaction, oxaloacetate (OAA),
is moderately unstable and is quickly reduced to a more stable C4 acid-either malate or
aspartate-which is transported out of the mesophyll cell into an adjacent bundle-sheath cell
situated around the leaf veins. Once in the bundle-sheath cell, the acid undergoes a
decarboxylation to form CO2 and pyruvate. The resulting CO2 so released reacts with
ribulose1,5-bisphosphate to form two molecules triose sugars 3-phosphoglycerate via the
Calvin cycle in the bundle-sheath chloroplast. The pyruvate is returned to the mesophyll cells
where it is converted to PEP with concomitant formation of a molecule of AMP from ATP.
This step, which uses up two high-energy phosphate bonds, is the reason why, overall, C4

plants require 30 molecules of ATP for each molecule of glucose synthesized whereas C3
plants only require 18.
Under optimal conditions, C4 crop species can assimilate CO2 at rates two to three times
that of C3 species. All this productivity does not, however, come “free”. There is an energy
cost to building to the CO2 concentration in the bundle-sheath cells. For every CO2
assimilated, two ATP must be expended in the regeneration of PEP. This is an addition to the
ATP and NADPH required in the PCR cycle. Thus the net energy requirement for
assimilation of CO2 by the C4 cycle is five ATP and two NADPH.
C4 plants are generally of tropical or subtropical origin representing nearly 1,500
species spread through at least 18 different angiosperm families (3 monocots, 15 dicots).
Interestingly, no one family has been found to express the C4 syndrome exclusively-all 18
families contain both C3 and C4 representatives. This suggests that the C4 cycle has arisen
rather recently in evolution of angiosperms and in a number of diverse taxon at different
times. Under conditions of high fluence rates and high temperature (30° to 40°C) the
photosynthetic rate of C4 species may be two to three times greater than that of C3 species.
They appear to be better equipped to withstand drought and are able to maintain active
photosynthesis under conditions of water stress that would lead to stomatal closure and
consequent reduction of CO2 uptake by C3 species. All of these features appear to be a
consequence of the CO2-concetrating capacity of C4 plants and the resulting suppression of
photorespiratory CO2 loss.
2.4.2.2. Photorespiration of C4 plants
In C4 plants photorespiration is hardly detectable, possibly because synthesis of
glycolate, the substrate for photorespiration, is much lower in C4 plants (about 10% of that of
C3 plants). This could be because the concentration of CO2 in the bundle sheath cells is so
high that oxidation (instead of carboxylation) of ribulose bisphosphate is prevented.
2.5. The structural, physiological and ecological differences between C3 and
C4 plants Unlike C3 plants, photosynthesis of C4 plants is not inhibited by O2, and they have a
very low CO2 compensation concentration. The CO2 compensation concentration is the
ambient carbon dioxide concentration at which the rate of CO2 uptake (for photosynthesis) is
balanced by the rate of CO2 evolution (by respiration). For C3 plants, values fall into the range

of 20 to 100 μl CO2 per litre. Comparable values for C4 plants are in the range of 0 to 5 μl
CO2 per litre. The C4 plants are particularly well suited to exploit, for active photosynthetic
metabolism, the naturally low CO2 concentration of air at high light fluxes and high
temperatures. This can be observed very well in C4 plants like maize and sugar cane.
Especially in plants of arid habitants, the C4 cycle is used as a mechanism to reduce water loss
by stomatal transpiration, which inevitably is coupled to the CO2 uptake into the leaf.
Photosynthesis in most situations is limited by available CO2 and water. In C3 plants,
even moderate water stress will initiate closure of the stomata and reduce the available supply
of CO2. The low CO2 compensation concentration of C4 plants means that they can maintain
higher rates of photosynthesis at lower CO2 levels .Thus C4 plants gain an advantage over C3
plants when the stomata are partially closed to conserve water during a period of water stress.
An effective measure of this advantage is the value of the transpiration ratio. The
transpiration ratio relates the uptake of CO2 to the loss of water by transpiration from the leaf.
Transpiration ratios for C4 plants are typically in the range of 200 to 350 (grams of the loss of
water by transpiration in order to produce 1 g dry matter), while for C3 plants value in the
range of 500 to 1000 are often cited. The low transpiration ratio for C4 plants reflects their
capacity to maintain high rates of photosynthesis while effectively conserving water.
The most C4 plants tend to have a higher temperature optimum (30-40°C) than C3 plants
(20-25°C). This difference is due at least in part to the higher temperature stability of some of
the C4 cycle enzymes. Maximal activity of PEPcase, for example, is in the range of 30 to
35°C compared with 20 to 25°C for Rubisco.
Another interesting feature of C4 plants is their general low-temperature sensitivity.
Maize, for example, will not grow at temperatures below 12 to 15°C. This lower limit for
growth is probably set by the enzyme pyruvate, phosphate dikinase, which is cold labile and
experiences a substantial loss of activity below 12°C.
Although C4 plants are not competitive in all situations-some C3 plants may even equal
or exceed C4 plants in productivity-given the right combination of high temperature, high
light and low water, the C4 confers a definite advantage. This advantage is reflected in the
observation that many of more aggressive weeds are C4 species. These include crabgrass
(Digitaria sanguinalis), Russian thistle (Salsoa kali), and several species of pigweed
(Amaranthus) that often take over during the hot, dry months in the middle of summer. Many
of the more highly productive crop species also fall within the C4 group, including sugarcane
(Saccharum officinarum), sorghum (Sorghum bicolor), maize (Zea mays), and millet
(Panicum miliaceum).

Amongst the xerophytic plants of arid, semi-arid or saline habitats there is a group of
plants which are striking in the remarkable structural and functional adaptations of their
photosynthetic apparatus to the special demands of the environment. These plants are able to
fix CO2 and synthesise organic substances at different times of the day. The mechanism is
known as Crassulacean acid metabolism (CAM).
2.6. Crassulacean acid metabolism, an alternative to C4 photosynthesis The CAM (Figure 2.12.) was so named because it was originally studied most
extensively in the family Crassulaceae. This specialised pattern of photosynthesis has now
been found in some 23 different families of flowering plants (including the Asteraceae,
Cactaceae, Crassulaceae, Euphorbiaceae and Liliaceae), one family of ferns (the
Polypodiaceae), and in the primitive plant Welwitschia.
The CAM plants keep their stomata closed during the day to reduce water loss by
transpiration. Carbon dioxide can therefore only enter at night using the carboxylating part of
the C4 cycle when, it combines with the three-carbon compound phosphoenol pyruvate (PEP)
to give the four-carbon oxaloacetate (OAA). As in the C4 plants, the enzyme PEP carboxylase
is central to CAM operation. The oxaloacetate is rapidly reduced by NAD-dependent malate-
dehydrogenase to malate, which can be stored in the cell vacuoles. During the daylight hours,
malate is retrieved from the vacuole, decarboxylated and the CO2 diffuses into the chloroplast
where is converted to triose phosphates by the C3 PCR (Calvin) cycle.
Fig. 2.12. The mechanism of CO2 fixation in CAM plants (from Mohr et Schopfer 1995).

2.6.1. Ecological significance of CAM
The CAM represents a particularly significant adaptation to exceptionally dry habitats.
Many CAM plants are true desert plants, growing in shallow, sandy soils with little available
water. Nocturnal opening of the stomata allows for CO2 uptake during periods when
conditions leading to evaporative water loss are at a minimum. Then, during the daylight
hours when the stomata are closed to reduce water loss, photosynthetic can proceed by using
reservoir of stored CO2. This interpretation is supported by the transpiration ratio for CAM
plants, in the range of 50 to 100, which is substantially lower than that for either C3 or C4
plants. There is a price to be paid, however. Rates for daily carbon assimilation by CAM
plants are only about one-half that of C3 plants and one-third that of C4 plants. The CAM
plants can be expected to grow more slowly under conditions of adequate moisture.
While some species, in particular the cacti, are obligatory CAM plants, many other
succulent exhibit a facultative approach to CAM. Under conditions of abundant water supply,
assimilates carbon as a typical C3 plant-there is no significant uptake of CO2 at night and no
diurnal variation in leaf cell acidity. Under conditions of limited water availability or high salt
concentration in the soil, CAM metabolism is switched on. Although carbon assimilation by
CAM is slower than with conventional C3 photosynthesis, its higher water use efficiency
permits photosynthesis to continue in times of water stress and the plant is better able to
complete its reproductive development.