universidad de chile facultad de medicina escuela … · glÁndulas salivales labiales de pacientes...
TRANSCRIPT

UNIVERSIDAD DE CHILE FACULTAD DE MEDICINA ESCUELA DE POSTGRADO
INFLUENCIA DE LA SEÑALIZACIÓN DE LA INTEGRINA α6β1 Y GTPASAS DE LA FAMILIA RHO EN EL CONTROL DEL FENOTIPO ACINAR POLARIZADO. POSIBLE ROL EN
LOS CAMBIOS DE POLARIDAD PRESENTES EN GLÁNDULAS SALIVALES LABIALES DE PACIENTES CON
SÍNDROME DE SJÖGREN
MÓNICA ISABEL BRITO WITTWER
TESIS PARA OPTAR AL GRADO DE DOCTOR EN CIENCIAS BIOMÉDICAS
Directoras de Tesis: Prof. Dra. María Julieta González Burgos Prof. Dra. Lisette Leyton Campos
2008

ii
UNIVERSIDAD DE CHILE FACULTAD DE MEDICINA ESCUELA DE POSTGRADO
INFORME DE APROBACION TESIS DE DOCTORADO EN CIENCIAS BIOMÉDICAS
Se informa a la Comisión de Grados Académicos de la Facultad de Medicina, que la Tesis de Doctorado en Ciencias Biomédicas presentada por la candidata
MÓNICA ISABEL BRITO WITTWER
ha sido aprobada por la Comisión Informante de Tesis como requisito para optar al Grado de Doctor en Ciencias Biomédicas en Examen de Defensa de Tesis rendido el día 22 de Agosto de 2008.
Prof. Dra. María. Julieta González Prof. Dra. Lisette Leyton Directora de Tesis Directora de Tesis
Dpto. Biología Celular y Molecular Dpto. Biología Celular y Molecular Fac. de Medicina, Universidad de Chile Fac. de Medicina, Universidad de Chile
COMISION INFORMANTE DE TESIS
Prof. Dr. Enrique Brandan Prof. Dr. Jorge Martínez
Prof. Dr. Héctor Gatica Prof. Dra. Jimena Sierralta Presidenta Comisión de Examen

iii
Esta tesis fue realizada en el laboratorio de Biología Celular del Programa de
Biología Celular y Molecular del Instituto de Ciencias Biomédicas de la Facultad de
Medicina de la Universidad de Chile, bajo la dirección de las profesoras Dra. María
Julieta González Burgos y Dra. Lisette Leyton Campos. Financiada por los proyectos
Fondecyt 1050192 y 1070699, Beca de Postgrado Conicyt 103318 y laboratorios
farmacéuticos.
Todos los individuos participantes en este estudio fueron diagnosticados y
clasificados por el Dr. Sergio Aguilera Covarrubias, médico Reumatólogo de la
Clínica INDISA, y co-investigador del proyecto Fondecyt 1050192. Esta institución
facilita, sin costo los pabellones quirúrgicos e insumos para realizar las biopsias de
glándulas salivales labiales, que se practican gratuitamente a los pacientes con fines
diagnósticos. Además, financia los exámenes de laboratorio que no pueden ser
cancelados por los pacientes.

iv
-.Como el resultado de todo mi esfuerzo, esta tesis es dedicada a mi madre y hermano.-

v
AGRADECIMIENTOS
Al terminar este proceso, quiero expresar mi agradecimiento a todos aquellos
que estuvieron presentes de distintas maneras durante estos años, y me ayudaron a
llegar hasta este punto.
Agradezco a mi madre y hermano, Delia y Jorge, por su cariño y apoyo
incondicional, y por haber entendido que el camino que tomé no es el más fácil ni el
más recompensado, pero es mi camino.
A mis Directoras de Tesis, Dra. María Julieta González y Dra. Lisette Leyton,
por compartir conmigo sus experiencias, conocimientos y reflexiones, las que me han
permitido ir formando una visión científica propia.
A la Comisión evaluadora, Dra. Jimena Sierralta, Dr. Jorge Martínez, Dr.
Enrique Brandan y Dr. Héctor Gatica, quienes con su visión crítica permitieron
fortalecer este trabajo.
Agradezco el apoyo generoso de todos los integrantes de los laboratorios de:
Dr. Remigio López, Dr. Norbel Galanti, Dr. Mario Galindo, Dr. Jorge Sanz, Dra.
Patricia Iturra, Dra. Nancy Olea, Dr. Bruce Baum, Dr. Matthew Hoffman y Dr. Arnoud
Sonnenberg.
A los Drs. Sergio Aguilera y Rafael Romo, por su gran aporte en la parte
clínica de esta tesis.
Además agradezco a las Dras. Cecilia Alliende y Cecilia Leyton, que con su
experiencia, pero sobre todo su alegría diaria, permitieron que esto funcionara.
A Javier Esparza, Mónica Peralta, Osvaldo Almeida, Mónica Astudillo y Paola
Vergara, por haberme ayudado desde sus distintas áreas.

vi
A todos los compañeros y amigos que han compartido conmigo los últimos 6
años: Paola, Claudio M, Yoon-Jeoung, Patricia, José, Marisol, Cecilia, Isabel, Sergio,
Verónica, Marianela, Claudio F, Gonzalo, Melina, Daniela, Mariana y Don Ramón…
porque los días fueron mas dulces con ustedes ahí.
Y finalmente, a mis amigas del alma Paola y Angélica, por el gran cariño que
continuamente me demuestran y que permitió que en los momentos difíciles
surgieran fuerzas de la nada para concluir este proyecto.

vii
INDICE DE MATERIAS
PáginasINTRODUCCIÓN 1
I. Generalidades del síndrome de Sjögren…………………………..……................ 1
II. Mecanismos etiopatogénicos del síndrome de Sjögren…………..…………........ 2
III. La célula epitelial como centro del síndrome de Sjögren………...……............... 3
IV. Control de la polaridad ejercida por la lámina basal…..……………..…….......... 4
V. La integrina α6 como señalizador que controla polaridad…................................ 6
VI. Señalización a través de integrinas………………….....…………........................ 7
VII. Señales involucradas en la polaridad epitelial……………...………….............. 8
VIII. Importancia de los cultivos en tres dimensiones….………….…………….........
10
HIPÓTESIS
13
OBJETIVOS
14
MATERIALES Y MÉTODOS
I. Materiales…………………………........................................................................ 15
1. Sales y reactivos………….….......................................................................... 15
2. Partidores….……….........……………............................................................ 15
3. Anticuerpos…….………..………………......................................................... 16
II. Diagnóstico de pacientes y obtención de GSL…………………….………............ 20
1. Diagnóstico de pacientes…...………………………......................................... 20
2. Criterio para el diagnóstico del SS primario……...……………………........... 20
3. Características de los individuos del estudio……………………………......... 21
4. Biopsia de GSL…..……...………………………………...….……….…............ 21
III. Métodos…..…..…….………………………………………………………............. 23
1. Obtención de fracciones enriquecidas en acinos y ductos……..………......... 23
2. Obtención de extractos proteicos………..….………...…………..……........... 24
3. Cuantificación de proteínas………………………………….......…….............. 24

viii
4. Electroforesis en geles de poliacrilamida…..………………………….........… 25
5. Westernblot…..……………………………………………………………........... 25
6. Inmunoprecipitación……..…………………………………………...….........… 28
7. Inmunofluorescencia en cortes por congelación de GSL……………..........… 28
8. Cultivo de células HSG en plano………..………………………...…............... 29
9. Cultivo de acinos de células HSG en 3D…...….…………………….............… 29
10. Inmunofluorescencia en acinos de células HSG........................................... 32
11. Microscopía electrónica de transmisión de acinos de HSG…..…...........… 32
12. Extracción de RNA total……………………………...…………...…............... 33
13. Electroforesis de RNA……..……………………………...……….........…..… 34
14. Síntesis de cDNA…...……………………………………………….........……. 35
15. PCR…………..…………………………………………………….........…….... 35
16. Tratamiento con anticuerpo bloqueante……..…………………………......... 37
17. Expresión y purificación de las proteínas RBD-GST y PBD-GST.............… 37
18. Determinación de los niveles de Rho A, Cdc42 y Rac 1 activos...........……. 38
19. Análisis estadístico………………………………...........................…………...
39
RESULTADOS 40
I. Análisis de la expresión y localización de las integrinas α6 y β1 en GSL de
controles y pacientes con síndrome de Sjögren………………..…………………….. 41
1. Niveles proteicos de las integrinas α6 y β1………………..…..…….............… 41
2. Inmunoprecipitación de las integrinas α6 y β1……………………...….........… 42
3. Localización de las integrinas α6 y β1……………………………..…............… 44
II. Determinación de los niveles de FAK fosforilada en glándulas salivales labiales
de controles y pacientes con Síndrome de Sjögren…............................................. 47
III. Establecimiento y caracterización de un sistema de cultivo en tres
dimensiones de acinos de células HSG…………………...…………………………... 49
1. Caracterización morfológica de los acinos de células HSG…..……..........… 49
2. Caracterización molecular del fenotipo polarizado de acinos de células
HSG....................................................................................................................
55

ix
IV. Importancia de la integrina α6 en la organización acinar y el establecimiento
del fenotipo polarizado……………..…………………………………………………… 61
1. Expresión de las integrinas α6 y β1 en acinos de células HSG. ............…... 61
2. Efecto del bloqueo de la integrina α6 sobre la organización y mantención
de fenotipo polarizado de acinos de células HSG…………................................ 65
V. Relación entre la señalización de la integrina α6 y las GTPasas de la familia
Rho en acinos de células HSG……………..………………….…………..……………
79
DISCUSIÓN 84
I. Relación las integrinas α6 y β1 con el SS……………………………...….........….. 85
II. Utilización del cultivo tridimensional de acinos de células HSG como modelo
de estudio de morfogénesis acinar y polaridad: ¿posible utilización como modelo
experimental para el SS?......................................................................................... 89
III. Rol de la integrina α6 en la desorganización acinar observada en GSL de
pacientes con Síndrome de Sjögren ………………..…………………………............ 93
IV. Rol de la integrina α6 en el control de la sobrevida en GSL, ¿posible relación
con el desarrollo del Síndrome de Sjögren ………………….…………….........……. 97
V. Relación de las Rho GTPasas con la desorganización acinar presente en el
SS y con la señalización río abajo de la integrina α6………..……..................……..
99
CONCLUSIONES
103
BIBLIOGRAFÍA
104

x
INDICE DE FIGURAS
Página
Figura 1 Esquema explicativo del cultivo en 3D de células HSG…………...... 31
Figura 2 Determinación de los niveles proteicos e inmunoprecipitación de
las integrinas β1 y α6…..……......……………………………………… 43
Figura 3 Localización de las integrinas α6 y β1 en acinos de GSL de
individuo control y paciente con SS……..…………..............………... 46
Figura 4 Niveles de FAK fosforilada (Tyr397) en extractos de glándulas
salivales labiales de controles y pacientes con SS............................ 48
Figura 5 Curso temporal del cultivo de acinos de células HSG en extracto de
lámina basal…………………………………………………….........…... 51
Figura 6 Análisis morfológico de acinos de células HSG……..….............…… 54
Figura 7 Comparación de la organización de acinos crecidos en extracto de
lámina basal con respecto a células crecidas en plano y acinos de
GSL………………………...........................……………………………. 57
Figura 8 Adhesiones célula-célula en acinos de células HSG...............…...… 59
Figura 9 Caracterización molecular del fenotipo polarizado en acinos de
células HSG………...……………...……………...............................… 60
Figura 10 Expresión de las integrinas α6 y β1 en acinos de células HSG…… 63
Figura 11 Co-localización de las integrinas α6 y β1 en acinos de células
HSG……………………………………….………………………………. 64
Figura 12 Efecto del bloqueo de la integrina α6 en la organización de acinos
de células HSG…………………….………………………………..….... 68
Figura 13 Especificidad del efecto del bloqueo de la integrina α6 en la
organización de acinos de células HSG……………...…….............… 69
Figura 14 Efecto del bloqueo de la integrina α6 en la organización de la
lámina basal en acinos de células HSG….........................……..…… 71

xi
Figura 15 Efecto del tiempo de bloqueo de la integrina α6 en la organización
de acinos de células HSG………………………………………………. 74
Figura 16 Efecto del tiempo de bloqueo pulsado de la integrina α6 en la
organización de acinos de células HSG…...……........................…… 75
Figura 17 Detección de F-actina en acinos de células HSG luego de la
inhibición de la integrina α6………………..............……..….………… 77
Figura 18 Redistribución de la integrina β1 en acinos de células HSG como
respuesta a la inhibición de la integrina α6…………………………… 78
Figura 19 Efecto del bloqueo de la integrina α6 en los niveles de Cdc42
activo…………………………………………….......................…...…… 81
Figura 20 Efecto del bloqueo de la integrina α6 en los niveles de Rho A
activo…………………………………..................…………....………… 82
Figura 21 Efecto del bloqueo de la integrina α6 en los niveles de Rac 1
activo………………………………….…………………….…………..… 83

xii
INDICE DE TABLAS
Página
Tabla 1 Características de los partidores y tamaño del producto de
amplificación esperado…………….………………..………………..…
17
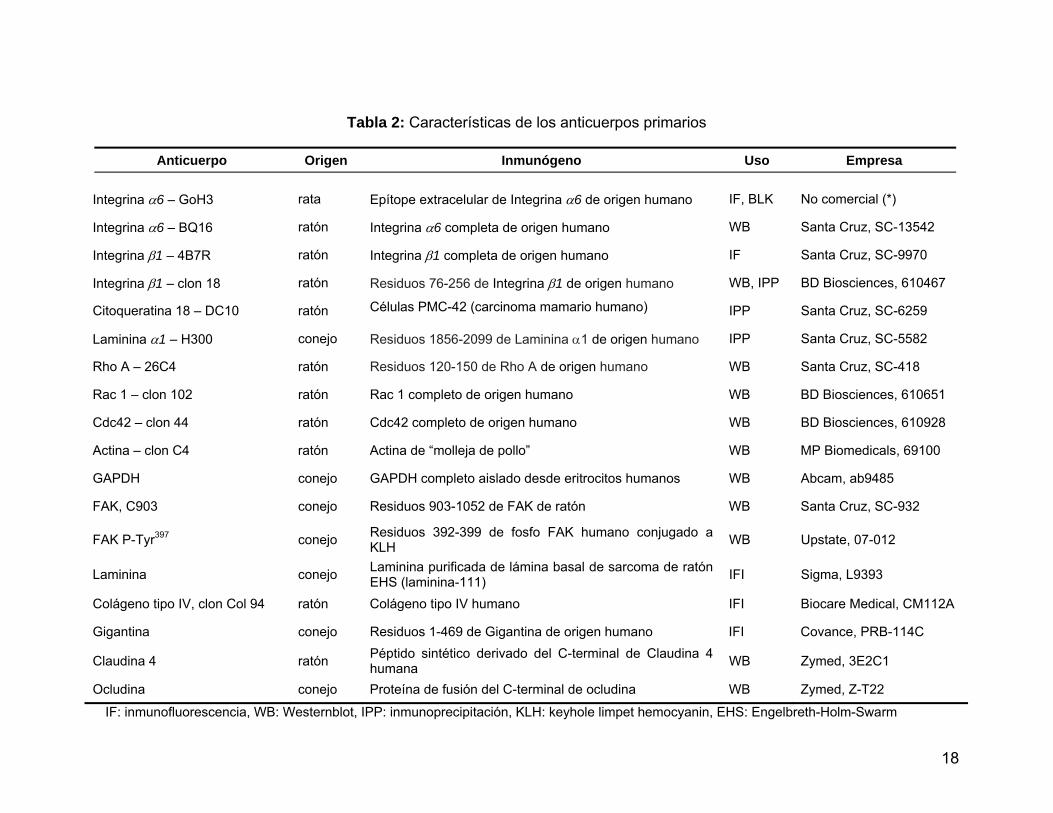
Tabla 2 Características de los anticuerpos primarios…..……………...……... 18
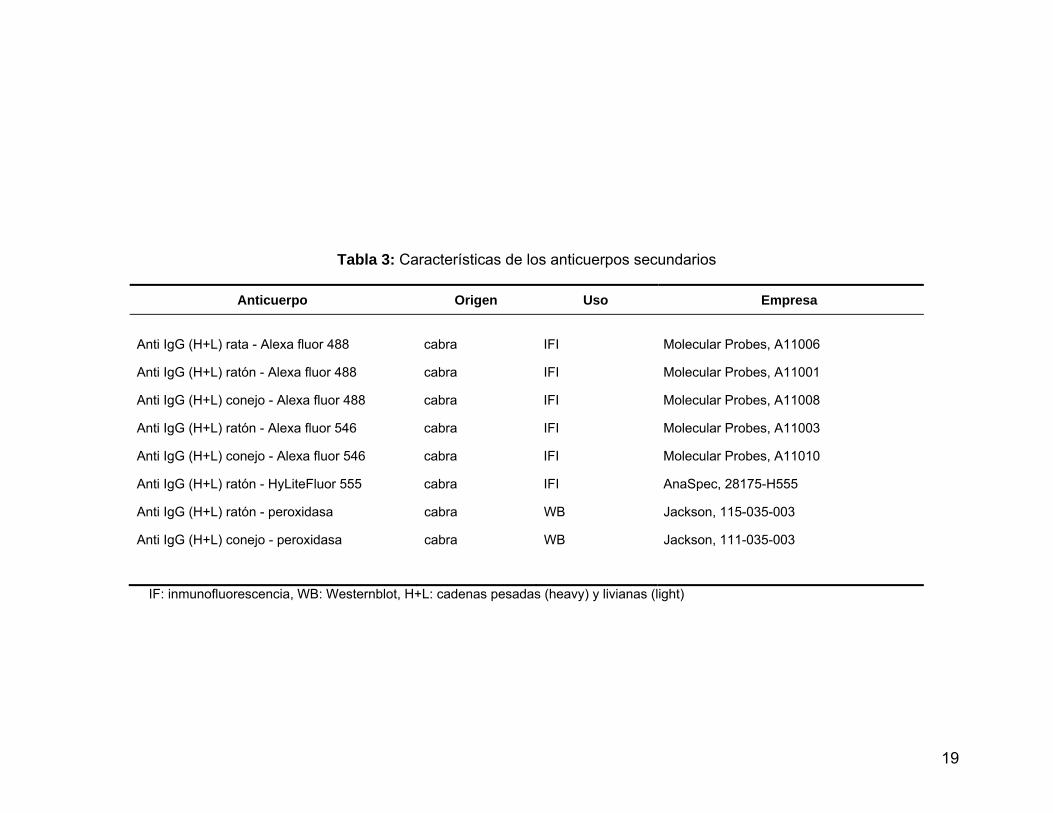
Tabla 3 Características de los anticuerpos secundarios.…………...………... 19
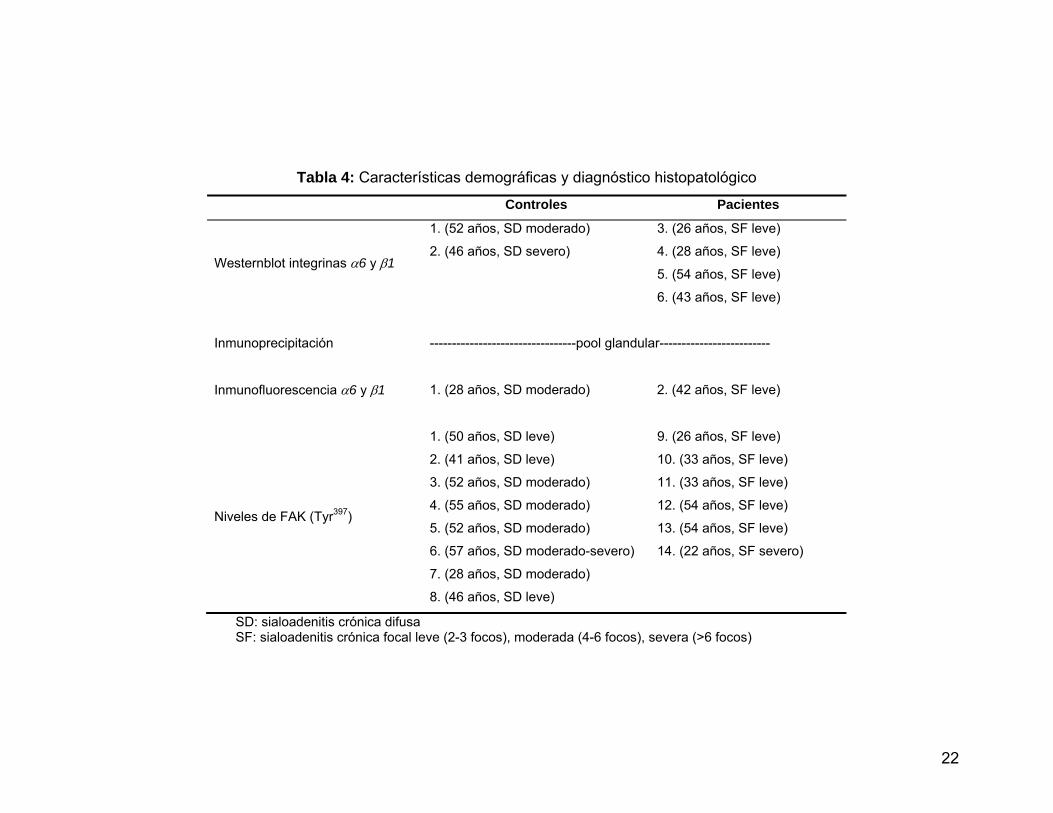
Tabla 4 Características demográficas y diagnóstico histopatológico……..… 22
Tabla 5 Condiciones de Westernblot……………………………………..…….. 27
Tabla 6 Programas de PCR………...………………………………..………….. 36

xiii
LISTA DE ABREVIATURAS
2D Cultivo en dos dimensiones (plano) 3D Cultivo en tres dimensiones aPKC Proteina kinasa C atipica Aº Amstrong ATP Adenosina -5'-trifosfato b-FGF Factor de crecimiento fibroblástico básico BLK Ensayo de bloqueo BSA seroalbúmina de bovino cDNA DNA codificante Cla-4 Claudina 4 CK18 Citokeratina 18 CTP Citidina-5'-trifosfato CYS Cistatina DABCO Biciclo(2,2,2)-1,4-diasa octano DMEM-F12 Mezcla 1:1 medio Dulbecco modificado de Eagle y solución
F12 de Ham DNA ADN, ácido desoxiribonucleico dATP 2´-Desoxi-Adenosina 5´-trifosfato dCTP 2´-Deoxi-Citidina 5´- trifosfato dGTP 2´-Desoxi-Guanosina 5´-trifosfato dTTP 2´-Desoxi-Timidina 5´-trifosfato EDTA Ácido etileno diamino tetracético EGF Factor de crecimiento epidermal EGTA Acido etilen-glicol bis(beta-aminoetil ether)-N,N,N',N'-t
tetracético EHS Sarcoma de ratón Engelbreth-Holm-Swarm FAK Kinasa de adhesión focal FasL Ligando de Fas FITC Fluoresceina iso-tiocianato GAP Proteína activadora de la actividad GTPásica GAPDH Gliceraldehído 3-fosfato deshidrogenasa GDI Inhibidor de la disociación de nucleótidos de guanina GDP Guanosina- 5'-difosfato GEF Factor intercambiador de nucleótidos de guanina GSL Glándulas salivales labiales GTP Guanosina- 5'-trifosfato HGF Factor de crecimiento de hepatocitos HSG Línea celular derivada de glándulas salivales humanas IgG Inmunoglobulina IGF1 Factor de crecimiento tipo insulina 1 IL-1 Interleukina-1 IL-6 Interleukina-6 INFγ Interferon α IPP Inmunoprecipitación

xiv
kDa kiloDalton LM111 Laminina-111 (Laminina-1) mRNA RNA mensajero MAPK Proteína kinasa activada por mitógenos MDCK Células epiteliales de riñón canino Madin-Darby MMP Metaloproteinasa de matriz extracelular MOPS Acido 3-(N-morfolino) propano sulfónico MUC1 Mucina 1 MUC5B Mucina 5b MUC7 Mucina 7 nm nanómetro NOD Ratones diabéticos no obesos NOD-scid Ratones NOD con inmunodeficiencia severa combinada NFkB Factor nuclear kappa beta pb Pares de bases pFAK FAK fosforilada PAGE-SDS Electroforesis en gel de poliacrilamiada con SDS PBD-GST Dominio de unión a Cdcd42 y Rac de PAK fusionado a GST PBS Tampón fosfato salino PDGF Factor de crecimiento de plaquetas PFA paraformaldehído PI3K fosfatidilinositol 3-kinasa PIPES Ácido piperacina-1,4-bis (2-etano) sulfónico PI4,5K Fosfatidilinositol 4,5-kinasa RNA ARN ácido ribonucleico RBD-GST Dominio de unión a RhoA de Rotekina fusionado a GST RIPA Tampón de radioinmunoprecipitiación RNAseout Inhibidor de ribonucleasas ROCK Rho kinasa RT-PCR Trascripción reversa acoplada de PCR SBF Suero fetal de bovino SDS Sodio dodecil sulfato SS Síndrome de Sjögren TAE Tampón Tris-Acetato- EDTA TBS Tampón Tris salino TGFβ Factor de crecimiento transformante β TNF-α Factor de necrosis tumoral - α TIMPs Inhibidores tisulares de metaloproteinasas TRITC Tetrametilrodamina isotiocianato Tris Tris (hidroximetil) aminometano TTP Timidia 5-trifosfato ZO-1 Zonnula occludens 1

xv
RESUMEN
El Síndrome de Sjögren (SS) es una patología autoinmune caracterizada por
una severa sequedad oral y ocular. La presencia de estos síntomas ha sido asociada
con una disfunción y disminución del parénquima secretor. Existen evidencias
morfológicas y moleculares que permiten plantear que las células acinares de
glándulas salivales labiales (GSL) de pacientes con SS presentan un patrón de
diferenciación alterado. Esto se asociaría con cambios en el fenotipo polarizado,
desanclaje de la lámina basal y apoptosis celular. En conjunto, estos eventos serían
responsables de la capacidad secretora alterada de las glándulas exocrinas en estos
pacientes.
Aunque la etiopatogénesis del SS es desconocida, existe mucha información
que sugiere que la lámina basal que rodea las células acinares tendría un rol
fundamental, ya que controla la polarización y sobrevida de las células epiteliales
durante el proceso de desarrollo glandular.
La pregunta central abordada en esta tesis fue: ¿Qué señales o receptores
podrían estar alterados y darían cuenta de los cambios morfológicos que sufren las
glándulas salivales de estos pacientes con SS?. Utilizando la técnica de Westernblot
se determinó que tanto la integrina α6 como β1 están presentes en células acinares
de pacientes con SS. En algunos pacientes la integrina α6 se encuentra disminuida y
la integrina β1 presenta una forma proteica de mayor movilidad electroforética. Existe
la posibilidad de que ambas integrinas estén involucradas en el desanclaje observado
en las células epiteliales de GSL y en los cambios en el fenotipo polarizado
encontrado en las células acinares de pacientes con SS. Sin embargo, la
señalización de integrinas no da cuenta del fenómeno, ya que la kinasa de adhesión
focal (FAK), una tirosina kinasa no receptora blanco de las señales derivadas de
integrinas, no presenta cambios en sus niveles de expresión ni activación,
determinado mediante Westernblot usando un anticuerpo fosfoespecífico para FAK.
Debido a la complejidad del modelo utilizado en estos estudios, nos
propusimos desarrollar un sistema de cultivo celular en 3D, donde pueden formarse
acinos de células derivadas de glándulas salivales humanas. El desarrollo exitoso de

xvi
este tipo de cultivo produjo acinos que permiten el estudio de polarización y
morfogénesis acinar. Sin embargo, por razones que en este momento no son
completamente entendidas, los acinos formados fueron no funcionales ya que no
expresan productos de secreción.
Igualmente, pudimos estudiar si las integrinas de la familia α6 pudiesen estar
involucradas en los cambios morfológicos que ocurren en las glándulas de pacientes
con SS. Usando anticuerpos bloqueantes de función, se determinó que las señales
recibidas de la integrina α6 son requeridas para el mantenimiento de la organización
acinar y la sobrevida celular, ya que la unión de anticuerpos contra α6 induce muerte
celular y fenotipo despolarizado. El efecto del anticuerpo depende del estado de
diferenciación alcanzado por las células, ya que el cambio fenotípico es menos
dramático cuando el anticuerpo es agregado tardíamente en el cultivo. Estos
resultados sugieren que, in vivo, las señales disminuidas o alteradas recibidas por las
integrinas de la familia α6, podrían ser responsables en parte de la morfología
alterada que se encuentra en GSL de pacientes con SS.
Otro punto interesante que se desprende de estos estudios, es el posible rol
que tendría la GTPasa Cdc42 en el proceso de morfogénesis acinar. Esta GTPasa
presenta una tendencia a la disminución en los niveles de activación durante este
proceso, pudiendo estar implicada en el desarrollo del fenotipo polarizado.
Finalmente, el establecimiento en nuestro laboratorio de este sistema de
cultivo en 3D, constituye un modelo importante en el estudio de morfogénesis acinar
salival, además de permitir estudiar los mecanismos moleculares implicados en el
control y mantenimiento del fenotipo acinar polarizado.

xvii
ABSTRACT
The Sjögren’s syndrome (SS) is an autoimmune disease characterized by
severe dryness of the mouth and eyes. Presence of these symptoms has been
associated with a decreased quantity and functionality of the remnant parenchyma.
Reportedly, morphological and molecular evidence support the idea that acinar cells
of labial salivary glands (LSG) from SS patients display an altered differentiation
pattern that yield cells of a non-polarized phenotype, that are detached from the basal
lamina and are undergoing apoptosis. Overall, these events might be responsible for
the altered secretory capacity of the exocrine glands in these patients.
Although the precise origin of SS is unknown, available information suggests a
critical role for the basement membrane surrounding the acinar cells, since this
specialized extracellular matrix controls polarization and survival of epithelial cells
during the process of gland development.
The main question this thesis sought to adress was to define the signals or
receptors that might be altered and could account for the morphological aberrations
found in the salivary glands of these SS patients. Western blot analysis revealed that
both α6 and β1 integrins subunits were present in acinar cells of SS patients.
However, α6 integrin was expressed at lower levels in some patients while an faster
movility form of the β1 protein was detected in most patients samples. Thus, changes
in both integrin subunits might be in part responsable for detachment of the epithelial
cells in the LSG and absence of the polarized phenotype found in the acinar cells of
SS patients. However, integrin signaling did not appear to account for the
phenomenum, since neither changes in the expression levels nor in activation of the
focal adhesion kinase (FAK), a non-receptor tyrosine kinase downstream target that
is activated as a consequence of integrin engagement, were detected by Western
blots performed with anti-phospho specific FAK antibodies.
Due to the complexity of the model utilized in these studies, we sought to
develop an in vitro 3D cell culture system, that permitted acini formation in vitro from
human salivary gland cells. These cultures yielded complete acini that were expected

xviii
to permite the study acini polarization and morphogenesis. However, for reasons that
remain to be defined, the acini formed were non- functional and did not express
secretory products.
In any case, this cultures did allow us to ask wether whether α6β1 integrin was
involved in the morphological changes observed in glands of SS patients. Using
inhibitory antibodies, α6 was shown to be required to maintain acini organization and
cell survival since binding of the anti-α6 antibodies induced cell death and a non-
polarized phenotype. The effect of the antibody depended on the stage of
differentiation of the cells, since phenotypical changes were less dramatic when the
antibody was added to the culture at later time points. These results suggest that,
perturbation in vivo of α6 integrins dependent signalling could, in part, be responsible
for the morphological changes observed in LSG of SS patients.
Another interesting point addressed in this thesis was to identify a possible role
for small GTPase Cdc42 in the process of acinar morphogenesis. This GTPase
decrease their activity levels during this process, probably implicating them in the
control of the polarized phenotype.
Finally, the development of these 3D cultures in our lab represent an important
model to investigate processes underlying acini morphogenesis as well as identify
study molecular mechanisms relevant to the control and maintenance of the polarized
acini phenotype.

1
INTRODUCCIÓN
I. Generalidades del síndrome de Sjögren
El síndrome de Sjögren (SS) es una enfermedad autoinmune sistémica, que
afecta principalmente a las glándulas exocrinas [1]. La manifestación clínica más
frecuente es una severa sequedad oral y ocular, debido a un deterioro progresivo y
crónico de las glándulas salivales y lacrimales [2]. Aunque la etiología es
desconocida, se ha planteado que factores inmunológicos, genéticos, psico-
endocrinos e infecciosos podrían contribuir al desarrollo de la enfermedad [3].
Las glándulas salivales están afectadas en casi todos los pacientes con SS [2]
y presentan una importante infiltración linfocítica periductal focal mayoritariamente
del tipo T CD4+ [4]. El carácter autoinmune del SS está dado por la presencia de
autoanticuerpos circulantes en el suero de los pacientes, siendo los más importantes
anti Ro (SSA) y anti La (SSB) [5]. La presencia de infiltrado glandular y de estos
autoanticuerpos son utilizados como parte del criterio para el diagnóstico de este
síndrome [6].
El SS es la segunda patología reumática autoimmune más común, después de
la artritis reumatoídea, con una prevalencia estimada entre 0,5% y 5% [7]. Afecta
más a mujeres que a hombres (9:1) y se diagnostica frecuentemente entre los 40 y
50 años de edad [3,8]. La prevalencia de este síndrome está subestimada pues
existe un desconocimiento dentro de los profesionales de salud en los criterios
diagnósticos [4].
El SS puede presentarse como patología única, denominándose SS primario,
o acompañado por otras enfermedades autoinmunes del tejido conectivo, en este
caso llamado SS secundario. Además, los pacientes con SS pueden desarrollar una
serie de manifestaciones extra glandulares que involucran distintos sistemas [1,9].
Por ejemplo, el SS se asocia con neuropatías periféricas, mialgias, artralgias,
desórdenes de la tiroides, desórdenes pulmonares, renales y renales, existiendo
además un incremento significativo en la incidencia de linfoma en pacientes con SS
primario [7].

2
El SS fue descrito inicialmente como una exocrinopatía autoinmune, y
actualmente es considerado una epitelitis autoinmune, dado que el epitelio de las
glándulas exocrinas parece ser el blanco de la respuesta inflamatoria. Por esta
razón, prácticamente todas las glándulas exocrinas se pueden ver afectadas en un
paciente con SS [3].
Como consecuencia directa de la sequedad oral, los pacientes con SS
presentan una mayor incidencia de caries dentales e infecciones orales [10]. Es
importante destacar que esta patología tiene un impacto negativo en la comunicación
y las habilidades alimenticias del paciente, pues la sequedad oral le genera
dificultades para hablar, masticar y tragar, afectando así aspectos críticos para una
calidad de vida satisfactoria [7]. Dado que esta enfermedad tiene un carácter crónico
y complejo, propicia además un deterioro en el estado físico general, psicológico y
social del paciente [11]. El tratamiento del SS es básicamente sintomático, donde se
debe incluir un grupo interdisciplinario buscando lograr un manejo integral de la
enfermedad [3].
Si bien el SS es una patología sistémica, nos hemos centrado en entender la
patogenia de esta enfermedad en las glándulas salivales. En particular, la sequedad
oral, que sufren estos pacientes, considerando el alto impacto que tiene en su
calidad de vida.
II. Mecanismos etiopatogénicos del síndrome de Sjögren
El modelo clásico de la etiopatogénesis del SS se basa en los conceptos de
autoinmunidad, donde el sistema inmune no es capaz de discriminar entre moléculas
propias y no propias. Así, el sistema inmune respondería contra autoantígenos que
se expresan en las células acinares y ductales de las glándulas exocrinas, en
especial salivales y lacrimales. Este proceso podría iniciarse por la combinación de
factores intrínsecos, como es la predisposición individual, y extrínsecos como son la
acción de agentes exógenos, como por ejemplo virus. Los autoantígenos
estimularían la actividad de las células T y B, promoviendo la síntesis de citoquinas y
quimioquinas, que favorecerían la persistencia del proceso autoinmune e inflamación

3
crónica con la consiguiente destrucción y pérdida de la función glandular [2].
Los mecanismos que llevan a la destrucción y pérdida de la función glandular
en los pacientes con SS aún son desconocidos, no obstante se postulan algunas
hipótesis. La destrucción del parénquima se ha relacionado preferentemente con la
actividad del infiltrado linfocitario, el cual provocaría daños citotóxicos en las células
epiteliales a través de procesos que incluyen la inducción de la apoptosis en células
epiteliales [12] y la denervación glandular por daño en las fibras nerviosas simpáticas
y parasimpáticas [13]. Por otro lado, la disfunción glandular se observa en un gran
número de pacientes que presentan falta de secreción, pero mantienen una
proporción importante de parénquima. Este hallazgo, se ha asociado con la pérdida
del acoplamiento neuro-glandular producido por la inhibición de la liberación de
acetilcolina, donde IL-1 y TNF-α serían responsables de esta inhibición [14]. Además,
se ha reportado la acción inhibitoria de autoanticuerpos contra el receptor
muscarínico del subtipo M3 [15,16].
III. La célula epitelial como centro de la patogenia del síndrome de Sjögren
La célula epitelial juega un rol central en la patogénesis del SS, tanto en el
período de iniciación como en la perpetuación de la etapa autoinmune [9]. Se ha
descrito que mecanismos como: la presentación antigénica, la apoptosis, o la
producción de citoquinas se encuentran en el centro de la patogénesis del SS y en
ellos la célula epitelial tiene un papel activo. Por esto, se plantea que la célula
epitelial no solo sería un blanco del sistema inmune sino que también un inductor de
la respuesta autoinmune [17].
Por otra parte, estudios realizados en ratones diabéticos no obesos (NOD),
uno de los modelos murinos para el estudio del SS, han permitido postular el
desarrollo bifásico de la patología, donde la parte inicial parece ser dependiente sólo
de alteraciones en la homeostasis glandular, y la fase final dependiente del
componente inmune [18,19]. Evidencias indican que en las glándulas salivales de
estos ratones están aumentadas las metaloproteinasas de matriz extracelular (MMP-
2 y 9) de forma independiente del componente inmune. Este aumento se ha

4
determinado también en ratones que carecen de linfocitos B y T funcionales (NOD-
scid) [20].
Nuestro grupo ha determinado que en glándulas salivales labiales (GSL) de
pacientes con SS existe una desorganización morfológica de la lámina basal que
rodea acinos y ductos [21]. En estos pacientes, el patrón de distribución de lamininas
y colágeno tipo IV se observa desorganizado y con una inmunorreactividad
aparentemente menor [22], asociado a una mayor síntesis y actividad proteolítica de
metaloproteinasas de matriz extracelular [23].
Por otro lado, en GSL de pacientes con SS ocurre con frecuencia cambios
importantes en la calidad del parénquima glandular. Existen evidencias morfológicas
y moleculares de alteraciones en el patrón de diferenciación y el fenotipo polarizado
de células acinares de GSL de pacientes con SS. Se describen cambios en la
polaridad nuclear y en la desorganización del polo apical, con pérdida parcial de las
microvellosidades, independiente de la cercanía y cantidad de células inflamatorias
[21]. Además, se observan alteraciones en la destinación proteica de proteínas
localizadas en el polo apical de las células acinares, como por ejemplo, redistribución
de aquaporina-5 [24], MUC1 [25] y ezrina [26], lo que podría dar cuenta de una
incorrecta organización de los dominios de membrana plasmática característicos de
la célula acinar.
Planteamos que esta “desorganización de la lámina basal” alteraría su
interacción con receptores localizados en el dominio basal de la membrana
plasmática de la célula acinar, modificándose los procesos celulares regulados por la
lámina basal.
IV. Control de la polaridad celular ejercida por la lámina basal
Las células acinares presentan una polaridad ápico-basal bien definida,
caracterizada por asimetría en la forma, distribución proteica y vectorialidad del
proceso de secreción [27]. El evento primario que desencadena la formación del
fenotipo polarizado, es el establecimiento de una asimetría física y molecular en la
superficie celular, iniciado por el contacto de las células epiteliales indiferenciadas

5
con la lámina basal y con células vecinas. Ambos tipos de asociación, inducen la
organización de adhesiones celulares especializadas, las cuales inicialmente están
formadas por integrinas y sus ligandos (célula-lámina basal) y E-cadherinas (célula-
célula), promoviendo así la definición de los ejes celulares de polaridad ápico-basal y
lateral [28].
Particular influencia en el control del establecimiento de los ejes de polaridad
se le ha conferido a uno de los ligandos de integrina, la laminina-111 (conocida como
laminina-1) [29,30]. Esta proteína regula la orientación del eje de polaridad ápico-
basal de manera dependiente de su correcto ensamblaje [31] y el establecimiento del
fenotipo polarizado [32]. En GSL se ha descrito la expresión de ella en la lámina
basal de acinos y ductos [22,33].
Existen evidencias de que la cadena α1 de laminina-111 está sobre expresada
en extractos de GSL provenientes de pacientes con SS [34]. Sin embargo, el análisis
por inmunofluorescencia de la distribución de esta cadena muestra una disminución
drástica de la señal [33] y la detección de laminina-111 muestra que está
desorganizada en acinos de pacientes con SS [22]. Estas evidencias permiten
plantear que la cadena α1 sería correctamente sintetizada en pacientes con SS, pero
la organización de laminina-111 en la red de la lámina basal se encontraría alterada,
pudiendo dar cuenta de un ensamblaje incorrecto, existiendo evidencia que la
perturbación del ensamblaje de laminina genera un patrón de localización por
inmunofluorescencia débil y desorganizado [30,31,35].
Por otra parte, en pacientes con SS existen altos niveles de degradación de
nidógeno 1 y 2 [34]. Se ha demostrado que alteraciones en la interacción de laminina
con nidógeno se asocia a la pérdida de estabilidad de la red de laminina y resulta en
patrones de diferenciación alterados [36] y desorganización de la lámina basal [37].
Así, las señales dependientes de laminina-111 estarían alteradas en las GSL de
pacientes con SS y podrían ser causantes de las alteraciones del fenotipo polarizado.
Esta interacción incorrecta de las células acinares con los componentes de lámina
basal permite plantear la hipótesis de que la desorganización de la lámina basal en
pacientes con SS, controla negativamente el desarrollo de la célula acinar, alterando
el proceso de diferenciación y polaridad epitelial.

6
V. La integrina α6 como señalizador que controla polaridad
Entre los receptores más estudiados para laminina-111 se encuentran
diversas integrinas y α-distroglicán, catalogándose las integrinas como los receptores
“clásicos” [38]. Las integrinas son proteínas transmembrana formadas por 2
subunidades (α y β) asociadas no covalentemente, hasta el momento en mamíferos
se describen 24 integrinas producto de la combinación de distintas subunidades α y β
[39]. Ambas subunidades están constituidas por un gran dominio extracelular, un
segmento transmembrana único y un dominio citoplasmático corto. El dominio
extracelular de la cadena α tiene 7 motivos repetidos que se pliegan formando un
dominio globular con el sitio de unión al ligando en la superficie, presentando además
sitios de unión a cationes divalentes [40].
En los epitelios, las integrinas α6β1 y α6β4 juegan un papel importante,
descritas ambas como receptores exclusivos para lamininas [41]. Sin embargo, la
integrina α6β4 generalmente está formando parte de los hemidesmosomas e
interacciona preferentemente con laminina-332 (conocida como laminina-5). En esta
tesis se analizará la integrina α6β1 como el receptor de laminina-111 [42].
La cadena α6 está formada por una cadena liviana y pesada unidas por un
enlace disúlfuro [43], existiendo dos variantes de procesamiento, denominadas α6A y
α6B (según el largo de su dominio citoplasmático), y ambas se expresan en GSL
[44]. La subunidad β1 es bastante promiscua, ya que forma parte de al menos 10
integrinas diferentes, y a pesar que muchas integrinas de la familia β1 pueden unir el
mismo ligando, cada una transduce una información única. En glándulas salivales
humanas, la subunidad β1 se ha detectado en el dominio de la membrana plasmática
basal de acinos y ductos [45]. No existen estudios acerca de la expresión del dímero
α6β1 en GSL, pero durante el desarrollo postnatal de la parótida se ha descrito su
expresión, con altos niveles después del nacimiento y una disminución posterior [46].
Considerando que se ha demostrado la expresión tanto de laminina-111 como de la
integrina α6 en acinos, es esperable que al ser α6β1 su receptor clásico, éste se
exprese funcionalmente en este tejido.

7
VI. Señalización a través de integrinas
En general, las vías de señalización descritas para las integrinas son
compartidas por los distintos dímeros, no encontrándose exclusividad para cada uno
[47]. Este conocimiento se basa en el análisis de procesos de adhesión y migración
de células cultivadas en plano sobre proteínas de matriz extracelular; difiriendo de lo
que ocurre in vivo o en sistema de cultivo en 3D donde los componentes de matriz no
son únicos [48]. Las colas citoplasmáticas de las integrinas son generalmente cortas
y carecen de actividad enzimática, transduciendo señales por asociación a proteínas
adaptadoras que conectan la integrina con el citoesqueleto de actina, kinasas
citoplasmáticas y receptores de factores de crecimiento [47]. Después de la unión a
su ligando, las integrinas se agregan y reclutan un gran número de proteínas hacia
su dominio citoplasmático, organizando las llamadas adhesiones focales,
describiéndose más de 50 proteínas que forman parte de ellas [49]. La kinasa de
adhesión focal (FAK) es considerada un punto central, ya que es activada por la
mayoría de las integrinas, actuando como “anclaje” de muchas moléculas
señalizadoras [50]. FAK es una tirosina kinasa no receptora, cuya asociación a
integrinas no está clara, pudiendo ser directa o a través de la interacción con otras
proteínas como paxilina y talina. Su activación depende de la fosforilación de al
menos 7 residuos de tirosinas, que provienen de su autofosforilación y fosforilación
vía Src [51]. Estos residuos fosforilados sirven de anclaje para kinasas de la familia
Src y proteínas adaptadoras que permiten la activación de la vía de MAPK y
GTPasas de la familia Rho, entre otras [47]. Las GTPasas están asociadas al
desarrollo del fenotipo polarizado en distintos sistemas. Además, la adhesión celular
a través de integrinas, juega un papel fundaméntela en el control de la sobrevida
celular. En esto, proteínas como PI3-Kinasa, central en el control de la sobrevida, se
activa de una manera controlada por FAK, constituyendo un posible mecanismo por
el cual una falta de función de FAK puede la anoikis, un tipo particular de muerte
celular promovida por el desanclaje [52].

8
VII. Señales involucradas en la polaridad epitelial
Las GTPasas de la familia Rho pertenecen a la superfamilia Ras, proteínas G
monoméricas [53]. En humanos se han descrito 20 distintas Rho GTPasas, siendo
las más estudiadas las de las subfamilias Rho (A, B y C), Rac (1, 2 y 3) y Cdc42
(Cdc42Hs y G25K) [54]. La principal función de estas proteínas es la regulación del
ensamblaje del citoesqueleto de actina [55]. De esta manera, ellas juegan un papel
en una variedad de procesos celulares dependientes del citoesqueleto de actina,
tales como: migración, citoquinesis, secreción y morfogénesis, entre otros [53,56]. La
activación de estas proteínas depende de la unión e hidrólisis de los nucleótidos de
guanina GDP y GTP. Estos procesos están regulados por la acción de proteínas
denominadas GEFs, GAPs y GDIs, las cuales promueven el intercambio GDP/GTP,
la hidrólisis de GTP o inhiben la disociación de GDP, respectivamente [57]. Teniendo
presente su efecto sobre la organización del citoesqueleto, cada una de ellas induce
la formación de estructuras específicas como las fibras de estrés y adhesiones
focales (Rho), lamelipodios y velos de membrana (Rac) y filopodios (Cdc42) [54]. Los
efectores identificados a la fecha incluyen más de 30 proteínas blanco, como kinasas
y proteínas andamio. El mecanismo de activación mas común que ejercen las
proteínas de la familia Rho es la ruptura de interacciones intramoleculares
autoinhibitorias y la exposición de dominios funcionales en la proteína efectora [53].
Algunos de los efectores involucrados en la reorganización del citoesqueleto son: la
kinasa ROCK y mDía para Rho, las kinasas PAK y PI4,5K y el complejo WAVE para
Rac, y por último PAK y N-WASP para Cdc42 [53,58].
Conjuntamente con el papel fundamental que cumple la reorganización del
citoesqueleto de actina en la definición del fenotipo polarizado epitelial, existen otros
procesos fundamentales que también son regulados por estas proteínas [56]. Se ha
asociado a distintos miembros de la familia Rho con el control del establecimiento del
fenotipo polarizado. En una línea celular derivada de cáncer mamario que ha perdido
el control de la polaridad, se demostró que la inhibición de Rac1 mediante el uso de
mutantes inactivos, es capaz de restablecer el eje de polaridad [59]. En un cultivo 3D
de cistos de células MDCK, la orientación del eje de polaridad ápico-basal requiere
de la actividad de Rac1 y de la polimerización correcta de laminina-111. Un

9
dominante negativo de Rac1 altera el ensamblaje de laminina sintetizada
endógenamente y por ende la definición de polaridad, dándose una inversión de la
polaridad apical [31]. Si bien en ambos casos atribuyen funciones distintas a Rac1
(como regulador directo/indirecto), queda claro su papel en el control del eje de
polaridad, ya que por sí solo define polaridad. En cuanto a destinación proteica, en
células MDCK polarizadas, Rac1 regula los niveles de endocitosis desde ambos
dominios de la membrana plasmática y la expresión de un dominante activo de Rac1
altera específicamente tráficos de membrana post-endocíticos y biosintéticos que
dirigen al dominio apical, pero no al baso-lateral [60].
El estudio de la polaridad celular en C. elegans y Drosophila ha revelado una
serie de proteínas de polaridad que se requieren para una localización correcta de
las proteínas celulares y para la división asimétrica que se da en el desarrollo
temprano. Estas incluyen el complejo PAR-aPKC, que es considerado un complejo
molecular conservado, fundamental para el establecimiento de la polaridad celular en
una variedad de contextos biológicos [61]. En mamíferos, éste es un complejo
ternario formado por las proteínas ASIP/PAR-3, aPKC y PAR-6 [61]. Además, se ha
descrito que se requiere de aPKC activa para el establecimiento de la polaridad
celular, dado que la sobre expresión de un dominante negativo de aPKC en células
MDCKII en proceso de polarización, causa redistribución de proteínas ubicadas en el
dominio apical, demostrándose también que la formación de las uniones estrechas
está severamente alterada en estas células [62]. Mediante ensayos de doble híbrido
se estableció que PAR-6 interacciona directamente con las formas activas de Rac y
Cdc42 [63], y PAR-6 regula la actividad de aPKC a través de su interacción con la
región regulatoria de la kinasa, la cual es modificada por Cdc42-GTP, uniéndose al
dominio CRIB/PDZ de PAR-6 [64].
Recientemente se ha descrito en células MDCKII que PAR-3 controla el
ensamblaje de las uniones estrechas, promoviendo la inactivación de Rac mediante
el secuestro o inactivación del GEF Tiam1 [65]. Esta interacción directa con Tiam1 y
Tiam2 (STEF) también se ha descrito en células N1E-115 de neuroblastoma donde
se encuentra formando un complejo con PAR-3–aPKC–PAR-6–Cdc42-GTP,
regulando así la activación de Rac dependiente de Cdc42, siendo importante en el

10
establecimiento de la polaridad neuronal [66]. También se ha descrito que Rho
kinasa (ROCK), un efector de Rho, es necesaria para el ensamblaje de uniones
estrechas en epitelio intestinal polarizado, teóricamente modulando el citoesqueleto
de actina [67].
Se ha descrito en literatura que en glándulas salivales de ratón las GTPasas
de la familia Rho son controladas por integrinas. La mutación del gen de la integrina
α3 en ratones, desencadena un fenotipo diferenciado aberrante en su glándula
submandibular y alteraciones en la expresión y localización de Rho GTPasas. RhoA
está disminuido en expresión, pero presenta un aumento en su asociación a la
membrana plasmática basolateral, mientras Cdc42 se encuentra disminuido tanto en
los niveles proteicos como en el reclutamiento hacia la superficie basolateral en
glándulas mutantes [68]. Estas GTPasas son activadas durante la citodiferenciación
de glándula submandibular de ratón normal, colaborando con E-cadherina y
β-catenina en el establecimiento de la polaridad celular y uniones intercelulares [69].
VIII. Importancia de los cultivos en tres dimensiones
Los tejidos y órganos son tridimensionales (3D), pero nuestra capacidad de
comprender su formación, función y fisiología frecuentemente proviene de estudios
en cultivos celulares bidimensionales (2D). Sin embargo, las células cultivadas en
plano difieren considerablemente de aquellas crecidas en un ambiente tridimensional
más fisiológico, en su morfología, estado diferenciado y en la naturaleza de las
interacciones que establecen entre ellas y con la matriz extracelular [70].
El paso del cultivo en monocapa a células en tres dimensiones (3D) es
motivado por la necesidad de trabajar con modelos celulares que imitan las funciones
de los tejidos, ya que funciones celulares esenciales presentes en los tejidos se
pierden en cultivos tradicionales en 2D, limitando su capacidad de predecir
respuestas celulares de los organismos [71].
Muchos modelos en 3D se han establecido a partir de células aisladas (por
ejemplo líneas de células, tejidos disgregados y células madre), donde una estrategia
ampliamente utilizada es propagar las células en cultivo y a continuación sembrarlas

11
dentro de una matriz 3D como células individuales. Estas matrices han sido
generadas a partir de moléculas purificadas como el colágeno tipo I, biomateriales
sintéticos e incluso matrices de proteínas extracelulares nativas [70]. Tanto el tipo
celular, como el microambiente de la matriz extracelular pueden influir
sustancialmente en los resultados experimentales obtenidos. Dentro de esto último,
tanto la composición como, la rigidez de la matriz extracelular que rodea a las células
tienen efectos importantes sobre la señalización y el comportamiento celular [72,73].
Para el crecimiento y diferenciación de células epiteliales, geles reconstituidos de un
extracto que contiene componentes de la lámina basal y factores de crecimiento
(denominado Matrigel, matriz EHS o Cultrex) son mucho más efectivos en el proceso
de diferenciación [74].
In vivo, las células acinares están en un contexto espacial tridimensional,
donde las señales provenientes de la lámina basal se integran con otras
provenientes de células vecinas, en un contexto muy distinto al establecido en
cultivos en plano, y se han descrito diferencias en la señalización y organización de
células crecidas en plano y en 3D [75].
Se ha desarrollado el cultivo de estructuras tipo acinos en células epiteliales
de distinto origen, que incluyen células derivadas de epitelio renal (MDCK) [31,76],
mamario (MCF10) [77] y también de epitelio de glándulas salivales (HSG) [78-80].
La organización del tipo acinar no es la misma en todos los tipos celulares, ya que
se obtienen estructuras esferoidales de diversos tamaños, donde la naturaleza
celular influye por ejemplo en el tamaño del lumen y en la morfología acinar. Este
tipo de cultivo permite estudiar fenómenos como el desarrollo acinar, la polarización
de las células y apoptosis durante la maduración acinar, entre otros. Además, la
utilización de una matriz extracelular que confiere a la célula epitelial las señales de
polarización, permite la organización de los ejes de polaridad.
Se han establecido líneas celulares derivadas desde biopsias de GSL de
pacientes con SS, cultivadas sobre matriz de colágeno que se han utilizado para
estudiar el SS, sin embargo, la forma de cultivo no asemeja lo que sucede
tridimensionalmente. La utilización del modelo de cultivo acinar de células HSG

12
como modelo del SS permite un aproximación experimental a lo que sucede en
acinos de pacientes. Considerando todos estos antecedentes que relacionan las alteraciones
en el fenotipo polarizado de acinos de GSL, con la desorganización de la lámina basal de acinos de pacientes con SS, planteamos que señales
derivadas de la integrina α6β1 y Rho GTPasas podrían estar jugando un papel
en estos procesos. Una lámina basal desorganizada puede gatillar señales incorrectas desencadenando el fenotipo alterado.

13
HIPÓTESIS
La pérdida de polaridad observada en las células epiteliales acinares de glándulas
salivales labiales de pacientes con síndrome de Sjögren puede explicarse por una
incorrecta señalización a través de la integrina α6β1 que resulta en cambios en los
niveles de activación de GTPasas de la familia Rho

14
OBJETIVO GENERAL
Demostrar que la alteración de la señalización de la integrina α6β1 modifica el
fenotipo polarizado de células epiteliales acinares en forma dependiente de GTPasas
de la familia Rho
OBJETIVOS ESPECÍFICOS
1. Definir la expresión y localización de las integrinas α6 y β1, así como la
activación de FAK en acinos aislados de GSL de individuos controles y de
pacientes con SS
2. Demostrar que el fenotipo polarizado en células derivadas de glándula salival
humana es controlado por la señalización de la integrina α6
a) Desarrollar un sistema de cultivo en 3D de células derivadas de glándula
salival que presenten un fenotipo polarizado, descrito como la expresión
asimétrica de proteínas de membrana plasmática y organelos como el
complejo de Golgi
b) Evaluar el control que ejerce la integrina α6 sobre el fenotipo polarizado de
células HSG cultivadas en 3D
3. Determinar la influencia de las GTPasas de la familia Rho en el establecimiento
del fenotipo polarizado en células HSG
a) Determinar los niveles de las GTPasas de la familia Rho activas en
condiciones de bloqueo de la integrina α6

15
MATERIALES Y MÉTODOS I. Materiales
1. Sales y reactivos
La totalidad de las sales, solventes orgánicos y reactivos generales utilizados
fueron adquiridos en las empresas Sigma (St. Louis, MO, EEUU), Merck (Darmstadt,
Germany), BioRad Laboratories (Hercules, CA, EEUU) y US Biological (Swampscott,
MA, EEUU). La proteína A-Sefarosa “nProtein A Sepharose 4 fast Flow” y Glutation
Sefarosa 4B son de Amersham Biosciences Corporation (Piscataway, NJ, EEUU). La
membrana de nitrocelulosa es de BioRad Laboratories (Hercules, CA, EEUU), la
membrana de PVDF- Immobilon-P es de Millipore (Billerica, MA, EEUU). El reactivo
de quimioluminiscencia “SuperSignal West Pico Chemiluminiscent substrate” es de
Pierce Biotechnology, Inc. (Rockford, IL, EEUU). Los medios y soluciones de cultivo
celular son de la compañía Gibco–Invitrogen (Carlsbad, CA, EEUU), y el suero fetal
bovino caracterizado es Hyclone (South Logan, UT, EEUU). El extracto de lámina
basal normal “Cultrex® Basement Membrane Extract without phenol red” es de
Trevigen (Gaithersburg, MD, EEUU).
2. Partidores
En la Tabla 1 se muestran las secuencias de cada una de las parejas de
partidores utilizados. Los partidores para cistatina (Cys), zonnula ocludens 1 (ZO1),
claudina 4 (Cla-4) y mucina 1 (MUC1) fueron diseñados considerando la secuencia
del mRNA disponible en la base de datos GenBank (NCBI). Utilizando el programa
Gene Runner versión 3.00 (Hasting Software Inc, NY, EEUU) se establecieron
parejas de partidores específicos que no forman estructuras secundarias ni
dimerizan. Para los genes de integrina α6 [81], integrina β4 [81], MUC5B [82], MUC7
[82], gliceraldehído 3-fosfato deshidrogenasa (GAPDH) [34] y amilasa [83] se
utilizaron partidores previamente descritos en la literatura. Todos los partidores
fueron sintetizados por Invitrogen (Carlsbad, CA, EEUU).

16
3. Anticuerpos
Los anticuerpos utilizados se indican en las Tabla 2 y 3, y se entregan los
datos correspondientes a la especie de procedencia, inmunógeno así como la
aplicación en la cual se utilizaron. Los anticuerpos son de las empresas: Santa Cruz
Biotechnology Inc. (Santa Cruz, CA, EEUU), BD Biosciences (San José, CA, EEUU),
MP Biomedicals (Solon, OH, EEUU), Abcam Inc, (Cambridge, MA, EEUU), Upstate
Biotechnology Inc. (Lake Placid, NY, EEUU), Covance Inc. (Princeton, NJ, EEUU),
Biocare Medical (Concord, CA, EEUU), Molecular Probes Invitrogen (Carlsbad, CA,
EEUU), AnaSpec (San José, CA) y Jackson Immunochemical Research Laboratory
(West Grove, PA, EEUU). El anticuerpo contra la integrina α6-GoH3 es un anticuerpo
no comercial generosamente donado por el Dr. Arnoud Sonnenberg (The
Netherlands Cancer Institute, Amsterdam, The Netherlands).

17
Tabla 1: Características de los partidores y tamaño del producto de amplificación esperado
Gen Número de acceso (NCBI) Partidores
Tamaño del producto de
amplificación (pb)
MUC5B
NM_002458 Directo : 5’ CTG CGA GAC CGA GGT CAA CAT C 3’
415 Reverso : 5’ TGG GCA GCA GGA GCA CGG AG 3’
MUC7
BC025688 Directo : 5’ CCA CAC CTA ATT CTT CCC CAA CTA C 3’
407 Reverso : 5’ CTG GCT TGT GGG ATA GAG GCA TT 3’
MUC1 NM_182741
Directo : 5’ TCT ACT CTG GTC CAC AAC GG 3’ 381
Reverso : 5’ ATT GTA CCA CCA CAG ATC C 3’
amilasa NM_004038
Directo : 5’ AAT TGA TCT GGG TGG TGA GC 3’ 474
Reverso : 5’ CTT ATT TGG CGC CAT CGA TG 3’
cistatina NM_001037160
Directo : 5’ CAT CAG CGA GTA TAA CAA G 3’ 525
Reverso : 5’ GAT GCT ACT GTT TAA TTG C 3’
ZO-1 NM_003257
Directo : 5’ CCA CCG GAG TCT GCC ATT AC 3’ 199
Reverso : 5’ CAG GCG AAA GGT AAG GGA C 3’
claudina 4 NM_001305
Directo : 5’ GGC TGC TTT GCT GCA ACT GTC 3’ 107
Reverso : 5’ GAG CCG TGG CAC CTT ACA CG 3’
GAPDH BT006893
Directo : 5’ ACA ACA GCC TCA AGA TCA TCA 3’ 670
Reverso : 5’ GTC CAG GGG TCT TAC TCC 3’ integrina α6
NM_000210 NM_001079818
Directo : 5’ CTAACGGAGTCTCACAACTC 3’ 817 y 701
Reverso : 5’ ACT CTG AAA TCA GTC CTC AG 3’
pb: pares de bases
18
Tabla 2: Características de los anticuerpos primarios
Anticuerpo Origen Inmunógeno Uso Empresa
Integrina α6 – GoH3 rata Epítope extracelular de Integrina α6 de origen humano IF, BLK No comercial (*)
Integrina α6 – BQ16 ratón Integrina α6 completa de origen humano WB Santa Cruz, SC-13542
Integrina β1 – 4B7R ratón Integrina β1 completa de origen humano IF Santa Cruz, SC-9970
Integrina β1 – clon 18 ratón Residuos 76-256 de Integrina β1 de origen humano WB, IPP BD Biosciences, 610467
Citoqueratina 18 – DC10 ratón Células PMC-42 (carcinoma mamario humano) IPP Santa Cruz, SC-6259
Laminina α1 – H300 conejo Residuos 1856-2099 de Laminina α1 de origen humano IPP Santa Cruz, SC-5582
Rho A – 26C4 ratón Residuos 120-150 de Rho A de origen humano WB Santa Cruz, SC-418
Rac 1 – clon 102 ratón Rac 1 completo de origen humano WB BD Biosciences, 610651
Cdc42 – clon 44 ratón Cdc42 completo de origen humano WB BD Biosciences, 610928
Actina – clon C4 ratón Actina de “molleja de pollo” WB MP Biomedicals, 69100
GAPDH conejo GAPDH completo aislado desde eritrocitos humanos WB Abcam, ab9485
FAK, C903 conejo Residuos 903-1052 de FAK de ratón WB Santa Cruz, SC-932
FAK P-Tyr397 conejo Residuos 392-399 de fosfo FAK humano conjugado a KLH WB Upstate, 07-012
Laminina conejo Laminina purificada de lámina basal de sarcoma de ratón EHS (laminina-111) IFI Sigma, L9393
Colágeno tipo IV, clon Col 94 ratón Colágeno tipo IV humano IFI Biocare Medical, CM112A
Gigantina conejo Residuos 1-469 de Gigantina de origen humano IFI Covance, PRB-114C
Claudina 4 ratón Péptido sintético derivado del C-terminal de Claudina 4 humana WB Zymed, 3E2C1
Ocludina conejo Proteína de fusión del C-terminal de ocludina WB Zymed, Z-T22
IF: inmunofluorescencia, WB: Westernblot, IPP: inmunoprecipitación, KLH: keyhole limpet hemocyanin, EHS: Engelbreth-Holm-Swarm
19
Tabla 3: Características de los anticuerpos secundarios
Anticuerpo Origen Uso Empresa Anti IgG (H+L) rata - Alexa fluor 488 cabra IFI Molecular Probes, A11006
Anti IgG (H+L) ratón - Alexa fluor 488 cabra IFI Molecular Probes, A11001
Anti IgG (H+L) conejo - Alexa fluor 488 cabra IFI Molecular Probes, A11008
Anti IgG (H+L) ratón - Alexa fluor 546 cabra IFI Molecular Probes, A11003
Anti IgG (H+L) conejo - Alexa fluor 546 cabra IFI Molecular Probes, A11010
Anti IgG (H+L) ratón - HyLiteFluor 555 cabra IFI AnaSpec, 28175-H555
Anti IgG (H+L) ratón - peroxidasa cabra WB Jackson, 115-035-003
Anti IgG (H+L) conejo - peroxidasa cabra WB Jackson, 111-035-003 IF: inmunofluorescencia, WB: Westernblot, H+L: cadenas pesadas (heavy) y livianas (light)

20
II. Diagnóstico de pacientes y obtención de GSL
1. Diagnóstico de pacientes
El protocolo de ingreso de los individuos al estudio se encuentra dentro del
proyecto Fondecyt 1050192 (Investigador principal, Dra. María Julieta González),
aprobado por el Comité de Ética de la Facultad de Medicina de la Universidad de
Chile. Los pacientes son diagnosticados por su médico tratante, Dr. Sergio Aguilera
Covarrubias, Reumatólogo de la Clínica Indisa y coinvestigador del proyecto
mencionado. Los pacientes con SS primario fueron diagnosticados según el criterio
de consenso Europeo-Americano [6]. Los controles corresponden a individuos que
consultaron al médico por sequedad oral u ocular, pero que no cumplen el criterio
diagnóstico para el SS.
2. Criterio para el diagnóstico del SS primario
Según el criterio de consenso Europeo - Americano [6], durante el diagnóstico
de SS primario se consideran los siguientes puntos:
a. Síntomas subjetivos de sequedad ocular
b. Síntomas subjetivos de sequedad oral
c. Síntomas objetivos de sequedad ocular (Test de Schrimer o Rosa de
Bengala)
d. Biopsia de GSL (Sialoadenitis focal con score de foco >1 por 4mm2 de tejido
glandular. Score de foco está definido como el número de focos linfocitarios
que contenga más de 50 linfocitos).
e. Flujo salival total no estimulado <1,5 mL en 15 minutos
f. Presencia de autoanticuerpos anti Ro (SSA), La (SSB) o ambos en el suero.
Según este criterio, un individuo es diagnosticado con SS primario si no
presenta otra enfermedad autoinmune asociada y además cumple una de las
siguientes condiciones:

21
• presenta 4 de los 6 puntos previamente nombrados, siempre y cuando el
diagnóstico histopatológico (d) o la serología (f) sean positivos
• presenta 3 de los 4 criterios objetivos (c, d, e, f) positivos
3. Características de los individuos del estudio
En esta tesis se utilizaron las muestras de distintos individuos según las
determinaciones a realizar, y en la Tabla 4 se describen las características
demográficas de los individuos, además del diagnóstico histopatológico de la biopsia
de GSL.
4. Biopsia de GSL
La obtención de GSL se realizó mediante una técnica operatoria previamente
descrita [84], donde bajo anestesia local infiltrativa se realizó una incisión de 1,5-2cm
de longitud en la cara interna del labio inferior, paralela al borde bermellón y lateral a
la línea media. Las glándulas obtenidas fueron congeladas inmediatamente en N2
líquido y mantenidas a -80ºC para la posterior extracción de proteínas y RNA total.
Otras glándulas fueron embebidas en OCT (Tissue-Tek OCT Compound) congeladas
en isopentano previamente enfriado en N2 para la obtención de cortes por
congelación. Otro grupo de glándulas fueron mantenidas a 4ºC para ser
inmediatamente disgregadas y obtener fracciones enriquecidas en acinos y ductos.
22
Tabla 4: Características demográficas y diagnóstico histopatológico
Controles Pacientes
Westernblot integrinas α6 y β1
1. (52 años, SD moderado)
2. (46 años, SD severo)
3. (26 años, SF leve)
4. (28 años, SF leve)
5. (54 años, SF leve)
6. (43 años, SF leve)
Inmunoprecipitación ---------------------------------pool glandular-------------------------
Inmunofluorescencia α6 y β1 1. (28 años, SD moderado) 2. (42 años, SF leve)
Niveles de FAK (Tyr397)
1. (50 años, SD leve)
2. (41 años, SD leve)
3. (52 años, SD moderado)
4. (55 años, SD moderado)
5. (52 años, SD moderado)
6. (57 años, SD moderado-severo)
7. (28 años, SD moderado)
8. (46 años, SD leve)
9. (26 años, SF leve)
10. (33 años, SF leve)
11. (33 años, SF leve)
12. (54 años, SF leve)
13. (54 años, SF leve)
14. (22 años, SF severo)
SD: sialoadenitis crónica difusa SF: sialoadenitis crónica focal leve (2-3 focos), moderada (4-6 focos), severa (>6 focos)

23
III. Métodos
1. Obtención de fracciones enriquecidas en acinos y ductos
La disgregación de GSL se realizó utilizando una metodología previamente
estandarizada en nuestro laboratorio que permite obtener fracciones glandulares
donde el componente acinar y ductal es mayoritario [26,85]. Se determinó mediante
RT-PCR un enriquecimiento de hasta un 80% en marcadores de células epiteliales,
siendo la contaminación con células inflamatorias directamente proporcional a la
cantidad de infiltrado observado en el análisis histopatológico de las glándulas [26].
La disgregación se realizó a partir de 5-7 GSL de cada individuo mantenidas a 4ºC
en medio de cultivo mínimo Dulbecco–DMEM (Gibco, Invitrogen) inmediatamente
después de su obtención. Las glándulas se cortaron en trozos de aproximadamente
2 mm de diámetro y se incubaron con 5 mL de una solución de colagenasa tipo IV
(2mg/mL en DMEM) durante 30 minutos a 37ºC con agitación constante. Luego, las
glándulas se disgregaron mecánicamente haciéndolas pasar por pipetas Pasteur de
2 mm de diámetro previamente siliconizadas (Sigmacote, Sigma). Para eliminar los
linfocitos, la suspensión se centrifugó a 1,9 ×g por 10 minutos a 4ºC, ya que a esta
velocidad de centrifugación los linfocitos se mantienen en suspensión. Se rescató el
pellet celular y luego se resuspendió en 5mL de solución de colagenasa tipo IV
fresca, y se reincubó por 45 minutos a 37ºC con agitación constante para continuar
la disgregación. Posteriormente, se realizó una segunda disgregación mecánica con
pipetas siliconizadas de diámetros decrecientes (1-0,5mm) hasta obtener una
suspensión homogénea de acinos y ductos. Esta suspensión se dejó decantar por 5
minutos y se centrifugó a 12,5 ×g a 4ºC, rescatándose el pellet de acinos y ductos, el
cual se lavó 5 veces con solución Hanks pH 7,4 centrifugándolo a 31×g a 4ºC. En
cada etapa de la disgregación se tomaron alícuotas de la suspensión y se analizaron
morfológicamente mediante microscopía de luz, a fin de detectar la cantidad de
linfocitos presentes. El pellet obtenido se utilizó para la extracción de proteínas.

24
2. Obtención de extractos proteicos
• Extractos proteicos de la fracción enriquecida en acinos y ductos de GSL: el pellet
obtenido se resuspendió en tampón RIPA (Tris-HCl 50mM pH 7,4, NaCl 150mM,
deoxicolato de sodio 1%, TX100 1%, SDS 0,1%, EDTA 10mM, benzamidina
10μg/mL, aprotinina 10μg/mL leupeptina 10μg/mL, PMSF 1mM, Na3VO4 1mM) y
se homogeneizó utilizando un homogenizador vidrio-vidrio, 3 ciclos rápidos de
congelación-descongelación y 3 pasajes por jeringa de tuberculina. Este
homogeneizado se centrifugó a 16000 xg durante 15 minutos a 4ºC y se rescató
la fracción soluble.
• Extractos proteicos de GSL totales: 1-2 GSL mantenidas a -80ºC se congelaron
en N2 líquido y luego se les agregó tampón RIPA y se pulverizaron en un mortero
de porcelana. Posteriormente, la homogenización y el rescate de la fracción
soluble se realizó de manera semejante a la descrita previamente.
• Extractos proteicos de células HSG cultivadas en plano: Las células se lavaron
una vez con PBS y luego se les agregó tampón RIPA y las células se despegaron
con una espátula. Finalmente, el homogeneizado se centrifugó a 16000 xg
durante 15 minutos a 4ºC y se rescató la fracción soluble.
• Extractos proteicos de acinos de células HSG: Se eliminó el medio de cultivo y los
acinos se resuspendieron en tampón RIPA y se homogenizaron por pipeteo.
Finalmente, el homogeneizado se centrifugó a 16000 xg durante 15 minutos a
4ºC y se rescató la fracción soluble. Es importante destacar que en este extracto
está incluido el extracto de lámina basal utilizado para el cultivo.
En todos los casos, se cuantificaron las proteínas de acuerdo al procedimiento
descrito por Bradford [86].
3. Cuantificación de proteínas
Se utilizó el método descrito por Bradford [86], que consiste en la incubación
de una alícuota del extracto proteico con el reactivo de Bradford (Biorad) durante 2
minutos, a temperatura ambiente y la posterior detección del producto coloreado
formado midiendo la absorbancia de la muestra a 595nm. En paralelo se realizó una
curva estándar de seroalbúmina de bovino (BSA) con un rango de concentración

25
proteica entre 12 y 0,75μg/mL. Las mediciones se realizaron en duplicado tanto para
la muestra como para la curva estándar.
4. Electroforesis en geles de poliacrilamida
Las proteínas se separaron mediante geles SDS-PAGE en base al método
descrito por Laemmli [87]. Los extractos proteicos (25-100μg) se mezclaron en una
relación 1:1 con tampón de carga 2x (Tris 125mM pH 6,8, SDS 4,6%, glicerol 20%,
β-mercaptoetanol 10% y azul de bromofenol 0,02%) y se hirvieron por 5 minutos
para denaturar y reducir las proteínas. El caso particular de la detección de la
integrina α6 se realizó en condiciones no reductoras, utilizando un tampón de carga
sin β-mercaptoetanol. La separación de proteínas se realizó en un minigel de
poliacrilamida (8-15%) en tampón de corrida (Glicina 192mM, Tris 25mM, SDS 0,1%)
a 25 mA constantes. El gel se tiñó con solución de tinción (Azul de Coomassie R-250
0.25%, isopropanol 45%, ácido acético glacial 8%) durante 30 minutos y se destiñó el
exceso con una solución de ácido acético 13% e isopropanol 20%. Alternativamente
las proteínas del gel se transfirieron a una membrana sintética para la realización de
un Westernblot (WB).
5. Westernblot
Las proteínas separadas en geles SDS-PAGE se electrotransfirieron a una
membrana de nitrocelulosa (poro 0,45μm, Biorad) o PVDF (poro 0,45μm, Immobilon-
P-Millipore Billerica, MA, EEUU) aplicando una corriente de 70mA constantes
durante 15 horas en tampón de transferencia (Glicina 154mM, Tris 25mM, metanol
20%) a 4ºC. Se comprobó la correcta transferencia de las proteínas a la membrana
tiñéndola durante 5 minutos con una solución de rojo Ponceau (Ponceau S 0,1% en
ácido acético 0,5%). Posteriormente se bloquearon los sitios inespecíficos en la
membrana incubándola durante 1 hora a temperatura ambiente en solución de
bloqueo (leche descremada 6-10% o BSA libre de proteasas e inmunoglobulinas 5%
en tampón TBST (Tris 10mM pH 7,5, NaCl 100mM, Tween 20 0,1%)) a temperatura
ambiente. Luego, la membrana se incubó con los anticuerpos primarios diluidos en
solución de bloqueo a temperatura ambiente o toda la noche a 4ºC. En la Tabla 5 se

26
muestran los detalles del bloqueo y las diluciones de anticuerpo utilizadas para las
distintas proteínas detectadas. Luego, se eliminó el exceso de anticuerpo primario
realizando 5 lavados de 5 minutos en tampón TBST. Posteriormente, se incubó la
membrana con un anticuerpo secundario conjugado a peroxidasa por 1 hora a
temperatura ambiente, eliminando luego el exceso de anticuerpo secundario con 5
lavados de 5 minutos en tampón TBST. Finalmente, se detectaron las proteínas por
el método de detección de quimioluminiscencia (Pierce) siguiendo las instrucciones
del fabricante.

27
Tabla 5: Condiciones de Westernblot
Proteína Bloqueo Anticuerpo primario Anticuerpo secundario
Rho A Leche descremada 6%, 1h TA Rho A, 26C4, 1:250 O/N a 4ºC
anti - IgG de ratón – peroxidasa, 1:5000,
1h TA
Rac 1 Leche descremada 6%, 1h TA Rac 1, clon 102, 1:250 O/N a 4ºC
Cdc42 Leche descremada 6%, 1h TA Cdc42, clon 44, 1:250 O/N a 4ºC
Claudina 4 Leche descremada 6%, 1h TA Claudina 4, 1:750 1h a TA
GAPDH Leche descremada 6%, 1h TA GAPDH, 1:5000 O/N a 4ºC
anti - IgG de conejo – peroxidasa,
1:5000, 1h TA
FAK Leche descremada 6%, 1h TA FAK, C903, 1:1000 4h a TA
FAK Tyr397 BSA 5 %, 3h TA FAK P-Tyr397, 1:1000 O/N a 4ºC
Ocludina Leche descremada 6%, 1h TA Ocludina 1:500 1h a TA
Integrina α6 (*) Leche descremada 10% , 1h TA Integrina α6 –BQ16, 1:2000 1h TA anti - IgG de ratón – peroxidasa,
1:10000, 1h TA Integrina β1 Leche descremada 6%, 1h TA Integrina β1 – clon 18 , 1:3500 1h a TA
Actina Leche descremada 6%, 1h TA Actina-clon C4, 1:20000 1h a TA
TA: temperatura ambiente, O/N: toda la noche, h: hora. (*) Realizado en condiciones no reductoras

28
6. Inmunoprecipitación
Previo a la inmunoprecipitación, el extracto proteico glandular se pre-clarificó
para eliminar las proteínas que se unen inespecíficamente a la proteína A. Para esto,
500μg de extracto proteico se incubaron con 50μL de proteína A-Sefarosa 50% slurry
(Amersham) en tampón de inmunoprecipitación (TX100 1%, NaCl 150mM, Tris-HCl
10mM pH 7,4, EDTA 1mM, benzamidina 10μg/mL, aprotinina 10μg/mL, leupeptina
10μg/mL, PMSF 1mM) durante 30 minutos a 4ºC con agitación constante. Luego, se
rescató el extracto proteico pre-clarificado y se incubó con 2μg de anticuerpo
inmunoprecipitante (Integrina α6–BQ16, Citoqueratina 18, cadena α1 de laminina-
111) en tampón de inmunoprecipitación durante toda la noche a 4ºC con agitación
constante. Posteriormente, se rescataron los inmunocomplejos agregando 50μl de
proteína A-Sefarosa 50% slurry e incubando durante 1 hora a 4ºC con agitación.
Luego se rescatan los correspondientes perlas de proteína-A Sefarosa tras
centrifugación y se realizan 3 lavados con tampón de inmunoprecipitación.
Finalmente, se elimina el máximo de solución y se resuspenden las perlas en 40μL
de tampón de carga 2x (Tris 125mM pH 6,8, SDS 4,6%, glicerol 20%, β-
mercaptoetanol 10% y azul de bromofenol 0,02%), se hierven durante 5 minutos y se
analiza por WB la presencia de la Integrina β1 en los inmunocomplejos.
7. Inmunofluorescencia en cortes por congelación de GSL
Se obtuvieron criosecciones de 5μm de las GSL embebidas en OCT, las
cuales fueron adheridas a portaobjetos silanizados y mantenidas a -80ºC. Previo a
comenzar la inmunofluorescencia, las criosecciones se estabilizaron a temperatura
ambiente por 20 minutos y posteriormente se fijaron 20 minutos en paraformaldehído
(PFA) 4% en tampón fosfato salino - PBS (Na2HPO4.2H2O 38mM, NaH2PO4.2H2O
17mM). Los cortes se lavaron 3 veces con PBS durante 5 minutos y luego se
bloquearon los sitios de unión inespecífica con caseína al 0.25% en PBS, incubando
a temperatura ambiente por 45 minutos. Luego se incubó con el anticuerpo primario
(integrina α6–GoH3 dilución 1:5 e integrina β1-4B7R dilución 1:15 en PBS) durante 2
horas a temperatura ambiente. Se realizaron 3 lavados de 5 minutos con PBS y se
incubaron durante 45 minutos con los anticuerpos secundarios acoplados a

29
fluoróforos (anti IgG de rata acoplado a Alexa 488 y anti IgG de ratón acoplado a
HyLiteFluor 555, diluidas 1:200 en PBS). Finalmente, se realizaron 3 lavados de 5
minutos con PBS, 1 con H2O destilada y se montaron los cortes en medio de montaje
Mowiol. Las muestras se observaron en el microscopio confocal (LSM-140 Axiovert-
10.0. Zeiss, CESAT, ICBM, Facultad de Medicina, Universidad de Chile), realizando
cortes confocales de 0.8 μm de grosor.
8. Cultivo de células HSG en plano
Las células HSG se obtuvieron del laboratorio del Dr. Bruce Baum (NIDCR,
NIH, EEUU) y son de origen epitelial transformadas y aisladas desde ductos
intercalares de glándula submandibular humana irradiada [88,89]. Estas células
crecen en medio de cultivo DMEM:Ham’s F12 (1:1) (DMEM-F12), suero bovino fetal
(SBF) 5%, penicilina 100U/mL y estreptomicina 100μg/mL a 37ºC en medio húmedo
con 5% de CO2. El pasaje de las células se realizó una vez por semana, donde las
células se lavaron con PBS (pH 7,2, sin CaCl2 ni MgCl2) y luego se incubaron con
Tripsina-EDTA 0,05% durante 1 minuto a 37ºC, y finalmente se resuspendieron las
células en medio completo. Para el conteo de las células luego de la trispsinización,
éstas se tiñen con el colorante vital azul de tripán, y luego se cuentan en una cámara
de Neubauer.
9. Cultivo de acinos de células HSG en 3D
Se desarrolló un sistema de cultivo en tres dimensiones (3D) a partir de la
metodología previamente descrita para células de mama [77], que se adaptó para las
células HSG. Para esto se utilizó una preparación soluble de lámina basal extraída
de sarcoma de ratón Engelbreth-Holm-Swarm (EHS), rico en proteínas de matriz
extracelular. En la Figura 1 se muestra un esquema general de la metodología
utilizada para este tipo de cultivo. En un portaobjetos con 8 pocillos (Labtek Chamber
Slide System, Nunc, Rochester, NY, EEUU) se genera una base con 35μL de
matrigel 10mg/mL (Cultrex Trevigen Inc., Maryland, MD, EEUU) disuelto en DMEM-
F12 con penicilina 100U/mL y estreptomicina 100μg/mL. Esta solución se incubó 1
hora a 37oC para que gelifique, y luego sobre este gel se sembraron 6250 células por

30
pocillo, las cuales se resuspendieron en 400μL de matrigel 0,3mg/mL disuelto en
DMEM-F12 con 5% de SBF, penicilina 100U/mL y estreptomicina 100μg/mL. Las
células se crecieron a 37ºC con 5 % de CO2 y cada 2-3 días se realizó un cambio de
medio de cultivo. Inicialmente las células se localizan sobre el matrigel y a medida
que pasan las horas se van diferenciando y pasando a formar estructuras tipo acinos,
los cuales podrían estar sobre o sumergidos en el matrigel. Estos acinos fueron
utilizados posteriormente para la realización de los ensayos de bloqueo, extracción
de proteínas totales para WB, extracción de RNA para RT-PCR, inmunofluorescencia
y análisis ultraestructural por microscopía electrónica de transmisión (MET). En
ciertas condiciones de cultivo se agregaron EGF 10ng/mL, Hidrocortisona 0,4μg/mL,
Insulina 5μg/mL y Extracto de pituitaria 25μg/mL al medio de cultivo, utilizando la
misma metodología descrita previamente.

31
Células HSG cultivadas en plano
Tripsinización
6250 Células libres
• Extracción de proteínas WB • Extracción de RNA RT-PCR • Inmunofluorescencia • Análisis MET • Determinación de Rho GTPasas
Matrigel 10mg/mL Matrigel 0,3mg/mL
Figura 1: Esquema explicativo del cultivo en 3D de células HSG: Las células HSG se mantienen en plano crecidas en medio de cultivo DMEM-F12 con 5% de SBF, penicilina 100U/mL y estreptomicina 100μg/mL a 37ºC en medio húmedo con 5% de CO2. En un portaobjetos con 8 pocillos (Labtek Chamber Slide System, Nunc, Rochester, NY, EEUU) se genera una base con 35μL de matrigel 10mg/mL (Cultrex Trevigen Inc., Maryland, MD, EEUU) disuelto en DMEM-F12 sin suero. Esta solución se incubó 1 hora a 37oC para que gelifique, y luego sobre este gel se sembraron 6250 células por pocillo, las cuales se resuspendieron en 400μL de matrigel 0,3mg/mL disuelto en DMEM-F12 completo. Las células se crecieron a 37ºC con 5 % de CO2 y cada 2 días se realizó un cambio de medio de cultivo. Estos acinos fueron utilizados posteriormente para la realización de los ensayos de bloqueo, extracción de proteínas totales para WB, extracción de RNA para RT-PCR, inmunofluorescencia y análisis ultraestructural por microscopía electrónica de transmisión.

32
10. Inmunofluorescencia en acinos de células HSG cultivados en matrigel
La reacción se realizó directamente en los portaobjetos con pocillos, sin sacar
los acinos del matrigel, siguiendo una metodología descrita previamente [77]. Se
eliminó el medio de cultivo y los acinos se fijaron con PFA 2% en PBS por 20
minutos a temperatura ambiente o acetona enfriada a -20ºC por 5 minutos. Luego las
células se permeabilizaron con TX-100 0.5% en PBS durante 10 minutos a 4ºC. Los
acinos se lavaron con tampón PBS con Glicina 100mM 3 veces por 10 minutos y
luego se bloquearon los sitios de unión inespecífica con suero de cabra 10% en
tampón IF (NaN3 7,7mM, BSA 0,1%, TX100 0,2%, Tween20 0,05%) durante 1,5
horas a temperatura ambiente. Luego, se incubó con el anticuerpo primario diluido en
la solución de bloqueo toda la noche a temperatura ambiente (Integrina α6-GoH3
1:25, Integrina β1-4B7R 1:25, Laminina 1:25, Colágeno tipo IV 1:50, Gigantina
1:500). Para determinar F-actina, se utilizó Faloidina acoplada a TRITC (Sigma),
diluida 1:50 y se incubó de manera semejante a los anticuerpos primarios. Se
realizaron 3 lavados de 20 minutos cada uno con tampón IF y luego se incubó,
durante 45 minutos con los anticuerpos secundarios respectivos acoplados a
fluoróforos y diluidos en solución de bloqueo. Posteriormente, se realizaron 3 lavados
de 20 minutos con tampón IF y se tiñeron los núcleos durante 15 minutos con una
solución de yoduro de propidio 1μg/mL y RNAsaA 1μg/mL en H2O, a temperatura
ambiente. Finalmente, se realizó 3 lavados de 10 minutos con PBS, 1 con H2O
destilada y se montaron los cortes en Mowiol. Las muestras se analizaron en
microscopio confocal (LSM-140 Axiovert-10.0. Zeiss, CESAT, ICBM, Facultad de
Medicina, Universidad de Chile), realizando cortes confocales de 0.8μm de grosor.
11. Microscopía electrónica de transmisión de acinos de HSG
Acinos de 3 y 5 días de cultivo se lavaron 2 veces con tampón cacodilato de
sodio 0,1M pH 7,4 y se fijaron durante 3 horas con glutaraldehído 3% en tampón
cacodilato a temperatura ambiente. Luego, se realizaron 3 lavados de 10 minutos
con tampón cacodilato y se post-fijaron en una solución de OsO4 1,5% en tampón
cacodilato durante 30 minutos. Posteriormente, los acinos se incubaron en una
solución de tiocarbohidrazida (TCH) 1% durante 5 minutos y nuevamente se

33
incubaron en la solución OsO4 1,5% en tampón cacodilato durante 3 minutos. El
empleo de TCH en la postfijación se basa en un protocolo descrito previamente y
permite aumentar el contraste de las membranas celulares [90]. Luego se lavaron los
acinos durante 30 minutos con cambios sucesivos de tampón cacodilato. Finalmente,
se deshidrató la muestra con concentraciones crecientes de etanol (30%, 50%, 70%,
96%, y 100%), durante 7 minutos cada uno y para la incubación con etanol 100% se
realizaron 3 cambios. La impregnación se realizó con resina Epoxy (Epon-812,
Electron Microscopy Sciences, EEUU) utilizando una mezcla 1:1 con etanol 100%
durante 1,5 horas, con varios cambios. Luego se agregó la resina 100% y se dejó a
37ºC durante toda la noche, polimerizándose posteriormente a 70ºC por 48 horas.
Las secciones semifinas de 1μm de grosor se realizaron en un ultramicrotómo
(Sorvall Poter-Blum, modelo MT2-B) y luego fueron teñidas con azul de toluidina
preparado en borato de sodio 1% con el propósito de controlar si en las secciones
estaban presentes los acinos. Posteriormente, se realizaron secciones finas de
600Aº las cuales se montaron en grillas de 200 mesh y se contrastaron
sucesivamente con acetato de uranilo alcohólico 4% y citrato de plomo 0,4%, durante
1 y 5 minutos, respectivamente, a temperatura ambiente. Las secciones contrastadas
fueron observadas en un microscopio electrónico de transmisión, marca Zeiss,
modelo EM-109. Este microscopio cuenta con un sistema de captación de imágenes,
empleando una cámara fotográfica analógica para negativos de 35mm y de 60mm.
La resolución del equipo es de 10A° y el poder máximo de magnificación es de
250.000x.
12. Extracción de RNA total
La extracción de RNA total se realizó empleando el método de extracción por
afinidad en columnas de silica-gel RNeasy, acoplada a la digestión de DNA
genómico en columnas (Qiagen, Stanford, EEUU), según las indicaciones del
fabricante. En el caso de GSL, 1-2 GSL se disgregaron en tampón de lisis RLT
utilizando un homogeneizador cónico vidrio-vidrio. Para completar el proceso, el
producto de la lisis se pasó por las columnas de homogeneización por exclusión
QIAshredder (Qiagen, Stanford, EEUU), centrifugando durante 2 minutos a 13.000

34
×g. Acinos de células HSG provenientes de 12 pocillos de cultivo, se utilizaron para
la extracción de RNA. Se eliminó el medio de cultivo y se resuspendieron los acinos
(incluido el matrigel) en 600μl de tampón de lisis RLT. Una placa de 10cm de
diámetro de células HSG crecidas en plano se resuspendió en 600μl de tampón RLT.
En los tres casos, al homogeneizado se agregó etanol al 70%, que provee las
condiciones de unión selectiva del RNA a la columna. Esta solución se depositó en
las columnas RNeasy y se centrífugó durante 15 segundos a 13.000 ×g a
temperatura ambiente. Las columnas se lavaron una vez con 700μL de tampón RW1,
centrifugando durante 15 segundos a 13.000 ×g. Luego se realizaron dos lavados
con 500μL de tampón RPE (etanol 80%, composición protegida por patente No. 5,
234,809, EEUU), centrifugando a 13.000 ×g durante 15 segundos y 3 minutos,
respectivamente. Finalmente, el RNA se eluyó con 30μL de agua libre de nucleasas.
La concentración y pureza del RNA se determinó calculando la razón de absorbancia
de la muestra a 260 y 280 nm y realizando un PCR directamente sobre el RNA. Se
emplearon muestras de RNA total con una relación de absorbancia 260/280 de 1,8 a
2,0 y que no generaran productos al amplificarlas por PCR utilizando partidores para
GAPDH. La concentración del RNA se determinó empleando la siguiente fórmula:
[RNA] = 200 × (Absorbancia 260 nm) × 40μg/mL.
13. Electroforesis de RNA
Para verificar la calidad del RNA extraído y descartar aquellas preparaciones
que presentaran degradación, se realizaron geles denaturantes de RNA (agarosa-
formaldehído). 1μg de RNA total se diluyó en tampón de carga de RNA 6X (azul de
bromofenol 0,04% p/v, xileno cyanol 0,04% p/v, formaldehído 6% formamida 40%,
glicerol 3%, bromuro de etidio 0,1μg/μL, en tampón MOPS (MOPS 0,2M pH 8,0,
acetato de sodio 0,05M, EDTA 1mM, formaldehído 8%) se analizó en geles de
agarosa 1,5% p/v (Biotools B & M Labs, S.A. Madrid, España) en tampón MOPS La
separación electroforética se realizó a 75mA durante 45 minutos. Los geles se
fotografiaron sobre un transiluminador UV, empleando una cámara digital

35
14. Síntesis de cDNA
El cDNA se obtuvo transcribiendo 1μg de RNA total utilizando partidores de
oligos(dT) y random hexámeros (Promega Co., Madison, Wi, EEUU) con el sistema
comercial de transcripción reversa Superscript II (GibcoBRL, Invitrogen, CA, EEUU),
de acuerdo a las especificaciones del fabricante. La reacción de transcripción reversa
contiene 1,25mM de cada desoxinucleótido: dATP, dTTP, dCTP, dGTP (Promega
Co., Madison, Wi, EEUU), MgCl2 3mM, partidores oligos (dT) 1μM, partidores random
hexámeros 1μM, RNAseout 2U/μL (GibcoBRL, CA, EEUU) y transcriptasa reversa
Superscript II 10U/μL (GibcoBRL, CA, EEUU) en tampón de reacción (Tris-HCl 50mM
pH 8,3, KCl 75mM, MgCl2 3mM), el volumen final de reacción fue de 20μL. De esta
reacción de transcripción se utilizó 1μL para estandarizar las condiciones de los PCR
para cada par de partidores analizados.
15. PCR
La reacción de PCR se realizó en un volumen final de 25μL, conteniendo Taq-
DNA polimerasa 0,05U/μL (Invitrogen, CA, EEUU), desoxinucleótidos 0,2mM cada
uno (Promega, Madison, Wi, EEUU), MgCl2 1,5mM, 1μL de cDNA en tampón de
reacción (Tris-HCl 10mM pH 8,3, KCl 50mM). Las condiciones del PCR y los
partidores utilizados se detallan en las Tablas 1 y 5.
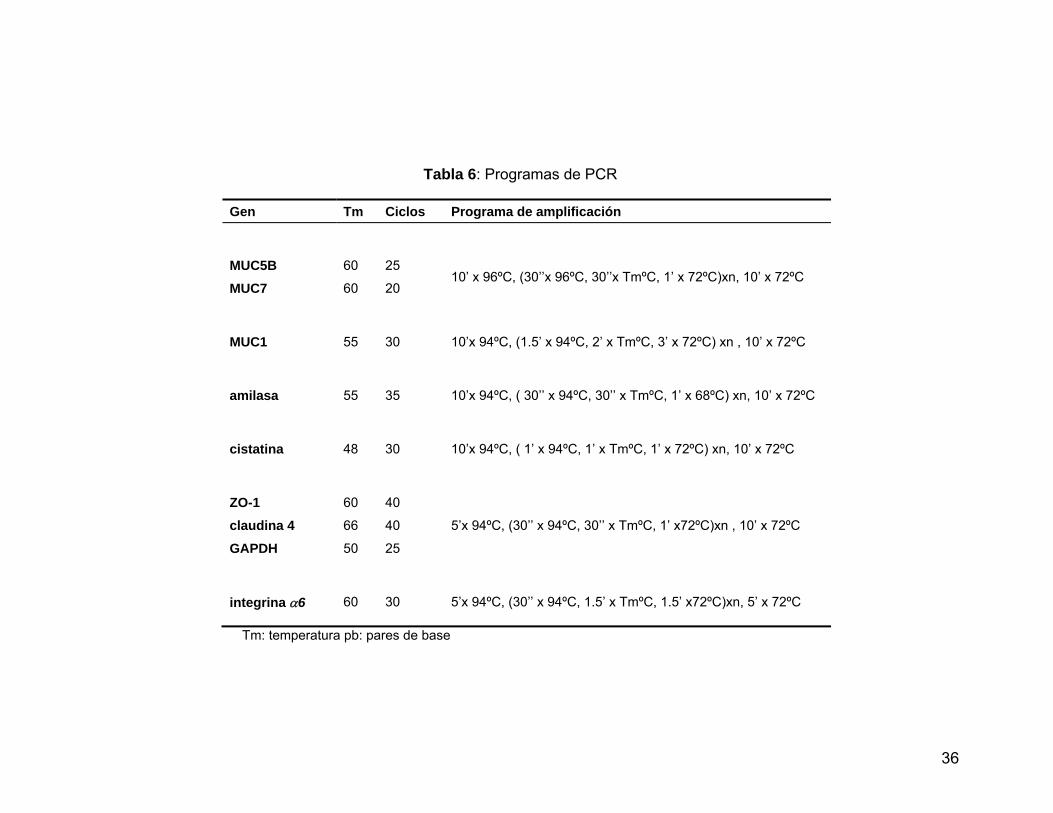
36
Tabla 6: Programas de PCR
Gen Tm Ciclos Programa de amplificación
MUC5B 60 25 10’ x 96ºC, (30’’x 96ºC, 30’’x TmºC, 1’ x 72ºC)xn, 10’ x 72ºC
MUC7 60 20
MUC1 55 30 10’x 94ºC, (1.5’ x 94ºC, 2’ x TmºC, 3’ x 72ºC) xn , 10’ x 72ºC
amilasa 55 35 10’x 94ºC, ( 30’’ x 94ºC, 30’’ x TmºC, 1’ x 68ºC) xn, 10’ x 72ºC
cistatina 48 30 10’x 94ºC, ( 1’ x 94ºC, 1’ x TmºC, 1’ x 72ºC) xn, 10’ x 72ºC
ZO-1 60 40
5’x 94ºC, (30’’ x 94ºC, 30’’ x TmºC, 1’ x72ºC)xn , 10’ x 72ºC claudina 4 66 40
GAPDH 50 25
integrina α6 60 30 5’x 94ºC, (30’’ x 94ºC, 1.5’ x TmºC, 1.5’ x72ºC)xn, 5’ x 72ºC
Tm: temperatura pb: pares de base

37
16. Tratamiento con anticuerpo bloqueante
Los acinos de células HSG se cultivaron durante 2 días y luego se les agregó
el anticuerpo bloqueante de la integrina α6 GoH3 (0,1, 1, 3 o 5μg/mL) disuelto en
medio de cultivo completo con matrigel 0,3mg/mL (ver sección 9 métodos), durante
los distintos tiempos de incubación a 37ºC. En caso del bloqueo pulsado, una vez
retirado el medio de bloqueo, este se reemplazó por medio de cultivo completo con
matrigel 0,3mg/mL. Como control se utilizó IgG de rata purificada y disuelta en la
misma solución y concentración. Posteriormente, se utilizaron estos acinos para
realizar las inmunofluorescencias.
17. Expresión y purificación de las proteínas RBD-GST y PBD-GST
Se utilizaron los plasmidios pGEX_PBD-GST y pGEX_RBD-GST, que
codifican para el dominio de interacción a Rac y Cdc42 de PAK1 fusionado a GST
(PBD-GST) y el dominio de interacción a RhoA de Rhotekina fusionado a GST (RBD-
GST), respectivamente. En ambos plasmidios la expresión de las proteínas de fusión
está controlada por el promotor lac y codifican para resistencia a ampicilina. El
protocolo de expresión y purificación es igual para ambas proteínas. Una colonia
aislada de la cepa de bacterias E. coli (XL1Blue) transformada con el plasmidio
pGEX_PBD-GST o pGEX_RBD-GST se creció en 10mL de medio Luria Bertani
(Peptona 1%, extracto de levadura 0.5%, NaCl 1%) con ampicilina (100μg/mL),
durante toda la noche a 37ºC. Luego se inoculó 400mL de LB-ampicilina con 8mL de
cada cultivo de toda la noche, y se creció a 37ºC hasta llegar a una absorbancia
cercana a 0,2 medida a 600nm. Inmediatamente alcanzada esta absorbancia, se
indujo la expresión de la proteína de fusión agregando al cultivo IPTG 0,3mM y
creciendo durante 17 horas a 25ºC. Las bacterias se centrifugaron por 15 minutos a
4000 xg y el pellet bacteriano se congeló a -80ºC. Las proteínas de fusión se
purificaron mediante afinidad utilizando una resina de Glutation Sefarosa 4B
(Amersham). El pellet bacteriano se resuspendió en 10 mL de tampón de lisis a 4ºC
(Tris-HCl 50mM pH 7,4, NaCl 50mM, MgCl2 5mM, DTT 1mM, PMSF 1mM,
benzamidina 10ug/mL, aprotinina 10ug/mL, leupeptina 10ug/mL y Tritón X-100 1%).
La suspensión se sonicó 10 veces con pulsos de 20 segundos y se centrifugó a

38
16000 xg por 15 minutos a 4ºC, rescatándose la fracción soluble. Posteriormente, el
sobrenadante se incubó con agitación durante 1 hora a 4ºC con 1mL de resina
Glutatión Sefarosa 4B (50% slurry) previamente equilibrada en tampón de lisis. Se
lavaron las perlas 3 veces con tampón de lisis y luego 3 veces con tampón de lisis
sin Tritón X-100, separando en cada oportunidad las perlas por centrifugación
durante 1 minuto a 1300 xg. Finalmente, las perlas de Glutatión Sefarosa unidas a
las proteínas de fusión se mezclaron con glicerol hasta una concentración del 50%,
se congelaron en N2 y se guardaron alicuotadas a -80ºC. Una alícuota de cada
proteína se utilizó para determinar la concentración de proteína de fusión mediante el
método de Bradford y determinar la pureza del extracto analizándose en un gel de
poliacrilamida.
18. Determinación de los niveles de RhoA, Cdc42 y Rac1 activos
Se realizó una precipitación por afinidad de RhoA-GTP, Cdc42-GTP y Rac1-
GTP, utilizando las proteínas de fusión RBD-GST y PBD-GST asociadas a partículas
de Glutation Sefarosa 4B. Para células HSG crecidas en plano, una placa de 5cm de
diámetro crecida hasta confluencia, se lisó en 3mL de tampón A (Tris 50mM pH 7,6,
NaCl 500mM, SDS 0.1%, DOC 0.5%, Tritón X-100 1%, MgCl2 0.5mM, PMSF 1mM,
benzamidina 10μg/mL, aprotinina 10μg/mL y leupeptina 10μg/mL). Esta suspensión
se centrifugó a 16000 xg por 10 minutos a 4ºC, rescatándose la fracción soluble.
500μL de esta fracción se incubaron con 30μg de RBD-GST o PBD-GST y se
incubaron durante 1 hora con agitación a 4ºC. Posteriormente, se decantaron las
perlas por centrifugación y se lavaron 2 veces con tampón B (Tris 50mM pH 7,6,
NaCl 150mM, Tritón X-100 1%, MgCl2 0.5mM, PMSF 1mM, benzamidina 10μg/mL,
aprotinina 10μg/mL y leupeptina 10μg/mL), se recuperaron las perlas y se mezclaron
con 30 μL de tampón de carga de proteínas 2x, se hirvieron y se corrieron en un gel
de poliacrilamida al 14% en condiciones reductoras. Las proteínas unidas se
separarán en gel de poliacrilamida y se detectarán por Westernblot, comparándose
con los niveles totales de Rho GTPasas y actina presentes en 50 μL de la fracción
soluble. Para los acinos de células HSG, se utilizaron 4 pocillos de cultivo para
obtener una determinación. Previo a la obtención de los extractos, se realizaron los

39
ensayos de bloqueo. Se eliminó el medio de cultivo y se agregó 350μL de medio de
cultivo completo (0,3mg/mL de matrigel) con 3μg/mL de GoH3 o IgG de rata como
control, y se incubó durante 30 minutos a 37ºC. Luego se eliminó el medio de
incubación y los 4 pocillos de cultivo se resuspendieron en 500μL de tampón A. La
medición de la actividad de Rho GTPasas se realizó de manera semejante a lo
descrito para los 500μL de extracto de células HSG crecidas en plano.
19. Análisis estadístico
El análisis estadístico de los resultados se realizó utilizando el programa
GraphPad Prism 4.02 (GraphPad Software Inc., San Diego, CA, EEUU). Se usó el
test no paramétrico de Mann Whitney para analizar la significancia estadística
(p<0,05) de los resultados.

40
RESULTADOS
Los resultados de esta tesis incluyen datos obtenidos en dos modelos de
estudio distintos. El primero corresponde a un estudio descriptivo en biopsias de
glándulas salivales labiales humanas, de individuos controles y de pacientes con SS,
donde se analizó la expresión y localización in vivo de la integrina α6β1 y de señales
río abajo de integrinas. El segundo grupo de resultados se obtuvieron de un modelo
in vitro de cultivo de células derivadas de glándula salival humana, HSG. En este
sistema, se implementó y caracterizó un cultivo celular en tres dimensiones usando
extractos proteicos de lámina basal. Este modelo permitió estudiar la relación de la
integrina α6 con la organización acinar polarizada y con la activación de GTPasas de
la familia Rho. A continuación se presentan los resultados agrupados de acuerdo a
estos dos modelos utilizados.

41
I. Análisis de la expresión y localización de las integrinas α6 y β1 en glándulas
salivales labiales de controles y pacientes con síndrome de Sjögren
1. Niveles proteicos de las integrinas α6 y β1
Estudios realizados en nuestro laboratorio muestran que las integrinas α6 y β1
se expresan en extractos totales de GSL [44]. En estos extractos, además de las
células del parénquima glandular, están representadas todas las células estromales
e infiltrantes. Como se ha descrito que la integrina α6β1 es el mayor receptor para
laminina en linfocitos T [91], quisimos evaluar la expresión cualitativa de las
integrinas α6 y β1 de forma particular en el componente epitelial (acinos y ductos), en
un número reducido de individuos controles (n=2) y pacientes con SS (n=4).
Mediante disgregación enzimática y mecánica de biopsias de GSL, se obtuvieron
fracciones enriquecidas en componente epitelial donde se identificó la presencia de
las integrinas α6 y β1.
Utilizando un anticuerpo dirigido contra el dominio extracelular de la integrina
β1, se detectaron 2 formas de la integrina β1 de masa molecular estimada en 110 y
140 kDa. La forma de mayor movilidad electroforética podría corresponder a una
forma inmadura parcialmente glicosilada de la integrina β1 [92] (Figura 2A). En todos
los individuos se detectó la integrina β1 de menor movilidad electroforética, con una
intensidad equivalente en los distintos individuos, mientras que la forma de mayor
movilidad sólo se encontró en los pacientes con SS, aunque en muy bajos niveles
(Figura 2A, 3-6). Esta diferencia podría relacionarse con alteraciones en algunos
procesos de modificación postraduccional descritos previamente en las glándulas de
estos pacientes [93].
La integrina α6 presentó una movilidad electroforética de alrededor de 180
kDa (Figura 2A). En los pacientes estudiados hay un comportamiento variable de
expresión de esta integrina, ya que dos pacientes presentan una expresión similar a
los controles (Figura 2A, 4 y 5) y en los otros dos, la expresión está disminuida
(Figura 2A, 3 y 6). Un comportamiento similar se obtuvo en un estudio con 12
pacientes, donde los niveles proteicos de la integrina α6 se determinaron en

42
extractos totales de GSL [44]. Estas diferencias de expresión de α6 no se asociaron
al número de células inflamatorias, pues los pacientes presentaron un score de foco
variable de células inflamatorias.
Estos datos nos muestran que en el parénquima secretor de GSL de
individuos controles y pacientes con SS es posible detectar las integrinas α6 y β1. En
estos pacientes con SS existe una forma de mayor movilidad electroforética de la
integrina β1, no detectada en los individuos controles, y además existen individuos
donde la integrina α6 presenta una disminución importante en sus niveles de
expresión.
2. Inmunoprecipitación de las integrinas α6 y β1
Varios estudios reportan que la integrina α6 forma dímeros exclusivamente
con las integrinas β1 y β4 [94]. Previamente se describió que en GSL humanas, α6
forma un complejo con la integrina β4 [44], pero no existe información acerca de su
asociación con la integrina β1. En un pool de extractos de GSL se demostró
mediante inmunoprecipitación que las integrinas α6 y β1 forman un complejo (Figura
2B). Los resultados muestran que se inmunoprecipitó la integrina β1 cuando se
utilizaron anticuerpos dirigidos contra la integrina α6 o contra la cadena α1 de
laminina-111, mientras que no se encontró β1 cuando se usó un anticuerpo control
dirigido contra citoqueratina 18 (CK18).
Estos resultados permiten plantear que en GSL humanas las integrina α6, β1 y
β4 podrían dar origen a los heterodímeros α6β1 y α6β4. Interesantemente, en este
sistema se probó que en un inmunocomplejo donde se encuentra la integrinas β1 se
puede detectar la cadena α1 de laminina-111.

43
Figura 2. Determinación de los niveles proteicos e inmunoprecipitación de las integrinas β1 y α6. (A) Detección por WB de las integrinas β1 y α6 en extractos de GSL enriquecidos en componente epitelial de controles (carriles 1 y 2) y pacientes con SS (carriles 3-6). Se usó la detección de la GAPDH como control de carga, a la izquierda de cada gel se indica los marcadores de masa molecular. (B) Bandas para la integrina β1 fueron detectadas cuando se inmunoprecipitó con anticuerpos contra la integrina α6 y la cadena α1 de laminina-111, (carril 2 y carril 3, respectivamente), pero no con anticuerpos contra CK18, (carril 4), extracto total de GSL (carril 1) usado como control de su expresión.
B
kDa 150
100
IPP (-) α6 LM111 CK18
WB β1
1 2 3 4
Integrina β1
150
100
Controles Pacientes
37
250
150
37
Integrina β1
GAPDH
GAPDH
Integrina α6
kDa
1 2 3 4 5 6
A

44
3. Localización de las integrinas α6 y β1
Los acinos de GSL humanas son mixtos, ya que están formados por células
secretoras serosas y mucosas. La diferencia de cada tipo celular radica en su
morfología, organización y la composición proteica/glicoproteica de sus productos de
secreción [88]. A pesar de esta importante diferencia estructural y funcional, no se
han descrito diferencias en la composición de la lámina basal adyacente a estos dos
tipos celulares. En este trabajo, se analizó mediante inmunofluorescencia de
microscopía confocal, la localización de las integrinas α6 y β1 en acinos de GSL.
En criosecciones obtenidas de una GSL, proveniente de un individuo control y
un paciente con SS, se muestra la inmunolocalización de la integrina α6 y la integrina
β1 en acinos mixtos (Figura 3). En el control, la integrina α6 se localiza
preferentemente en las células acinares serosas (s), donde se observa una intensa
señal en el dominio de membrana plasmática baso-lateral (Figura 3A). En las células
mucosas (m), la intensidad de la integrina α6 es mas baja que en células serosas, y
también se localiza en la región de la membrana plasmática baso-lateral (3A). En los
acinos de pacientes con SS que mantienen su integridad morfológica, la localización
de la integrina α6 es similar a la descrita para controles (Figura 3B). Sin embargo, en
acinos que muestran una morfología alterada, se observa una disminución de la
señal presente en las células serosas, especialmente en el dominio de la membrana
plasmática lateral, donde no se definen claramente los límites celulares (Figura 3C).
En acinos de controles y de pacientes con morfología conservada, la integrina
β1 tiene una localización en el dominio baso-lateral de la membrana plasmática,
tanto en las células mucosas como serosas, y parece estar levemente aumentada en
las células serosas (Figura 3D y E). En la Figura 3F, se muestra un acino proveniente
de un paciente con SS, donde el componente seroso está marcadamente
desorganizado, y se observa señal citoplasmática para β1 en algunas células (flecha
roja). También, en este mismo paciente, la expresión de β1 parece estar aumentada
con respecto al control en forma generalizada. En G, H e I se muestra la
superposición de las señales para la integrina α6 en verde y β1 en rojo. En acinos
controles, α6 y β1 co-localizan preferentemente en el componente seroso (Figura

45
3G), tanto en el dominio basal (flecha celeste), como en el lateral (flecha amarilla),
mostrando un patrón de co-localización discontinuo (Figura 3G’). A partir de este
patrón de co-localización podríamos inferir la presencia de otros dímeros formados
con la integrina β1 y α6 como son α6β4 y α3β1 por ejemplo, ya que existen zonas en
que no co-localizan α6 y β1.
La co-localización de α6 y β1 en las células serosas daría cuenta de la acción
que la integrina α6β1 ejerce, tanto como receptor de componentes de la LB (co-
localización en dominio basal de la membrana plasmática) como participando en las
adhesiones intercelulares (co-localización en el domino lateral de la membrana
plasmática) en este tipo celular particular. Las integrinas presentes en el dominio
basal de la membrana plasmática, interaccionan con ligandos de la lámina basal
reportados en estudios previos. Sin embargo, los ligandos de las integrinas
localizadas en el dominio lateral aún no son claros.
Estos resultados constituyen la primera evidencia de la existencia del dímero
α6β1 en GSL, donde su ligando laminina-111 se expresa exclusivamente en la
lámina basal de acinos [33]. Las integrinas α6 y β1 co-localizan preferentemente en
los acinos serosos, donde se ubican en el dominio baso-lateral de la membrana
plasmática. No se observan diferencias en la localización de estas integrinas entre
acinos controles y de pacientes que presentan una morfología aparentemente
conservada. No obstante, en acinos de GSL de pacientes que presentan cambios
morfológicos importantes, es posible ver señal citoplasmática para la integrina β1 y
una disminución en la señal para α6 en el dominio de la membrana plasmática
lateral.

46
Figura 3. Localización de las integrinas α6 y β1 en acinos de GSL de individuo control y paciente con SS. Detección por inmunofluorescencia de las integrinas α6 (A, B y C, verde) y β1 (D, E y F, rojo) en secciones de 5μm obtenidas de la GSL de un individuo control (A, D, G) y de un paciente con SS (B, C, E, F, H, I). G’, H’ e I’ corresponden a ampliaciones digitales de los recuadros de G, H e I. El análisis se realizó por microscopía confocal (cortes confocales de 0.8 μm). Las barras corresponden a 10μm. (s: acino seroso, m: acino mucoso).
A
IHG
FE
CB
D
I’H’G’
Integrina β1
Integrina β1 Integrina α6
Integrina α6
Control Paciente
m s m
s m
s
m s m
s m
s
s
s
m s m
s
m
s
s

47
II. Determinación de los niveles de FAK fosforilada en glándulas salivales labiales de controles y pacientes con Síndrome de Sjögren.
Las cascadas de señalización gatilladas por integrinas tienen como elemento
común la fosforilación de la kinasa de adhesión focal (FAK) en múltiples residuos
[95]. Dentro de éstos, el residuo de Tyr357 (pFAK-Tyr357) es producto de la
autofoforilación y se da muy tempranamente dentro de sus fosforilaciones [51]. Como
medida de esta activación de señales río abajo de las integrinas, se determinó en
extractos de GSL de controles (n=8) y pacientes con SS (n=6) los niveles de pFAK-
Tyr357 y FAK mediante Westernblot, normalizados con respecto a la señal de actina.
En la Figura 4A, se muestran las bandas correspondientes a FAK, pFAK-Tyr357
y actina en controles (carriles 1-8) y pacientes con SS (carriles 9-14), donde FAK y
FAK fosforilada presentan masas moleculares estimadas de 120 kDa. La expresión
relativa de FAK total es heterogénea, tanto en controles como en pacientes con SS
(Figura 4C). El valor promedio (± desviación estándar) de los niveles relativos de FAK
en controles y pacientes con SS es de 1,06 ± 0,89 y 0,63 ± 0,71, respectivamente,
sin existir diferencias estadísticamente significativas entre ambos grupos (p=0.197).
La fosforilación del residuo de Tyr357 en FAK también es heterogénea dentro
de ambos grupos (Figura 4B), no asociándose estas diferencias con los niveles de
expresión de FAK. En algunos individuos existe una relación inversa entre el nivel de
expresión de la proteína y la fosforilación de este residuo (ver controles 2 y 3). Al
analizar la razón entre FAK fosforilada y FAK total se mantiene esta variación entre
los distintos individuos del grupo control y de pacientes (Figura 4D). El promedio (±
desviación estándar) en el grupo de controles y pacientes con SS es de 0,33 ± 0,32 y
0,50 ± 0,31 respectivamente, y estas diferencias no son estadísticamente
significativas (p=0.245).
Siendo FAK un indicador de la activación inicial desencadenada por todas las
integrinas, y dado que en el tejido la expresión de la integrina α6β1 no es exclusiva,
este resultado da cuenta de todas las señales activadas por integrinas y de la
transactivación dependiente de factores de crecimiento.
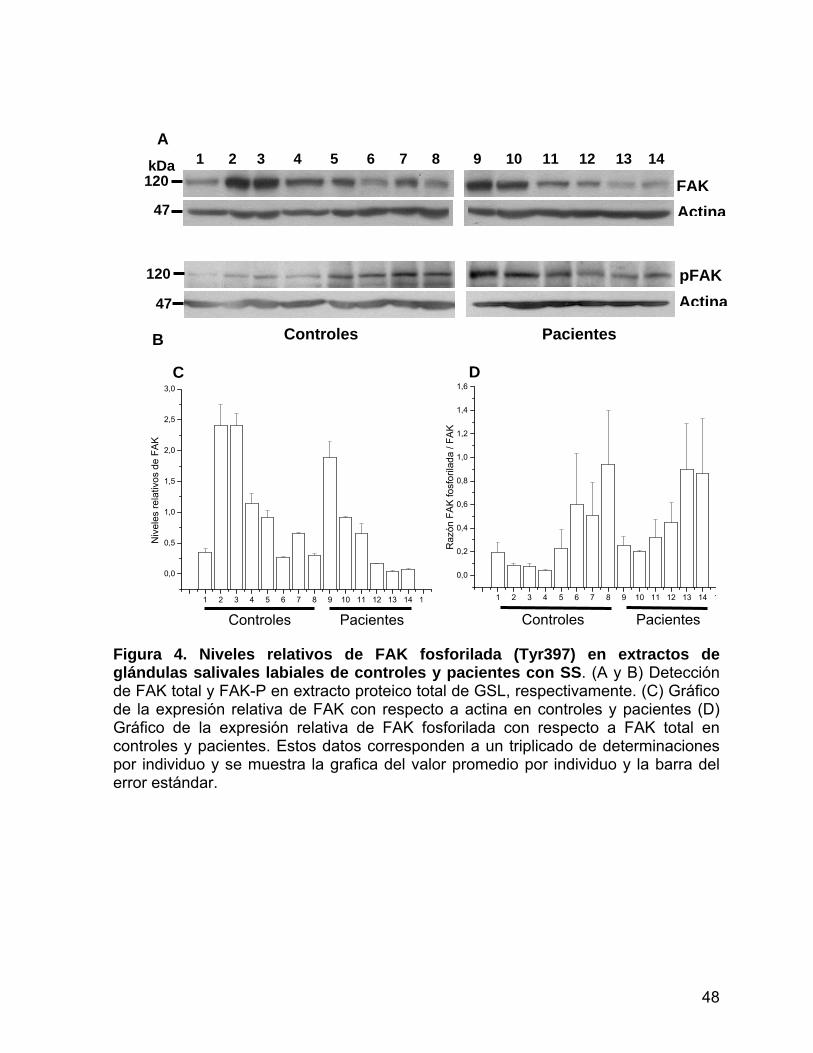
48
Figura 4. Niveles relativos de FAK fosforilada (Tyr397) en extractos de glándulas salivales labiales de controles y pacientes con SS. (A y B) Detección de FAK total y FAK-P en extracto proteico total de GSL, respectivamente. (C) Gráfico de la expresión relativa de FAK con respecto a actina en controles y pacientes (D) Gráfico de la expresión relativa de FAK fosforilada con respecto a FAK total en controles y pacientes. Estos datos corresponden a un triplicado de determinaciones por individuo y se muestra la grafica del valor promedio por individuo y la barra del error estándar.
Controles Pacientes
FAKActina
A 1 2 3 4 5 6 7 8 9 10 11 12 13 14
B
pFAK Actina
kDa 120
120
47
47
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
0,0
0,5
1,0
1,5
2,0
2,5
3,0
Niv
eles
rela
tivos
de
FAK
Controles Pacientes
C
Controles Pacientes
D
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
Raz
ón F
AK
fosf
orila
da /
FAK

49
III. Establecimiento y caracterización de un sistema de cultivo en tres dimensiones de acinos de células HSG
1. Caracterización morfológica de los acinos de células HSG
En esta tesis se postula un mecanismo que permitiría explicar los cambios de
polaridad descritos en acinos de GSL de pacientes con SS. Este mecanismo
relaciona los cambios de polaridad y organización acinar con señales alteradas
desde la lámina basal, y surge de la inferencia de datos descriptivos obtenidos de
glándulas salivales de pacientes con SS. No obstante, no se ha comprobado
experimentalmente la relación causa-efecto que permitiría sustentar este
planteamiento. Por esta razón, fue necesario el desarrollo de un modelo de estudio
posible de intervenir y que permita estudiar probables cambios de polaridad.
Con este propósito, como elemento central de esta tesis, se desarrolló un
sistema de estudio de organización y polaridad acinar in vitro, cercano a lo que
ocurre en la glándula, utilizando cultivos en tres dimensiones (3D) de células
derivadas de glándula submandibular humana (HSG). Los modelos de cultivo en 3D
permiten el crecimiento de células inmersas en una matriz semi-sólida de proteínas
componentes de la matriz extracelular, y permiten la organización de estructuras tipo
acinos a partir de células epiteliales libres.
Las células HSG son de origen epitelial transformadas, aisladas desde ductos
intercalares de glándula submandibular humana irradiada [88,89]. Estas células
pueden diferenciarse hacia distintos fenotipos dependiendo de los factores a los
cuales son expuestas [96,97].
Se han descrito diversas metodologías para el cultivo de acinos de células
HSG en matrigel [78,79,98], pero ninguno de estos estudios ha entregado un análisis
detallado de las características moleculares y morfológicas de los acinos generados.
En esta tesis se utilizó un protocolo descrito para las células MCF-10A,
derivadas de glándula mamaria [77]. Las células se cultivan sobre una base de
extracto de lámina basal concentrado (Figura 1). Como este protocolo no estaba
probado para células HSG fue necesario controlar el desarrollo temporal del cultivo
de acinos, a fin de definir el tiempo de cultivo adecuado para obtener acinos

50
polarizados.
Se analizó la morfología de los acinos de células HSG durante 5 días de
cultivo, comparándolos con el patrón de crecimiento de las células en plano (Figura
5). Las células cultivadas en plano crecen adheridas a la placa hasta alcanzar
confluencia (Figura 5, columna 1). En cambio, las células cultivadas en matrigel se
organizan como estructuras esféricas de diversos tamaños y formas (Figura 5,
columna 2). El tamaño de los acinos va aumentando durante los días de cultivo y se
estabiliza al cuarto día. Las estructuras iniciales están formadas por 1-3 células y
terminan organizando grupos de distintos tamaños, formados por más de 10 células,
donde se visualizan hendiduras centrales que corresponderían al lumen (Figura 5,
columnas 3 y 4).

51
Figura 5. Curso temporal del cultivo de acinos de células HSG en extracto de lámina basal. Microfotografías de células HSG cultivadas en plano (columna 1) y en extracto de lámina basal (columnas 2, 3 y 4) durante 5 días de cultivo (d1-d5). Las imágenes de las columnas 1 y 2 tienen un aumento de 50x y de 200x en las columnas 3 y 4.
d1
d2
d3
d4
d5
plano (2D) Acinos de células HSG crecidos en 3D
1 2 3 4

52
La morfología celular en acinos de 3 y 5 días de cultivo fue analizada en
secciones de 1μm teñidas con azul de toluidina (Figura 6A-F). Se observó que al
tercer día de cultivo los acinos tienen un número menor de células que a los 5 días, y
existen fenotipos nucleares diversos en un mismo acino, observándose algunas
células en activa mitosis (Figura 6B y C, flecha roja). Se distinguen lúmenes
pequeños que no están presentes en todos los acinos (Figura 6A y C, asterisco). Al
quinto día de cultivo, se observan acinos formados por una capa única de células
posicionada alrededor de un lumen extenso, el cual está presente en la mayoría de
los acinos (Figura 6D-F). Las células tienen forma piramidal y la proporción de
células mitóticas es mas baja que en acinos de tres días de cultivo. Se observan
restos celulares en algunos lúmenes, lo que podría ser parte del proceso apoptótico
necesario para la formación del lumen (flecha negra).
Estos acinos se han mantenido en cultivo durante 7 días, no obstante la
morfología de las células cambia desde una forma piramidal, clásica de las células
acinares a una cuboídea y baja, característica de las células ductales y lejana al
fenotipo que interesa en este estudio. Estas observaciones permitieron definir el día
5 como el más óptimo para hacer los ensayos de polaridad, ya que claramente está
definido el eje ápico-basal de las células, donde la evidencia más sólida es la
formación del lumen acinar.
Al analizar la ultraestructura de las células acinares, al quinto día de cultivo,
mediante microscopía electrónica de transmisión (Figura 6G y H) es posible describir
algunas características del fenotipo acinar. Se observa una cromatina
descondensada y homogénea, lo que se relaciona con una alta actividad
transcripcional (Figura 6H). Las cisternas del complejo de Golgi se observan
abundantes y bien organizadas (Figura 6G, flechas rojas), además existe una gran
cantidad de mitocondrias, las cuales presentan distintos tamaños y algunas se
encuentran en fisión, sugiriendo un alto requerimiento energético (Figura 6G, flechas
negras). También podemos detectar que está comenzando la organización del
retículo endoplásmico rugoso, donde claramente se observan cisternas largas
aplanadas con ribosomas asociados a sus membranas (Figura 6H, flechas negras).

53
Este análisis morfológico nos permite afirmar que los acinos de células HSG
de 5 días de cultivo están formados por células con algunas características acinares
similar a los acinos glandulares in vivo, donde se ha desarrollado un lumen, indicativo
de la estructuración de un eje de polaridad ápico-basal. Todos los experimentos
posteriores son realizados en acinos de 5 días de cultivo, a menos que se indique
alguna variación que se describe particularmente.

54
Núcleo
H G
d5
d3
A
*
*
B
*
D
*
C
E F
*
Figura 6. Análisis morfológico de acinos de células HSG. Secciones de 1 μm de acinos de 3 (A-C) y 5 (D-F) días de cultivo incluidos en Epon, fueron teñidos con azul de toluidina y observados al microscopio de luz, captándose las imágenes con un aumento de 300x. Las flechas rojas indican células en mitosis, las negras indican restos celulares luminales y los asteriscos la localización del lumen. En G y H se muestran secciones de acinos (600Aº) de células HSG al quinto día de cultivo procesados para microscopia electrónica de transmisión. Flechas rojas indican el complejo de Golgi, las negras el retículo endoplásmico y las azules mitocondrias (d : días de cultivo).

55
2. Caracterización molecular del fenotipo polarizado de los acinos de células HSG
Las células HSG, al organizarse en acinos presentan un cambio fenotípico
importante. Éste se ve reflejado en la organización del citoesqueleto de actina y la
morfología nuclear (Figura 7). La actina polimerizada (F-actina) se distribuye en la
zona cortical de todas las células del acino de células HSG, concentrándose en la
zona apical, delimitando el lumen (Figura 7F). El citoesqueleto de actina de los
acinos de HSG adquiere la misma distribución que tiene en acinos de GSL humanas
(comparar con Figura 7D). Es importante destacar que no se observan las clásicas
fibras de estrés descritas en células crecidas en plano (Figura 7E), por lo que es
claro que las señales son distintas y mediarían procesos estables de asociación con
la lámina basal.
Por otra parte, al analizar la morfología nuclear de acinos de HSG, éstos se
ven con cromatina descondensada, con nucléolo único, a diferencia de lo observado
en las células cultivadas en plano (Figura 7B). Esta reorganización de la cromatina
daría cuenta del cambio “funcional” esperado en los acinos reconstituidos, y donde
debe ocurrir una transformación, desde un estado preferentemente proliferativo a
uno más diferenciado.
La organización de las adhesiones célula-célula, especialmente las uniones
estrechas, son fundamentales en la definición del fenotipo epitelial polarizado, por lo
que consideramos importante detectar su presencia. Mediante microscopía
electrónica de transmisión se observaron distintos tipos de uniones intercelulares en
acinos de cinco días de cultivo (Figura 8). Se observan desmosomas en botón de
distintos tamaños, además existen extensas zonas de adhesión célula-célula tipo
uniones comunicantes. Mediante Westernblot se determinó la expresión de claudina-
4, una proteína de uniones estrechas, en células HSG crecidas tanto en 2D como
3D, sin embargo, la expresión es mucho mayor en células crecidas en 3D. Además,
se detectó una forma de ocludina exclusivamente en células HSG cuando son
crecidas en 3D (Figura 8E). Por RT-PCR, se ha encontrado la expresión de ZO1 y
claudina 4, detectándose el mensajero de esta última exclusivamente en células
HSG cultivadas en 3D (Figura 8D). Estos resultados son importantes si
consideramos que la organización de estructuras de adhesión, en particular de

56
uniones estrechas, es un proceso relevante en la definición de los ejes de polaridad
celular.
A pesar de la diferenciación morfológica observada en los acinos de células
HSG, éstos no expresan ninguna proteína de secreción. En la Figura 8D se muestra
que los mensajeros de MUC7, MUC5B y Amilasa no se expresan en células HSG
bajo ninguna condición de cultivo; ya que la adición al cultivo de EGF, extracto de
pituitaria e insulina no induce la expresión de estos genes. Por lo tanto, la
diferenciación morfológica observada en los acinos de células HSG no se
correlaciona con la diferenciación funcional secretora.
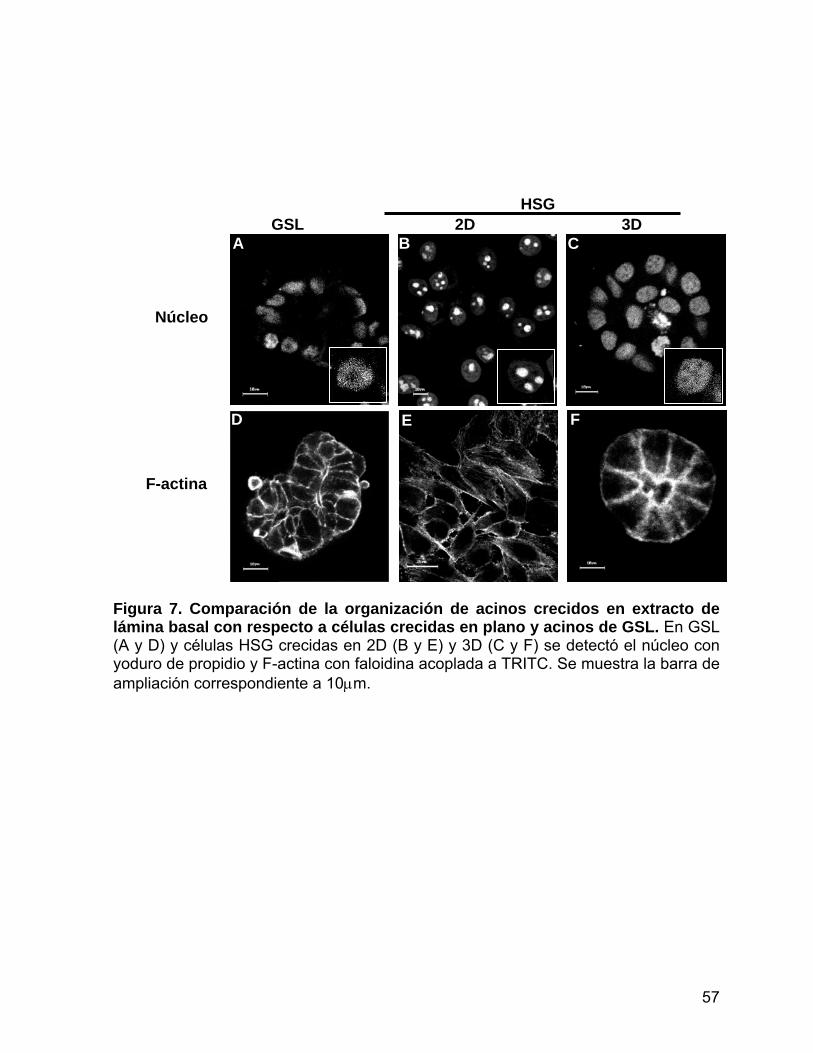
57
GSL 2D 3D HSG
Núcleo
F-actina
BA C
E F D D
Figura 7. Comparación de la organización de acinos crecidos en extracto de lámina basal con respecto a células crecidas en plano y acinos de GSL. En GSL (A y D) y células HSG crecidas en 2D (B y E) y 3D (C y F) se detectó el núcleo con yoduro de propidio y F-actina con faloidina acoplada a TRITC. Se muestra la barra de ampliación correspondiente a 10μm.

58
Si bien el cultivo de acinos se realiza en un extracto de lámina basal, fue
importante evaluar si se organiza correctamente una lámina basal “endógena”
alrededor de los acinos, para ello se determinó la presencia de uno de sus
componentes, colágeno tipo IV. En la Figura 9, se muestra la detección por
inmunofluorescencia de colágeno tipo IV, el cual se concentra rodeando las células
acinares. Esto sugiere que en este tipo de cultivo se organiza una lámina basal. La
organización de una LB rodeando los acinos, es importante en la determinación de
los ejes de polaridad, ya que las señales dependientes de la lámina basal son
fundamentales en este proceso. La Figura 9 también muestra la localización basal de
los núcleos, muy cercana a la lámina basal y que claramente concuerda con la
descripción de núcleos basales en acinos de GSL.
En células epiteliales polarizadas el complejo de Golgi se localiza cerca de la
porción apical de la superficie celular, en posición supra-nuclear [99]. La disposición
de este organelo es un marcador de polaridad correcta en una célula epitelial. En los
acinos de HSG con 5 días de cultivo, se estudió la localización del complejo de Golgi
a través de la detección de la proteína gigantina, la cual es un marcador específico
de este organelo [100]. En la Figura 9D-F se observa un corte confocal a través del
centro del lumen acinar donde se ve la marca de gigantina (verde) hacia la porción
apical y supra-nuclear.
Estos resultados nos permiten afirmar que en nuestro sistema de cultivo las
células HSG se organizan en estructuras tipo acinos que presentan una distribución
de organelos polar, con el complejo de Golgi supranuclear, núcleos basales y una
lámina basal rodeando los acinos, sin embargo, no se expresan proteínas de
secreción.

59
Figura 8. Adhesiones célula-célula en acinos de células HSG. Acinos de células HSG de 5 días de cultivo fueron procesados para microscopía electrónica de transmisión y se realizaron secciones de 600 Aº para su observación (A-C). Las flechas negras muestran desmosomas en botón y las flechas azules zonas de uniones tipo comunicantes. En D se muestra RT-PCR de Claudina-4 (Clau-4), ZO-1, MUC7, MUC5B, Amilasa (Amy), Cistatina (Cys) y GAPDH a partir de RNA total de extractos de GSL y células HSG crecidas en 2D y 3D. En E se muestra la detección de claudina-4, ocludina y GAPDH para extractos totales por Westernblot. 3D (+) indica la adición de EGF, extracto de pituitaria e insulina en el cultivo, y 3D (-) corresponde al cultivo en extracto de lamina basal sin estos factores.
D
B A C
ZO1
Clau-4
GAPDH
GSL 2D 3D- 3D+ (-) HSG
107
199
670
MUC7
Amy MUC5
407
415
474
pb
525 Cys
E
20
37
Ocludina
Clau-4
GSL 2D 3DHSG
GAPDH
Kda
65 72
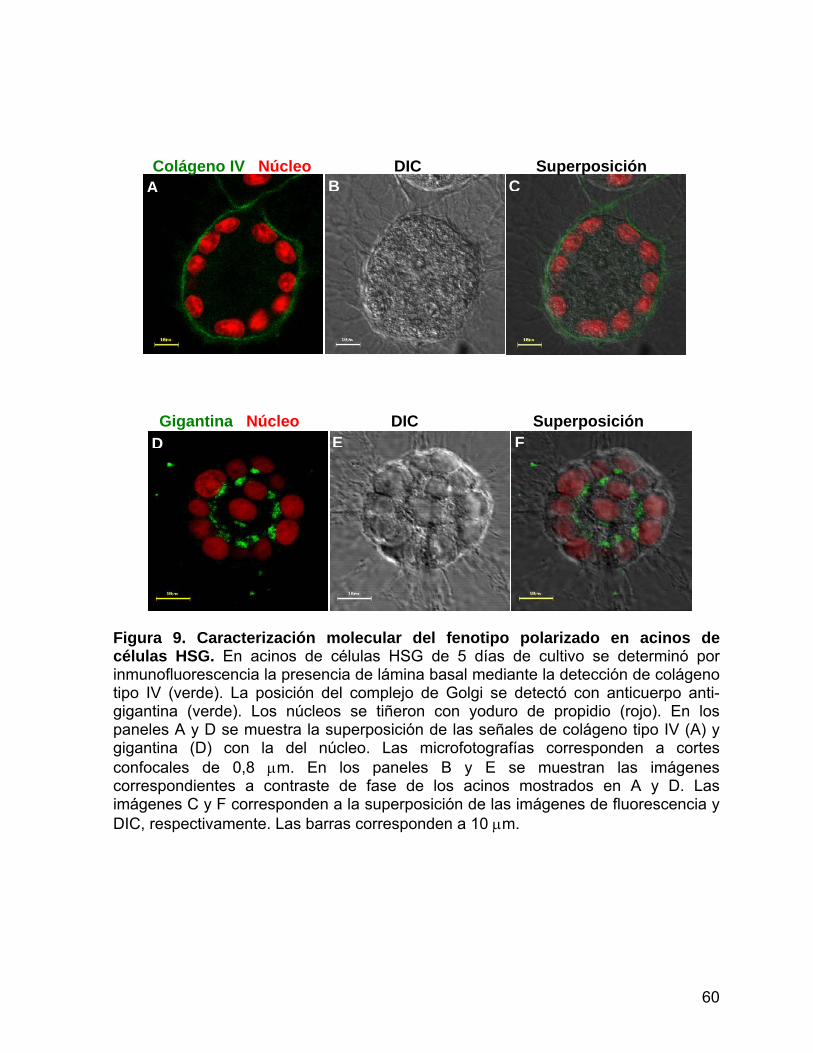
60
Figura 9. Caracterización molecular del fenotipo polarizado en acinos de células HSG. En acinos de células HSG de 5 días de cultivo se determinó por inmunofluorescencia la presencia de lámina basal mediante la detección de colágeno tipo IV (verde). La posición del complejo de Golgi se detectó con anticuerpo anti- gigantina (verde). Los núcleos se tiñeron con yoduro de propidio (rojo). En los paneles A y D se muestra la superposición de las señales de colágeno tipo IV (A) y gigantina (D) con la del núcleo. Las microfotografías corresponden a cortes confocales de 0,8 μm. En los paneles B y E se muestran las imágenes correspondientes a contraste de fase de los acinos mostrados en A y D. Las imágenes C y F corresponden a la superposición de las imágenes de fluorescencia y DIC, respectivamente. Las barras corresponden a 10 μm.
Colágeno IV Núcleo DIC Superposición A B C
Gigantina Núcleo DIC Superposición D E F

61
IV. Importancia de la integrina α6 en la organización acinar y el establecimiento
del fenotipo polarizado
1. Expresión de las integrinas α6 y β1 en acinos de células HSG
Una evaluación fundamental, en el contexto de esta tesis, fue la expresión de
las integrinas α6 y β1 en los acinos de células HSG y su relación con lo que ocurre in
vivo (Figura 10). En GSL se ha descrito la expresión de las variantes de
procesamiento α6A y α6B [44] y por esto se analizó mediante RT-PCR si ellas se
expresan en acinos de células HSG. Se detectaron ambas variantes en acinos, pero
interesantemente, las células crecidas en 2D expresan exclusivamente el mensajero
de la variante α6B (Figura 10A). La presencia del mensajero de α6B en células
crecidas en 2D no se correlaciona con su traducción. Al observar el patrón de
expresión proteica de α6 mediante Westernblot se observa una expresión diferencial
entre las células HSG cultivadas en 2D y 3D, pues esta proteína no se detecta en
células crecidas en 2D (Figura 10). Con respecto a la gran expresión que se observa
en el extracto de acinos de HSG, es importante considerar que este extracto incluye
el matrigel presente en el cultivo y que se detecta una banda de tamaño equivalente
a la detectada para α6 en matrigel puro (Figura 10B), por lo que los niveles de
expresión no son comparables.
La expresión inequívoca de α6 en los acinos 3D se analizó mediante
inmunofluorescencia (Figura 10C). Aquí se observa que presenta una clara
distribución en el dominio basal de la membrana plasmática y débil señal
citoplasmática. Esta expresión asimétrica resulta fundamental en la definición
operacional de fenotipo polarizado en el sistema de cultivo utilizado, ya que daría
señales de la diferenciación operacional que se da en los dominios de membrana
plasmática. Considerando que in vivo, las GSL expresan la integrina α6, y que ésta
sólo se expresa en HSG crecidas en 3D, se refuerza la importancia que se plantea
para la organización tridimensional de los acinos en la correcta expresión y
señalización.

62
Luego se analizó la localización en conjunto de las integrinas α6 y β1 en
nuestro sistema de cultivo para sostener la posible existencia del dímero α6β1, que
se muestra en la Figura 11. Como se había descrito previamente, la integrina α6
presenta una evidente localización asociada a membrana plasmática basal y β1 se
localiza homogéneamente en la membrana plasmática baso-lateral, pero presenta
una señal citoplasmática intensa. Observamos que existe co-localización de ambas
proteínas en el dominio basal de membrana plasmática, asociada a la zona de la
lámina basal.
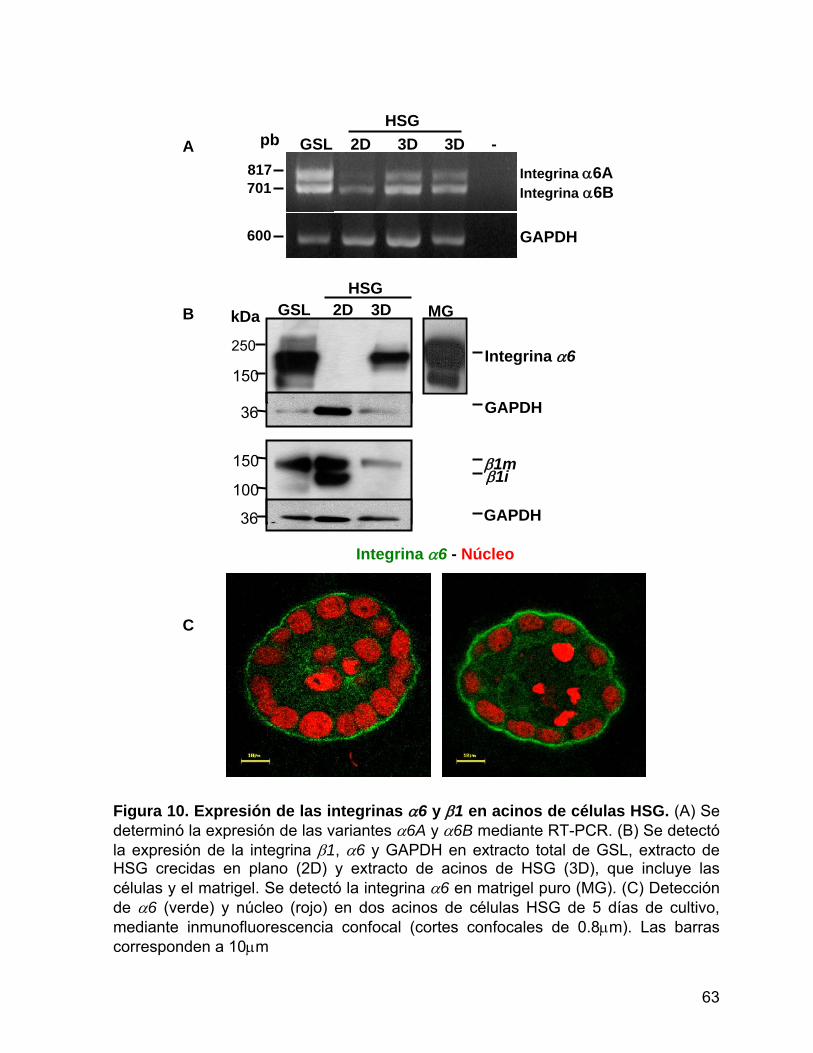
63
GAPDH
Integrina α6
β1i
GAPDH
β1m
36
36
150
100
250
150
GSL 2D 3D kDa
HSG
A
B
C
Integrina α6 - Núcleo
MG
GAPDH
Integrina α6A Integrina α6B
GSL 2D 3D 3D -
701 817
600
pb HSG
Figura 10. Expresión de las integrinas α6 y β1 en acinos de células HSG. (A) Se determinó la expresión de las variantes α6A y α6B mediante RT-PCR. (B) Se detectó la expresión de la integrina β1, α6 y GAPDH en extracto total de GSL, extracto de HSG crecidas en plano (2D) y extracto de acinos de HSG (3D), que incluye las células y el matrigel. Se detectó la integrina α6 en matrigel puro (MG). (C) Detección de α6 (verde) y núcleo (rojo) en dos acinos de células HSG de 5 días de cultivo, mediante inmunofluorescencia confocal (cortes confocales de 0.8μm). Las barras corresponden a 10μm

64
Figura 11. Co-localización de las integrinas α6 y β1 en acinos de células HSG. En dos acinos de células HSG de 5 días de cultivo se detectó por inmunofluorescencia la integrina β1 (A y D) e integrina α6 (B y E). En C y D se muestra la superposición de ambas imágenes donde la integrina β1 se muestra en rojo y α6 en verde. C’ y F’ corresponden a ampliaciones digitales de los recuadros de C y F. El análisis se realizó por microscopía confocal (cortes confocales de 0.8 μm). Las barras corresponden a 10μm.
Integrina β1
Integrina β1 Integrina α6
Integrina α6
B
C
E
F
C’ F’
A D

65
2. Efecto del bloqueo de la integrina α6 sobre la organización y mantención de
fenotipo polarizado de acinos de células HSG
Se evaluó la importancia que tiene la integrina α6 en la formación y el
establecimiento del fenotipo polarizado de acinos de células HSG. La estrategia
experimental utilizada fue bloquear la interacción de esta integrina con sus ligandos
presentes en la lámina basal mediante el uso de un anticuerpo contra α6 (GoH3).
Este anticuerpo presenta la capacidad bloqueante de esta interacción, la cual ha sido
ampliamente descrita [101] y además ha sido utilizado previamente con esta finalidad
en cultivos de células crecidas en matrigel [102]. El ensayo consiste en cultivar las
células HSG en presencia del anticuerpo GoH3 en condiciones similares al cultivo
acinar, y luego analizar la organización acinar y la polaridad de sus células mediante
el uso de marcadores específicos.
Al bloquear la integrina α6 desde el inicio del cultivo de acinos, se impide
completamente su formación. En la Figura 12 se muestran dos imágenes de
microscopía de luz de acinos al tercer día de cultivo, donde se comparan los acinos
en condición control, sin bloqueo (Figura 12A), y en condiciones de bloqueo
utilizando 3 μg/mL de GoH3 (Figura 12B). Se observa que las células se mantienen
inmersas en el matrigel, pero no forman acúmulos y existe un número bajo de células
al final de los 5 días de cultivo, lo que indicaría que las células no proliferan. Todas
las diluciones de este anticuerpo utilizadas están dentro del rango que se describe en
la literatura, y ellas inducen este cambio. Este resultado sugiere que la interacción de
las integrinas de la familia α6 con componentes de la lámina basal es fundamental en
la formación de los acinos de células HSG, no descartando que integrinas de otras
familias u otros receptores de membrana plasmática pudieran estar involucrados en
el control de la formación acinar.
Para determinar la influencia que tiene la integrina α6 en la mantención del
fenotipo de los acinos en formación, se probaron distintas concentraciones de GoH3
dentro del rango descrito en la literatura (1 - 5 µg/mL). GoH3 se agregó al cultivo al
comienzo del tercer día, y se mantuvo hasta finalizar los 5 días de cultivo (Figura
12C-E). Para definir un correcto establecimiento del eje de polaridad ápico-basal se

66
analizó la morfología acinar por contraste de fase, y la relación posicional del
complejo de Golgi (gigantina, verde) con respecto al núcleo (yoduro de propidio,
rojo).
Los acinos tratados con GoH3 se desorganizan dependientes de la
concentración de anticuerpo utilizada. Los acinos tratados con 3 y 5 μg/mL de GoH3
pierden su forma esférica y se observan como grupos celulares y células libres. El
efecto es más dramático cuando se utiliza una concentración de GoH3 de 5μg/mL,
donde se oberva una pérdida total de la organización acinar (Figura 12D’ y E’). Esta
desorganización morfológica se correlaciona con el patrón desordenado que
presentan los núcleos y el complejo de Golgi en las células. En las Figuras 12D y E
se puede apreciar que no se distingue una zona luminal claramente definida, y los
núcleos no se encuentran en el dominio basal de las células. Además, la posición del
complejo de Golgi se observa completamente al azar con respecto al núcleo,
mostrando que no existe ningún eje de polaridad definido. Además, al utilizar 5μg/mL
de GoH3 se observa una disminución aparente en el número total de células en el
cultivo. Este efecto es moderado cuando se utiliza 1μg/mL de GoH3, ya que los
acinos aún mantienen una organización esférica (Figura 12C’) y el lumen se observa
definido (Figura 12C). Además, en esta condición experimental, el complejo de Golgi
en algunas células conserva su posición supranuclear, mientras en otras células este
organelo no se visualiza en el corte.
Junto con esta desorganización acinar, el bloqueo de la integrina α6 promueve
la apoptosis celular. La morfología nuclear de estas células muestra la presencia de
cuerpos apoptóticos de distintos tamaños además de condensaciones de cromatina
tipo semilunar en la periferia de núcleos que aún permanecen organizados (Figura
12C-E, recuadros pequeños). Al utilizar 5 µg/mL de GoH3 se observa una importante
disminución en el número de células, y se podría inferir que la mayoría de las células
ya han sufrido apoptosis, mientras las células remanentes muestran evidencias de
desorganización y no existen estructuras tipo acinos. En células mamarias crecidas
en matrigel, se ha descrito que la presencia de anticuerpos bloqueantes de la
integrina β1 induce el proceso apoptótico, sugiriendo fuertemente que en este
sistema, la integrina β1 transmite señales desde la matriz extracelular que son

67
requeridas para la sobrevida [103].
La especificidad del efecto del anticuerpo bloqueante GoH3, se corroboró
utilizando inmunoglobulinas (IgGs) purificadas desde suero de rata, en
concentraciones de 3 y 5 μg/mL, bajo las mismas condiciones que el caso
experimental (Figura 13). Los acinos mantienen una morfología esférica, sin cambios
en la localización del complejo de Golgi supranuclear y la morfología nuclear es
homogénea, sin signos apoptóticos.

68
1μg/mL 5μg/mL GoH3
C
C’
3μg/mL GoH3
D
D’
Gigantina - Núcleo
A B
E
E’
Figura 12. Efecto del bloqueo de la integrina α6 en la organización de acinos de células HSG. (A-B) Imágenes de acinos de células HSG de tres días de cultivo que fueron cultivados en presencia (B) o ausencia (A) de anticuerpo bloqueante GoH3 a una concentración de 3μg/mL desde el inicio del cultivo. Las fotografías presentan un aumento de 200x. (C-D) Acinos de células HSG fueron cultivados durante 2 días en condiciones normales y posteriormente fueron tratados con anticuerpo bloqueante GoH3 a una concentración de 1μg/mL (C), 3μg/mL (D) o 5μg/mL (E) durante 3 días. En verde se muestra la detección de gigantina (Golgi) y en rojo el núcleo teñido con yoduro de propidio. En paralelo se muestra la imagen correspondiente de los acinos en contraste de fase (C’, D’, E’). La barra corresponde a 10μm.
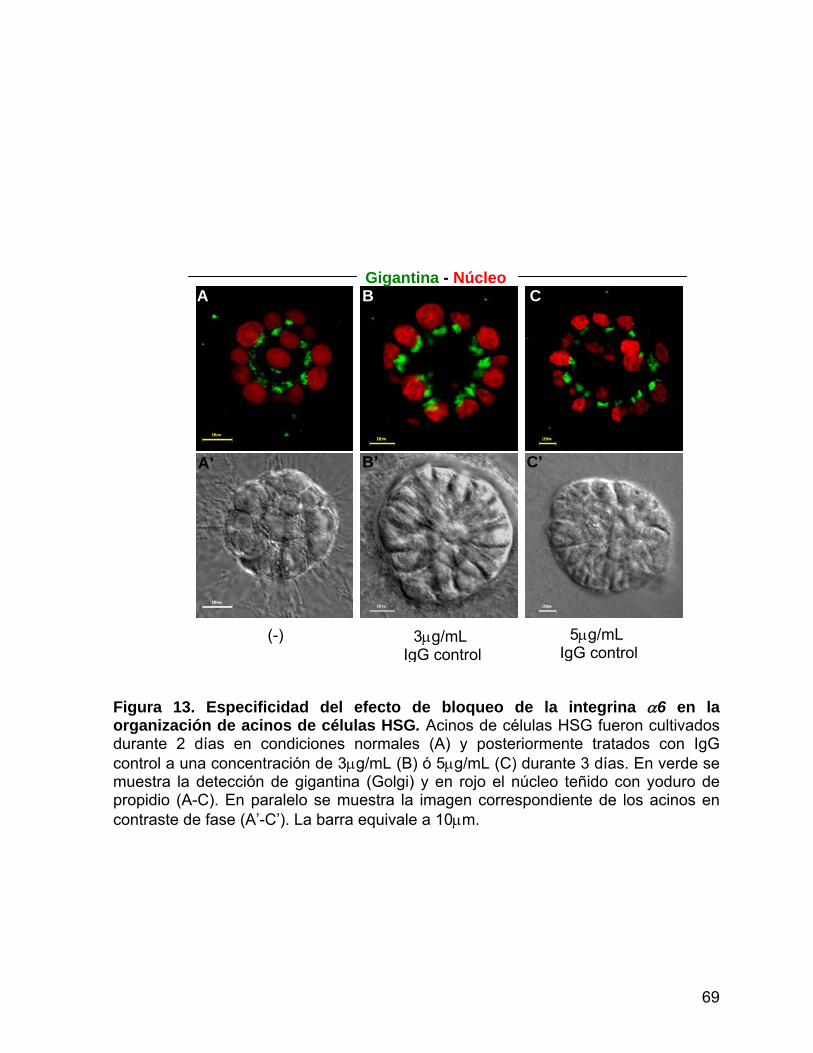
69
Figura 13. Especificidad del efecto de bloqueo de la integrina α6 en la organización de acinos de células HSG. Acinos de células HSG fueron cultivados durante 2 días en condiciones normales (A) y posteriormente tratados con IgG control a una concentración de 3μg/mL (B) ó 5μg/mL (C) durante 3 días. En verde se muestra la detección de gigantina (Golgi) y en rojo el núcleo teñido con yoduro de propidio (A-C). En paralelo se muestra la imagen correspondiente de los acinos en contraste de fase (A’-C’). La barra equivale a 10μm.
A B C Gigantina - Núcleo
A’ B’ C’
3μg/mL IgG control
5μg/mL IgG control
(-)

70
Se realizó un ensayo para demostrar la presencia del anticuerpo bloqueante
GoH3 en la región basal de membrana plasmática de los acinos en formación. Este
control permite determinar que el anticuerpo no queda retenido en el matrigel, a
pesar que se ha detectado por Westernblot la presencia de la integrina α6 en esta
matriz. Con este propósito, en acinos de células HSG bloqueados durante los últimos
3 días de cultivo con 3μg/mL de GoH3, se detectó la presencia de este anticuerpo
mediante inmunofluorescencia. Se omitió el anticuerpo primario y se utilizó
exclusivamente el anticuerpo secundario que reconoce al anticuerpo GoH3 (anti IgG
de rata acoplado a Alexa 488). En la Figura 14 se muestra la detección por
inmunofluorescencia del anticuerpo contra α6 en una situación sin bloqueo (Figura
14A) y bloqueando con GoH3 (Figura 14B y C). Se observa claramente que en la
situación control no se detecta el anticuerpo unido, lo que descarta la unión
inespecífica del anticuerpo secundario. Mientras que en la condición de bloqueo se
detecta marca verde alrededor de un agregado celular semi-organizado y alrededor
de células individuales en los casos de alta desorganización (Figura 14B y C). Estos
datos nos permiten confirmar que el anticuerpo bloqueante tiene accesibilidad a la
zona basal de la membrana plasmática acinar, donde se encuentra su blanco a
bloquear, la integrina α6, y más aún, el anticuerpo no queda retenido en el extracto
de la matriz extracelular.
Por otra parte, se controló si la presencia de este anticuerpo bloqueante altera
la organización de la lámina basal existente, detectando mediante
inmunofluorescencia la localización de colágeno tipo IV y laminina-111, componentes
de lámina basal. En la Figura 14 se muestra la detección de ambas proteínas en las
condiciones de bloqueo (E, F y H) y sin bloqueo (D y G). Se puede observar que la
señal para colágeno tipo IV se mantiene alrededor de las células acinares, sin
cambios sustantivos en la situación de bloqueo de la integrina, y sólo se observa una
señal un poco más difusa pero se mantiene delimitando los acinos. Igualmente, la
disposición fibrilar de laminina-111 en el matrigel se mantiene, muy concentrada y
organizada en forma de trama alrededor del acino.

71
Figura 14. Efecto del bloqueo de la integrina α6 en la organización de la lámina basal en acinos de células HSG. Células HSG fueron cultivadas en 3D durante 2 días, y posteriormente cultivados en presencia (B, C, E, F, H, y J) o ausencia (A, D, G e I) de 3 μg/mL de anticuerpo bloqueante de la integrina α6 (GoH3) durante 3 días. Se analizó mediante inmunofluorescencia la localización del anticuerpo bloqueante GoH3 usando exclusivamente un anticuerpo secundario anti IgG de rata acoplado a un fluoróforo (verde, A-C). Además, se detectó la presencia de Colágeno tipo IV (verde, D-F) y laminina-111 (verde, G-J) junto a yoduro de propidio (rojo) para teñir los núcleos. I y J corresponden a la señal para laminina-111 en zonas donde no existen células. La barra equivale a 10μm en A, B y D, y 25μm en las otras imágenes.
E D F
Colágeno tipo IV
α IgG rata Núcleo
Laminina-111 Núcleo
H G
J I
A B C Sin bloqueo 3μg/mL GoH3

72
Como se describió previamente, la incubación con el anticuerpo bloqueante
desde el inicio del cultivo, no permite la organización de los acinos (Figura 12B), y si
se inicia el bloqueo al tercer día de cultivo estos se desorganizan totalmente (Figura
12C-E). Se realizaron distintos tiempos de bloqueo de la integrina α6 en acinos de
células HSG, los cuales fueron analizados morfológicamente y con una tinción doble
de núcleo y complejo de Golgi, para evidenciar la correcta estructuración del eje de
polaridad ápico-basal. En la Figura 15 se muestran los resultados correspondientes
al bloqueo de la integrina α6 realizados de forma continua durante las etapas
terminales del cultivo, considerado los últimos 3, 2 ó 1 día(s) (Figura 15, ver
esquema). Si se inicia el bloqueo de la integrina α6 al finalizar el segundo día de
cultivo, los acinos se desorganizan totalmente (Figura 15, IV) pierden su forma
esférica (Figura 15F’ y G’) y sus células pierden cualquier organización polarizada.
Sin embargo, si se comienza al día 4 ó 5 de cultivo, los acinos mantienen su
morfología esférica (Figura 15B’, C’, D’ y E’). Sin embargo, la disposición de las
células dentro del acino y la relación núcleo-Golgi están muy alteradas al bloquear la
integrina al día 4 (Figura 15D y E), manteniéndose mas normales si el bloqueo es
exclusivamente durante el quinto día de cultivo (Figura 15B y C). Esto sugiere, que la
acción de la integrina α6 es más relevante en los primeros estadios de la formación
del acino, durante el proceso mismo de diferenciación, o significa que un día de
bloqueo no es suficiente para desencadenar la desorganización.
Además, se determinó si las células son capaces de continuar organizando un
fenotipo normal, luego de la interrupción transitoria de la interacción de la integrina
α6 con la lámina basal. Para esto se utilizó pulsos temporales de bloqueo de la
integrina α6 con GoH3. Así, a partir del tercer día de cultivo se bloqueó durante 1, 2 ó
24 hora(s) y posteriormente se reestableció la condición experimental sin anticuerpo
(Figura 16, esquema). Se muestra que en los tres casos se mantiene la morfología
esférica de los acinos según lo observado al contraste de fase (Figura 16B’, C’, D’,
E’, F’ y G’). Sin embargo, ya con 24 horas de bloqueo la morfología celular ha
cambiado, se observa que un grupo importante de las células presenta la localización
del complejo de Golgi en posición lateral y no supranuclear (Figura 16F y G),
mostrando cambios importantes en el eje de polaridad.

73
En conjunto estos resultados sugieren que dependiendo del estado de
diferenciación, las células acinares presentan distinta sensibilidad a la falta de
señales a través de la integrina α6. Esto último se puede concluir ya que tiempos de
bloqueo semejantes, pero aplicados en distintos períodos del curso temporal de la
generación del acino, provocaron efectos morfológicos distintos. Si el bloqueo se
realiza al final del cultivo (Figura 15B y C) no hay cambios en la polaridad celular tan
evidentes como cuando se realizan durante el tercer día de cultivo (Figura 16F y G).
Con estos experimentos se determinó que, tanto la duración del bloqueo de la
integrina α6, como el período en que los acinos inician su diferenciación, son
determinantes en el grado de la desorganización acinar. Mientras más temprano se
interrumpe la interacción de la integrina α6 con la lámina basal, más severo es el
efecto de la desorganización acinar. Sin embargo, las células son capaces de
“resistir” un bloqueo transitorio, ya que si este es removido, las señales se
reestablecen y se logran organizar acinos con un grado de organización normal
(Figura 16B, C, D y E).
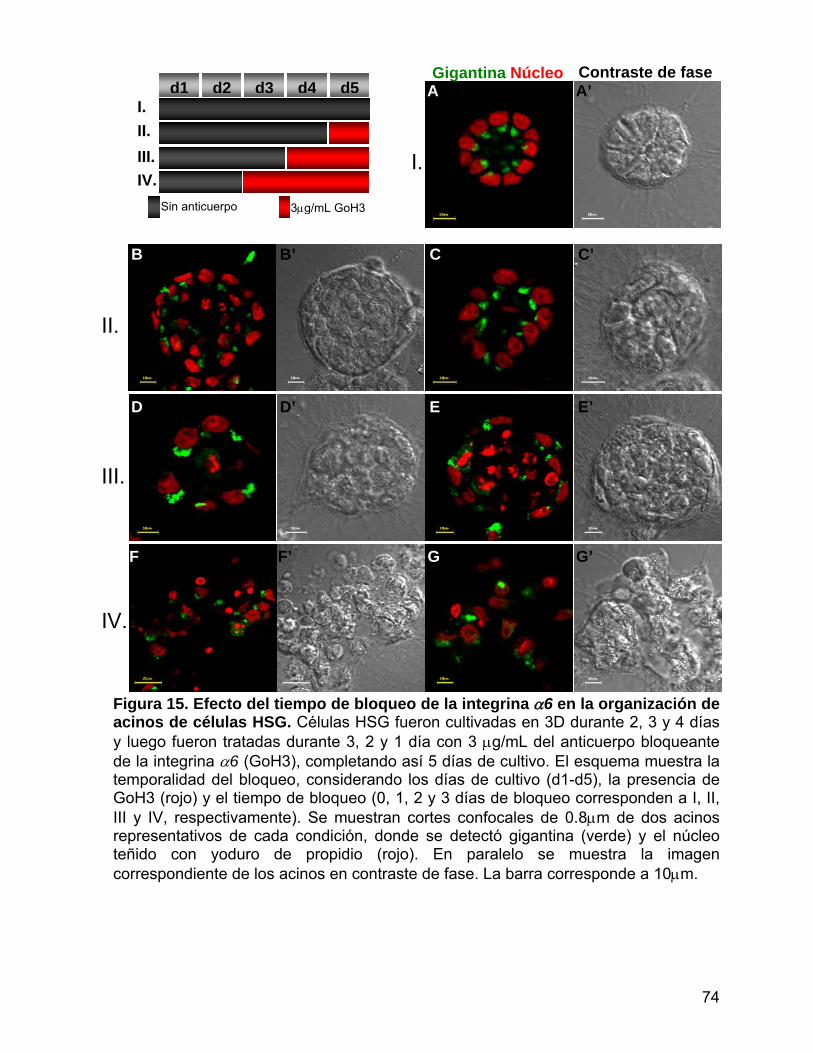
74
Figura 15. Efecto del tiempo de bloqueo de la integrina α6 en la organización de acinos de células HSG. Células HSG fueron cultivadas en 3D durante 2, 3 y 4 días y luego fueron tratadas durante 3, 2 y 1 día con 3 μg/mL del anticuerpo bloqueante de la integrina α6 (GoH3), completando así 5 días de cultivo. El esquema muestra la temporalidad del bloqueo, considerando los días de cultivo (d1-d5), la presencia de GoH3 (rojo) y el tiempo de bloqueo (0, 1, 2 y 3 días de bloqueo corresponden a I, II, III y IV, respectivamente). Se muestran cortes confocales de 0.8μm de dos acinos representativos de cada condición, donde se detectó gigantina (verde) y el núcleo teñido con yoduro de propidio (rojo). En paralelo se muestra la imagen correspondiente de los acinos en contraste de fase. La barra corresponde a 10μm.
I.
A A’
II.
III.
IV.
B B’ C C’
D D’ E E’
F F’ G G’
d1 d2 d3 d4 d5I. II. III. IV.
3μg/mL GoH3Sin anticuerpo
Gigantina Núcleo Contraste de fase

75
Figura 16. Efecto del tiempo de bloqueo pulsado de la integrina α6 en la organización de acinos de células HSG. Células HSG fueron cultivadas en 3D durante 2 días hasta la formación de acúmulos celulares. Luego fueron tratadas durante 1, 2 y 24h con 3 μg/mL del anticuerpo bloqueante de la integrina α6 (GoH3), tras lo cual se retiró del medio de cultivo y se continuó el cultivo en condiciones controles hasta completar los 5 días. El esquema muestra la temporalidad del bloqueo, considerando los días de cultivo (d1-d5), la presencia de GoH3 (rojo) y el tiempo de bloqueo (0, 1, 2 y 24 horas corresponde a I, II, III y IV, respectivamente). Se muestran cortes confocales de 0.8μm de dos acinos representativos de cada condición, donde se detectó gigantina (verde) y el núcleo teñido con yoduro de propidio (rojo). En paralelo se muestra la imagen correspondiente de los acinos en contraste de fase. La barra corresponde a 10μm.
A A’
I.
B B’ C C’
II.
D D’ E E’
III.
F F’ G G’
IV.
d1 d2 d3 d4 d5I. II. III. IV.
3μg/mL GoH3Sin anticuerpo
Gigantina Núcleo Contraste de fase

76
Dentro de los estudios morfológicos realizados a estos acinos desorganizados
por el bloqueo de la integrina α6, se determinó que esta desorganización se
correlaciona con cambios importantes en la distribución de actina polimerizada. Así,
en las células tratadas con el anticuerpo bloqueante, se observan acumulaciones
citoplasmáticas de actina polimerizada y en otras células, la actina delimita la
membrana plasmática, mostrando de esta manera los cambios en la forma celular
(Figura 17). Además, bajo estas condiciones, la señal citoplasmática para la integrina
β1 en algunos grupos de células se pierde, manteniéndose sólo la señal de la
membrana plasmática (Figura 18).
Todos estos resultados permiten plantear que dentro de las funciones que
tendrían las integrinas de la familia α6 en la organización de acinos se encuentran
además de la inducción de la diferenciación y polaridad, el control de la sobrevida
celular, donde pudiesen activarse señales de muerte celular. Este proceso de
desorganización de los acinos se correlaciona con cambios importantes en la
distribución de actina polimerizada y la integrina β1 en algunos grupos de células se
pierde.
Es importante destacar que la severidad del cambio morfológico depende del
tiempo y duración de la inhibición de la integrina α6, siendo las células más sensibles
durante el comienzo del establecimiento del fenotipo polarizado
.
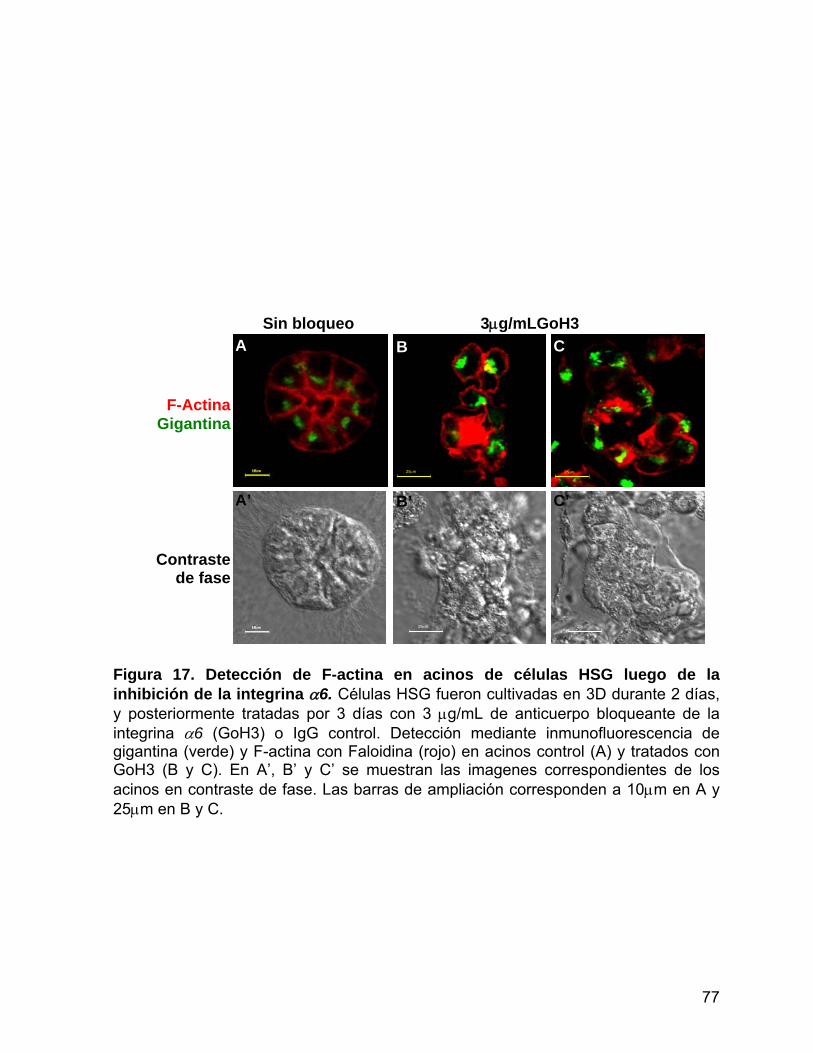
77
Figura 17. Detección de F-actina en acinos de células HSG luego de la inhibición de la integrina α6. Células HSG fueron cultivadas en 3D durante 2 días, y posteriormente tratadas por 3 días con 3 μg/mL de anticuerpo bloqueante de la integrina α6 (GoH3) o IgG control. Detección mediante inmunofluorescencia de gigantina (verde) y F-actina con Faloidina (rojo) en acinos control (A) y tratados con GoH3 (B y C). En A’, B’ y C’ se muestran las imagenes correspondientes de los acinos en contraste de fase. Las barras de ampliación corresponden a 10μm en A y 25μm en B y C.
F-Actina Gigantina
A’ B’ C’
A B C
Contraste de fase
25μm 25μm
Sin bloqueo 3μg/mLGoH3 Bl A I t i 6
25μm 25μm

78
Figura 18. Redistribución de la integrina β1 en acinos de células HSG como respuesta a la inhibición de la integrina α6. Células HSG fueron cultivadas en 3D durante 2 días y posteriormente tratadas por 3 días con 3 μg/mL de anticuerpo bloqueante de la integrina α6 (GoH3) o IgG control. Detección mediante inmunofluorescencia de la integrina β1 (verde) contra tinción nuclear con yoduro de propidio (rojo) en acinos control (A) y tratados con GoH3 (B y C). En A’, B’ y C’ se muestran las imagenes correspondientes de los acinos en contraste de fase Las barras de ampliación corresponden a 10μm en A y 25μm en B y C.
Sin bloqueo 3μg/mL GoH3 Bl A I t i 6
Integrina β1 Núcleo
B’ C’
B C
Contraste de fase
A’
10μm
A

79
V. Relación entre la señalización de la integrina α6 y las GTPasas de la familia
Rho en acinos de células HSG
La familia de las GTPasas de la familia Rho participan en el establecimiento
del fenotipo epitelial polarizado [31,59,65,66]. Considerando que el bloqueo de la
integrina α6 promueve la desorganización acinar y pérdida del fenotipo polarizado, se
evaluó si este bloqueo se relaciona con cambios en los niveles de RhoA, Rac1 y
Cdc42 activos. En acinos de células HSG se analizaron los niveles de estas
GTPasas activas, mediante ensayos de precipitación por afinidad y Westernblot
específicos, utilizando las proteínas de fusión PBD-GST (dominio de interacción a
Rac y Cdc42 de PAK1 fusionado a GST) [104] y RBD-GST (dominio de interacción a
RhoA de Rhotekina fusionado a GST) [105].
Se bloqueó la integrina α6 en acinos de células HSG de 5 días de cultivo,
durante un período de 30 minutos a 37ºC, e inmediatamente se determinaron las
fracciones totales y activas de RhoA, Rac1 y Cdc42. El propósito de este ensayo fue
evaluar el efecto directo de la inhibición de la integrina α6 en la activación de estas
GTPasas, descartando eventos compensatorios a desarrollarse durante los 5 días de
cultivo. Resultó difícil obtener datos reproducibles con esta técnica, realizándose
varias repeticiones con resultados dispares, considerándose para esta tesis aquellos
más consistentes.
En la Figura 19A se muestra una imagen representativa del Westernblot para
la determinación de los niveles de Cdc42 activo y total tras el bloqueo de la integrina
α6 (carril 3) en comparación con el efecto obtenido de utilizar una IgG como control
(carril 2). Además, se muestran los niveles basales de Cdc42 en condiciones de
cultivo en plano (carril 4) y en tres dimensiones (carril 1). En la Figura 19B y C, se
muestran los gráficos que comparan el valor relativo de Cdc42 según el tipo de
cultivo (cultivo en plano versus 3D) y el ensayo de bloqueo (sin bloqueo, control con
IgG y bloqueado con GoH3), respectivamente. Los valores promedio (± error
estándar) para cada condición fueron obtenidos de tres experimentos independientes
(n=3).

80
El Westernblot para Cdc42 muestra una diferencia en las intensidades de las
bandas correspondientes a Cdc42 activo entre el cultivo en plano y en 3D. Los
niveles relativos de Cdc42 activo disminuyen en promedio aproximadamente 9
veces, al comparar el cultivo en plano (31,1x105 ± 7,5x105) con el cultivo en 3D
(3,5x105 ± 1,8x105), pero estas diferencias no son estadísticamente significativas
(p=0.05). Por otra parte, en los acinos de células HSG, el promedio de los niveles
relativos de Cdc42 activo es de 113,3x105 ± 51,1x105 cuando se bloquea la integrina
α6 y 22,3x105 ± 17,0x105 al utilizar una IgG control, una diferencia que tampoco
resulta estadísticamente significativa (p=0,127). Por lo tanto considerando estos
resultados, no existirían diferencias en los niveles de Cdc42 activo tras el bloqueo de
la integrina α6. No hay que perder de vista que estos resultados son producto de un
bajo número de repeticiones, siendo necesario un mayor número de experimentos
para obtener un resultado concluyente.
Al analizar los resultados obtenidos para RhoA, tampoco se observan
diferencias estadísticamente significativas entre los niveles de RhoA activo en el
cultivo en plano con respecto al cultivo en 3D a partir de 2 experimentos
independientes (p=0,121), con valores promedios (± error estándar) de 13,2x104 ±
2,8x104 y 2,1x104 ± 1,6x104 respectivamente. Además, el bloqueo de la integrina α6
no genera cambios en los niveles de Rho activo, ya que se obtienen valores
promedios (± error estándar) de 20,9x104 ± 19,9x104 al utilizar el anticuerpo
bloqueante versus 10,7x104 ± 1,7x104 en la situación control. Estos datos no son
diferentes estadísticamente presentando un valor de p=1, lo que indica que
estadísticamente son semejantes. A partir de estos experimentos, podemos decir
que Rho A no estaría involucrada en el control del fenotipo polarizado dependiente
de la integrina α6, ni se requerirían variaciones necesarias para el establecimiento
del cultivo en 3D. Los resultados observados en Rac son bastante más dispares que
los obtenidos para Cdc42 y Rho, pues existe mayor variabilidad, no existe
reproducibilidad en ninguno de los datos obtenidos y hay una alta dispersión en ellos,
lo que genera un alto error estándar en todas las determinaciones. Así no existirían
resultados concluyentes para esta GTPasa, lo que queda claro al observar la grafica
(Figura 21).

81
Figura 19: Efecto del bloqueo de la integrina α6 en los niveles de Cdc42 activo. Células HSG fueron cultivadas en 3D durante 5 días y luego tratadas por 30 minutos con 3 μg/mL del anticuerpo bloqueante de la integrina α6, IgG control o sin anticuerpo. En el extracto celular se determinó los niveles de Cdc42 activo mediante un ensayo de precipitación por afinidad utilizando la proteína de fusión PBD-GST unida a partículas de Glutatión Sefarosa. (A) Se detectó mediante WB la fracción de Cdc42 activo, total y actina en los extractos de acinos de células HSG tratados con GoH3 (carril 3), IgG control (carril 2) y sin tratar (carril 1), junto a los niveles en las células HSG cultivadas en plano (carril 4). En B y C se muestran los gráficos que comparan los niveles relativos de Cdc42 activo presentes en el extracto según el cultivo (B) y las condiciones de bloqueo (C). Se normalizó la carga de Cdc42 total con los niveles de actina del extracto. Estos resultados representan el promedio de 3 experimentos independiente (promedio ± error estándar).
A
B
kDa
25
Cdc42 activo
(-) IgG control
GoH3 (-)
3D 2D
Cdc42 total
Actina
1 2 3 4
20
25
20
50
Cp=0,05 p=0,127
3D 2D0
5
10
15
20
25
30
35
40
Niv
eles
Rel
ativ
os d
e C
dc42
act
ivo/
tota
l (x1
05 )
3D 3D-IgG 3D-GoH3
0
20
40
60
80
100
120
140
160
180N
ivel
es R
elat
ivos
de
Cdc
42 a
ctiv
o/to
tal (
x105 )
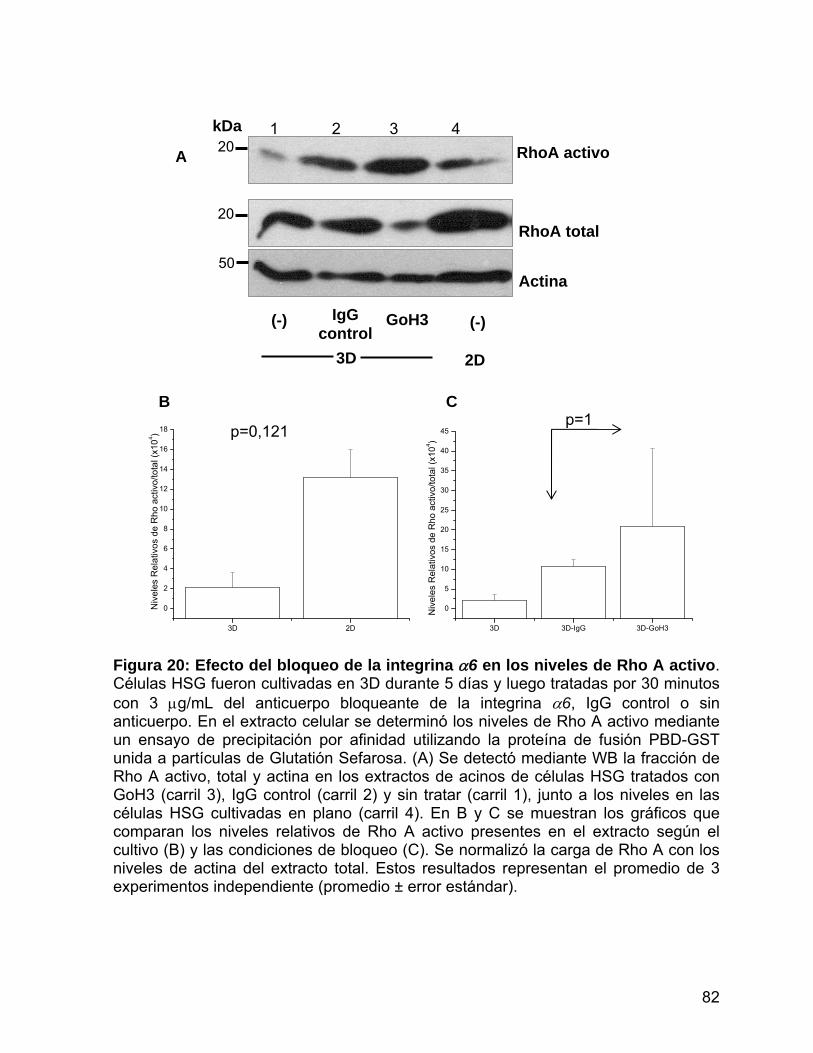
82
Figura 20: Efecto del bloqueo de la integrina α6 en los niveles de Rho A activo. Células HSG fueron cultivadas en 3D durante 5 días y luego tratadas por 30 minutos con 3 μg/mL del anticuerpo bloqueante de la integrina α6, IgG control o sin anticuerpo. En el extracto celular se determinó los niveles de Rho A activo mediante un ensayo de precipitación por afinidad utilizando la proteína de fusión PBD-GST unida a partículas de Glutatión Sefarosa. (A) Se detectó mediante WB la fracción de Rho A activo, total y actina en los extractos de acinos de células HSG tratados con GoH3 (carril 3), IgG control (carril 2) y sin tratar (carril 1), junto a los niveles en las células HSG cultivadas en plano (carril 4). En B y C se muestran los gráficos que comparan los niveles relativos de Rho A activo presentes en el extracto según el cultivo (B) y las condiciones de bloqueo (C). Se normalizó la carga de Rho A con los niveles de actina del extracto total. Estos resultados representan el promedio de 3 experimentos independiente (promedio ± error estándar).
RhoA activo
(-) IgG control
GoH3 (-)
3D 2D
RhoA total
Actina
1 2 3 4
A
B
kDa
20
50
20
C p=1
p=0,121
3D 2D
0
2
4
6
8
10
12
14
16
18
Niv
eles
Rel
ativ
os d
e R
ho a
ctiv
o/to
tal (
x104 )
3D 3D-IgG 3D-GoH3
0
5
10
15
20
25
30
35
40
45N
ivel
es R
elat
ivos
de
Rho
act
ivo/
tota
l (x1
04 )
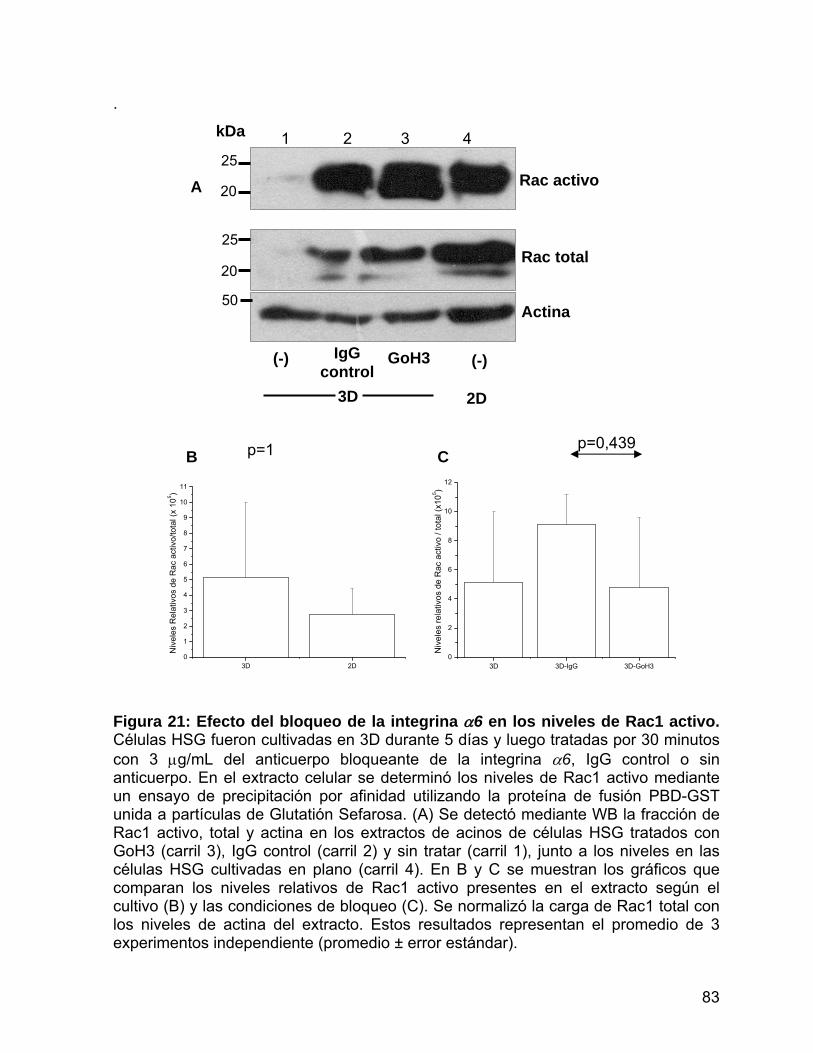
83
.
Figura 21: Efecto del bloqueo de la integrina α6 en los niveles de Rac1 activo. Células HSG fueron cultivadas en 3D durante 5 días y luego tratadas por 30 minutos con 3 μg/mL del anticuerpo bloqueante de la integrina α6, IgG control o sin anticuerpo. En el extracto celular se determinó los niveles de Rac1 activo mediante un ensayo de precipitación por afinidad utilizando la proteína de fusión PBD-GST unida a partículas de Glutatión Sefarosa. (A) Se detectó mediante WB la fracción de Rac1 activo, total y actina en los extractos de acinos de células HSG tratados con GoH3 (carril 3), IgG control (carril 2) y sin tratar (carril 1), junto a los niveles en las células HSG cultivadas en plano (carril 4). En B y C se muestran los gráficos que comparan los niveles relativos de Rac1 activo presentes en el extracto según el cultivo (B) y las condiciones de bloqueo (C). Se normalizó la carga de Rac1 total con los niveles de actina del extracto. Estos resultados representan el promedio de 3 experimentos independiente (promedio ± error estándar).
1 2 3 4
Rac activo
(-) IgG control
GoH3 (-)
3D 2D
Rac total
Actina
A
B
25
20
25
20
50
kDa
C p=0,439 p=1
3D 2D0
1
2
3
4
5
6
7
8
9
10
11
Niv
eles
Rel
ativ
os d
e R
ac a
ctiv
o/to
tal (
x 10
5 )
3D 3D-IgG 3D-GoH30
2
4
6
8
10
12
Niv
eles
rela
tivos
de
Rac
act
ivo
/ tot
al (x
105 )

84
DISCUSIÓN
El origen y desarrollo del síndrome de Sjögren son aún desconocidos, sin
embargo existen nuevas hipótesis para describir su etiopatogenia. Éstas no se
centran exclusivamente en la acción del infiltrado inflamatorio, sino que le dan un
papel importante a la célula epitelial como centro de la patología [17]. Se han descrito
cambios morfológicos en el parénquima de las glándulas salivales de los pacientes
con SS, que incluyen desorganización acinar, anoikis [21] y cambios importantes en
la polaridad celular [106], como resultado de la señales alteradas provenientes de
una lámina basal desorganizada [21,22]. Dada la importancia que tiene la correcta
polarización epitelial para el proceso secretor, la pregunta que se abordó en esta
tesis es: ¿Qué señales participan en el desarrollo de esta morfología alterada?. El
entendimiento adecuado de la generación y progresión de la enfermedad permitiría
eventualmente el desarrollo de tratamientos dirigidos hacia la causa del fenotipo seco
y no solo a tratar los síntomas de sequedad.
La discusión del trabajo realizado será presentada en cuatro secciones que
engloban los resultados obtenidos de GSL y del modelo de cultivo en 3D, los cuales
son los siguientes:
1. Relación de las integrinas α6 y β1 con el síndrome de Sjögren
2. Utilización del cultivo tridimensional de acinos de células HSG como modelo de
estudio de morfogénesis acinar y polaridad: ¿posible utilización como modelo
experimental para el síndrome de Sjögren?
3. Rol de la integrina α6 en la desorganización acinar observada en GSL de
pacientes con síndrome de Sjögren
4. Rol de la integrina α6 en el control de la sobrevida en GSL, ¿posible relación con
el desarrollo del síndrome de Sjögren?
5. Relación de las Rho GTPasas con la desorganización acinar presente en el SS y
con la señalización río abajo de la integrina α6

85
I. Relación de las integrinas α6 y β1 con el síndrome de Sjögren
La integrina β1 se expresa en la fracción epitelial de GSL de controles y
pacientes con SS, sin cambios evidentes en sus niveles proteicos (Figura 2). De
manera semejante, en extracto total de GSL de pacientes con SS no existen
diferencias en los niveles de la integrina β1, independiente de la cantidad de infiltrado
inflamatorio presente [44]. Las integrinas de la familia β1 mas ampliamente
expresadas son α5β1 y α6β1, selectivas para fibronectina y laminina, así como α2β1
y α3β1, que pueden unir múltiples ligandos tales como colágeno y laminina [107].
Debido a que en GSL humanas no se ha determinado cuales de estas integrinas se
encuentran más representadas, los niveles de la subunidad β1 detectados en esta
tesis representan a todos sus heterodímeros presentes en el componente epitelial de
GSL, por lo que la discusión se asociará a la familia de integrinas β1 en general.
Resulta interesante la aparición de una banda de β1 con mayor movilidad
electroforética exclusivamente en los pacientes con SS (Figura 2). Dado el bajo
número de individuos analizados, sería necesario realizar este análisis en un mayor
número de individuos, a fin de demostrar o descartar que la aparición de esta forma
de β1 sea un evento particular de los pacientes con SS.
Considerando la migración de la forma glicosilada y sin glicosilar de β1 [92], la
banda de menor movilidad electroforética pudiese corresponder a un estado no
glicosilado o parcialmente glicosilado, lo que requeriría de una comprobación
experimental. Por ejemplo, se podría tratar extractos epiteliales de GSL de individuos
controles con N-endoglicosidasas (ej. Endo-H) y luego analizarlos por Westernblot
contra β1, así debiese aparecer una banda de movilidad electroforética semejante a
la que aparece en pacientes con SS. Si este fuera el resultado, este ensayo
permitiría definir la presencia en pacientes con SS de una forma de la integrina β1
con residuos glicosilados disminuidos.
La probable existencia de esta integrina β1 “inmadura” en pacientes con SS
podría tener diversas implicancias en el comportamiento celular de GSL de los
pacientes. Se ha descrito que la modificación in vitro del patrón de glicosilación de

86
las integrinas altera su conformación y capacidad adhesiva a ligandos, dependiendo
del tipo celular, ligando e integrina involucrada. Por ejemplo, se ha reportado que en
la línea celular de neutrófilos HL60, el tratamiento con la exoglicosidasa
neuraminidasa, que elimina residuos de ácido siálico presentes en la integrina β1,
incrementa fuertemente su capacidad de unión a fibronectina [108]. Sin embargo, en
células de melanoma G361, la remoción del mismo tipo de residuos sialilados en la
integrina α5, inhibe su adhesión celular a fibronectina [109]. Considerando estas
evidencias, los posibles cambios en la glicosilación de las integrinas β1 presentes en
el componente epitelial de GSL de pacientes con SS, podrían modificar su capacidad
adhesiva a la lámina basal. Esta modificación podría ser parte del fenómeno de
desanclaje de la célula acinar a su lámina basal que se observa en las GSL de
pacientes con SS [21]. Así, la pérdida de anclaje celular en pacientes con SS podría
resultar, tanto de la degradación y desorganización del sustrato adhesivo presente en
la lámina basal [22,23,33,34], como de modificaciones postraduccionales en las
integrinas que actúan como receptores de componentes de lámina basal. Ensayos
de adhesión en placa de células de GSL aisladas desde controles, pacientes con SS
y controles tratados con N-endoglicosidasas permitirían evaluar in vitro esta
proposición.
Dado que las láminas basales son organizaciones complejas, formadas por la
asociación de múltiples proteínas [48], y en GSL se han descrito al menos cinco
lamininas que forman parte de ella [33], resulta importante tomar en cuenta que las
moléculas de adhesión que median la adhesión a esta lámina basal son múltiples.
Así, esta posible glicosilación alterada de la integrina β1 que pudiese generar
cambios adhesivos se sumaría a cambios en la localización de la integrina β4 que
han sido descritos para pacientes con SS [44]. También, cuando las alteraciones
morfológicas en acinos de pacientes con SS son importantes, es posible ver una
señal citoplasmática para la integrina β1 (Figura 3). Esto resulta importante ya que de
existir una fracción de la integrina β1 parcialmente glicosilada, solo si se localiza en
la superficie celular podría tener efectos negativos en los procesos de adhesión y
señalización de la célula epitelial de los pacientes con SS. Se requeriría determinar
qué fracción de esta integrina β1 se encuentra en superficie, la fracción madura o la

87
inmadura, por ejemplo mediante ensayos de inmunoprecipitación usando
previamente marcación de la superficie celular con biotina.
Considerando que la glicosilación diferencial de la integrina β1 ocurre durante
cambios fenotípicos celulares [107] como activación de keratinocitos [110],
diferenciación mieloide [111], y transformación oncogénica y metástasis [112], es
posible pensar que los cambios en la integrina β1 en pacientes con SS podrían ser
parte de modificaciones generalizadas en el patrón diferenciado de las células
acinares. Esto se relaciona con la modificación en los patrones de glicosilación de la
mucina salival MUC5B en pacientes con SS, donde el residuo glicosilado sulfo-
Lewisa se encuentra disminuido significativamente en los acinos mucosos [93].
Por otra parte, ya que las señales derivadas de la lámina basal a través de
integrinas son requeridas para adquirir el patrón diferenciado y polarizado [28], esta
glicosilación alterada de la integrina β1 también podría estar involucrada en la
desdiferenciación epitelial. Se ha descrito que la integrina β1 tendría un rol en el
desarrollo y diferenciación de glándula salival en humano, pues su expresión
aumenta a medida que progresa la morfogénesis glandular y es coincidente con la
aparición de marcadores de diferenciación [113]. Considerando estos antecedentes,
es plausible pensar que esta integrina podría también ser necesaria para la
mantención del fenotipo acinar diferenciado.
Estos cambios en la integrina β1, se deben asociar con los cambios en la
integrina α6 en los acinos de los pacientes con SS. Resulta interesante, que en el
grupo de los pacientes con SS estudiados existen comportamientos opuestos de
expresión de estas integrinas, ya que dos pacientes presentan una expresión normal
y en los otros la expresión está disminuida (Figura 2). Esto se relaciona con
evidencia previa donde se encontró una disminución muy significativa en los niveles
proteicos de la integrina α6 en extractos totales de GSL de 4/12 pacientes con SS
estudiados [44]. Se ha descrito que las citoquinas IL1-α y TNF-α disminuyen la
expresión de la integrina α6 en cultivos prolongados de sinoviocitos obtenidos de
pacientes con artritis reumatoide [114]. Además, TGFβ1 regula negativamente la
expresión de la integrina α6 en una línea celular epitelial de lente humana HLE B-3
[115] y en keratinocitos humanos normales [116]. Considerando que en acinos y

88
ductos de pacientes SS están sobre expresadas las citoquinas TGF-β1, IL-6, IL1-α,
INF-γ y TNF-α [117], la expresión diferencial de estas citoquinas en algunos
pacientes podrían promover esta disminución en los niveles de la integrina α6. Para
evaluar esta posibilidad, sería necesario probar que el tratamiento, in vitro, de células
derivadas de GSL de individuos controles o HSG con estas citoquinas, modula
directamente la expresión de la integrina α6.
Los resultados entregados en esta tesis, constituyen la primera evidencia de la
existencia del dímero α6β1 en GSL, donde su ligando laminina-111 se expresa en
láminas basales de acinos y ductos [22,33]. Las integrinas α6 y β1 co-localizan
preferentemente en los acinos serosos, donde se localizan en el dominio basal y
lateral de la membrana plasmática (Figura 3). Estos resultados indicarían que la
integrina α6β1 podría jugar un rol importante en las señales de este tipo celular
seroso. Los productos de secreción de los acinos serosos son diferentes a los
presentes en acinos mucosos. Resulta interesante que la otra integrina de la familia
α6, la integrina α6β4, también colocaliza preferentemente en células serosas [44].
Esto indicaría que a pesar que no han sido estudiadas las diferencias en la
composición de la lámina basal de estos tipos celulares, sí existirían diferencias en
los receptores involucrados con su señalización y adhesión. Lo cual podría modular
los procesos diferenciadores que resultan en la secreción de componente seroso o
mucoso.
En conjunto, todos estos cambios descritos en integrinas de la familia β1 y α6
podrían participar en el desarrollo de los procesos de desanclaje y desdiferenciación
celular. Esto debería correlacionarse con cambios en la señalización derivada de
estas integrinas, sin embargo, al determinar las señales de FAK fosforilada en
Tyr357 se observan niveles variables entre los distintos individuos, tanto controles
como pacientes, los cuales considerados como grupo no presentan diferencias
significativas. Es importante considerar que FAK es un indicador de la activación
inicial desencadenada por todas las integrinas, y dado que en el tejido la expresión
de α6β1 no es única, este resultado da cuenta de todas las señales activadas por
integrinas y de la transactivación dependiente de factores de crecimiento [51].
Además, resulta importante destacar que en células en cultivo, la fosforilación de

89
FAK en Tyr357 es un evento muy inicial en la cascada de integrinas, siendo su
fosforilación máxima determinada en los primeros minutos de unión de la integrina
con su ligando. En nuestras determinaciones estamos trabajando con un sistema
donde la interacción de proteínas de lámina basal con integrinas es un proceso
temporalmente estable, donde existen múltiples interacciones integrina-ligando.
II. Utilización del cultivo tridimensional de acinos de células HSG como modelo de estudio de morfogénesis acinar y polaridad: ¿posible utilización como modelo experimental para el síndrome de Sjögren?
En el SS existe una desorganización acinar y cambios en el fenotipo
polarizado de GSL, lo que se debería a alteraciones en las señales derivadas de la
lámina basal. ¿Qué modelo celular usar para estudiar estos cambios morfológicos?
Se sabe que factores como la arquitectura tejido específica, las señales mecánicas y
las comunicaciones célula/célula y célula/matriz extracelular se alteran en los
sistemas de cultivo en plano, limitando su potencial de predecir respuestas celulares
en organismos reales. Considerando estas limitantes, surgen los cultivos en 3D, los
cuales reestablecen las interacciones fisiológicas entre las células y la matriz
extracelular, pudiendo así emular la organización de tejidos reales mejor que los
cultivos convencionales en plano [71].
Los modelos de cultivo en 3D permiten la organización de estructuras tipo
acinos, a partir de células epiteliales crecidas inmersas en un gel semi-sólido de
proteínas de matriz extracelular, cuya composición determina el tipo de cultivo que se
obtiene [70]. Se utilizan proteínas puras (laminina-111 o colágeno tipo I) o extractos
de lámina basal que contienen proteínas (laminina, colágeno tipo IV, heparán sulfato
y nidógeno) [118] y factores de crecimiento (TGF-β, EGF, IGF1, bFGF y PDGF)
[119]. Este extracto de lámina basal es conocido comercialmente como matrigel,
cultrex o extracto EHS [74].
Uno de los logros importantes de esta tesis, fue implementar en nuestro
laboratorio un sistema de cultivo en 3D de células HSG en extracto de lámina basal.
En este cultivo, estas células adquieren una organización esférica que se asemeja a

90
los acinos de glándulas salivales, las unidades secretoras funcionales (Figura 5). Ya
que en GSL, la co-localización de las integrinas de la familia α6, α6β1 (Figura 3) y
α6β4 [44], es preferentemente observada en las células serosas del acino, resulta
importante que las células HSG son derivadas desde ductos intercalares de glándula
submandibular humana irradiada [89], e in vivo las células de ductos intercalares
tienen características pluripotentes y dan origen a células acinares serosas [88].
Además, la diferenciación acinar de células HSG depende de la acción de laminina-
111 [79], cuya red se encuentra desorganizada en la lámina basal de pacientes con
SS [22].
Se han descrito diversas metodologías para el cultivo de acinos de células
HSG en extractos de lámina basal, donde se obtienen diversas morfologías del
cultivo [78,79,98]. En esta tesis se utilizó un protocolo descrito para las células MCF-
10A, derivadas de glándula mamaria [77], donde las células se cultivan sobre una
base de extracto de lámina basal concentrado (Figura 1). Esta metodología de cultivo
permite la diferenciación morfológica de las células HSG, donde se da un cambio
fenotípico importante hacia un fenotipo diferenciado-polarizado.
Dentro de los elementos que permiten comprobar la adquisición de un fenotipo
polarizado, se encuentra el desarrollo de una lámina basal organizada alrededor de
los acinos. Si bien dentro del extracto de lámina basal se describe la presencia de
colágeno tipo IV [118], en los acinos de HSG se detecta una señal de colágeno tipo
IV restringida al dominio adyacente a la membrana plasmática basal (Figura 9). Esta
señal clara de colágeno tipo IV puede deberse a la formación de una red organizada
similar a la lámina basal en los tejidos [48], o tal vez sea producto de la secreción de
las propias células HSG, como se ha descrito en otros cultivos en 3D [120].
Otra característica importante para definir el fenotipo polarizado es la
formación del lumen acinar. El desarrollo del lumen es posterior a la polarización
ápico-basal y precede a la supresión proliferativa durante el desarrollo acinar [121].
Durante el desarrollo de glándula submandibular de ratón, la formación de lumen se
relaciona con la presencia de células apoptóticas [122], y en acinos mamarios su
formación depende de la apoptosis selectiva de las células localizadas centralmente
y que no están en contacto con la matriz extracelular [121]. En nuestros acinos, la

91
formación del lumen acinar se alcanzó al quinto día de cultivo, con presencia de
restos celulares luminales (Figura 6 y 10), dando así claras señales que el eje de
polaridad ápico-basal estaría definido.
Durante la diferenciación epitelial se da un cambio importante en la
organización nuclear, existiendo cambios en la localización de dominios de cromatina
condensada [121]. Este cambio se aprecia claramente en los acinos de células HSG
al ser comparados con el cultivo en 2D (Figura 7), donde el núcleo se asemeja más
al fenotipo de GSL in vivo. Asimismo, en células epiteliales polarizadas el complejo
de Golgi se localiza cerca del polo apical de la célula, en posición supra-nuclear [99],
localización que se observa en los acinos de células HSG.
También resulta importante la distribución asimétrica de proteínas de
membrana plasmática, lo que da cuenta de la definición de los dominios de
membrana plasmática apical y basolateral, que se observa en acinos de células
mamarias [77], MDCK [31,76] y por supuesto en GSL [25,26]. Dentro de las proteínas
que se analizaron para acinos de HSG, la integrina α6 presenta una localización
polarizada hacia el dominio de membrana plasmática basal (Figura 10).
Esta “separación” en dominios de la membrana plasmática se mantiene por la
formación de uniones estrechas [28]. La formación de uniones estrechas en acinos
de HSG es controversial. Nosotros encontramos que en los acinos de HSG se
organizan distintos tipos de adhesiones organizadas como desmosomas en botón de
distintos tamaños, extensas zonas de adhesión célula-célula tipo uniones
comunicantes y estructura tipo unión estrecha (Figura 8). Por otra parte, la
descripción de la expresión de ciertas proteínas de uniones estrechas como claudina
4 y ocludina resulta importante, ya que existen datos que indican que las células
HSG no son capaces de desarrollar uniones estrechas en cultivo en Transwell [123],
lo que es esencial para el normal funcionamiento del acino.
A pesar de la diferenciación morfológica observada en los acinos de HSG, no
se observó diferenciación funcional, ya que las células no sintetizan proteínas de
secreción como amilasa, cistatina o mucinas (Figura 8). Este resultado no era
esperado, ya que existen distintos trabajos que describen la expresión de amilasa en
cultivos de HSG sobre extractos de lámina basal [79,124]. Más aún, este extracto

92
promueve la expresión de marcadores de diferenciación en otros tipos celulares. Por
ejemplo, en células epiteliales mamarias murinas aumenta la expresión de caseína
[125], y en hepatocitos se promueve la expresión de albúmina [126].
La falta de expresión de proteínas de secreción en los acinos de HSG bajo las
condiciones establecidas en esta tesis, podría deberse a varios factores. Uno de
ellos corresponde a las diferencias en la condiciones del cultivo con las previamente
descritas. Se ha descrito recientemente que la concentración proteica de los
extractos de lámina basal son determinantes en promover la diferenciación
morfológica o funcional. Utilizar una alta concentración proteica induce la formación
de una red reticular y un incremento en expresión de amilasa, mientras que una baja
concentración de este extracto promueve la formación de estructuras complejas que
asemejan acinos asociados pero sin expresión de amilasa, planteando así que la
completa diferenciación morfológica y funcional parece difícil de alcanzar, ya que los
dos procesos tendrían requerimientos de cultivo opuestos [80]. Nuestro propósito era
lograr estructuras acinares discretas, sin formación de túbulos asociados, y esto
resultó en desmedro de la diferenciación funcional. Si bien se puede aumentar la
concentración de extracto de lámina basal en el cultivo, esto impide la formación de
estructuras acinares independientes, lo que era el fin del cultivo planteado a realizar.
En los primeros estudios que describen la formación de acinos de HSG, se
muestra la formación de estructuras acinares discretas sin ductos unidos similares a
las que nosotros obtuvimos, donde sí se expresa cistatina, a pesar de utilizar
extractos de lámina basal muy diluidos [79]. Esto resulta intrigante, ya que en los
preparados actuales de extracto de lámina basal, concentraciones tan diluidas de
estas proteínas no permiten la formación de un gel [127]. Esto podría indicar que las
propiedades fisicoquímicas del extracto de lámina basal actualmente en el mercado
es distinta, y por esto fue posible obtener diferenciación morfológica y funcional a la
vez en los experimentos iniciales [98].
Se ha demostrado que factores de crecimiento como EGF, TGF-α y HGF
incrementan la actividad del promotor de amilasa en células HSG [128]. También se
ha descrito la utilización de EGF, hidrocortisona, insulina y extracto de pituitaria en
cultivos primarios de células de glándulas salivales [129,130]. Considerando estos

93
antecedentes, se utilizaron estos factores para promover la expresión de amilasa,
pero no se generó ningún cambio en la expresión de las proteínas de secreción.
En el transcurso de esta tesis, las células se infectaron con micoplasma y
fueron tratadas con antibióticos, lo que puede haber alterado su patrón de expresión
génica. Al respecto, se ha descrito que la infección con esta bacteria intracelular
produce cambios en la expresión génica en células MCF-7 [131], MSB-1 y HD-11
[132].
A la luz de las características del cultivo en 3D de células HSG obtenido,
postulamos que este modelo se puede utilizar para estudios de morfología acinar y
diferenciación morfológica, pero no funcional a nivel de secreción. Por tanto, para ser
un modelo in vitro de secreción, las células deberían expresar alguna proteína de
secreción, además de mantener el fenotipo acinar.
Dentro de los modelos de estudio del SS se encuentran los ratones NOD que
desarrollan un fenotipo similar al SS [18,19], conjuntamente se han establecido
líneas celulares derivadas de glándulas salivales de pacientes con SS, pero
mantenidas en cultivo en 2D [133]. Esto permite plantear este sistema de cultivo de
acinos de células HSG como un modelo experimental útil para estudiar los cambios
morfológicos descritos en acinos de pacientes con SS.
III. Papel de la integrina α6 en la desorganización acinar observada en GSL de
pacientes con síndrome de Sjögren
En células HSG existe una expresión proteica diferencial de la integrina α6
entre las células cultivadas en 2D y 3D, expresándose sólo cuando las células están
organizando acinos, a pesar de la presencia del mensajero en las células cultivadas
en 2D (Figura 10). Se ha demostrado que cambios en el sustrato de adhesión tienen
efectos importantes en la biosíntesis celular. Por ejemplo, la adhesión de células
HSG a fibronectina o colágeno tipo I a través de integrinas β1, resulta en cambios en
el patrón de expresión de proteínas y mRNA [134]. En el caso particular de la
integrina α6, previamente se describió su ausencia de expresión en cultivos en 2D
[135], y se ha descrito su expresión en cultivos en 3D [79]. También en otros tipos

94
celulares se ha demostrado la influencia de la matriz extracelular en el patrón de
expresión génica. Por ejemplo, el cultivo de células epiteliales mamarias en
presencia de laminina y prolactina, induce la transcripción de β-caseína, de manera
dependiente de las integrinas α6 y β1 [136]. Por otra parte, la síntesis de la integrina
α6 en cultivos de glándula submandibular fetal de ratón aumenta como consecuencia
del tratamiento con EGF [137], uno de los factores de crecimiento presente en el
Matrigel [119]. Por lo tanto, tanto los componentes adhesivos, como los factores
solubles estarían regulando este cambio en la expresión génica.
Considerando que in vivo, las GSL expresan la integrina α6, y que sólo se
expresa en células HSG crecidas en 3D, se refuerza la importancia de esta integrina
para la organización tridimensional de los acinos y para la correcta expresión y
señalización de moléculas. Resulta importante considerar que al utilizar un producto
biológico como el matrigel, estamos ante la presencia de factores no controlables en
términos experimentales. Este compuesto se extrae a partir del sarcoma de ratón
EHS, donde la metodología de purificación puede ser más o menos eficaz. Quedó de
evidencia esto al demostrarse la presencia de la integrina α6 en el extracto de
matrigel, un producto o subproducto del proceso de purificación que no permite
asegurar cuales son los componentes finales de la matriz utilizada. Esto resulta
importante de tener en consideración para evaluar los factores que influyen el
desarrollo, ya que si no se hubiese evaluado la presencia de esta integrina en el
matrigel puro, la interpretación de los resultados hubiese estado errada.
En esta tesis se determinó que el cultivo de las células HSG en extracto de
lámina basal, en presencia de un anticuerpo inhibitorio de la integrina α6 desde el
inicio del cultivo, impide la formación de acinos, manteniéndose las células inmersas
en el matrigel como células aisladas (Figura 12). Esta evidencia se correlaciona con
evidencias de que el bloqueo de la integrina α6 en glándula submandibular en
desarrollo inhibe su ramificación [29]. Esto demuestra que la integrina α6 es
fundamental en la morfogénesis acinar inducida por laminina-111.
Se ha descrito que la adhesión de las células HSG al matrigel está mediada
por las integrinas β1, α2 y α6 [124]. En este sistema, la integrina β1 estaría
controlando la formación acinar, pues el uso de anticuerpos que bloquean su función

95
perturban la formación acinar [138] e inhiben la formación de estructuras tipo ductos
asociada con la diferenciación [124]. Además, el bloqueo de la integrina α6
disminuye el tamaño de los acinos formados [79]. Se ha determinado que sitios
ubicados en el dominio-G de la cadena α1 de laminina-111 contribuirían a la
diferenciación celular, que involucran su adhesión con integrinas de la familia β1 y
sindecán-1 [138]. Así, hay evidencias de varios receptores de adhesión involucrados
en el proceso de diferenciación acinar, que incluirían las integrina β1, sindecán-1 y la
integrina α6. Esto sugiere que la formación de acinos de células HSG en repuesta a
extractos de lámina basal es dependiente de integrinas de la familia α6, no
descartando que integrinas de otras familias estuviesen involucradas en el control del
proceso de diferenciación. Estos datos permiten complementar los datos de la
literatura que hablan de las moléculas de membrana plasmática importantes para la
organización de acinos en esta línea celular. O sea, la dependencia de laminina-111
para la diferenciación de acinos de células HSG dependería en parte de integrinas de
la familia α6.
En los pacientes con SS, los síntomas clínicos de la patología aparecen
preferentemente durante la etapa adulta, entre la cuarta y quinta década de la vida
[4], sin embargo, se desconoce cuándo comienza el desarrollo de la patología. Dado
que, los cambios morfológicos evidenciados en ratones NOD, un modelo animal de
SS, son descritos durante el desarrollo embrionario de la glándula salival [139],
podría darse que esta desorganización acinar partiera durante la morfogénesis acinar
como resultado de alteraciones en las señales derivadas de integrinas α6, β1 y
sindecán-1.
Por otra parte, si se desencadena la desorganización de la lámina basal y
posterior a ésto, se alteran sus señales a través de la integrina α6, ¿qué sucede con
el patrón diferenciado/polarizado?. Para poder evaluar esta pregunta, en esta tesis
estudiamos la influencia que tiene la integrina α6 en la mantención del fenotipo en
acinos en formación. Para esto, se permite a los acinos de células HSG adquirir un
cierto grado de organización y luego se bloquean las señales de la integrina α6,
evaluando si los acinos formados son capaces de mantener su fenotipo polarizado y

96
así comprobar, la real importancia de la integrina α6 en este proceso.
Cuando se bloquea la integrina α6 en acinos de células HSG de dos días de
cultivo, estos se desorganizan y se presentan como acúmulos celulares donde
incluso se ven células desagrupadas (Figura 12). Esta desorganización morfológica
se correlaciona con la ausencia de un eje de polaridad definido ya que la posición del
aparato de Golgi aparece completamente al azar con respecto al núcleo. Este tipo de
alteraciones se ha visto en acinos de células mamarias HMT-3522 crecidas en
matrigel, donde se observa una inhibición importante de la formación de estructuras
acinares al bloquear las integrinas β1 y α3, pero no al bloquear las integrina α2 o α6,
dando cuenta que el control de la morfogénesis acinar depende del tipo celular
involucrado [103].
En los acinos de células HSG logramos definir que, tanto la duración como el
estado de diferenciación de los acinos al momento de iniciar el bloqueo, son
determinantes en la intensidad de la desorganización acinar. Mientras más
tempranamente se interrumpe esta interacción, más importante y severo es el efecto
de la desorganización acinar (Figura 15 y 16). Es importante destacar que el reciclaje
de integrinas es un proceso altamente efectivo, y el ciclo endo-exocítico es bastante
rápido. Se ha planteado que la tasa de internalización y reciclaje de las integrinas es
alta y se daría cada 30 minutos [140]. Además se ha descrito que para integrinas de
la familia β3, esta internalización puede ser inducida por el uso de anticuerpos anti β3
[141]. Es por esto que los ensayos donde se bloquea la integrina α6 con anticuerpo y
luego se retira el medio que contiene este anticuerpo bloqueante permitirían
rápidamente reconstituir la situación control, dado el reciclaje del complejo integrina-
anticuerpo.
Es necesario aplicar otras estrategias experimentales mas directas que
permitan confirmar el efecto desorganizador de la estructura acinar que tiene la
pérdida de función de la integrina α6. Por ejemplo, se podría ver el efecto directo sin
la utilización de anticuerpos interferentes, aplicando una estrategia que elimine la
expresión de la integrina de manera endógena, por ejemplo mediante la utilización de
RNA de interferencia o la utilización de adenovirus que bloqueen su expresión. Es
importante que se requiere el control de la expresión de estos bloqueantes, ya que

97
por lo descrito en esta tesis, es importante controlar temporalmente la eliminación de
la integrina α6 para lograr la formación de los acinos.
Así, los resultados de esta tesis sugieren que en un estado de menor
diferenciación, las células acinares son más sensibles a la falta de señales a través
de la integrina α6, ya que bloqueos de la integrina α6 semejantes en duración, pero
distintos en temporalidad, resultan en efectos morfológicos diferentes.
¿Qué pasaría si esta desorganización de la lámina basal fuese un evento
transitorio, y las células tuviesen la capacidad de reestablecer los equilibrios en
cuanto a la señalización, o al equilibrio síntesis/degradación de la lámina basal Así, al
responder favorablemente, no se establecerían estos cambios en la polaridad y
desorganización. Sin embargo, si la homeostasis glandular está desbalanceada y no
puede reestablecerse ante algún tipo de injuria, se llegaría al fenotipo desorganizado.
Esto se observa muy claramente en los experimentos de bloqueo pulsado, donde en
períodos cortos de falta de señalización de la integrina α6, la célula aún responde
positivamente al desarrollo del fenotipo polarizado, sin embargo, si el bloqueo se
mantiene en el tiempo, se llega a la desorganización (Figura 16). Si pensamos que la
señal de la integrina α6 alterada da estos niveles de desorganización, ¿podríamos
pensar que la restitución de esta señal en particular disminuiría el fenómeno de la
alteración morfológica?
IV. Rol de la integrina α6 en el control de la sobrevida en GSL, ¿posible relación
con el desarrollo del síndrome de Sjögren?
Los acinos de células HSG cultivados en nuestro laboratorio se han mantenido
por períodos de siete días sin signos evidentes de muerte celular, a excepción de las
células que participan en el proceso de la formación del lumen. Esto se contradice
con evidencias recientemente publicadas, donde se describió que el cultivo de
células HSG en matrigel induce que las células pierdan su inmortalidad y desarrollen
apoptosis (entre los 4 y 5 días de cultivo). Esto estaría acompañado por la
desaparición gradual de la proteína antiapoptótica survivina, y un aumento de
actividad de la caspasa 3 [142]. Se ha descrito en cultivos que las células HSG

98
entran en apoptosis cuando son cultivadas con TNF-α [143], presencia de altos
niveles de Ca2+ [144] e inducción de especies reactivas del oxígeno [145]. Dado que
el cultivo realizado por estos autores utiliza matrigel depletado de factores de
crecimiento, y en nuestro sistema, donde la apoptosis no es un fenómeno que se
desencadene morfológicamente, utilizamos matrigel normal, pudiesen ser los
factores depletados los necesarios para lograr una mayor sobrevida.
Conjuntamente con la desorganización acinar, el bloqueo de la integrina α6 en
acinos de células HSG promueve la apoptosis celular (Figura 12). Esto podría
relacionarse con la cantidad disminuida de parénquima acinar presente en algunos
pacientes con SS [4]. Durante mucho tiempo se ha postulado que ciertos
mecanismos apoptóticos alterados estarían relacionados con el daño glandular que
existe en pacientes con SS [146], ya que se describe una mayor proporción de
moléculas pro-apoptóticas (CD95/Fas, Bax) que anti-apoptóticas (Bcl-2 y Bcl-X) en
células acinares y ductales de pacientes con SS [12,147]. Aunque existen algunos
estudios que son controversiales [148]. Si el bloqueo de la interacción de la integrina
α6 promueve la apoptosis en acinos de células HSG, podemos plantear que en la
progresión del SS una señalización alterada a través de la integrina α6 podría
desencadenar la anoikis y consiguiente pérdida de parénquima. En nuestros
pacientes la ocurrencia de anoikis ha sido demostrada [21]. En otros modelos en 3D
se han asociado bloqueo de integrinas con inducción de apoptosis, ya que la
presencia de anticuerpos bloqueantes de la integrina β1 en células mamarias
cultivadas en matrigel induce apoptosis, sugiriendo que en este sistema, la integrina
β1 transmite señales desde la matriz extracelular que son requeridas para la
sobrevida [103].
Se han asociado señales particulares, tanto de integrinas, como de moléculas
río abajo en el control de la sobrevida celular. La correcta formación de
hemidesmosomas por la integrina α6β4 y la mantención de la actividad de NFkB
fueron demostrados como importantes en la mediación de la resistencia ante la
apoptosis en acinos de células de mama [102]. La alteración de la función de FAK a
través de su degradación o por la competencia de otras proteínas promueve la
anoikis, también la alteración de los efectores de FAK y PI3K como las Rho GTPasas

99
y CAS, modulan la respuestas para la adhesión celular [52]. En células HSG, la
supervivencia celular y la morfología de las células no son afectadas por la inhibición
de PI3K durante la diferenciación [142].
Actualmente, no solo la observación morfológica es signo de la progresión de
fenómenos apoptóticos. Existen una serie de marcadores moleculares del fenómeno
que permitirían darle más consistencia a la determinación del fenómeno. Por
ejemplo, determinar la activación de caspasas iniciadoras o terminales mediante
ensayos de Westernblot, demostrar la degradación de DNA, ensayos de TUNEL, así
como evaluar la relación entre proteínas pro y antiapoptóticas, posteriormente al
bloqueo de la integrina α6. Además resultaría muy importante tratar de correlacionar
en acinos de pacientes SS, las zonas de desanclaje o desorganización morfológica
con un aumento de señales apoptóticas.
V. Relación de las Rho GTPasas con la desorganización acinar presente en el
SS y con la señalización río abajo de la integrina α6.
La desorganización acinar inducida por el bloqueo de la integrina α6 responde
a modificaciones en su señalización, pues al impedir la interacción con sus ligandos
de matriz extracelular, se bloquean las señales derivadas de estas interacciones.
Tratar de definir cuales señales estarían involucradas con este proceso resulta
importante al momento de relacionarla con la etiopatogenia del SS, ya que hasta el
momento, no existe ningún estudio que relacione los cambios morfológicos del SS
con señales particulares.
Las Rho GTPasas, RhoA, Rac y Cdc42 se han asociado con la definición del
fenotipo polarizado [59,65] y participan en la organización del citoesqueleto de actina
[56]. La actividad particular de cada una de estas proteínas se relaciona con la
formación de estructuras protrusivas (Rac1 y Cdc42) y la contractibilidad y
ensamblaje de adhesiones focales (RhoA) [56].
Estas estructuras particulares se dan en células cultivadas en 2D sobre una
placa, pero ¿qué sucede en el contexto de un tejido rodeado por matriz extracelular,
ya sea in vivo o en cultivos en 3D?. En esta tesis se intentó responder esta pregunta,

100
sin embargo dado dificultades metodológicas, y una baja reproducibilidad de los
experimentos, no se obtuvieron evidencias concluyentes para Cdc42. Con los
resultados de esta tesis, se determinó, sin embargo, que hay una tendencia a la
disminución en los niveles de Cdc42 activo en el cultivo de las células HSG en 3D
comparado con las células en 2D (Figura 19), pero las diferencias no son
estadísticamente significantes (p=0.05). Si se llegase a demostrar claramente esta
disminución, los resultados indicarían que sería necesario un estado de baja
activación de Cdc42 para lograr la organización acinar, determinada al quinto día de
cultivo.
En la definición de la organización del citoesqueleto, las células responden a
la composición de la matriz extracelular, pero además sensan y reaccionan ante sus
propiedades físicas que incluyen la tridimensionalidad y la rigidez [70,149]. Por lo
tanto, este posible estado “inhibido” de Cdc42 respondería a la suma de los eventos
producidos por estas dos condiciones. In vitro, las células epiteliales de mama
sensan la rigidez o densidad de su medio ambiente a través de la contractilidad
mediada por ROCK, y se requiere un descenso en la función de RhoA para que
ocurra la tubulogénesis [150]. Es interesante que la regulación hacia abajo de RhoA
a través de P190-B, una proteína GAP de Rho, se ha relacionado con el desarrollo
de la glándula mamaria regulando la formación de la yema terminal y la morfogénesis
ductal [151,152]. Esto sugiere que in vivo, una correcta formación de estructuras
diferenciadas en la mama requiere de la precisa regulación de la actividad de RhoA,
lo que es consistente con las observaciones in vitro.
Se ha determinado que las Rho GTPasas son fundamentales en la
formación acinar en células HSG, ya que al inactivarlas mediante la utilización de la
toxina A de Clostridium difficile, se impide la formación de acinos [78]. Este resultado
parece contradictorio a la posible inactivación de Cdc42 que nosotros describimos en
el cultivo de células HSG en 3D. Sin embargo, se ha descrito que la desregulación de
la actividad de RhoA, dada por la expresión de una forma constitutivamente activa o
un dominante negativo, perturba la tubulogénesis mamaria in vitro [150]. Además, en
otros estudio se ha visto que acinos de células epiteliales mamarias que expresan el
dominante negativo de Cdc42 desde el comienzo del cultivo, pierden su estructura

101
polarizada [153]. Esto indicaría que la formación acinar requeriría de la acción
regulada de estas GTPasas, por ello cualquier desbalance hacia la sobreactivación o
inhibición resulta en una formación acinar alterada.
Si consideramos el importante cambio que se observa en la morfología del
citoesqueleto de actina durante la transición desde el cultivo 2D hasta la formación
acinar (Figura 7), es esperable que se inhiba la formación acinar al bloquear las Rho
GTPasas. Una transición morfológica como ésta es resultado de modificaciones
drásticas en el citoesqueleto de actina, donde además se requieren procesos de
migración celular para la formación acinar de células HSG [79].
Además, es muy importante señalar que nuestras determinaciones se
realizaron al final del quinto día de cultivo, luego que las células acinares se han
“estabilizado” en el tiempo y el fenotipo polarizado ya está establecido. Es probable
que en los primeros estadíos de la formación acinar sean dependientes de la forma
activa de estas GTPasas para lograr los cambios en la estructura necesarios que
hacen pasar las células de un estado extendido sobre la placa, a uno menos
tensionado, para luego requerirse en estados basales activos más bajos.
La determinación de estas GTPasas resultó técnicamente difícil de reproducir,
ya que se obtuvieron resultados de muchos experimentos con datos contradictorios.
Finalmente solo se consideraron aquellos que presentaron más consistencia. Por
esto, con estos ensayos de bloqueo no es posible decir con certeza estadística que
existan cambios en los niveles de GTPasas activas. Se requiere un mayor número de
determinaciones para poder afirmar o descartar esto.
Para finalizar, es importante destacar que técnicamente existe una gran
variabilidad en la obtención de datos producto del ensayo de precipitación, sobre
todo en las condiciones de tratamiento con anticuerpos bloqueantes. En esta tesis se
incluyen los resultados más sólidos y reproducibles, ya que de otros experimentos
realizados, se obtuvieron resultados aún mas dispares. Por esta razón, las
repeticiones realizadas son menores. Esto indicaría que esta aproximación
experimental sería muy sensible a errores experimentales, dando cuenta de estos
resultados tan dispares. En general se ha planteado que el análisis bioquímico en
estos cultivos en 3D es mas difícil, dado que la densidad de las células es

102
aproximadamente dos ordenes de magnitud mas bajos que en los cultivos en 2D,
haciendo los análisis bioquímicos de proteínas presentes en baja abundancia,
problemáticos [154]. En estos ensayos fue necesario trabajar con varios pocillos de
cultivo para lograr determinar los niveles de estas proteínas activas. Por tanto, resta
decir que para corroborar estos posibles planteamientos para las GTPasas es
necesaria la obtención de un mayor número de repeticiones que permitan dar
sustentación estadística a las “posibilidades” que aquí se afirman.

103
CONCLUSIONES
1. En acinos de pacientes con SS se detectó una forma de la integrina β1 con menor
movilidad electroforética
2. En pacientes con SS la integrina α6 presenta niveles relativos variables. Algunos
pacientes tienen niveles de proteína más bajos.
3. No existen cambios globales en los niveles de FAK fosforilada en GSL de
pacientes con SS.
4. El sistema de cultivo en 3D de acinos de células HSG implementado durante esta
tesis parece ser un buen modelo de estudio de la morfogénesis acinar y fenotipo
polarizado, pero no permite el análisis de la secreción.
6. El bloqueo de la integrina α6 en acinos de células HSG induce una
desorganización acinar que depende del estado diferenciado de las células y del
tiempo de bloqueo.
8. La metodología utilizada en esta tesis no permite determinar de manera
concluyente si el bloqueo de la integrina α6 en acinos de células HSG modifica los
niveles de Rho GTPasas activas.

104
BIBLIOGRAFÍA 1. Delaleu, N., R. Jonsson, and M.M. Koller, Sjogren's syndrome. Eur J Oral Sci, 2005.
113(2): p. 101-13. 2. Ramos-Casals, M. and J. Font, Primary Sjogren's syndrome: current and emergent
aetiopathogenic concepts. Rheumatology (Oxford), 2005. 44(11): p. 1354-67. 3. Anaya, J., et al., Síndrome de Sjögren, in Autoinmunidad y Enfermedad Autoinmune,
C.p.I. Biológicas, Editor. 2005. 4. Anaya, J., M. Ramos, and M. García, Síndrome de Sjögren. 2001: Corporación para
Investigaciones Biológicas. 5. Routsias, J.G. and A.G. Tzioufas, Sjogren's syndrome--study of autoantigens and
autoantibodies. Clin Rev Allergy Immunol, 2007. 32(3): p. 238-51. 6. Vitali, C., et al., Classification criteria for Sjogren's syndrome: a revised version of the
European criteria proposed by the American-European Consensus Group. Ann Rheum Dis, 2002. 61(6): p. 554-8.
7. Fox, P.C., Autoimmune diseases and Sjogren's syndrome: an autoimmune exocrinopathy. Ann N Y Acad Sci, 2007. 1098: p. 15-21.
8. Fox, R.I. and M. Stern, Sjogren's syndrome: mechanisms of pathogenesis involve interaction of immune and neurosecretory systems. Scand J Rheumatol Suppl, 2002(116): p. 3-13.
9. Mitsias, D.I., E.K. Kapsogeorgou, and H.M. Moutsopoulos, Sjogren's syndrome: why autoimmune epithelitis? Oral Dis, 2006. 12(6): p. 523-32.
10. al-Hashimi, I., The management of Sjogren's syndrome in dental practice. J Am Dent Assoc, 2001. 132(10): p. 1409-17; quiz 1460-1.
11. Mavragani, C.P., N.M. Moutsopoulos, and H.M. Moutsopoulos, The management of Sjogren's syndrome. Nat Clin Pract Rheumatol, 2006. 2(5): p. 252-61.
12. Kong, L., et al., Fas and Fas ligand expression in the salivary glands of patients with primary Sjogren's syndrome. Arthritis Rheum, 1997. 40(1): p. 87-97.
13. Konttinen, Y.T., et al., Peptide-containing nerves in labial salivary glands in Sjogren's syndrome. Arthritis Rheum, 1992. 35(7): p. 815-20.
14. Main, C., P. Blennerhassett, and S.M. Collins, Human recombinant interleukin 1 beta suppresses acetylcholine release from rat myenteric plexus. Gastroenterology, 1993. 104(6): p. 1648-54.
15. Waterman, S.A., T.P. Gordon, and M. Rischmueller, Inhibitory effects of muscarinic receptor autoantibodies on parasympathetic neurotransmission in Sjogren's syndrome. Arthritis Rheum, 2000. 43(7): p. 1647-54.
16. Bacman, S., et al., Autoantibodies against lacrimal gland M3 muscarinic acetylcholine receptors in patients with primary Sjogren's syndrome. Invest Ophthalmol Vis Sci, 1998. 39(1): p. 151-6.
17. Mitsias, D.I., E.K. Kapsogeorgou, and H.M. Moutsopoulos, The role of epithelial cells in the initiation and perpetuation of autoimmune lesions: lessons from Sjogren's syndrome (autoimmune epithelitis). Lupus, 2006. 15(5): p. 255-61.
18. Humphreys-Beher, M.G., et al., Salivary gland changes in the NOD mouse model for Sjogren's syndrome: is there a non-immune genetic trigger? Eur J Morphol, 1998. 36 Suppl: p. 247-51.
19. Cha, S., A.B. Peck, and M.G. Humphreys-Beher, Progress in understanding autoimmune exocrinopathy using the non-obese diabetic mouse: an update. Crit Rev Oral Biol Med, 2002. 13(1): p. 5-16.

105
20. Yamachika, S., et al., Excessive synthesis of matrix metalloproteinases in exocrine tissues of NOD mouse models for Sjogren's syndrome. J Rheumatol, 1998. 25(12): p. 2371-80.
21. Goicovich, E., et al., Enhanced degradation of proteins of the basal lamina and stroma by matrix metalloproteinases from the salivary glands of Sjogren's syndrome patients: correlation with reduced structural integrity of acini and ducts. Arthritis Rheum, 2003. 48(9): p. 2573-84.
22. Molina, C., et al., Basal lamina disorganisation of the acini and ducts of labial salivary glands from patients with Sjogren's syndrome: association with mononuclear cell infiltration. Ann Rheum Dis, 2006. 65(2): p. 178-83.
23. Perez, P., et al., Differential expression of matrix metalloproteinases in labial salivary glands of patients with primary Sjogren's syndrome. 2000. 43(12): p. 2807-2817.
24. Steinfeld, S., et al., Abnormal distribution of aquaporin-5 water channel protein in salivary glands from Sjogren's syndrome patients. Lab Invest, 2001. 81(2): p. 143-8.
25. Castro, I., Niveles de expresión y localización subcelular de MUC1 en glándulas salivales labiales de pacientes son síndrome de Sjögren. 2005, Universidad de Chile.
26. Perez, P., Mecanismo de disfunción secretora en glándulas salivales de pacientes con síndrome de Sjögren: participación de ezrina en la desorganización del polo apical de células acinares. 2007, Universidad de Chile.
27. Nelson, W.J., Epithelial cell polarity from the outside looking in. News Physiol Sci, 2003. 18: p. 143-6.
28. Yeaman, C., K.K. Grindstaff, and W.J. Nelson, New perspectives on mechanisms involved in generating epithelial cell polarity. Physiol Rev, 1999. 79(1): p. 73-98.
29. Kadoya, Y., et al., Antibodies against domain E3 of laminin-1 and integrin alpha 6 subunit perturb branching epithelial morphogenesis of submandibular gland, but by different modes. J Cell Biol, 1995. 129(2): p. 521-34.
30. Schuger, L., et al., Laminin fragment E4 inhibition studies: basement membrane assembly and embryonic lung epithelial cell polarization requires laminin polymerization. Int J Dev Biol, 1998. 42(2): p. 217-20.
31. O'Brien, L.E., et al., Rac1 orientates epithelial apical polarity through effects on basolateral laminin assembly. Nat Cell Biol, 2001. 3(9): p. 831-8.
32. Gudjonsson, T., et al., Normal and tumor-derived myoepithelial cells differ in their ability to interact with luminal breast epithelial cells for polarity and basement membrane deposition. J Cell Sci, 2002. 115(Pt 1): p. 39-50.
33. Laine, M., et al., Segment-specific but pathologic laminin isoform profiles in human labial salivary glands of patients with Sjogren's syndrome. Arthritis Rheum, 2004. 50(12): p. 3968-73.
34. Kwon, Y.J., et al., Involvement of specific laminins and nidogens in the active remodeling of the basal lamina of labial salivary glands from patients with Sjogren's syndrome. Arthritis Rheum, 2006. 54(11): p. 3465-75.
35. Colognato, H., D.A. Winkelmann, and P.D. Yurchenco, Laminin polymerization induces a receptor-cytoskeleton network. J Cell Biol, 1999. 145(3): p. 619-31.
36. Kadoya, Y., et al., Importance of nidogen binding to laminin gamma1 for branching epithelial morphogenesis of the submandibular gland. Development, 1997. 124(3): p. 683-91.
37. Tunggal, J., et al., Expression of the nidogen-binding site of the laminin gamma1 chain disturbs basement membrane formation and maintenance in F9 embryoid bodies. J Cell Sci, 2003. 116(Pt 5): p. 803-12.
38. Ekblom, P., P. Lonai, and J.F. Talts, Expression and biological role of laminin-1. Matrix Biol, 2003. 22(1): p. 35-47.

106
39. Brakebusch, C., et al., Integrins in invasive growth. J Clin Invest, 2002. 109(8): p. 999-1006.
40. Humphries, M.J., et al., Mechanisms of integration of cells and extracellular matrices by integrins. Biochem Soc Trans, 2004. 32(Pt 5): p. 822-5.
41. Belkin, A.M. and M.A. Stepp, Integrins as receptors for laminins. Microsc Res Tech, 2000. 51(3): p. 280-301.
42. Miner, J.H. and P.D. Yurchenco, Laminin functions in tissue morphogenesis. Annu Rev Cell Dev Biol, 2004. 20: p. 255-84.
43. Fernandez, C., et al., Regulation of the extracellular ligand binding activity of integrins. Front Biosci, 1998. 3: p. d684-700.
44. Velozo, J., et al., Severe alterations in expression and localization of {alpha}6{beta}4 integrin in salivary gland acini from Sjogren's syndrome patients. Ann Rheum Dis, 2008.
45. Loducca, S.V., et al., Salivary gland tumours: immunoexpression of integrins beta 1, beta 3 and beta 4. J Oral Pathol Med, 2003. 32(5): p. 305-9.
46. Lazowski, K.W., et al., Temporal and spatial expression of laminin, collagen types IV and I and alpha 6/beta 1 integrin receptor in the developing rat parotid gland. Differentiation, 1994. 56(1-2): p. 75-82.
47. Giancotti, F.G. and E. Ruoslahti, Integrin signaling. Science, 1999. 285(5430): p. 1028-32.
48. Sasaki, T., R. Fassler, and E. Hohenester, Laminin: the crux of basement membrane assembly. J Cell Biol, 2004. 164(7): p. 959-63.
49. Giancotti, F.G. and G. Tarone, Positional control of cell fate through joint integrin/receptor protein kinase signaling. Annu Rev Cell Dev Biol, 2003. 19: p. 173-206.
50. Boudreau, N.J. and P.L. Jones, Extracellular matrix and integrin signalling: the shape of things to come. Biochem J, 1999. 339 ( Pt 3): p. 481-8.
51. Parsons, J.T., Focal adhesion kinase: the first ten years. J Cell Sci, 2003. 116(Pt 8): p. 1409-16.
52. Reddig, P.J. and R.L. Juliano, Clinging to life: cell to matrix adhesion and cell survival. Cancer Metastasis Rev, 2005. 24(3): p. 425-39.
53. Bishop, A.L. and A. Hall, Rho GTPases and their effector proteins. Biochem J, 2000. 348 Pt 2: p. 241-55.
54. Burridge, K. and K. Wennerberg, Rho and Rac take center stage. Cell, 2004. 116(2): p. 167-79.
55. Hall, A., Rho GTPases and the actin cytoskeleton. Science, 1998. 279(5350): p. 509-14.
56. Etienne-Manneville, S. and A. Hall, Rho GTPases in cell biology. Nature, 2002. 420(6916): p. 629-35.
57. Mackay, D.J. and A. Hall, Rho GTPases. J Biol Chem, 1998. 273(33): p. 20685-8. 58. Schwartz, M., Rho signalling at a glance. J Cell Sci, 2004. 117(Pt 23): p. 5457-8. 59. Liu, H., et al., Polarity and proliferation are controlled by distinct signaling pathways
downstream of PI3-kinase in breast epithelial tumor cells. J Cell Biol, 2004. 164(4): p. 603-12.
60. Jou, T.S., et al., Selective alterations in biosynthetic and endocytic protein traffic in Madin-Darby canine kidney epithelial cells expressing mutants of the small GTPase Rac1. Mol Biol Cell, 2000. 11(1): p. 287-304.
61. Ohno, S., Intercellular junctions and cellular polarity: the PAR-aPKC complex, a conserved core cassette playing fundamental roles in cell polarity. Curr Opin Cell Biol, 2001. 13(5): p. 641-8.

107
62. Suzuki, A., et al., Atypical protein kinase C is involved in the evolutionarily conserved par protein complex and plays a critical role in establishing epithelia-specific junctional structures. J Cell Biol, 2001. 152(6): p. 1183-96.
63. Joberty, G., et al., The cell-polarity protein Par6 links Par3 and atypical protein kinase C to Cdc42. Nat Cell Biol, 2000. 2(8): p. 531-9.
64. Yamanaka, T., et al., PAR-6 regulates aPKC activity in a novel way and mediates cell-cell contact-induced formation of the epithelial junctional complex. Genes Cells, 2001. 6(8): p. 721-31.
65. Chen, X. and I.G. Macara, Par-3 controls tight junction assembly through the Rac exchange factor Tiam1. Nat Cell Biol, 2005. 7(3): p. 262-9.
66. Nishimura, T., et al., PAR-6-PAR-3 mediates Cdc42-induced Rac activation through the Rac GEFs STEF/Tiam1. Nat Cell Biol, 2005. 7(3): p. 270-7.
67. Walsh, S.V., et al., Rho kinase regulates tight junction function and is necessary for tight junction assembly in polarized intestinal epithelia. Gastroenterology, 2001. 121(3): p. 566-79.
68. Menko, A.S., et al., Loss of alpha3beta1 integrin function results in an altered differentiation program in the mouse submandibular gland. Dev Dyn, 2001. 220(4): p. 337-49.
69. Menko, A.S., et al., Regulation of cadherin junctions during mouse submandibular gland development. Dev Dyn, 2002. 224(3): p. 321-33.
70. Yamada, K.M. and E. Cukierman, Modeling tissue morphogenesis and cancer in 3D. Cell, 2007. 130(4): p. 601-10.
71. Pampaloni, F., E.G. Reynaud, and E.H. Stelzer, The third dimension bridges the gap between cell culture and live tissue. Nat Rev Mol Cell Biol, 2007. 8(10): p. 839-45.
72. Discher, D.E., P. Janmey, and Y.L. Wang, Tissue cells feel and respond to the stiffness of their substrate. Science, 2005. 310(5751): p. 1139-43.
73. Cukierman, E., R. Pankov, and K.M. Yamada, Cell interactions with three-dimensional matrices. Curr Opin Cell Biol, 2002. 14(5): p. 633-9.
74. Kleinman, H.K. and G.R. Martin, Matrigel: basement membrane matrix with biological activity. Semin Cancer Biol, 2005. 15(5): p. 378-86.
75. Schmeichel, K.L. and M.J. Bissell, Modeling tissue-specific signaling and organ function in three dimensions. J Cell Sci, 2003. 116(Pt 12): p. 2377-88.
76. Yu, W., et al., Beta1-integrin orients epithelial polarity via Rac1 and laminin. Mol Biol Cell, 2005. 16(2): p. 433-45.
77. Debnath, J., S.K. Muthuswamy, and J.S. Brugge, Morphogenesis and oncogenesis of MCF-10A mammary epithelial acini grown in three-dimensional basement membrane cultures. Methods, 2003. 30(3): p. 256-68.
78. Crema, V.O., D.E. Hamassaki, and M.F. Santos, Small Rho GTPases are important for acinus formation in a human salivary gland cell line. Cell Tissue Res, 2006. 325(3): p. 493-500.
79. Hoffman, M.P., et al., Role of laminin-1 and TGF-beta 3 in acinar differentiation of a human submandibular gland cell line (HSG). J Cell Sci, 1996. 109 ( Pt 8): p. 2013-21.
80. Vag, J., et al., Morphological and functional differentiation of HSG cells: role of extracellular matrix and trpc 1. J Cell Physiol, 2007. 212(2): p. 416-23.
81. Chopra, A., B. Maitra, and N.J. Korman, Decreased mRNA expression of several basement membrane components in basal cell carcinoma. J Invest Dermatol, 1998. 110(1): p. 52-6.
82. Choudhury, A., et al., Retinoic acid-dependent transforming growth factor-beta 2-mediated induction of MUC4 mucin expression in human pancreatic tumor cells follows retinoic acid receptor-alpha signaling pathway. J Biol Chem, 2000. 275(43): p. 33929-36.

108
83. Joraku, A., et al., Tissue engineering of functional salivary gland tissue. Laryngoscope, 2005. 115(2): p. 244-8.
84. Daniels, T.E., Labial salivary gland biopsy in Sjogren's syndrome. Assessment as a diagnostic criterion in 362 suspected cases. Arthritis Rheum, 1984. 27(2): p. 147-56.
85. Gonzalez, M., N. Galanti, and R.O. Lopez, Changes in the content and distribution of sialic acid on the basal surface of isoproterenol-stimulated mouse parotid acinar cells. Eur J Cell Biol, 1989. 49(1): p. 171-9.
86. Bradford, M.M., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, 1976. 72: p. 248-54.
87. Laemmli, U.K., Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 1970. 227(5259): p. 680-5.
88. Dobrosielski-Vergona, K., Biology of the Salivary Glands. 1993. 89. Shirasuna, K., M. Sato, and T. Miyazaki, A neoplastic epithelial duct cell line
established from an irradiated human salivary gland. Cancer, 1981. 48(3): p. 745-52. 90. Willingham, M.C. and A.V. Rutherford, The use of osmium-thiocarbohydrazide-
osmium (OTO) and ferrocyanide-reduced osmium methods to enhance membrane contrast and preservation in cultured cells. J Histochem Cytochem, 1984. 32(4): p. 455-60.
91. Wei, J., L.M. Shaw, and A.M. Mercurio, Integrin signaling in leukocytes: lessons from the alpha6beta1 integrin. J Leukoc Biol, 1997. 61(4): p. 397-407.
92. Bellis, S.L., E. Newman, and E.A. Friedman, Steps in integrin beta1-chain glycosylation mediated by TGFbeta1 signaling through Ras. J Cell Physiol, 1999. 181(1): p. 33-44.
93. Alliende, C., et al., Reduced sulfation of MUC5B is linked to xerostomia in Sjogren's Syndrome patients. Ann Rheum Dis, 2007.
94. Hynes, R.O., Integrins: bidirectional, allosteric signaling machines. Cell, 2002. 110(6): p. 673-87.
95. Giancotti, F.G., Complexity and specificity of integrin signalling. Nat Cell Biol, 2000. 2(1): p. E13-4.
96. Azuma, M., et al., Effects of retinoic acid on morphological features and biological markers of a neoplastic human salivary intercalated duct cell line in culture. Cancer Res, 1988. 48(24 Pt 1): p. 7219-25.
97. Azuma, M., et al., Emergence of differentiated subclones from a human salivary adenocarcinoma cell clone in culture after treatment with sodium butyrate. Cancer Res, 1986. 46(2): p. 770-7.
98. Vag, J., et al., Morphological and functional differentiation of HSG cells: Role of extracellular matrix and trpc 1. J Cell Physiol, 2007.
99. Polishchuk, R.S. and A.A. Mironov, Structural aspects of Golgi function. Cell Mol Life Sci, 2004. 61(2): p. 146-58.
100. Linstedt, A.D. and H.P. Hauri, Giantin, a novel conserved Golgi membrane protein containing a cytoplasmic domain of at least 350 kDa. Mol Biol Cell, 1993. 4(7): p. 679-93.
101. Matlin, K.S., B. Haus, and A. Zuk, Integrins in epithelial cell polarity: using antibodies to analyze adhesive function and morphogenesis. Methods, 2003. 30(3): p. 235-46.
102. Weaver, V.M., et al., beta4 integrin-dependent formation of polarized three-dimensional architecture confers resistance to apoptosis in normal and malignant mammary epithelium. Cancer Cell, 2002. 2(3): p. 205-16.
103. Howlett, A.R., et al., Cellular growth and survival are mediated by beta 1 integrins in normal human breast epithelium but not in breast carcinoma. J Cell Sci, 1995. 108 ( Pt 5): p. 1945-57.

109
104. Bagrodia, S., et al., A novel regulator of p21-activated kinases. J Biol Chem, 1998. 273(37): p. 23633-6.
105. Ren, X.D., W.B. Kiosses, and M.A. Schwartz, Regulation of the small GTP-binding protein Rho by cell adhesion and the cytoskeleton. Embo J, 1999. 18(3): p. 578-85.
106. Ewert, P., Expresión y localización de proteínas de uniones estrechas (Tight Junctions) de células acinares de glándulas salivales de pacientes con síndrome de Sjogren. 2006, Universidad de Chile.
107. Bellis, S.L., Variant glycosylation: an underappreciated regulatory mechanism for beta1 integrins. Biochim Biophys Acta, 2004. 1663(1-2): p. 52-60.
108. Pretzlaff, R.K., V.W. Xue, and M.E. Rowin, Sialidase treatment exposes the beta1-integrin active ligand binding site on HL60 cells and increases binding to fibronectin. Cell Adhes Commun, 2000. 7(6): p. 491-500.
109. Nadanaka, S., et al., Occurrence of oligosialic acids on integrin alpha 5 subunit and their involvement in cell adhesion to fibronectin. J Biol Chem, 2001. 276(36): p. 33657-64.
110. Kim, L.T., et al., Altered glycosylation and cell surface expression of beta 1 integrin receptors during keratinocyte activation. J Cell Sci, 1992. 103 ( Pt 3): p. 743-53.
111. Semel, A.C., et al., Hyposialylation of integrins stimulates the activity of myeloid fibronectin receptors. J Biol Chem, 2002. 277(36): p. 32830-6.
112. Asada, M., et al., Increased expression of highly branched N-glycans at cell surface is correlated with the malignant phenotypes of mouse tumor cells. Cancer Res, 1997. 57(6): p. 1073-80.
113. Lourenco, S.V., et al., Expression of beta-1 integrin in human developing salivary glands and its parallel relation with maturation markers: in situ hybridisation and immunofluorescence study. Arch Oral Biol, 2007. 52(11): p. 1064-71.
114. Rinaldi, N., et al., Loss of laminin and of the laminin receptor integrin subunit alpha 6 in situ correlates with cytokine induced down regulation of alpha 6 on fibroblast-like synoviocytes from rheumatoid arthritis. Ann Rheum Dis, 1998. 57(9): p. 559-65.
115. Lim, J.M., et al., Downregulated expression of integrin alpha6 by transforming growth factor-beta(1) on lens epithelial cells in vitro. Biochem Biophys Res Commun, 2001. 284(1): p. 33-41.
116. Decline, F., et al., Keratinocyte motility induced by TGF-beta1 is accompanied by dramatic changes in cellular interactions with laminin 5. Cell Motil Cytoskeleton, 2003. 54(1): p. 64-80.
117. Sun, D., M.R. Emmert-Buck, and P.C. Fox, Differential cytokine mRNA expression in human labial minor salivary glands in primary Sjogren's syndrome. Autoimmunity, 1998. 28(3): p. 125-37.
118. Kleinman, H.K., et al., Basement membrane complexes with biological activity. Biochemistry, 1986. 25(2): p. 312-8.
119. Vukicevic, S., et al., Identification of multiple active growth factors in basement membrane Matrigel suggests caution in interpretation of cellular activity related to extracellular matrix components. Exp Cell Res, 1992. 202(1): p. 1-8.
120. Petersen, O.W., et al., Interaction with basement membrane serves to rapidly distinguish growth and differentiation pattern of normal and malignant human breast epithelial cells. Proc Natl Acad Sci U S A, 1992. 89(19): p. 9064-8.
121. Debnath, J., et al., The role of apoptosis in creating and maintaining luminal space within normal and oncogene-expressing mammary acini. Cell, 2002. 111(1): p. 29-40.
122. Tucker, A.S., Salivary gland development. Semin Cell Dev Biol, 2007. 18(2): p. 237-44.

110
123. Aframian, D.J., et al., Absence of tight junction formation in an allogeneic graft cell line used for developing an engineered artificial salivary gland. Tissue Eng, 2002. 8(5): p. 871-8.
124. Lam, K., et al., HSG cells differentiated by culture on extracellular matrix involves induction of S-adenosylmethione decarboxylase and ornithine decarboxylase. J Cell Physiol, 2005. 203(2): p. 353-61.
125. Streuli, C.H., et al., Laminin mediates tissue-specific gene expression in mammary epithelia. J Cell Biol, 1995. 129(3): p. 591-603.
126. Liu, J.K., C.M. DiPersio, and K.S. Zaret, Extracellular signals that regulate liver transcription factors during hepatic differentiation in vitro. Mol Cell Biol, 1991. 11(2): p. 773-84.
127. Trevigen, C. 128. Zheng, C., et al., Growth factor regulation of the amylase promoter in a differentiating
salivary acinar cell line. J Cell Physiol, 1998. 177(4): p. 628-35. 129. Manoussakis, M.N., et al., Expression of B7 costimulatory molecules by salivary gland
epithelial cells in patients with Sjogren's syndrome. Arthritis Rheum, 1999. 42(2): p. 229-39.
130. Ogawa, N., et al., Involvement of the interferon-gamma-induced T cell-attracting chemokines, interferon-gamma-inducible 10-kd protein (CXCL10) and monokine induced by interferon-gamma (CXCL9), in the salivary gland lesions of patients with Sjogren's syndrome. Arthritis Rheum, 2002. 46(10): p. 2730-41.
131. Miller, C.J., et al., Mycoplasma infection significantly alters microarray gene expression profiles. Biotechniques, 2003. 35(4): p. 812-4.
132. Lam, K.M., Mycoplasma gallisepticum--induced alterations in cytokine genes in chicken cells and embryos. Avian Dis, 2004. 48(1): p. 215-9.
133. Dimitriou, I.D., et al., Establishment of a convenient system for the long-term culture and study of non-neoplastic human salivary gland epithelial cells. Eur J Oral Sci, 2002. 110(1): p. 21-30.
134. Lafrenie, R.M., S.M. Bernier, and K.M. Yamada, Adhesion to fibronectin or collagen I gel induces rapid, extensive, biosynthetic alterations in epithelial cells. J Cell Physiol, 1998. 175(2): p. 163-73.
135. Azuma, M., et al., Expression of integrin subunits in normal and malignant human salivary gland cell clones and its regulation by transforming growth factor-beta 1. Cancer Lett, 1996. 109(1-2): p. 91-9.
136. Muschler, J., et al., Division of labor among the alpha6beta4 integrin, beta1 integrins, and an E3 laminin receptor to signal morphogenesis and beta-casein expression in mammary epithelial cells. Mol Biol Cell, 1999. 10(9): p. 2817-28.
137. Kashimata, M. and E.W. Gresik, Epidermal growth factor system is a physiological regulator of development of the mouse fetal submandibular gland and regulates expression of the alpha6-integrin subunit. Dev Dyn, 1997. 208(2): p. 149-61.
138. Hoffman, M.P., et al., Laminin-1 and laminin-2 G-domain synthetic peptides bind syndecan-1 and are involved in acinar formation of a human submandibular gland cell line. J Biol Chem, 1998. 273(44): p. 28633-41.
139. Cha, S., et al., Abnormal organogenesis in salivary gland development may initiate adult onset of autoimmune exocrinopathy. Exp Clin Immunogenet, 2001. 18(3): p. 143-60.
140. Caswell, P.T. and J.C. Norman, Integrin trafficking and the control of cell migration. Traffic, 2006. 7(1): p. 14-21.
141. Gawaz, M., et al., The NITY motif of the beta-chain cytoplasmic domain is involved in stimulated internalization of the beta3 integrin A isoform. J Cell Sci, 2001. 114(Pt 6): p. 1101-13.

111
142. Szlavik, V., et al., Matrigel-induced acinar differentiation is followed by apoptosis in HSG cells. J Cell Biochem, 2008. 103(1): p. 284-95.
143. Kulkarni, K., K. Selesniemi, and T.L. Brown, Interferon-gamma sensitizes the human salivary gland cell line, HSG, to tumor necrosis factor-alpha induced activation of dual apoptotic pathways. Apoptosis, 2006. 11(12): p. 2205-15.
144. Atsumi, T., S. Fujisawa, and K. Tonosaki, (Meth)acrylate monomer-induced cytotoxicity and intracellular Ca(2+) mobilization in human salivary gland carcinoma cells and human gingival fibroblast cells related to monomer hydrophobicity. Biomaterials, 2006. 27(34): p. 5794-800.
145. Atsumi, T., K. Tonosaki, and S. Fujisawa, Induction of early apoptosis and ROS-generation activity in human gingival fibroblasts (HGF) and human submandibular gland carcinoma (HSG) cells treated with curcumin. Arch Oral Biol, 2006. 51(10): p. 913-21.
146. Patel, Y.I. and N.J. McHugh, Apoptosis-new clues to the pathogenesis of Sjogren's syndrome? Rheumatology (Oxford), 2000. 39(2): p. 119-21.
147. Manganelli, P., et al., Quantitative analysis of apoptosis and bcl-2 in Sjogren's syndrome. J Rheumatol, 1997. 24(8): p. 1552-7.
148. Van Blokland, S.C., et al., Apoptosis and apoptosis-related molecules in the submandibular gland of the nonobese diabetic mouse model for Sjogren's syndrome: limited role for apoptosis in the development of sialoadenitis. Lab Invest, 2003. 83(1): p. 3-11.
149. Yamada, K.M., R. Pankov, and E. Cukierman, Dimensions and dynamics in integrin function. Braz J Med Biol Res, 2003. 36(8): p. 959-66.
150. Wozniak, M.A., et al., ROCK-generated contractility regulates breast epithelial cell differentiation in response to the physical properties of a three-dimensional collagen matrix. J Cell Biol, 2003. 163(3): p. 583-95.
151. Chakravarty, G., et al., P190-B, a Rho-GTPase-activating protein, is differentially expressed in terminal end buds and breast cancer. Cell Growth Differ, 2000. 11(7): p. 343-54.
152. Chakravarty, G., et al., p190-B RhoGAP regulates mammary ductal morphogenesis. Mol Endocrinol, 2003. 17(6): p. 1054-65.
153. Akhtar, N. and C.H. Streuli, Rac1 links integrin-mediated adhesion to the control of lactational differentiation in mammary epithelia. J Cell Biol, 2006. 173(5): p. 781-93.
154. O'Brien, L.E., et al., Morphological and biochemical analysis of Rac1 in three-dimensional epithelial cell cultures. Methods Enzymol, 2006. 406: p. 676-91.