Çukurova Ünİversİtesİ fen bİlİmlerİ enstİtÜsÜİmmobilizasyon yöntemleri çeşitli...
TRANSCRIPT

ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
YÜKSEK LİSANS TEZİ
Fadile YENER
PEKTİNAZ ENZİMİNİN FARKLI İKİ DESTEK ÜZERİNE İMMOBİLİZASYONU VE KARAKTERİZASYONU
KİMYA ANABİLİM DALI
ADANA, 2007

ÇUKUROVA ÜNİVERSİTESİ
FEN BİLİMLERİ ENSTİTÜSÜ
PEKTİNAZ ENZİMİNİN FARKLI İKİ DESTEK ÜZERİNE İMMOBİLİZASYONU VE KARAKTERİZASYONU
Fadile YENER
YÜKSEK LİSANS TEZİ
KİMYA ANABİLİM DALI
Bu tez ..../...../2007Tarihinde Aşağıdaki Jüri Üyeleri Tarafından Oybirliği/Oyçokluğu İle Kabul Edilmiştir.
İmza............……… İmza...................…. ….. İmza.................…………. Doç..Dr.Güzide YÜCEBİLGİÇ Prof.Dr.S.Seyhan TÜKEL Prof.Dr.Nuray ŞAHAN DANIŞMAN ÜYE ÜYE
Bu tez Enstitümüz Kimya Anabilim Dalında hazırlanmıştır.
Kod No: Prof. Dr Aziz ERTUNÇ Enstitü Müdürü
Bu Çalışma Çukurova Üniversitesi Bilimsel Araştırmalar ve Projeler (BAP) Birimi tarafından Desteklenmiştir.
Proje No: FEF.2006.YL.70
Not: Bu tezde kullanılan özgün ve başka kaynaktan yapılan bildirişlerin, çizelge, şekil ve fotoğrafların kaynak gösterilmeden kullanımı, 5846 sayılı Fikir ve Sanat Eserleri Kanunundaki hükümlere tabidir.

I
ÖZ
YÜKSEK LİSANS TEZİ
Fadile YENER
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
KİMYA ANABİLİM DALI
Danışman : Doç. Dr. Güzide YÜCEBİLGİÇ
Yıl : 2007 Sayfa: 47 Jüri : Doç. Dr. Güzide YÜCEBİLGİÇ Prof. Dr. S.Seyhan TÜKEL Prof. Dr. Nuray ŞAHAN Pektinaz enzimi alginat ve florisil üzerine immobilize edilmiştir. Belirlenen
optimum koşullarda serbest ve alginat üzerine immobilize edilen Pektinaz için
maksimum aktivite sırasıyla 69,4 ve 20,7 U/mg protein olarak ölçülmüştür. Km
değerleri sırasıyla 8,04 ve 0,36 mg/ml olarak belirlenmiştir. İmmobilize Pektinaz’ın
tekrar kullanılabilirliği kesikli reaktör modelinde araştırılmış ve 20 kullanımdan
sonra immobilize Pektinaz’ın başlangıç aktivitesinin % 94’ünü koruduğu
belirlenmiştir. Serbest ve immobilize Pektinaz’ın farklı sıcaklıklarda belirlenen
termal kararlılıkları karşılaştırılmıştır. Serbest Pektinaz’ın 5oC ve 25oC’de 5 hafta
depolanma süresi sonunda kalan aktivitesi sırasıyla başlangıç aktivitesinin % 57 ve
% 64’üdür. Bununla birlikte immobilize enzim 5oC ve 25oC’de 5 hafta depolanma
süresi sonunda sırasıyla başlangıç aktivitesinin % 75 ve % 76’sını korumuştur.
Florisil destek üzerine yapılan immobilizasyonda, denenen koşullarda enzimin
desteğe bağlandığı, bağlandıktan sonra aktivite göstermediği gözlenmiştir.
Anahtar Kelimeler: Pektik enzimler, exopoligalakturonaz, alginat, immobilizasyon, florisil.
PEKTİNAZ ENZİMİNİN FARKLI İKİ DESTEK ÜZERİNE İMMOBİLİZASYONU VE KARAKTERİZASYONU

II
ABSTRACT
MSc THESIS
Fadile YENER
DEPARTMENT OF CHEMISTRY INSTITUTE OF NATURAL AND APPLIED SCIENCES
UNIVERSITY OF ÇUKUROVA
Supervisor : Doç. Dr. Güzide YÜCEBİLGİÇ Year : 2007 Pages: 47 Jury :Doç. Dr. Güzide YÜCEBİLGİÇ Prof. Dr. S.Seyhan TÜKEL Prof. Dr. Nuray ŞAHAN
Pectinase enzyme was immobilized onto alginate and florisil. Maximum
activity of free pectinase and immobilized pectinase onto alginate were measured as
69,4 and 20,7 U/mg protein respectively, at predetermined optimum conditions. The
KM values for free and immobilized pectinase were determined as 8,04 and 0,36
mg/ml,respectively. Reusability of the immobilized pectinase was investigated in a
batch type reactor. After 20 reuses, the residual activity of immobilized pectinase
was about % 94 of its initial activity. Thermal stabilities of free and immobilized
pectinase determined at different temperatures were compared. The residual activity
for immobilized pectinase stored at 5oC and 25oC for five weeks were 57 % and 64
% of its initial activity, respectively. However, the immobilized enzyme protected 75
% and 76 % of its original activity at 5oC and 25oC, respectively for five weeks. In
the immobilization onto the florisil support, it was observed that the enzyme was
bound to the support however, no activity was observed.
Keywords: Pectic enzymes, exopolygalacturonase, alginate, immobilization,
florisil.
IMMOBILIZATION OF PECTINASE ONTO TWO DIFFERENT SUPPORTS AND THEIR CHARACTERIZATION

III
TEŞEKKÜR
Yüksek Lisans Tezinin yönetiminde ve oluşumunda aynı zamanda tez
çalışmam süresince her türlü desteğini gördüğüm, sabır ve anlayışından dolayı
değerli Danışmanım Doç. Dr. Güzide YÜCEBİLGİÇ’e,
Çalışmalarımda benden yardım ve desteklerini esirgemeyen hocalarım Prof.
Dr. S.Seyhan TÜKEL’e ve Yrd. Doç. Dr. Ramazan BİLGİN’e,
Çalışmalarım süresince her türlü yardım ve manevi desteklerini gördüğüm
Özlem ALPTEKİN, Dilek ALAGÖZ, Deniz YILDIRIM, Özlem YILDIRIM’a ve
aynı laboratuarı paylaştığım arkadaşlarıma,
Şu an çalışmakta olduğum Adli Tıp Anabilim Dalında bana her türlü yardım
ve desteği gösteren Nebile DAĞLIOĞLU ve Dilek AKÜNAL’a,
Karşılaştığım her zorlukta hep yanımda olan, varlığıyla bu sürecin
kolaylaşmasını sağlayan, çalışmalarımda yardımlarını esirgemeyen, hayatımı
paylaşmaya karar verdiğim Ümit YALDIZ’a,
Benden maddi, manevi desteklerini esirgemeyen, beni büyük bir ilgi ve
sevgiyle bugünlere getiren babam Abdullah YENER, annem Nursel YENER’e ve
hayatımı anlamlı hale getiren kardeşlerime teşekkürlerimi sunmayı bir borç bilirim.

IV
İÇİNDEKİLER SAYFA
ÖZ…………………………………………………………………………………..I
ABSTRACT………………………………………………………………………..II
TEŞEKKÜR……………………………………………………………………….III
İÇİNDEKİLER……………………………………………………………………IV
ÇİZELGELER DİZİNİ …………..………………………………………………VI
ŞEKİLLER DİZİNİ…..…………………………………………………………..VII
SİMGELER VE KISALTMALAR ..…………………………………………...VIII
1.GİRİŞ……………………………………………………………………………...1 1.1. Enzim İmmobilizasyon Yöntemleri…………………………………………...3
1.1.1.Çözünmez Formda İmmobilizasyon Yöntemleri………………………...3
1.1.1.1. Bağlama………………………………………………………...4
1.1.1.2. Tutuklama………………………………………………………5
1.1.2. Çözünür Formda İmmobilizasyon………………………………………7
1.1.3. İmmobilize Edilen Enzimden Beklenilen Özellikler................................7
1.2. Pektin (Pektik bileşikler)....................................................................................8
1.2.1. Pektinin Endüstriyel Önemi...................................................................13
1.3. PektolitikEnzimler….....………………………………………...................... 13
1.3.1. Pektolitik Enzimlerin Genel Özellikleri.................................................13
1.3.2. Endüstride Pektolitik Enzimlerin Kullanılması .....................................16
1.4. Alginat…………………………………………………………...................... 17
1.5. Florisil...............................................................................................................18
2.ÖNCEKİ ÇALIŞMALAR……………………………………………………… 19
3.MATERYAL VE METOD……………………………………………………....22
3.1. Materyal……………………………………………………………………..22
3.2. Metod…..........................................................................................................22
3.2.1. Protein Tayini.........................................................................................22
3.2.2. Pektinaz’ın Alginat Destek Üzerine İmmobilizasyonu..........................23
3.2.3. Pektinaz’ın Florisil Destek Üzerine İmmobilizasyonu..........................23

V
3.2.4. Serbest veya İmmobilize Pektinaz’ın Aktivitesinin Belirlenmesi.........24
3.2.5. Galakturonik Asit Tayini.......................................................................25
3.2.6. Serbest ve İmmobilize Enzimlerin Karakterizasyonu............................26
3.2.6.1. Serbest ve immobilize Enzimlerin Optimum pH’larının
Belirlenmesi..............................................................................26
3.2.6.2. Serbest ve İmmobilize Enzimlerin Optimum Tampon
Derişimlerinin Belirlenmesi.....................................................26
3.2.6.3. Serbest ve İmmobilize Enzimlerin Optimum Sıcaklıklarının
Belirlenmesi..............................................................................26
3.2.6.4. Serbest ve İmmobilize Enzimlerin KM ve Vmax Değerlerinin
Belirlenmesi..............................................................................27
3.2.6.5. Serbest ve İmmobilize Enzimlerin Termal Kararlılıklarının
Belirlenmesi..............................................................................27
3.2.6.6. Serbest ve İmmobilize Enzimlerin Depolama
Kararlılıklarının Belirlenmesi...................................................27
3.2.6.7. İmmobilize Pektinaz’ın Tekrar Kullanım Kararlılığının
Belirlenmesi..............................................................................27
4.BULGULAR VE TARTIŞMA……………………………………………..........28
4.1.Bulgular.................................................................................................................28
4.1.1. Serbest ve İmmobilize Pektinaz’ın Karakterizasyonu.............................28
4.1.2. Pektinaz’ın Florisile İmmobilizasyonu....................................................36
4.2. Tartışma...............................................................................................................37
4.2.1.Pektinaz’ın Alginat Destek Üzerine İmmobilizasyonu ve
Karakterizasyonu......................................................................................37
4.2.2. Pektinaz’ın Florisil Destek Üzerine İmmobilizasyonu ..........................40
5.SONUÇLAR VE ÖNERİLER..............................................................................41
5.1.Sonuçlar............................................................................................................41
5.2. Öneriler............................................................................................................42
KAYNAKLAR..........................................................................................................44
ÖZGEÇMİŞ................................................................................................................47

VI
ÇİZELGELER DİZİNİ SAYFA
Çizelge 1.1. Meyve pektin içeriği.............................................................................8
Çizelge 1.2. Pektik bileşikler ve özellikleri.............................................................10
Çizelge 1.3. Pektik enzimlerin sınıflandırılması.....................................................14
Çizelge 4.1. Serbest ve immobilize Pektinaz’ın kinetik parametreleri...................32
Çizelge 4.2. Pektinaz’ın florisile immobilizasyonu................................................36

VII
ŞEKİLLER DİZİNİ SAYFA Şekil 1.1. Enzim immobilizasyon yöntemleri……………………………....... 3
Şekil 1.2. Enzimin polimerler arasında tutuklanması………...…………....... 6
Şekil 1.3. Düşük ve yüksek esterleşme dereceli pektin…………………....... 9
Şekil 1.4. Pektin molekülünde düz ve saçaklı bölgeleri gösteren model......... 11
Şekil 1.5. Pektin molekülü............................................................................... 12
Şekil 1.6. Poligalakturonaz enziminin etki mekanizması................................ 15
Şekil 1.7. Pektat-liyaz enziminin etki mekanizması........................................ 15
Şekil 1.8. Pektin-liyaz enziminin etki mekanizması........................................ 16
Şekil 1.9. Pektinesteraz enzimin etki mekanizması......................................... 16
Şekil 4.1. Serbest ve immobilize Pektinaz aktivitesinin pH’ya bağlı olarak
değişimi…………………………………………………………... 28
Şekil 4.2. Serbest ve immobilize Pektinaz aktivitesinin tampon derişimine
bağlı olarak değişimi……………………………………..……… 29
Şekil 4.3. Serbest ve immobilize Pektinaz aktivitesinin sıcaklığa bağlı
olarak değişimi……………………………………….………….. 30
Şekil 4.4. Serbest Pektinaz’ın Arrhenius grafiği…………………………… 30
Şekil 4.5. İmmobilize Pektinaz’ın Arrhenius grafiği……………………… 31
Şekil 4.6. Serbest Pektinaz için Lineweaver-burk grafiği…………………... 31
Şekil 4.7. İmmobilize Pektinaz için Lineweaver-burk grafiği……………… 32
Şekil 4.8. Serbest Pektinaz enziminin termal kararlılık grafiği…………….. 33
Şekil 4.9. İmmobilize Pektinaz enziminin termal kararlılık grafiği……….... 34
Şekil 4.10. Serbest Pektinaz enziminin depolama kararlılığı………………… 34
Şekil 4.11. İmmobilize Pektinaz enziminin depolama kararlılığı……………. 35
Şekil 4.12. İmmobilize Pektinaz’ın tekrar kullanım kararlılığı………………... 36

VIII
SİMGELER ve KISALTMALAR KM : Michaelis-Menten hız sabiti (mg/ml) Vmax : Maksimum hız (µmol.dk-1.mg protein-1) Ea : Aktivasyon enerjisi (J mol-1) Vmax / KM : Katalitik etkinlik U : Ünite V : Tepkime hız sabiti (µmol.dk-1.mg protein-1) [S] : Subsrat derişimi DNSA : Dinitrosalisilik asit

1.GİRİŞ Fadile YENER
1
1.GİRİŞ
Enzimler, canlı hücrelerde oluşan ve organizmadaki tüm reaksiyonların çok
yumuşak koşullarda gerçekleşmesini sağlayan ve bunları regüle eden biyolojik
katalizatörlerdir. Enzimlerle katalizlenen reaksiyonlar katalizlenmemiş karşıtlarına
göre 107-1016 kez daha hızlı gerçekleşirler. Enzimler son derece özgün olup
birbirinin aşağı yukarı aynısı olan enantiomerleri bile ayırt edebilirler.
Enzimlerin kimyasal katalizörlere göre üstünlüklerini incelediğimizde;
enzimler ileri derecede substrat spesifikliğine sahip olmaları nedeniyle, istenmeyen
yan ürünlerin oluşumunu engeller. Enzimlerin katalizlediği reaksiyonlarda verim %
100’dür. Stereospesifik reaksiyonları enzimler katalizleyebilirler. Enzimler ılımlı
koşullarda ( pH 7.0, oda sıcaklığı) çalıştığı için reaksiyon maliyetini düşürürler ve
çevresel sorunlar yaratmazlar. Yeterli koşulların sağlanması koşuluyla etkilerini
doğal ortamlarının dışında da gösterebiliyor olmaları, enzimlerden pek çok alanda
yararlanabilme imkanı vermektedir. Enzimlerden başlıca endüstriyel, tıbbi, bilimsel
ve analitik amaçlı olmak üzere üç farklı alanda yararlanılmaktadır. Her bir alanda
kullanılan enzimlerin fiyatları beklenen özelliklerine göre birkaç dolardan birkaç
bin dolara kadar değişebilmektedir.
Enzimler canlılar tarafından üretildiklerinden endüstriyel veya analitik amaçlı
kullanımları için doku, kan, mikroorganizma gibi canlı veya canlı kökenli
kaynaklardan saflaştırılırlar. Enzim üretiminde hammadde olarak canlıların kullanımı
ekonomik açıdan sınırlayıcı bir durum olsa da bu sorun mikrobiyal kaynaklar
sayesinde büyük ölçüde çözülmüş görünmektedir. Bununla birlikte enzimlerin
mikrobiyal kaynaklardan saflaştırılması oldukça masraflı bir iştir. Endüstriyel
uygulamalarda serbest enzimin aktivitesini kaybetmeden geri kazanılması çok
zordur. Serbest enzim, reaksiyon ortamından istenilen anda uzaklaştırılamadığından
reaksiyonun kontrolü çok güçtür. Reaksiyonun istenilen anda durdurulması için
inhibitör katılması düşünülebilir. Ancak serbest enzim tarafından kirletilmiş olan
reaksiyon ürünlerine böylece yeni bir kirlilik unsuru eklenmiş olacaktır. Ürün veya
ürünlerin bu kirlilik unsurlarından arıtılması maliyeti çok arttırmaktadır. Katalizör
olarak kullanılan serbest enzimi reaksiyon ortamından aktivitesini yitirmeden

1.GİRİŞ Fadile YENER
2
çıkarabilmek olanaksız olduğundan enzimin yeniden kullanılması da söz konusu
değildir. Bu ise enzimlerin spesifik ama o ölçüde pahalı katalizör olmaları nedeniyle
maliyeti yükselten önemli bir etmendir. Ayrıca serbest enzimler sürekli üretim
sistemlerine de uygulanamazlar. Bu nedenle saflaştırılan enzimlerden olabildiğince
faydalanmak için immobilizasyon teknikleri geliştirilmiştir.
Enzimler, suda çözünmeyen bir taşıtıcıya fiziksel veya kimyasal olarak
bağlanarak, suda çözünmeyen ürün veren bir kopolimerizasyona enzim
molekülünün monomer olarak katılmasıyla ve suda çözünmeyen mikrokapsüllerde
tutuklamakla immobilize edilirler (Telefoncu, 1997).
İmmobilize enzimin serbest enzime göre üstünlükleri;
Reaksiyon sonunda ortamdan kolayca uzaklaştırılabilir (süzme, santrifüjleme
v.b.) ve ürünlerin enzim tarafından kirletilmesi gibi bir problem yaratmaz.
Çevre koşullarına ( pH, sıcaklık v.s.) karşı daha dayanıklıdır.
Birçok kez ve uzun süre kullanılabilir.
Sürekli işlemlere uygulanabilir.
Doğal enzime kıyasla daha kararlıdır.
Ürün oluşumu kontrol altında tutulabilir.
Birbirini izleyen çok adımlı reaksiyonlar için uygundur.
Bazı durumlarda serbest enzimden daha yüksek bir aktivite gösterebilir.
Enzimin kendi kendini parçalama olasılığı azalır.

1.GİRİŞ Fadile YENER
3
1.1.Enzim İmmobilizasyon Yöntemleri
İmmobilizasyon yöntemleri çeşitli şekillerde sınıflandırılmakla beraber Şekil
1.1’de verildiği gibi sınıflandırmak mümkündür (Telefoncu, 1997).
Şekil 1.1. Enzim İmmobilizasyon Yöntemleri (Telefoncu, 1997).
1.1.1. Çözünmez Formda İmmobilizasyon Yöntemleri
Çözünmez formda immobilizasyon yöntemleri bağlama ve tutuklama olarak
ikiye ayrılmaktadır.

1.GİRİŞ Fadile YENER
4
1.1.1.1.Bağlama
Bağlama yöntemi; çapraz bağlama, enzim kopolimerizasyonu ve taşıyıcıya
bağlama olmak üzere üç gruba ayrılmaktadır.
Çapraz bağlama; Küçük moleküllü iki veya çoklu fonksiyonel grupları olan
reaktifler enzim molekülleri arasında çapraz bağlar yapmaktadırlar. En çok
kullanılan çapraz bağlama reaktifleri; glutaraldehit, klorformat ve karbonilimidazol,
heterosiklik halojenürler, bioksiranlar, divinilsülfonlar, p-benzokinon, geçiş metal
iyonları ve epiklorohidrinlerdir.
Enzim kopolimerizasyonu; Enzimler bir kopolimerizasyon reaksiyonunda
monomerlerden biri gibi davranarak matrikse bağlanmaktadır. Yöntem polimer
matrikse tutuklanmaya benzemekle birlikte enzim kaçışının önlenmesi gibi üstünlüğü
vardır.
Taşıyıcıya bağlama; Adsorpsiyon, iyonik bağlama, şelat bağlama, kovalent
bağlama ve biyospesifik bağlama olarak beş gruba ayrılmaktadır. Bir protein olan
enzim molekülünün yapısından yararlanılır. Molekül yüzeyindeki fonksiyonel
gruplar, iyonik gruplar ve hidrofobik bölgeler bu bağlamada rol alırlar. Enzim
immobilizasyonunda doğal veya sentetik birçok organik ve inorganik materyal
kullanılmaktadır. Taşıyıcı membran, suda çözünmeyen katı veya polimer olabilir.
Enzim immobilizasyonunda kullanılacak taşıyıcıda aranan nitelikler şunlardır:
Hidrofilik karakter,
Suda çözünmeme,
Gözenekli yapı,
Mekanik stabilite uygun partikül formu,
Kimyasal ve termal stabilite,
Kovalent bağlamada kullanılacak taşıyıcıların yumuşak koşullarda
reaksiyon verebilen fonksiyonel grupları taşıması,

1.GİRİŞ Fadile YENER
5
Mikroorganizmalara karşı dirençlilik,
Ucuzluk,
Zehirsizlik,
Rejenere olabilme.
Enzim immobilizasyonunda kullanılacak taşıyıcılar reaktif değillerse
yardımcı bir reaktif ile aktifleştirilmesi gerekir. İmmobilizasyon çok yumuşak
koşullarda (oda sıcaklığı, nötral pH vb.) gerçekleştirilmelidir.
Taşıyıcı suda çözünmemeli ancak büyük ölçüde hidrofobik karakterli de
olmamalı, suda ıslanabilmeli, ayrıca mekanik kararlı olmalıdır. Bu tür taşıyıcıların
seçiminde enzim-taşıyıcı bağının aktivite için zorunlu gruplar üzerinden olmaması
yanında taşıyıcının enzim tarafından parçalanmaması, mikroorganizma üremesine
olanak vermemesi, pH ve çözücülere karşı dayanıklı olması gibi özellikler taşımasına
dikkat edilmelidir..
1.1.1.2. Tutuklama Yöntemleri
Tutuklama yönteminde, enzim molekülü belirli bir mekanda durmaya
zorlanmaktadır. Enzim bulunduğu çevreden dışarıya çıkamaz. Bu işlem polimer
matriks içindeki kafeslerde gerçekleştirilebileceği gibi yarı geçirgen membranlar
içinde mikrokapsülleme ve miseller ile de gerçekleştirilebilir. Bu yöntemi, kovalent
bağlama ve çapraz bağlama ile immobilizasyondan ayıran en önemli özellik enzim
molekülünün fiziksel veya kimyasal olarak herhangi bir taşıyıcıya bağlanmamış
olmasıdır. Tutuklama yöntemi; polimer matrikste tutuklama, mikrokapsülleme,
lipozom tekniği olmak üzere üçe ayrılır.
Polimer matrikste tutuklama yöntemi, yüksek derecede çapraz bağlı bir
polimerin enzim çözeltisi içinde oluşturulması temeline dayanır (Şekil 1.2).
Polimerleşme sonucu enzim molekülleri çapraz bağ ağları arasında tutuklanmakta ve
böylece ana çözeltiye geçmeleri engellenmektedir. Bu amaçla en çok kullanılan
polimer N,N’-metilenbisakrilamid ile çapraz bağlanmış poliakrilamiddir.

1.GİRİŞ Fadile YENER
6
Şekil 1.2. Enzimin polimerler arasında tutuklanması
Mikrokapsülleme yöntemi, enzim moleküllerinin yarı geçirgen bir membran
içinde tutuklanması esasına dayanır. Mikrokapsüllerin büyüklüğü 1-100 µ arasında
değişmektedir. Yarıgeçirgen membranın gözenek çapları; substrat moleküllerinin
kapsül içine girişine ve ürün moleküllerinin dışarı çıkışına olanak verecek bir
büyüklükte olmalıdır. Substrat molekülleri ne kadar küçükse bu yöntem ile
immobilize edilmiş enzimin verimliliği o ölçüde yüksek olacaktır.
Lipozom tekniği, sıvı-yüzey yapıcı membran temeline dayanır. Yöntemin en
önemli üstünlüğü; süreksiz, dönüşümlü ve tamamen fiziksel oluşudur. Aynı anda bir
adımda birçok enzimin immobilizasyonuna olanak sağlar ve oldukça büyük bir
kontakt yüzeyine sahiptir. Önemli sakıncaları ise; substrat ve ürünün membranda
geçişinin çözünürlüğe bağımlı olması, işlem sırasında enzimin inaktive olması ve sıvı
olan membrandan enzimin kaçma olasılığıdır.

1.GİRİŞ Fadile YENER
7
1.1.2. Çözünür Formda İmmobilizasyon
Çözünür formda immobilizasyon yöntemi enzimin herhangi bir taşıyıcıyla
fiziksel veya kimyasal etkileşiminden çok yarı geçirgen bir zarla çevrelendiği ve
enzime geniş bir hareket alanının sağlandığı bir yöntemdir. Enzim taşıyıcıya
herhangi bir bölgesinden bağlanmadığı için moleküler geometrisi, esnekliği ve
dolayısıyla katalitik etkinliği değişmemektedir. Fakat substratın membrandan geçip
enzime ulaşmasında kısıtlamalara rastlanmaktadır. Bu nedenle küçük moleküllü
substrata sahip olan enzimlerin bu yöntemle immobilize edilmesi tercih edilebilir.
1.1.3. İmmobilize Edilen Enzimden Beklenilen Özellikler
Kullanılan immobilizasyon tekniği ne olursa olsun immobilize edilen
enzimden beklenilen özellikler şunlardır:
Yüksek kararlılık
Tekrar kullanılabilirlik
Sürekli üretime olanak vermesi
Reaksiyon kontrolüne olanak vermesi
Yüksek saflık
Yüksek ürün yüzdesi
Ekonomik olması
1.GİRİŞ Fadile YENER
8
1.2.Pektin (Pektik Bileşikler)
Meyve ve sebzelerin teknolojik açıdan en önemli polisakkaritleri pektindir.Bu
ürünlerde pektin miktarı % 0.5-% 1.0 düzeyindedir. Metilasyon derecesi, moleküler
ağırlık ve pektin içeriği meyve çeşidine göre farklılık gösterir (Çizelge 1.1).
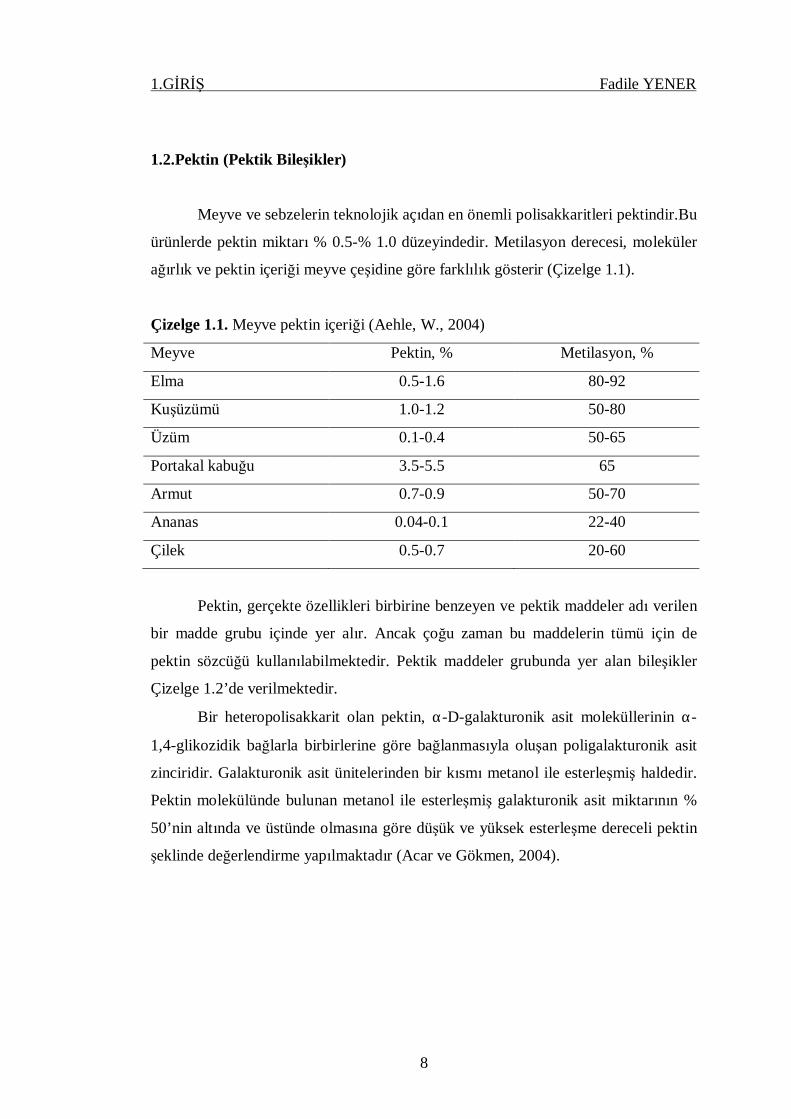
Çizelge 1.1. Meyve pektin içeriği (Aehle, W., 2004)
Meyve Pektin, % Metilasyon, %
Elma 0.5-1.6 80-92
Kuşüzümü 1.0-1.2 50-80
Üzüm 0.1-0.4 50-65
Portakal kabuğu 3.5-5.5 65
Armut 0.7-0.9 50-70
Ananas 0.04-0.1 22-40
Çilek 0.5-0.7 20-60
Pektin, gerçekte özellikleri birbirine benzeyen ve pektik maddeler adı verilen
bir madde grubu içinde yer alır. Ancak çoğu zaman bu maddelerin tümü için de
pektin sözcüğü kullanılabilmektedir. Pektik maddeler grubunda yer alan bileşikler
Çizelge 1.2’de verilmektedir.
Bir heteropolisakkarit olan pektin, α-D-galakturonik asit moleküllerinin α-
1,4-glikozidik bağlarla birbirlerine göre bağlanmasıyla oluşan poligalakturonik asit
zinciridir. Galakturonik asit ünitelerinden bir kısmı metanol ile esterleşmiş haldedir.
Pektin molekülünde bulunan metanol ile esterleşmiş galakturonik asit miktarının %
50’nin altında ve üstünde olmasına göre düşük ve yüksek esterleşme dereceli pektin
şeklinde değerlendirme yapılmaktadır (Acar ve Gökmen, 2004).

1.GİRİŞ Fadile YENER
9
Yüksek esterleşme dereceli pektin (%75)
OCOOCH3
H
H
H
H
OHO
OH
H
H OH
HOH
COOCH3
O
OH
HHO
H
H
H
OHO
OH
H OH
HOH
COOCH3OH
HH
COOH
H O
Düşük esterleşme dereceli pektin (%25)
O
O
H
H
OH
H OH
COOH
OH
H
O
OCOOCH3
H
H
H
OH
OH
O
HO
H
H
H
OH
OH
H
COOH
O
H OH
HOHH O
COOH
OH
Şekil 1.3. Düşük ve yüksek esterleşme dereceli pektin (Acar ve Gökmen, 2004)
Pektinin esterleşme oranı kaynağına göre değişim göstermektedir. Örneğin
elma pektininde esterleşmiş karboksil gruplarının oranı % 95.3 iken vişne pektininde
% 55.4’dür.

1.GİRİŞ Fadile YENER
10
Çizelge 1.2. Pektik bileşikler ve özellikleri (Ekşi, 1988)
Bileşik adı Özelliği
Pektik asit
(poligalakturonik asit)
Esteleşmemiş galakturonik asit birimlerinden oluşan
zincir. Asit formda suda çözünmez, kısmen
nötralizasyondan sonra suda çözünür ve Ca ile jel
oluşturur.
Pektat Pektik asitin nötral veya asidik tuzudur.Çok az sayıda
metoksil grubu içerir. (Na-pektat suda çözünmekte,ancak
Ca-pektat suda çözünmemektedir.)
Pektin Kısmen ve tümüyle metil alkolde esterleşmiş (metilize
olmuş) poligalakturonik asittir. Metoksil grubu sayısı
(esterleşme derecesi) ve polimerizasyon derecesine bağlı
olarak özellikleri değişmektedir.
Pektinat Düşük ve yüksek esterleşme oranı içeren pektinin
tuzudur.Molekülde metoksil gruplarıda bulunur.
Protopektin Pektin zincirleri, esterleşmemiş –COOH grupları
üzerinden birbirine metal iyonu (Mg, Ca) ile
bağlanmıştır.Kısıtlı sayıda fosforik asit üzerinden ester
köprüsü de içeren doğal pektin olup, suda çözünmez.
Pektin türevleri Ana valenslerine asetil vb. gibi özel grupların bağlandığı
pektindir.
Pektin molekülünde doğrusal poligalakturonik asit ana zincirinde α-1,2-
glikozidik bağlarla bağlanmış ramnoz molekülleri yer almaktadır. Ramnoz
moleküllerinin pektin molekülü içinde dağılımı muntazam değildir. Bu nedenle bu
konuda yapılan çalışmalarda, pektin molekülünde düz (smooth) ve saçaklı (hairy)
bölgelerin varlığından söz edilmektedir (Şekil 1.4).Bazı araştırmacılara göre pektin
bitkide bulunduğu yere bağlı olarak orta lamel pektini veya primer hücre duvarı
pektini olarak da sınıflandırılmaktadır (Acar ve Gökmen, 2004; Bonnin ve ark.,
2002).

1.GİRİŞ Fadile YENER
11
Şekil 1.4. Pektin molekülünde düz (smooth) ve saçaklı (hairy) bölgeleri gösteren model (Acar ve Gökmen, 2004).
Pektin molekülünde yer alan ramnoz moleküllerine galaktan gibi nötral
şekerler ya uzun yan zincirler halinde ya da monomerler halinde bağlanmışlardır
(Şekil 1.5).

1.GİRİŞ Fadile YENER
12
OO
OO O
O
O
O
O
O
O
O
O
O
O
O
O
O
O
O O
O
O
O
O
O
O
O
O
O
O
GUA GUA
GUA
GUA GUA GUA GUA GUA
GUA GUA
RAM
RAMRAM
RAM
RAM
Galaktan
Galaktan
Şekil 1.5. Pektin molekülü (Acar, 2004)
Pektik maddeler, bitki duvarlarında oluşan yapısal polisakkaritlerdir.Bu
maddeler büyük oranda anhidrogalakturonik asit birimlerinden oluşan karmaşık,
kolloidal karbonhidrat türevlerinden meydana gelen yüksek su tutma kapasiteli
polisakkaritlerdir. Bu durumda pektik maddeleri oluşturan birim, poligalakturonik
asit olup, düz bir zincir yapmak üzere birbirleriyle α -1,4 bağı yapmışlardır. 30000
– 300000 Da moleküler ağırlıklı heteropolisakkaritlerdir. Pektik maddeler pektinik
asit, pektin, pektik asit ve bunların tuzlarını içeren bir grup maddeye verilen genel
addır (Willatns ve ark., 2001).
Pektin selüloz ve hemiselüloz gibi yüksek bitkilerin hücre duvarında bulunan
bir polisakkarittir. Pektinin yapısında poligalakturonik asit, rhamnogalakturonik asit,
galaktanlar ve arabinogalaktanlar vardır (Celestino ve ark., 2006). Pektinler pektik
maddeler, protopektinler ve pektik asitler olarak sınıflandırılırlar. Pektik maddeler,
tamamen veya bir kısmı esterleşmiş poligalakturonik asitleri içerirler. Tamamen
esterleşmiş poligalakturonik asitler teorik olarak % 16.3 metoksil grubu içerirler.
Protopektinler selüloz, xylan ve glukomannan gibi düğer hücre duvarı maddelerine
kimyasal ve fiziksel olarak bağlı suda çözünmez pektinlerdir. Pektik asitler, çok az
esterleşmiş veya hiç esterleşmemiş poligalakturonik asitlerdir. Bunlar şekerlerle,

1.GİRİŞ Fadile YENER
13
asitlerle veya divalent iyonlarla katı jel formuna getirilebilirler. Meyve ve sebzelerin
yapıları pektik maddelerin özellikleri ve içerikleriyle düzenlenir.
1.2.1. Pektinin Endüstriyel Önemi
Pektin gıda endüstrisinde reçel ve marmelat üretiminde jelleşmeyi sağlamak
amacıyla kullanılmaktadır. Pektin ( E 440) bir gıda katkı maddesidir, Meyve ve
sebze teknolojisinde berrak meyve suyu üretiminde önemli bir yere sahiptir. Meyve
suyu üretiminde mayşe enzimasyonu uygulansa bile pres suyu, kolloidal olarak
çözünen pektin içerir. Pektin koruyucu özelliğinden dolayı, diğer bileşikleri de
meyve ham suyu içinde askıda tutmaktadır. Bu nedenle berrak meyve suyu üretimi
için öncelikle pektik bileşiklerin enzimatik yolla degredasyonu gereklidir (Ekşi,
1988).
Pektin endüstriyel olarak elma ve turunçgil meyveleri posasından elde edilir.
Ham maddedeki protopektin asit ekstraksiyonu ile çözünür hale getirilir ve daha
sonra izopropil alkol ile çözünür pektin sulu çözeltiden ayrılır. Pektin ticarette sıvı
veya kurutulmuş toz halde bulunur (Acar ve Gökmen, 2004).
1.3. Pektolitik Enzimler
1.3.1.Pektolitik Enzimlerin Genel Özellikleri
Pektolitik enzimler galakturonik asidin doğal polimeri olan pektinleri
parçalarlar. Bu enzimlerin üretimi daha çok yüzey kültür tekniği ile katı besiyerlerde
gerçekleştirilir. Üretimde A. niger, A. oryzae, A. wentii, A. flavus, Rhizopus
liquefaciens ve Penicillum türleri kullanılır. Pektolitik enzimlerin biyosentezi
indüksiyon veya katabolik represyon mekanizması ile kontrol edilir (Telefoncu,
1997).
Pektik enzimler, genelde polisakkaritleri parçalayan zincir kırıcı enzimler
olarak bilinmektedirler. Bitki hücrelerinin orta lamelinin ve primer hücre duvarının
yapısını oluşturan pektik asit ve pektini parçalayan bu enzimler bakteri, mantar,

1.GİRİŞ Fadile YENER
14
böcek, nematod ve protozoada bulunmaktadırlar. Bitkisel kaynaklı pektik enzimler
oldukça yüksek molekül ağırlıklı pektik asitleri tercih ederler ve pektatların
indirgeyici olmayan uçlarına atak yaparak monogalakturonat oluştururlar. Oligo ve
digalakturonatları da parçalarlar. Substratlarını tek zincir etki mekanizmasına göre
ürüne çevirirler. Kalsiyum (Ca++) iyonu varlığında sitimüle olurlar. Pektatların
tamamen hidrolize olmamasının nedeni, substratların yapılarındaki düzensizlik
nedeniyle olduğu şeklinde yorumlanmaktadır.
Pektik maddelere etki eden ve kısaca pektik enzimler denilen grupta da çok
sayıda enzim yer almaktadır (Sunnotel ve Nigam, 2002). Pektik enzimleri, pektin
molekülünün galakturonan omurgasına olan etkilerine göre pektinesterazlar (ester
bağını hidrolize edici enzimler) ve depolimerazlar (zincir kırıcı enzimler) olarak iki
sınıfa ayırarak incelemek olasıdır (Aksöz ve ark., 1985).
Depolimerazlar, pektik maddeler üzerinde iki farklı mekanizmayla çalışırlar.
Birincisi, glikozidik bağları hidroliz eden hidrolitik reaksiyon. İkincisi, glikozidik
bağları su katılması olmaksızın kıran trans-eliminasyon reaksiyonu.
Depolimerazların bu sınıfı liyazlar olarak adlandırılır. Bu farklı kırılma reaksiyonları
pektinazların sınıflandırılmasında kullanılır (Soares ve ark., 1999).
Çizelge 1.3. Pektik enzimlerin sınıflandırılması
EC numarası Kırma noktası Hidrolazlar Endopoligalakturonaz 3.2.1.15 Zinciri arasından rastgele Exopoligalakturonaz 3.2.1.67 Zincirin sonundan Liyazlar Pektat-liyaz 4.2.2.2 Zinciri arasından rastgele Pektin-liyaz 4.2.2.10 Zinciri arasından rastgele Esteraz Pektin-esteraz 3.1.1.11 Deesterifikasyon

1.GİRİŞ Fadile YENER
15
Endopoligalakturonazlar (Endo-PG; 3.2.1.15), mantar, bakteri, maya gibi
birçok mikroorganizmada ve bitkilerde bulunur. Poligalakturonaz için en iyi subsrat
pektik asitlerdir. Pektik asitler endopoligalakturonaz tarafından hidrolizle
depolimerize olurlar. Pektik asitlerdeki serbest karboksil ucuna yakın glikozidik bağı
kırarlar. Pektat çözeltisinin viskozitesindeki hızlı azalma PG aktivitesinin bir
göstergesidir. Birkaç bağın kırılması bile viskozitede % 50 azalmaya neden olur.
Subsratın esterifikasyon derecesinin düşmesiyle PG aktivitesi düşer.
Exopoligalakturonazlar (Exo-PG, EC 3.2.1.67), yüksek bitkilerde,
mantarlarda, bakterilerde ve bazı böceklerin bağırsak sistemlerinde bulunurlar.
Yüksek moleküllü pektik asit zincirini sonundan kırarlar. Hidroliz ile
depolimerizasyon sonunda monomerler açığa çıkarırlar.
Şekil 1.6. Poligalakturonaz enziminin etki mekanizması
Pektat-liyazlar (PATE; EC 4.2.2.2 ), bazı bakteri ve patojenik mantarlar
tarafından üretilirler. Serbest karboksil ucuna yakın glikozidik bağını eliminasyon
tepkimesiyle kırarlar. Pektat-liyazlar için en iyi subsratlar düşük metoksilli pektinler
veya tamamen deesterifiye olmuş pektik asitlerdir. Enzimin aktif olması için Ca+2
iyonuna ihtiyacı vardır.
Şekil 1.7. Pektat-liyaz enziminin etki mekanizması
O
OH
OH
OH
COOH
O
O
OH
OH
COOH
O
O
OH
OHOH
COOH
O
OH
OH
COOH
O
Pektat-liyaz
O
OH
OH
OH
COOH
O
O
OH
OH
OH
COOH
O
O
OH
OH
OH
COOH
O
OH
OH
COOH
O
OH
+ H2O
Poligalakturonaz

1.GİRİŞ Fadile YENER
16
Pektin –liyazlar (PTE; EC 4.2.2.10 ), yüksek bitkilerde bulunmazlar ama bazı
mantarlarda bulunurlar. Depolimerazların bu türleri, ticari enzimlerin önemli
elemanlarıdır. Özellikle pratik uygulamalarda önemlidir. Yüksek dereceli esterleşmiş
poligalakturonik asitleri depolimerize ederler. Pektin liyazlar glikozidik bağını
kırdıklarında metilenmiş galakturonik asit artıkları açığa çıkar. En iyi subsratları
fazla esterleşmiş pektinlerdir ve Ca +2 iyonu tarafından aktif hale getirilirler.
Şekil 1.8. Pektin-liyaz enzimininin etki mekanizması
Pektin-esterazlar (PE; EC 3.1.1.11), yüksek bitkilerde, çeşitli mantarlarda,
bazı maya ve bakterilerde bulunurlar. Pektinleri enzimatik bir şekilde deesterifike
ederler. Deesterifikasyon sonunda metanolün ayrılmasıyla düşük esterli pektinlere ve
pektik asitlere dönüşürler.Pektin zincirinde tercih ettiği kırma noktası, serbest
karboksil ucuna yakın esterli galakturonik asit artıklarıdır.
Şekil 1.9. Pektin-esteraz enzimininin etki mekanizması 1.3.2. Endüstride Pektolitik Enzimlerin Kullanılması
Pektinazlar 60 yıldan daha fazla süredir gıda ve meyve suyu üretiminde
kullanılmaktadır. Ticari pektinaz üretiminde kullanılan asıl mikroorganizma
A.niger’dir. Pektinazlar elma, armut, üzüm, turunçgiller, kuru erik gibi meyve
sularının domates suyu gibi sebze sularının hazırlanmasında kullanılmaktadır.
O
OH
OH
OH
COOCH3
O
O
OH
OH
O
O
OH
OHOH
COOCH3
O
OH
OH
COOCH3
O
COOCH3
Pektin-liyaz
O
OH
OH
OH
COOCH3
O
O
OH
OH
O
COOCH3
O
OH
OH
COOH
O
O
OH
OH
COOH
OHOCH3
+ H20
Pektin-esteraz

1.GİRİŞ Fadile YENER
17
Yüksek poligalakturonaz aktivitesine sahip pektik enzimlerin kullanımı ile meyve
sularındaki bulanıklığın kararlı hale gelmesi başarılmıştır.
Pektin parçalayan enzimler gıda endüstrisinde özellikle meyve sularının
berraklaştırılmasında, meyve suyu üretiminde filtrasyonu kolaylaştırıp verimin
artırılmasında, C vitamini sentezi için çıkış maddesi olan galakturonik asit eldesinde,
şarap endüstrisinde, yağların ekstraksiyonunda, pigmentlerin ve selüloz fiberlerin
hazırlanmasında, kahve ve çay fermantasyonunda fonksiyonel gıda maddeleri olarak
oligosakkaritlerin üretiminde yeni uygulamalar için kullanılmaktadırlar (Telefoncu,
1997; Phutela ve ark., 2005; Patil ve ark., 2006; Botella ve ark., 2007).
1.4.Alginat
Alginat deniz yosunlarında bulunan bir polisakkarit olup, aralarında β (1-4)
glikozidik bağları olan D-manuronik (M) asit ve α (1-4) glikozidik bağları olan L-
guluronik asit (G)'ten meydana gelen organik bir bileşiktir (Sardar ve Gupta, 1998).
Endüstriyel olarak üretildiği mikroorganizmalar şunlardır:
§ Laminaria hyperborea
§ Macrocystis pyrifera
§ Ascophyllum nodosum
§ L.digitata, vb.
Alginat molekülünün bileşimi elde edildiği organizmaya ve dokuya göre
farklılıklar gösterir. Alginat iki ve üç değerlikli katyonlar ile reaksiyona girerek jel
oluşturur. Katyonlar (Ca+2, Sr+2, Ba+2, Al+3, Fe+3 gibi) moleküldeki guluronik asitleri
birbirlerine bağlayarak jel olşumuna sebep olurlar.
2 Na (Alginate) + Ca++ Ca (Alginate)2 + 2 Na+
İyonik bağlı jel 0-100oC arasındaki sıcaklıklarda kararlıdır. Bu nedenle
ısıtmak jeli sıvılaştırmaz. Yüksek sodyum veya potasyum içeren bir çözeltiyle
muamele edilirse kolaylıkla çözünür.

1.GİRİŞ Fadile YENER
18
Jel oluşturma özellikleri alginat molekülünün kompozisyon ve sırasına
bağlıdır. G bloklarının uzunluğu jel formasyonunun başlıca yapısal durumunu
belirler. Eğer molekülde G miktarı fazla ise sert ve kırılgan bir jel oluşur, eğer M
miktarı fazla ise daha yumuşak ve elastik jel oluşumu gözlenir.Genellikle hücrelerin
immobilizasyonunda kullanılan bu metod enzimler için kullanıldığında, bazı özel
tekniklerin kullanılması gereklidir. Alginat ile bead, kapsül ve fiber oluşturularak
enzim veya hücre immobilize edilebilir.
Hiçbir toksik etkisi olmadığından alginat, gıda, ilaç, tekstil, kağıt
endüstrisinde geniş kullanım alanına sahiptir (Smidsrod ve ark., 1990).
1.5.Florisil
Florisil (magnezyum silikat) yüksek mekanik dayanıklılığa sahip, organik
çözücülere dayanıklı, mikrobiyal saldırılara karşı dirençli inorganik desteklerdir.
Pektinaz enziminin alginat ve florisil destek üzerine immobilizasyonu,
immobilize enzimlerin karakterizasyonu (optimum pH, optimum sıcaklık, termal
kararlılık, depolama kararlılığı, tekrar kullanılabilirliği); immobilize enzimlerin
kinetik parametrelerinin (aktivasyon enerjisi, Km, Vmax, katalitik etkinliği)
belirlenmesi ve bu parametrelerin serbest Pektinaz ile kıyaslanması bu çalışmanın
temelini oluşturmaktadır.

2. ÖNCEKİ ÇALIŞMALAR Fadile YENER
19
2. ÖNCEKİ ÇALIŞMALAR
Roy ve ark., 2003 Aspergillus niger’den saflaştırılan Pektinaz enzimini
kovalent olmayan bir şekilde alginat üzerine immobilize etmişlerdir. Substrat olarak
kitini kullanmışlar, immobilize enzimin serbest enzime göre % 56 aktivite
gösterdiğini bildirmişlerdir. İmmobilize enzim serbest enzimle karşılaştırıldığında
optimum sıcaklığın değişmeden kaldığını optimum pH’nın ise geniş bir aralık
gösterdiğini rapor etmişlerdir. Alginat üzerine immobilize edilen Pektinaz’ın termal
kararlılığını 55 ve 65oC olarak bulmuşlardır. Serbest ve immobilize enzimin kinetik
parametrelerini çalışmışlar, Vmax değerinin değişmediğini KM değerinin
immobilizasyonla yükseldiğini göstermişlerdir. İmmobilize enzimin KM ve Vmax
değerlerini sırasıyla 6 mg ml-1 ve 3.4 nmol min-1 olarak bulmuşlardır. İmmobilize
enzimin ilk kullanımdan sonra aktivitesinin % 55 azaldığını göstermişler ve bu
azalmanın sebeplerini incelemişlerdir.
Ortega ve ark., 2004 Üç farklı ticari Pektinaz enzim kompleksindeki
(Rapidase C80, Pectinase CCM and Pectinex 3XL) poligalakturonaz aktivitesinin
termal inaktivasyonunu ve kinetik özelliklerini incelemişlerdir. Bütün örneklerde PG
aktivitesinin Michaelis-Menten kinetikleri ve onların katalitik etkinlikleri
incelemişlerdir. Pectinex 3XL için optimum pH’yı 4.7, diğer ikisi için optimum
pH’yı 4.0 olarak hesaplamışlardır. Rapidase C80, Pectinase CCM, Pectinex 3 XL
için aktivasyon enerjilerini sırasıyla 26.5, 45.6, 4.2 kj mol-1 hesaplamışlar ve termal
inaktivasyon eğrilerinin 40-60oC arasında doğrusal olmadığını rapor etmişlerdir.
Ayrıca enzim komplekslerinin serbest enerji (∆G#), entalpi (∆H#) ve entropi (∆S#)
gibi termodinamik aktivasyon parametrelerini hesaplamışlardır. En yüksek katalitik
etkinliği (Vmax/KM) 44.4 değeri ile Rapidase C80 gösterdiği için üç enzim kompleksi
arasındaki en etkin enzimin Rapidase C80 olduğunu belirlemişlerdir.
Sardar ve Gupta, 2005 Domatesten saflaştırdıkları Pektinaz’ı Concanavalin
A bağlı agaroza immobilize etmişlerdir. İmmobilize ve serbest enzimin katalitik
etkinliğini(Vmax/KM) sırasıyla 0.4 ve 0.3 olarak bulmuşlar ve immobilize enzimin
katalitik etkinliğinin daha yüksek olduğunu rapor etmişlerdir. İmmobilize enzimin

2. ÖNCEKİ ÇALIŞMALAR Fadile YENER
20
termal kararlılığının 50oC ve depolama kararlılığının 4oC’de yüksek olduğunu rapor
etmişlerdir. İmmobilize enzimi aktivitesi kaybolmadan 3 kez kullanabilmişlerdir.
Busto ve ark., 2006 Meyve suyu üretiminde kullanılan ticari pektinazlardan
pektinliyazı alginat boncuklar üzerine immobilize etmişler ve optimum koşulları 0,17
g alginat ml-1, % 1,2 enzim konsantrasyonu ve pH 3,6 asetik- HCl/glisin –HCl veya
pH 6,4 tris HCl / imidazol tamponu olarak belirlemişlerdir. İmmobilizasyonun
maksimum %’ sini (%10,6) Rapidase C 80 ile belirlemişlerdir. Serbest ve immobilize
enzimin optimum pH ve sıcaklık değerlerini 7,2 ve 55oC olarak bulmuşlardır.
İmmobilizasyonla termal kararlılığın özellikle 40oC ’de önemli şekilde
değişmediğini göstermişlerdir. Yüksek esterli pektinin viskozitesindeki azalmayı
serbest enzim için 1,09’dan 0,70 mm2 sn -1 ve immobilize enzim için 1,09’dan 0,72
mm2sn -1 olarak bulmuşlardır. Ayrıca enzimin 4 kez kullanılabileceğini ve viskozite
azalmasındaki verim kaybını sadece % 9,2 olarak belirlemişlerdir.
Aslan ve Tanrıseven, 2007 Aspergillus aculeatus’dan elde edilen ticari
enzim kompleksi Pectinex Ultra SP-L’yi Eupergit C üzerine immobilize etmişler ve
galaktooligosakkaritleri elde etmek için kullanmışlardır. İmmobilizasyonu % 100
bağlanma yüzdesi ve serbest enzimden % 24 daha yüksek GOS
(galaktooligosakkarit) verimi ile sonuçlandırmışlardır. Optimum koşulların
immobilizasyondan çok etkilenmediğini göstermişler, optimum pH ve optimum
sıcaklığı her iki enzim için sırasıyla 4.0-5.0 ve 55-60oC olarak hesaplamışlardır.
Serbest ve immobilize enzimi kullanarak % 30’luk laktoz çözeltisinden üretilen GOS
miktarını reaksiyon karışımındaki toplam şekerin sırasıyla % 12.8 ve % 15.8’i olarak
belirlemişlerdir. İmmobilize enzim 20 gün boyunca azalma olmaksızın aktivite
gösterdiği ve immobilizasyon yüzdesi yüksek olduğu için immobilize Pectinex Ultra
SP-L’nin galaktooligosakkaritlerin üretimi için kullanılabileceğini bildirmişlerdir.
Li ve ark., 2007 Pektinaz enzimini gluteraldehit kullanılarak sodyum alginat
destek üzerine kovalent bir şekilde immobilize etmişler ve aktivitesinin % 66’nı
koruduğunu bildirmişlerdir. İmmobilizasyondan sonra aktivite için en uygun pH
değerinin 3’den 3.5’a değiştiğini fakat optimum sıcaklığın değişmeden 40oC’de
kaldığını göstermişlerdir. İmmobilize enzimin serbest enzime göre daha yüksek
termal kararlılığa ve tekrar kullanılabilirliğe sahip olduğunu, 11 kullanımdan sonra

2. ÖNCEKİ ÇALIŞMALAR Fadile YENER
21
ilk aktivitesinin % 80’nini koruduğunu rapor etmişlerdir. Farklı substrat
değişimlerinde Vmax ve KM değerlerini çalışmışlar, Vmax değerini serbest enzim için
0,5 µmol/mg dk, immobilize enzim için 0,4µmol/mg dk bulmuşlardır. KM değerini
ise serbest enzim için 0,7 mg ml-1, immobilize enzim için 0,6 mg ml-1 olarak
bulmuşlardır.
Lei ve Bi., 2007b Pektinaz enzimini ATRP (atom transfer radikal
polimerizasyonu) kullanılarak sentezlenen gözenek boyutu belirli PS-b-PAA
(polisitiren-b-poliakrilik asit) kopolimeri üzerine immobilize etmişlerdir. Polimer
üzerindeki karboksil gruplarının enzim immobilizasyonu için çok basit, hafif ve
zaman tasarruflu bir süreç sunduğunu göstermişlerdir. İmmobilize pektinazın bazı
özelliklerini ( pH, termal kararlılık, sıcaklık, depolama kararlılığı) çalışmışlar,
bunlardan optimum pH’yı 6,0 optimum sıcaklığı 65 oC olarak bulmuşlardır. Serbest
ve immobilize enzimin aktivitelerini farklı substrat derişimlerinde ölçmüşler ve KM
ve Vmax değerlerinin değişmediğini görmüşlerdir. Bunun sonucunda immobilize
enzimin serbest enzimle benzer hızda çalıştığını göstermişlerdir. Fakat immobilize
enzimin KM değerinin, serbest enziminkine göre daha yüksek olduğunu ortaya
çıkarmışlardır.
Lei ve ark., 2007a Pektinazı SiO2 üzerine daha önceden adsorbe edilmiş
kitozan yüzeyine immobilize etmeye çalışmışlardır. Kitozanın enzim
immobilizasyonu için uygun bir destek olduğunu ve gluteraldehit gibi reaktiflerle
kolay bir şekilde karşı bağ yaparak jel haline gelebildiğini bunun içinde iyi bir destek
olduğunu belirtmişlerdir. Glutaraldehitle aktifleştirilmiş silika kaplı kitozan destek
olarak kullanıldığında immobilize enzimin termal ve pH denaturasyonuna karşı daha
dirençli olduğunu belirtmişlerdir. Bu immobilize enzimin aktivitesini kaybetmeden
geniş bir pH aralığında (3-4.5) uygulanabileceğini göstermişlerdir. Bu partiküller
üzerine adsorbe edilen immobilize enzimin aktivitesini Michaelis-Menten
parametreleriyle analiz etmişlerdir. KM değerlerini serbest ve immobilize enzim için
sırasıyla 8.28 g pektin ml-1 ve 10.03 g pektin ml-1 olarak bulmuşlardır. KM
değerindeki ufak değişimin sebebini destek ve polimer zincirlerinin üç boyutlu
yapısıyla oluşan difüzyon etkileri olarak açıklamışlardır.

3.MATERYAL VE METOD Fadile YENER
22
3.MATERYAL VE METOD 3.1.Materyal
Araştırmada kullanılan tüm reaktifler analitik saflıkta olup Merck veya
Sigma, St. Louis, MO. Firmasından sağlanmıştır.Araştırmada kullanılan kimyasallar,
araç ve gereçler şunlardır:
Kimyasallar: Pektinaz (Novenzymes), alginat, galakturonik asit, sodyum
hidroksit, sodyum karbonat, sodyum potasyum tartarat, 3,5-dinitrosalisilik asit, fenol,
sodyum sülfit, florosil (60-100 mesh), glutaraldehit, 3-aminopropil trietoksisilan,
nitrik asit, aseton, bakır (II) sülfat, folin-ciocalteu çözeltisi, sodyum klorür, fosforik
asit, asetik asit, sitrik asit, sığır serum albumin (BSA).
Araç ve gereçler: UV- Vis spektrofotometre (ATI UNICAM), pH metre
(HANNA 8417), magnetik karıştırıcı, inkübatör (ES 500), santrifüj, analitik terazi,
otomatik pipet, girdap karıştırıcı, termostatlı çalkalayıcı su banyosu, kriyostat.
3.2.Metod
3.2.1.Protein Tayini
Protein tayini Lowry (1951) yöntemiyle yapılmıştır. Bunun için içerikleri
bildirilen A, B ve C çözeltileri hazırlanmıştır.
1) Çözelti A: 20 g Na2CO3 ve 4 g NaOH saf suda birlikte çözülüp son hacim 1 L’ye
tamamlanarak hazırlanmıştır.
2) Çözelti B: 0,5 g CuSO4.5H2O, % 1’lik sodyum sitrat çözeltisinde çözülüp son
hacim aynı çözelti ile 100 mL’ye tamamlanarak hazırlanmıştır.
3) Çözelti C: 50 mL A çözeltisi ile 1 mL B çözeltisi karıştırılarak hazırlanmıştır.
(Kullanılacağı zaman hazırlanmasına dikkat edilmiştir).
4) Folin-Ciocalteu çözeltisi: Folin-Ciocalteu saf su ile 1:2 oranında seyreltilerek
hazırlanmıştır.

3.MATERYAL VE METOD Fadile YENER
23
5) Standart protein çözeltisi:100 mL’de 14 mg sığır albümini olacak şekilde 0,9’luk
NaCl çözeltisi ile hazırlanmıştır.
6) Standart protein eğrisinin çizimi: 8 adet deney tüpü alınarak tüplere sırasıyla 0,
50, 100, 125, 250, 500, 750 ve 1000 µL olacak şekilde standart protein
çözeltisinden konulmuştur. Her tüp içeriğinin hacmi su ile 1 mL’ye
tamamlanmış, her tüpe 5 mL C çözeltisi ilave edilmiştir. 10 dakika oda
sıcaklığında bekletildikten sonra her tüpe 1:2 oranında seyreltilmiş Folin-
Ciocalteu çözeltisinden 0,5 mL eklenmiştir. 30 dakika oda sıcaklığında bekletilip
tüp içeriklerinin absorbansları köre karşı 750 nm’de okunmuştur, bu değerler
derişime karşı grafiğe geçirilmiştir. Örneklerin protein içerikleri aynı yöntemle
standart protein eğrisi kullanılarak değerlendirilmiştir.
3.2.2.Pektinaz’ın Alginat Destek Üzerine İmmobilizasyonu
Enzimlerin immobilizasyonunda Mondal ve ark.(2003), tarafından önerilen
yöntem kullanılmıştır.30 µl enzim çözeltisi (3,75 mg/ml) pH 5,0 sodyum asetat
tamponu ile 1 ml’ye tamamlanmıştır.Sonra % 2’lik 2 ml alginat çözeltisi eklenmiş
ve son hacim asetat tamponu ile 4 ml’ye tamamlanmıştır. Reaksiyon karışımı 3M
asetik asitle pH 3,8’e getirilmiştir.25oC’de bir saat bekletildikten sonra 2M CaCl2
çözeltisinden 400 µl eklenmiş ve 20 dakika sonunda santrifüj edilmiştir.
İmmobilizasyon işleminden sonra immobilize enzim saf su ile süzüntüde protein
kalmayıncaya kadar asetat tamponu ile yıkanmıştır.Toplam süzüntüde protein tayini
yapılarak g destek başına tutuklanan mg enzim miktarı hesaplanmıştır.Protein tayini
Lowry metoduna göre yapılmıştır.
3.2.3.Pektinaz’ın Florisil Destek Üzerine İmmobilizasyonu
Pektinaz’ın immobilizasyonunda önce florisil destek yüzeyinin aktifleştirilmesi
gerekmektedir (Tükel ve Alptekin, 2004). Bu nedenle 3- aminopropiltrietoksisilan
kullanılarak desteğin alkilamin türevi oluşturulmuştur. Alkilamin türevinin
hazırlanmasında (silanlama) Weetall, H.H. (1976) tarafından literatürde bildirilen
yöntem kullanılmıştır. Destek materyali %5 (v/v)’lik nitrik asit ile 80-90 ºC’de 60

3.MATERYAL VE METOD Fadile YENER
24
dakika yıkanmış ve takiben su ile yıkanıp 120 ºC’de kurutulmuştur. Kurutulmuş
desteğin 1 gramı hacimce % 4’lük 3- aminopropiltrietoksisilanın aseton
içerisindeki çözeltisine eklenerek 45 oC’de 24 saat bekletilmiştir. Saf su ile
yıkanarak 115 oC’de etüvde 1 gece kurutulmuştur.1 g silanlanmış desteğe 25 ml %
2,5 (v/v)`luk glutaraldehitin 50 mM pH 7,0 fosfat tamponundaki çözeltisi eklenmiş
120 dakika beklendikten sonra, reaksiyona girmemiş glutaraldehit saf su ile
yıkanarak uzaklaştırılmış ve 60 oC’de 1 sa kurutulmuştur.Bu basamaktan sonra
enzim destek üzerine doğruda immobilize edilmeye çalışılmıştır ( Costa ve ark.,
2001).
3.2.4.Serbest veya İmmobilize Pektinaz’ın Aktivitesinin Belirlenmesi
Pektinaz’ın aktivite tayini için Debing ve ark.(2006), önerdiği yöntem
kullanılmıştır. Pektinaz enzim kompleksinde exopoligalakturonaz aktivitesi
ölçülmüştür. Serbest enzim için 50oC’de 10 dakika bekletilen reaksiyon karışımında
pektinin hidrolizi sonucu oluşan galakturonik asit miktarı Dinitrosalisilik asit
(DNSA) yöntemi ile belirlenmiştir.
DNSA yöntemi ilk kez Sumner ve ark. (1921) tarafından geliştirilmiştir.
Yöntem bir indirgen şekerin 3,5-Dinitrosalisilik asit ile yükseltgenmesi esasına
dayanmaktadır.
Bu çalışmada kullanılan DNSA reaktifinin bileşimi (w/v) olarak aşağıda
verilmiştir (Wang ve ark., 1997).
% 1 DNSA
% 0,2 Fenol
% 0,05 Sodyum Sülfit
% 1 Sodyum Hidroksit
% 30 Sodyum-Potasyum Tartarat
DNSA yöntemi ile galakturonik asit tayini yapmak için 300 µl galakturonik
asit çözeltisi ile 300 µl DNSA reaktifi bir tüpte karıştırılır, kaynar su banyosunda 15
dakika tutulur.Tüm tüpler aynı anda kaynar su banyosundan alındıktan sonra buz

3.MATERYAL VE METOD Fadile YENER
25
banyosunda 2-3 dakika soğutulur. Çözelti üzerine 4,4 ml damıtık su eklenerek
seyreltilir ve girdap karıştırıcı ile karıştırıldıktan sonra spektrofotometrede 530
nm’de okunur.
Exopoligalakturonaz aktivitesini belirlemek için 0,2 mg/ml derişimindeki
serbest Pektinaz’ın 50 µl’si 250 µl % 1’lik pektin çözeltisi ile 50oC’de 10 dk
etkileştirilmiştir.Reaksiyonu durdurmak için ortama 300 µl DNSA reaktifi
eklenmiştir.Açığa çıkan galakturonik asit miktarı belirli galakturonik asit
derişimlerine karşı 530 nm’de absorbans değerlerinin grafiğe geçirilmesiyle elde
edilen standart galakturonik asit eğrisinden yararlanılarak bulunmuştur.Enzimin
aktivitesi µmol galakturonik asit dk-1 mg-1 protein olarak hesaplanmıştır.
İmmobilize Pektinaz aktivitesinin ölçümünde, immobilize Pektinaz içeren 0,2
g destek % 1’lik pektin çözeltisinin 250 µl’si ile 45 oC’de 10 dk etkileştirilmiş ve
reaksiyon DNSA reaktifi ile durdurulmuştur.Açığa çıkan galakturonik asit miktarı
standart eğriden yararlanılarak bulunmuştur.Enzimin aktivitesi µmol galakturonik
asit dk-1 g-1destek olarak hesaplanmıştır.
3.2.5.Galakturonik Asit Tayini
Standart galakturonik asit eğrisinin çizimi: 6 adet deney tüpüne sırasıyla 0,5,
1, 2, 3, 4 v 5 mM derişimlerde olacak şekilde 300 µl galakturonik asit çözeltisi ve
kör için ise 300 µl saf su konulmuştur. Her bir tüpe DNSA reaktifinden 300µl
eklendikten sonra 15 dakika kaynar su banyosunda bekletilmiştir.Bu sürenin sonunda
2-3 dakika buz içerisinde bekletilen tüplere 4,4 ml saf su eklenerek seyreltilmiş ve
tüplerin absorbansları 530 nm’de okunmuştur.Bu değerler derişime karşı grafiğe
geçirilmiştir.Örneklerin galakturonik asit içerikleri aynı yöntemle standart
galakturonik asit grafiği kullanılarak değerlendirilmiştir (Debing, J., 2006).

3.MATERYAL VE METOD Fadile YENER
26
3.2.6 Serbest ve İmmobilize Enzimlerin Karakterizasyonu.
3.2.6.1. Serbest ve İmmobilize Enzimlerin Optimum pH'larının Belirlenmesi
Serbest ve immobilize Pektinaz’ın aktivitesi farklı pH'larda (3.5, 4, 4.5, 5.0
asetat, 6.0 sitrat) sırasıyla 50 mM ve 25 mM tampon içinde hazırlanmış % 1’lik
pektin çözeltisi kullanılarak belirlenmiştir. 10 dakika sonunda oluşan galakturonik
asit miktarı ölçülerek aktiviteler hesaplanmış ve sonuçlar % maksimum aktivite
olarak pH'ya karşı grafiğe geçirilmiştir.
3.2.6.2. Serbest ve İmmobilize Enzimlerin Optimum Tampon Derişimlerinin
Belirlenmesi
Serbest ve immobilize Pektinaz’ın aktiviteleri daha önce belirlenmiş olan
optimum pH'larda ve farklı tampon derişimlerinde (5, 10, 25, 50, 75, 100 mM)
hazırlanmış substrat çözeltileri kullanılarak ölçülmüştür.
3.2.6.3. Serbest ve İmmobilize Enzimlerin Optimum Sıcaklıklarının
Belirlenmesi
Serbest ve immobilize Pektinaz’ın aktiviteleri daha önce belirlenmiş olan
optimum pH ve tampon derişimleri için farklı sıcaklıklarda (35, 40, 45, 50, 60, 70°C)
belirlenmiştir.
Her enzim örneği için aktivitenin sıcaklıkla arttığı ve azaldığı bölgeler için
Arrhenius grafiği çizilmiş ve elde edilen iki doğrunun ortak çözümü ile enzimin
denatüre olmaya başladığı sıcaklık belirlenmiştir. Aktivitenin sıcaklıkla arttığı bölge
için elde edilen grafiğin eğimi yardımıyla enzimin aktivasyon enerjisi hesaplanmıştır.

3.MATERYAL VE METOD Fadile YENER
27
3.2.6.4. Serbest ve İmmobilize Enzimlerin KM ve Vmax Değerlerinin Belirlenmesi
Serbest ve immobilize Pektinaz’ın aktiviteleri optimum şartlarda farklı
substrat derişimlerinde ölçülmüş ve Lineweaver-Burk grafiği yardımıyla KM ve Vmax
değerleri belirlenmiştir. Ayrıca serbest ve immobilize Pektinaz enzimi için katalitik
etkinlik parametresi olarak belirlenen Vmax/Km değerleri hesaplanmıştır.
3.2.6.5. Serbest ve İmmobilize Enzimlerin Termal Kararlılıklarının Belirlenmesi
Serbest ve immobilize Pektinaz 4 farklı sıcaklıkta (serbest enzim için; 5, 25,
50, 60°C ve immobilize enzim için; 5, 25, 45, 60oC) 18 saat süreyle inkübe edilmiş
ve belirli aralıklarla (başlangıç, 1, 3, 7 ve 18 saat) kalan aktiviteleri belirlenmiştir.
3.2.6.6. Serbest ve İmmobilize Pektinaz’ın Depolama Kararlılıklarının
Belirlenmesi
Serbest ve immobilize Pektinaz’ın başlangıç aktiviteleri belirlendikten sonra
4oC ve oda sıcaklığında (25oC) ağzı kapalı şişelerde bekletilmiş ve bu enzimlerin
kalan aktiviteleri 5 hafta boyunca belirli zaman ararlıklarla ölçülmüştür.
3.2.6.7. İmmobilize Pektinaz’ın Tekrar Kullanım Kararlılığının Belirlenmesi
İmmobilize enzimin tekrar kullanım kararlılığı belirlenirken kesikli ve
karıştırmalı kolon reaktör kullanılmıştır.
1 g immobilize enzim bulunan kolona 1 ml % 1’lik pektin çözeltisi eklenmiş
ve 10 dakika sonunda alttaki musluğun açılması ile kolan boşaltılmıştır. Enzim
aktivitesi oluşan galakturonik asit miktarı ölçülerek hesaplanmıştır. Kolon
boşaltıldıktan sonra tekrar 1 ml subsrat çözeltisi eklenerek aynı şekilde aktivite
belirlenmiş ve bu işlem 24 kez tekrarlanmıştır. Kalan aktivite ilk kullanımdaki
aktivitenin %’si olarak hesaplanmıştır.

4.BULGULAR VE TARTIŞMA Fadile YENER
28
4.BULGULAR ve TARTIŞMA
4.1.Bulgular
4.1.1.Serbest ve İmmobilize Pektinaz’ın Karakterizasyonu
Serbest ve immobilize Pektinaz’ın farklı pH’daki aktiviteleri % maksimum
aktivite olarak Şekil 4.1’de verilmektedir.
0
20
40
60
80
100
120
3 4 5 6 7
pH
% M
aksim
um A
ktiv
ite
Şekil 4.1.Serbest ve İmmobilize Pektinaz aktivitesinin pH’ya bağlı olarak değişimi
(•):İmmobilize Pektinaz, (ο):Serbest Pektinaz Şekil 4.1’de görüldüğü gibi serbest ve immobilize Pektinaz’ın her ikisi de en
yüksek aktiviteyi pH 5.0’da göstermektedir.
Serbest ve immobilize Pektinaz’ın pH 5.0’da hazırlanmış olan farklı
derişimlerdeki tampon çözeltileri kullanılarak aktiviteleri belirlenmiş ve sonuçlar %
maksimum aktivite olarak Şekil 4.2’de gösterilmektedir.

4.BULGULAR VE TARTIŞMA Fadile YENER
29
0
20
40
60
80
100
120
0 25 50 75 100 125 150 175 200
Tampon Derişimi(mM)
% A
ktiv
ite
Şekil 4.2.Serbest ve immobilize Pektinaz aktivitesinin tampon derişimine bağlı olarak değişimi (o):İmmobilize Pektinaz, (•):Serbest Pektinaz
Şekil 4.2’de görüldüğü gibi serbest enzimin en yüksek aktiviteyi gösterdiği
tampon derişimi 50 mM olarak belirlenirken immobilize enzim için bu değer 25 mM
olarak bulunmuştur.
Serbest ve immobilize Pektinaz’ın aktivitesi farklı sıcaklıklarda ölçülmüş,
aktivitenin sıcaklığa bağlı olarak değişimi % maksimum aktivite olarak Şekil 4.3’de
gösterilmiştir. Serbest Pektinaz’ın maksimum aktivite gösterdiği sıcaklık 50°C olarak
belirlenirken immobilize Pektinaz’ın maksimum aktivite gösterdiği sıcaklık 45°C
olarak bulunmuştur.

4.BULGULAR VE TARTIŞMA Fadile YENER
30
0
20
40
60
80
100
120
0 20 40 60 80
Sıcaklık(oC)
% M
aksim
um A
ktiv
ite
Şekil 4.3.Serbest ve immobilize Pektinaz aktivitesinin sıcaklığa bağlı olarak değişimi
(•):İmmobilize Pektinaz, (o):Serbest Pektinaz
Serbest Pektinaz’ın sıcaklığa bağlı olarak değişen aktivite değerleri Arrhenius
bağıntısına göre grafiğe geçirildiğinde Şekil 4.4 elde edilmiştir. Grafikte aktivitenin
arttığı ve azaldığı bölgeler ayrı ayrı değerlendirilmiştir.
Aktivitenin sıcaklıkla arttığı doğrunun eğimini kullanarak serbest
Pektinaz’ın aktivasyon enerjisi 32 J mol-1K-1 olarak hesaplanmıştır. Her iki doğrunun
ortak çözümü ile enzimin denaturasyona başladığı sıcaklık 50,1 oC olarak
belirlenmiştir.
y = -3,8466x + 15,622
y = 5.884x - 14.492
2
2.5
3
3.5
4
2.8 2.9 3 3.1 3.2 3.3
1/T (K-1)x10-3
ln V
(U m
g PG
-1)
Şekil 4.4. Serbest Pektinaz’ın Arrhenius grafiği
4.BULGULAR VE TARTIŞMA Fadile YENER
31
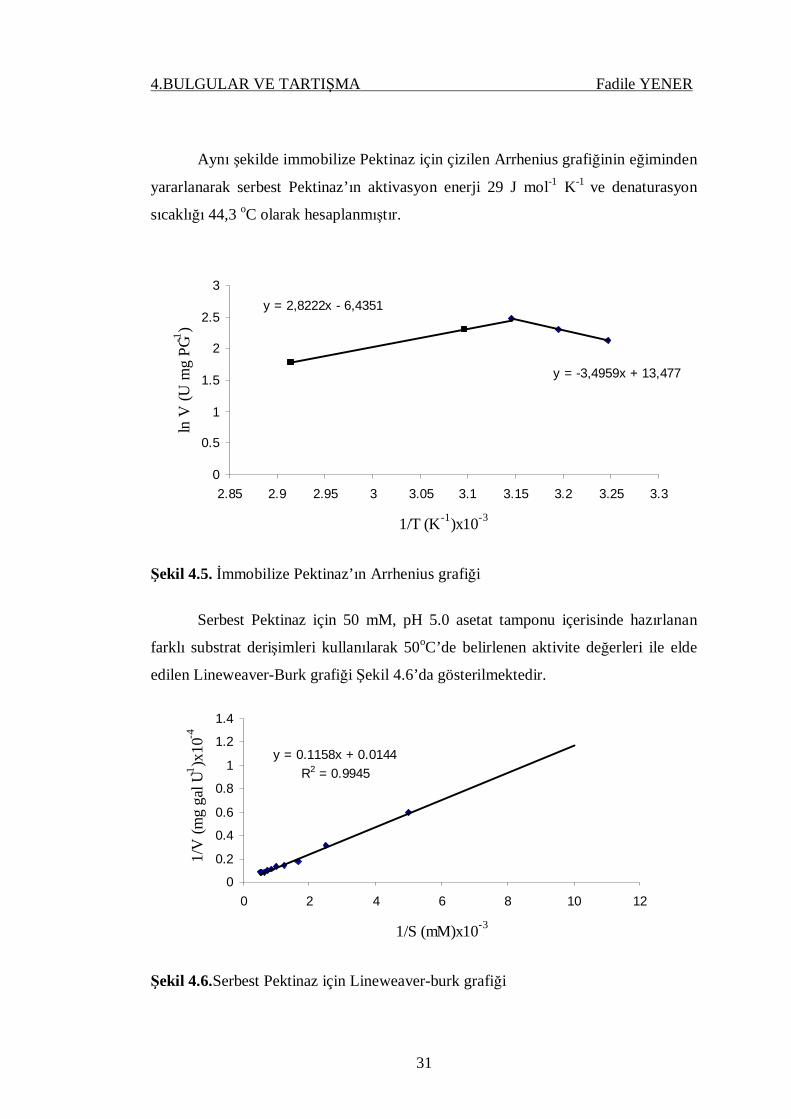
Aynı şekilde immobilize Pektinaz için çizilen Arrhenius grafiğinin eğiminden
yararlanarak serbest Pektinaz’ın aktivasyon enerji 29 J mol-1 K-1 ve denaturasyon
sıcaklığı 44,3 oC olarak hesaplanmıştır.
y = -3,4959x + 13,477
y = 2,8222x - 6,4351
0
0.5
1
1.5
2
2.5
3
2.85 2.9 2.95 3 3.05 3.1 3.15 3.2 3.25 3.3
1/T (K-1)x10-3
ln V
(U m
g PG
-1)
Şekil 4.5. İmmobilize Pektinaz’ın Arrhenius grafiği
Serbest Pektinaz için 50 mM, pH 5.0 asetat tamponu içerisinde hazırlanan
farklı substrat derişimleri kullanılarak 50oC’de belirlenen aktivite değerleri ile elde
edilen Lineweaver-Burk grafiği Şekil 4.6’da gösterilmektedir.
y = 0.1158x + 0.0144R2 = 0.9945
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0 2 4 6 8 10 12
1/S (mM)x10-3
1/V
(mg
gal U
-1)x
10-4
Şekil 4.6.Serbest Pektinaz için Lineweaver-burk grafiği

4.BULGULAR VE TARTIŞMA Fadile YENER
32
İmmobilize Pektinaz için önceden belirlenmiş optimum koşullarda hazırlanan
farklı substrat derişimlerinde, belirlenen aktivite değerleri kullanılarak elde edilen
Lineweaver-Burk grafiği Şekil 4.7’de gösterilmektedir.
y = 0.0178x + 0.0484R2 = 0.9981
0
0.05
0.1
0.15
0.2
0.25
0 2 4 6 8 10 12
1/S (mM)x10-3
1/V
(mg
Gal
U-1)x
10-4
Şekil 4.7. İmmobilize Pektinaz için Lineweaver-Burk grafiği
Serbest ve immobilize Pektinaz için KM, Vmax ve Vmax/Km değerleri her bir
enzim için çizilen Lineweaver-Burk grafiğinden hesaplanarak bulunmuştur.Bu
sonuçlar Çizelge 4.1’de gösterilmektedir.
Çizelge 4.1. Serbest ve immobilize Pektinaz’ın kinetik parametreleri
Serbest Pektinaz İmmobilize Pektinaz
KM (mg/ml) 8,04 0.37
Vmax (µmolgalakturonik
asit/mg protein.dk)
69,4 20,7
Vmax / KM 8.63 55.9
Çizelgede 4.1’de görüldüğü gibi immobilize Pektinaz’ın KM ve Vmax
değerlerinin ikiside azalmıştır. Buna göre katalitik etkinlik olarak tanımlanan Vmax /
KM değerinin serbest enzim için 8,63 iken, immobilize enzim için bu değer 55,9

4.BULGULAR VE TARTIŞMA Fadile YENER
33
olarak hesaplanmıştır. İmmobilize enzimin katalitik etkinliği serbest enzime göre
yaklaşık 6.5 kat artmıştır.
Şekil 4.8 ve 4.9’da sırasıyla serbest ve immobilize Pektinaz için termal
kararlılık grafikleri verilmektedir.
0
20
40
60
80
100
120
0 5 10 15 20
İnkübasyon süresi (saat)
Kal
an A
ktiv
ite (%
)
Şekil 4.8.Serbest Pektinaz enziminin termal kararlılık grafiği.(▲):5oC, (□):25oC,
(■):50oC, (∆):60oC
Serbest Pektinaz’ın farklı inkübasyon süreleri sonunda kalan aktivitelerini
gösteren Şekil 4.8 incelendiğinde 5oC’de 16 saat inkübe edilen enzimin termal
kararlılığı yüksek olup inkübasyon sonunda kalan aktivitesi yaklaşık % 81’dir.16 saat
sonunda 25 ve 50oC’de inkübe edilen serbest Pektinaz enzimi başlangıç
aktivitelerinin sırasıyla % 75 ve % 68’ini korurken, 60oC’de 16 saat sonunda
başlangıç aktivitesinin % 19’nu koruduğu belirlenmiştir.
İmmobilize Pektinaz’ın termal kararlılığını veren Şekil 4.9 incelendiğinde,
immobilize enzimin termal kararlılığının serbest enzime göre çok farklı olmadığı
gözlenmektedir. İmmobilize Pektinaz’ın 5oC’de 16 saat inkübasyonu sonunda kalan
aktivitesi yaklaşık % 83 iken, 60oC’de 16 saat inkübasyonu sonunda kalan aktivitesi
% 27’dir.

4.BULGULAR VE TARTIŞMA Fadile YENER
34
0
20
40
60
80
100
120
0 5 10 15 20
İnkübasyon süresi (saat)
Kal
an A
ktiv
ite (%
)
Şekil 4.9. İmmobilize Pektinaz enziminin termal kararlılık grafiği.(▲):5oC,
(□):25oC, (■):45oC, (∆):60oC. Serbest Pektinaz’ın 5oC ve oda sıcaklığındaki (25oC) depolama kararlılığı
Şekil 4.10’da gösterilmektedir.
0
20
40
60
80
100
120
0 5 10 15 20
Zaman (Gün)
Kal
an A
ktiv
ite %
Şekil 4.10. Serbest Pektinaz’ın Depolama Kararlılığı. (□):5oC, (■):25oC

4.BULGULAR VE TARTIŞMA Fadile YENER
35
Şekil 4.10’da görüldüğü gibi serbest Pektinaz enzimi 5oC ve 25oC
sıcaklıklarında 5 hafta depolanmıştır.Depolanan serbest enzim her iki sıcaklıkta da
benzer bir aktivite değişimi göstermektedir. 5oC ve 25oC’de depolanan iki enzimde
ikinci haftanın sonunda başlangıç aktivitelerinin % 92 ‘sini korumaktadırlar.
Şekil 4.11’de immobilize Pektinaz’ın 5oC ve 25oC depolama kararlılıkları
gösterilmektedir.
0
20
40
60
80
100
120
0 5 10 15 20
Zaman (Gün)
Kal
an A
ktiv
ite %
Şekil 4.11. İmmobilize Pektinaz’ın Depolama Kararlılığı. (□):5oC, (■):25oC
Şekil 4.11’de görüldüğü gibi serbest enzim 5 hafta boyunca 5oC ve 25oC
sıcaklıklarında depolanmıştır. 5 hafta boyunca depolanan immobilize enzim yine
benzer bir aktive göstermiş 5 inci haftanın sonunda 5oC’de başlangıç aktivitesinin %
75’ini, 25oC’de ise % 76’sını korumuştur.

4.BULGULAR VE TARTIŞMA Fadile YENER
36
İmmobilize Pektinaz’ın kullanım sayısına bağlı olarak aktivitesindeki
değişimi Şekil 4.12’de görülmektedir.
0
20
40
60
80
100
120
0 5 10 15 20 25
kulanım sayısı
Kal
an a
ktiv
ite (%
)
Şekil 4.12. İmmobilize Pektinaz’ın tekrar kullanım kararlılığı
İmmobilize Pektinaz, 20 kullanımdan sonra başlangıç aktivitesinin % 94’ünü
koruyabilmektedir.
4.1.2. Pektinaz’ın Florisile İmmobilizasyonu
Pektinaz enzimi yaklaşık % 72 bağlanma yüzdesiyle florisil destek üzerine
bağlanabilmiştir. Fakat aktivite ölçümlerinden hiçbir sonuç alınamamıştır.
Çizelge 4.2. Pektinaz’ın Florisile İmmobilizasyonu
Enzim derişimi
(mg/ml)
Bağlanan protein
(mg protein)
% Bağlanma Aktivite
1 0.72 72 -
2 1.51 75 -
3 2.17 72 -
5 3.15 70 -
10 6.07 70 -

4.BULGULAR VE TARTIŞMA Fadile YENER
37
4.2. Tartışma
4.2.1.Pektinaz’ın Alginat Destek Üzerine İmmobilizasyonu ve Karakterizasyonu
Pektinaz’ ın alginat desteğe immobilizasyonunda Mondal ve Gupta (2003)
tarafından belirlenen yöntem kullanılmıştır. Pektinaz’ ın standart koşullar altında
alginat üzerine immobilizasyonu sonucunda başlangıçta ortamda bulunan protein
miktarının % 83’ ünün bağlandığı hesaplanmıştır. Enzim tutuklama yöntemi ile
alginat desteğe immobilize edilmiştir. İmmobilize enzimin aktivitesi substrat olarak
pektin kullanılarak Miller, (1959) tarafından belirlenen ve birçok araştırmacı
tarafından da kullanılan yöntem ile belirlenmiştir (Roy ve ark., 2003; Sardar ve ark.,
2005; Debing ve ark., 2006; Lei ve ark., 2007). Alginat kalsiyum iyonları varlığında
jel formuna dönüşmekte ve kalsiyum-alginat jeller oluşmaktadır. Ca-alginat jeller
genellikle tüm hücreyi ya da enzimlerin tutuklanması için kullanılmaktadırlar.
Alginat geri dönüştürülebilir şekilde çözülür-çözülmez bir polimerdir.Bu polimer
Ca+2 iyonları ile çöktürülebilir ve diyaliz ile veya EDTA’nın eklenmesiyle geri
çözülebilir (Roy ve ark., 2003). Şimdiye kadar Pektinaz’ı naylon, iyon-değiştirici
reçine, deri ve kitin gibi çeşitli desteklere immobilize etmişlerdir. Alginat da son
zamanlarda geniş bir şekilde uygulama alanı bulan, enzimi jel halinde tutuklama
yöntemi ile immobilize edebilen bir destek ortamıdır (Li ve ark., 2007).
Roy ve ark., (2003), kitini hidroliz etmek için alginata immobilize edilmiş
Pektinaz kompleksini kullanmışlardır. Kovalent bağlarla yapılan immobilizasyon ile
kıyaslama yapmışlardır. Alginatın toksik etkisinin bulunmaması ve ekonomik
olmasından dolayı kitin hidrolizi için kolaylıkla kullanılabileceğini rapor etmişlerdir.
Alginat destek üzerine immobilize edilen Pektinaz’ın aktivitesi ve serbest
Pektinaz’ın aktivitesi farklı ortam pH’ larında ölçülmüş olup immobilizasyon
sonucunda immobilize Pektinaz’ ın optimum pH değerinin serbest Pektinaz’ ın
optimum pH’sına göre değişmediği, 5.0 olarak kaldığı belirlenmiştir (Şekil 4.1).
Literatürde immobilize ve serbest Pektinaz’ın pH değerinin 4.0 ile 5.0
arasında olduğu bildirilmektedir. Ancak farklı desteklere değişik yöntemlerle
immobilize edilmiş olan Pektinaz enzimi için pH 3.0-5.0 aralığında değişen farklı

4.BULGULAR VE TARTIŞMA Fadile YENER
38
optimum pH değerleri rapor edilmiştir (Li ve ark., 2007). Optimum pH‘daki bu
kaymaların büyüklüğü ve yönü enzimin immobilize edildiği desteğin özelliklerine,
desteğin aktifleştirilmesi için kullanılan metoda ve immobilizasyon yöntemine bağlı
olarak değişmektedir.
Enzimatik reaksiyonlarda reaksiyon ortamında bulunan iyonlar enzim
stabilizasyonunun sağlanmasında önemli rol oynamaktadır. Düşük iyon derişimine
sahip tampon çözeltiler enzim stabilizasyonunu sağlamada yetersiz kalırken yüksek
tampon derişimi de enzimle substrat derişimini azaltacağı için aktivitenin düşmesine
neden olabilmektedir. Ancak Pektinaz enziminin aktivitesi tampon derişiminden
reaksiyon ürününün galakturonik asit olması nedeniyle de tampon çözeltilerin
tamponlama kapasitesi önem kazanmaktadır. Şekil 4.2 ‘de serbest Pektinaz
aktivitesinin tampon derişiminden fazla etkilenmediği, immobilize Pektinaz
enziminin ise tampon derişiminden etkilendiği görülmektedir. Literatürde Pektinaz
aktivitesi üzerine iyonik gücün etkisi araştırılmamıştır. % maksimum aktivite-
sıcaklık grafiğine göre serbest enzimin immobilize enzimle farklı sıcaklıklarda
optimum gösterdiği görülmektedir. Serbest enzim 50oC’ de optimum sıcaklık
gösterirken immobilize enzim 45oC’ de göstermektedir. Bu sonuçlar serbest enzim
için literatürdeki enzimlerle benzer sonuçlar göstermektedir. (Chellegetti ve ark.,
2002; Silva ve ark., 2005; Barense ve ark., 2001).
Aktivasyon ile ilgili bulgulara göre immobilize enzimin katalizlediği
reaksiyonun aktivasyon enerjisi (Ea) 29 Jmol-1K-1 ve serbest enzimin katalizlediği
reaksiyonun aktivasyon enerjisi 32 Jmol-1K-1 olarak hesaplanmıştır.İmmobilize
enzimin denaturasyona uğradığı sıcaklık 44,3oC olarak hesaplanmıştır.Aktivasyon
enerjisi ile ilgili araştırmaya pek rastlanmamıştır.
Serbest Pektinaz’ ın alginat desteğe immobilizasyonu sonucu hem KM hem de
Vmax değerlerinin azaldığı görülmektedir (Çizelge 4.1). İmmobilizasyon işlemi
sonunda enzimin desteğe bağlanma miktarının yüksek olmasına rağmen (yaklaşık
%83) immobilize Pektinaz’ ın aktivitesinin daha düşük görülmesi desteğe tutuklanan
enzim molekülünün hareketinin kısıtlanmış olmasına ve üç boyutlu yapısında
oluşabilecek değişime bağlanabilir. Bunun dışında destek çevresinde proteinlerin üst
üste birikerek difüzyon problemlerinin görülmesi söylenebilir.

4.BULGULAR VE TARTIŞMA Fadile YENER
39
Literatürde enzimin katalitik etkinliğinin bir ölçüsü olarak Vmax / KM değeri
kullanılmıştır. Vmax/KM değerinin endüstriyel uygulamalarda en etkili enzimin seçimi
için kullanışlı bir parametre olduğunu rapor etmişlerdir (Roy, 2003; Ortega, 2004;
Sardar, 2005).
Bulgularımızda serbest enzim için Vmax / KM değeri 8,6 immobilize enzim
için Vmax / KM değeri 55,9 olarak bulunmuştur. İmmobilize enzim serbest enzimle
karşılaştırıldığında daha yüksek katalitik etkinlik göstermiştir.Yani immobilize
enzim daha fazla poligalakturonik asit hidroliz etmiştir. Sardar ve ark., (2005),
Pektinaz’ı Con-A bağlı agaroz desteğe immobilize ettiklerinde immobilize enzim
serbest enzime göre yaklaşık 1.3 kat daha yüksek katalitik etkinlik göstermiştir.
Ortega ve ark., (2004), üç farklı ticari Pektinaz enzim kompleksindeki (Rapidase
C80, Pectinase CCM, Pectinex 3XL) poligalakturonaz aktivitesinin kinetik
özelliklerini karşılaştırmışlardır. 44,4 değeri ile en yüksek Vmax/KM değerini gösteren
Rapidase C80’ni en etkili enzim kompleksi olarak rapor etmişlerdir.
Enzimin kararlılığı verimliliğini etkileyen faktörlerden bir tanesidir. Enzimler
solüsyonları içinde depolanmaları süresince çok kararlı değillerdir ve zamanla
aktivitelerini kaybederler. Belirli zaman aralıklarında enzim aktivitelerinin
ölçülmesiyle depolama kararlılıkları belirlenir. Serbest ve immobilize enzimin 5oC
ve 25oC’ de depolama kararlılıklarının yüksek olduğu Şekil 4.10 ve Şekil 4.11’ de
görülmektedir.
Şekil 4.8 ve 4.9 immobilize ve serbest Pektinaz’ ın farklı sıcaklıklardaki
inkübasyon süresi ile kalan aktivite miktarları arasındaki ilişkiyi göstermektedir.
5oC’de 16 saat inkübe edilen enzimin termal kararlılığı yüksek olup inkübasyon
sonunda kalan aktivitesi yaklaşık % 81’dir.16 saat sonunda 25 ve 50oC’de inkübe
edilen serbest Pektinaz enzimi başlangıç aktivitelerinin sırasıyla % 75 ve % 68’ini
korurken, 60oC’de 16 saat sonunda başlangıç aktivitesinin % 19’nu korumaktadır.
İmmobilize Pektinaz’ın termal kararlılığını veren Şekil 4.9 incelendiğinde,
immobilize enzimin termal kararlılığının serbest enzime göre çok farklı olmadığı
gözlenmektedir. İmmobilize Pektinaz’ın 5oC’de 16 saat inkübasyonu sonunda kalan
aktivitesi yaklaşık % 83 iken, 60oC’de 16 saat inkübasyonu sonunda kalan aktivitesi
% 27’dir.

4.BULGULAR VE TARTIŞMA Fadile YENER
40
İmmobilize enzimlerin tekrar tekrar kullanılabilmesı enzimin endüstride
kullanılması açısından önemli bir faktördür. Artan kararlılık immobilize enzimi
serbest enzimden daha avantajlı hale getirmektedir (Lei ve ark., 2007c). Pektinaz’ın
tekrar kullanımını gösteren grafik Şekil 4.12’de verilmektedir. Optimum koşullarda
20 kullanımdan sonra kalan aktivite başlangıç aktivitesinin % 94’üdür.Literatürde
Concanavalin A’ ya bağlı agaroz’a immobilize edilmiş Pektinaz’ın aktivitesini
kaybetmeksizin 3 kez tekrar kullanılabildiği bildirilmektedir (Sardar ve ark., 2005) .
4.2.2. Pektinaz’ın Florisil Desteğe İmmobilizasyonu
Pektinaz enzimi gluteraldehitle aktifleştirilmiş florisil destek üzerine yaklaşık
% 72 bağlanma yüzdesiyle bağlanmıştır. Fakat enzim bağlandıktan sonra aktivitesini
kaybetmiş yani inaktif hale gelmiştir. Enzimin aktivite göstermemesi amino grubu
üzerinden olan bağlanma bölgesinin enzimin aktif bölgesi olduğunu
düşündürmektedir.

5.SONUÇLAR ve ÖNERİLER Fadile YENER
41
5.SONUÇLAR 5.1.Sonuçlar 1. Serbest ve immobilize Pektinaz’ın en yüksek aktivite gösterdikleri pH değerleri
5.0 olarak belirlenmiş olup immobilizasyon işleminin enzimlerin % aktivite- pH
profilini etkilemediği belirlenmiştir.
2.İmmobilize Pektinaz’ın optimum aktivite gösterdiği sıcaklık 45ºC olarak
belirlenmiştir.
3. İmmobilize Pektinaz için Vmax değeri 69,4 µmolgalakturonikasit dk-1 mg protein -1
bulunurken, KM değeri 8,04 olarak bulunmuştur. Katalitik etkinlik olarak verilen
Vmax/KM değeri serbest enzime göre çok daha büyük olduğu belirlenmiştir.
4. İmmobilize Pektinaz için hem oda sıcaklığında hem de 5°C’de belirlenen
depolama kararlılıklarına göre immobilize enzimin 5°C’deki depolama
kararlılığının daha yüksek olduğu saptanmıştır
5. İmmobilize Pektinaz, 20 kullanım sonunda başlangıç aktivitesinin % 94’ünü
korumuştur.
6. İmmobilize Pektinaz, farklı sıcaklıklardaki farklı inkübasyon süreleri sonunda
kalan aktiviteleri incelendiğinde, 5°C’de 16 saat inkübasyon sonunda başlangıç
aktivitesinin % 83’ünü koruduğu belirlenmiştir.
7. Florisil üzerine immobilize edilen Pektinaz’ın çeşitli denemeler sonucunda pektini
hidroliz etmemesini, enzim molekülünün desteğe aktif bölgesinden bağlanmış
olmasına ve bağlanan enzim molekülünün konformasyonunda oluşabilecek
değişikliklere bağlayabiliriz. Bu nedenle florisil Pektinaz immobilizasyonu için
uygun bir destek değildir diyebiliriz.

5.SONUÇLAR ve ÖNERİLER Fadile YENER
42
5.2. Öneriler
1. Kompleks bir enzim olan Pektinaz’ın poligalakturonaz aktivitesi dışındaki
aktiviteleride farklı çalışma koşullarında ölçülüp kıyaslama yapılabilir.
2. Pektinaz için uygun bir yöntem olan tutuklama yöntemi daha farklı
polimerlerle denenebilir.
3. Serbest veya immobilize Pektinaz’ın farklı pH ve derişimlerdeki tampon
çözeltileri içerisindeki depolama kararlılıkları araştırılabilir.
4. Florisil destek kullanılarak yapılan immobilizasyon işleminin ortam koşulları
değiştirilerek(pH gibi) immobilizasyon işlemi tekrarlanabilir.
5. Florisille başarılı olamayan kovalent bağlı immobilizasyonu kıyaslayabilmek
için yine bağlanma ile immobilize olabilecek destekler araştırılıp
uygulanabilirlikleri incelenebilir.
6. Pektinaz enzimi immobilizasyon için daha sık kullanılan polisakkarit destek
ortamlarına değişik ara kollar bağlanarak immobilizasyon yapılabilir.

43
KAYNAKLAR
ACAR, J., GÖKMEN, V., 2004. Meyve ve Sebze İşleme Teknolojisi. Cilt 1-Meyve
ve Sebze Suları Üretimi, 674 s.
AEHLE, W., 2004. Enzyme in Industry. Production and Applications. 484 s.
AKSÖZ, E., AKSÖZ, N., 1985. Pektik Enzimler (derleme).Biyokimya
Dergisi,10:38-51.
ASLAN, Y., TANRISEVEN, A., 2007. Immobilization of Pectinex Ultra SP-L to
Produce Galactooligosaccharides. Journal of Molecular Catalysis, 45:73-77.
BARENSE, R.I., CHELLEGATTI, M.A.S.C., FONSECA, M.J., SAID, S., 2001.
Partıal Purification and Characterization of Exopolygalacturonase II and III
of Penicillum Frequentans. Brazilian Journal of Microbiology, 32:327-330.
BONNIN, E., GOFF, A.L., KORNER, R., VIGOROUX, R., ROEPSTORFF, P.,
THİBAULT, J.-F., 2001. Hydrolysis of Pectins with Different Degrees and
Patterns of Methylation by Endopolygalacturonase of Fusarium moniliforme.
Biochimica et Biophysica acta, 1596:83-94.
BOTELLA, C., DIAZ, A., ORY, I., WEBB, C., BLANDINO, A., 2006. Xylanase
and Pectinase Production by Aspergillus awamori on Grape Pomace in Solid
State Fermentation. Process Biochemistry, 42:98-101.
BUSTO, M.D., TRAMONTIN, K.E.G., ORTEGA, N., MATEOS, M.P., 2006.
Preparation and Properties of an Immobilized Pectinlyase for the Treatment
of Fruit Juices. Bioresoure Technology, 97:1477-1483.
CHELLEGATTI, M.A., SAID, S., 2002. Purification and partial characterization of
exopolygalacturonase I from Penicillum frequentans. Microbiological
Research, 157:19-24.
CESTINO, S.M.C., FREITAS, S.M., MEDRANO, F.J., SOUSA, M.V., FILHO,
E.X.F., 2006.Purification and Characterization of a Novel Pectinase from
Acrophialophora nainiana with Emphasis on Its Physicochemical Properties.
Journal of Biotechnology, 123:33-42.

44
COSTA, S.A., TZANOV, T., PAAR, A., GUDELJ, M., GUBITZ, G.M., PAULO,
A.C., 2001. Immobilization of Bacillus SF on Alumina for the Treatment of
Textile Bleacing Effluents. Enzyme and Microbial Technology, 28:815-819.
DEBING, J., PEIJUN, L., STAGNITTI, F., XIANZHE, X., LI, L., 2006. Pectinase
Production by Solid Fermentation form Aspergillus niger by a New
Prescription Experiment. Ecotoxicology and Environmental Safety, 64:244-
250.
EKŞİ, A., 1988. Meyve Suyu Durultma Tekniği, Gıda Teknolojisi Derneği Yayın
no:9, 127s.
LEI, Z., BI, S., 2007a. The silica-coated chitosan particle from a layer-by-layer
approach for pectinase immobilization. Enzyme and Microbial Technology,
40:1442-1447.
LEI, Z., BI, S., 2007b. Preparation and Properties of Immobilized Pectinase onto the
Amphiphilic PS-b-PAA Diblock Copolymers. Journal of Biotechonology,
128:112-119.
LEI, Z., BI, S., HU, B., YANG, H., 2007c. Combined Magnetic and Chemical
Covalent Immobilization of Pectinase on Composites Membranes Improves
Stability and Activity. Food Chemistry,(baskıda).
LI, T., W, N., LI, S., ZHAO, Q., GUO, M., ZHANG, C., 2007. Optimization of
Covalent Immobilization of Pectinase on Sodium Alginate Support.
Biotechnol Lett, 29:1413-1416.
LOWRY, O.H., ROSENBROUGH, N.J., FARR, A.L., RANDALL, R.J., 1951.
Protein Measurement with Folin Phenol Reagent.Journal of Biological
Chemistry, 193:265-275.
MILLER, G.L., 1959. Use of Dinitrosalicyclic Acid Reagent for Determination of
Reducing Sugar. Analytical Chemistry, 31:426-428.
MONDAL, K., MEHTA, P., GUPTA, M.N., 2004. Affinity Precipitation of
Aspergillus niger Pectinase by Microwave-treated Alginate. Protein
Expression and Purification, 33:104-109.

45
ORTEGA, N., DIEGO, S., MATEOS, M.P., BUSTO, M.D., 2004. Kinetic properties
and thermal behaviour of polygalacturonase used in fruit juice clarification.
Food Chemistry 88:209-217.
PATIL, S.R., DAYANAND, A., 2006. Exploration of Regional Agrowastes for the
Production of Pectinase by Aspergillus niger. Food Technol. Biotechnol.
44.289-292.
PHUTELA, U., DHUNA, V., SANDHU, S., CHADHA, B.S., 2005. Pectinase and
Polygalacturonase Production by a Thermophilic Aspergillus fumigatus
Isolated from Decomposting Orange Peels. Brazilian Journal of
Microbiology, 36:63-69.
ROY, I., SARDAR, M., GUPTA, M.N., 2003. Evaluation of Smart Bioconjugate of
Pectinase for Chitin Hydrolysis. Biochemical Engineering Journal, 16:329-
335.
ROY, I., SARDAR, M., GUPTA, M.N., 2003. Hydrolysis of Chitin by Pectinex.
Enzyme and Microbial Technology, 32:582-588.
SARDAR, M., GUPTA, M.N., 1998. Alginate Beads as an Affinity Material for
Alpha Amylases. Bioseparation, 7:159-165.
SARDAR, M., GUPTA, M.N., 2005. Immobilization of Tomato Pectinase on Con A-
Seralose 4B by Bioaffinity Latering. Enzyme and Microbial Technology,
37:355-359.
SILVA, D., TOKUIOSHI, K., MARTINS, E.S., SILVA, R.D., GOMES, E., 2005.
Production of Pectinase by Solid-State Fermentation with Penicillium
Viridicatum RFC3. Process Biochemistry, 40:2885-2889.
SMIDSROD, O., SKJAK, B.G., 1990. Alginate as Immobilization Matrix for Cells.
Trends Biotechnol., 8:71-78.
SOARES, M.M.C.N., SILVA, R., GOMES, E., 1999. Screening of Bacterial Strains
for Pectinolytic Activity: Characterization of the Polygalacturonase
Produced by Bacillus sp. Revista de Microbiologia, 30:299-303.
SUMNER, J.B., 1921.Dinitrosalicyclic acid:A reagent for the estimation of sugar in
normal and diabetic urine. J.Biol.Chem. 47:5-9.

46
SUNNOTEL, O., NIGAM, P., 2002. Pectinolytic Activity of Bacteria Isolated from
Soil Two Fungal Strains During Submerged Fermentation. World Journal of
Microbiology and Biotechnology, 18:835-839.
TELEFONCU, A., 1997. Enzimoloji.Yüksek Lisans Yazokulu. 21-27 Eylül 1997.
Kuşadası, Aydın, Türkiye.446 s.
TUKEL, S.S., ALPTEKIN, O., 2004. Immobilization Kinetics of Catalase onto
Magnesium Silicate. Process Biochemistry, 39:2149-2155.
WANG, G., MICHAILIDES, T.J., BOSTOCK, R.M., 1997. Improved Detection of
Polygalacturonase Activity Due to Mucor piriformis with a Modified
Dinitrosalicylic Acid Reagent. Techniques, 87:161-163.
WEETHALL, H.H., 1976. Covalent Coupling Methods for Inorganic Support
Metarials. Methods of Enzmology, 44:134-148.
Willants, W.G., Mc-Cartney, L., Mackie, W., Knox, J.P., 2001. Pectin: cell biology
and structural prospects for functional analysis. Plant Molecular Biology, 47:
1-27.

47
ÖZGEÇMİŞ
1982 yılında Gaziantep’de doğdum.İlk ve orta öğrenimimi Adana’da
tamamladıktan sonra 2000 yılında Çukurova Üniversitesi Fen-Edebiyat Fakültesi
Kimya Bölümünde üniversite eğitimime başladım ve 2004 yılında mezun olduktan
sonra aynı yıl Çukurova Üniversitesi Fen-Edebiyat Fakültesi Kimya Bölümünde
Yüksek Lisans eğitimime başladım.