this paper is available on line at ... · proteomic analysis of mitotic rna polymerase ii reveals...
TRANSCRIPT

Proteomic Analysis of Mitotic RNA PolymeraseII Reveals Novel Interactors and AssociationWith Proteins Dysfunctional in Disease*□S
Andre Moller‡��, Sheila Q. Xie‡, Fabian Hosp§, Benjamin Lang¶, Hemali P. Phatnani�§§,Sonya James‡¶¶, Francisco Ramirez**, Gayle B. Collin‡‡, Jurgen K. Naggert‡‡,M. Madan Babu¶, Arno L. Greenleaf�, Matthias Selbach§, and Ana Pombo‡��
RNA polymerase II (RNAPII) transcribes protein-codinggenes in eukaryotes and interacts with factors involved inchromatin remodeling, transcriptional activation, elonga-tion, and RNA processing. Here, we present the isolationof native RNAPII complexes using mild extraction condi-tions and immunoaffinity purification. RNAPII complexeswere extracted from mitotic cells, where they exist disso-ciated from chromatin. The proteomic content of nativecomplexes in total and size-fractionated extracts was de-termined using highly sensitive LC-MS/MS. Protein asso-ciations with RNAPII were validated by high-resolutionimmunolocalization experiments in both mitotic cells andin interphase nuclei. Functional assays of transcriptionalactivity were performed after siRNA-mediated knock-down. We identify >400 RNAPII associated proteins inmitosis, among these previously uncharacterized proteinsfor which we show roles in transcriptional elongation. Wealso identify, as novel functional RNAPII interactors, twoproteins involved in human disease, ALMS1 and TFG, em-phasizing the importance of gene regulation for normaldevelopment and physiology. Molecular & Cellular Pro-teomics 11: 10.1074/mcp.M111.011767, 1–16, 2012.
The transcription of protein coding genes is a highly regu-lated process that depends on the enzymatic activity of RNApolymerase II (RNAPII)1, a complex comprised of 12 subunits.
The ability of cells to transcribe specific subsets of mRNAsrelies on activities influencing the accessibility of gene pro-moters within their chromatin context, the formation of pre-initiation complexes, the initiation of RNA synthesis, and thetransition to states of productive elongation (1). Central tothese activities is the RNAPII complex itself, which interactswith chromatin regulators, transcription initiation factors, andwith complexes required for processive elongation (2). Fur-thermore, chromatin-bound RNAPII recruits enzymatic activ-ities involved in the cotranscriptional processing of nascentRNA, such as capping, splicing, and 3� processing of primarytranscripts (3).
Recruitment depends on specific phosphorylation of thecarboxy-terminal domain (CTD) of the largest subunit of RNA-PII, RPB1, consisting of a highly repetitive heptapeptide se-quence (Y1S2P3T4S5P6S7; 52 repeats in mammals). S2, S5,and S7 residues become phosphorylated during transcription:S5 and S7 during the stage of transcriptional initiation, and S2during transcriptional elongation (4).
The regulation of transcription and its downstream pro-cesses is of central importance in many human diseases (5,6), and many proteins involved in these processes are aber-rantly expressed or dysfunctional in cancer (7, 8). Compre-hensive knowledge of the mammalian RNAPII protein interac-tion landscape may therefore further our understanding ofdisease genesis in humans, and ultimately help prevent itsprogression.
In this study, we have purified intrinsically soluble RNAPIIfrom mitotic cells, a strategy based on observations thatRNAPII dissociates from chromatin as the cell enters mitosis((9); Fig. 1A), and that functionally relevant components, suchas splicing factors, can remain associated with RNAPII duringmitosis (10). We speculated that other cotranscriptional inter-actions with RNAPII may also be transcription-independentand more stable than anticipated, and that identification of theRNAPII proteome in native conditions during mitosis would
From the ‡MRC Clinical Sciences Centre, Imperial College Schoolof Medicine, Hammersmith Hospital Campus, London W12 0NN,United Kingdom; §Max-Delbruck Center for Molecular Medicine,13092 Berlin, Germany; ¶MRC Laboratory of Molecular Biology,Cambridge CB2 0QH, United Kingdom; �Department of Biochemistry,Duke University, Medical Center, Durham, North Carolina 27710;**South Ruislip, Middlesex, UK; ‡‡The Jackson Laboratory, Bar Har-bor, Maine 04609, USA
Received June 6, 2011, and in revised form, November 19, 2011Published, MCP Papers in Press, December 22, 2011, DOI
10.1074/mcp.M111.0117671 The abbreviations used are: RNAPII, RNA Polymerase II; ABA,
antibody blocking assay; ANOVA, analysis of variance; AP, alkalinephosphatase; CTD, carboxy-terminal domain; GO, gene ontology;GTF, general transcription factor; IP, immunoprecipitation; MIG, mi-totic interchromatin granules; nPAI, normalized PAI; PAI, peptideabundance index; PBMCs, PHA-activated peripheral blood mononu-
clear cells; PFA, paraformaldehyde; PHA, phytohaemagglutinin; qRT-PCR, quantitative real-time PCR; RNAi, RNA interference; RNAPIIA,hypophosphorylated RNAPII; RNAPIIO, hyperphosphorylated RNAPII;SD, standard deviation.
Research© 2012 by The American Society for Biochemistry and Molecular Biology, Inc.This paper is available on line at http://www.mcponline.org
Molecular & Cellular Proteomics 11.6 10.1074/mcp.M111.011767–1

allow the identification of novel RNAPII-associated proteins.We combined low stringency lysis and immunoprecipitationusing highly specific elution conditions with mass spectrom-etry and appropriate control purifications. We show that mi-totic RNAPII is present in complexes with sizes expected forthe canonical 12 subunit complex (�700 KDa) and in muchlarger (�2 MDa) complexes, enriched in S2 and S5 phos-phorylation. Our mass-spectrometric analyses of complexesfrom total and size-fractionated lysates revealed �400 RNA-PII associated proteins, including novel and known RNAPIIinteractors. We have validated various interactions by in situexperiments using fluorescence microscopy, measuring tran-scriptional activity after RNAi and colocalization betweenRNAPII and candidate interactors. The results allow us to infera functional importance for several of the RNAPII interactorsin interphase, including proteins that have been implicated inhuman disease.
EXPERIMENTAL PROCEDURES
Mitotic Cell Collection, Lysis, Gel Filtration and Immunoprecipita-tion—Mitotic cells were collected and frozen in conditions that pre-serve viability. Mitotic cell lysates cleared of chromosomes and largerorganelles were treated with calf intestinal phosphatase and TurboDNase, unless otherwise stated. Fractionation of mitotic lysates wasperformed on a Superose 6 10/300 GL column (GE Healthcare).
For immunoprecipitation of RNAPII associated proteins, total ly-sate, fraction 2 or fraction 8 samples were incubated with antibody8WG16 (mouse ascites, Covance, Princeton, NJ) or control mouseIgG2a antibody (monoclonal cell culture supernatant; DakoCytoma-tion). Proteins were eluted by resuspension in elution buffer contain-ing a CTD repeat peptide (ab12795; Abcam, Cambridge, MA) andethanol precipitated. Full details are available in Supplementary Text.
Mass Spectrometry—Immunoprecipitated protein pellets were sol-ubilized in 6 M urea/2 M thiourea/10 mM HEPES (pH 8.0) buffer,reduced with 1 mM dithiothreitol, alkylated with 5.5 mM iodoacet-amide and digested using unmodified lysyl endopeptidase (LysC,1:100 w/w, Wako, Osaka, Japan) and subsequently sequencing grademodified trypsin (1:100 w/w, Promega, Mannheim, Germany) in 50mM ammoniumbicarbonate buffer. Peptides were purified using stopand go extraction (STAGE) tips (11). Liquid chromatography-tandemMS (LC-MS/MS) analysis was performed online as previously de-scribed (12). In brief, peptide mixtures were separated by reversedphase chromatography using the Eksigent NanoLC - 1D Plus system(Eksigent, Dublin, CA) on in-house manufactured 10 cm fritless silicamicrocolumns with an inner diameter of 75 �m. Columns werepacked with ReproSil-Pur C18-AQ 3 �m resin (Dr. Maisch GmbH,Ammerbuch, Germany) (13). Separation was performed using a 10–60% acetonitrile gradient (155 min) with 0.5% acetic acid at a flowrate of 500 nl/min. Eluting peptides were directly ionized by electro-spray ionization and transferred into the orifice of a LTQ-Orbitrapclassic hybrid mass spectrometer (Thermo Fisher, Waltham, MA).Mass spectrometry was performed in the data dependent mode withone full scan in the Orbitrap (m/z � 300–1700; r � 60,000; targetvalue � 1 � 106). The five most intense ions with a charge stategreater than one were selected (target value 5000; monoisotopicprecursor selection enabled) and fragmented in the LTQ using colli-sion-induced dissociation (35% normalized collision energy, wide-band activation enabled). Dynamic exclusion for selected precursorions was set to 60 s. The MaxQuant software package (version1.0.12.5) was used to identify proteins (14, 15) with enabled polymerdetection and top six MS/MS peaks selection per 100 Da. Peak lists
were searched on a MASCOT search engine (version 2.2, Matrix-Science, Boston, MA) against an in-house curated concatenatedtarget-decoy database (16) of forward and reversed proteins in theInternational Protein Index (IPI) human protein database (version 3.43,72,346 entries), supplemented with common contaminants. Carbam-idomethylation of cysteine was selected as fixed modification, oxida-tion of methionine and acetylation of the protein N terminus wereused as variable modifications. Trypsin/p � DP was selected asprotease (full specificity) with a maximum of three missed cleavages.Mass tolerance for fragment ions was set to 0.5 Da. The original masstolerance of precursor ions was 7 p.p.m., but narrowed down furtherto the individual mass tolerances calculated by MaxQuant. A mini-mum of six amino acids per identified peptide and at least onepeptide per protein group were required. Only unique peptidesor peptides assigned to the protein group with the highest number ofpeptides are reported. False discovery rate was set to 1% at both thepeptide and protein level. The data associated with this study may bedownloaded from Proteome Commons (http://proteomecommons.org/) Tranche using the following hash: MfbhtAHFbsvG3IcVs8hJp-IYNFUH1uw9cesgvnoFMh08mvFtaPMaZ9GK0QjXgnnJU6Kcb7Sjfz-LA6lRhFN9r5n947550AAAAAAAAHSg��.
Gene Ontology (GO) Analysis—Over- and under-representation ofGO terms was analyzed using GOstat (17) in conjunction with thegoa_human annotation data set and Benjamini-Hochberg correctionfor multiple testing.
RNAi and Br-UTP Incorporation Assay—HeLa cells were cotrans-fected with two different siRNAs against each gene of interest or withcontrol siRNA using the Lipofectamine 2000 reagent (Invitrogen,Carlsbad, CA). At 72 h post-transfection, in situ transcription run-onswere performed in the presence of Br-UTP and labeled Br-RNAindirectly immunolabeled as described in (18). Confocal microscopyimages were analyzed using ImageJ software. SiRNA results wereanalyzed by ANOVA (using SAS, version 9.1), testing for all maineffects (siRNA treatment, day of transfection and Br-UTP labeling set)of which only the siRNA treatment and day of transfection weresignificant (both p � 0.0001). The ANOVA estimates were used tocompare siRNA effects relative to control siRNA using Student’s ttest. The p values represented in Fig. 4 are adjusted for multipletesting using the method of false discovery rate by Benjamini andHochberg. Full details are available in supplementary Text.
RNAi and RT-qPCR—Cells were treated with siRNAs as above,before isolation of total RNA using TRIzol reagent (Invitrogen). qPCRswere performed using the SensiMax NoRef kit (Quantace) and aCFX96 Real-Time System (BioRad). Transcript levels were normalizedto beta-actin. Full details are available in supplementary Text.
Protein Immunolocalization And Antibody Blocking Assay (ABA)Using Tokuyasu Cryosections—HeLa cells, human PHA-activated pe-ripheral blood mononuclear cells (PBMCs), and pancreatic and hypo-thalamic mouse tissues were used for ABA. Human female PBMCswere purified using the Leuco-Sep kit (human; Harlan Sera-Lab,Loughborough, United Kingdom) and activated with phytohemagglu-tinin (PHA; Sigma) for 72 h. PBMCs and cultured HeLa cells werefixed in 4% and then 8% depolymerised paraformaldehyde (PFA;EM-grade) in 250 mM HEPES (pH 7.6; 10 min and 2 h, respectively).
Mice used in the study were maintained in the Research Facility atThe Jackson Laboratory. Experimental procedures were carried outunder Institutional Animal Care and Use Committee approval. Briefly,pancreatic and hypothalamic tissues were collected following intrac-ardiac perfusion with 4% PFA/HEPES solution. Tissues were dis-sected into 1.5 mm pieces in cold 4% PFA/HEPES solution and fixedfor an additional 2 h 8% PFA/HEPES. Fixed cell pellets and dissectedtissues were embedded in sucrose solution and frozen in liquid nitro-gen. Cryosections (150 nm thick) on coverslips were processed forimmunofluorescence labeling or ABA, as previously described (18,
Proteomic Analysis of Mitotic RNA Polymerase II
10.1074/mcp.M111.011767–2 Molecular & Cellular Proteomics 11.6

19). For ABA, cryosections were pretreated with alkaline-phospha-tase to ensure maximized binding of 8WG16 to detected total RPB1,and processed as previously described (20).
Confocal microscopy images from cryosections were collectedusing settings calibrated on the negative control (no antibody) sam-ples without saturation of the intensity signal. Nuclear fluorescenceintensities were determined in Photoshop. Statistical significance wastested by ANOVA (using SAS, version 9.1). ANOVA estimates wereused to compare the blocking effects of antibodies to proteins ofinterest relative to control blocking antibodies using Student’s t test.The p values represented in Figs. 3 and 4 are adjusted for multipletesting using the method of false discovery rate by Benjamini andHochberg. Full experimental details and primary antibodies used areavailable in Supplementary Text.
RESULTS
RNAPII is Present in �2 MDa Complexes During Mito-sis—To enhance our understanding of the protein interactionlandscape of RNAPII, we purified RNAPII complexes undermild conditions from mitotic cells, where RNAPII is mostlyexcluded from the condensed chromosomes and distributedthroughout the mitotic cytoplasm (Fig. 1A).
Mitotic cells were lysed in the presence of the natural,nonionic surfactant saponin, which preserves RNAPII functionduring interphase (18), and using a near physiological saltconcentration of 150 mM NaCl. The resulting lysate wastreated with DNase to remove residual DNA, which mightotherwise act as a scaffold for indirect protein-protein inter-actions (Fig. 1B). The lack of chromatin contamination wasconfirmed by testing for the presence of histone H2B in themitotic lysate (Fig. 1C). Although H2B is detectable in wholecell lysates of unsynchronized cells (95% interphase), no H2Bis detectable in the mitotic lysate clarified of chromosomes.
To investigate the size of RNAPII complexes in mitosis, weperformed size fractionation of the mitotic lysate followed byWestern blotting using various antibodies directed againstsubunits of RNAPII (RPB1, RPB2, and RPB3) and CTD phos-pho-epitopes (S2p and S5p; Fig. 1D). We identify RNAPIIenrichment in two parts of the elution spectrum: at �700 KDa(around fraction 8), a size expected for the canonical 12subunit RNAPII complex, and at �2 MDa (fraction 2). RNAPIIcomplexes in the 2–4 MDa range have previously been re-ported in whole cell and nuclear extracts from asynchronouspopulations of mammalian cells (21–23), but not from mitoticcells, and were shown to contain general transcription factors(GTFs) and mediator subunits as well as DNA repair proteins(21) or transcription elongation factors (22).
Interestingly, using antibodies directed against phosphory-lated RPB1, we detect an enrichment of hyperphosphorylatedRPB1 in the larger RNAPII containing complexes, suggestingthat RPB1 phosphorylation in mitotic cells may be associatedwith the formation of higher-order complexes between RNA-PII and additional factors. This mirrors current models for therole of RNAPII phosphorylation during the transcription cyclein recruiting specific chromatin remodelers and the RNA pro-cessing machinery (3, 4).
Scheme for the Purification of RNAPII Complexes FromMitotic Extracts—To purify RNAPII-containing protein com-plexes from total and size-fractionated extracts, we devisedan immunoprecipitation strategy (Fig. 1E) based on the bind-ing of the antibody 8WG16 to the CTD of RPB1 (24). This isfollowed by elution using a CTD peptide consisting of tworepeats of the CTD heptad sequence that effectively com-petes for binding to the 8WG16 paratope and specificallyelutes RNAPII complexes from 8WG16-bound agarose beadspost-immunoprecipitation (Fig. 1F). The highly repetitive na-ture of the CTD allows for a high-affinity interaction with8WG16. Because of the known propensity of 8WG16 to pref-erentially bind hypophosphorylated RPB1 (19, 25, 26), weinitially introduced an alkaline phosphatase (AP) treatmentinto our purification strategy ((19), supplementary Fig. S1).Subsequently, we investigated RNAPII interactors in the ab-sence of AP treatment (supplementary Fig. S2). As an addi-tional specificity control, we performed parallel immunopre-cipitations with a matched isotype control antibody. Sixdifferent samples were initially analyzed by mass-spectrom-etry from AP-treated mitotic extracts: the pre-fractionation(total) lysate, and the gel filtration fractions containing the �2MDa (fraction 2) or the �700 kDa (fraction 8) RNAPII com-plexes, together with their three matched isotype controls.
Mass-Spectrometric Identification of RNAPII-associatedProteins—To investigate RNAPII-associated proteins in mito-sis, we first purified the total mitotic lysate directly, withoutsize fractionation. The eluates from 8WG16 and control IPswere subsequently trypsinized and analyzed by LC-MS/MS. Atotal of 511 proteins were identified in the 8WG16 sample, ofwhich 387 proteins were represented by at least two uniquepeptides (Table I). Half of these proteins (n � 178) werespecific to the 8WG16 IP (i.e. not identified in the isotypecontrol IP). A second category of “more abundant” proteins(n � 117), represented with more peptides in the 8WG16 IPthan in the isotype control (�2 peptides difference) were alsoconsidered, all of which also exhibit higher mean peptideintensities (2- to 297-fold; average 25-fold; S.D. 41-fold; sup-plementary Table S1). In total, we identified 295 RNAPII-associated proteins in the mitotic lysate, as either “specific”(i.e. �2 peptides in the 8WG16 sample and no identification inthe control) or “more abundant” (�2 peptides difference).
Among the RNAPII-associated proteins in mitosis, we findall twelve subunits of RNAPII and a number of proteins knownto directly interact with RNAPII (e.g. GDOWN1 (27), SAFB1(28), RMP (29), SFPQ, and NONO (30). Previous proteomicstudies of RNAPII-associated proteins (31–33) reported onlyeight (31) or five (32) RNAPII subunits, highlighting the value ofmilder purification conditions to achieve a more completecharacterization of RNAPII complexes.
To further assess the quality of the purification, we com-pared the abundance of RNAPII-associated proteins relativeto RNAPII subunits. As a rough estimate of protein abundancewe used the ratio of observed unique peptides to theoretically
Proteomic Analysis of Mitotic RNA Polymerase II
Molecular & Cellular Proteomics 11.6 10.1074/mcp.M111.011767–3

observable unique peptides, similar to the protein abundanceindex (PAI) (34). According to this calculation, the top 21%most abundant proteins lie within the PAI range of the RNAPIIsubunits (0.6–1.2) and a further 18% have a PAI 0.3–0.6 (atleast half the PAI of the least abundant RNAPII subunit;POLR2H, PAI � 0.6). Several proteins of unknown functionhave PAIs in range of 0.3–1.2 (TFG, MRT4, ZNF326,
FAM103A1), whereas the least abundant, known RNAPII in-teractor identified by our approach has a PAI of 0.08 (SAFB1;(28)), suggesting that several of the RNAPII-associated pro-teins identified are likely to be bona fide interactors.
To identify proteins specific to larger (�2 MDa) and smaller(�700 kDa) RNAPII complexes, we extended the proteomicanalysis to samples from fractions 2 and 8 obtained after gel
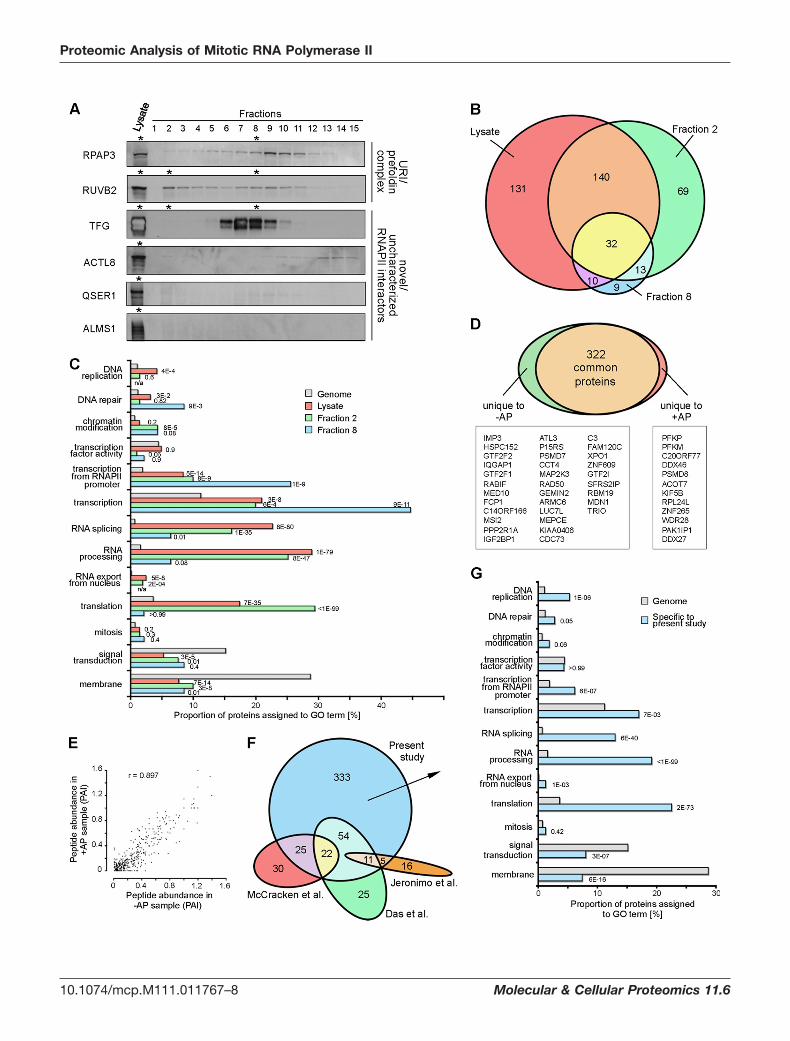
FIG. 1. Isolation of native RNAPII complexes from mitotic cells. A, Indirect immunofluorescence of RNAPII (pseudocolored green) inultrathin Tokuyasu cryosections shows nucleoplasmic localization during interphase and dissociation from chromatin in mitosis. Nucleic acidswere counterstained with TOTO-3 (red). Most RNAPII is present throughout the mitotic cytoplasm; in �45% of mitotic cell profiles (n � 66),1–3 larger RNAPII aggregates (MIGs; (59)) are also present. Scale bar: 6 �m. B, Scheme for the extraction of RNAPII from mitotic cells inphysiological conditions. C, Western blotting of unsynchronized and mitotic cell lysates confirms separation from chromatin, as histone H2Bis not detected in the mitotic lysate. Loading control was ribosomal subunit S6. D, Western blots of gel filtration fractions reveal presence ofmitotic RNAPII in a range of differently sized supercomplexes, with enrichments at �700 kDa (size expected for core RNAPII) and at �2 MDa.Larger complexes show hyperphosphorylation of RPB1 (RNAPIIO) on both S5 (antibody 4H8) and S2 (antibody H5) residues of the CTD,whereas the �700 kDa RNAPII is depleted of phosphorylation. E, Schematic representation of the experimental workflow leading to themass-spectrometric characterization of four different RNAPII populations, investigated using an RNAPII-specific antibody (8WG16), incomparison with an isotype control antibody. See supplementary Fig. S1 for evidence that alkaline phosphatase treatment of mitotic extractsremoves CTD phosphorylation. F, Western blot against subunit RPB3, after immunoprecipitation with 8WG16 antibodies to RPB1, showsefficient elution of RNAPII complexes using a peptide containing two repeats of the CTD consensus sequence.
Proteomic Analysis of Mitotic RNA Polymerase II
10.1074/mcp.M111.011767–4 Molecular & Cellular Proteomics 11.6

filtration of mitotic lysates and immunoprecipitated with8WG16 or a matched isotype antibody (Fig. 1E; Table I). Thisallowed us to identify 131 and 38 “specific” (�2 peptides) or68 and 10 “more-abundant” RNAPII associated proteins infractions 2 and 8, respectively. Within this group of specific ormore-abundant proteins, 112 out of 199 (56%; fraction 2) and27 out of 48 (56%; fraction 8) have PAI values that are �50%of the PAI of the least abundant RNAPII subunit in the respec-tive fraction.
Comparing the lists of specific or more abundant proteins,generated in the lysate, fraction 2 and fraction 8 samples,reveals several protein complexes which are present in thecomplete lysate (U1, U5, SMN, core snRNP) but absent inboth fractions 2 and 8 (Table II), implying that these proteincomplexes co-purify with RNAPII in intermediate fractions orare lost during gel filtration. (A complete representation ofproteins shared by, or exclusive to, specific fractions and/orthe lysate is listed in supplementary Table S2.) In addition,some complexes are best identified in the full lysate, such asthe URI/prefoldin complex, involved in nutrient sensing andTOR signaling (35), and recently implicated in the nuclearimport of RNAPII complexes (36), for which we identify allreported subunits (37, 38). In fraction 2, only subunits RUVB1and RUVB2 are present; Western blot analysis confirmed thedifferential distribution of the URI/prefoldin subunits RPAP3,peaking at fraction 9, and RUVB2, which has a profile similarto the RNAPII subunits (Fig. 2A; see also Fig. 1D). AlthoughRUVB1 and RUVB2 are known constituents of various com-plexes, including SRCAP, hINO80, and TRRAP/TIP60 (37),other subunits of these complexes were not identified. Thissuggests that RUVB1 and RUVB2 are part of a further, as yetunidentified complex, that is specifically associated with thehigher-molecular weight RNAPII complex, or that they directlyinteract with RNAPII itself.
Several proteins identified in fraction 2 were not detected infraction 8, suggesting that their association occurs predomi-nantly in the context of protein complexes of higher molecularweight. There are also several proteins and complexes thatwere not identified in the complete lysate but were identifiedin fraction 2 (mediator, integrator, SWI/SNF, NuRD, amino-
acyl-tRNA synthetase multienzyme complex) or fraction 8(the deubiquitinating enzyme USP9X, several T-complexsubunits). The detection of additional RNAPII-associatedproteins and protein complexes after size fractionation in-dicates that enrichment of specific RNAPII complexes isrequired for a subset of interactors to achieve efficientco-immunoprecipitation.
To investigate the differential association of proteins withRNAPII complexes in total and size-fractionated lysates, weanalyzed the presence of four RNAPII-associated proteins byWestern blotting after size fractionation (TFG, ACTL8, QSER1,ALMS1; Fig. 2A). The four proteins show distributions that arein agreement with their identification in the mass-spectromet-ric data of specific samples (Table II). TFG is present in thetotal lysate and fraction 8, but absent in the larger complexesof fraction 2. ACTL8 is detected in the total lysate but is onlyweakly detectable in fraction 8 and mainly present in fractionsthat would contain single RNAPII subunits. QSER1 andALMS1 are detected in the total lysate but not in the gelfiltration fractions analyzed, possibly because of loss duringthe size fractionation process. These results exemplify theimportance of using complementary strategies to identifyRNAPII-associated proteins, which may vary in their stabilityor abundance.
We next compared the proteins identified as specific ormore abundant in the total lysate and the size fractions. Here,we included specific proteins identified with only one peptide,if their presence in multiple 8WG16 IP samples supportedtheir correct identification. We found 32 proteins to be presentin all three samples (Fig. 2B and Table III), including eleven ofthe twelve canonical RNAPII subunits. Furthermore, 172 pro-teins were common between the total lysate and fraction 2,and 42 proteins between the total lysate and fraction 8.
Mitotic RNAPII Complexes are Enriched in Proteins withKnown Functions in Transcription, RNA Processing andTranslation—To explore the functions of the proteins associ-ated with RNAPII in mitosis, we analyzed the Gene Ontology(GO) terms of the specific and more-abundant proteins iden-tified in the total lysate, fraction 2, and fraction 8 (Fig. 2C).Despite RNAPII being transcriptionally silent and dissociated
TABLE INumber of RNAPII-associated proteins in mitotic extracts. Overview of the number of identified proteins, including total number of proteinsidentified, total number of proteins identified with two or more peptides, number of proteins identified as specific to the RNAPII immunopre-cipitation (i.e. absent from the control immunoprecipitation), number of proteins specific to the RNAPII immunoprecipitation identified with twoor more peptides, and number of proteins identified as probably more abundant in the RNAPII specific immunoprecipitation than in the controlimmunoprecipitation (peptide difference � 2). See supplementary Table S1 for complete lists of proteins identified in the RNAPII specific and
control precipitations
�AP -AP
Lysate Fraction 2 Fraction 8 Lysate
total proteins 511 363 144 775proteins �2 peptides 387 242 90 5238WG16-specific (total) 269 228 65 2468WG16-specific (�2 peptides) 178 131 38 116more abundant in 8WG16-IP relative to control-IP 117 68 10 143
Proteomic Analysis of Mitotic RNA Polymerase II
Molecular & Cellular Proteomics 11.6 10.1074/mcp.M111.011767–5
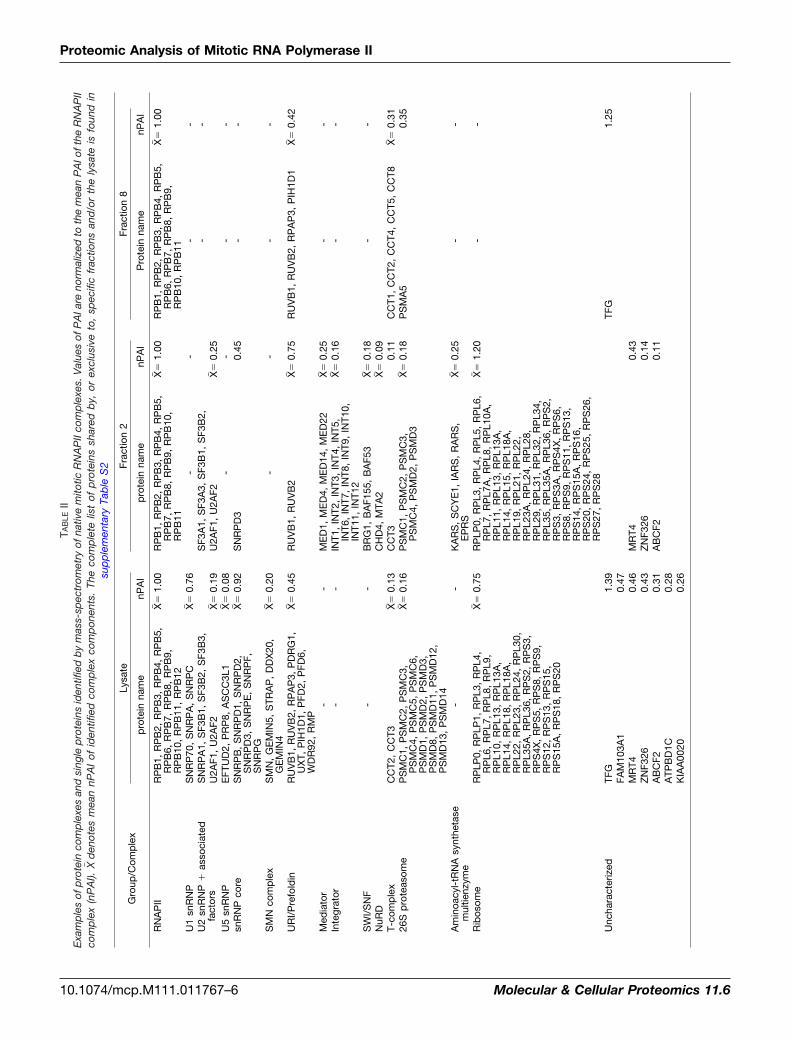
TAB
LEII
Exa
mp
les
ofp
rote
inco
mp
lexe
san
dsi
ngle
pro
tein
sid
entif
ied
by
mas
s-sp
ectr
omet
ryof
nativ
em
itotic
RN
AP
IIco
mp
lexe
s.V
alue
sof
PA
Iare
norm
aliz
edto
the
mea
nP
AIo
fthe
RN
AP
IIco
mp
lex
(nP
AI).
X�d
enot
esm
ean
nPA
Iof
iden
tifie
dco
mp
lex
com
pon
ents
.Th
eco
mp
lete
list
ofp
rote
ins
shar
edb
y,or
excl
usiv
eto
,sp
ecifi
cfr
actio
nsan
d/o
rth
ely
sate
isfo
und
insu
pp
lem
enta
ryTa
ble
S2
Gro
up/C
omp
lex
Lysa
teFr
actio
n2
Frac
tion
8
pro
tein
nam
enP
AI
pro
tein
nam
enP
AI
Pro
tein
nam
enP
AI
RN
AP
IIR
PB
1,R
PB
2,R
PB
3,R
PB
4,R
PB
5,R
PB
6,R
PB
7,R
PB
8,R
PB
9,R
PB
10,
RP
B11
,R
PB
12
X��
1.00
RP
B1,
RP
B2,
RP
B3,
RP
B4,
RP
B5,
RP
B7,
RP
B8,
RP
B9,
RP
B10
,R
PB
11
X��
1.00
RP
B1,
RP
B2,
RP
B3,
RP
B4,
RP
B5,
RP
B6,
RP
B7,
RP
B8,
RP
B9,
RP
B10
,R
PB
11
X��
1.00
U1
snR
NP
SN
RP
70,
SN
RP
A,
SN
RP
CX�
�0.
76-
--
-U
2sn
RN
P�
asso
ciat
edfa
ctor
sS
NR
PA
1,S
F3B
1,S
F3B
2,S
F3B
3,S
F3A
1,S
F3A
3,S
F3B
1,S
F3B
2,-
-U
2AF1
,U
2AF2
X��
0.19
U2A
F1,
U2A
F2X�
�0.
25U
5sn
RN
PE
FTU
D2,
PR
P8,
AS
CC
3L1
X��
0.08
--
--
snR
NP
core
SN
RP
B,
SN
RP
D1,
SN
RP
D2,
SN
RP
D3,
SN
RP
E,
SN
RP
F,S
NR
PG
X��
0.92
SN
RP
D3
0.45
--
SM
Nco
mp
lex
SM
N,
GE
MIN
5,S
TRA
P,
DD
X20
,G
EM
IN4
X��
0.20
--
--
UR
I/P
refo
ldin
RU
VB
1,R
UV
B2,
RP
AP
3,P
DR
G1,
UX
T,P
IH1D
1,P
FD2,
PFD
6,W
DR
92,
RM
P
X��
0.45
RU
VB
1,R
UV
B2
X��
0.75
RU
VB
1,R
UV
B2,
RP
AP
3,P
IH1D
1X�
�0.
42
Med
iato
r-
-M
ED
1,M
ED
4,M
ED
14,
ME
D22
X��
0.25
--
Inte
grat
or-
-IN
T1,
INT2
,IN
T3,
INT4
,IN
T5,
INT6
,IN
T7,
INT8
,IN
T9,
INT1
0,IN
T11,
INT1
2
X��
0.16
--
SW
I/S
NF
--
BR
G1,
BA
F155
,B
AF5
3X�
�0.
18-
-N
uRD
CH
D4,
MTA
2X�
�0.
09T-
com
ple
xC
CT2
,C
CT3
X��
0.13
CC
T30.
11C
CT1
,C
CT2
,C
CT4
,C
CT5
,C
CT8
X��
0.31
26S
pro
teas
ome
PS
MC
1,P
SM
C2,
PS
MC
3,P
SM
C4,
PS
MC
5,P
SM
C6,
PS
MD
1,P
SM
D2,
PS
MD
3,P
SM
D8,
PS
MD
11,
PS
MD
12,
PS
MD
13,
PS
MD
14
X��
0.16
PS
MC
1,P
SM
C2,
PS
MC
3,P
SM
C4,
PS
MD
2,P
SM
D3
X��
0.18
PS
MA
50.
35
Am
inoa
cyl-
tRN
Asy
nthe
tase
mul
tienz
yme
--
KA
RS
,S
CY
E1,
IAR
S,
RA
RS
,E
PR
SX�
�0.
25-
-
Rib
osom
eR
PLP
0,R
PLP
1,R
PL3
,R
PL4
,R
PL6
,R
PL7
,R
PL8
,R
PL9
,R
PL1
0,R
PL1
3,R
PL1
3A,
RP
L14,
RP
L18,
RP
L18A
,R
PL2
2,R
PL2
3,R
PL2
4,R
PL3
0,R
PL3
5A,
RP
L36,
RP
S2,
RP
S3,
RP
S4X
,R
PS
5,R
PS
8,R
PS
9,R
PS
12,
RP
S13
,R
PS
15,
RP
S15
A,
RP
S18
,R
PS
20
X��
0.75
RP
LP0,
RP
L3,
RP
L4,
RP
L5,
RP
L6,
RP
L7,
RP
L7A
,R
PL8
,R
PL1
0A,
RP
L11,
RP
L13,
RP
L13A
,R
PL1
4,R
PL1
5,R
PL1
8A,
RP
L19,
RP
L21,
RP
L22,
RP
L23A
,R
PL2
4,R
PL2
8,R
PL2
9,R
PL3
1,R
PL3
2,R
PL3
4,R
PL3
5,R
PL3
5A,
RP
L36,
RP
S2,
RP
S3,
RP
S3A
,R
PS
4X,
RP
S6,
RP
S8,
RP
S9,
RP
S11
,R
PS
13,
RP
S14
,R
PS
15A
,R
PS
16,
RP
S20
,R
PS
24,
RP
S25
,R
PS
26,
RP
S27
,R
PS
28
X��
1.20
--
Unc
hara
cter
ized
TFG
1.39
TFG
1.25
FAM
103A
10.
47M
RT4
0.46
MR
T40.
43Z
NF3
260.
43Z
NF3
260.
14A
BC
F20.
31A
BC
F20.
11A
TPB
D1C
0.28
KIA
A00
200.
26
Proteomic Analysis of Mitotic RNA Polymerase II
10.1074/mcp.M111.011767–6 Molecular & Cellular Proteomics 11.6

from chromatin during mitosis, we found a highly significantenrichment for GO processes directly linked to transcriptionor downstream of transcription, namely, RNA splicing, pro-cessing, export, and translation. An association betweenmitotic RNAPII and SR proteins or Sm snRNPs had previ-ously been reported (10). In contrast, DNA-dependent pro-cesses that are largely independent from transcription (re-pair, replication) and such processes that may beconsidered to occur “upstream” from transcription (tran-scription factor activity, chromatin, chromatin modification)were less or not enriched. These analyses suggest thatRNAPII interactions in mitosis are linked to the transcrip-tional activity of RNAPII in interphase, albeit independent ofchromatin or ongoing transcription.
Our investigation of the GO term “translation” is due to thepresence of ribosomal proteins, mostly identified as moreabundant relative to the control antibody. Although the largenumber of cellular ribosomes (�10 million per HeLa cell; (39))makes ribosomal proteins likely candidates for nonspecificinteractors, the higher apparent abundance of translation-related proteins in the 8WG16 sample compared with thecontrol antibody sample, and their identification above back-ground in other RNAPII purifications (31, 33) or phosphoCTD-interactor purifications (40) may reflect an association be-tween RNAPII and ribosomes previously reported duringinterphase (41, 42). In comparison, cytoskeletal proteins, animportant class of contaminant proteins identified in a recentstudy as binding directly to commonly used immunoprecipi-tation matrices (43), were not enriched in our specific immu-noprecipitations, but were instead enriched in our controlimmunoprecipitations (see Supplementary Data).
To investigate whether RNAPII interacts with proteins in-volved in mitotic processes, we also analyzed the presence ofproteins annotated with mitosis-related GO terms. The num-ber of such proteins in the total lysate sample is low (1–5proteins) and their enrichment was not statistically significant(p values: 0.12–0.56). Equivalent results were obtained forfractions 2 and 8. RNAPII interactions with proteins involved inmitosis are therefore not prevalent.
Phosphorylation of Mitotic RNAPII is Not Required for theMaintenance of Protein Interactions—To investigate whetherthe identification of RNAPII interactors was influenced by thealkaline phosphatase (AP) treatment used to maximize immu-noprecipitation efficiency with 8WG16, we analyzed a totallysate sample prepared in the absence of AP treatment andpresence of phosphatase inhibitors. Although many RNAPIIinteractions identified in the context of co-transcriptional pro-cesses, such as RNA processing, are thought to be depen-dent on CTD phosphorylation, we identified only 13 additionalproteins that had not been identified in the AP-treated sample(Figs. 2D, 2E). Several of these proteins are functionally linkedwith each other within the context of transcriptional elonga-tion, a part of the transcription cycle in which the RPB1subunit of RNAPII is highly phosphorylated, or are known to
TAB
LEII—
cont
inue
d
Gro
up/C
omp
lex
Lysa
teFr
actio
n2
Frac
tion
8
pro
tein
nam
enP
AI
pro
tein
nam
enP
AI
Pro
tein
nam
enP
AI
AC
TL8
0.21
C20
OR
F77
0.20
C22
OR
F28
0.19
C1O
RF7
70.
19Q
SE
R1
0.18
LUC
7L2
0.17
KIA
A04
600.
14LA
RP
10.
06LA
RP
10.
09C
15O
RF4
40.
16O
ther
pro
tein
sR
PA
P2
0.72
RP
AP
20.
11R
PA
P2
0.25
XA
B1
0.49
XA
B1
0.56
MA
TR3
0.33
MA
TR3
0.45
NU
MA
10.
05A
LMS
10.
20FA
M12
0A0.
31FA
M12
0A0.
30M
YB
BP
1A0.
23
Proteomic Analysis of Mitotic RNA Polymerase II
Molecular & Cellular Proteomics 11.6 10.1074/mcp.M111.011767–7
Proteomic Analysis of Mitotic RNA Polymerase II
10.1074/mcp.M111.011767–8 Molecular & Cellular Proteomics 11.6

interact with each other (2): the TFIIF subunits GTF2F1 (8peptides) and GTF2F2 (6 peptides), the CTD phosphataseFCP1 (11 peptides), as well as CDC73 (2 peptides), a subunitof the PAF complex. Two other proteins with roles in tran-scriptional elongation appear in both �AP and –AP lysates,but are more abundant in the –AP sample: the DSIF subunitSPT5H (8 and 1 peptides, respectively) and the TFIID subunitTAF9 (3 and 1 peptides, respectively). The small overall dif-ference between the mass-spectrometric results of the �APand –AP samples (Figs. 2D, 2E) suggests that the mainte-nance of the association of most proteins with mitotic RNAPIIis independent of the phosphorylation state of RPB1 and its
associated proteins, although phosphorylation may be impor-tant for recruitment. The presence of hypophosphorylatedRNAPII (RNAPIIA) in early gel filtration fractions (Fig. 1D) is alsoconsistent with phosphorylation-independent interactions be-tween RNAPII and associated proteins within the larger sizecomplexes.
Meta-analysis of RNAPII Mass-Spectrometric Data—Tocompare our data for mitotic RNAPII interactors to that ofthree recent studies investigating RNAPII associated proteinsin cycling human cells (31–33), we determined the degree ofoverlap between nonredundant lists of the proteins identifiedin the four studies. In brief, Das et al. (2007) analyzed RNAPII
FIG. 2. Analysis of RNAPII-associated proteins identified by mass-spectrometry. A, Western blots against the URI/prefoldin subunitsRPAP3 and RUVB2 and against TFG, ACTL8, QSER1 and ALMS1, confirm respective mass-spectrometric detection (asterisks) or absence inthe total lysate, fraction 2 and fraction 8. B, Proportional Venn diagram comparing RNAPII-associated proteins among total lysate and fractions2 and 8. C, GO analysis for proteins identified in the total lysate, fraction 2 and fraction 8 reveals very strong enrichment for GO termsassociated with transcription and downstream processes. D, Proportional Venn diagram comparing the proteins identified in RNAPIIcomplexes treated �AP, and lists of proteins specific to AP or �AP. Overlap includes proteins present in both samples that were specific(identified with �2 peptides) or more abundant in only one of the two samples. E, Comparison of peptide abundance indices in total lysates�AP, revealing that phosphorylation of RPB1 has little contribution to the relative abundance of proteins in the two samples (Pearsonproduct-moment correlation coefficient of 0.897). See supplementary Fig. S2 for evidence that antibody 8WG16 can immunoprecipitate RPB1phosphorylated on Ser2 and/or Ser5 of the CTD. F, Venn diagram comparing the RNAPII associated proteome identified in the present studyto three previous proteomic analyses of RNAPII. For Jeronimo et al. (2007), only proteins directly co-immunoprecipitated with RNAPII subunitswere included.
TABLE IIIProteins identified in the full lysate, fraction 2 and fraction 8. PAI values in brackets indicate identification of only one peptide
Name IdentifierPAI
PAILysate Fraction 2 Fraction 8
RPB5 IPI00291093 1.00 0.62 0.69 2.31RPB7 IPI00218895 1.20 0.50 0.60 2.30RPB4 IPI00007283 1.00 0.50 0.50 2.00RPB8 IPI00003309 0.60 0.60 0.80 2.00TFG IPI00294619 1.33 (0.07) 0.60 2.00RPB3 IPI00018288 0.88 0.44 0.50 1.81RPB10 IPI00003311 1.00 0.40 0.40 1.80RPB2 IPI00027808 1.07 0.35 0.36 1.78RPB11 IPI00873238 1.14 0.29 0.29 1.71RPB1 IPI00031627 1.05 0.36 0.27 1.68RPB9 IPI00006113 0.89 0.33 0.44 1.67RPB6 IPI00015119 1.00 (0.20) 0.40 1.60ILF2 IPI00005198 0.67 0.71 0.10 1.48RUVB1 IPI00021187 0.71 0.38 0.25 1.33RUVB2 IPI00009104 0.79 0.28 0.21 1.28GDOWN1 (GRINL1A) IPI00552141 0.76 0.24 0.28 1.28EF1A1 IPI00396485 0.28 0.60 0.32 1.20FUS IPI00260715 0.71 0.12 (0.06) 0.88RPAP2 IPI00293375 0.69 0.05 0.12 0.86HNRPK IPI00216746 0.52 0.26 (0.04) 0.81NONO IPI00304596 0.42 0.23 0.12 0.77RBP56 IPI00873762 0.18 0.15 0.13 0.45HNRPD IPI00028888 0.27 (0.07) (0.07) 0.40PRP19 IPI00004968 0.18 0.14 (0.05) 0.36SERPING1 IPI00879931 0.07 0.11 0.11 0.29EWS IPI00009841 0.09 0.09 0.09 0.26C1R IPI00296165 0.11 0.07 0.07 0.24C1QB IPI00477992 (0.08) (0.08) (0.08) 0.23NAP1L1 IPI00023860 (0.07) (0.07) (0.07) 0.21TIF1B IPI00438229 0.06 (0.03) 0.06 0.14CP IPI00017601 (0.02) (0.02) 0.06 0.10C4B IPI00889723 0.02 (0.01) 0.02 0.06
Proteomic Analysis of Mitotic RNA Polymerase II
Molecular & Cellular Proteomics 11.6 10.1074/mcp.M111.011767–9

complexes from a HeLa cell nuclear extract after 8WG16immunoprecipitation and total elution with gel loading buffer,Jeronimo et al. (2007) from EcR-293 whole cell extracts afterimmunoprecipitation of TAP-tagged RNAPII subunits, andMcCracken et al. (2005) from a HeLa cell nuclear extract after8WG16 immunoprecipitation and high salt elution.
The meta-analysis revealed a set of proteins exclusive toeach purification stratagem (Fig. 2F, supplementary Table S3),with 333 proteins specific to the present study, as comparedwith 30, 25, and 16 for the McCracken et al., Das et al. andJeronimo et al. data sets, respectively. Of note, among theknown RNAPII-associated proteins exclusively identified inour study, many have high PAI values (e.g. SMN, NONO,PRMT1), showing that the differences in protein detectionbetween the present and previous studies are not simply dueto higher detection sensitivity of our mass spectrometrymethod, but most likely reflect inherent differences in thepurification strategies.
Our proteomic analyses of RNAPII complexes, extracted inmild conditions from mitotic extracts, identify novel RNAPII-associated proteins, including previously characterized(e.g. ALMS1, DBC1), and uncharacterized proteins (e.g.FAM103A1, ABCF2, ZNF326, QSER1). GO analysis of the 333RNAPII interactors exclusively identified in our study revealedenrichment for various terms associated with transcriptionand related processes (Fig. 2F).
No proteins were identified as common to all four studies,although several proteins, mostly involved in splicing and othertranscription related processes, were found in at least three ofthe four studies. Several proteins co-purified with RNAPII in twoof the studies, including proteins involved in translation.
RNAPII Interactors Colocalize With RNAPII During Mito-sis—To determine whether the RNAPII-associated proteinsidentified in vitro also associate with RNAPII in situ, weperformed antibody-blocking assays as a high-resolutionmethod to determine colocalization between candidate pro-teins and RNAPII (20, 41, 44). Antibody-blocking assaysmeasure the decrease in binding of one antibody after pre-incubation with a test antibody; if the two target epitopes liewithin the size range of the antibody molecules used to detectthem (�9 nm in the case of IgG type antibodies), binding ofthe first (blocking) antibody produces steric hindrance thatprevents binding of the second (test) antibody, resulting indecreased fluorescence intensity.
We performed antibody-blocking assays as previouslyshown using ultrathin (�150 nm thick) cryosections of fixedcells (20, 44), which optimally preserve RNAPII organizationand avoid extraction of relevant nuclear proteins (45). Briefly,cryosections were first immunostained using either a set ofavailable antibodies against proteins of interest or a controlantibody raised against biotin that specifically detects mito-chondria in the cytoplasm, before being immunostained using8WG16 (20). To measure the colocalization of candidate in-teractors with RNAPII independently of its phosphorylation
state, cryosections were pretreated with alkaline phosphataseto maximize 8WG16 detection of RNAPII ((19); data notshown). Images from mitotic cells were collected in the sameconditions for control and antibody-blocked sections. Weanalyzed both known (RUVB1, RUVB2, ILF2, TFG) and novel(QSER1, ALMS1, RPS6, RPL26) RNAPII interactors. A positivecontrol antibody detecting Sm proteins, known to associatewith RNAPII, was also employed (19). To investigate whetherRNAPII interactors identified in HeLa cells also associate withRNAPII in primary human cells, we performed antibody-block-ing experiments in both HeLa cells and human primary Tlymphocytes (PHA-activated peripheral blood mononuclearcells).
As expected, the antibody-blocking experiments showedcolocalisation between the CTD of RPB1 and RPB2 in bothHeLa cells and lymphocytes (Fig. 3). Furthermore, statisticalanalysis showed significant decreases in the staining intensityof 8WG16 after incubation with antibodies to RNAPII-interac-tors TFG, QSER1, and ALMS1, as well as with the positivecontrols RPB2 and Sm (all in comparison with antibiotin),demonstrating their close proximity to the CTD of RPB1 inmitotic HeLa cells and lymphocytes (Figs. 3A, 3B). ILF2 andRUVB2 blocking effects were only statistically significant inHeLa cells (p � 0.001 and p � 0.05, respectively). The colo-calization of RNAPII interactors in both HeLa cells and primarylymphocytes shows that their association with RNAPII is un-likely to be cell-type or cancer specific. It is worthwhile notingthat lack of colocalization in this assay does not exclude thepossibility that proteins interact with RNAPII in a complex, ifthe respective epitopes tested lie more than 10 nm apart, forexample if they interact with the RNAPII complex further awayfrom the CTD (detected by 8WG16), or if their association withRNAPII is of an indirect nature.
Interactor Requirement for Transcript Elongation During In-terphase—Our strategy to identify RNAPII-associated pro-teins uses mitotic cell extracts. To investigate whether thenovel RNAPII interactors have functions in the transcriptionalactivity of RNAPII complexes during interphase, we con-ducted a small-scale functional screen to determine changesin levels of ongoing transcription and nuclear organization atthe single-cell level.
We transfected HeLa cells with siRNAs against 25 RNAPIIinteractors and employed an in situ transcription assay basedon the incorporation of 5-bromouridine 5�-triphosphate (Br-UTP) by engaged RNAPII complexes (18). For this analysis,we chose proteins (1) previously known to associate withRNAPII (RUVB1, RUVB2, MATR3, GDOWN1, RPAP3, RPAP2,ILF2, NUMA1, HNRPD, XAB1/GPN1, TFG), (2) exclusivelyidentified in our study as RNAPII associated, but which haveeither been previously characterized (CCT3, WDR92,MYBBP1A/MBB1A, ALMS1, FAM120A/F120A), or (3) thatwere completely uncharacterized (QSER1, C20ORF77/RPRD1B, KIAA0460/RPRD2, ATPBD1C/GPN3, FAM103A1/F103A, ABCF2, ACTL8, C1ORF77/CA077, ZNF326).
Proteomic Analysis of Mitotic RNA Polymerase II
10.1074/mcp.M111.011767–10 Molecular & Cellular Proteomics 11.6

The levels of transcript elongation measured for eachknockdown after immunofluorescence detection of Br-la-beled RNA were compared with levels obtained with negative
control siRNA. Treatments with siRNAs that lead to a knock-down of the RNAPII subunits RPB2 and RPB4 resulted inroughly 60–70% of the levels of newly-made transcripts ob-
FIG. 3. Antibody-blocking assay reveals in situ colocalisation between RPB1 and several candidate proteins in mitosis. A, Cryosectionswere first treated with AP to maximize 8WG16 binding to total RPB1, and indirectly immunolabeled in the absence (negative control) or presenceof antibodies to candidate proteins (QSER1 or ALMS1), positive blocking antibody against RPB2, or an unrelated anti-biotin antibody that detectsmitochondria in the cytoplasm (arrowhead, negative control). After cross-linking of the first immunocomplex, sections were indirectly immunola-beled with 8WG16 (red). Nucleic acids were counterstained with DAPI (blue), and confocal images collected using the same settings without signalsaturation. Scale bar: 6 �m. Pre-incubation with anti-RPB2 reduces the intensity of 8WG16 (RPB1) signal throughout the nucleoplasm. Controlpre-incubation with antibiotin control antibody before labeling with 8WG16 has little or no effect. B, Antibody inhibition assays (ABA) in mitotic cellsshow a decrease in 8WG16 detection after blocking with antibodies against RPB2, TFG, QSER1, ALMS1, and Sm proteins, in both HeLa cells andprimary human PHA-activated peripheral blood mononuclear cells. In HeLa cells, colocalisation is also detectable after blocking with antibodiesagainst RUVB2 and ILF2. Asterisks denote statistical significance relative to the anti-biotin sample at significance levels of p � 0.05 (*), p � 0.01(**), and p � 0.001 (***). Average and SD of two coverslips are represented. Number of nuclear profiles was �5 for each coverslip. Statisticalsignificance was tested using ANOVA and Benjamini-Hochberg correction for multiple testing; uncorrected p values are indicated in supplementaryTable S5.
Proteomic Analysis of Mitotic RNA Polymerase II
Molecular & Cellular Proteomics 11.6 10.1074/mcp.M111.011767–11

FIG. 4. Many RNAPII interactors identified in mitosis are functionally and cytologically associated with RNAPII during interphase. A,Br-UTP transcription assay after siRNA-mediated knockdown of candidate proteins. Representative images for Br-RNA labeling after RNAiagainst a selection of targets. Scale bar: 10 �m. Graph represents mean nuclear Br-RNA intensities per cell (average and SD from two
Proteomic Analysis of Mitotic RNA Polymerase II
10.1074/mcp.M111.011767–12 Molecular & Cellular Proteomics 11.6

tained with control siRNA (Figs. 4A, 4B). Greater effects werelikely not observed because of incomplete depletion of RNA-PII subunits, despite good transcript depletion detected byqRT-PCR of total RNA (Fig. 4A), or due to the death of moreseverely affected cells, presence of untransfected cells, andbackground transcription by RNAPIII. Pre-incubation of per-meabilized HeLa cells with a concentration of the transcriptioninhibitor �-amanitin (1 �g/ml) that completely inhibits tran-scription by RNAPII, but which leaves RNAPIII unaffected,yields levels of Br-RNA detection of 7–9% of the levels ob-served in untreated cells (supplementary Fig. S3), as expected(18).
We find that 14 of the 25 RNAPII interactors tested bysiRNA knockdown are involved in sustaining normal levels oftranscript elongation in interphase (Fig. 4A), including six outof 13 proteins that were exclusively identified as RNAPII in-teractors in this study. Interestingly, we identify proteins thatare required to support transcriptional activity (RUVB1,RUVB2, HNRPD, QSER1, ALMS1, TFG, ILF2, FAM120A,ACTL8), as well as proteins that have an inhibitory effect onlevels of transcription elongation (XAB1, MYBBP1A, RPAP2,C20ORF77, FAM103A1), as shown by an increase in tran-scriptional activity after RNAi. Knockdown of target mRNAswas confirmed by qPCR for all candidate proteins (Fig. 4A,supplementary Table S4). Two proteins, which were also iden-tified by Jeronimo et al. to be associated with nuclear RNAPIIcomplexes, RPAP2 and XAB1, were shown here to be func-tionally relevant to the activity of RNAPII, with knockdown ofeither of these proteins leading to an increase in transcrip-tional activity. A yeast homolog of RPAP2, Rtr1, has recentlybeen identified as a potential RNAPII-S5p specific phospha-tase (46). In those cases in which RNAi knockdown does notaffect elongation, candidate proteins may be involved in otheraspects of transcription, or levels of protein knockdown maybe insufficient to reveal a phenotype, depending on the sta-bility of each protein studied.
Among the proteins that colocalize with RNAPII during mi-tosis are several with a clear involvement in nuclear transcrip-tion (QSER1, ALMS1, ILF2, RUVB2, TFG; Figs. 3B, 4A). Wefirst asked whether several of these RNAPII interactors were
localized inside the nucleus during interphase. Importantly, alltested antibodies against candidate RNAPII interactors led todetectable staining within the cell nucleus (Fig. 4B and datanot shown). This finding is especially significant for ALMS1, aprotein defective in Alstrom syndrome. Although ALMS1 con-tains nuclear localization signals and a putative leucine zipper(47, 48), previous studies reported its localization to centro-somes and the basal bodies of cilia (49). Nuclear localizationof ALMS1 was confirmed using two independent antibodies,raised against epitopes in the internal part of the protein or theC-terminal region. The two antibodies also show nuclear andcytoplasmic staining in cryosections of murine pancreatic andhypothalamic tissue samples (Fig. 4B), both of which areassociated with Alstrom syndrome pathology (50). Immuno-localisation in ultrathin cryosections (�150 nm thickness) op-timally preserves nuclear architecture and the localization ofnuclear proteins such as TBP and RNAPII (45), raising thepossibility that the nuclear localization of ALMS1 might nothave been detected in previous studies because of the cellu-lar preparation before immunolabeling, which could have hin-dered antibody accessibility or resulted in protein extraction.
To test whether the physical association between RNAPIIinteractors identified in mitosis is retained during interphase,we repeated the antibody-blocking experiment using inter-phase cells of both primary PHA activated peripheral lympho-cytes and HeLa cells. The blocking experiments reveal thatthe colocalization and physical association with RNAPII ob-served for the proteins RUVB2, ILF2, TFG, QSER1, andALMS1 during mitosis are retained during interphase, both inHeLa cells and in primary lymphocytes (Fig. 4C), thus showingthat the effects on global transcription observed for theseproteins are likely of a direct nature.
In summary, the results of the transcription and colocaliza-tion assays serve to validate the low stringency, mitosis-based approach to identifying proteins that functionally inter-act with RNAPII in both mitosis and interphase.
DISCUSSION
We set out to isolate RNAPII and its associated proteinsfrom mitotic cells as a strategy to circumvent the harsh ex-
coverslips). Two independent experiments are shown per candidate protein. Knockdowns of RPB2 and RPB4 served as positive controls.Transcriptional activity is expressed relative to matched AllStars Negative siRNA control (set to 100%). Asterisks mark statistical significancerelative to control siRNA at significance levels of p � 0.05 (*), p � 0.01 (**), and p � 0.001 (***). Statistical significance was tested using ANOVAand Benjamini-Hochberg correction for multiple testing. See supplementary Fig. S3 for images from the remaining treatments and a differentgraphical representation of the results. B, Immunolabeling of cryosections demonstrates nuclear localization of QSER1, TFG and ALMS1 inHeLa cells. Nuclear localization of ALMS1 was also tested in murine pancreatic and hypothalamic tissue, using antibodies directed againstinternal or C-terminal epitopes of human ALMS1. Nucleic acids were counterstained with DAPI (not show); nuclear perimeters are representedas white dotted lines. Scale bars: 6 �m. C, Antibody inhibition assay (ABA) in interphase cells shows colocalisation between RPB1 andassociated proteins identified in mitosis. Measurements of average 8WG16 intensity across the nucleoplasm in both HeLa cells and humanPHA-activated peripheral blood mononuclear cells show a decrease in 8WG16 detection after blocking with antibodies against RPB2, TFG,QSER1, ALMS1, RUVB2, ILF2, and Sm proteins. Asterisks denote statistical significance relative to the anti-biotin sample at significance levelsof p � 0.05 (*), p � 0.01 (**), and p � 0.001 (***). Error bars: SD of two coverslips. Number of nuclear profiles was �5 for each coverslip.Statistical significance was tested using ANOVA and Benjamini-Hochberg correction for multiple testing; uncorrected p values are indicatedin supplementary Table S5.
Proteomic Analysis of Mitotic RNA Polymerase II
Molecular & Cellular Proteomics 11.6 10.1074/mcp.M111.011767–13

traction conditions usually employed in purifications of RNAPIIfrom unsynchronized, largely interphasic cells. We combinedour isolation method with control immunoprecipitations, a spe-cific elution regimen, and highly sensitive mass spectrometry,leading to the identification of a large number of novel andknown interactors. Validation of RNAPII associations by func-tional and cytological methods for several candidate proteinsconfirms a significant proportion of novel interactors.
In line with earlier reports, we find that mitotic RNAPII isassociated with factors involved in RNA splicing and process-ing (10), and is phosphorylated on the S2 and/or S5 residuesof the RPB1 C-terminal domain (51). However, the demon-stration that phosphorylation is restricted to the subpopula-tion of larger RNAPII complexes (Fig. 1D) may suggest a directrelationship between these two observations. In light of thegenerally held view of recruitment of RNA processing factorsby phosphorylated RNAPII complexes during transcription,association of RNA processing factors with RNAPII duringmitosis may also directly result from RNAPII phosphorylation,and serve to facilitate the co-ordinated postmitotic nuclearre-import. However, some of the associations observed be-tween RNAPII and RNA processing factors during mitosis, inlight of their insensitivity to RNAPII dephosphorylation, areindicative of more permanent interactions.
A group of 32 proteins was consistently identified in thethree samples treated with alkaline phosphatase. The pres-ence of these proteins in all of these samples suggests thatthey bind to RPB1 directly or indirectly by interactions amongeach other. Most of these proteins (25 out of 32) have previ-ously been implicated in transcription-related processes.These include recently identified RNAPII associated proteinssuch as GDOWN1 and RPAP2, and proteins that so far hadonly been implicated in the transcription of specific genes(ILF2, RUVB1, RUVB2). SiRNA knockdown demonstrated aglobal requirement for ILF2 and RUVB1/2 in transcriptionalelongation, as well as for MYBBP1A, which was exclusivelyfound in the prefractionation lysate and has previously beenshown to be important for the transcription of selected genes(52). RUVB1 and RUVB2 have been identified as constituentsof a variety of protein complexes (53). It will be interesting tosee whether they perform comparable functions in all of thesecomplexes, be it a role in complex assembly, DNA binding, orother processes.
Especially noteworthy is the identification of three previ-ously uncharacterized proteins as functional interactors ofRNAPII: QSER1, C20ORF77, and ACTL8. Our results imply arole for these proteins within the context of transcription, asthey copurify with RNAPII in mitotic cells and support thenormal transcriptional activity of RNAPII in interphase. ForQSER1, we demonstrated colocalization with RNAPII in bothinterphase and mitosis (Figs. 3B, 4C); for ACTL8, the availableantibodies did not stain cryosections, whereas for C20ORF77preliminary antibody-blocking experiments also show somelevel of blocking (not shown). C20ORF77 appears to be or-
thologous to yeast Rtt103 and C. elegans CIDS-1, proteinsshown to be involved in transcriptional termination (54, 55). Aninvolvement of C20ORF77 in transcription termination is inagreement with the observed increase in Br-UTP incorpora-tion during transcript elongation upon knockdown of theprotein.
Importantly, we have also functionally verified two novelinteractors of RNAPII that are involved in human disease. Thegene coding for ALMS1 is mutated in patients suffering fromAlstrom syndrome (47, 48), a progressive and multisystemicdisorder that is characterized by neurosensory, developmen-tal, and metabolic dysfunctions (50), including diabetes andobesity. The ALMS1 protein has previously been shown tolocalize to centrosomes and to the basal body of cilia (49), afact that fits well with some of the pathology observed inAlstrom syndrome. In our study, we find that ALMS1 co-purifies with RNAPII, and that it is important for sustaining thetranscriptional activity of RNAPII. We also find that it localizesin the interphase nucleus in both human cells and murinetissues, where it lies in close proximity to RNAPII, as shown byantibody-blocking assays. This data implies that the molecu-lar basis of Alstrom syndrome is more complicated than hasbeen previously appreciated, involving control of gene ex-pression, and may lead to a better understanding of thedysfunctions observed in this syndrome.
The second verified RNAPII interactor involved in diseaseformation is TFG, which forms oncogenic fusion proteins invarious cancer types involving either tyrosine kinases or thetranscription factor NOR1 (56). TFG contains a SYGQ-richregion that shares similarity with the SYGQ-rich regions of theTET family of proteins, TLS/FUS, EWS, and TAF15/RBP56, allof which co-purified with RNAPII in all of our samples. Themembers of the TET family are known to associate with RNA-PII and to take part in transcription-related processes (57),and may have roles in motor neuron disease (58). They alsoform oncogenic fusion proteins with transcription factors inwhich their SYGQ-rich regions are fused to the DNA-bindingdomains of their fusion partners. The SYGQ-rich regions arethought to act as transactivation domains in these fusionproteins. The co-purification of TFG with RNAPII, its effect ontranscriptional activity, and colocalization with RNAPII sug-gest that the pathology of the TFG-NOR1 fusion protein islikely based on its ability to transactivate RNAPII.
The large number of RNAPII interactors found associatedwith native complexes easily isolated from the mitotic cyto-plasm raises the possibility that other protein complexes in-volved in chromatin-associated processes may, at least par-tially, dissociate from mitotic chromosomes intact. Therefore,our approach of using mitotic lysates, as starting materials forthe isolation of protein complexes and their associated fac-tors, may be more widely applicable to other nuclear com-plexes, and may lead to the identification of proteins other-wise lost due to the unphysiological purification conditionsoften required to extract nuclear proteins.
Proteomic Analysis of Mitotic RNA Polymerase II
10.1074/mcp.M111.011767–14 Molecular & Cellular Proteomics 11.6

In summary, using mild lysis conditions in the preparation ofRNAPII complexes from mitotic cells we have significantlyexpanded the proteome of RNAPII associated proteins, pro-viding an important resource for understanding RNAPII func-tion and regulation in humans. We also show that severalpreviously uncharacterized proteins and two proteins involvedin disease formation in humans are important for the tran-scriptional activity of RNAPII, thus opening the door for moredetailed investigations into their functions.
Acknowledgment—We thank Marc Vigneron (Strasbourg) for pro-viding the antibody 7C2.
* AM, SQX, BL, SM, MMB, and AP thank the Medical ResearchCouncil (UK) for support. BL thanks the Herchel Smith ResearchFund. GBC and JKN were funded by a National Institute of Healthgrant, HD036878.
□S This article contains supplemental Figs. S1 to S3 and TablesS1 to S5.
�� To whom correspondence should be addressed: MRC ClinicalSciences Centre, Imperial College School of Medicine, HammersmithHospital Campus, Du Cane Road, London W12 0NN, UK. Tel.:�44.2083838232; Fax: �44.2083838306; E-mail: [email protected] or [email protected].
§§ Present address: Department of Biochemistry and MolecularBiophysics, Columbia University Medical Center, New York, NY10032, USA.
¶¶ Present address: Tenovus Laboratory, Cancer Sciences Divi-sion, Southampton University School of Medicine, General Hospital,Southampton, United Kingdom.
Author contributions: AM, designed and performed experiments,designed and performed data and bioinformatic analyses, wrote pa-per; SQX, designed and performed experiments and performed dataanalyses (protein immunofluorescence localization and antibodyblocking); FH, performed mass-spec runs and processed raw mass-spec data; BL, designed and performed bioinformatic analyses (GO,comparisons with other studies, other analyses with negative results);HP, designed and assisted in initial mitotic extract preparation, gelfiltration and western blotting studies; SM, characterized initial gelfiltration fractions by western blotting; FR, performed ANOVA forantibody blocking and siRNA assays; GC and JKN, provided tissuesand antibodies; MMB, designed bioinformatic analyses (GO, compar-isons with other studies, other analyses with negative results); ALG,designed initial mitotic extract preparation and gel filtration studies;MS, designed and performed mass-spec runs and processed rawmass-spec data; AP, designed and performed initial mitotic extractpreparation, gel filtration and western blotting studies, designed ex-periments and bioinformatic studies, wrote paper.
REFERENCES
1. Fuda, N. J., Ardehali, M. B., and Lis, J. T. (2009) Defining mechanisms thatregulate RNA polymerase II transcription in vivo. Nature 461, 186–192
2. Sims, R. J., 3rd, Belotserkovskaya, R., and Reinberg, D. (2004) Elongationby RNA polymerase II: the short and long of it. Genes Dev. 18,2437–2468
3. Phatnani, H. P., and Greenleaf, A. L. (2006) Phosphorylation and functionsof the RNA polymerase II CTD. Genes Dev. 20, 2922–2936
4. Brookes, E., and Pombo, A. (2009) Modifications of RNA polymerase II arepivotal in regulating gene expression states. EMBO Rep. 10, 1213–1219
5. Villard, J. (2004) Transcription regulation and human diseases. Swiss Med.Wkly 134, 571–579
6. Faustino, N. A., and Cooper, T. A. (2003) Pre-mRNA splicing and humandisease. Genes Dev. 17, 419–437
7. Grosso, A. R., Martins, S., and Carmo-Fonseca, M. (2008) The emerging
role of splicing factors in cancer. EMBO Rep. 9, 1087–10938. Chaudhary, K., Deb, S., Moniaux, N., Ponnusamy, M. P., and Batra, S. K.
(2007) Human RNA polymerase II-associated factor complex: dysregu-lation in cancer. Oncogene 26, 7499–7507
9. Parsons, G. G., and Spencer, C. A. (1997) Mitotic repression of RNApolymerase II transcription is accompanied by release of transcriptionelongation complexes. Mol. Cell Biol. 17, 5791–5802
10. Kim, E., Du, L., Bregman, D. B., and Warren, S. L. (1997) Splicing factorsassociate with hyperphosphorylated RNA polymerase II in the absenceof pre-mRNA. J. Cell Biol. 136, 19–28
11. Rappsilber, J., Ishihama, Y., and Mann, M. (2003) Stop and go extractiontips for matrix-assisted laser desorption/ionization, nanoelectrospray,and LC/MS sample pretreatment in proteomics. Anal. Chem. 75,663–670
12. Selbach, M., Schwanhausser, B., Thierfelder, N., Fang, Z., Khanin, R., andRajewsky, N. (2008) Widespread changes in protein synthesis inducedby microRNAs. Nature 455, 58–63
13. Ishihama, Y., Rappsilber, J., Andersen, J. S., and Mann, M. (2002) Micro-columns with self-assembled particle frits for proteomics. J. Chromatogr.A 979, 233–239
14. Cox, J., and Mann, M. (2008) MaxQuant enables high peptide identificationrates, individualized p.p.b.-range mass accuracies and proteome-wideprotein quantification. Nat. Biotechnol. 26, 1367–1372
15. Cox, J., Matic, I., Hilger, M., Nagaraj, N., Selbach, M., Olsen, J. V., andMann, M. (2009) A practical guide to the MaxQuant computational plat-form for SILAC-based quantitative proteomics. Nat. Protoc. 4, 698–705
16. Elias, J. E., and Gygi, S. P. (2007) Target-decoy search strategy for in-creased confidence in large-scale protein identifications by mass spec-trometry. Nat. Methods 4, 207–214
17. Beissbarth, T., and Speed, T. P. (2004) GOstat: find statistically overrepre-sented Gene Ontologies within a group of genes. Bioinformatics 20,1464–1465
18. Pombo, A., Jackson, D. A., Hollinshead, M., Wang, Z., Roeder, R. G., andCook, P. R. (1999) Regional specialization in human nuclei: visualizationof discrete sites of transcription by RNA polymerase III. EMBO J. 18,2241–2253
19. Xie, S. Q., Martin, S., Guillot, P. V., Bentley, D. L., and Pombo, A. (2006)Splicing speckles are not reservoirs of RNA polymerase II, but contain aninactive form, phosphorylated on serine2 residues of the C-terminaldomain. Mol. Biol. Cell 17, 1723–1733
20. Ferrai, C., Xie, S. Q., Luraghi, P., Munari, D., Ramirez, F., Branco, M. R.,Pombo, A., and Crippa, M. P. (2010) Poised transcription factories primesilent uPA gene prior to activation. PLoS Biol. 8, e1000270
21. Maldonado, E., Shiekhattar, R., Sheldon, M., Cho, H., Drapkin, R., Rickert,P., Lees, E., Anderson, C. W., Linn, S., and Reinberg, D. (1996) A humanRNA polymerase II complex associated with SRB and DNA-repair pro-teins. Nature 381, 86–89
22. Pan, G., Aso, T., and Greenblatt, J. (1997) Interaction of elongation factorsTFIIS and elongin A with a human RNA polymerase II holoenzyme ca-pable of promoter-specific initiation and responsive to transcriptionalactivators. J. Biol. Chem. 272, 24563–24571
23. Kimura, H., Tao, Y., Roeder, R. G., and Cook, P. R. (1999) Quantitation ofRNA polymerase II and its transcription factors in an HeLa cell: littlesoluble holoenzyme but significant amounts of polymerases attached tothe nuclear substructure. Mol. Cell Biol. 19, 5383–5392
24. Thompson, N. E., Aronson, D. B., and Burgess, R. R. (1990) Purification ofeukaryotic RNA polymerase II by immunoaffinity chromatography. Elu-tion of active enzyme with protein stabilizing agents from a polyol-responsive monoclonal antibody. J. Biol. Chem. 265, 7069–7077
25. Bregman, D. B., Du, L., van der Zee, S., and Warren, S. L. (1995) Tran-scription-dependent redistribution of the large subunit of RNA polymer-ase II to discrete nuclear domains. J. Cell Biol. 129, 287–298
26. Patturajan, M., Schulte, R. J., Sefton, B. M., Berezney, R., Vincent, M.,Bensaude, O., Warren, S. L., and Corden, J. L. (1998) Growth-relatedchanges in phosphorylation of yeast RNA polymerase II. J. Biol. Chem.273, 4689–4694
27. Hu, X., Malik, S., Negroiu, C. C., Hubbard, K., Velalar, C. N., Hampton, B.,Grosu, D., Catalano, J., Roeder, R. G., and Gnatt, A. (2006) A Mediator-responsive form of metazoan RNA polymerase II. Proc. Natl. Acad. Sci.U.S.A. 103, 9506–9511
28. Nayler, O., Stratling, W., Bourquin, J. P., Stagljar, I., Lindemann, L., Jasper,
Proteomic Analysis of Mitotic RNA Polymerase II
Molecular & Cellular Proteomics 11.6 10.1074/mcp.M111.011767–15

H., Hartmann, A. M., Fackelmayer, F. O., Ullrich, A., and Stamm, S.(1998) SAF-B protein couples transcription and pre-mRNA splicing toSAR/MAR elements. Nucleic Acids Res. 26, 3542–3549
29. Dorjsuren, D., Lin, Y., Wei, W., Yamashita, T., Nomura, T., Hayashi, N., andMurakami, S. (1998) RMP, a novel RNA polymerase II subunit 5-inter-acting protein, counteracts transactivation by hepatitis B virus X protein.Mol. Cell Biol. 18, 7546–7555
30. Emili, A., Shales, M., McCracken, S., Xie, W., Tucker, P. W., Kobayashi, R.,Blencowe, B. J., and Ingles, C. J. (2002) Splicing and transcription-associated proteins PSF and p54nrb/nonO bind to the RNA polymeraseII CTD. RNA 8, 1102–1111
31. Das, R., Yu, J., Zhang, Z., Gygi, M. P., Krainer, A. R., Gygi, S. P., and Reed,R. (2007) SR proteins function in coupling RNAP II transcription topre-mRNA splicing. Mol. Cell 26, 867–881
32. Jeronimo, C., Forget, D., Bouchard, A., Li, Q., Chua, G., Poitras, C.,Therien, C., Bergeron, D., Bourassa, S., Greenblatt, J., Chabot, B.,Poirier, G. G., Hughes, T. R., Blanchette, M., Price, D. H., and Coulombe,B. (2007) Systematic analysis of the protein interaction network for thehuman transcription machinery reveals the identity of the 7SK cappingenzyme. Mol. Cell 27, 262–274
33. McCracken, S., Longman, D., Marcon, E., Moens, P., Downey, M., Nicker-son, J. A., Jessberger, R., Wilde, A., Caceres, J. F., Emili, A., andBlencowe, B. J. (2005) Proteomic analysis of SRm160-containing com-plexes reveals a conserved association with cohesin. J. Biol. Chem. 280,42227–42236
34. Ishihama, Y., Oda, Y., Tabata, T., Sato, T., Nagasu, T., Rappsilber, J., andMann, M. (2005) Exponentially modified protein abundance index (em-PAI) for estimation of absolute protein amount in proteomics by thenumber of sequenced peptides per protein. Mol. Cell Proteomics 4,1265–1272
35. Gstaiger, M., Luke, B., Hess, D., Oakeley, E. J., Wirbelauer, C., Blondel, M.,Vigneron, M., Peter, M., and Krek, W. (2003) Control of nutrient-sensitivetranscription programs by the unconventional prefoldin URI. Science302, 1208–1212
36. Boulon, S., Pradet-Balade, B., Verheggen, C., Molle, D., Boireau, S., Geor-gieva, M., Azzag, K., Robert, M. C., Ahmad, Y., Neel, H., Lamond, A. I.,and Bertrand, E. (2010) HSP90 and its R2TP/Prefoldin-like cochaperoneare involved in the cytoplasmic assembly of RNA polymerase II. Mol. Cell39, 912–924
37. Sardiu, M. E., Cai, Y., Jin, J., Swanson, S. K., Conaway, R. C., Conaway,J. W., Florens, L., and Washburn, M. P. (2008) Probabilistic assembly ofhuman protein interaction networks from label-free quantitative proteom-ics. Proc. Natl. Acad. Sci. U.S.A. 105, 1454–1459
38. Cloutier, P., Al-Khoury, R., Lavallee-Adam, M., Faubert, D., Jiang, H.,Poitras, C., Bouchard, A., Forget, D., Blanchette, M., and Coulombe, B.(2009) High-resolution mapping of the protein interaction network for thehuman transcription machinery and affinity purification of RNA polymer-ase II-associated complexes. Methods 48, 381–386
39. Gorlich, D., and Mattaj, I. W. (1996) Nucleocytoplasmic transport. Science271, 1513–1518
40. Phatnani, H. P., Jones, J. C., and Greenleaf, A. L. (2004) Expanding thefunctional repertoire of CTD kinase I and RNA polymerase II: novelphosphoCTD-associating proteins in the yeast proteome. Biochemistry43, 15702–15719
41. Iborra, F. J., Escargueil, A. E., Kwek, K. Y., Akoulitchev, A., and Cook, P. R.(2004) Molecular cross-talk between the transcription, translation, andnonsense-mediated decay machineries. J. Cell Sci. 117, 899–906
42. Brogna, S., Sato, T. A., and Rosbash, M. (2002) Ribosome components are
associated with sites of transcription. Mol. Cell 10, 93–10443. Trinkle-Mulcahy, L., Boulon, S., Lam, Y. W., Urcia, R., Boisvert, F. M.,
Vandermoere, F., Morrice, N. A., Swift, S., Rothbauer, U., Leonhardt, H.,and Lamond, A. (2008) Identifying specific protein interaction partnersusing quantitative mass spectrometry and bead proteomes. J. Cell Biol.183, 223–239
44. Pombo, A., Hollinshead, M., and Cook, P. R. (1999) Bridging the resolutiongap: Imaging the same transcription factories in cryosections by lightand electron microscopy. J. Histochem. Cytochem. 47, 471–480
45. Guillot, P. V., Xie, S. Q., Hollinshead, M., and Pombo, A. (2004) Fixation-induced redistribution of hyperphosphorylated RNA polymerase II in thenucleus of human cells. Exp. Cell Res. 295, 460–468
46. Mosley, A. L., Pattenden, S. G., Carey, M., Venkatesh, S., Gilmore, J. M.,Florens, L., Workman, J. L., and Washburn, M. P. (2009) Rtr1 is a CTDphosphatase that regulates RNA polymerase II during the transition fromserine 5 to serine 2 phosphorylation. Mol. Cell 34, 168–178
47. Collin, G. B., Marshall, J. D., Ikeda, A., So, W. V., Russell-Eggitt, I., Maffei,P., Beck, S., Boerkoel, C. F., Sicolo, N., Martin, M., Nishina, P. M., andNaggert, J. K. (2002) Mutations in ALMS1 cause obesity, type 2 diabetesand neurosensory degeneration in Alstrom syndrome. Nat. Genet. 31,74–78
48. Hearn, T., Renforth, G. L., Spalluto, C., Hanley, N. A., Piper, K., Brickwood,S., White, C., Connolly, V., Taylor, J. F., Russell-Eggitt, I., Bonneau, D.,Walker, M., and Wilson, D. I. (2002) Mutation of ALMS1, a large gene witha tandem repeat encoding 47 amino acids, causes Alstrom syndrome.Nat. Genet. 31, 79–83
49. Hearn, T., Spalluto, C., Phillips, V. J., Renforth, G. L., Copin, N., Hanley,N. A., and Wilson, D. I. (2005) Subcellular localization of ALMS1 supportsinvolvement of centrosome and basal body dysfunction in the pathogen-esis of obesity, insulin resistance, and type 2 diabetes. Diabetes 54,1581–1587
50. Marshall, J. D., Beck, S., Maffei, P., and Naggert, J. K. (2007) Alstromsyndrome. Eur. J. Hum. Genet. 15, 1193–1202
51. Xu, Y. X., and Manley, J. L. (2007) Pin1 modulates RNA polymerase IIactivity during the transcription cycle. Genes Dev. 21, 2950–2962
52. Hara, Y., Onishi, Y., Oishi, K., Miyazaki, K., Fukamizu, A., and Ishida, N.(2009) Molecular characterization of Mybbp1a as a co-repressor on thePeriod2 promoter. Nucleic Acids Res. 37, 1115–1126
53. Jha, S., and Dutta, A. (2009) RVB1/RVB2: running rings around molecularbiology. Mol. Cell 34, 521–533
54. Kim, M., Krogan, N. J., Vasiljeva, L., Rando, O. J., Nedea, E., Greenblatt,J. F., and Buratowski, S. (2004) The yeast Rat1 exonuclease promotestranscription termination by RNA polymerase II. Nature 432, 517–522
55. Cui, M., Allen, M. A., Larsen, A., Macmorris, M., Han, M., and Blumenthal,T. (2008) Genes involved in pre-mRNA 3�-end formation and transcrip-tion termination revealed by a lin-15 operon Muv suppressor screen.Proc. Natl. Acad. Sci. U.S.A. 105, 16665–16670
56. Hisaoka, M., Ishida, T., Imamura, T., and Hashimoto, H. (2004) TFG is anovel fusion partner of NOR1 in extraskeletal myxoid chondrosarcoma.Genes Chromosomes Cancer 40, 325–328
57. Law, W. J., Cann, K. L., and Hicks, G. G. (2006) TLS, EWS and TAF15: amodel for transcriptional integration of gene expression. Brief Funct.Genomic Proteomic 5, 8–14
58. Lagier-Tourenne, C., and Cleveland, D. W. (2009) Rethinking ALS: the FUSabout TDP-43. Cell 136, 1001–1004
59. Prasanth, K. V., Sacco-Bubulya, P. A., Prasanth, S. G., and Spector, D. L.(2003) Sequential entry of components of the gene expression machineryinto daughter nuclei. Mol. Biol. Cell 14, 1043–1057
Proteomic Analysis of Mitotic RNA Polymerase II
10.1074/mcp.M111.011767–16 Molecular & Cellular Proteomics 11.6