the timing of adolescent growth spurts of ten body dimensions in boys and girls of the wrocŀaw...
TRANSCRIPT

P. Bergman M. Gor~cy Institute of Anthropology, Polish Academy of Sciences, 50-951 Wroclaw, ul. Kugnicza 35, Poland
Received 15 October 1983 and accepted 2 March 1984
Keywords: twins, age at peak velocity, sequence of growth velocity peaks, heritability, h~.
The Timing of Adolescent Growth Spurts of Ten Body Dimensions in Boys and Girls of the Wroc~aw Longitudinal Twin Study
Age at peak velocity of growth in ten body dimensions was estimated by graphic interpolation in 186 twin pairs from the Wroc~aw longitudinal twin study. The sequence of these peak velocity ages was determined for boys and girls and sex dimorphism assessed. The results were similar to those from the literature. The h 2 and F for ages at peak velocity of the several characters were estimated. A significant influence of genetic factors on timing of the peak adolescent growth spurt was shown. The heritabilities (h") range from 0"6 to 0"9; the F coefficients with one exception are statistically highly significant. There is a suggestion of some relationship between the time of appearance of growth spurts in some characters and their recency of phylogenetic origin.
1. Introduction
Today, data on pubertal growth spurts of different body dimensions in twins are still few, and so there is little information on their genetic control. Ljung et al. (1977) and Fischbein (1977) published some data on pubertal growth spurts in height and weight of Swedish twins. Sklad (1977) investigated height of twins from Warsaw. Only pubertal growth spurts in height and in hand dimensions have been reported so far from the Wroclraw longitudinal twin study (WLTS) (Bergman et al., 1981; Bergman & Szmyd, 1981). It is the aim of the present work to estimate the age of the pubertal growth spurts, as indicated by the age at peak velocity, in ten body dimensions, and to examine the extent of their genetic determination, using the data of the WLTS.
2. Material, Traits and Methods
Out of a series of 224 twin pairs studied longitudinally since 1967 in the Anthropological Institute of the Polish Academy of Sciences in Wroc~aw, Poland, 186 pairs have been selected for the present study. Whether a pair was chosen or not depended upon completion of the relevant part of the growth curves. The composition of the material by zygosity and sex is shown in Table 2. Zygosity was assessed as follows:
By the established diagnostic methods of polysymptomatic similarity. By the index of dermatoglyphic similarity of the fingers, palms and soles, following Orczykowska-Swigtkowska (1964).
By the blood group systems as set out in Table 1.
The tbllowing measurements were considered:
(1) Body height (H). (2) Cervical sitting height (Ch). (3) Subischial leg length (L1). (4) Upper limb length (U1). (5) Body weight (W). (6) Biacromial diameter (A).
Journal of Human Evolution (1984) 13, 339-347
0047 2484/84/040339 + 09 $03.00/0 �9 1984 Academic Press Inc. (London) Limited
340 P. BERGMAN AND M. OOR~CY
(7) Biiliac diameter (Ic). (8) Transverse chest diameter (T1). (9) Anteroposterior chest diameter (Xi).
(10) H a n d length (Hd). (The data on hand length were reported by Bergman & Szmyd, 198I.)
F rom the measurements made regularly upon these twins, growth velocities were calculated, and then individual velocity curves were drawn for all the characters for each individual. On these curves the ages of peak growth velocity were identified by graphical interpolation following Bielicki & Welon (1973). This method was used to allow comparison of the results of the twin study (WLTS) with those of the Wroc~aw growth study (WGS, Welon & Bielicki 1979).
For each measurement the mean (M) and s tandard deviation (SD) of the age of pubertal peak velocity were calculated. For the estimation of the influence of genetic and nongenetic factors on the age at peak velocity, the intrapair variances o f the monozygot ic and dizygotic twins (VMz and VDZ) and from these the heritability coefficient h 2 following Clark (1956) and F ratio were calculated according to the following formulae:
VDz- V~z VDz h 2 = , F =
VDz VMz
Table 1 Distribution of numbers of twin pairs by sex, zygosity, and number of blood characters investigated
No. of twin pairs investigated No. of blood Boys Girls characters investigated MZ DZ MZ DZ
6 (ABO, MN, Rh, Hp, Gc, Gm) 7 (ABO, MN, Rh, Kell, Hp, Gc, Gm)
12 (ABO, MN, Rh, Kell, Hp, Gc, Gin, AP, PGM, AK, Inv, 6PGD)
13 (ABO, MN, Rh, Kell, Hp, Gc, Gin, AP, PGM, GPT, AK, Inv, 6PGD)
Total 186 pairs
11 15 13 10 14 19 15 18
14 7 10 7
12 11 5 5 51 52 43 40
Table 2 Means (M) and standard deviations (SD) of ages at peak velocity of 10 body dimensions in Wroctaw Twins
Girls (MZ + DZ) Boys (MZ + DZ)
No. of pairs No. of pairs M SD
MZ DZ MZ DZ M SD
( 1 ) Body height 44 38 11-84 1-08 50 50 13.92 1.00 (2) Cervical sitting height 42 38 12"02 1'20 49 45 14.19 1'05 (3) Subischial leg length 41 35 11-22 1"01 47 48 13'09 1'20 (4) Upper limb length 42 36 11"57 1'40 49 46 13"76 1"01 (5) Body weight 43 41 12'60 1"06 48 49 14'29 1'10 (6) Biacromial diameter 38 37 12'03 1'11 51 51 14'12 1'09 (7) Bi-iliae diameter 39 39 12'06 1'25 49 47 13'87 1.17 (8) Transverse chest 37 37 12'01 1.21 39 35 13"90 1.03 (9) Antero-posterior chest 34 36 11"87 1.25 35 34 13"80 1-17
(10) Hand length 18 14 11'73 0"93 22 18 13"89 1.08
TIMING OF ADOLESCENT GROWTH SPURTS 341
3. Results and Conclusions
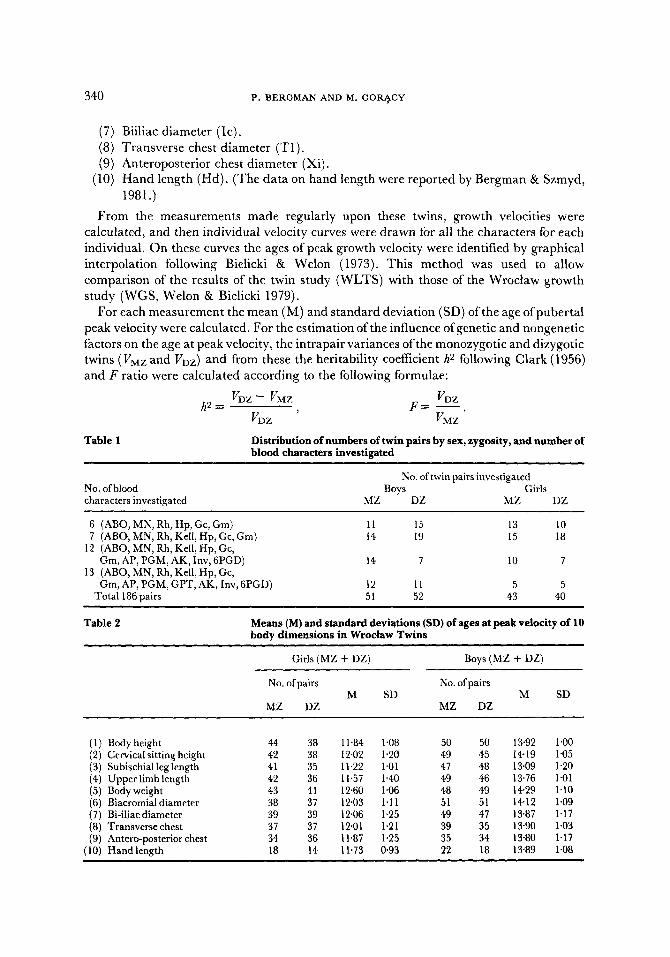
The Sequence of Growth Spurts Within each sex, no significant difference between monozygotic and dizygotic mean ages at peak velocity appeared in any of the ten measurements. For the sake of simplicity the mean and standard deviation for each character were therefore calculated in males and in females neglecting zygosity (Table 2, Figure 1). In each sex, the growth spurt appears first in lower limb length (L1) and last in weight (W). The mean difference in age between the growth spurts of these two characters is 1.4 years in girls and 1"2 years in boys.
Figure 1. Peak velocity sequence for 10 body diameters in Wroc[aw twins as compared with other data. O: Swedish singleton controls; STM: sitting height; • median; other explanations in text.
HGS s
WGS s
STM s
WLTS s
WLTS d
STM d
WGS d
HGS d
L I [ HI STI "~ I IC
Lt Ic Hd A H Ch W I I X I I I I
(• H W
Ch Ul Hxi TI AIc L,~ , ? ,+_ ,,,, W
55% "~.
LI
70% UI Xi IcTI H A Ch W
Hd
i G O l H H w W
I X I I I Ll Hd Ic W H
A Ch 1 I 1 I t LI H Ic A ST
,,'s ,~ ,2'~ ,~ d.5 ,'~ , ,s Age
The growth spurt in height (H) takes place later than that in lower limb length (L1) by about 0"6 years in girls and 0"8 years in boys, and correspondingly earlier than the growth spurt in weight (by about 0"8 years in girls and 0"4 years in boys) (Figure 1). The growth spurt in cervical sitting height (Ch) occurs about 0"2-0"3 years after that in height, and always earlier than that in weight. The sequence of the growth spurts in these four characters, L1-H-Ch-W, is in accordance with the generally established regularities of growth rhythms. In this respect there are no differences between twins and singletons. In both girls and boys mean peak velocity of biacromial diameter (A) appears after that of height, those of upper limb length (U1) and hand length (Hd) before it. The sequence of growth spurts in these seven dimensions is almost the same in both sexes. Only the mean peak velocities of the three remaining characters, anteroposterior chest (Xi), transverse

342 P. BERGMAN AND M. GOR~CY
chest (T1) and bi-iliac diameter (Ic) appear in boys very shortly before, and in girls shortly after, those of height. However several of these age differences are biologically meaningless, since a difference of 0"25 years in male and 0"35 years in females is necessary for minimal statistical significance.
The mean sex differences for the age of peak velocity range from 1"7 years in body weight to 2"2 years in upper limb length and cervical sitting height. Sex differences however are also manifest in the rhythm of the growth spurts. In girls there is quite a large time difference between the growth spurt in the group of advanced characters and that in weight; in boys on the other hand there is a large difference between the lower limb length and the later characters. Also the time differences between the mean peak velocity ages of height and of lower limb length and weight are very interesting. Considering the whole of the time sequence of the means, in girls about 45% of this occurs between the peak velocities of lower limb length and of stature, and about 55% between the latter and the peak velocity of weight. The corresponding percentages in boys are 70% and 30% (Figure 1). The rhythm of growth spurts thus reveals a quite pronounced sex difference.
In Figure 1 are compared the data on the twins and those from some other authors. The differences in methods of analysis however only allow general comparisons. Thus the method of graphic interpolation has been applied only in the Wroc~aw growth study (Welon & Bielicki, 1979) and in the present one. The Harpend-growth study (HGS) data (Tanner et al., 1976) were analysed using logistic functions and the Swedish material (Ljung et al., 1977; Fischbein, 1977) using a modified mid-year-velocity method. There is quite good accordance between our twins and those of the Harpend-growth study, especially as regards the sequence of the characters. Only the growth spurt of lower limb length is earlier in the twins. Comparison with the Swedish series of both twin (STM) and non-twin material shows similar patterns of height and weight peak growth velocities, though our girls are slightly earlier.
Quite similar results are observed when our girls are compared with those of the WGS for many characters. The only two exceptions lie in the peak growth spurts for biiliac (Ic) and biacromial (A) diameters. In both sexes in the WGS these come before height, while in our twins, and also in the Harpend-growth study, they come after it. There is a quite marked difference between male twins and the WGS boys, especially as regards peak velocity in leg length (L1) and height (H). It is however stressed that to the authors' knowledge the occurrence of the peak growth spurt of weight in advance of height has so far only been observed in the WGS boys (Figure 1). Also other authors have emphasized the rule that the height peak is always later than that of weight (Vital and Health Statistics, 1973; Miiller-K6nig, 1977). The mean sex differences in the twins in the pubertal growth spurts of all characters are very similar to those in other groups (less than 2'2 years). Again however the WGS sample is different, showing differences up to 2'7 years.
Thus one can say that the Wroclaw twins do not differ very much from other groups studied (other than the WGS boys) either in the age at which the maximum growth spurts occur or in the sequence among the characters. One exception however is the rather early age of peak velocity in the lower limb length in boys. Some of the differences that are observed are partly attributable to the differences between the methods used and partly to differences in the environments of the samples.
In our opinion it is impossible to formulate a rigid model to explain the average progression of the growth spurts, either for the sequence of characters themselves or for the

TIMING OF ADOLESCENT GROWTH SPURTS 343
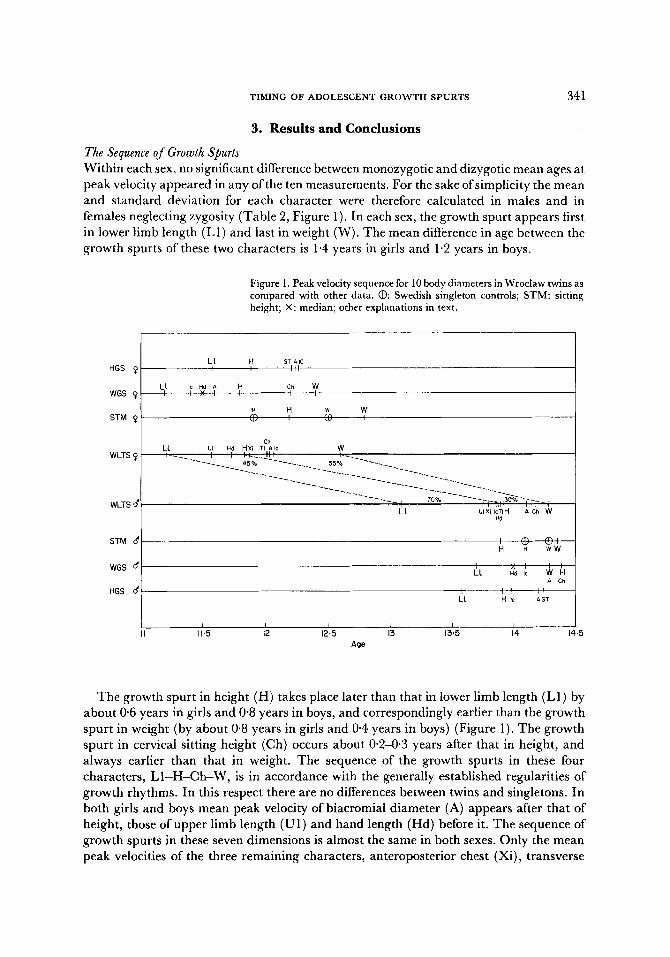
actual ages at which the peak velocity occurs, not only generally but also for any given population. Peak growth spurts might well be expected to occur in different sequences of characters in different individuals, affecting even those characters whose location in the mean sequence is the most stable, for example L1 -H -Ch -W . This also partly applies to the sex differences. Individual variations in the growth rhythm are possible which do not reflect the mean sequence. These are shown graphically in Figure 2.
The age distribution of the peak velocities of all the characters overlap to the extent of plus or minus one standard deviation. Such a graph demonstrates reality better than the average sequence of peak velocities (Figure 2). The rhythm of growth spurts is rather more fluid, more extensive in time and not so discontinuous.
LL
UI
H
A
Ic
TI
XL
Hc
Ct"
W
Figure 2. Mean peak velocity age ( + 1 SD) in WrocIaw twins.
I'1 12 GirLs
I )( ~,
I
�9 I
�9 I
�9 I
�9 I
I
I
�9 I
8
'[ 13 [4 15
Boys
I
:~ �9 I x
, , ' I *,
�9 ~ | )c
x ~ I >(
I ,, , I ~
i~1 ~ L , , I 13 14 15
Age (years)
The age distributions of the growth spurts of some characters in boys and girls also overlap by plus or minus one standard deviation.
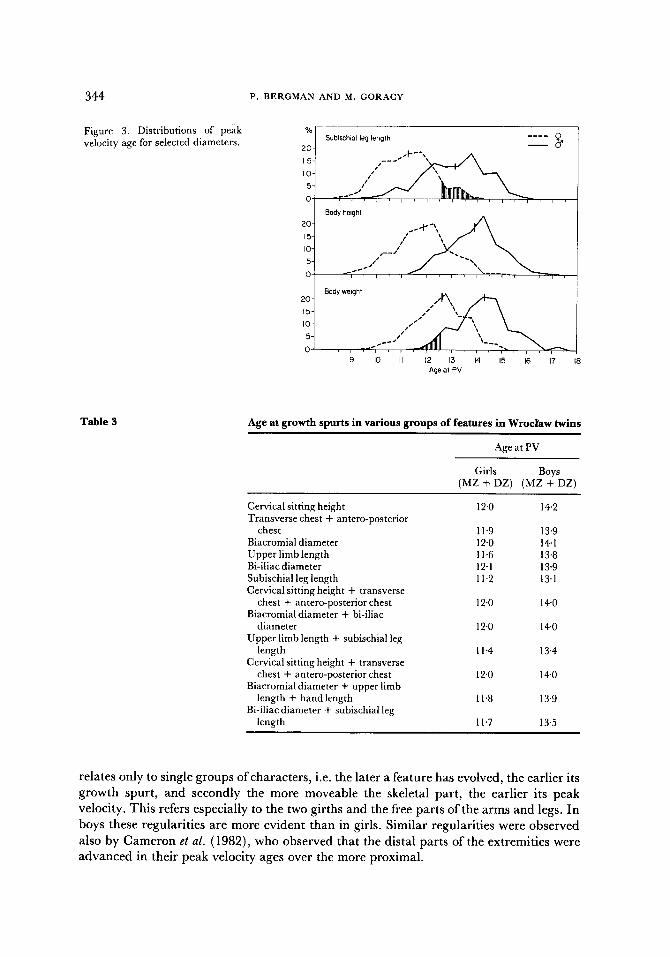
In Figure 3 the full distributions of the peak velocity ages of some characters are shown to overlap each other to an appreciable degree. Even the distributions of the earliest character in girls (L1) and latest in boys (W) overlap (Figure 3, the shaded areas).
With the exception of height and weight, the characters analysed here represent dimensions of skeletal parts. Skeletal parts differ from each other in function (moveable parts) and in phylogeny (older or younger in phylogenetic evolution). Both aspects are reflected in the anatomical organisation of the skeleton; the trunk (the dorsal spine and thorax), the skull, the appendicular skeleton (the shoulder and the pelvic girdles and the free parts of the limbs). One may ask whether the sequence of the peak growth velocities is related in some way with the evolutionary age of these skeletal parts. As can be seen in Table 3, there appears to be some relationship but it cannot be taken as a general rule; it
344
Figure 3. Distributions of peak velocity age for selected diameters.
P. BERGMAN AND M. GORACY
2O
15
I
O -
Subischial leg length . . . .
2 0 -
15-
I0 -
5-
0
20
15
I0.
5
C
s/ �9 ~
-,---~ , . . . . . . ~ . . . .
Body height
Body weight /~",, ~ s , s ~
9 I0 II 12 13 14 15 16 17 18 Age et PV
Table 3 Age at growth spurts in various groups of features in Wroclaw twins
Age at PV
Girls Boys (MZ-~DZ) (MZ+DZ)
Cervical sitting height 12'0 14"2 Transverse chest + antero-posterior
chest 11'9 13"9 Biacromial diameter 12.0 14'1 Upper limb length 11.6 13-8 Bi-iliac diameter 12-1 13'9 Subischial leg length 11 "2 13" 1 Cervical sitting height + transverse
chest + antero-posterior chest 12'0 14.0 Biacromial diameter + bi-iliac
diameter 12'0 14.0 Upper limb length + subischial leg
length 11'4 13'4 Cervical sitting height + transverse
chest + antero-posterior chest 12'0 14'0 Biacromial diameter + upper limb
length + hand length 11'8 13"9 Bi-iliac diameter + subischial leg
length 11"7 13-5
relates only to single groups of characters, i.e. the later a feature has evolved, the earlier its growth spurt, and secondly the more moveable the skeletal part , the earlier its peak velocity. This refers especially to the two girths and the free parts of the arms and legs. In boys these regularities are more evident than in girls. Similar regularities were observed
also by Cameron et al. (1982), who observed that the distal parts of the extremities were advanced in their peak velocity ages over the more proximal.
T I M I N G OF A D O L E S C E N T G R O W T H S P U R T S 345
Table 4 Variances of intrapair differences and heritability coefficients o f mean ages at peak velocities in 10 body dimensions in Wroc|aw Twins.
Girls Boys
VMZ VDz V~; h 2 F VMz Vl)z VG h 2 F
(1) Bodyheight 0"133 0.494 0'361 0.73 3.714 0'070 0'480 0'410 0'85 6"860 (2) Cervical sitting height 0.131 0'345 0-214 0-62 2.634 0.198 0.599 0'401 0.67 3.025 (3) Subischialleglength 0.065 0-340 0"275 0.81 5.231 0'209 0-649 0.440 0"68 3"105 (4) Upperlimblength 0'096 0"491 0"395 0-80 5.115 0.151 0-408 0"257 0"63 2.702 (5) Bodyweight 0-174 0-583 0"409 0-70 3-351 0"130 0-7t0 0-580 0-82 5-462 (6) Biacromialdiameter 0-162 0'267 0'105 0"39 1-648" 0.119 0.366 0"247 0'68 3-076 (7) Bi-iliacdiameter 0-091 0"369 0.278 0 '75 4.055 0'112 0'502 0-390 0"78 4"482 (8) Transverse chest 0"073 0.589 0'516 0"87 3"068 0.114 0.446 0'332 0"74 3"912 (9) Antero-posteriorchest 0-079 0"621 0.542 0 '87 7.861 0'097 0.435 0"338 0.78 4.486
(10) Handlength 0"173 0.442 0'269 0"61 2 '555 0"080 0.729 0'649 0"89 9"106
* Not significant.
Genetic Basis of Peak Velocity Ages T h e ages of peak veloci ty of the ten body d imensions give h 2 values which are not very different from each o ther (Tab le 4). In boys they vary from 0"63 to 0"89, and in girls f rom 0"39 to 0"87. Were it not for the lowest h 2 value for shoulder b r e a d th in girls (0"39), the r ange of the h 2 values in both sexes would be between 0"6 and 0"9. For the indiv idual charac te rs
no grea t sex difference can be seen. The Fcoeff icients , o ther than tha t for shoulder b r e a d th in girls, are s ta t is t ical ly highly significant (P <0"01). One may therefore speak of a s t rong
genet ic control of the t ime of peak growth velocity. T h e size of h 2 does not depend on the t ime of the peak velocity. The highest h 2 va lues
occur in the middle of the sequence, for hand length and s ta ture in boys and for both chest d imens ions in girls. The re is a s l ight dependence ofh 2 values on the na ture of the charac te r . T h e h 2 values are somewha t h igher for those characters represent ing more s table skeletal pa r t s (Ic, T1, Xi) and for those which a r e made up of many components (height and weight) . Fo r those charac ters represent ing moveable skeletal par ts , the h 2 values a re r a the r lower. These regular i t ies however are not very dist inct . The re is no evidence of any re la t ionsh ip of the h 2 value with the phylogenet ic age of the character , in cont ras t ~o wha t
was seen in the peak velocity ages. T o compare our results on height and weight with those from the Swedish twin ma te r i a l
(F ischbein , 1977) it is necessary first to calculate the value o fh 2 based on corre la t ions as
shown in the following formula:
rMZ -- roz hr 2 -
1 -- rDz
T h e results are shown in T a b l e 5. The h~ values in the Swedish twin mate r ia l are c lear ly lower than ours. This f inding in our opinion can only be expla ined by differences in the env i ronments . O u r twins live in the same town, that is to say they have very s imi la r l iving condi t ions , whereas the Swedish twins came from the whole of Sweden and therefore more
va r ied l iving condit ions.
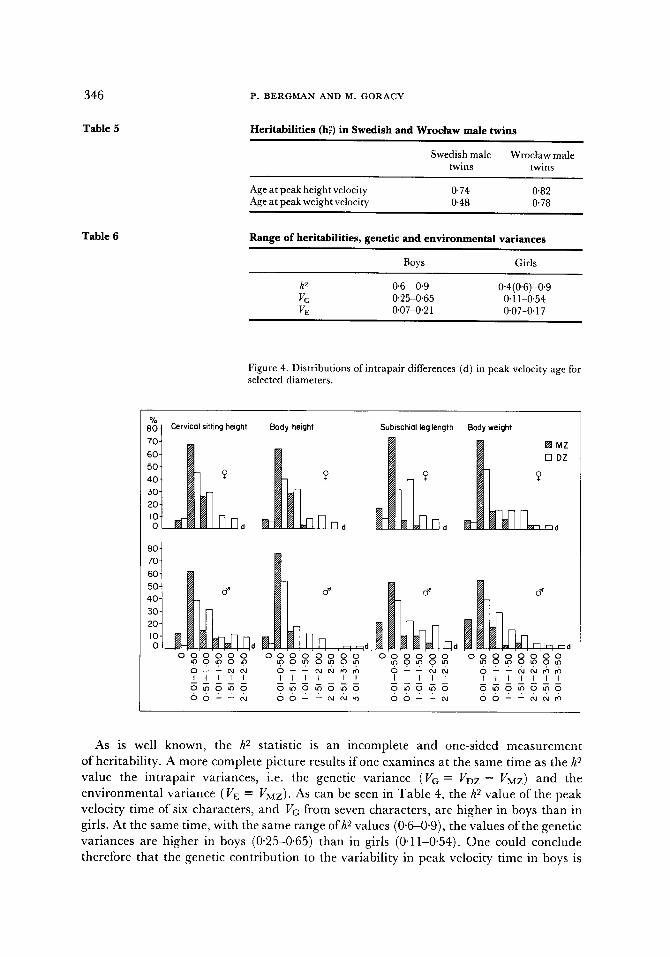
346
Table 5
Table 6
P. BERGMAN AND M. GORACY
Heritabilities (hl) in Swedish and Wrociaw male twins
Swedish male Wroctaw male twins twins
Age at peak height velocity 0-74 0'82 Age at peak weight velocity 0.48 0'78
Range of heritabilities, genetic and environmental variances
Boys Girls
h 2 0"6 -0'9 0"4(0'6)-0.9 Vc. 0"25-0.65 0" 11-0.54 VE 0"07-0'21 0-07-0-17
Figure 4. Distributions ofintrapair differences (d) in peak velocity age for selected diameters.
8• Cervical sitting height Body height Subischial leg length Body weight
60 [] DZ 5O
4o ? ? ? 30 20
50 c~ o ~ 4o 30-1
H , . . . . . . . no . . . . . . 0 0 0 0 0 0 0 0 ~ 9 ~ o ~ ~ 1 7 6 1 7 6 1 7 6 1 7 6 1 7 6 ~ 1 7 6 1 7 6 ~ 1 7 6 1 7 6 1 7 6 1 7 6
0 ~ - - 0 . 1 C ~ 0 - - - - O J 0 - - - - e , I C ~ 0 - - - -
, , , , , , , , , ? ? 7 I , , , , , , , 7 7 7 7 0 r 0 r 0 0 ~ 0 ~ 0 ~ 0 0 r 0 ~ 0 0 LL'~ 0 r 0 ~ 0
0 0 - - - - Cd 0 0 - - ~ OJ e'J fr 0 0 - - - - 04 0 0 - - - - 0 , 10a ~
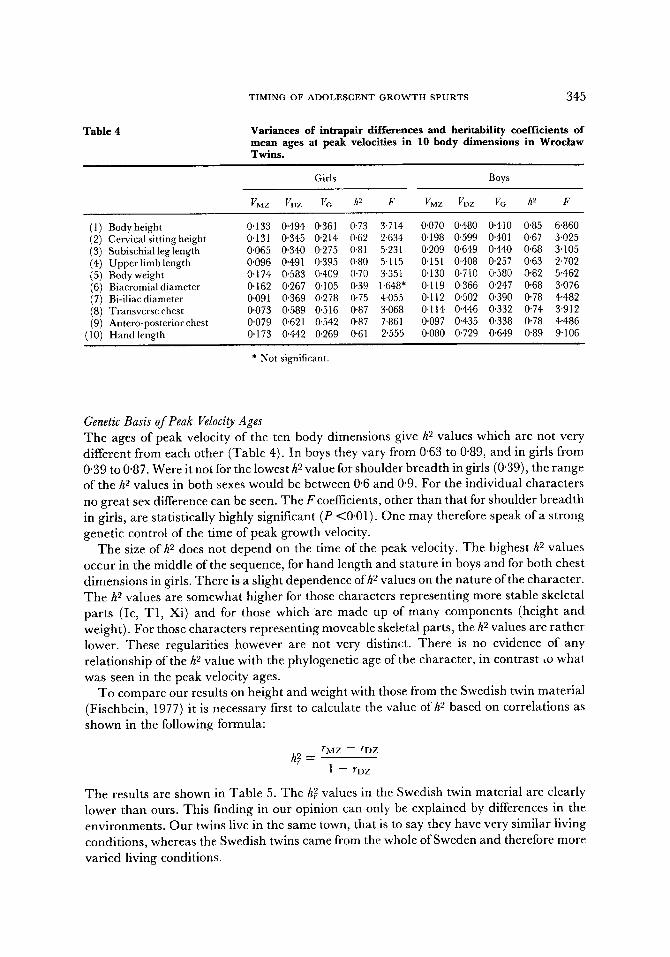
As is well known, the h 2 statistic is an incomplete and one-sided measurement of heritability. A more complete picture results if one examines at the same time as the h 9 value the intrapair variances, i.e. the genetic variance (VG = VDZ- VMZ) and the environmental variance (VE = VMZ). As can be seen in Table 4, the h 2 value of the peak velocity time of six characters, and Vc from seven characters, are higher in boys than in girls. At the same time, with the same range ofh 2 values (0"6-0"9), the values of the genetic variances are higher in boys (0'25-0.65) than in girls (0"11-0"54). One could conclude therefore that the genetic contribution to the variability in peak velocity time in boys is

TIMING OF ADOLESCENT GROWTH SPURTS 347
la rger than that in girls. Yet relat ive to the h 2 values, the env i ronmenta l var iance VE also is often h igher in boys (0"07-0'21) than in girls (0"07-0"17). Consequen t ly girls are charac te r ized not only by a lower genet ical ly de te rmined var iabi l i ty , bu t also by a lower env i ronmen ta l va r iab i l i ty (i.e. ecosensit ivi ty) , in the t ime of peak velocity than are boys. These conclusions are clear f rom Tab le 6.
F igure 4 shows the d i s t r ibu t ions of the in t rapa i r differences (d) of the age at peak veloci ty of some i m p o r t a n t charac ters in the monozygot ic and dizygot ic twins. The range of va r i a t ion of these differences is much higher in dizygot ic twins than in monozygot ic ; d izygot ic pa r tne r s vary much more often, and with a grea ter age difference, in reach ing the peak veloci ty than do the monozygot ic co-twins. The major i ty of the monozygot ic twins show an age difference of 0-0.5 years, pa r t of them from 0"5-1 year but only in some characters ; only a few pairs show a difference greater than one year. The greates t n u m b e r of d izygot ic twins however show a difference of 0"5-1'5 years and some of them even o f two years and over. These d i s t r ibu t ions of the in t r apa i r differences provide a good i l lus t ra t ion of the impl ica t ions of the da t a summar i zed in Tab le 3.
References Bergman, P., Grzesiowski, F. & Szmyd, A. (1981). Verwendung der logistischen Funktion zur quantitativen
Beschreibung des K6rperlS.ngenwachstums bei Zwillingen. Homo 32, 81-89. Bergman, P. & Szmyd, A. (1981). Growth of the hand in Wroclaw twins. Studies in Physical Anthropology 7, 47-61. Bielicki, T. & Welon, Z. (1973). The sequence of growth velocity peaks of principal body dimensions in girls.
Materiaty i Prace Antropologiczne 86, 3-10. Cameron, N., Tanner, J. M. & Whitehouse, R. H. (1982). A longitudinal analysis of the growth of limb segments
in adolescence. Annals of Human Biology 9, 211-220. Clark, P.J. (1956). The heritability of certain anthropometric characters ascertained from measurements of
twins. American Journal of Human Genetics 8, 49-54. Fischbein, S. (1977). Onset of puberty in MZ and DZ twins. Acta Geneticae Medicae et Gemellologiae 26, 151-157. Ljung, B-O., Fischbein, S. & Lindgren, G. (1977). A comparison of growth in twins and singleton controls of
matched age followed longitudinally from 10 to 18 years. Annals of Human Biology 5, 405-415. Mfiller-K6nig, T. (1977). Das Wachstum von K6rperh6he und Gewicht bei Knaben und M~idchen im Schulalter
mit besonderer Ber/icksichtlgung der sozialen Schichtung dargestellt durch Querschnitts- und Liingsschnittsuntersuchungen. Inaugural-Dissertation, UniversitS.t Kiel, 257 S.
Orczykowska-Swi~tkowska, Z. (1964). Studies on inheritance of dermatoglyphic patterns (in Polish). Materialy i Prace Antropologiczne 65, 91-122.
SkIad, M. (1977). The rate of growth and maturing of twins. Acta Geneticae Medicae et Gemellologiae 26, 221-237. Tanner,J. M., Whitehouse, R. H., Marubini, E. & Resele, L. F. (1976). The adolescent growth spurts of boys and
girls of the Harpenden Growth Study. Annals of Human Biology 3, 109-126. Vital and Health Statistics. (1973). Height and Weight of Youths 12-17 Years. United States. Series 11, No. 124. U.S.
Department of Health, Education, and Welfare. Welon, Z. & Bielicki, T. (1979). The timing of adolescent growth spurts of 8 body dimensions in boys and girls of
the Wroclraw Growth Study. Studies in Physical Anthropology 5, 75-79.