the rabbit placent as a organa n of diffusional...
TRANSCRIPT

The Rabbit Placenta as an Organof Diffusional Exchange
COMPARISON WITH OTHER SPECIES BY DIMENSIONAL ANALYSIS
By J. Job Faber, M.B., Ph.D., and Frederick M. Hart, B.S.
ABSTRACTThe umbilical arteries of intact, in-situ placentas of one series of anesthe-
tized rabbits were perfused with deoxygenated blood equilibrated with 10%or 20% acetylene and in another series with oxygenated blood without acetylene.The transfer of acetylene, and probably also of oxygen, were shown notto be limited by diffusion. Above the normal physiological rates of fetalplacental flow, oxygen uptake in the fetal placental circulation did not in-crease appreciably with an increase in the rates of perfusion; below it, itdecreased almost proportionally with a decrease in the rate of perfusion.
To explore the applicability of these results to the human placenta, theexperimental data were reduced to dimensionless form. It was shown thatthree dimensionless variables suffice to describe the characteristics of diffusionalexchange in the placenta. The dimensionless diagrams were very similar for theideal countercurrent exchanger and for the rabbit placenta but these differedfrom diagrams made from data published on human, sheep, cow, andmonkey placentas. The authors propose the dimensionless diagram as aconcise description of the passive transfer characteristics of a placenta, at leastuntil measurements of vascular geometry, regional permeabilities and flowpatterns allow the calculation of mean diffusion gradients.
ADDITIONAL KEY WORDS perfused fetal placentasoxygen transfer acetylene transfer pressure-flow curvesplacental oxygen consumption maternal placental blood flowfetal placental shunt flow anesthetized rabbits
• The behavior of the placenta as an organof oxygen exchange depends on such factorsas the rates of maternal and fetal blood flow,hemoglobin contents and half-saturation pres-sures, arterial oxygen pressures, and the oxy-gen permeability and vascular geometry of theplacenta. Many of these factors have been
From the Department of Physical Medicine andRehabilitation, University of Washington, Seattle,Washington.
This work was supported in part by a grant(HD980) from the National Institute of Child Healthand Human Development and in part by a researchand training grant (R&T 3) from the Office ofVocational Rehabilitation.
The authors' present address is Department ofPhysiology, University of Oregon Medical School,Portland, Oregon.
Parts of this paper appeared in abstract form inThe Physiologist 25(2): 189, 1965, and in the Fed-eration Proceedings 8: 107, 1965.
Accepted for publication June 30, 1966.
measured in intact animal and human preg-nancies. Very little is known about the dy-namics of placental transfer consequent to al-terations in the conditions of perfusion, exceptthat such alterations do occur and affect gastransport. In the experiments to be described,the effects of variations in the rate of umbili-cal blood flow on the transfer of oxygen andof acetylene were measured in rabbits in whichthe fetuses were removed and the fetal side ofthe placenta artificially perfused in situ.
The similarity of function between animaland human placentas leads one to supposethat the observed behavior of animal placen-tas is indicative of the behavior of the humanplacenta. By invoking the principle of simili-tude (1, 2), one can delineate the conditionsunder which this supposition is likely to betrue. The application of the principle re-quires the use of dimensionless combinations
816 CircaUtiom Rvurcb, Vol. XIX, Octohtr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

DIFFUSIONAL PLACENTAL TRANSFER 817
of the variables. The disadvantage that thesemay be hard to visualize is outweighed bythe advantage that the number of necessarydimensionless combinations is always lessthan the number of original variables (1, 2).Complete experimental investigation of thebehavior of the system when represented bydimensionless variables is correspondingly, lesslaborious. A further advantage is found in theconvenience with which theoretical modelsand empirical relations can be compared whenexpressed in nondimensional form.
TheoryASSUMPTIONS
The mathematical formulation of heat ex-change between two moving fluids separatedby a barrier is directly applicable to diffusion-al exchange, with minor modifications of themeaning assigned to the variables. Bowmanet al. (3) summarized the behavior of heatexchangers of a great variety of flow patterns.Mathematical formulations of the theoreticalbehavior of idealized placentas have beenpublished by Wilkin (4), and by Bartels andMoll (5). Bowman et al. (3) used three di-mensionless parameters to illustrate their cal-culations, as did Bartels and Moll (5). Theassumptions made in the derivation of thedifferential equations of heat transfer are, ac-cording to Bowman et al. (3):
1. The overall heat transfer coefficient isconstant throughout the exchanger. (Thecorresponding requirement in placentasis that the permeability constant is thesame everywhere.)
2. The flow rates are constant.3. The specific heats of the fluids are con-
stant. (The oxygen capacities are con-stant and the hemoglobin dissociationcurves are linear.)
4. There is no condensation or boiling ofthe liquids (no gas bubbles present).
5. There is no heat loss from the exchanger(no exchange with spaces outside theplacenta).
For exchange through the placenta, Wilkin(4) required some additional assumptions:
6. There is no diffusion in the direction of
the blood stream, either in the barrier orin the bloods.
7. There is no transport by fluid filtrationacross the barrier.
Bartels and Moll (5) required the additionalassumptions that:
8. Any subdivision of the placenta receivesthe same fraction of the maternal andfetal blood flows and has the same frac-tion of the total permeability. There areno shunts at the fetal side of the placen-ta.
9. The placenta does not metabolize thesubstance under consideration.
These assumptions, except no. 3, are also im-plied in the calculation of the average gradi-ent across the placenta by the Bohr integra-tion (6-8).
It is useful for further calculations to rep-resent the assumption that the oxygen-hemo-globin dissociation curve is linear by the equa-tion
where C is the oxygen content of the bloodin moles per milliliter; [O2] is the concentra-tion of oxygen in physical solution (moles permilliliter) and / is a number. The oxygen con-tent of blood is also given by the relationship
C = Hb-H-Sat+[O2], (2)
where Hb is the hemoglobin content (gramsper milliliter), H is the Hiiffner coefficient(moles of oxygen bound per gram of fullysaturated hemoglobin) and Sat is the frac-tional saturation of hemoglobin; [O2] will beneglected in this equation. Combination ofequations 1 and 2 gives
f = Hb.H.Sat/[O2]. (3)
At half saturation ([O2] = [O2]*; Sat = 0.5),equation 3 gives
f = Hb.H/2[O2U, (4)
where [On]* is the concentration of oxygenin physical solution at which the hemoglobinis half saturated with oxygen. The number /is the ratio of content and concentration evalu-ated at half saturation. For substances that are
CircuUtio* Reitrcb, Vol. XIX, Octobtr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

818 FABER, HART
present in solution only and do not react withblood, / is equal to 1. This definition of / willbe used in subsequent calculation.
CHOICE OF DIMENSIONLESS VARIABLES
According to Buckingham's rule (1), the
were chosen. A "transport fraction," T, is givenby
T= ( [* ]*• - [* ] " • ) / ( [* ]*" - [* ] " ) • (5)
For oxygen, the definition of T will be:
To., = (Sat*" - Sat1'")[(>,] w"I(Sat*"[O]y*- SatF°[O2]VlF). (6)
number of dimensionless variables to be usedfor a complete description usually equals thenumber of elementary variables minus thenumber of dimensions occurring among them.1
In equations similar to those derived by Wil-kin (4) and Barrels and Moll (5), which areanalogous to the equations used as a firstapproximation in the calculation of heat trans-fer in heat exchangers (3), the elementaryvariables entering into the problem are thematernal and fetal placental flow rates(Qu and QF milliliters per second respective-ly), the constants fF and fM, denned by equa-tion 4, the arterial and venous concentra-tions of oxygen in physical solution ([O2]
Ma,[O2]
Fa, [O2]M°, and [O2]
F* moles per milli-liter) and the placental permeability to oxy-gen (p milliliters per second). The superscriptsM and F stand for maternal and fetal respec-tively, and a and v for the anatomic arterialand venous ends of the placental capillariesrespectively.
In the above-mentioned equations, thethe variables Q and f always occur as a prod-uct {QMf and QFf)- The equations can berewritten in a form in which the oxygen con-centrations occur only in the combinations( [ O 2 ] ^ - [ O 2 ] " ° ) and ( [ O 2 ] ^ - [ O 2 ] ^ ) .With these four variables and the placentalpermeability, p, there are only five elementaryvariables and two dimensions (length andtime) so that three dimensionless variablesmust be chosen.
The following combinations of variables inwhich the physical meaning is relatively clear
lrThis rule has exceptions. A general theorem canbe found in reference 2.
In equations 5 and 6, O < T < 1.It follows from equations 3 and 4 that [O2]2. Sat- [OL,]v4. Therefore, TV, could be de-fined for oxygen in the same way in which itis defined for the substance x. But the defini-tion of 7V, as given for oxygen, eliminatesthe adverse effects on the accuracy of the cal-culation of very high oxygen pressures in thematernal blood. In the experiments to be de-scribed, in which fairly high maternal oxygenpressures were used, the arterial oxygencontent exceeded the oxygen capacity of theblood by about 10%. This will be taken intoaccount. In experiments in which animals areventilated with room air, the percentage willbe so small as to be negligible.
The choice of T is based on the considera-tion that the concentration of a substance inthe maternal circulation diffusing from thematernal to the fetal blood cannot drop belowthe concentration in the fetal artery. Thedenominator of T represents the maximumpossible reduction in concentration in thematernal flow, and the numerator the actualreduction. Thus, T equals the reduction inconcentration in the maternal blood streamexpressed as a fraction of the maximum pos-sible reduction.
A "flow ratio," R, is denned by the relation
R = fQ"if(^t (O<R). (7)
R is determined by the rates of blood flow, thehemoglobin concentrations, and the half satu-ration pressures: it represents the ability ofthe fetal circulation to "carry away" dividedby the ability of the maternal circulation to"deliver" at any given oxygen concentration.For substances that are present in physicalsolution only, R is the ratio of the fetal andmaternal rates of blood flow.
CircmUiion Rtitmrcb, Vol. XIX, Octoitr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

DIFFUSIONAL PLACENTAL TRANSFER 819
One expects the third dimensionless variableto contain the placental permeability, p. Theamount of oxygen diffusing across the placen-ta per unit time is equal to the placental per-meability to oxygen, p, multiplied by theaverage oxygen gradient across the placentalbarrier, which will be represented byA[Oo]. The oxygen diffusing across the pla-centa per unit time is taken from the maternalblood stream, and therefore is equal to (CMa —
moles/sec. Thus: 2[O2]v4Jf(Sfl*ifo- SatMv) •
]viJ'(Sat3"1 - S^ J
= pl(t*Q*). (8)Therefore, a third dimensionless variable, the"permeability variable," d, is defined by
d = p/(Q><f') (O<d). (9)
The number d is the ratio of placental perme-ability and the rate of maternal flow for thesubstance under consideration.
In those placentas in which A[C>2] can beevaluated by a modified Bohr integration (6-8),d is by definition related to the Bohr integral.
There is a certain degree of freedom in thechoice of dimensionless variables. Engineeringpractice (3) makes use of variables equiva-lent to T=([x]i><1- [x]Mv)/([x]Ma~ [x]F°),T' = ([x]Fv- [x]Fa)I([X]"" - [*]*•),andd' =([i]37"-[x]"°)/A[xJ. Bartels and Moll (5)used T" as defined above, d" ~*pl (f*'QF), andR' = fQu/(fQ") to illustrate the theoreticalbehavior of a number of ideal exchangers.They graphed T" versus d" at various valuesof fl'. This practice is adequate to depict thecomputed behavior of models, but not to de-pict experimental results, since usually in thelatter case the value of d" is not known.
The choice of the set T, R, and d is some-what arbitrary. From another complete set,one could calculate T, R, and d and viceversa. However, it is convenient to considerthe maternal placental blood flow as the pri-mary determinant of fetal life. Thus, a trans-port fraction was chosen as defined above bythe fractional clearance of the maternal blood
stream, the permeability variable as the pla-cental permeability per unit maternal rate offlow, and the flow ratio as the ratio of fetaland maternal flows.
The variables T, R, and d can replace thevariables that are contained in them: p, f,fM> QF, Qu, SatUa, Sat11", SatFa. A relationshipbetween T, R, and d, whether derived theo-retically or empirically, contains all the infor-mation regarding the behavior of the placentaas an organ of exchange that is contained inthe corresponding relationships between theoriginal variables. For graphical representationT and R will be plotted on ordinate andabscissa, and the appropriate value of d willbe indicated, if known. These graphs will bereferred to as T-R diagrams.
THE T-R DIAGRAM OF IDEAL EXCHANGERS
Four simple vascular structures were con-sidered. Placentas with parallel fetal and ma-ternal vessels are common in rodents. The onesin which the fetal and maternal flows are inopposite directions will be referred to as coun-tercurrent placentas. These will be comparedwith exchangers in which the flows are in thesame direction or concurrent. Exchangers inwhich the fetal blood is contained in vesselsthat are suspended in a well mixed pool of thematernal blood are called pool flow placentas;this may be the appropriate model of a hu-man placenta. Since in many placentas, thevascular arrangement is almost random, thecross-current exchanger in which fetal andmaternal vessels cross at right angles will alsobe considered. The appropriate transport equa-tions for these exchangers, which can befound in the literature (3-5, 9), have beenconverted into dimensionless form and plot-ted in Figure 1. T was calculated at values ofR between 0.2 and 6 and of d between 0.03and 30.
The pool flow exchanger is very inefficientbecause fetal blood equilibrates against ma-ternal venous blood. An even more inefficientexchanger is conceivable if one assumes adouble pool flow exchanger in which bothmaternal and fetal compartments are thorough-ly stirred and the diffusional gradient is thatbetween maternal venous and fetal venous
OrcmUiian Rtiureb, Vol. XIX, October 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

820 FABER, HART
a '3o//
'L̂—
d
d
.
d
d
d
. 1
• .3
..1
•J33
_ _ —
cross curor t
— :
i i
0 2 1
FIGURE 1
The T-R diagrams of four ideal exchangers. T = transport fraction; R = flow ratio; d = perme-ability variable.
blood. The T-R diagram of this hypotheticalplacenta showed that a double pool flow pla-centa is only a few percent less efficientthan a pool flow placenta; its T-R diagram istherefore not shown inTigure 1.
Figure 1 demonstrates that at values forthe permeability variable d below 0.3, thegraphs are nearly identical; apparently theanatomical arrangement, at least in these foursimplified cases, becomes largely irrelevant ifthe placenta is very impermeable to the sub-stance under consideration, d<0.3. Figure 1further illustrates that values of d larger than10 are almost without additional effect on thetransport fraction T. In very permeable pla-centas a further increase in permeability doesnot increase transfer.
A comparison between the concurrent andpool flow T-R diagrams shows a nearly iden-
tical behavior of these two types of exchangersin terms of diffusional effectiveness. A similarsituation exists between the counterciuTentand the cross-current diagrams. Thus, ana-tomical information is needed to discriminatebetween two functionally similar types of pla-centas.
INTERPRETATION OF THE T-R DIAGRAMS
The rate of oxygen transfer from mother tofetus, 02, in moles per second is given by:02 = (Cu° - C"»)Q* = (Sat"* - Sat*") • Qu •Hbu • H. By using equation 4, and the defini-tion of T (equation 6), one finds:
(10)
The rate of oxygen transfer is proportionalto the difference between maternal arterial
CircuUtKm Ktiurcb, Vol. XIX, Oaobtr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

DIFFUSIONAL PLACENTAL TRANSFER 821
and fetal arterial concentrations, to T, and tothe maternal placental flow rate. These rela-tionships are the basis of the interpretation ofthe T-R diagrams.
MethodsExperimental perfusion of the fetal side of the
placenta allowed adjustment of the fetal rate ofblood flow, adjustment of the gaseous composi-tion of fetal arterial blood, and monitoring ofoxygen tension and acetylene concentrations infetal placental inflow and outflow. The fetus wasremoved from the placenta, which remained insitu, and the fetal half of the placenta was per-fused with blood pumped from a disc tonometerin which it was equilibrated against a gas ofknown composition. Oxygen uptake and acetyleneloss were calculated from the arteriovenous differ-ences in oxygen and acetylene contents and theflow rate of perfusion.
Placentas of 30 New Zealand white rabbits ata gestational age of 27 to 29 days were success-fully perfused. Term in rabbit is 30 to 31 days.Excluded from the study were those perfusionsin which leaks developed in excess of 5% of theperfusion rate, which showed a steadily risingarterial pressure at constant rate of perfusion(due to insufficient filtering of the inflowingblood), or in which we failed to cannulate bothumbilical arteries and umbilical vein. The up-take of oxygen was measured in 20 rabbits; theloss of acetylene was measured in 7 of these.Placental oxygen consumptions were also mea-sured in placentas, none of which had been ex-posed to acetylene. Umbilical arterial blood pres-sure, venous blood pressure, and rates of flow weremeasured in 5, 8, and 6 fetuses, respectively.Umbilical arterial and venous oxygen pressureswere measured in 4 and 5 fetuses respectively;uterine venous oxygen pressure and maternal ar-terial oxygen pressure were each measured in 2animals; the latter was measured 13 times atdifferent rates and depths of artificial ventilation.Twenty-three of the animals were pretreatedwith oral reserpine for 1 or 2 days (0.3 mg/kgbody weight per day) in the expectation thiswould reduce vascular reactivity. The last 7perfusions performed without reserpine pretreat-ment showed that the treatment did not affectoxygen transfer nor appreciably reduce vascularreactivity.
Anesthesia (pentobarbital 30 mg/kg iv) wasgiven immediately before the experiment andmaintained during the experiment by additionalinjections. The animal was hyperventilated by arespirator pump with a gas mixture of 5% carbondioxide in oxygen to insure that the arterial oxy-gen saturation was 10035. The abdomen was
CircnUtum Kutrcb, Vol. XIX, Oaokt 1966
opened and the animal was placed in a bathfilled with saline solution at 39.1°C, the normalbody temperature of a pregnant rabbit (10).The composition of the saline solution was ap-proximately that of mammalian extracellular fluidexcept that bicarbonate ion was replaced bychloride ion.
Two 5-cm squares of gauze were glued withEastman 910 adhesive to the uterus. To preventcontraction of the uterus under the placenta, theuterus was slightly stretched and the gauze at-tached to die abdominal wall. The uterus wasopened and the fetal membranes were clippedaway. The umbilical vessels were bathed with asolution of 1 mg of nitroglycerine in 10 ml ofisotonic saline, which produced visible vasodila-tion. The dilated vessels were fixed with a solu-tion of 10% formaldehyde, which was drippedon the vessels and carefully washed off afterapplication. After this procedure, the umbilicalarteries were still expected to show good pulsa-tions.
The following experiments were made on intactfetuses. The hydrostatic pressures in the umbili-cal vessels were measured with Statham pres-sure gauges, which were connected to a thin tygontube filled with saline and with a 25-gauge needlemounted at the end. Zero pressures were setwith the needle at the level of the placenta. Theneedles were inserted in the upstream directionand the pressures were recorded on the poly-graph. Umbilical rates of blood flow were re-corded by insertion of an 18-gauge needle at-tached to a tuberculin syringe without plungerinto the umbilical vein toward die placenta. Col-lections of 0.2 ml of blood in the syringe weretimed. All blood from the placenta flowed into thesyringe, since the vessel fitted snugly around theneedle. Visual checks were made to ascertainthat no blood escaped past the needle into thefetus. The normal venous pressure in this vesselis about 6 mm Hg. The resistance to flow of theneedle and syringe was about 4 mm Hg whenblood flow was 2 ml/min, and the level of dieblood in the syringe was about 2 to 3 cm abovethe level of the placenta. The disturbance ofnormal venous pressure was therefore negligible.In a few fetuses, oxygen tensions were measuredin die umbilical vessels witii microelectrodes(Beckman #315780) inserted into the vessels.The electrodes totally obstructed the blood flow.However, as their 95$ response time is about 15sec, it appears unlikely that die measured oxygentensions were different from the real ones. Wesucceeded only twice in obtaining a measurementof the oxygen tension in the uterine vein. Theelectrodes tend to tear diese extremely diin-walled vessels and to produce large hemorrhages.Several attempts to cannulate tiiese veins directly
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

822 FABER, HART
or via the inferior caval vein failed. During theplacental perfusion experiments, no samples couldbe obtained at all since the gauze that held theuterus in position obstructed access to the uterineveins.
In the placental perfusion experiments, the fol-lowing procedures were used. The umbilical ves-sels were cannulated with sharp cut-off hypoder-mic needles mounted in thin tygon tubing. Theumbilical arteries accommodated 20-gaugeneedles, and the vein an 18-gauge needle. Thetwo arterial cannulations were performed whilea flow rate of less than 0.5 ml of blood/ min wasmaintained.2 (If venous pressure rose aboveabout 15 mm Hg, leaks developed between thefetal and maternal circulations.) The fetus wasremoved from the preparation after cannulationof the umbilical vein. The placenta was at thelevel of the fluid in the animal bath and keptmoist with warm saline. In experiments in whichtransfer of acetylene was measured, the placentalsurface was covered by a layer of white petroleumjelly since without this precaution more than10% of the acetylene was lost through the ex-posed surface of the placenta.
In order to measure placental oxygen consump-tion after termination of the perfusion experi-ments, the maternal circulation to the placentawas occluded by a hemostat placed on the largeligament under the placenta. The effectiveness ofthis procedure was immediately apparent becauseall oxygen from the umbilical venous outflowdisappeared. Oxygen was then admitted to thedisc tonometer and oxygen tension arteriovenousdifference across the placenta was measured.
-An initial perfusion with saline to wash out allfetal red cells destroyed the preparation and wastherefore discontinued. The washed preparationsfailed to show any oxygen uptake.
Oxygen tensions were converted to oxygen con-tents and multiplied by the rate of umbilicalblood flow to arrive at a value for the oxygenconsumption of the placenta. The measurementwas usually repeated at different rates of bloodflow.
PERFUSION CIRCUIT
The perfusion circuit consisted of a disc tonom-eter, in which the blood was equilibrated with agas of known composition, a constant flow pump(Sigmamotor model T8) and two electrode cu-vettes in which the oxygen and carbon dioxidetensions and hydrostatic pressures were measuredin the blood entering and leaving the placenta(Fig. 2).
The circuit was filled with 100 ml of heparin-ized adult rabbit blood. The disc tonometer equil-ibrated the blood at room temperature with agas mixture consisting of approximately 5% carbondioxide in nitrogen saturated with water vapor.In some experiments 10 or 20% of the mixture wasreplaced by acetylene. Equilibration at room tem-perature reduced the rate at which the tonome-ter hemolyzed the red cells, and at 39.1°C pro-duced a carbon dioxide pressure of about 40 to 50mm Hg. The tonometer had sufficient capacityto keep the blood entering the placenta deoxygen-ated and at a constant carbon dioxide pressure.The blood was pumped through a cotton woolfilter from the tonometer into the arterial elec-trode cuvette.11 A coil made of 120 cm of thin-walled stainless steel tubing was submerged inthe animal bath between the filter and the elec-trode cuvette in order to bring the temperature ofthe inflowing blood to 39.1°C, the temperatureat which the oxygen and carbon dioxide pressures
:iWithout this filter, the placental perfusion pres-sures at constant flow were abnormally high.
forcedventilationof animalwith mixtureof 5%COjand 95% O
constanttemperature
bath
filter
6constant flow pump
gas mixingpump
FIGURE 1
Diagram of the perfusion circuit viewed from above. Not shown are the electrodes and pressuregauges which are located in the electrode cuvettes, the recording equipment, and the rabbit.
CircuUltca Rtsurcb, Vol. XIX, October 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

DIFFUSIONAL PLACENTAL TRANSFER 823
of the blood were measured. Oxygen tensionswere measured with Beckman Spinco macroelec-trodes (type 315790). Carbon dioxide tensionswere measured with National Welding (type9987100) electrodes. Tygon tubing with hypo-dermic needles at one end conducted the bloodbetween the electrode cuvettes and the placen-ta. The blood returning from the placenta drainedinto a graduated cylinder outflow meter on theside of the tonometer. A stopcock between theflowmeter and the tonometer allowed timing ofcollections of blood. The inflow rate of the sigma-motor pump was recorded by a microswitch onthe drive shaft of the pump connected to acardiotachometer. The cardiotachometer readingwas calibrated against the venous outflow meterbefore the experiment. The accuracy of this mea-surement was about 5%. The hydrostatic pressureof the blood flowing to and from the placentaswas measured with Statham gauges. Zero pressurelevel was taken at the level of the placenta. Aftereach experiment, the placenta was removed andthe arterial and venous needles connected to-gether. The resistance to flow was determined inthis "short circuit" at various flows, and latersubtracted from the total resistance to flow ofthe circuit and the placenta together to obtainthe placental resistance.
All measurements were recorded on an ink-writing Beckman-Offner Dynograph, except pH,which was measured at 39.1°C in blood samplestaken from the circuit at regular intervals and in-jected into a Spinco Model 160 pH meter, andacetylene concentrations, which were measuredin small (5 /iliter) samples taken at regularintervals and injected into a Wilkens 600 D gaschromatograph. Accuracy of the acetylene deter-mination was ± 5%. The venous acetylene con-centrations were expressed as percentages of thearterial acetylene concentrations.
The electrodes were calibrated before and aftereach experiment by pumping saline through thecircuit from containers in which it had beenequilibrated at 39.1°C with calibration gasesfrom premixed tanks (usual composition about0% O2, 8% CO;,, and 10% O2, 2% CO2, in nitrogen).Experimental accuracies were ± 2 mm Po2 and± 2 mm Pco2.
CONVERSION OF OXYGEN TENSIONINTO OXYGEN CONTENTS
Oxygen saturations were determined from therecorded oxygen pressures by reference to theoxygen dissociation curve of blood of New Zea-land white rabbits (7). Since the position of thedissociation curve depends on the pH and tem-perature of the blood, the observed oxygen ten-sions were corrected for pH and temperature(11). According to Bartels and Harms (12) thecorrection factors for human blood (11) alsoCircuUiton RuMrcb, Vol. XIX, Oaobtr 1966
apply to rabbit blood. Rabbits normally have anaverage hematocrit of 41.5% and a hemoglobincontent of 11.9 g/100 ml of blood (13), i.e.,about 0.287 g of hemoglobin/100 ml per 1%hematocrit. Assuming the Hiiffner coefficient(1.34 ml of oxygen capacity/g of hemoglobin) tobe valid for rabbit hemoglobin also, we arrivedat the formula that the oxygen capacity is 0.38volume percent per percent hematocrit (Hcrt)and that the oxygen content equals Sat-Hcrt-38ml of oxygen STPD/100 ml of blood. The oxygencontents were adjusted for the amount of dis-solved oxygen by addition of Po2 . 0.00296 (mlof oxygen dissolved/100 ml of blood).
Results and CalculationsOXYGEN UPTAKE BY INTACT FETUSES
Some preliminary experiments were per-formed to establish the normal and maximaloxygen transfers to intact rabbit fetuses of 27to 29 days gestation. These measurementsserved as standards against which the resultsof the placental perfusions were compared.
The results of these measurements areshown in Table 1. The measured oxygen ten-sions agreed fairly well with those Barron andBattaglia calculated from measured oxygensaturations and the oxygen-hemoglobin disso-ciation curve of fetal rabbit blood. The tworecorded values of the oxygen tension in theuterine vein were somewhat lower than thoseobtained by Barron and Battaglia (Table 1).Since we used the mean of the latter figures inthe calculation of the maximum oxygen de-livery by the maternal blood stream, we haveset the criteria for the intactness of the per-fusion preparations too high rather than toolow, as will be explained later.
From the umbilical flow rates in millilitersper minute per kilogram of fetal weight re-corded in Table 1 (SEM 14.5%) and theaverage arteriovenous difference in oxygencontents published by Barron and Battaglia(7) (SEM 8%), it could be calculated that ata gestational age of 27 to 29 days the aver-age oxygen uptake of a rabbit fetus is 5.0ml/min per kg of fetal weight.
OXYGEN CONSUMPTION OF THE PLACENTA
The placental oxygen consumptions mea-sured in the perfusion experiments in whichthe maternal circulation was occluded are tab-
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

824 FABER, HART
TABLE 1
Oxygen Uptake and Umbilical Hemodynamics in 27 to 29 Day Rabbit Fetuses (Mean ±1 SEM)
Fetal (umbilical)artery
Fetal (umbilical)
Maternalartery
Maternal (uterine)vein
Fetal arterymean blood pressure
Fetal vein
blood pressure
Umbilical blood flow
Average fetal weightUmbilical blood flow per kg
fetal weight
Po2
0 2 SatO~ Content
Poo
O2~SatO., Content
Po2
O2 SatO., Content
Po2
O2 SatO2 Content
From
15.720.2
reference 7
±0.6± 1.4
2.83 ± 0.2
42.881.911.3
88.394.513.3
22.826.2
3.7
± 1.9± 1.0±0.8
±2.8±0.9±1.2
±1.4±2.5±0.46
Our
19 :
42.6 :
220-
dau± 1
±5.2
•615*
17.0 ±3.0
15.8 :±0.9
mm Hg%
ml O2/100
mm Hg%
ml CX./100
mm Hg%
ml 0^/100
mm Hg%
ml OV100
mm Hg
ml
ml
ml
ml
blood
blood
blood
blood
6.3 ± 1 . 3 mm Hg
2.03 ± 0.43 ml/min
39.0 ±2 .7 g
59 ± 8.6 ml/min per kg
*This pressure depends on the degTee to which the animal "fights" the respiration pump and inhales roomair. A value of 500 mm Hg is a representative average for the experiments (rabbits breathing 5% CO2-95% O2).
Po2 = partial pressure of oxygen; Sat = percentage saturation of hemoglobin with oxygen.
TABLE 2
Oxygen Consumption of the Perfused in Situ Placenta
Gestationdays
28
28
27
27
27
27
28
MeanSEM
Oxygenartery
mm
16215212712113326013210516916010910079
10599
tensionvein
Hg
6130394449595548
105887456356758
Flowml/min
1.030.510.671.150.972.101.700.904.152.285.352.331.022.651.26
Oxygen
perplacenta
0.0610.0620.0620.0920.0700.1450.0510.0390.0430.0380.0730.0740.0690.0490.037
0.0650.007
consumption
per k£ fetalweight
1.61.61.92.92.45.01.41.11.31.22.22.32.11.71.4
2.00.25
of the placentaper 100 g
wet weightof placenta
0.780.801.522.251.122.310.540.410.960.861.101.121.040.740.55
1.070.15
(ml/min)per 100 g
dry weightof placenta
5.85.98.9
13.17.5
15.53.12.46.55.87.57.67.14.33.3
7.00.9
Circulation Rutrtb, Vol. XIX, Oaokn 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

DIFFUSIONAL PLACENTAL TRANSFER 825
ulated in Table 2. The oxygen consumptionper 100 g of tissue of the rabbit placenta insitu (1.07, SEM 0.15 ml/min) is about twotimes higher than that of the human placentain vitro (14-16). It has been shown, how-ever, that the oxygen consumption of mincedtissues may be abnormally low (17), so thatno conclusions may be drawn from this differ-ence between rabbit and human placentas.
The average rabbit placenta consumesabout 2.0 ml of oxygen/kg of its fetus. Sincethe equivalent figure for the fetus is 5.0 ml,the total oxygen consumption of the conceptusis about 7.0 ml of oxygen/min per kg of fetus.
This oxygen consumption is achieved by adecrease in saturation from 95% in the mater-nal arterial blood to an average of 26% in theuterine vein (Table 1). In the experiments tobe reported, the average maternal arterial oxy-gen tension was about 500 mm Hg. At theaverage maternal hematocrit of 39%, theamount of free oxygen dissolved at this pres-sure (1.48 ml/100 ml of blood) is just equal tothe amount of oxygen necessary to increasethe saturation by 10%. It is convenient, there-fore, to assume a maternal arterial saturationof 110* in all our experiments, to allow for theamounts of dissolved oxygen.
If the maternal blood saturation were de-creased from 110% to 0$ during one passagethrough the placenta, instead of from 95% to
the usual 26%, the amount delivered would be7.0.110/(95-26) = 11.2 ml of oxygen/minper kg of fetus. The placenta takes about 2.0ml, so that about 9 ml would remain avail-able. This is probably a slight overestimate,for the reason that the actual oxygen satura-tion in the uterine vein may be somewhatlower than the 26% assumed. Conversely, ifunder certain conditions of rate of perfusionthe preparation yields the maximum of about9 ml of oxygen/min per kg of fetus, one mayassume that the oxygen tension and satura-tion in the uterine vein are very nearly zero.
According to these calculations, the pla-centa appears to consume more than 18% ofall oxygen delivered to the placenta by thematernal blood stream. In the computationmade later in this paper, the figure of 20%is used to allow for a slight overestimate inthe figure of 11.2 ml of oxygen/min per kg offetus calculated above.
TRANSFER OF OXYGEN ACROSS THE PLACENTA
The 20 perfusion experiments, from whichoxygen recoveries were computed, were donewith completely deoxygenated rabbit blood.The oxygen tensions in the umbilical veinshowed large spontaneous fluctuations (Fig.3). These were almost certainly due to varia-tions in maternal placental blood flow, sincethey were absent when the fetal circulationwas perfused with oxygenated blood after oc-
4 0 50 60 7020 30
Minutes
FIGURE 3
Spontaneous fluctuations in the oxygen pressure in the umbilical vein at various rates ofumbilical blood flow. Oxygen tension of the blood in the umbilical artery was zero.
OrcuUiitm Rtstarch. Vol. XIX, Octobtr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

826 FABER, HART
elusion of the maternal circulation. Oxygentensions were recorded for periods that wellexceeded the time necessary to establish anew steady state after a change in flow rate,i.e., about 20 min or longer. The oxygen ten-sions measured during the latter part of theseperiods were averaged.
The measurements on intact fetuses (Table1) showed that at flow rates of about 2 ml/min, the driving blood pressure (A-V) is about15.8 - 6.3 = 9.5 mm Hg. The graph in Figure4 shows that the perfused placentas demon-strated a driving pressure at a flow of 2 ml/minof 13.5 mm Hg (SEM 1.8). Precise pressuremeasurements were not made in all perfu-sions. By this criterion, at least the major por-tion of the placental vascular bed appearedto be perfused. Visual inspection of the pla-centa confirmed this. The maximum rate ofoxygen recovery from the preparations ap-proached the 9 ml of oxygen/min per kg offetus calculated above (Fig. 5). This furthersuggested that the placentas were intact.
To calculate the values of T and R, the oxy-gen tensions or saturations in the uterine veins
50
40
30
20
10
0
Umbilical Driving Pressure,
Blood Perfusions, with
" Cotton Wool Filter
I I80 2 4 6
Umbilical Flow, ml /min
FIGURE 4
Umbilical arteriovenous blood pressure difference atvarious rates of flow.
Flow. ml /mm
FIGURE 5
Oxygen uptake in milliliters per minute per kilogramfetus at various rates of umbilical perfusion. Oxygentension in the umbilical artery blood was zero.
had to be known. As these could not be mea-sured directly, an indirect approach was used.Mossman (18) established histologically thatthe parallel maternal and fetal vessels of therabbit placenta are arranged as a counter-current exchanger. This is fully compatiblewith the physiological measurements of Bar-ron and Battaglia (7), which showed thatthe oxygen tensions in the umbilical vein farexceed those in the uterine vein. In such ex-changers, the transfer of oxygen from the ma-ternal to the fetal blood can be complete, pro-vided that:
1. The hemoglobin flow rate (in grams perminute) through the fetal vessels exceedsthe hemoglobin flow rate through the ma-ternal vessels.
2. The oxygen tension in the fetal arterialblood is zero.
3. No measurable diffusional limitations ex-ist to the transfer of oxygen in the pla-centa.
A rough calculation indicated that the firstcondition is likely to be satisfied if the rateof fetal blood flow is greater than 3 ml/min.The second condition was satisfied in our ex-periments, and the third will be shown to beapproximately correct by a comparison of the
Circulation Rtstarcb, Vol. XIX, Oaohtr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

DIFFUSIONAL PLACENTAL TRANSFER 827
loss of acetylene from the fetal blood to thegain of oxygen.
Acetylene is much more easily transferredin a placenta than oxygen because it is notbound to hemoglobin. No stores of acetyleneare available except the amounts free in solu-tion. In terms of the analysis presented at thebeginning, the / of oxygen is in the order of100 and the / of acetylene is 1. Therefore,do., is about one one-hundredth of daret (seeequations 4 and 9). We will calculate one ap-parent maternal placental blood flow (QoJ")based on the assumption that the oxygen ten-sion in the uterine vein is zero at flow ratesof perfusion greater than 3 ml/min and thatthe saturation in the uterine artery is 1.1. Thus:Qo.?' = 1.25 (rate of O± uptake by perfusion)I02 content*".The factor 1.25 takes into account that one-fifth of the oxygen delivered to the placentais consumed by the placental tissues. A secondapparent maternal blood flow will be calculat-ed from the rate at which acetylene is lostfrom the perfusing blood, based on the as-sumption that the acetylene concentration inthe uterine vein is equal to the one in the fetalartery at perfusion rates greater than 3ml/min. Thus: Qacetir = percent loss of acety-lene from perfusion times rate of perfusion.If there are any measurable diffusional limi-tations to the transfer of acetylene, the rate oftransfer of acetylene will far exceed that rateof transfer of oxygen; and Qo2
i! will be morethan 10 times less than Qarr/'- If the two ap-parent flows can be shown to be equal, it maybe assumed that diffusional limitations do notexist either for oxygen or for acetylene, thatthe apparent blood flows are real blood flowsand that the oxygen tension in the uterinevein is very nearly zero at perfusion rates inexcess of 3 ml/min.
In 14 measurements in six preparations inwhich the rates of perfusion were greater than3 ml/min (range 3.0 to 10.4 ml/min) theaverage Qo.2
M was 3.15 ml/min and the aver-age Qarct31 was 3.23 ml/min. The SE of themean difference of 0.08 ml/min was 0.20 ml/min and the probability that the entire differ-ence was due to random errors was 70$. There
CircuUtum RtsMrcb, Vol. XIX, Oaokt, 1966
is more than 95$ confidence that Qu..11 is notmore than 13$ less than Qactt*1- The apparentmaternal blood flow calculated from the lossof acetylene is therefore the real maternalblood flow, and the apparent maternal bloodflow calculated from the oxygen uptake is un-likely to underestimate the real maternal bloodflow by more than 13$, if at all.
The average rate of maternal placentalblood flow calculated from the results of allperfusions in which oxygen uptakes were mea-sured at perfusion rates greater than 3 ml/min was 2.38, SEM 0.17 ml/min per placenta.The difference between the figure for thewhole group and that for the perfusions inwhich acetylene was used is mostly due to adifference in size of fetuses and placentas inthe two groups. The maternal blood flows ex-pressed per kilogram of fetal weight were 64and 72 ml/min per kg, respectively. Thesefigures represent the rates of the blood flowthat exchanges with fetal vessels or providesthe placental tissues. Any shunt flows that mayexist in the maternal circulation are not in-cluded.
At very low rates of perfusion (1.05 to 2.45ml/min), the average acetylene loss duringone passage through the placenta was 86$.About 14$ of the perfused blood appears tohave been shunted, if all acetylene was re-moved from the other 86$, which is a reason-able assumption at these low rates of flow. Insheep placentas perfused with plasma, anaverage fetal shunt of 19$ has been demon-strated (19) by a different method.
EVALUATION OF T AND R
According to the definition of R in equation7, the maternal and fetal blood flows and thefactors / (equation 4) need to be known tocalculate R. The average maternal placentalblood flow was calculated for each experi-ment from the maternal blood flowsQo2
u, defined above, measured at perfusionrates in excess of 3 mm/min. This averageflow was divided by 1.25 to remove the frac-tion of the maternal blood flow satisfying theoxygen requirements of the placental tissuesfrom the calculation of H. The factors f andf" were calculated from the hematocrits and
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

828 FABER, HART
the half-saturation concentrations, which wereobtained from the rabbit oxygen-hemoglobindissociation curve (7), after corrections hadbeen applied for the pH of fetal and maternalbloods and for temperature. It was assumedthat maternal rate of blood flow was the sameat rates of perfusion below 3 ml/min.
According to equation 10, T is proportion-al to the rate of oxygen uptake, provided thatthe oxygen saturation of the bloods in thematernal and fetal arteries do not change andthat the rate of maternal placental blood flowdoes not change. The latter condition hasalready been formulated, and the former wassatisfied by the experimental methods. At theaverage rate of oxygen uptake recorded at theperfusion rates in excess of 3 ml/min in anyone experiment, T was assumed to be 1 sinceat these flows all oxygen is removed from thematernal blood stream (SatUv = SatPa = O);therefore, T = 1 according to its definition(equation 6). At other recorded rates of oxy-gen uptake T was taken to be proportionallydifferent.
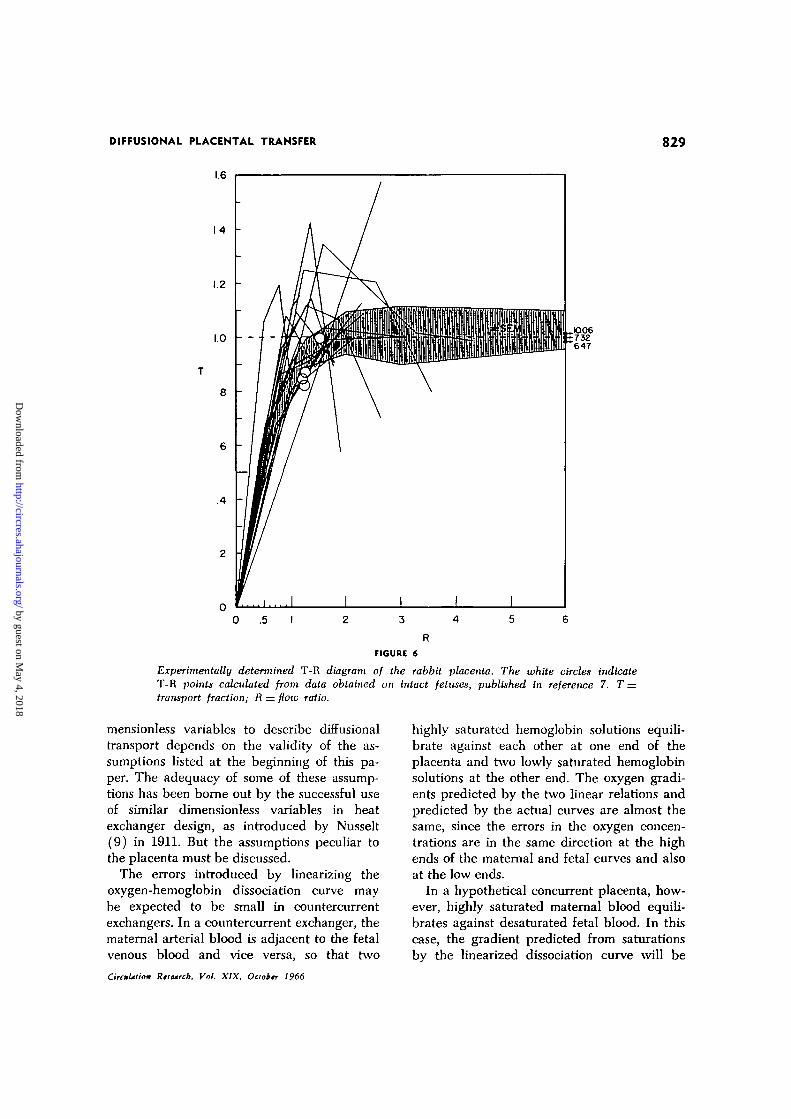
The values of T and R thus calculated areshown in Figure 6. Also shown in this figureare the four T-R points calculated from thesaturation data on intact fetuses of 27 to 29days gestation in reference 7. T was calculatedfrom its definition (equation 6). R could becalculated from the formula:
age oxygen gradients obtained by Bohrintegrations of the four fetuses in reference 7and by using equation 8, are 2.6, 3.6, 3.0 and3.9 (average 3.3). It should be noted thatthese Bohr integrations were performed on theassumption that there were no shunts oneither side of the rabbit placenta.
CONFIDENCE LIMITS OF T AND R
It was shown above that there is 95$ con-fidence that the rate of maternal placentalblood flow as measured with acetylene isnot more than 13? greater than the rate cal-culated on the basis of oxygen uptake.Since R was calculated from the rate of flowbased on oxygen uptake, there is 95$ con-fidence that if there is an overestimate in R,it is not more than 13%. A similar argumentwould lead to the conclusion that there is95$ confidence that an underestimate in Ris not more than 8$, if it were not for the logi-cal impossibility of the oxygen tension in thematernal vein being less than zero. R cannotbe underestimated.
T was calculated on the assumption thatthe equilibration of oxygen between the ma-ternal vein and the fetal artery was completeat high rates of perfusion (Sat*1* = SatFa = 0).It follows from the consideration that if thetrue quantity (Sat*"- Sat"") in equation 11is 13$ less than its assumed value, the samequantity in equation 6 will cause the true T
(11)
which follows from equations 4 and 7.Since the rabbit placenta is a countercurrent
exchanger, its T-R diagram can be comparedwith that of the ideal countercurrent exchang-er in Figure 1. It should be taken into accountthat the values of R in the diagram of therabbit placenta are approximately 14% too high,as that proportion of the perfusate seemedto bypass exchange with maternal vessels. Ifthe R scale is reduced by that degree, theaverage T-R curve of the exchanging part ofthe rabbit placenta seems to coincide withthe ideal curve at d = 10 in Figure 1. The val-ues of d, calculated from the published aver-
to be 13$ lower than the T calculated on thebasis of complete oxygen equilibration. Theupper and lower 95$ confidence ratios of Tare therefore 1.00 and 0.87, respectively.
These confidence limits of T and R are onlythose of the entire group of curves in Figure 6.The individual scatter of T-R lines in thisfigure represents an additional degree of un-certainty, as indicated by the shaded area inthe figure.
Discussion
THE VALIDITY OF THE REPRESENTATIONON THE T-R DIAGRAM
The validity of the use of only three di-CircmUtion Rtstmrcb, Vol. XIX, Octobtr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from
DIFFUSIONAL PLACENTAL TRANSFER 829
.4 -
FIGURE 6
Experimentally determined T-R diagram of the rabbit placenta. The white circles indicateT-R points calculated from data obtained on intact fetuses, published in reference 7. T =transr>ort fraction; R = flotv ratio.
mensionless variables to describe diffusionaltransport depends on the validity of the as-sumptions listed at the beginning of this pa-per. The adequacy of some of these assump-tions has been borne out by the successful useof similar dimensionless variables in heatexchanger design, as introduced by Nusselt(9) in 1911. But the assumptions peculiar tothe placenta must be discussed.
The errors introduced by linearizing theoxygen-hemoglobin dissociation curve maybe expected to be small in countercurrentexchangers. In a countercurrent exchanger, thematernal arterial blood is adjacent to the fetalvenous blood and vice versa, so that two
CircmUiion Rtsturcb, Vol. XIX, Oaobtr 1966
highly saturated hemoglobin solutions equili-brate against each other at one end of theplacenta and two lowly saturated hemoglobinsolutions at the other end. The oxygen gradi-ents predicted by the two linear relations andpredicted by the actual curves are almost thesame, since the errors in the oxygen concen-trations are in the same direction at the highends of the maternal and fetal curves and alsoat the low ends.
In a hypothetical concurrent placenta, how-ever, highly saturated maternal blood equili-brates against desaturated fetal blood. In thiscase, the gradient predicted from saturationsby the linearized dissociation curve will be
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

830 FABER, HART
higher than the real gradient, since error atthe low end of the fetal curve is added to theerror at the high end of the maternal curve.In order to investigate the extent of this effect,a T-R diagram was calculated for a con-current exchanger on the basis of sigmoid dis-sociation curves with a difference in fetal andmaternal half-saturation pressures of 1.25times. Fetal and maternal saturations were as-sumed and Bohr integrations performed byhand. Equations 8 and 9 yielded the value ofdo.,, and the definition of To2 (equation 6)was used to find T. The assumed saturationswere chosen in such a way that a number ofintegrations were made at each value of R.At these values the calculated ds were plottedagainst the calculated T"s. The T"s at d"s of 0.1,0.3, 1.0, 3.0, and oo were obtained by curvilin-ear interpolation and plotted versus R in Fig-ure 7. It can be seen that a given pair ofvalues of d and R no longer determines aunique T, but that T is still determined withina narrow range. Therefore, the T-R diagramis still almost equivalent to the raw data, inspite of the error in the assumption that theoxygen hemoglobin dissociation curve is lin-ear. It can also be seen, however, that in the
range where R ~ 1, the shape of the diagramis somewhat different from the T-R diagram ofthe same exchanger perfused with a linearlysoluble solute, as depicted in Figure 1.
The assumption that no diffusion relativeto fluid transport takes place in the directionof the blood stream is clearly necessary, sincein the case of infinitely fast diffusion in thisdirection, the concentration at the two ends ofthe vessel would be the same. All placentas,no matter what the actual arrangement oftheir vessels, would then obey the T-R dia-gram of double pool flow placentas. It isevident from a comparison of Figures 1 and6 that rabbit placentas are not double poolflow placentas.4
Fluid filtration across the barrier of theplacenta is immeasurably small. Oxygentransport by this mechanism is likely to be
4A calculation based on an approximate linearconcentration gradient along the placental bloodvessels showed that, in rabbits, the transport alongthe capillary by diffusion is only about one hundredthof the transport by bulk flow so that the concentra-tion gradient along the vessel depends almost entirelyon the latter form of transport—as assumed in thederivations.
ideal
rate
ICry f
i
concurrent
^ ^
1
placenta
1
d
_——
—3
d
. •
d
d
i
1 '
. 3
• 1 __------r- " !
. 3
> 1
i i
FIGURE 7
T-R diagram of an ideal concurrent placenta bated on Bohr integrations. The shaded areasindicate the range of values of To obtained at any given d0 . The diagram is based on 37integrations.
CircmUtion Rutrcb, Vol. XIX, Oaottr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

DIFFUSIONAL PLACENTAL TRANSFER 831
negligible. In the case of diffusion of non-h'pid-soluble substances, reevaluation of thistransport mechanism will be necessary (20,21).
The eighth assumption, that all parts ofthe placenta are equivalent, is merely a sim-plification used to compare the behavior of aplacenta with the theoretical behavior of asimple exchanger (5). Equations exist, how-ever, for a great number of fairly inhomo-geneous heat exchangers and these have beenexpressed (3) as a relation between the samethree dimensionless variables used for simpleexchangers, such as the ones whose behavioris shown in Figure 1. The calculation of Tand R from experimental data does not re-quire an assumption regarding the vasculargeometry of the placenta, except in the casethat no samples can be obtained from one ofthe four vessels. The calculation of an abso-lute value of d would require an assumptionregarding the geometry of the vessels, althoughexperimental T-R curves can be made atknown ratios of d without it if eitherQu ^ jn c a n jjg varied experimentally.
The comparison of the experimental T-Rdiagram with the T-R diagram of idealizedcountercurrent exchangers is misleading if nocorrections are made for the oxygen consump-tion of the placenta. The fraction of the ma-ternal flow rate satisfying the placental oxy-gen demands was therefore deleted in thecalculation of Ro.,. The value of do2 will alsobe affected since the oxygen consumption ofthe placenta disturbs the gradient betweenthe maternal and fetal bloods. However, sincedOn is experimentally determined from the re-lations between To,, and Ro2 and the vasculargeometry, the error in do2 does not invalidatethe prediction of Tc»o from Ro2 or vice versa.
There remains a question regarding theaccuracy of the T-R diagram. The dimension-less variables are more sensitive to experi-mental errors than the elementary variablesof which they consist. This is particularlytrue of those variables which consist of one ormore differences between two measured vari-ables. This inaccuracy cannot be avoided inany experimental description of the properties
CircmUsion Ristrcb. Vol. XIX, Ociobtr 1966
of the placenta as an exchanger, which de-pends on measured concentrations and rates offlow. However, the variability in the mea-sured variables is only partly due to errors ofmeasurement. Much of the variability can beascribed to the variability in the rates ofblood flow and the arterial concentrations pro-vided by different fetuses and mothers. Sincethe relationships between T, R, and d are aproperty of the placenta alone, only that partof the variability in the measured variablesthat is due to experimental errors occurs inthe derived relationships between the di-mensionless variables.
Compared to the use of a modified Bohrintegration (6-8) as a means to describe pla-cental efficiency, the T-R diagram seems tooffer two advantages:
1. It does not require any assumptionsregarding the vascular geometry, but canbe constructed from purely experimentaldata.
2. Once obtained, it can be used to predicttransfers under different circumstances,for instance, a different rate of fetalblood flow, without requiring anotherlaborious numerical approximation. Thisrequires the use of equation 10.
OXYGEN UPTAKE AS A FUNCTION OF FETAL FLOW
Inspection of Figures 5 and 6 shows thatoxygen uptake is already two-thirds of themaximum possible oxygen uptake at the nor-mal rate of umbilical blood flow of about 2ml/min or a normal R of about 1.3. Nearlymaximum oxygen uptake is reached at abouttwo times the normal fetal rate of placentalblood flow. The normal fetus operates hisplacenta at a flow rate where the curvature ofthe T-R relation is maximal. An increase infetal flow rate beyond normal effects only asmall increase in oxygen recovery, whereas adecrease in fetal flow rate is followed by aproportional decrease in oxygen recovery. Fe-tal flow rate is thus ideally matched to thecombined effects of the vascular structure andthe permeability of the placenta.
This conclusion may not, however, be ex-tended to the human placenta, as it is unlikelythat the human placenta is functionally a
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

832 FABER, HART
countercurrent exchanger. Similar experimentsperformed on monkeys with a villous placen-ta would yield a T-R diagram that could beapplied to the human placenta if the degreeof fetal shunting of the placental vessels isabout the same in the two species, and pro-vided evidence could be adduced to show thedoo values are approximately the same. Someidea of the behavior of the human placentacan be obtained, however, when publishedsaturations and half-saturation pressures ofthe fetal and maternal bloods—measured atbirth—are converted to To2 and Ro2 values andplotted. Figure 8 shows such a plot togetherwith the results of some values from otherspecies. Whether the low human and the even
lower primate Ton values are due to a lowvalue of do* of the villous placenta (do, "= 1)or to a less advantageous vascular structure,or both, is not certain. This question couldprobably be decided on the basis of perfusionexperiments in which oxygen uptake is com-pared to acetylene loss in villous placentas.Comparison with Figure 1 indicates that ineither case an increase in fetal flow rate be-yond the median value of about 0.7 Ro2 willbe more beneficial, and a decrease less dis-advantageous, than in rabbits. Similar conclu-sions appear to hold a fortiori for sheep.
AcknowledgmentThe authors wish to thank Mrs. Sadami Ono, who
o-Sheep at sea level c-Sheep after 10 days at altitude•-Sheep permanently at altitude F-Rabbits t-Meno-Cows A-Rhesus monkeys
FIGURE 8
The T-R diagrams of various placentas. Data were taken from the literature. Sheep at sea levelfrom ref. 6, sheep at altitude for 10 days from ref. 22, and sheep permanently at altitude fromref. 23. The ranges of fetal ages were similar in the three groups, and no systematic relationwith age was apparent. The data for Rhesus monkeys were taken from refs. 24 and 25, thedata for cows from ref. 26, the data for humans from ref. 27 and the data for rabbits fromref. 7. The approximate areas on the diagram that appear to be typical for sheep, man, andrabbits are shaded.
Circulation Research, Vol. XIX, October 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

DIFFUSIONAL PLACENTAL TRANSFER 833
rendered assistance with numerous computations,only some of which are referred to in this paper;Dr. John \V. Prothero, who discussed the theoryand suggested improvements in the presentation; andDr. James Metcalfe, who also helped clarify presen-tation of the research.
References1. BUCKINGHAM, E.: On physically similar systems:
Illustrations of the use of dimensional equa-tions. Physiol. Rev. 4: 345, 1914.
2. LANCHAAR, H. L.: Dimensional Analysis andTheory of Models. New York, John Wiley &Sons, Inc., 1962, pp. 18-20.
3. BOWMAN, R. A., MUELLER, A. C , AND NAGLE,W. M.: Mean temperature difference in de-sign. J. Heat Transfer 62: 283, 1940.
4. WELKIN, P.: Les theories explicatives due me-canisme des echanges transplacentaires. InLe Placenta Humain, ed. by Jean Snoeck.Paris, Masson et Cie, 1958, pp. 248-279.
5. BAHTELS, H., AND MOLL, W.: Passage of inertsubstances and oxygen in the human placenta.Pfliigers Arch. Ges. Physiol. 280: 165, 1964.
6. BARHON, D. H.: Some aspects for the transferof oxygen across the syndesmochorial placen-ta of the sheep. Yale J. Biol. Med. 24:169, 1951.
7. BARRON, D. H., AND BATTACLIA, F. C : Oxygenconcentration gradient between the plasmasin the maternal and fetal capillaries of theplacenta of the rabbit. Yale J. Biol. Med. 2S:197, 1956.
8. LAMPORT, H.: Transport of oxygen in the sheep'splacenta; the diffusion constant of the pla-centa. Yale J. Biol. Med. 27: 26, 1954.
9. NUSSELT, W.: Der Wanneubergang im Kreuz-strom. Z. Ver. dt. Ing. 55: 2021, 1911.
10. HART, F. M., AND FABER, J. J.: Fetal and ma-ternal temperatures in rabbits. J. Appl. Phys-iol. 20: 737, 1965.
11. SEVERINCHAUS, J. W.: Oxyhemoglobin dissocia-tion curve correction for temperature and pHvariation in human blood. J. Appl. Physiol.12: 485, 1958.
12. BARTELS, H., AND HARMS, H.: Sauerstoffdisso-ziationskurven des Blutes von Saugetieren.Pflugers Arch. Ces. Physiol. 268: 334, 1959.
13. ALTMAN, P. L., AND DITTMER, D. S.: BiologyData Book. Federation of American Societiesof Experimental Biology, p. 269, 1964.
14. GHILAIN, A.: La consommation d'oxygenepar Ie placenta humain in vitro. Bull. Soc.
Roy. Beige Gynecol. Obstet. 32: 451, 1962.15. FRIEDMAN, E. A., AND SACHTLEBEN, M. R.:
Placental oxygen consumption in vitro. Am.J. Obstet. Gynecol. 79: 1058, 1960.
16. MACKAY, R. B.: Studies of the oxygen consump-tion of fresh placental tissue from normaland abnormal pregnancies. J. Obstet. Gy-naecol. Brit. Empire 65: 791, 1958.
17. FARR, D. A., AND FUHRMAN, F. A.: Role ofdiffusion of oxygen in the respiration oftissues at different temperatures. J. Appl.Physiol. 20: 637, 1965.
18. MOSSMAN, H. W.: Rabbit placenta and theproblems of placental transmission. Am. J.Anat. 37: 433, 1926.
19. METCALFE, J., MOLL, W., BARTELS, H., HTL-PERT, P., AND PARER, J. T.: Transfer of carbonmonoxide and nitrous oxide in the artificiallyperfused sheep placenta. Circulation Res. 16:95, 1965.
20. KEDEM, O., AND KATCHALSKY, A.: Thermody-namic analysis of the permeability of biologicalmembranes to non-electrolytes. Biochim. Bio-phys. Acta. 27: 229, 1958.
21. KEDEM, O., AND KATCHALSKY, A.: Physicalinterpretation of the phenomenological coeffi-cients of membrane permeability. J. Gen.Physiol. 45: 143, 1961.
22. KAISER, I. H., CUMMINCS, J. N., REYNOLDS,S. R. M., AND MARBARCER, J. P.: Acclimati-zation response of the pregnant ewe and fetallamb to diminished ambient pressure. J. Appl.Physiol. 13: 171, 1958.
23. METCALFE, J., MESCHIA, G., HELLECERS, A. E.,PRYSTOWSKY, H., HUCKABEE, W., AND BARRON,D. H.: Observations on the placental ex-change of the respiratory gases in pregnant eweat high altitudes. Quart. J. Expd. Physiol.47: 74, 1962.
24. BEHRMAN, R. E., HELLER, C. J., BATTACLLA,F. C., AND HELLECERS, A. E.: Comparison ofthe oxygen affinity of maternal and fetal bloodof the Macaca mulatto. Quart. J. Expd.Physiol. 48: 258, 1963.
25. BATTACLLA, F. C, ET AL.: Gestational changesin arteriovenous difference of oxygen acrossthe uterus of the rhesus monkey. Am. J. Ob-stet. Gynecol. 93: 32, 1965.
26. Roos, J. AND ROMIJN, C : Some conditions offoetal respiration in the cow. J. Physiol.(London) 92: 249, 1938.
27. WULF, H.: Oxygen and carbon dioxide tensiongradients in the human placenta at term. Am.J. Obstet. Gynecol. 88: 38, 1964.
CircuUtton Rtsetrcb, Vol. XIX, Octoktr 1966
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from

J. JOB FABER and FREDERICK M. HARTby Dimensional Analysis
The Rabbit Placenta as an Organ of Diffusional Exchange: Comparison with Other Species
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1966 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.19.4.8161966;19:816-833Circ Res.
http://circres.ahajournals.org/content/19/4/816World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Research Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on May 4, 2018
http://circres.ahajournals.org/D
ownloaded from