the interaction of dna with pancreatic ribonuclease a
TRANSCRIPT

202 BIOCHIMICA ET BIOPHYSICA ACTA
BBA 96086
T H E INTERACTION OF DNA W I T H PANCREATIC RIBONUCLEASE A
H I R O S H I S E K I N E , E I I C H I N A K A N O AND K E N J I S A K A G U C H I
Noda Institute for Scientific Research, Noda-shi, Chiba-ken (Japan)
(Received J u l y Is t , 1968)
SUMMARY
I. The complex formation of calf thymus DNA with pancreatic ribonuclease A (ribonucleate pyrimidinenucleotido-2'-transferase (cycling), EC 2.7.7.16) was stu- died through ultracentrifugation, competitive inhibition of ribonuclease activity and gel filtration on Sephadex G-2oo. Ultracentrifugation data indicated that under low ionic strength ribonuclease formed a soluble complex with DNA at least in the range of pH 7.5 to pH 8.5 and this ability to form the complex was strongly affected by the presence of 0.05 M NaC1.
2. The activity of ribonuclease was competitively inhibited by DNA when as- sayed in o.oi M phosphate buffer (pH 7.5). The denatured DNA was much more effective than native DNA in inhibiting ribonuclease activity.
3. Through gel filtration on Sephadex G-2oo, ribonuclease was found to form a complex with DNA in a definite combining ratio in o.oi M phosphate buffer (pH 7.5). The combining ratios were calculated to be about one ribonuclease molecule per 500 ooo-Iooo ooo daltons of native DNA and 8ooo-lo ooo daltons of denatured DNA. In the presence of o.I M NaC1 the complex dissociated into free enzyme and DNA. The combining ratio of ribonuclease to native DNA was about IOO times smaller than that of denatured DNA and the discrepancy is discussed in connection with specific sites on DNA. The thermal denaturation profile of DNA was not affected by ribo- nuclease in the amount of this combining ratio.
INTRODUCTION
From many studies it has been widely accepted that a certain specific protein is apt to bind to a specific base sequence or to other specific sites of nucleic acids. For example, aminoacyl-tRNA synthetase combines with a definite tRNA 1. Ribo- somes contain only definite ribosomal proteins 2,~. I t is observed from electromicro- scopical and kinetic studies that the DNA-dependent RNA polymerase is fixed on specific points of polyoma DNA 4,5. However, more information is necessary to un- derstand the mode of interaction of DNA with proteins and the character of these complexes.
In the course of our study on the interaction of DNA with several proteins, we found that under certain conditions ribonuclease, which originally has a property to bind with RNA, formed a soluble complex with DNA at a definite combining
Bioehim. Biophys. Acta, 174 (1969) 2o2-21o

DNA-RIBONUCLEASE A INTERACTION 203
ratio. Concerning the interaction of ribonuclease with DNA, FELSENFELD, SANDEEN AND VON HIPPEL s reported the destabilizing effect of ribonuclease on the thermal denaturation of DNA.
The present paper is concerned with some properties of this soluble complex.
MATERIALS AND METHODS
Calf thymus DNA was prepared according to the method of SAITO AND MIU- RA'S 7 pH 9 phenol extraction with a little modification and was stored in the stand- ard saline citrate solution at --20 ° . After being dialyzed against an appropriate buffer, this preparation was used throughout these experiments unless otherwise stated. The protein content of this preparation was less than 0.5 %. Heat denatured DNA was prepared by heating this stock solution at IOO ° for IO min followed by rapid cooling. The amount of DNA was determined by colorimetric analysis of phos- phorus or by calculating from the absorbance at 260 m#. DNA preparations were checked for their denaturation degrees by the method of SHACK 8.
RNA was extracted from commercial baker 's yeast using sodium lauryl sul- fate and further purified by KIRBY'S method 9. The amount of RNA was determined by the same method used for DNA.
Commercial preparations of bovine pancreatic ribonuclease (Worthington Bio- chemical Corp. and Sigma Chemical Co.) were further fractionated by CM-cellulose column chromatography according to the method of TABORSKY 1° and the main com- ponent (TABORSKY'S fraction D) was used in our experiments after dialysis against an appropriate buffer. The amount of enzyme protein was usually determined colo- rimetrically by the method of LOWRY et al. 11. When very small amounts of ribonu- clease protein had to be determined, they were calculated from ribonuclease activity.
The activity of ribonuclease was assayed by a modified method of KUNITZ TM. In the usual assay of fractions obtained from gel filtration, for example, the final reaction mixture contained 1.2 mg RNA in I.O ml of 0.04 M phosphate buffer at pH 7.5. After incubation for 5 or IO min at 37 °, the reaction was terminated by the addition of an equal volume of IO % perchloric acid containing 0.3 ~o uranyl acetate. The mixture was allowed to stand at o ° for IO min and then centrifuged at 3000 rev./min for 15 min. 0.3 ml of the supernatant fluid was added to 4.0 ml of water and the absorbance was read at 260 m~. The absorbance was linear against ribonu- clease concentration up to o.12/~g of ribonuclease.
Gel filtration of DNA-ribonuclease soluble complex was performed using a column of approx. 2 c m x 50 cm packed with Sephadex G-2oo in a cold room. Ther- mal denaturation curves of DNA were obtained according to the method of MAR- MUR AND DOTY TM and by an auto-recording apparatus developed by WADA AND YABUK114.
RESULTS
Soluble complex/ormation o] ribonuclease with DNA In general, a protein forms an insoluble complex with a nucleic acid at pH
values below its iso-electric point, and this complex can be easily precipitated by
Biochim. Biophys. Acta, 174 (1969) 2o2-21o

204 H. SEKINE, E. NAKANO, K. SAKAGUCHI
centrifugation at 18oo ×g for 2o min (3ooo rev./min, a centrifuge of 18 cm radius). There are also observations, however, that at low salt concentrations a complex re- mains in solution at a lower pH if the components are first mixed at a higher pH. As shown by the experiment of HOFSTEE 15, this soluble complex can be completely sedimented by centrifugation at 127 ooo x g for 60 rain (4 ° ooo rev./min by Spinco Model L ultracentrifuge, Type 50 rotor), whereas little or no free protein is sedimented. In our experiments, ribonuclease was simply mixed with DNA in o.oi M buffers (acetate, phosphate and borax) at various pH values without any further t reatment such as mixing at higher pH followed by gradual lowering of pH. In the case of NaCl-free mixtures, as shown in Fig. IA, the range of pH was found to be consider-
60C
40C
.@'5 r r ~_
4
A
o° -
J o. ,0 . - - - ' 0 " " .0 o*"
0""
5 6 7 8 9 10 4 5 6 7 8 9 10 pN
E
200~ v
t~
~ o
.c < z C~
Fig. i. Effect of p H on the soluble complex formation of ribonuclease with DNA in the absence of NaC1 (A) or in the presence of NaCl (B). In 2.7 ml of o.oi M buffers described below with or wi thout o.o 5 M NaC1, 600/zg of ribonuclease w&s mixed with 20o/ ,g of DNA. Buffers used were ace ta te-sodium acetate (pH 4-6), Na2HPO4-KHsPO 4 (pH 6-8), NasBaOT-HC1 (pH 8.5) and N%B4OT-NaOH (above p H 9). After the mixtures were centrifuged, the amoun t s of ribonuclease ( .) and DNA (- - -) in the superna tan t fluid were determined. O, result of centr i fugat ioa at 18oo x g for 20 min; O, result of centrifugation at 127 ooo x g for 60 min.
ably wide where all the DNA and ribonuclease remained in the 18oo x g super- natant fluid and where both DNA and ribonuclease were sedimented by ultracen- trifugation at 127 ooo xg. I t is not clear why the DNA added to the mixture can not be completely recovered in 127 oooxg-supernatant fluid even at high pH values where ribonuclease can be completely recovered. However, the results in Fig. I show that under NaCl-free conditions ribonuclease forms a soluble complex with DNA at least in the range of pH 7.5 to pH 8.5 and that this ability to form a soluble complex is strongly affected by the presence of 0.05 M NaCI.
Inhibition o/ ribonuclease activity by DNA The formation of the soluble complex of ribonuclease with DNA suggests that
ribonuclease activity would be inhibited by DNA. The decrease of ribonuclease ac- t ivi ty by the addition of DNA into the reaction mixture was assayed with changes in pH, in the amount of substrate and in ionic strength. In the absence of NaC1 the activity of ribonuclease was markedly inhibited both by native and denatured DNA, although the degree of inhibition decreased along with the increase of pH values. The presence of o.I M NaC1 in the reaction mixture resulted in the release of the
Biochim. Biophys. Acta, 174 (1969) 2o2-21o
DNA-RIBONUCLEASE A INTERACTION 205
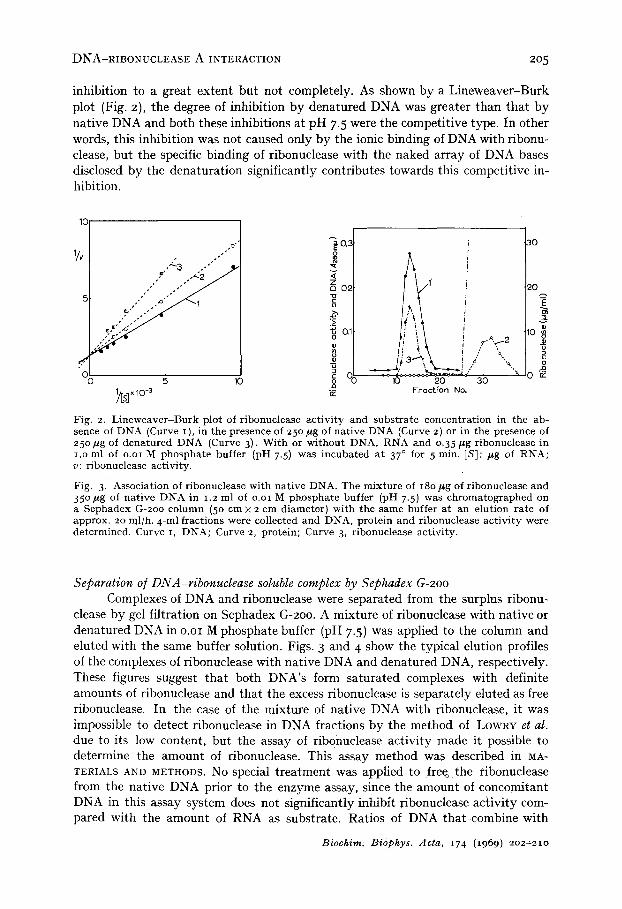
inhibition to a great extent but not completely. As shown by a Lineweaver-Burk plot (Fig. 2), the degree of inhibition by denatured DNA was greater than that by native DNA and both these inhibitions at pH 7.5 were the competitive type. In other words, this inhibition was not caused only by the ionic binding of DNA with ribonu- clease, but the specific binding of ribonuclease with the naked array of DNA bases disclosed by the denaturation significantly contributes towards this competitive in- hibition.
O'
...- 4....--"°"1
0 5 10 ~$] x lO -3
2Q3 o
< z a Q2
~ 0.1
~ q iff
; 3O
1 ~,. / 20
il i 10 20 3 0
Fraction No.
Fig. 2. L ineweaver -Burk plot of ribonuclease activity and subs t ra te concentrat ion in the ab- sence of DNA (Curve I I, in the presence of 250/*g of native DNA (Curve 2) or in the presence of 250/*g of denatured DNA (Curve 3). Wi th or wi thout DNA, RNA and o.35/~g ribonuclease in i .o ml of o.oi M phospha te buffer (pH 7.5) was incubated at 37 ° for 5 min. [Sl: #g of RNA; v: ribonuclease activity.
Fig. 3. Association of ribonuclease with nat ive DNA. The mixture of 18o/*g of ribonuclease and 35 ° #g of nat ive DNA in 1.2 ml of o.oi M phospha te buffer (pH 7.5) was chromatographed on a Sephadex G-2oo column (5o cm x 2 cm diameter) with the same buffer at an elution rate of approx. 20 ml/h. 4-ml fractions were collected and DNA, protein and ribonuclease act ivi ty were determined. Curve I, DNA; Curve 2, protein; Curve 3, ribonuclease activity.
Separation o/DNA-ribonuclease soluble complex by Sephadex G-2oo Complexes of DNA and ribonuclease were separated from the surplus ribonu-
clease by gel filtration on Sephadex G-2oo. A mixture of ribonuclease with native or denatured DNA in o.oi M phosphate buffer (pH 7-5) was applied to the column and eluted with the same buffer solution. Figs. 3 and 4 show the typical elution profiles of the complexes of ribonuclease with native DNA and denatured DNA, respectively. These figures suggest that both DNA's form saturated complexes with definite amounts of ribonuclease and that the excess ribonuclease is separately eluted as free ribonuclease. In the case of the mixture of native DNA with ribonuclease, it was impossible to detect ribonuclease in DNA fractions by the method of LOWRY et al. due to its low content, but the assay of ribonuclease activity made it possible to determine the amount of ribonuclease. This assay method was described in MA- TERIALS AND METHODS. No special t reatment was applied to free~ the ribonuclease from the native DNA prior to the enzyme assay, since the amount of concomitant DNA in this assay system does not significantly inhibit ribonuclease activity com- pared with the amount of RNA as substrate. Ratios of DNA that combine with
Biochim. Biophys. Acta, 174 (1969) 2o2~2io

206 H. SEKINE, E. NAKANO, K. SAKAGUCHI
ribonuclease were calculated based on the quantitative analyses of ribonuclease and DNA. Results obtained from several experiments are summarized in Table I. The amount of DNA needed to form a saturated complex with one mole of ribonuclease was calculated to be 500 ooo-Iooo ooo daltons for native DNA and 8ooo-lo ooo daltons for denatured DNA. When the mixture of ribonuclease with native DNA
T A B L E I
C O M B I N I N G R A T I O O F D I ~ A W I T H R I B O N U C L E A S E O B T A I N E D F R O M G E L F I L T R A T I O N A N A L Y S E S
In these ca lcula t ions , 13 7oo was used as the tool. wt. of r ibonuclease aS.
Expt. DNA (daltons/mole ribonuclease) No.
Native DNA-ribonuclease Denatured DNA-ribonuclease ( x io5) ( x io~)
i 6.5-7. 5 2 4.5-5.5 3 8.5-1o.o 4 8.5-1o.5 5 7 .6-11.1 6 8.8-1o. 3 P h a g e T 4 DNA 7.5
prepared from phage T 4 (ref. 7) was applied to gel filtration under the same condition, the combining ratio was found to be 75 ° ooo daltons of DNA per mole of ribonuclease. The lengths of these DNA molecules were checked by the electron microscope according to the method of KLEINSCHMIDT et al. 16 and their average molecular weight was found to exceed I . lO 7. The effects of NaC1 on the formation of these complexes were also examined. When gel filtration was carried out in the buffer solution con- taining o.I M NaC1, formation of the complex was not observed. Re-chromatography in the presence of o.I M NaC1 of fractions containing the complex obtained in Figs. 3 and 4 also resulted in the dissociation of the two components of the complex.
0,8
0.6
E o o.4
~ 0,2
O - -
P' "o
1o
30
4o- 2
2o }
o iff 20 30 40
Fraction No.
Fig. 4. Associa t ion of r ibonuclease wi th d e n a t u r e d DNA. The same procedure was appl ied ex- cep t t h a t the m i x t u r e of iooo #g of r ibonuclease and 260/zg of d e n a t u r e d DNA in 2.8 ml of buffer so lu t ion was charged on the column. 0 - 0 , DNA; O - - - O , prote in .
Biochim. Biophys. Aeta, 174 (1969) 2o2-21o

DNA-RIBONUCLEASE A INTERACTION 207
Thermal denaturation As shown in Fig. 5, at low ionic strength, thermal denaturation of DNA was
markedly affected by the presence of ribonuclease between 4 °0 and 7 o°. This de- stabilizing and renaturating effect of ribonuclease and its release by salt were origi-
1.4
o v
oo
o
1.1
~ I . 0 ft.
A
#' ', :
E~ :#' ,7.
40 5o 60 7b 8b ~o 4b 5o6b 7'o Bo 9b Temperoture
Fig. 5- Effect of ribonuclease on the thermal denatura t ion curve of DNA. Absorbance increase a t 260 mk* was plotted as a function of tempera ture with respect to the absorbance at 25 °. %: absorbance at 25% a: absorbance at specified temperature , DNA (% o.51 ) in o.ooi M phosphate buffer (pH 7.5) containing 0.002 M NaC1 (A) and o.oi M NaC1 (B) was heated wi thout (O-C) ) or with o. 13 mg ribonuclease per ml ( 0 - - -O) . The absorbance of ribonuclease was subtracted.
nally observed by FELSENFELD, SANDEEN AND VON HIPPEL e. In our experiments (repeated more than IO times) a complete renaturation of DNA, caused probably by the thermal transition of ribonuclease to a more unfolded form 17 at 67-7 o°, was consistently observed on calf thymus, E. coli and phage T 4 DNA. As for the relation with NaC1 concentration, this effect decreased along with the increase of NaC1 con- centration and no destabilizing effect on DNA was detected with NaC1 above o.I M. The same result was also obtained for DNA prepared from E. coli Monod's strainL Attempts to find this effect of ribonuclease on DNA in the soluble complex obtained from gel filtration (Fig. 3) were unsuccessful. Ribonuclease mixed with native DNA in the same ratio as that found in the soluble complex by gel filtration (Table I) also had no effect on the thermal denaturation of DNA. Ribonuclease at this gel filtration concentration involves so few base (less than o.I %) that the destabilizing effect would not be detectable in the optical melting experiment, though ribonu- clease at this concentration may also have a similar effect (locally) on the thermal denaturation of DNA, as do the more massive amounts of ribonuclease.
DISCUSSION
Under the condition of low ionic strength, DNA forms a soluble complex with ribonuclease in a considerably wide range of pH, although a further decrease of pH resalts in the formation of an insoluble complex as shown in Fig. I. The marked p H dependence suggests the predominant contribution of coulombic interaction between ribonuclease and DNA to the formation of the complex. In other words, the negati- vely-charged DNA more easily forms a complex with ribonuclease when ribonuclease
Biochim. Biophys. Acta, 174 (1969) 2o2-21o

208 H. SEKINE, E. NAKANO, K. SAKAGUCHI
is positively charged. Thus, the soluble complex is formed only at the pH where ribonuclease has only a few positive charges; at lower pH values, where more positive charges are available, the complex becomes insoluble due to the formation of large three-dimensional networks as proposed by HOFSTEE 15 in his report on the DNA- protein complex. On the presence of o.I M NaC1, little or no interaction of DNA with ribonuclease was observed for the soluble complex formation or for the inhibition of ribonuclease activity or for thermal denaturation. These effects of NaC1 may also be considered as confirmation that the interaction occurs mainly between the nega- tively-charged DNA and the positively-charged ribonuclease. HOFSTEE 19,~0 reported that a-chymotrypsin formed a soluble complex with DNA at low ionic strength and that in the saturated complex at pH 7.5 the combining ratio of protein to DNA was about 2o:1 (w/w) when estimated from kinetic and ultracentrifugation data. In his data, similar to our results, c~-chymotrypsin activity is inhibited by DNA and the inhibition is released by the presence of o.I M KC1 as a non-specific ion. Namely, the soluble complex is dissociated into the two components by o.I M KC1. However, the type of inhibition was competitive in the case of ribonuclease contrary to that of ~-chymotrypsin which was reported to be a non-competitive one. Dif- ference was also observed in the combining ratio, which was roughly calculated for ribonuclease to DNA to be about I : I (w/w); i.e. one protein molecule per about 35 nucleotide units (from the ultracentrifugation data at pH 7.5 depicted in Fig. I).
On the other hand, the participation of the non-ionic binding force in this interaction was suggested by the fact that denatured DNA was much more effective than native DNA in this competitive inhibition as shown in Lineweaver-Burk plot of Fig. 2. Ribonuclease can attack only single stranded RNA, but its hybrid with the complementary DNA 21 or double stranded RNA 2z is resistant to this attack. One explanation of this phenomenon can be the failure of ribonuclease to form a complex with the double stranded RNA or DNA-RNA hybrid. Our present inhibi- tion experiments and the results obtained from the gel filtration technique strongly support this idea although RNA is substituted with DNA. Generally, on the inter- action of DNA with ribonuclease, the coulombic affinity between the anionic nu- cleotide chain and the partially cationic enzyme may be a fundamental binding force. However, the effective addition of the enzymic binding force of ribonuclease with single stranded nucleic acids plays an important role in the formation of this complex.
I t is also obvious from Table I that in the complex obtained by gel filtration the combining ratio of ribonuclease to native DNA is smaller than that calculated from the ultracentrifugation data. Results of gel filtration show that ribonuclease combines with native DNA in a ratio of about one protein molecule per 15oo-3ooo nucleotide units. In contrast, in the case of denatured DNA, the ratio is about one protein mole- cule per 30 nucleotide units, a value which is more in accordance with that calculated from the ultracentrifugation data of native DNA. In addition to these facts, ribonu- clease added to DNA in the same ratio as indicated in the native DNA-ribonuclease complex by gel filtration had no detectable effect on the thermal denaturation curve of DNA. These results suggest that the complex obtained by gel filtration would be different from that detected by ultracentrifugation in the mode of interaction. In the latter case, the binding of ribonuclease to DNA would be rather non-specific. On the other hand, in the former case, where the ratio of ribonuclease to DNA is
Biochim. Biophys. Acta, 174 (1969) 2o2-21o

D~A-RIBONUCLEASE A INTERACTION ,209
very low, such as one protein molecule per 500 ooo-Iooo ooo daltons, the interac- tion would be more specific, which suggests the presence of specific binding sites for ribonuclease in the native DNA molecule.
According to SADRON'S 23-25 remark, there exist specific points on the DNA molecule with an average distance of 500 ooo daltons. These specific points are demon- strated as flexible points by light scattering method, or as weak points where the DNA molecule is susceptible to deoxyribonuclease ~6 and to ultrasonic irradiation. As to these points, he proposed two models. In the first one, there exists chemical hete- rogeniety at this point and this point is the junction between DNA subunits composed of a peptidic link as suggested by KIRBY 2~ and by LESKO AND EMERY ~8. In the sec- ond one, it is proposed that there is no chemical heterogeniety at this point and the double stranded DNA has a local detwisting of the helix. Our results also suggest that there are specific points on native DNA at each 500 ooo-Iooo ooo daltons. From the fact that denatured DNA combines with ribonuclease in a ratio about IOO times higher than native DNA, the specific binding site for ribonuclease in the native DNA molecule (though there is no direct proof that this site is identical with the SANDRON'S specific point) would be more easily explained by the second model than by the first one proposed by SADRON.
ACKNOWLEDGEMENTS
The authors wish to express their sincere thanks to Professors Y. IKEDA and M. TSUBOI of the University of Tokyo for their valuable discussions. They are also grateful to Professor C. SADRON for his kind suggestions. The kind permission to use the temperature-shifting spectrophotometer (by Drs. T. ANDO and S. YABUKI) is gratefully acknowledged. This work was part ly supported by a grant from the Ministry of Education, Japan.
R E F E R E N C E S
I G. I,. BROWN, in J. N. DAVlDSON AND W. E. COHN, Progress in Nucleic Acid Research, Vol. 2, Academic Press, New York, 1963, p. 259.
2 P. F. SPAHR, J. Mol. Biol., 4 (1962) 395. 3 E. C. Cox AND J. G. FLAKS, Federation Proc., 23 (1964) 22o. 4 L. V. CRAWFORD, E. M. CRAWFORD, J. p. RICHARDSON AND H. S. SLA.gTER, J. Mol. Biol., 14
(1965) 593. 5 D. PETTIJOHN AND T. KAMIYA, J. Mol. Biol., 29 (1967) 275. 6 G. FELSENFELD, G. SANDEEN AND P. H. VON HIePEL, Proc. Natl. Acad. Sci. U.S., 5 ° (1963)
644. 7 H. SAITO AND K. MIURA, Biochim. Biophys. Acta, 72 (1963) 619. 8 J. SHACK, J. Biol. Chem., 233 (1958) 677. 9 K. S. KIRB.g, Biochem. J., 64 (1956) 4o5 .
io G. TABORSKY, J. Biol. Chem., 234 (1959) 2652. I I O. H. LOWRY, N. J. ROSEBROUGH, A. L. FARR AND R. J. RANDALL, J. Biol. Chem., 193 ~I95 I)
265. 12 M. KONITZ, J. Gen. Physiol., 24 (194 o) 15. 13 J. MARMUR AND P. DOT'g, J. Mol. Biol., 5 (1962) lO9. 14 S. YABUKI ANn A. WADA, Seibutsu Butsuri Japan, 6 (1966) 83. 15 B. H. J. HOFSTEE, Biochim. Biophys. Acta, 91 (1964) 337- I6 A. K. KLEINSCHMIDT, D. LANG, D. JACHERTS AND R. K. ZAHN, Biochim. Biophys. Acta, 61
(1962) 857.
Biochim. Biophys. Acta, 174 (1969) 2o2-21o

210 I~. SEKINE, E. NAKANO, K. SAKAGUCHI
27 W. F. HARRINGTON AND J. SHELLMAN, Compt. Rend. Tray. Lab. Carlsberg, 3 ° (I956) 21. 18 D. G. SMITH, J. H. STEIN AND S. MOORE, J. Biol. Chem., 238 (1963) 227. 19 B. H. J. HOFSTEE, Biochim. Biophys. Acta, 55 (1962) 44 o. 2o B. H. J. HO~STEE, J. Biol. Chem., 238 (1963) 3235 . 2i A. LANDY, J. ABELSON, H. M. GOODMAN AND J. D. SMITH, J. Mol. Biol., 29 (1967) 457- 22 A. R. BELLAMY, L. SttAPIRO, J. T. AUGUST AND W. K. JOKLIK, J. Mol. Biol., 29 (1967) i. 23 C. SADRON, Seminar at the Institute o/ Applied Microbiology, University o/ Tokyo, 1966. 24 C. SAI)RON, J. POUYET, A. M. FREUND AND M. CHAMPAGNE, J. Chim. Phys., IO (1965) 1187. 25 C. SADRON, Bull. Soc. Chim. Biol., 5 ° (1968) 16. 26 G. BERNARDI AND C. SADRON, Biochemistry, 3 (1964) 1411. 27 K. S. KIRBY, Biochem. J., 66 (1957) 495. 28 S. A. LESKO, Jr. AND A. J. EMERY, Jr., Biochem. Biophys. Res. Commun., 23 (1966) 707 .
Biochim. Biophys. Acta, I74 (1969) 2o2-21o