tesis doctoral aranzazu urresola olabarrieta
TRANSCRIPT

1
Tesis Doctoral Aranzazu Urresola Olabarrieta
Topografía Precisa de las Subunidades de los Canales de
Potasio Kv3.1b y Kv3.3 en la Capa Molecular de la Corteza
Cerebelosa de la Rata
Resumen
......................................................................................................... 3
Introducción
......................................................................................................... 8
Hipótesis de trabajo
........................................................................................................ 43
Objetivos
......................................................................................................... 48
Material y métodos
......................................................................................................... 50
Resultados
........................................................................................................ 60
Discusión
......................................................................................................... 73
Conclusiones
......................................................................................................... 94
Bibliografía
........................................................................................................ 98

2
Tesis Doctoral Aranzazu Urresola Olabarrieta
RESUMEN

3
Tesis Doctoral Aranzazu Urresola Olabarrieta
RESUM EN
Los canales de potasio dependientes de voltaje Kv3.1 y Kv3.3
intervienen en la generación de potenciales de acción rápidos, que
permite a las neuronas que los poseen disparar potenciales de acción
a alta frecuencia (Rudy y McBain 2001). Son canales activados a
potenciales de membrana relativamente positivos, y se caracterizan
por sus cinéticas de activación extraordinariamente rápidas,
moldeando así la repolarización del potencial de acción (Rudy and
McBain 2001). Estudios de hibridación in situ e inmunohistoquímica
han demostrado que Kv3.1 y Kv3.3 se expresan en abundancia en la
corteza cerebelosa (Goldman-Wohl y cols., 1994; McMahon y cols.,
2004), en particular, en las células granulares (Weiser y cols., 1994,
1995; Sekirnjak y cols., 1997; Grigg y cols., 2000; Li y cols., 2001;
Rudy McBain 2001; Ozaita y cols., 2002; Alonso-Espinaco y cols.,
2008; Puente y cols., 2010). Desde el punto de vista funcional, los
ratones que carecen de los canales Kv3.1 y Kv3.3 presentan un
síndrome cerebeloso caracterizado por ataxia severa y temblor,
además de ser particularmente sensibles al alcohol (Espinosa y cols.,
2001, 2004; Matsukawa y cols., 2003, Joho y cols., 2006). Estas
evidencias en conjunto sugieren que la presencia de Kv3.1 y Kv3.3 es
crítica en el mantenimiento de una función cerebelosa normal.

4
Tesis Doctoral Aranzazu Urresola Olabarrieta
Por otra parte, se ha visto que ratones sin Kv3.3, pero con Kv3.1,
cursan con incoordinación motora (McMahon y cols., 2004, Joho y
cols., 2006; Hurlock y cols., 2008), que desaparece tras la expresión de
nuevo de Kv3.3 en las neuronas de Purkinje de estos ratones, al igual
que ocurre al introducir Kv3.3 en ratones que carecen de los dos alelos
de Kv3.3 y un alelo de Kv3.1 (Hurlock y cols., 2008). No obstante, el
aprendizaje de habilidades motoras no se ve afectado en los ratones
mutantes sin Kv3.3 (Hurlock y cols., 2008). Merece una especial
mención el hecho de que la mutación del gen que codifica el canal
Kv3.3 es la responsable de la ataxia espinocerebelosa 13 de carácter
familiar, caracterizada por un cuadro de ataxia y degeneración
cerebelosa de aparición en individuos adultos (Waters y cols., 2006;
Waters y Pulst, 2008; Schorge y cols., 2010).
La coexpresión en las células granulares de los canales Kv3.1 y
Kv3.3, ambos con propiedades biofísicas similares, es asumida como
una redundancia functional (Ho y cols., 1997; Sánchez y cols., 2000),
ya que los alelos de Kv3.1 y Kv3.3 eliminados genéticamente en una
manera dosis-dependiente, causa anormalidades en la plasticidad a
corto plazo de la transmisión sináptica de las fibras paralelas con las
células de Purkinje, lo cual está unido a la aparición de deficiencias
motoras (Matsukawa y cols., 2003). A pesar de los hallazgos que

5
Tesis Doctoral Aranzazu Urresola Olabarrieta
apuntan hacia un papel cofuncional de los canales Kv3.1 y Kv3.3 en
las fibras paralelas, no se conoce la localización ultraestructural
precisa ni la contribución de cada una de estas subunidades en los
compartimentos específicos (terminales sinápticas/ porciones
intervaricosas) de los axones de las células granulares (las fibras
paralelas), lo que es fundamental en la fisiología sináptica
excitadora de las fibras paralelas con las espinas dendríticas de las
células de Purkinje.
Esta investigación por tanto se ha concebido con el fin de
estudiar estas cuestiones en la corteza cerebelosa de la rata,
utilizando para ello antisueros específicos frente al canal Kv3.1b y
Kv3.3 en combinación con técnicas inmunocitoquímicas de alta
resolución para microscopía electrónica.
Los resultados de esta Tesis Doctoral han demostrado la
presencia de numerosas inmunopartículas que señalan la
localización de Kv3.1b y Kv3.3, en membranas de las terminales
sinápticas y segmentos intervaricosos de las fibras paralelas. En
particular, Kv3.1b está aproximadamente en el 85% de los botones
sinápticos, y en cerca del 47% de las porciones intervaricosas de las
fibras. Sin embargo, sólo el 28% de las intervaricosidades y un 23%
de las terminales sinápticas de las fibras paralelas contienen Kv3.3.

6
Tesis Doctoral Aranzazu Urresola Olabarrieta
Por otro lado, el análisis también revela que las espinas dendríticas
de las células de Purkinje son el principal recipiente de Kv3.3 (54%).
En conclusión, aunque ambas subunidades de canales de
potasio comparten localización en las mismas porciones
presinápticas de las fibras paralelas, los resultados de esta Tesis
Doctoral indican que el canal de potasio Kv3.1b en la corteza
cerebelosa contribuye en gran medida a la construcción de la
arquitectura molecular de las fibras paralelas, mientras que Kv3.3
ejerce sobre todo una influencia postsináptica a través de su
localización mayoritaria en las espinas dendríticas de las células de
Purkinje.

7
Tesis Doctoral Aranzazu Urresola Olabarrieta
INTRODUCCIÓN

8
Tesis Doctoral Aranzazu Urresola Olabarrieta
INTRODUCCIÓN
El cerebelo es una parte del sistema nervioso central que ejerce
una función crucial en el control motor. Desde el punto de vista
clínico, la lesión del cerebelo ocasionada generalmente por tumores o
por causas vasculares, da origen a la aparición del denominado
síndrome cerebeloso. Este síndrome en su forma más florida se
caracteriza por pérdida del equilibrio y la postura, alteración de los
movimientos oculares (nistagmo), dificultad en el habla (disartria,
palabra escandida), temblor intencional, ataxia (marcha inestable,
temblorosa), hipotonía, dismetría, adiadococinesia (incapacidad de
realizar movimientos rápidos alternantes) y falta de coordinación de
los movimientos finos. Además, estos pacientes tienen una gran
dificultad en el aprendizaje de actos motores. La base celular de esta
sintomatología radica en la afectación de los circuitos a nivel de la
corteza cerebelosa y de los núcleos profundos. Ambos están
interconectados por las células de Purkinje, únicas células de la
corteza cerebelosa que proyectan a los núcleos profundos. A partir de
estos últimos se organizan las proyecciones a diferentes centros
nerviosos, sobre todo a la corteza cerebral a través del tálamo, a
centros subcorticales como el núcleo rojo y la formación reticular y a
la médula espinal, ejerciendo así su efecto sobre el control del
movimiento.

9
Tesis Doctoral Aranzazu Urresola Olabarrieta
El cerebelo, para realizar su función, recibe abundante
información del estado de nuestro cuerpo a través de dos tipos de
proyecciones nerviosas: las fibras musgosas y las fibras trepadoras,
originadas estas últimas exclusivamente en la oliva inferior. Las
primeras terminan en la capa granular de la corteza cerebelosa
formando parte de los glomérulos cerebelosos. A este nivel, hay un
relevo sináptico desde las fibras paralelas de las células granulares a
las espinas dendríticas de las células de Purkinje. Por su parte, las
fibras trepadoras terminan directamente sobre el árbol dendrítico de
las células de Purkinje. Ambos sistemas aferentes convergentes en
las células de Purkinje son glutamatérgicos, por lo tanto excitadores,
mientras que las células de Purkinje son gabaérgicas, inhibidoras. A
su vez, las fibras musgosas y trepadoras durante su recorrido dan
colaterales axónicas excitadoras a las neuronas de los núcleos
profundos, que en su mayoría son excitadoras.
En definitiva, la función del cerebelo va a depender de la
integridad de los circuitos cerebelosos y de la correcta activación de
los diferentes receptores excitadores (de glutamato) e inhibidores (de
GABA) que existen en el cerebelo en conjunto. Además, se ha
observado que determinadas subunidades de canales de potasio, en
concreto las denominadas Kv3.1 y Kv3.3 son fundamentales para el

10
Tesis Doctoral Aranzazu Urresola Olabarrieta
correcto funcionamiento del cerebelo, ya que ratones knockout que
carecen de los genes que codifican estas subunidades de canales,
presentan un cuadro cerebeloso caracterizado por ataxia, temblor,
mioclonias y una sensibilidad extraordinaria al alcohol (Espinosa y
cols., 2001). Por otra parte, se ha demostrado que los ARN
mensajeros que codifican para Kv3.1 y Kv3.3 se expresan mucho en
las células granulares de la corteza cerebelosa (Weiser y cols, 1994;
Grigg y cols, 2000; Li y cols., 2001; Rudy y McBain, 2001).
En los últimos años se ha descubierto un gran número de
canales de potasio, la mayoría de los cuales se encuentran en el
sistema nervioso central. La subfamilia Kv3 forma canales
dependientes de voltaje que modulan la excitabilidad neuronal. En
los vertebrados, los genes de los Kv3 están relacionados con los genes
Shaw de Drosophila y están asignados a la subfamilia 3 de los Shaw.
Esta subfamilia de canales de potasio Kv3 consta de cuatro genes
independientes: Kv3.1, Kv3.2, Kv3.3 y Kv3.4. Tres de estos cuatro
genes (Kv3.1, Kv3.2 y Kv3.3) se expresan principalmente en el
sistema nervioso central (SNC) en forma de heteromultímeros.
Los canales que contienen las subunidades Kv3.1 y Kv3.3 se
caracterizan por presentar una activación e inactivación muy rápidas

11
Tesis Doctoral Aranzazu Urresola Olabarrieta
y por abrirse ante potenciales de membrana que oscilan entre –10mV
y 0mV. Por lo tanto, los canales de potasio con Kv3.1-Kv3.3
participan en la rápida repolarización que sigue tras un potencial de
acción, confiriendo a las células que las contienen las características
de disparo rápido, conocido como fast-spiking. Si estas circunstancias
se ven alteradas como ocurre en los ratones knockout que carecen de
dichas subunidades, las células afectadas son incapaces de disparar
potenciales de acción a alta frecuencia (Rudy y McBain, 2001),
además de presentar alteraciones de la plasticidad sináptica a corto
plazo, en concreto, a nivel de las sinapsis excitadoras entre las fibras
paralelas y las células de Purkinje (Sabatini y Regehr, 1997; Zucker
y Regehr, 2002).
En un estudio previo en colaboración con el Dr. Thomas Knöpfel
(Laboratory for Neuronal Circuit Dynamics, RIKEN, Brain Science
Institute, Japón), utilizamos ratones transgénicos que contenían el
gen que codifica la proteína fluorescente amarilla aumentada
(Enhanced Yellow-Fluorescent Protein, EYFP), bajo el control del
promotor del canal de potasio Kv3.1 (pKv3.1). Así describimos los
tipos celulares que expresaban fluorescencia (y por lo tanto la
subunidad Kv3.1) detectada con microscopía confocal en el cerebelo

12
Tesis Doctoral Aranzazu Urresola Olabarrieta
(Metzger y cols., 2002). Este fue el punto de partida del presente
trabajo.
CANALES DE POTASIO
En las últimas décadas se ha descubierto un gran número de
canales de potasio, la mayoría de los cuales se encuentran en el
sistema nervioso central (Pongs, 1992; Jan y Jan, 1997; Coetzee y
cols., 1999). Un importante objetivo de la investigación actual es
comprender el significado funcional de los mismos. Es por ello que la
caracterización de los canales de potasio formados por diferentes
subunidades en sistemas heterólogos de expresión y la identificación
celular y subcelular de los patrones de expresión en tejido, resulta
crucial para entender el papel de estos canales en la función
neuronal.
Las técnicas de ADN recombinante han demostrado la
existencia de dos grupos de genes que codifican canales de potasio
dependientes de voltaje. Estas dos familias de genes, ambas
pertenecientes a la superfamilia S4 de proteínas de canales iónicos,
son el gen homólogo de eag de Drosophila y el gen homólogo de
Shaker de Drosophila. Esta segunda familia de proteínas,
denominada familia de genes Sh, se divide en cuatro subgrupos.

13
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 1. Estructura de las subunidades de
un canal de potasio dependiente de voltaje.
En la parte superior, se representa la subunidad α
formada por 6 hélices transmembrana unidos por
lazos intracelulares y extracelulares, con dos
extremos amino y carboxilo intracelulares. Entre
las hélices 5 y 6 está el poro del canal, en la hélice
4 está el sensor de voltaje.
Cada uno está compuesto
por un homólogo de cuatro
genes muy relacionados:
Shaker, Shab, Shaw y Shal
(Perney y Kaczmarek, 1991;
Pongs 1992; Salkoff y cols.,
1992; Jan y Jan, 1997; Rudy y
cols., 1999), que corresponden
a las distintas subunidades α
que se han denominado Kv1,
Kv2, Kv3 y Kv4. Esta familia
de proteínas Kv codifican
subunidades que forman
poros de canales de potasio
dependientes de voltaje
tetraméricos que se clasifican
en distintos grupos según las
analogías de secuencia (Coetzee y cols., 1999; Gutman y cols., 2005).
Cada subunidad individual de Sh forma tetrámeros funcionales de
canales de potasio dependientes de voltaje, con una dependencia de
voltaje, cinética y farmacología propias, observadas al expresar las
subunidades en ovocitos de Xenopus laevis o en otro sistema de

14
Tesis Doctoral Aranzazu Urresola Olabarrieta
expresión heterólogo (Perney y Kaczmarek, 1991; Salkoff y cols.,
1992; Rudy y cols., 1999). El centro tetramérico de estas subunidades
α puede estar compuesto por subunidades idénticas o por una
combinación de diferentes subunidades (Sheng y cols., 1993; Scott y
cols., 1994) (Figura 1).
Las subunidades de la misma familia Sh, pero no de diferentes
subfamilias, suelen formar canales heteroméricos, sugiriendo que
cada conjunto de subunidades forma un sistema de canales
diferentes (Coetzee y cols., 1999). Los canales heteroméricos, a
menudo, presentan propiedades intermedias a las que presentarían
esas subunidades individualmente formando homómeros (Christie y
cols., 1990; Isacoff y cols., 1990; McCormack y cols., 1990;
Ruppersberg y cols., 1990; Covarrubias y cols., 1991; Weiser y cols.,
1994). Además, los canales formados por alguna subunidad Sh
pueden interactuar con otras subunidades, viéndose modificada de
esta forma las propiedades de los canales (Rettig y cols., 1994; Scott
y cols., 1994). Teniendo en cuenta todos estos factores, estas
subunidades pueden dar lugar a la formación de un amplio número
de canales de potasio dependientes de voltaje funcionalmente
distintos. Esta gran diversidad proporciona un mecanismo que
permite lograr sutiles diferencias en la integración neuronal.

15
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 2. En los vertebrados, los genes de los canales de potasio activados
por voltaje Kv3 están relacionados con los genes Shaw de Drosophila y están
asignados a la subfamilia 3 de entre los Shaw, también denominados ShIII.
Los canales de potasio son relativamente desconocidos, a pesar
de que todas las proteínas Sh y eag que se han descrito se
encuentran en neuronas, constituyendo un grupo muy diverso de
canales iónicos (Figura 2). Esta diversidad es uno de los factores
principales que contribuye a las dinámicas propiedades
electrofisiológicas y a la especificidad funcional de las acciones
moduladoras de los neurotransmisores (Hille y cols., 1999).
Entre las subunidades que han atraído mayor atención figuran
las de la familia Kv3, ya que forman canales dependientes de voltaje
con propiedades electrofisiológicas específicas en sistemas

16
Tesis Doctoral Aranzazu Urresola Olabarrieta
heterólogos de expresión, lo cual les confiere un papel especial en la
modulación de la excitabilidad neuronal (Vega-Sáenz de Miera y
cols., 1994; Wang y cols., 1998a, Erisir y cols., 1999; Rudy y cols.,
1999; Rudy y McBain, 2001). El canal Kv3 se distingue de los demás
canales Kv por su rango positivo de activación y por su rápida
cinética de desactivación (Rudy y cols., 1999). La apertura de estos
canales Kv3 no tiene lugar hasta que el potencial de membrana es
más positivo que -10 mV. En la repolarización de la membrana, los
canales Kv3 se cierran muy rápidamente, con rangos de
desactivación 10 veces más rápidos que otros canales Kv (Grissmer y
cols., 1994; Coetzee y cols., 1999). En definitiva, los canales que
contienen subunidades Kv3 participan en la rápida repolarización
que sigue tras un potencial de acción, confiriendo a las células que
las contienen las características de disparo rápido, conocido como
fast-spiking. Según esto, se ha sugerido que los canales que
contienen subunidades Kv3 juegan un papel fundamental en la
reducción de la duración de los potenciales de acción, tanto en el
establecimiento de la frecuencia de los trenes de potenciales de
acción (controlando la resistencia entre espigas), como en la
modulación presináptica de la liberación de neurotransmisor
limitando, en definitiva, el transcurso postsináptico de la
despolarización inducida por la apertura del receptor-canal y

17
Tesis Doctoral Aranzazu Urresola Olabarrieta
haciendo posible una transmisión sináptica fásica de alta frecuencia.
Esta subfamilia de canales Kv3 consta de cuatro genes
independientes: Kv3.1, Kv3.2, Kv3.3 y Kv3.4 (Chandy y Gutman,
1993), que codifican al menos 12 tránscritos generados por
ensamblaje alternativo: Kv3.1a, Kv3.1b, Kv3.2a, Kv3.2b, Kv3.2c,
Kv3.2d, Kv3.3a, Kv3.3b, Kv3.4a, Kv3.4b, Kv3.4c, Kv3.4d (Vega-
Sáenz de Miera y cols., 1994; Rudy y cols., 1999; Rudy y McBain,
2001). Tres de los cuatro genes estudiados (Kv3.1, Kv3.2 y Kv3.3) se
expresan principalmente en el sistema nervioso central (Rudy y cols.,
1999), mientras que los tránscritos de Kv3.4 son más abundantes en
el músculo esquelético (Weiser y cols., 1994; Abott y cols., 2001;
Vullhorst y cols., 2001). Los datos obtenidos de la coexpresión de
algunas de dichas subunidades, sugieren que las más comunes
forman heteromultímeros en el SNC (Hernández-Pineda y cols.,
1999; Rudy y cols., 1999).
Por otra parte, la farmacología de los canales de la subfamilia
Kv3 se caracteriza por una relativa sensibilidad a 4-aminopiridina
(4-AP) e iones de tetraetilamonio (TEA) (Baranauskas y cols., 1999;
Erisir y cols., 1999; Hendriks y cols., 1999). De los tres subtipos que
se encuentran en el cerebro, el patrón de expresión de Kv3.1 y Kv3.2

18
Tesis Doctoral Aranzazu Urresola Olabarrieta
es en gran parte complementario, mientras que Kv3.1 y Kv3.3 a
menudo se coexpresan en muchas regiones cerebrales (Weiser y cols.,
1994; Grigg y cols., 2000). El gen codificante para Kv3.1 exhibe un
patrón de expresión restringido según el tejido. En experimentos
llevados a cabo en líneas celulares, se ha observado que la
transcripción está regulada por calcio, AMP cíclico y cascadas de
transducción generadas por factores de crecimiento (Gan y cols.,
1996).
Como ha sido mencionado más arriba, los canales de potasio
dependientes de voltaje Kv3.1 y Kv3.3 están ampliamente
coexpresados en el SNC (Drewe y cols., 1992; Perney y cols., 1992;
Rudy y cols., 1992; Goldman-Wohl y cols., 1994; Lenz y cols., 1994;
Weiser y cols., 1994, 1995; Du y cols., 1996; Perney y Kaczmarek,
1997; Sekirnjak y cols., 1997; Baranauskas y cols., 1999; Hernández-
Pineda y cols., 1999; Rudy y cols., 1999; Rudy y McBain, 2001), en
especial en las áreas implicadas en el control de la actividad motora
y, por tanto, también en poblaciones neuronales del cerebelo (ver
más abajo). Este hecho revela el gran potencial que tienen para
formar heterómeros entre los productos de estos genes. Además se ha
visto que los ratones knockout carentes de los genes que codifican
Kv3.1 y Kv3.3 presentan ataxia severa, movimientos temblorosos,

19
Tesis Doctoral Aranzazu Urresola Olabarrieta
mioclonía e hipersensibilidad al alcohol, aunque no un déficit de
aprendizaje o memoria (Espinosa y cols., 2001). Sin embargo, los
ratones knockout para un solo gen tienen alteraciones fisiológicas,
pero mucho más sutiles (Chan 1997; Ho y cols., 1997; Joho y cols.,
1999; Sánchez y cols., 2000; Porcello y cols., 2002; Macica y cols.,
2003).
Expresión de Kv3.1 y Kv3.3 detectada por hibridación in
situ
La distribución subcelular de los tránscritos de Shaw (ShIII) ha
sido analizada por técnicas de Northern blot e hibridación in situ.
Los estudios de hibridación in situ en el SNC han mostrado que
algunos ARNm de Shaw son tan abundantes como los tránscritos de
otros canales de potasio (Weiser y cols., 1994). La expresión de los
ARNm de los cuatro genes de la subfamilia Shaw, tiene lugar en
poblaciones neuronales específicas de proyección y de circuitos
locales (Perney y cols., 1992; Rudy y cols., 1992; Weiser y cols., 1994).
Cada gen de ShIII exhibe un patrón de expresión diferente,
pero una gran mayoría de las poblaciones neuronales que expresan
los tránscritos Kv3.1 también expresan el ARNm para Kv3.3. Los
genes que codifican Kv3.1 y Kv3.3 son, respectivamente, Kcnc1 y

20
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 3. Distribución de los ARNm de los canales de potasio activados por voltaje
Kv3 en el cerebro de los roedores. Kv3.1 y Kv3.3 presentan una amplia distribución en el
sistema nervioso central, incluidos el cerebelo y los ganglios basales implicados en el control y
la modulación de la actividad motora. Nótese la importante coexpresión de Kv3.1 y Kv3.3 en las
células granuleres del cerebelo. Rudy y cols. (1999).
Kcnc3. En general, el ARNm de Kv3.1 se expresa en gran cantidad
en el hipocampo y tálamo, mientras que los niveles del ARNm de
Kv3.3 son elevados en el tronco del encéfalo (Weiser y cols., 1994)
(Figura 3).
Ambos tránscritos se expresan ampliamente en el cerebelo.
Aquí, los ARN mensajeros que codifican para Kv3.1 y para Kv3.3 se
expresan en la corteza cerebelosa y en los núcleos cerebelosos
profundos. En las células granulares de la corteza se expresan

21
Tesis Doctoral Aranzazu Urresola Olabarrieta
principalmente ambos ARNm, sin embargo, en las células de
Purkinje sólo se expresa el ARNm codificante para Kv3.3 (Goldman-
Wohl y cols., 1994; Ozaita y cols., 2002; Sekirnjak y cols., 1997;
Weiser y cols., 1994, 1995) (Figura 3). En las células de la capa
molecular se observan ambos ARNm, pero sin ser tan abundantes
como en las demás poblaciones neuronales de la corteza. Los núcleos
profundos del cerebelo muestran también un patrón de alta
expresión de los tránscritos de Kv3.1 y Kv3.3 (Weiser y cols., 1994).
Por tanto, estos dos ARNm tienen un patrón de distribución que, si
bien a veces es similar, no siempre tiene por qué coincidir (Figura 3).
ARN mensajero codificante para Kv3.1
El ARNm que codifica para Kv3.1 se expresa en el cerebelo del
ratón y de la rata adultos (Drewe y cols., 1992; Perney y cols., 1992;
Rudy y cols., 1992; Weiser y cols., 1994, 1995; Du y cols., 1996;
Perney y Kaczmarek, 1997; Sekirnjak y cols., 1997) (Figura 3). En
concreto, el ARNm de Kv3.1 presenta un patrón heterogéneo (Lenz y
cols., 1994).
Así, los niveles más elevados de mensajero se expresan en el
cerebelo, aunque también en el globo pálido, subtálamo y en la
porción reticular de la sustancia negra. Muchos núcleos talámicos,

22
Tesis Doctoral Aranzazu Urresola Olabarrieta
pero sobre todo el núcleo reticular talámico, muestran una fuerte
señal de hibridación, al igual que algunas poblaciones de
interneuronas de la corteza cerebral e hipocampo (Perney y cols.,
1992). También se detecta la expresión del canal Kv3.1 en núcleos
del tronco del encéfalo, tales como el colículo inferior y los núcleos
cocleares y vestibulares y, sobre todo, en el núcleo medial del cuerpo
trapezoide (Grigg y cols., 2000) (Figura 3).
El ARNm que codifica para Kv3.1 se expresa mucho en las
capas granulares externa e interna desde los períodos postnatales
tempranos. Además, Kv3.1 se expresa en las fibras paralelas de
células granulares de dos días postnatal en cultivos de
microexplantes. Todo ello pone de manifiesto que estos canales
predominan en las células granulares inmaduras (Shibata y cols.,
1999). Otros experimentos realizados en neuroblastos cultivados del
núcleo magnocelular auditivo del romboencéfalo del pollo,
demuestran la existencia de corrientes de potasio muy sensibles a 4-
aminopiridina y tetraetilamonio (lo que indica que son del tipo
Kv3.1) detectables desde el segundo día embrionario y a lo largo de
toda la embriogénesis (Hendriks y cols., 1999).
Los canales de potasio Kv3.1 parecen tener un papel destacado

23
Tesis Doctoral Aranzazu Urresola Olabarrieta
durante el desarrollo neuronal temprano y en la maduración, ya que
la actividad neuronal espontánea es fundamental en la modulación
de la excitabilidad neuronal (Gan y Kaczmarek, 1998). En este
sentido, se ha constatado una regulación de la isoforma Kv3.1b en la
subpoblación de interneuronas hipocampales que expresan
parvalbúmina, apareciendo ambas proteínas de forma simultánea.
Además, la proteína Kv3.1b predominante se expresa de manera
selectiva en los somas, dendritas proximales y axones de esas células
localizadas en la capa piramidal o en su proximidad (Du y cols.,
1996). De hecho, los registros electrofisiológicos en interneuronas de
la capa piramidal se asemejan a los de los canales formados por esta
subunidad en otras neuronas. La subunidad Kv3.1b en hipocampo es
detectado por primera vez en el octavo día postnatal (P8). Esta señal
alcanza un máximo de intensidad en P14 y se mantiene hasta P40.
Por otra parte, el aumento de las corrientes de potasio Kv en
neuronas medulares de embriones de Xenopus durante el desarrollo,
es crítico para la maduración de la excitación y de la onda del
potencial de acción. En este sentido, se ha demostrado que la
inyección de oligonucleótidos antisentido de Kv3.1 en estos
embriones reduce de forma espectacular los niveles de ARNm de
Kv3.1 en la médula espinal en desarrollo, lo cual acarrea una

24
Tesis Doctoral Aranzazu Urresola Olabarrieta
reducción de la activación de la corriente de potasio Kv en esas
mismas neuronas cultivadas maduras. La corriente en estas
neuronas se asemeja a la existente en un estadio temprano de la
diferenciación, previo a la aparición del ARNm codificante para
Kv3.1 (Vincent y cols., 2000).
El gen Kv3.1 codifica dos proteínas distintas, las subunidades
Kv3.1a y Kv3.1b, formas corta y larga respectivamente (Luneau y
cols., 1991). Las dos proteínas comparten homología de secuencia en
su terminal amino, pero difieren en el terminal carboxilo (Perney y
cols., 1992). Ambas isoformas expresadas en ovocitos comparten
similares propiedades electrofisiológicas y farmacológicas (Schröter y
cols., 1991; Critz y cols., 1993). El ARNm que codifica para Kv3.1a es
menos abundante que la isoforma Kv3.1b. De todos modos, los
patrones de expresión de las variantes a y b son similares y está
claro que ambas coexisten en varios tipos neuronales. Generalmente,
Kv3.1b constituye la variante predominante en el cerebro adulto,
mientras que Kv3.1a está sobre todo en neuronas embrionarias y
perinatales (Perney y cols., 1992; Du y cols, 1996) y se expresa
predominantemente en axones. Existen excepciones como las células
mitrales del bulbo olfatorio y las neuronas mesencefálicas del

25
Tesis Doctoral Aranzazu Urresola Olabarrieta
trigémino, en las que el Kv3.1a se expresa en las membranas de
somas y dendritas (Ozaita y cols., 2002).
ARNm codificante para Kv3.3
La distribución del ARNm que codifica para Kv3.3 es amplia
por todo el cerebro, siendo especialmente abundante en el tronco del
encéfalo y cerebelo (Figura 3).
La señal de hibridación del ARNm de Kv3.3 es heterogénea,
siendo especialmente fuerte en las astas ventral y dorsal de la
médula espinal, y en las células de Purkinje, así como en las células
granulares de la corteza cerebelosa y en las poblaciones neuronales
de los núcleos cerebelosos profundos (Figura 3). También se detecta
expresión del ARNm para Kv3.3 en diversos centros del tronco del
encéfalo, como la zona incerta, los núcleos oculomotores, el colículo
inferior, el núcleo rojo, los núcleos pontinos, el núcleo motor del
trigémino y los núcleos vestibulares (Figura 3). Sin embargo, su
distribución es mucho menor en otras zonas del encéfalo e incluso, a
veces, inexistente. Este es el caso de los ganglios basales (putamen y
globo pálido), septum, tálamo dorsal, epitálamo e hipocampo (Weiser
y cols., 1994; Chang y cols., 2007).

26
Tesis Doctoral Aranzazu Urresola Olabarrieta
Distribución de las proteínas Kv3.1 y Kv3.3 en el SNC
Los canales de potasio juegan un papel primordial en la
regulación de diversos aspectos de la excitabilidad neuronal. Estos
canales están especialmente bien diseñados para una gran variedad
de funciones, debido a la gran diversidad de tipos de canales que
existen. Esta diversidad contribuye a la habilidad de algunas
neuronas (y posiblemente de diferentes compartimentos de una
misma neurona) de responder de manera única a un estímulo
recibido. La integración neuronal depende de la respuesta local de
las aferencias que se encuentran espacialmente segregadas y que
llegan a la célula, así como de la comunicación que se da entre estos
centros de integración con el axón. Por tanto, las implicaciones
funcionales de los canales de potasio varían dependiendo de su
precisa localización en la superficie neuronal (Weiser y cols., 1995).
Los canales de potasio dependientes de voltaje Kv3.1 y Kv3.3 están
ampliamente expresados en el cerebro, en especial en las áreas
implicadas en el control de la actividad motora y en áreas conocidas
por regular los estados de excitación (Espinosa y cols., 2004), como ya
hemos descrito.
Las proteínas Kv3 se distribuyen a lo largo de todo el SNC y
generalmente se coexpresan con otras proteínas Kv3, dando lugar a

27
Tesis Doctoral Aranzazu Urresola Olabarrieta
patrones de expresión similares. Este hecho sugiere que las
proteínas Kv3 forman frecuentemente parte de los complejos de
canales heteromultiméricos, como es el caso de las subunidades
Kv3.1 y Kv3.3 que a menudo se coexpresan. Sin embargo, la
distribución de las proteínas Kv3.1 y Kv3.3 a lo largo de las
estructuras del cerebro no es idéntica, en consonancia con los
patrones de expresión de sus correspondientes tránscritos de ARNm
(Weiser y cols., 1994).
Proteína Kv3.1
Estudios inmunocitoquímicos en secciones de cerebro de rata
(Weiser y cols., 1995; Hernández-Pineda y cols., 1999; Rudy y
McBain, 2001) han demostrado la localización selectiva de Kv3.1b en
células granulares del cerebelo, neuronas de proyección de los
núcleos cerebelosos profundos, porción reticular de la sustancia
negra, globo pálido y tálamo ventral (núcleo reticular talámico,
núcleo geniculado ventral lateral y zona incerta). La isoforma Kv3.1b
se encuentra además presente en varios centros nerviosos implicados
en el procesamiento de señales auditivas, como son el colículo
inferior, núcleos del lemnisco lateral, la oliva superior, el núcleo
medial del cuerpo trapezoide y algunas partes de los núcleos
cocleares (Weiser y cols., 1995; Härtig y cols., 2001). La

28
Tesis Doctoral Aranzazu Urresola Olabarrieta
inmunorreactividad frente a Kv3.1 también se localiza en otros
grupos neuronales del tronco del encéfalo (núcleo oculomotor común,
núcleos pontinos, núcleo reticulotegmental del puente, núcleos
vestibular y trigeminal y formación reticular) (Metzger y cols., 2002).
El uso de técnicas de inmunofluorescencia de doble marcado con
anticuerpos frente a la proteína Kv3.1b y la proteína ligadora de
calcio parvalbúmina o el neuropéptido somatostatina, ha puesto de
manifiesto que Kv3.1 está localizado en aproximadamente el 80% de
las interneuronas inhibidoras gabaérgicas que contienen
parvalbúmina (Weiser y cols., 1995; Du y cols., 1996; Sekirnjak y
cols., 1997) caracterizadas por ser de disparo rápido. En el
hipocampo, Kv3.1b se localiza en las células en cesto, aunque
también se encuentra en otras interneuronas positivas frente a
calbindina en el estrato oriens, y en algunas células que no contienen
proteínas ligadoras de calcio (Sekirnjak y cols., 1997). En la
neocorteza, la mayoría de las interneuronas positivas frente a
parvalbúmina que expresan el canal Kv3.1b (Weiser y cols., 1994,
1995; Du y cols., 1996), corresponden a células bipolares, en cesto y
en candelabro (Sekirnjak y cols., 1997). Por el contrario, Kv3.1b no se
encuentra en interneuronas positivas frente a la somatostatina.

29
Tesis Doctoral Aranzazu Urresola Olabarrieta
Finalmente, no todas las neuronas Kv3.1b positivas tienen el
canal en sus axones. Por ejemplo, a pesar de que muchas poblaciones
neuronales están fuertemente marcadas (como los núcleos pontinos,
el núcleo reticulotegmental del puente y la formación reticular) que
proyectan fibras musgosas al cerebelo, no hay evidencias de marcado
en estas fibras.
Localización subcelular de Kv3.1
La subunidad Kv3.1b se localiza predominantemente en
membranas celulares de somas y axones (especialmente en campos
axónicos terminales), pero también en arborizaciones dendríticas (Du
y cols., 1996; Weiser y cols., 1995) y en protrusiones espinosas
adyacentes a membranas postsinápticas (Wang y cols., 1998b).
Mediante el empleo de técnicas inmunocitoquímicas para
microscopía electrónica, se ha observado que la proteína Kv3.1 está
concentrada en núcleos auditivos del tronco del encefálo, donde
aparece delineando los cuerpos celulares y el neuropelo. También se
encuentra asociada al retículo endoplásmatico y a cisternas del
aparato de Golgi. En el núcleo medial del cuerpo trapezoide, además
del habitual marcado en somas celulares, procesos dendríticos y
espinas, se ha observado marcado en las terminales presinápticas,

30
Tesis Doctoral Aranzazu Urresola Olabarrieta
incluidos los cálices de Held (Elezgarai y cols., 2003). La localización
dual de Kv3.1, tanto presináptica como postsináptica, sugiere que
esta subunidad participa en la modulación de la transmisión
sináptica rápida en la vía auditiva. En el caso de la neocorteza y del
hipocampo, las células piramidales no son inmunorreactivas (como
ya se observaba a nivel de microscopía de luz), pero estas neuronas
aparecen delineadas por terminales presinápticas de interneuronas
que expresan Kv3.1b (Sekirnjak y cols., 1997). En el cerebelo, sólo las
células granulares contienen la subunidad que, al igual que las
interneuronas neocorticales e hipocampales, presentan la
inmunotinción distribuida en las membranas somáticas, axónicas y
en el citoplasma adyacente, así como en las porciones más
proximales de los procesos dendríticos (Sekirnjak y cols., 1997). Sin
embargo, existen pruebas electrofisiológicas y farmacológicas de la
presencia de canales Kv3 en las terminales sinápticas de las células
en cesto alrededor de los somas de las células de Purkinje (Southan y
Robertson, 2000).
Proteína Kv3.3
Su patrón de distribución más destacado se encuentra en el
tronco del encéfalo y la corteza cerebelosa (Chang y cols., 2007). La
proteína Kv3.3 se localiza en las células de Purkinje y en las células

31
Tesis Doctoral Aranzazu Urresola Olabarrieta
granulares. Las estructuras cerebrales en general muestran una
inmunorreactividad más débil. Por otro lado, se observa marcaje de
moderado a fuerte en el bulbo olfatorio, el hipocampo y el núcleo
reticular talámico (Chang y cols., 2007). Con frecuencia, la
subunidad Kv3.3 aparece tanto en el soma como en el neuropelo de la
misma estructura. La inmunorreactividad para Kv3.3 también es
muy pronunciada en la formación reticular, en los núcleos motores
troncoencefálicos y en la médula espinal (Chang y cols., 2007).
Además, Kv3.3 se localiza ampliamente en el tronco del encéfalo, del
mismo modo que la proteína Kv3.1b (Weiser y cols., 1995; Li et al.,
2001), sobre todo en los centros auditivos (Chang y cols., 2007). Por
tanto, al igual que en el cerebro anterior y el cerebelo, Kv3.3 y
Kv3.1b pueden ser componentes de un canal heteromultímero en
muchas de estas poblaciones neuronales.
La proteína Kv3.3 está por lo general asociada a somas y
axones. Casi todas las poblaciones neuronales del cerebro anterior y
mesencéfalo que contienen Kv3.3, corresponden al sistema inhibidor
gabaérgico.

32
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 4. Maquinaria que integra la unidad neuronal del cerebelo. El cerebelo
contiene numerosos módulos, cada uno de los cuales está integrado por circuitos
neuronales uniformemente estructurados. En el dibujo aparecen las conexiones
neuronales más representativas de la unidad cerebelosa. 5-HT: fibras serotoninérgicas;
BC: célula en cesto; CN/VN núcleos cerebelosos profundos/núcleos vestibulares; Go:
célula de Golgi; Gr: célula granular; IO: oliva inferior; Lg: célula de Lugaro; MF: fibra
musgosa; N-C; proyección núcleo-cortical; N-O: proyección núcleo-olivar; PCN: núcleo
precerebeloso; Pd: fibra peptidérgica; pRN: porción parvocelular del núcleo rojo; R-O:
proyección rubro-olivar; SC: célula estrellada; UB: célula unipolar en pincel. Ito
(2008).
CIRCUITOS NEURONALES DEL CEREBELO
Las conexiones neuronales del cerebelo están constituidas por
axones aferentes, que transmiten la información de otras partes del
SNC al cerebelo; circuitos cerebelosos intrínsecos corticales y
nucleares que integran y procesan la información, y axones eferentes
que transmiten la información procesada a otras partes del SNC
(Figura 4).

33
Tesis Doctoral Aranzazu Urresola Olabarrieta
Las fibras aferentes alcanzan la corteza cerebelosa tras dar
colaterales para los núcleos cerebelosos profundos y los núcleos
vestibulares. A su vez, la información es procesada en los circuitos
intrínsecos de la corteza cerebelosa, y el resultado es enviado por los
axones de las células de Purkinje a los núcleos profundos. En estos
núcleos la información es de nuevo procesada, surgiendo a partir de
ellos las fibras eferentes del cerebelo tanto en dirección ascendente
(tálamo, corteza cerebral), como descendente (médula espinal)
(Figura 4).
Los circuitos inhibidores de la corteza cerebelosa están
constituidos por tres tipos fundamentales de interneuronas: las
células de Golgi, las células estrelladas y las células en cesto (Figura
4). Pueden actuar directamente sobre las células de Purkinje (como
lo hacen las células estrelladas y las células en cesto), o
indirectamente a través de las células granulares, como en el caso de
las células de Golgi. Todas estas interneuronas utilizan GABA como
neurotransmisor inhibidor. Las células estrelladas y las células en
cesto son estimuladas por las fibras paralelas, y son las encargadas
de modular la activación de las células de Purkinje por las fibras
trepadoras produciendo un fenómeno de inhibición lateral. Esta
inhibición lateral hace más precisa la señal que llega a las células de

34
Tesis Doctoral Aranzazu Urresola Olabarrieta
Purkinje, de la misma manera que otros mecanismos de inhibición
lateral acentúan el contraste de las señales en otros muchos circuitos
neuronales de sistema nervioso.
Las células de Golgi reciben estímulos excitadores de las fibras
paralelas y, en menor cantidad, por las células de Lugaro (Figura 4).
Actúan a nivel de los glomérulos cerebelosos haciendo sinapsis Gray
tipo II (inhibidoras) sobre las dendritas de las células granulares.
Mediante estas sinapsis modulan la activación de las células
granulares por las fibras musgosas y, por consiguiente, regulando la
actividad de las células de Purkinje. Así, las células de Golgi crean
un circuito de retroalimentación negativa para las células
granulares.
El circuito funcional básico del cerebelo queda constituido por
dos arcos: uno principal o excitador, que pasa por los núcleos
profundos, y otro secundario o inhibidor, que pasa por la corteza
cerebelosa y regula al anterior (Figura 5). Este circuito se repite
unas 30 millones de veces en todo el cerebelo y está formado por las
células de Purkinje y las neuronas nucleares de proyección
correspondientes, así como las interneuronas relacionadas con ellas.

35
Tesis Doctoral Aranzazu Urresola Olabarrieta
Arco principal
El arco principal está constituido por las ramas colaterales de
las fibras musgosas y trepadoras, que terminan en las neuronas de
los núcleos profundos. Los axones de las neuronas de proyección de
los núcleos profundos salen del cerebelo a través de los pedúnculos
cerebelosos, para terminar en diferentes núcleos del tronco del
encéfalo y en el tálamo (Figura 5).
En los núcleos profundos hay sinapsis axosomáticas y, sobre
todo, sinapsis axodendríticas entre terminales sinápticas de las
colaterales axónicas de las fibras musgosas y trepadoras (elemento
presináptico) y una dendrita de una neurona de proyección o una
interneurona de los núcleos profundos (elemento postsináptico). Las
fibras musgosas y las fibras trepadoras usan el neurotransmisor
glutamato, aunque también pueden tener otros neurotransmisores.
Desde el punto de vista funcional, los núcleos profundos del
cerebelo poseen dos tipos básicos de neuronas de proyección: unas
neuronas pequeñas gabaérgicas (inhibidoras) que mandan su axón
hacia la oliva inferior, y otras neuronas glutamatérgicas
(excitadoras) que mandan sus axones a otros centros nerviosos.

36
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 5. Organización sináptica del módulo
básico de circuito cerebeloso. Las fibras musgosas
y trepadoras transmiten eferencias del cerebelo
mediante un arco excitador principal que pasa por los
núcleos profundos. Este arco está regulado por otro
inhibidor secundario que pasa a través de la corteza
cerebelosa. Kandel y cols., Principios de Neurociencia.
Mc Graw-Hill-Interamericana (2001).
Las neuronas de proyección de los núcleos profundos en
condiciones normales disparan permanentemente, a una frecuencia
de más de 100
potenciales de acción
por segundo. Esta
frecuencia puede
modularse al alza o a
la baja, dependiendo de
las señales excitadoras
e inhibidoras que
lleguen a la neurona.
Las señales excitadoras
provienen sobre todo de
las colaterales axónicas
de las fibras musgosas
y trepadoras, mientras
que las señales inhibidoras proceden de los axones de las células de
Purkinje, que forman parte del arco secundario. El equilibrio entre
estos dos efectos es ligeramente favorable a la excitación, lo que
explica por qué la frecuencia de descargas de las neuronas de
proyección se mantiene relativamente constante a un nivel moderado
de estimulación contínua.

37
Tesis Doctoral Aranzazu Urresola Olabarrieta
Arco secundario
El arco secundario pasa a través de la corteza cerebelosa y está
constituido en torno a una pieza neuronal fundamental: la célula de
Purkinje. En la célula de Purkinje terminan dos tipos de circuitos:
los circuitos excitadores o principales, que son los que la estimulan, y
los circuitos inhibidores, formados por interneuronas inhibidoras.
Finalmente, los axones de las células de Purkinje proyectan sobre las
neuronas de los núcleos cerebelosos y vestibulares, ejerciendo sobre
ellos una acción inhibidora mediante sinapsis gabaérgicas. De esta
forma se modula y regula el arco principal excitador (Figura 5).
Las células de Purkinje se estimulan por dos vías distintas:
mediante las fibras trepadoras (vía directa) y las fibras musgosas
(vía indirecta) (Figura 5). Las fibras trepadoras, al terminar sobre el
soma y el árbol dendrítico de las células de Purkinje, producen una
estimulación directa y muy específica mediante sinapsis Gray tipo I
(excitadora) que utilizan el neurotransmisor glutamato. Al formar
múltiples contactos con cada célula de Purkinje, una sola fibra
trepadora produce una acción excitadora mucho más eficaz que las
fibras musgosas.

38
Tesis Doctoral Aranzazu Urresola Olabarrieta
Las fibras musgosas no actúan de forma directa sobre las
células de Purkinje, sino que lo hacen a través de las células
granulares excitadoras. A nivel del glomérulo cerebeloso, las fibras
musgosas hacen sinapsis excitadoras sobre las dendritas de las
células granulares y los impulsos son vehiculizados por las fibras
paralelas hasta alcanzar las espinas dendríticas de las células de
Purkinje. Las fibras hacen sinapsis excitadoras asimétricas, que
acumulan vesículas esféricas con glutamato en su interior. En
conjunto, las fibras musgosas actúan sobre las células de Purkinje
con una amplia convergencia y divergencia, estableciendo por tanto
conexiones más inespecíficas que las fibras trepadoras.
Las células de Purkinje no cumplen el principio de que todos los
potenciales de acción producidos por una neurona son iguales, ya que
presentan dos tipos de potenciales de acción distintos dependiendo
de la vía de estimulación. Si se estimulan directamente a través de
las fibras trepadoras, generan una despolarización prolongada y un
potencial de acción de pico complejo con una frecuencia de descarga
de 3 o 4 herzios. La estimulación de las células de Purkinje a través
de la vía indirecta de las fibras musgosas, genera en ellas un
potencial de acción breve denominado pico sencillo, con una
frecuencia de descarga de 100 a 200 herzios. Para generar un pico

39
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 6. Circuito cerebeloso simplificado (sin interneuronas ni neuronas inhibidoras
de los núcleos profundos -DCN- proyectantes a la oliva inferior) ilustrando la
dirección del flujo de información y la localización de los canales Kv3. Las aferencias
cerebelosas musgosas (mossy fibers) y trepadoras (climbing fibers) estimulan las neuronas de
DCN y la corteza, donde las fibras trepadoras estimulan las células de Purkinje, y las células
granulares excitadas por fibras musgosas hacen lo propio sobre las células de Purkinje a través
de las fibras paralelas. Las células de Purkinje, por su parte, inhiben las neuronas de los núcleos
profundos propiciando la recuperación de la excitación. Hurlock y cols. (2009).
sencillo, es necesaria la suma temporal y espacial de la estimulación
producida por varias fibras paralelas. Por lo tanto en conjunto, la
información aportada por los dos tipos de fibras extrínsecas
(trepadoras y musgosas) que llegan al cerebelo, es diferente y es
procesada de manera distinta.
Localización de Kv3.1 y Kv3.3 en la corteza cerebelosa
Kv3.1b y Kv3.3 se localizan ampliamente en la corteza
cerebelosa, como hemos visto anteriormente, donde colocalizan en
varios tipos celulares (Figura 6).

40
Tesis Doctoral Aranzazu Urresola Olabarrieta
La inmunorreactividad para Kv3.3 en la capa molecular
coincide con el patrón de marcado de la calbindina en dendritas
proximales, lo que concuerda con la localización de Kv3.3 en
dendritas de células de Purkinje. En la parte más distal de sus
arborizaciones dendríticas, Kv3.3 sólo se colocaliza parcialmente con
la calbindina (Martina y cols., 2003). Por otra parte, las fibras
paralelas expresan mucho Kv3.3 (McMahon y cols., 2004), lo que
concuerda con la expresión del ARNm en las células granulares
(Goldman-Wohl y cols., 1994; Weiser y cols., 1994; MacMahon y cols.,
2004).
Existen evidencias de la localización de Kv3.1b en axones de la
corteza cerebelosa. La capa molecular es muy inmunopositiva para
Kv3.1b (Alonso-Espinaco y cols., 2008). La intensidad del marcado es
mayor que la observada en la capa granular, a pesar de ser donde se
concentra el ARNm (Weiser y cols., 1994). Este marcado se debe al
entramado de fibras paralelas, los axones de las células granulares
(Alonso-Espinaco y cols., 2008). Las colaterales de las fibras
paralelas también son inmunopositivas para Kv3.1b produciendo un
patrón de finas líneas horizontales. Las células en cesto y
estrelladas, así como las dendritas de las células de Purkinje de la
capa molecular son inmunonegativas para Kv3.1b (McMahon y cols.,
2004).

41
Tesis Doctoral Aranzazu Urresola Olabarrieta
El marcado de Kv3.3 se observa en las capas molecular, de
Purkinje y granular del cerebelo, pero es especialmente sobresaliente
en los somas de las células de Purkinje y en la capa molecular
(McMahon y cols., 2004; Chang y cols., 2007; Alonso-Espinaco y cols.,
2008). Los axones de las células de Purkinje tienen este canal en su
trayecto por la capa granular en dirección a la sustancia blanca
cerebelosa. Kv3.3 también se localiza en porciones proximales de las
dendritas primarias de las células de Purkinje (Chang y cols., 2007).
Es posible que las fibras paralelas contengan Kv3.3, dada su
presencia en los somas de las células granulares. Este es un objetivo
de la presente Tesis Doctoral.

42
Tesis Doctoral Aranzazu Urresola Olabarrieta
HIPÓTESIS DE TRABAJO

43
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 7. Trayectorias de la marcha de ratones
silvestres (WT1949) y de ratones que carecen de Kv3.1 y
Kv3.3 (DKO1684). Obsérvese el aumento de las fluctuaciones
respecto al centro, indicando una ataxia severa en los ratones
DKO. Matsukawa y cols. (2003).
HIPÓTESIS DE TRABAJO
Como ya ha sido explicado, las subunidades de los canales de
potasio dependientes de voltaje Kv3.1 y Kv3.3 poseen un alto umbral
de activación que tiene lugar durante la fase ascendente del
potencial de acción, modelando así su repolarización (Rudy y
McBain, 2001). Los ratones que carecen de Kv3.1 y Kv3.3 presentan
deficiencias motoras (temblor, mioclonias, ataxia severa), y son muy
sensibles al alcohol (Espinosa y cols., 2001). Sin embargo, los ratones
que carecen sólo de Kv3.1 o Kv3.3 no exhiben alteraciones
manifiestas de la función motora (Chan, 1997; Ho y cols., 1997,
Sánchez y cols., 2000). En comparación con otras regiones cerebrales,
la expresión de los ARN mensajeros codificantes para Kv3.1 y Kv3.3

44
Tesis Doctoral Aranzazu Urresola Olabarrieta
A CB
1
FWHM
0.0
-0.4
F/F
[%
]
2 3
200 µm
D1.61.51.41.31.21.11.00.90.80.7
WT Kv3.1,3.3 -/-,+/- Kv3.1,3.3 -/-,-/-
E F
*
1
2
3
V VI
0.0
0.5
1.0
1.5
* * * * ** * * * *
WTKv3.1,3.3 -/-,+/-Kv3.1,3.3 -/-,-/-
1 ms
*
WT TM
FW
HM
[m
s]
* *
DKO
G
50 850 16500.5
1.0
1.5
2.0
FW
HM
[m
s]
Distance ( m)
WTKv3.1,3.3 -/-,-/-
Figura 8. Mapas en códigos de colores de la amplitud de los potenciales de
acción de un ratón silvestre (WT), de un ratón que carece los dos alelos de Kv3.1 y uno de
Kv3.3 (Kv3.1,3.3-/-,+/) y de un ratón que carece de los dos genes (Kv3.1,3.3-/-,-/-).
Matsukawa y cols. (2003).
es muy importante en la corteza cerebelosa, lo que induce a pensar
que al menos algunas de las anomalías motoras observadas en los
ratones que carecen Kv3.1 y Kv3.3 podrían ser debidas a una
disfunción cerebelosa.
Las células granulares de la corteza cerebelosa contienen
ambas subunidades Kv3.1 y Kv3.3 (Weiser y cols., 1994; Grigg y
cols., 2000; Li y cols., 2001; Rudy y McBain, 2001), por lo que es
posible que debido a ello y a sus características biofísicas similares,
se dé una situación de redundancia funcional, que explicaría la falta
de cambios fenotípicos severos en ratones que carecen solamente de
Kv3.1 o Kv3.3. Las alteraciones motoras severas mencionadas son la
consecuencia de la alteración de la transmisión sináptica entre fibras

45
Tesis Doctoral Aranzazu Urresola Olabarrieta
A CB
1
FWHM
0.0
-0.4
F/F
[%
]2 3
200 µm
D1.61.51.41.31.21.11.00.90.80.7
WT Kv3.1,3.3 -/-,+/- Kv3.1,3.3 -/-,-/-
E F
*
1
2
3
V VI
0.0
0.5
1.0
1.5
* * * * ** * * * *
WTKv3.1,3.3 -/-,+/-Kv3.1,3.3 -/-,-/-
1 ms
*
WT TM
FW
HM
[m
s]
* *
DKO
G
50 850 16500.5
1.0
1.5
2.0
FW
HM
[m
s]
Distance ( m)
WTKv3.1,3.3 -/-,-/-
Figura 9. E: Anchura de los potenciales de acción trazados versus la
distancia del electrodo de estimulación en preparación de corteza cerebelosa de
ratón silvestre (WT) y doble Kv3.1 y Kv3.3 knockout (Kv3.1,3.3-/-,-/-). Nótese un
pequeño aumento en la anchura del potencial de acción medido cuanto más
alejado del lugar de estimulación, lo que indica unas velocidades de conducción
no homogéneas de los potenciales de acción. F: Media ± SEM de la forma de los
potenciales de acción. G: Media ± SEM de la anchura de los potenciales de
acción. Matsukawa y cols. (2003).
paralelas y células de Purkinje, y dependen del número de alelos de
Kv3.1 y Kv3.3 suprimidos genéticamente (Matsukawa y cols., 2003).
(Figuras 7, 8, 9).
Según la localización subcelular, ambas subunidades estarían
implicadas en la fase de repolarización de los potenciales de acción.
La alteración de la repolarización de los potenciales de acción
axónicos afecta la capacidad de mantener el disparo de potenciales
de acción de alta frecuencia (Rudy y McBain, 2001), así como la
fiabilidad y plasticidad a corto plazo en las sinapsis de las fibras
paralelas con las células de Purkinje (Sabatini y Regehr, 1997;
Zucker y Regehr, 2002).

46
Tesis Doctoral Aranzazu Urresola Olabarrieta
En base a estos hallazgos fisiológicos, nos planteamos la
hipótesis de la existencia de una colocalización de Kv3.1 y Kv3.3 en
compartimentos subcelulares neuronales dispuestos en la capa
molecular de la corteza cerebelosa, en particular, en las fibras
paralelas. Esta colocalización sería fundamental en la actividad
fisiológica de las sinapsis de los axones de las células granulares con
las células de Purkinje, viéndose seriamente alterada al faltar
alguna de estas subunidades de Kv3.

47
Tesis Doctoral Aranzazu Urresola Olabarrieta
OBJETIVOS

48
Tesis Doctoral Aranzazu Urresola Olabarrieta
OBJETIVOS
En base a la hipótesis propuesta, nos planteamos los siguientes
objetivos en esta Tesis Doctoral:
Estudiar la localización subcelular de Kv3.1 y Kv3.3 en la
capa molecular de la corteza cerebelosa de la rata, empleando
métodos inmunocitoquímicos de alta resolución para microscopía
electrónica.
Determinar semicuantitativamente la densidad de
inmunomarcado de ambas subunidades de canales de potasio en
aquellos compartimentos subcelulares en los que se localicen.

49
Tesis Doctoral Aranzazu Urresola Olabarrieta
MATERIAL Y MÉTODOS

50
Tesis Doctoral Aranzazu Urresola Olabarrieta
MATERIAL Y MÉTODOS
Los procedimientos empleados en esta Tesis Doctoral se han
ajustado estrictamente al cumplimiento de la normativa vigente en
materia de investigación para la protección de los animales
utilizados en experimentación y otros fines científicos (Real Decreto
1201/2005; BOE 21-10-2005), y aprobados por el Comité de Ética de
Bienestar Animal de la UPV/EHU (CEBA/94/2010/GRANDES
MORENO).
Introducción a las técnicas inmunocitoquímicas
En este trabajo utilizamos técnicas inmunocitoquímicas en
combinación con anticuerpos específicos frente a secuencias
peptídicas del fragmento intracelular carboxilo de las subunidades
de los canales de potasio Kv3.1b y Kv3.3. Las técnicas
inmunocitoquímicas se basan en la capacidad específica de los
anticuerpos para unirse al antígeno y en el uso de marcadores de los
anticuerpos. Concretamente, de cara a determinar inicialmente la
localización celular de las subunidades a nivel de microscopía de luz,
utilizamos el método de la avidina-biotina-peroxidasa (Hsu y cols.,
1981) junto con anticuerpos específicos frente a Kv3.1b y Kv3.3.
Este método de la avidina-biotina-peroxidasa (Figura 10) se
basa en la gran afinidad que presentan las moléculas de avidina y

51
Tesis Doctoral Aranzazu Urresola Olabarrieta
biotina entre sí. La avidina es una glicoproteína de elevado peso
molecular (68 kilodal-
tons) presente en
grandes cantidades en la
clara de huevo. Está
formada por cuatro
subunidades compuestas
cada una de ellas por
una cadena sencilla de
128 aminoácidos, que
configuran una estructura terciaria con cuatro regiones hidrófobas
de unión a la biotina. Además, la avidina se puede utilizar como
puente entre diferentes moléculas biotiniladas, como son, en este
caso, el anticuerpo y la peroxidasa.
Por su parte, la biotina es una vitamina de bajo peso molecular
(244 daltons) perteneciente al complejo B (vitamina H) que se
encuentra presente en la yema de huevo, además de en numerosos
tejidos animales y en vegetales. La biotinilación es un proceso
bioquímico que permite conjugar la biotina a diferentes moléculas,
tales como enzimas, lectinas, anticuerpos o ácidos nucleicos, entre
otros. Durante este proceso, la biotina es transformada previamente
Figura 10. Esquema de los pasos de la técnica
inmunocitoquímica de la avidina-biotina-
peroxidasa para microscopía de luz. DAB: 3-3´-
diaminobencidina.

52
Tesis Doctoral Aranzazu Urresola Olabarrieta
en una molécula activa a través de una esterificación en la cual su
grupo carboxílico queda unido de forma covalente a residuos NH2
proteicos. Esta posibilidad de obtener múltiples asociaciones permite
una gran variedad de reacciones citoquímicas de localización. El
pequeño tamaño de la biotina hace que el proceso de biotinilación no
modifique las propiedades inmunológicas, enzimáticas o físicas de las
moléculas marcadoras. Esto ofrece una gran variedad de
posibilidades a la hora de unir a cualquier molécula varias biotinas,
lo cual dota de una elevada sensibilidad a las técnicas que utilizan
esta vitamina. El número de moléculas de biotina que pueden unirse
a un anticuerpo se estima del orden de 150, por lo que se puede
incubar el anticuerpo primario a concentraciones muy bajas.
Con respecto a la técnica propiamente dicha, el tejido se incuba
con un anticuerpo primario frente a la proteína (subunidad de canal)
que queremos detectar. Sobre éste se aplica un segundo anticuerpo
biotinilado. El anticuerpo biotinilado se une a un complejo formado
por avidina conjugada a peroxidasa biotinilada. El cromógeno que se
emplea más habitualmente es la 3,3´-diaminobencidina (DAB), la
cual se oxida en un medio que contiene tanto la peroxidasa del
complejo avidina-biotina como el peróxido de hidrógeno, dando lugar
a un producto de reacción de color marrón (Figura 10).

53
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 11. Esquema de los pasos de la técnica
de inmuno-oro preinclusión para microscopía
electrónica. Fab: fragmento de unión de la
inmunoglobulina.
El objetivo principal de esta Tesis Doctoral es investigar la
distribución subcelular de las subunidades de los canales de potasio
Kv3.1b y Kv3.3 a nivel de
microscopía electrónica.
Este propósito se aborda
mediante un método de
inmuno-oro preinclusión
(Figura 11). La alta
precisión y resolución de
esta técnica se fundamenta
en la conjugación del
segundo anticuerpo a una partícula de oro, la cual permite, al ser
examinada en un microscopio electrónico, determinar el lugar de
localización de la proteína. Tiene la ventaja de que al ser un método
preinclusión, la reacción antígeno-anticuerpo no presenta problemas
con el osmio ni con la polimerización a 60ºC de las resinas. Otro
aspecto a favor es la utilización de partículas de oro de 1,4
nanómetros conjugadas a fracciones Fab’ de los anticuerpos
secundarios. Estas características permiten una mayor penetración
del anticuerpo en el tejido, aumentando así la sensibilidad del
método. Después de la unión antígeno-anticuerpo, las pequeñas
partículas de oro se intensifican con plata para aumentar su tamaño
y hacerlas visibles en el microscopio electrónico (Figura 11).

54
Tesis Doctoral Aranzazu Urresola Olabarrieta
Anticuerpos policlonales primarios empleados en
el estudio
• Anti-Kv3.1b (clon Kcnc1), Alomone Labs. Jerusalén, Israel.
La secuencia peptídica que reconoce este antisuero es
CKESPVIAKYMPTEAVRVT, que corresponde a los aminoácidos
567-585 del extremo carboxilo terminal intracelular de la isoforma
Kv3.1b de la rata. Ha sido producido en conejo y presenta reactividad
en rata, ratón, mono y perro. Peso molecular de la proteína que
reconoce: 97 kDa.
• Anti Kv3.3 (clon KNC3), Alomone Labs. Jerusalén, Israel.
Este anticuerpo policlonal ha sido sintetizado frente a la
secuencia peptídica KSPITPGSRGRYSRDRAC, que corresponde a
los aminoácidos 701-718 del extremo carboxilo terminal intracelular
de la subunidad Kv3.3 de la rata. Ha sido producido en conejo y
presenta reactividad en rata y ratón. Peso molecular de la proteína
que reconoce: 120 kDa.

55
Tesis Doctoral Aranzazu Urresola Olabarrieta
Técnica inmunocitoquímica para microscopía de luz
Técnica de la avidina-biotina peroxidasa
En este trabajo hemos utilizado ratas albinas (Sprague-Dawley,
200-250gr, n=12) sedadas con hidrocloruro de ketamina (25 mg/kg,
inyección intramuscular) y anestesiadas con pentobarbital sódico
(60mg/kg, inyección intraperitoneal), tras lo cual fueron prefundidas
a través del ventrículo izquierdo con tampón fosfato salino (TFS, pH
7,4, temperatura ambiente) durante 20 segundos, seguido por 1 litro
de una solución fijadora de 4% de formaldehído (preparado a partir
de paraformaldehído) y 0,2% de una solución saturada de ácido
pícrico en tampón fosfato (TF, 0,1M, pH 7,4, temperatura ambiente)
durante 15 minutos. A continuación, procedimos:
1. Extracción de los cerebros.
2. Crioprotección: sacarosa 15% peso/volumen en TF (0,1M) y a
continuación sacarosa 30% en TF.
3. Corte de secciones de tejido de 30 m de grosor, recogidas en pocillos
con TFS a temperatura ambiente.
4. Preincubación con suero normal de cabra (SC) al 1,5% en TFS
durante 1 hora.
5. Incubación con antisuero primario: Kv3.1b: 0,60 g/ml 1,5%
(SC)/TFS; Kv3.3: 2 g/ml 1,5% (SC)/TFS. Tiempo de incubación del
tejido con los antisueros primarios: 48 horas.
6. Lavado de secciones flotantes: en TFS durante 1 hora.
7. Incubación con el anticuerpo secundario biotinilado (IgG anti-conejo
sintetizado en cabra. Vector Laboratories Inc. Burlingame,
Califonia, EE.UU.): 1:200 en TFS durante 1 hora.
8. Lavado de las secciones flotantes: en TFS durante 1 hora.
9. Incubación con el complejo avidina-biotina-peroxidasa (ABC; Elite,
Vector Laboratories Inc. Burlingame, Califonia, EE.UU.):
previamente mezclado (1:50), durante 1 hora.
10. Lavado de las secciones en TFS.
11. Preincubación con 3,3'-diaminobencidina (DAB; Sigma Chemical
Co., St. Louis, MO, USA) al 0,05% en TF durante 5 minutos.

56
Tesis Doctoral Aranzazu Urresola Olabarrieta
12. Incubación con DAB al 0,05% en TF más peróxido de hidrógeno al
0,01% durante 5 minutos (o el tiempo óptimo de reacción
determinado por el investigador).
13. Lavado de secciones en TF durante 30 minutos.
14. Montado de secciones sobre portaobjetos.
15. Secado.
16. Deshidratación en alcoholes de graduación creciente. 17. Las muestras se cubren usando el medio de montaje DPX
(Fluka,Chemie AG, Buchs, Suiza).
Técnica inmunocitoquímica para microscopía
electrónica
Método preinclusión de inmuno-oro intensificado con plata
Ratas albinas (Sprague-Dawley, 200-250gr, n=18) previamente
sedadas y anestesiadas como ya hemos mencionado, fueron
perfundidas transcardiacamente con una mezcla fijadora fría de
formaldehído al 4%, ácido pícrico al 0,2% y glutaraldehído al 0,025%
en tampón fosfato (0,1M, pH 7,4). El protocolo aplicado fue el
siguiente:
1. Extracción de los cerebros.
2. Lavado de los bloques de tejido en TF durante 3 horas.
3. Crioprotección: inmersión en una solución de sacarosa al 15%,
seguida por otra al 30% en TFS.
4. Paso rápido por nitrógeno líquido (varios segundos).
5. Corte en vibrotomo de secciones de 70 m de grosor.
6. Preincubación de secciones flotantes en suero normal de cabra al 20%
en TFS durante 1 hora a temperatura ambiente.
7. Incubación con antisueros primarios frente a Kv3.1b (0,60 g/ml
1,5%(SC)/TFS), o Kv3.3 (2 g/ml 1,5%(SC)/TFS), a 4ºC durante 2 o 3
días.
8. Lavados durante 1 hora en TFS.
9. Incubación con la fracción Fab' del anticuerpo secundario unido a una
partícula de oro de 1,4 nanómetros durante 2 horas, en agitación y a
4ºC (Nanoprobes Inc. Yaphank, Nueva York, EE.UU.), diluído 1:100
en suero normal de cabra al 1,5% en TFS.
10. Lavado de las secciones en TFS.

57
Tesis Doctoral Aranzazu Urresola Olabarrieta
11. Postfijación con glutaraldehído al 1% durante 10 minutos en
agitación.
12. Lavado de las secciones en agua bidestilada.
13. Intensificación de las partículas de oro con plata, utilizando el
intensificador HQ Silver Kit (Nanoprobes Inc. Yaphank, Nueva York,
EE.UU.), durante 12 minutos.
14. Osmificación del tejido con tetraóxido de osmio al 2% durante 40
minutos.
15. Deshidratación en alcoholes de graduación creciente (50º, 70º, 96º y
100º).
16. Lavado con óxido de propileno.
17. Inclusión en resina Epon 812.
18. Polimerización de la resina (2 días en estufa a 60º C).
19. Corte de secciones de 50nm de grosor en ultramicrotomo.
20. Contraste con acetato de uranilo al 2% durante 30 minutos.
21. Contraste con citrato de plomo al 2,5% durante 30 minutos.
22. Observación en el microscopio electrónico.
Controles
Control de sustitución
Incubación del tejido con suero de cabra no inmunizada en lugar
del anticuerpo primario. El resto de los pasos inmunocitoquímicos
fueron similares y se realizaron en paralelo al tejido incubado con
antisuero primario. De esta manera, puede detectarse un posible
marcado por reacción cruzada del anticuerpo secundario y problemas
de precipitación inespecífica del cromógeno.
Control de preabsorción
Preincubación del antisuero primario con el péptido frente al
que ha sido sintetizado. Después el tejido es incubado con el

58
Tesis Doctoral Aranzazu Urresola Olabarrieta
antisuero preabsorbido. En principio, este control determina si el
anticuerpo primario detecta otros antígenos además del deseado.
Control de reacción cruzada
Con el fin de descartar la posibilidad de reconocimiento de
ambas proteínas por cada uno de los dos antisueros frente a KV3.1b
y Kv3.3, cada uno de los anticuerpos fue preabsorbido con la
secuencia peptídica reconocida por el otro. La incubación del tejido
fue con las mismas concentraciones de los antisueros empleadas en
la inmunodetección tisular.

59
Tesis Doctoral Aranzazu Urresola Olabarrieta
RESULTADOS

60
Tesis Doctoral Aranzazu Urresola Olabarrieta
RESULTADOS
Patrón de distr ibución de Kv3.1b y Kv3.3 en la corteza cerebelosa de la rata en el microscopio de luz
La inmunorreactividad frente a las dos subunidades es muy
evidente en la corteza cerebelosa (Figuras 12, 13). En particular, la
capa molecular presenta una intensidad de marcado muy superior al
resto de capas que conforman la corteza (Figuras 12, 13).
Asimismo en la capa granular, la inmunorreación en las
membranas de los cuerpos celulares de las neuronas granulares es
mucho más intensa para Kv3.1b que en el caso de los somas
granulares teñidos con el anticuerpo frente a Kv3.3 (Figuras 12, 13).
Por el contrario, los cuerpos celulares de las neuronas de Purkinje
sólo son inmunopositivos para Kv3.3 (Figura 13).
En general, el patrón de inmunorreactividad observado tanto
para Kv3.1b como para Kv3.3 concuerda con lo descrito por otros
autores (Ozaita y cols, 2002; Chang y cols, 2007) y nosotros mismos
en el ratón (Alonso-Espinaco y cols., 2008).

61
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 12. Localización inmunocitoquímica de Kv3.1b en el cerebelo de la
rata. Método de la avidina-biotina-peroxidasa a nivel de microscopía de
luz. A: en la imagen de un corte parasagital del cerebelo, la inmunorreactividad
frente a Kv3.1b (la isoforma más abundante en el cerebro adulto) se observa
concentrada en la superficie de la corteza cerebelosa. Nótese la uniformidad de
tinción en todos los lóbulos cerebelosos. B: lo más destacado es la intensa
inmunorreactividad en la capa molecular (CM) y la ausencia de marcado en los
somas de las células de Purkinje dispuestos en su capa (CP, asteriscos en B´). Este
patrón sugiere la presencia de Kv3.1b en las fibras paralelas (puntas de flecha en
B´), y no en las arborizaciones dendríticas de las células de Purkinje en CM.
Obsérvese en la capa granular (CG), procesos fibrosos inmunopositivos para Kv3.1b
(flechas en B´) de trayecto ascendente hacia CP que atraviesan la capa células de
Purkinje para perderse en CM, evocando la localización de Kv3.1b en los axones de
las células granulares en su camino hacia CM, donde terminan formando las fibras
paralelas. Barras de escala: A: 200μm. B: 20μm. B´: 10μm.

62
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 13. Localización inmunocitoquímica de Kv3.3 en la cerebelo de la
rata. Método de la avidina-biotina-peroxidasa a nivel de microscopía de
luz. A: en un corte parasagital completo del cerebelo, la inmunorreactividad frente
a Kv3.3 está concentrada en la superficie de la corteza cerebelosa. Nótese la
uniformidad de tinción en todos los lóbulos cerebelosos. El patrón de distribución de
Kv3.3 a este nivel de magnificación es comparable al presentado por Kv3.1b. Sin
embargo, obsérvese en B una intensa inmunorreacción en los somas de las células
de Purkinje dispuestos en su capa (CP), en particular en la membrana
citoplasmática (flechas en B´). El fuerte marcado en la capa molecular (CM en B)
indica la presencia de Kv3.3 en el árbol dendrítico de las células de Purkinje, así
como en las fibras paralelas (puntas de flecha en B´). La inmunorreactividad aun
siendo visible en la capa granular (CG en B), es más débil que en las capas más
superficiales descritas. Barras de escala: A: 200μm. B: 20μm. B´: 10μm.

63
Tesis Doctoral Aranzazu Urresola Olabarrieta
Compart imentos neuronales que cont ienen Kv3.1b y Kv3.3 en la capa molecu lar de la cor teza cerebelosa de la ra ta en el microscopio e lect rón ico
Con el fin de identificar los compartimentos subcelulares en los
que están localizados las subunidades de los canales de potasio Kv3.1b
y Kv3.3, empleamos una técnica de inmuno-oro preinclusión para
microscopía electrónica (Figuras 14, 15, 16).
Al estudiar la localización de Kv3.1b, observamos que las células
granulares de la corteza cerebelosa, tienen una gran densidad de
inmunopartículas. Esta subunidad se encuentra rodeando el
citoplasma del soma y dendritas de dichas células en la capa granular,
así como en los botones terminales de las fibras paralelas que
establecen sinapsis con las espinas dendríticas de las células de
Purkinje en la capa molecular (Figura 14). Típicamente, el marcado en
estos botones no se encuentra cerca de la especialización de las
membranas presinápticas, sino en porciones de membrana alejadas de
ella.

64
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 14. Localización subcelular de la subunidad del canal
de potasio Kv3.1b en la corteza cerebelosa de la rata adulta.
Método de inmuno-oro preinclusión para microscopía electrónica. En
la capa granular (a), la membrana somática y dendrítica de las
células granulares (CG) están densamente decoradas con
inmunopartículas que indican la rica distribución de la subunidad en
estos compartimentos. En b, la acumulación de partículas (flechas) es
en la membrana de un terminal de fibra paralela (ter FP), respetando
la típica especialización sináptica con la espina dendrítica (es) de una
célula de Purkinje. Un patrón de distribución similar es observado en
c, donde varias terminales típicas de fibras paralelas (ter FP)
contienen inmunopartículas (flechas) asociadas a sus membranas.
Nótese que las espinas dendríticas (es) de las células de Purkinje en
contacto con botones sinápticos de fibras paralelas, son Kv3.1b
inmunonegativas. Barras de escala: a: 2 m; b,c: 0,4 m.
a
CG
CG
CG
b
es
ter FP
c
ter FP
ter FP
ter
FP
ter FP
es
es
e
s

65
Tesis Doctoral Aranzazu Urresola Olabarrieta
A continuación, investigamos más en detalle la distribución
inmunocitoquímica de Kv3.1b y Kv3.3 en la capa molecular de la
corteza cerebelosa (Figuras 15, 16). Numerosas partículas de oro
intensificadas con plata se localizan en membranas de axones,
terminales sinápticas y espinas dendríticas. Pudimos observar un
fuerte marcado para Kv3.1b en varicosidades presinápticas de fibras
paralelas, caracterizadas por contener vesículas sinápticas
redondeadas y claras, y por hacer sinapsis asimétricas con espinas
dendríticas de células de Purkinje (Palay y Chan-Palay 1974; Grandes
y cols., 1994) (Figura 15). Por otra parte, el inmunomarcado para
Kv3.1b es particularmente denso en membranas de perfiles axónicos
seccionados de través (Figura 15) y a lo largo de membranas de axones
de pequeño tamaño cortados longitudinalmente, que corresponden a
segmentos intervaricosos de fibras paralelas (Figura 15). Es de
destacar que el marcado de membrana en todos los casos queda
confinado más allá de las especializaciones presinápticas, sin llegarse
a identificar en esta especialización de la sinapsis. Finalmente, no
pudimos demostrar a este nivel de resolución con la técnica empleada,
la localización de Kv3.1b en espinas ni en dendritas de las células de
Purkinje.

66
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 15. Compartimentación subcelular de Kv3.1b en la
capa molecular del cerebelo de la rata. Método de inmuno-oro
preinclusión para microscopía electrónica en secciones cerebelosas
parasagitales. Numerosas partículas de oro intensificadas con plata
(flechas) se observan en membranas de terminales sinápticas de
fibras paralelas (ter FP en a-d) que hacen contactos sinápticos
asimétricos con espinas dendríticas inmunonegativas de células de
Purkinje (es). Apréciese en c, una espina dendrítica completa (es;
sentido de flecha entrecortada) emergiendo de una dendrita (den) de
célula de Purkinje, que recibe una terminal paralela (ter FP)
inmunopositiva (flechas). Los segmentos intervaricosos finos de las
fibras paralelas (ax en a y c-d) también aparecen marcados para
Kv3.1b. En d, Kv3.1b se distribuye en la membrana de una terminal
de fibra paralela (ter FP) que está junto a un perfil axónico cortado
longitudinalmente (ax) que también contiene la subunidad.
Obsérvese que Kv3.1b está en las membranas perisinápticas y
extrasinápticas más allá de las zonas activas. Las especializaciones
sinápticas entre las terminales de las fibras paralelas (ter FP) y las
espinas dendríticas de las células de Purkinje (es) carecen por lo
general de inmunopartículas (puntas de flecha en a-d). Barras de
escala: 0,4µm.

67
Tesis Doctoral Aranzazu Urresola Olabarrieta
La subunidad Kv3.3 también está presente en las membranas de
las fibras paralelas (Figura 16), mientras que los contactos sinápticos,
al igual que Kv3.1b, están desprovistos de esta subunidad (Figura 16).
El principal marcado de Kv3.3 se da en las espinas dendríticas de las
células de Purkinje, que reciben contactos sinápticos excitadores de las
terminales sinápticas de las fibras paralelas (Figura 16). Como ocurre
con la distribución de ambas subunidades de Kv3 en otros perfiles
neuronales, las densidades postsinápticas de las espinas dendríticas
inmunorreactivas carecen de la presencia de Kv3.3, estando siempre
las partículas intensificadas con plata en un anillo periférico o más
alejadas de la especialización postsináptica (Figura 16).
No observamos la localización de Kv3.1b y Kv3.3 en botones
sinápticos de fibras trepadoras, que pudimos identificar según sus
típicas características ultraestructurales de varicosidades bulbosas o
perfiles más elongados, agrupando en su interior abundantes vesículas
sinápticas redondeadas y translúcidas dispuestas en la proximidad de
las uniones sinápticas con las dendritas de las células de Purkinje
(Palay y Chan-Palay 1974; Grandes y cols., 1994).
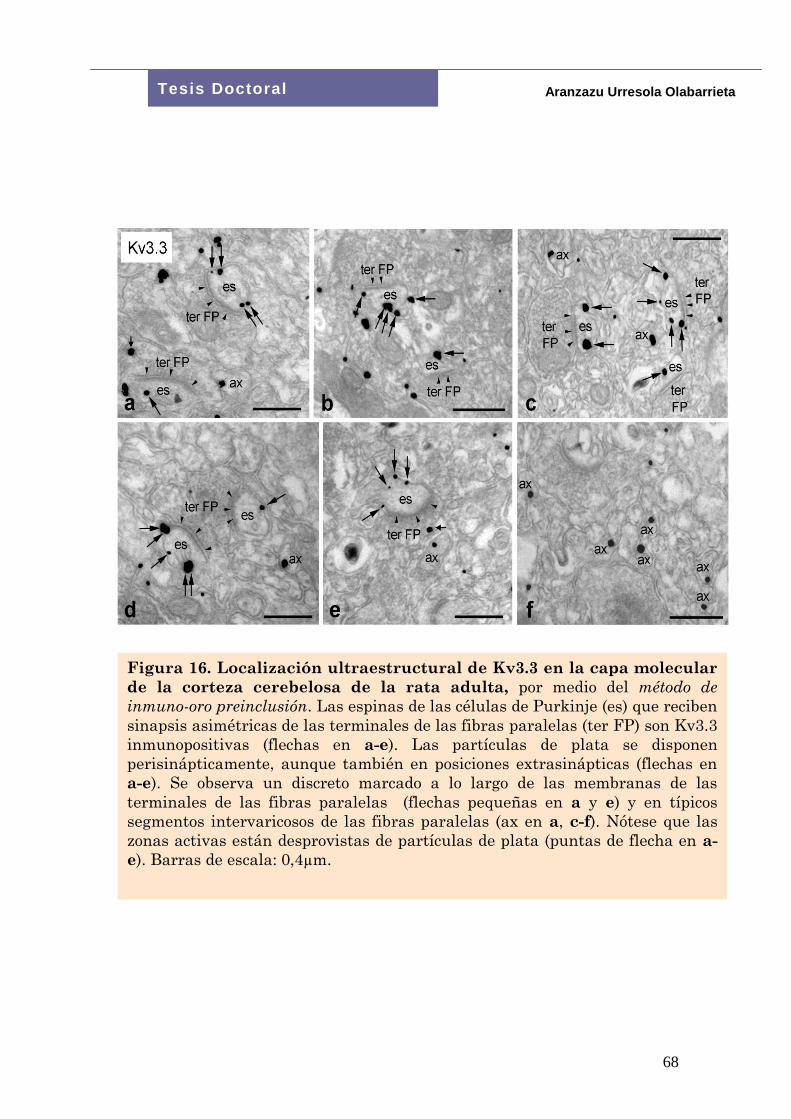
68
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 16. Localización ultraestructural de Kv3.3 en la capa molecular
de la corteza cerebelosa de la rata adulta, por medio del método de
inmuno-oro preinclusión. Las espinas de las células de Purkinje (es) que reciben
sinapsis asimétricas de las terminales de las fibras paralelas (ter FP) son Kv3.3
inmunopositivas (flechas en a-e). Las partículas de plata se disponen
perisinápticamente, aunque también en posiciones extrasinápticas (flechas en
a-e). Se observa un discreto marcado a lo largo de las membranas de las
terminales de las fibras paralelas (flechas pequeñas en a y e) y en típicos
segmentos intervaricosos de las fibras paralelas (ax en a, c-f). Nótese que las
zonas activas están desprovistas de partículas de plata (puntas de flecha en a-
e). Barras de escala: 0,4µm.

69
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 17. Especificidad de los antisueros frente a las
subunidades Kv3 estudiadas en la corteza cerebelosa de la
rata adulta. La preabsorción de los anticuerpos frente a Kv3.1b y
Kv3.3 con el péptido correspondiente, causa la desaparición completa
del inmunomarcado. Barras de escala: 0,4µm.
Contro les
Finalmente, la especificidad del marcado inmunocitoquímico fue
comprobada realizando la preincubación de los antisueros para Kv3.1b
y Kv3.3 con el péptido correspondiente frente al que el antisuero ha
sido sintetizado y, por lo tanto, es reconocido en el tejido. En estos
experimentos de preabsorción, el inmunomarcado desapareció en su
totalidad, indicando que las herramientas utilizadas en la
inmunodetección de Kv3.1b y Kv3.3 eran altamente específicas
(Figura 17). Por otra parte, tampoco fue posible detectar
inmunotinción en secciones de tejido cerebeloso procesadas
inmunocitoquímicamente sin antisuero primario, o en los
experimentos en los que el anticuerpo policlonal primario fue
sustituido por suero de cabra (no mostrado).

70
Tesis Doctoral Aranzazu Urresola Olabarrieta
Figura 18. Frecuencia relativa de las estructuras marcadas
para Kv3.1b y Kv3.3 en la capa molecular de la corteza
cerebelosa de la rata adulta. Las barras representan los
porcentajes de dominios neuronales de Kv3.1b y Kv3.3
inmunopositivos, sobre el número total de perfiles sinápticos
analizados en la capa molecular (n=325 para Kv3.1b y n=355 para
Kv3.3). Kv3.1b se localiza en membranas de terminales sinápticas
de fibras paralelas (ter FP; 85,16±3,155%) y en porciones
intervaricosas de estas fibras (FP; 47,50±4,578%). Kv3.3 se
distribuye en espinas dendríticas de las células de Purkinje (es CP;
53,98 ± 4,710%) y también en ter FP y en porciones intervaricosas
(FP) de las fibras paralelas (22,76 ± 3,796% y 27,73 ± 4,121%,
respectivamente). Los números dentro y fuera de los paréntesis en
cada columna indican el tamaño de muestra y la media de los
porcentajes. Barras de error indican la media ± SEM.
Anál is is estadís t ico
El análisis estadístico revela que Kv3.1b se localiza en
aproximadamente el 85% de las terminales sinápticas de las fibras
paralelas, y en el 47% de sus porciones intervaricosas (Figura 18). La
localización de Kv3.3, sólo alcanza el 28% de las intervaricosidades y
el 23% de los botones presinápticos de las fibras paralelas (Figura
18). Es de destacar que el 54% de las espinas dendríticas de las
células de Purkinje son Kv3.3 inmunopositivas.

71
Tesis Doctoral Aranzazu Urresola Olabarrieta
Por otra parte, la densidad de inmunomarcado para Kv3.1b (partículas
de oro intensificadas con plata por longitud de membrana) fue mucho
más alta en segmentos intervaricosos de las fibras paralelas, seguido
por las terminales sinápticas de estas fibras (***; p < 0,001; Tabla 1).
La densidad de Kv3.3 en estos compartimentos fue también
significativamente mayor que la densidad en las terminales sinápticas
de las fibras paralelas (***; p < 0,001; Tabla 1). Sin embargo, las
SUBUNIDAD CANAL
PERFIL PARTÍCULAS/
m
ÁREA ANALIZADA
( m2)
Nº TOTAL
DE PERFILES
MARCADOS
COMPARACIÓN
ESTADÍSTICA
Kv3.1b A. ter FP 1,23 ± 0,75 39,15 105
A versus B: ***; p < 0,001 B. FP 2,16 ± 0,16 8,90 57
A
Kv3.3
C. ter FP 0,99 ± 0,11 23,36 28 C versus D: ***; p < 0,001
C versus E: ***; p < 0,001
D versus E: ns; p > 0,05
D. FP 2,07 ± 0,26 5,52 33
E. es CP 1,73 ± 0,13 9,82 63
diferencias de densidad de Kv3.3 en las espinas dendríticas de las
células de Purkinje respecto a las intervaricosidades de las fibras
paralelas fueron estadísticamente no significativas (ns; p = 0,1947;
Tabla 1).
Tabla 1. Densidad de inmunopartículas en membranas de perfiles
neuronales en la capa molecular de la corteza cerebelosa de la rata
adulta. Las secciones parasagitales de cerebelo fueron procesadas con
anticuerpos específicos frente a Kv3.1b y Kv3.3 en combinación con un método
de inmuno-oro preinclusión. La densidad es el número de partículas de plata
de Kv3.1b y Kv3.3 por micra de membrana plasmática de terminales de fibras
paralelas (ter FP), porciones intervaricosas de fibas paralelas (FP) y espinas
dendríticas de células de Purkinje (es CP). La acumulación de Kv3.1b y Kv3.3
es aproximadamente 2 veces mayor en “FP” que en “ter FP” (***diferencias
estadísticamente significativas a p < 0,001). La densidad de Kv3.3 en “es CP”
es significamente más alta que en en “ter FP” (***diferencias
estadísticamente significativas a p < 0,001). Sin embargo, las diferencias de
densidad de Kv3.3 en “es CP” (1,73 ± 0,13) respecto a “FP” (2,07 ± 0,26) no son
estadísticamente significativas (p = 0,1947; ns: p > 0,05). El área total de los
diferentes perfiles y el número de perfiles inmunopositivos analizados
aparecen en las dos últimas columnas.

72
Tesis Doctoral Aranzazu Urresola Olabarrieta
DISCUSIÓN

73
Tesis Doctoral Aranzazu Urresola Olabarrieta
DISCUSIÓN
En este trabajo de Tesis Doctoral, hemos estudiado la
distribución de las subunidades de los canales de potasio Kv3, Kv3.1b
y Kv3.3, en compartimentos subcelulares específicos de la capa
molecular de la corteza cerebelosa de la rata adulta.
En los últimos años, el conocimiento de la localización y función
de Kv3.1 y Kv3.3 en la corteza cerebelosa ha aumentado
significativamente (Perney y cols., 1992; Goldman-Wohl y cols., 1994;
Weiser y cols., 1994, 1995; Sabatini y Regehr, 1997; Sekirnjak y cols.,
1997; Ozaita y cols., 2002; Zucker y Regehr, 2002; Martina y cols.,
2003; McKay y Turner, 2004; McMahon y cols., 2004; Akemann y
Knöpfel, 2006; Joho y cols., 2006; Chang y cols., 2007; Zagha y cols.,
2008; Hurlock y cols., 2008, 2009). Sin embargo, estos trabajos previos
estudiaron la localización de Kv3.1 y Kv3.3 con métodos de
inmunoperoxidasa, que tienen como consecuencia la identificación de
los lugares de distribución proteica por medio de un producto de
inmunorreacción difuso (Ozaita y cols., 2002; Chang y cols., 2007).
Los resultados de la presente Tesis Doctoral demuestran por
primera vez con un método de alta resolución de inmuno-oro
preinclusión, que existe un porcentaje más elevado de fibras paralelas

74
Tesis Doctoral Aranzazu Urresola Olabarrieta
marcadas para Kv3.1b que para Kv3.3, y que además la intensidad de
marcado para cada una de estas subunidades es mayor en ciertos
subcompartimentos neuronales analizados que en otros.
Debido a las propiedades biofísicas que presentan, los canales
Kv3 facilitan el disparo de potenciales de acción a gran frecuencia, y
las interacciones que presentan estos canales con corrientes de sodio,
sustentan el disparo de potenciales de acción espontáneos en la
ausencia de impulsos sinápticos (Perney y cols., 1992; Weiser y cols.,
1995; Massengill y cols., 1997; Martina y cols., 1998; Erisir y cols.,
1999; Hernández-Pineda y cols., 1999; Raman y Bean, 1999; Rudy y
McBain, 2001; McKay y Turner, 2004; Song y cols., 2005; Akeman y
Knöpfel, 2006). En un estudio electrofisiológico relativamente
reciente, se observó que las neuronas de las núcleos profundos del
cerebelo que carecen del enzima de síntesis del neurotransmisor
GABA, decarboxilasa del ácido glutámico (GAD-), disparan a tasas
más altas y exhiben potenciales de acción más rápidos que las
neuronas GAD+ (Uusisaari y cols., 2007). Estas observaciones se
correlacionan con la diferente distribución de Kv3.1b y Kv3.3 en
neuronas GAD- y GAD+ del núcleo dentado (Alonso-Espinaco y cols.,
2008). Por lo tanto, en conjunto, la localización de Kv3.1b y Kv3.3 en
específicos compartimentos subcelulares puede suponer el sustrato

75
Tesis Doctoral Aranzazu Urresola Olabarrieta
anatómico de las propiedades biofísicas diferenciales que caracterizan
a las neuronas GAD- y GAD+ de los núcleos profundos del cerebelo.
Nuestros resultados apuntan hacia la existencia de un mayor
porcentaje de fibras paralelas marcadas para Kv3.1b que para Kv3.3,
aun dándose un cierto solapamiento de localización de ambas
subunidades en estas fibras, como era de esperar a partir de la
expresión de los dos ARN mensajeros y correspondientes proteínas en
las células granulares. Ello está en consonancia con las observaciones
publicadas de que los potenciales de acción de las fibras paralelas son
más anchos en ratones sin Kv3.1 ni Kv3.3, y también en ratones
mutantes que carecen de ambos alelos de Kv3.1 y uno de Kv3.3
(Matsukawa y col., 2003). Estas subunidades probablemente tengan
una sinergia funcional, lo que permitiría en caso de lesión o
inactividad de una de ellas, que la otra realizase en solitario el mismo
trabajo. Curiosamente se ha observado que, aunque los dobles
mutantes presenten una severa disfunción motora, no se observa
diferencia anatómica alguna y, además, estos ratones muestran un
aumento locomotor y de la actividad exploradora. Tampoco sufren
alteración del aprendizaje ni de la memoria (Espinosa y cols., 2001).
Por tanto, no parece que participen directamente estas subunidades
de Kv3, en todos los procesos de intercambio de información entre

76
Tesis Doctoral Aranzazu Urresola Olabarrieta
neuronas, por lo que la ausencia de ellas puede ser corregida por
algún otro mecanismo.
A pesar de la imposibilidad en este estudio de llevar a cabo
experimentos directos de colocalización de ambas subunidades en la
capa molecular del cerebelo, debido a limitaciones metodológicas
(ambos antisueros Kv3.1b y Kv3.3 fueron sintetizados en conejo), es
factible asumir que Kv3.1b y Kv3.3 coinciden en al menos algunos de
los dominios de las fibras paralelas (botones sinápticos,
intervaricosidades) que contienen estas subunidades. Ello contrasta
con la situación dada en las células de Purkinje, que sólo expresan
niveles elevados del ARN mensajero que codifica para Kv3.3. De tal
forma, la falta de Kv3.3 en neuronas de Purkinje resulta en una
reducción de la tasa de disparo, potenciales de acción más amplios y
una más lenta hiperpolarización tardía de las células (McMahon y
cols., 2004; Akemann y Knöpfel 2006). Es interesante destacar que la
actividad de Kv3.3 en el soma de las células de Purkinje es muy
importante en la regulación de las espigas complejas evocadas en
estas células tras la activación de las fibras trepadoras, lo que se ve
afectado en ratones sin Kv3.3 (Zagha y cols., 2008). Hay que añadir
que la falta de sólo Kv3.3 o la carencia de Kv3.3 y un alelo de Kv3.1
causan un fenotipo atáxico en ratones, que puede revertirse tras la

77
Tesis Doctoral Aranzazu Urresola Olabarrieta
reinserción de Kv3.3 en las células de Purkinje (Hurlock y cols., 2008).
Este rescate no es observado en ratones que carecen de ambos Kv3.1 y
Kv3.3 (Hurlock y cols., 2009).
ANATOMÍA DE Kv3.1b y Kv3.3 EN LA CORTEZA CEREBELOSA
Estudios de hibridación in situ han demostrado que las
subunidades de los canales de potasio dependientes de voltaje Kv3.1 y
Kv3.3 se distribuyen ampliamente en distintas áreas del cerebro, con
una expresión relativamente alta en el cerebelo, tanto en la corteza
como en los núcleos profundos (Perney y cols., 1992; Goldman-Wohl y
cols., 1994; Weiser y cols., 1994). En primer lugar, quisimos
comprobar en esta Tesis si la distribución de Kv3.1b y Kv3.3 a nivel de
microscopía de luz mostraba el mismo patrón de distribución que los
correspondientes ARNm en la corteza cerebelosa. Asimismo, la
microscopía de luz nos permitió tener una primera visión general de la
distribución de ambas subunidades. La técnica inmunocitoquímica
aplicada sobre cortes parasagitales de cerebelo de ratas adultas,
demostró que Kv3.1b se distribuye en las capas molecular y granular
de la corteza cerebelosa, siendo el marcado más intenso en la capa
molecular. Sin embargo, los niveles de ARNm para Kv3.1 son más
altos en la capa granular que en la molecular, debido a que el ARNm
se localiza principalmente en las células granulares (Weiser y cols.,

78
Tesis Doctoral Aranzazu Urresola Olabarrieta
1994). El marcado de la capa molecular no puede explicarse por la
presencia de Kv3.1b en las células en cesto y estrelladas, ya que estas
neuronas expresan niveles bajos de ARNm para Kv3.1.
Estudios de microscopía electrónica han revelado que la
abundancia de Kv3.1 en la capa molecular se debe a su presencia en
los axones de las células granulares, las fibras paralelas (Weiser y
cols., 1995; Puente y cols, 2010). Por el contrario, no observamos en la
capa molecular la localización de Kv3.1b en las células en cesto,
estrelladas, ni en las dendritas de las células de Purkinje, al igual que
en un trabajo previo (Weiser y cols., 1995).
El ARNm codificante para Kv3.3 tiene un patrón de expresión
similar al del Kv3.1, siendo especialmente abundante en el tronco del
encéfalo y la corteza cerebelosa. En esta última, el ARNm se expresa
mayoritariamente en las células de Purkinje, y no tanto en las células
granulares, a diferencia de lo que ocurría con el ARNm para Kv3.1
(Goldman-Wohl y cols., 1994; Weiser y cols., 1994). En nuestros cortes
histológicos de rata procesados con el antisuero frente a Kv3.3 para
microscopía de luz, observamos células inmunopositivas en las capas
molecular, de Purkinje y granular, aunque con mayor intensidad de
marcado en la capa molecular y de Purkinje. Este patrón coincide con

79
Tesis Doctoral Aranzazu Urresola Olabarrieta
lo hallado por medio de técnicas de inmunoperoxidasa e
inmunofluorescencia, que han demostrado la distribución de Kv3.3
sobre todo en la capa molecular, las células de Purkinje y sus axones
que cruzan la capa granular en su camino a los núcleos cerebelosos
profundos (Chang y cols., 2007; Alonso-Espinaco y cols., 2008). Por
tanto, la diferencia fundamental que existe entre Kv3.1b y Kv3.3 en la
corteza cerebelosa es la localización de Kv3.3, y no de Kv3.1b. en las
neuronas de Purkinje. En definitiva, el patrón de distribución de estas
dos subunidades de canales potasio no es idéntico en la corteza
cerebelosa.
La isoforma Kv3.1b es la más larga de las dos que presenta
Kv3.1. La forma corta Kv3.1a es menos abundante que Kv3.1b
(Perney y cols., 1992; Gan y Kaczmarek, 1998). Asimismo, Kv3.1a es
reemplazada gradualmente por Kv3.1b en el adulto (Perney y cols.,
1992; Gan y Kaczmarek, 1998), siendo Kv3.1a prácticamente
inexistente en el animal maduro. Nuestros hallazgos en las células
granulares concuerdan con los datos existentes de la presencia de
Kv3.1b en dendritas proximales, somas, axones y terminales axónicas
(Weiser y cols., 1995; Sekirnjak y cols., 1997; Ozaita y cols., 2002).
Uno de los mecanismos que gobiernan la localización de Kv3.1 en
axones y dendritas, implica el dominio T1 del terminal amino de la

80
Tesis Doctoral Aranzazu Urresola Olabarrieta
subunidad y la proteína ankirina que reside sobre todo en el segmento
inicial del axón. Esta proteína, a su vez, se une a un motivo
identificado en el terminal carboxilo de Kv3.1 (Xu y cols., 2007). Este
motivo está presente en Kv3.1a y Kv3.1b, aunque su unión es más
eficaz al terminal carboxilo de Kv3.1b, lo que permite que esta
isoforma atraviese el segmento inicial del axón y se localice a lo largo
del mismo y en su campo terminal (Xu y cols., 2007). El motivo en el
terminal carboxilo que determina la localización axónica de Kv3.1 y el
dominio T1 del terminal amino de las subunidades Kv3, están muy
conservados desde moscas a humanos, por lo que su interacción debe
representar un mecanismo filogenético a través del cual se regula la
localización de los canales Kv3 (Xu y cols., 2007).
SIGINIFICADO FUNCIONAL DE LA LOCALIZACIÓN DE Kv3.1b y
Kv3.3 EN FIBRAS PARALELAS Y DE LA PRESENCIA DE Kv3.3 EN
CÉLULAS DE PURKINJE
Como ya hemos mencionado, las corrientes Kv3 están presentes
en neuronas especializadas en disparos de alta frecuencia (Perney y
cols., 1992; Weiser y cols., 1995; Massengill y cols., 1997; Martina y
cols., 1998; Erisir y cols., 1999; Hernández-Pineda y cols., 1999; Rudy
y McBain, 2001; McKay y Turner, 2004; Song y cols., 2005; Akeman y
Knöpfel, 2006; McDonald y Mascagni, 2006), en las que la
dependencia de voltaje y cinética de rápida decaída de las corrientes

81
Tesis Doctoral Aranzazu Urresola Olabarrieta
Kv3, ambas sintonizadas de forma precisa, facilitan una
repolarización rápida de los potenciales de acción (Erisir y cols, 1999;
Raman y Bean, 1999). En la corteza cerebral, la presencia de
corrientes Kv3 distingue las interneuronas gabaérgicas de disparo
rápido de las neuronas piramidales excitadoras (Massengill y cols.,
1997; Martina y cols., 1998). En los núcleos vestibulares y en los
núcles profundos del cerebelo, las neuronas gabaérgicas y no
gabaérgicas son capaces de soportar tasas de disparo muy elevadas
(Bagnall y cols., 2007; Uusisaari y cols., 2007), indicando el papel de
Kv3 en ambos tipos celulares de estos núcleos (Gittis y du Lac, 2007).
Es sabido que estos canales dependientes de voltaje presentan un
alto umbral de activación y son rápidamente activados durante la fase
de ascenso del potencial de acción. Por lo tanto, Kv3.1 y Kv3.3 actúan
modelando la forma de la repolarización del potencial de acción (Rudy
y McBain, 2001). Los ratones que carecen de los canales Kv3.1 y Kv3.3
tienen unos défictis motores muy manifiestos, fundamentalmente
presentan ataxia severa y temblor, además de una elevada
sensibilidad al alcohol (Espinosa y cols., 2001), como ya hemos
mencionado. Sin embargo, los ratones que carecen sólo de Kv3.1 o
Kv3.3 no presentan alteraciones de la función motora (Chan, 1997; Ho
y cols., 1997; Sánchez y cols., 2000). En cerebro, muchas de las

82
Tesis Doctoral Aranzazu Urresola Olabarrieta
regiones que expresan Kv3.1 también expresan Kv3.3 (Weiser y cols.,
1994). Los ARNm que codifican para Kv3.1 y Kv3.3 se expresan a
unos niveles relativamente elevados en la corteza y en los núcleos
profundos del cerebelo, por lo que al menos algunos de los déficits
motores observados en los ratones deficientes de Kv3.1 y Kv3.3
pueden deberse a una disfunción cerebelosa. Por ejemplo, las células
granulares expresan (Weiser y cols., 1994; Grigg y cols., 2000; Li y
cols., 2001; Rudy y McBain, 2001) y localizan ambas subunidades
Kv3.1 y Kv3.3, como hemos demostrado en esta Tesis Doctoral. Es
posible que en este tipo celular la expresión de estas dos subunidades
con similares características biofísicas resulte en una redundancia
funcional, lo que explicaría la falta de cambios fenotípicos manifiestos
en ratones que carecen Kv3.1 o Kv3.3. Las dos estarían implicadas en
la fase de repolarización del potencial de acción en el soma y/o el axón
de las células granulares. La alteración de la repolarización del
potencial de acción axónico afecta la capacidad de soportar una
transmisión sináptica de alta frecuencia (Rudy y McBain, 2001), así
como la fiabilidad y plasticidad a corto plazo a nivel de las sinapsis
que hacen las fibras paralelas con las células de Purkinje (Sabatini y
Regehr, 1997; Zucker y Regehr, 2002). De hecho, en estas mismas
sinapsis en ratones deficientes de Kv3.1 y Kv3.3, la transmisión de
trenes de potenciales de acción a 200Hz (pero no a 100Hz) está

83
Tesis Doctoral Aranzazu Urresola Olabarrieta
afectada; la anchura de los potenciales de acción de las fibras
paralelas aumenta; la facilitación del pulso por pares disminuye, así
como el umbral para la inducción de una corriente excitadora
postsináptica mediada por el subtipo 1 de receptor metabotrópico de
glutamato (Matsukawa y cols., 2003). Es de destacar que estos
cambios son dependientes del número de alelos afectados (Matsukawa
y cols., 2003), como ya ha sido comentado.
Las células de Purkinje disparan espontáneamente a una
frecuencia de 40Hz in vivo y en preparaciones de secciones de tejido
(Llinás y Sugimori, 1980). Estas células expresan (Goldman-Wohl y
cols., 1994) y localizan Kv3.3 en sus espinas dendríticas, como hemos
descrito. Los canales Kv3 en las neuronas de Purkinje impiden
activamente la propagación retrógrada de las espigas somáticas de
sodio, dan forma a las respuestas mediadas por la actividad de las
fibras trepadoras y facilitan el disparo en salvas (Martina y cols.,
2003; McKay y Turner, 2004). La tasa de disparos de potenciales de
acción espontáneos está sustancialmente reducida en células de
Purkinje carentes de Kv3.3 (Akemann y Knöpfel, 2006). Además,
estas células presentan unos potenciales de acción más anchos,
alcanzan potenciales más positivos y tienen una hiperpolarización
tardía más lenta (McMahon y cols., 2004; Akemann y Knöpfel, 2006).

84
Tesis Doctoral Aranzazu Urresola Olabarrieta
Esta reducción de la actividad espontánea de las células de Purkinje
que no expresan Kv3.3, no se debe a un simple aumento de la entrada
de calcio, o al incremento de la actividad de los canales de potasio
activados por calcio. Es de destacar que la tasa normal de disparo es
rescatada al reintroducir la conductancia de Kv3 en estas neuronas,
por medio de la técnica dinámica de clampaje (Akemann y Knöpfel,
2006).
El papel de los canales Kv3 es permitir una rápida repolarización
del potencial de acción, y por tanto posibilitar los disparos de alta
frecuencia (Wang y cols., 1998a; Erisir y cols., 1999; Wigmore y Lacey,
2000; Lien y Jonas, 2003). La expresión de los canales Kv3 en
dendritas podría tener un papel único en la fisiología de las células de
Purkinje (Martina y cols., 2003; McKay y Turner, 2004), a la vez de
ser importante en la modulación de los picos sinápticos de las
dendritas y en el flujo de calcio durante los picos. Recientemente, se
ha estudiado la función de Kv3.3 en la regulación de las espigas
complejas evocadas en las células de Purkinje tras la activación de las
fibras trepadoras (Zagha y cols., 2008). Los autores de este estudio
observaron que la expresión de estas espigas está regulada por la
actividad de Kv3.3 localizado a nivel somático y no dendrítico (Zagha y
cols., 2008). Muy importante es el hecho de que las propiedades

85
Tesis Doctoral Aranzazu Urresola Olabarrieta
eléctricas de Kv3.3 y los canales de sodio actúan coordinadamente
para limitar la acumulación de canales de sodio inactivados y, de esta
forma, permitir que tengan lugar disparos repetitivos rápidos. Es de
destacar que las células de Purkinje de los ratones carentes de Kv3.3
producen espigas complejas alteradas, lo que es probable que sea el
sustrato celular de los fenotipos de síndrome cerebeloso observado en
estos ratones (Zagha y cols., 2008).
Las alteraciones de los potenciales de acción de las neuronas de
Purkinje deficientes en Kv3.3, producen una alteración tónica y
rítmica de la transmisión sináptica gabaérgica sobre las neuronas de
los núcleos profundos del cerebelo (McMahon y cols., 2004). Hay que
subrayar que los datos obtenidos en otros experimentos indican que la
presencia de Kv3.3 es necesaria para que se manifieste el temblor
inducido por la inyección intraperitoneal de harmalina y que afecta de
forma selectiva al circuito olivocerebeloso (McMahon y cols., 2004). El
papel fundamental de las células de Purkinje en este temblor fue
previamente sugerido por la observación de que la harmalina en los
ratones pcd/pcd (carentes de células de Purkinje por degeneración
progresiva), causa un temblor más leve en términos de amplitud y
frecuencia (Milner y cols., 1995).

86
Tesis Doctoral Aranzazu Urresola Olabarrieta
Este circuito olivocerebeloso, que comprende la oliva inferior, las
células de Purkinje y los núcleos profundos del cerebelo, tiene un
papel principal en la generación de una actividad cerebelosa
coordinada y en el control de los movimientos intencionados (Welsh y
cols., 1995; De Zeeuw y cols., 1998; Fukuda y cols., 2001). Las
neuronas de la oliva inferior, que son el origen de las fibras
trepadoras, no expresan los ARNm para Kv3.1 ni Kv3.3 (Weiser y
cols., 1994). La vía constituida por las fibras musgosas-célula
granulares-células de Purkinje converge con el circuito olivocerebeloso
a nivel de las células de Purkinje. En un trabajo previo (Matsukawa y
cols., 2003), los autores observaron que la transmisión sináptica de las
fibras paralelas de las células granulares sobre las células de Purkinje
está afectada tanto por la ausencia de Kv3.1 como de Kv3.3. Por lo
tanto, es improbable que la falta de Kv3.3 en las células granulares
explique la ausencia de temblor inducido por harmalina en los ratones
deficientes de Kv3.3. Sin embargo, las alteraciones en estas sinapsis
podrían explicar los efectos de la pérdida de Kv3.1 sobre la amplitud y
frecuencia del temblor inducido por harmalina (McMahon y cols.,
2004).

87
Tesis Doctoral Aranzazu Urresola Olabarrieta
RESUMEN GENERAL
Las células nerviosas han evolucionado con el fin de llevar a cabo
funciones específicas en circuitos complejos, requiriendo una
especialización de su capacidad de procesar la información y
transformarla en un patrón de disparo de potenciales de acción. Esta
capacidad de las neuronas de procesar y transmitir información
depende de sus corrientes iónicas intrínsecas. Diversos estudios han
demostrado que el nivel de expresión de canales iónicos en una misma
célula varía considerablemente de animal a animal, pero el patrón de
salida de la información se mantiene constante a través de la
correlación de expresión de canales, que mantiene un balance de
corrientes específico para un tipo particular de neurona (Prinz y cols.,
2004; Schulz y cols., 2006). La distribución de canales iónicos en las
diferentes partes de la célula tiene un importante significado
funcional (Almers y Stirling, 1984; Llinás, 1988). La integración
neuronal depende de la respuesta local a los diversos estímulos que le
llegan a la célula y la comunicación de estos centros con el axón. Por
tanto, la implicación de un determinado grupo de canales iónicos va a
depender de su precisa localización en la membrana celular. Las
subunidades de los canales de potasio dependientes de voltaje Kv3.1 y
Kv3.3 confieren propiedades de disparo rápido a aquellas neuronas
que los expresan, y se distribuyen en muchas áreas cerebrales,

88
Tesis Doctoral Aranzazu Urresola Olabarrieta
incluyendo las implicadas en el control y la modulación de la actividad
motora y en la precisa ejecución de las habilidades motoras
adquiridas, como son el cerebelo, los ganglios basales y la médula
espinal (Drewe y cols., 1992; Perney y cols., 1992; Rettig y cols., 1992;
Rudy y cols., 1992; Goldman-Wohl y cols., 1994; Lenz y cols., 1994;
Weiser y cols., 1994, 1995; Sekirnjak y cols., 1997; Baranauskas y
cols., 1999; Hernández-Pineda y cols., 1999; Grigg y cols., 2000;
Deuchars y cols., 2001; Li y cols., 2001; Devaux y cols., 2003;
Matsukawa y cols., 2003; Brooke y cols., 2004). Por tanto, conocer la
precisa localización subcelular de Kv3.1 y Kv3.3 es muy importante de
cara a entender los mecanismos neurales en los que están implicados.
El hecho de que Kv3.1b y Kv3.3 se localicen en el cerebelo, tanto
en la corteza cerebelosa como en los núcleos profundos (Alonso-
Espinaco y cols., 2008; Puente y cols., 2010), como hemos descrito en
este estudio, plantea la pregunta de si las deficiencias motoras
observadas en los ratones que carecen de Kv3.1 y Kv3.3 pudieran ser
de origen cerebeloso. Se piensa que el circuito olivocerebeloso tiene un
importante papel en la generación de la actividad cerebelosa rítmica
que controla los movimientos voluntarios (Welsh y cols., 1995; De
Zeeuw y cols., 1998; Fukuda y cols., 2001). Una disfunción de este
sistema podría llevar a contracciones musculares anómalas,

89
Tesis Doctoral Aranzazu Urresola Olabarrieta
manifestándose con movimientos incoordinados, temblores y
mioclonias. El circuito olivocerebeloso comprende la oliva inferior, las
células de Purkinje y los núcleos cerebelosos profundos. Las fibras
trepadoras de la oliva inferior no expresan Kv3.1 ni Kv3.3 (Weiser y
cols., 1994). Estas fibras olivocerebelosas establecen sinapsis con las
células de Purkinje, las cuales sólo expresan Kv3.3, mientras que sus
dianas directas, los núcleos cerebelosos profundos, expresan ambos
Kv3.1 y Kv3.3 (Goldman-Wohl y cols., 1994; Weiser y cols., 1994;
Alonso-Espinaco y cols., 2008). En el núcleo dentado observamos en un
estudio previo de nuestro laboratorio la distribución de Kv3.1b y
Kv3.3 en células excitadoras e inhibidoras (Alonso-Espinaco y cols.,
2008). Una subpoblación de estas neuronas inhibidoras gabaérgicas
proyecta a la oliva inferior, cerrando así el circuito. Este circuito entra
en contacto con la vía de las fibras musgosas-células granulares-
células de Purkinje, en la cual Kv3.1 y Kv3.3 se expresan en las
células granulares (Weiser y cols., 1994). Por su parte, las neuronas
glutamatérgicas del núcleo dentado proyectan al tálamo ventral
lateral y la porción parvocelular del núcleo rojo contralateral, por lo
que ambas subunidades de canales de potasio están implicados en la
transmisión de señales excitadoras en estas vías motoras. También
observamos la presencia de Kv3.3 en axones y terminales sinápticas
de las células de Purkinje (Alonso-Espinaco y cols., 2008). Estos datos,

90
Tesis Doctoral Aranzazu Urresola Olabarrieta
junto con los descritos en la corteza cerebelosa en esta Tesis Doctoral,
ponen de manifiesto que Kv3.1b y Kv3.3 no siempre se encuentran
igualmente distribuidos.
También hay que tener en cuenta que los ratones que carecen de
los genes que codifican para Kv3.1 y Kv3.3 tienen deficiencias motoras
que no se observan en los ratones que sólo carecen de uno de estos dos
genes, lo que sugiere un solapamiento de la expresión de estas
proteínas y, por tanto, una sinergia funcional en algunas poblaciones
neuronales (Espinosa y cols., 2001; McMahon y cols., 2004; Joho y
cols., 2006). En este caso, es lógico pensar, como se ha apuntado
previamente por otros autores, en la puesta en marcha de un
mecanismo de compensación (Matsukawa y cols., 2003, Espinosa y
cols., 2001, 2004), debido además a que las cinéticas de activación de
estas dos subunidades de canales son muy similares. Esto está
avalado por la coexpresión de Kv3.1b y Kv3.3 en muchas estructuras
cerebrales, lo que sugiere que podrían ser componentes de complejos
de canales heteromultiméricos (Christie y cols., 1990; Isacoff y cols.,
1990; McCormack y cols., 1990; Ruppersberg y cols., 1990;
Covarrubias y cols., 1991; Weiser y cols., 1994). Sin embargo, los
resultados de este trabajo Doctoral, así como nuestros estudios en el
núcleo dentado que Kv3.1b y Kv3.3 (Alonso-Espinaco y cols., 2008),

91
Tesis Doctoral Aranzazu Urresola Olabarrieta
sugieren la posibilidad de que estas subunidades modulen acciones
distintas, estando a su vez regidas por vías diferentes de señalización
intracelular.
Los ratones que sólo carecen de uno de los dos genes que
codifican Kv3.1 o Kv3.3 también apoyan esta hipótesis, debido a que
muestran fenotipos diferentes, hecho que no ocurriría si se expresasen
ambas subunidades en las mismas vías y fueran funcionalmente
redundantes. En definitiva, estos ratones knockout deberían tener un
fenotipo lo más parecido al silvestre, sin embargo no es así. Por
ejemplo, la falta de Kv3.1 es principalmente responsable del
incremento de la actividad motora, una reducción de respuesta a
estímulos acústicos y de pérdida de sueño, debido esto último a la
importante presencia de Kv3.1 en el núcleo reticular talámico y en la
corteza cerebral, participando por tanto esta subunidad en la función
talamocortical. De hecho, los ratones knockout de Kv3.1 exhiben una
alteración de los ritmos corticales con aumento de las oscilaciones
rápidas gamma y disminución de las lentas delta registradas en la
corteza somatosensorial (Joho y cols., 1999). Por lo demás, exhiben
actividades locomotoras y exploratorias similares a las de los ratones
control y no presentan una mayor proclividad a desarrollar crisis
epilépticas. Tampoco se ve alterada su capacidad de adquisición

92
Tesis Doctoral Aranzazu Urresola Olabarrieta
(aprendizaje) ni de retención (memoria) (Ho y cols., 1997). La ausencia
de la subunidad Kv3.3 afecta la función cerebelosa, en particular a las
células de Purkinje y a las propiedades del sistema olivocerebeloso
(McMahon y cols., 2004; Zagha y cols., 2008).
Se ha visto en humanos que la carencia del gen que expresa la
proteína Kv3.3 causa una neurodegeneración cerebelosa que cursa con
ataxia cerebelosa en niños y adultos (Waters y cols., 2006; Schorge y
cols., 2010). No se conocen todavía los mecanismos que relacionan la
mutación de Kv3.3 con la atrofia cerebelosa, pero sí parece claro que
los eventos despolarizantes, la duración de los potenciales de acción y
la liberación de neurotransmisores están afectados en los
compartimentos somáticos, dendríticos y axónicos que expresan Kv3.3.
Es necesario iniciar nuevas investigaciones con el fin de
determinar si la alteración de la excitabilidad de las neuronas que
integran los circuitos neuronales corticocerebelosos y de proyección
corticonucleares que contienen Kv3.1 y Kv3.3 como hemos descrito en
este trabajo Doctoral en la corteza cerebelosa, contribuyen a la
aparición del síndrome cerebeloso observado en diferentes especies,
incluido el humano.

93
Tesis Doctoral Aranzazu Urresola Olabarrieta
CONCLUSIONES

94
Tesis Doctoral Aranzazu Urresola Olabarrieta
CONCLUSIÓN GENERAL
La convergencia de las fibras trepadoras y el sistema de fibras
musgosas-células granulares-fibras paralelas en las células de
Purkinje es crucial para un apropiado control motor por el cerebelo.
Ambos circuitos difieren en la presencia de Kv3.1b y Kv3.3 en las
fibras paralelas y en su ausencia de las fibras trepadoras (Weiser y
cols., 1994, Hurlock y cols., 2009). A la vista del hecho de la
localización de las subunidades Kv3.1b y Kv3.3 en las fibras
paralelas y sus terminales sinápticas, que sugiere una redundancia
funcional de estos canales (Matsukawa y cols., 2003), los resultados
anatómicos de esta Tesis Doctoral indican que la distribución
relativa de Kv3.1b contribuye, en mayor medida que Kv3.3, a la
arquitectura molecular de las fibras paralelas de las células
granulares, estando Kv3.3 principalmente localizada en las espinas
dendríticas de las células de Purkinje.

95
Tesis Doctoral Aranzazu Urresola Olabarrieta
CONCLUSIONES
1. Los subunidades de los canales de potasio dependientes de voltaje Kv3.1b y Kv3.3 están localizadas en las membranas de terminales sinápticas y segmentos intervaricosos de las fibras paralelas.
2. En concreto, Kv3.1b está en el 85% de los botones sinápticos, y en cerca del 47% de las porciones intervaricosas de las fibras paralelas.
3. Kv3.3, por su parte, se localiza en el 28% de las intervaricosidades y el 23% de las terminales sinápticas de las fibras paralelas.
4. La densidad de inmunomarcado para Kv3.1b y Kv3.3 es aproximadamente el doble en segmentos intervaricosos que en las terminales sinápticas de las fibras paralelas.
5. Los principales compartimentos de localización de Kv3.3 (54%) son las espinas dendríticas de las células de Purkinje.
6. La densidad de Kv3.3 es significativamente más alta en las espinas dendríticas de las células de Purkinje, que en las terminales sinápticas de las fibras paralelas.
7. Sin embargo, las diferencias de densidad de Kv3.3 entre las espinas dendríticas de las células de Purkinje y los segmentos intervaricosos de las fibras paralelas no son estadísticamente significativas.

96
Tesis Doctoral Aranzazu Urresola Olabarrieta
AGRADECIMIENTOS
Esta Tesis Doctoral ha sido realizada con fondos de las subvenciones
recibidas a Proyectos de Investigación financiados por el Gobierno
Vasco (GIC07/70-IT-432-07) y la Universidad del País Vasco/Euskal
Herriko Unibertsitatea.

97
Tesis Doctoral Aranzazu Urresola Olabarrieta
BIBLIOGRAFÍA

98
Tesis Doctoral Aranzazu Urresola Olabarrieta
BIBLIOGRAFÍA
Abbott,G.W., Butler, M.H., Bendahhou, S., Dalakas, M.C., Ptacek, L.J. y Goldstein, S.A. (2001) MiRP2 forms potassium channels in skeletal muscle with Kv3.4 and is associated with periodic paralysis. Cell 104, 217-231.
Akemann W. y Knöpfel, T. (2006) Interaction of Kv3 potassium channels and
resurgent sodium current influences the rate of spontaneous firing of Purkinje neurons. J. Neurosci. 26, 4602-4612.
Almers, W. y Stirling, C. (1984) Distribution of transport proteins over animal cell
membranes. J. Membr. Biol. 77, 169-186. Alonso-Espinaco, V., Elezgarai, I., Díez-García, J., Puente, N., Knöpfel, T. y Grandes,
P. (2008) Subcellular localization of the voltage-gated potassium channels Kv3.1b and Kv3.3 in the cerebellar dentate nucleus of glutamic acid decarboxylase 67-green fluorescent protein transgenic mice. Neuroscience 155, 1059-1069.
Bagnall, M.W., Stevens, R.J. y du Lac, S. (2007). Transgenic mouse lines subdivide
medial vestibular nucleus neurons into discrete, neurochemically distinct populations. J. Neurosci. 27, 23128-23130.
Baranauskas, G., Tkatch, T. y Surmeier, D.J. (1999) Delayed rectifier currents in rat globus pallidus neurons are attributable to Kv2.1 and Kv3.1/3.2 K(+) channels. J. Neurosci. 19, 6394-6404.
Brooke, R.E., Moores, T.S., Morris, N.P., Parson, S.H. y Deuchars, J. (2004) Kv3 voltage gated potassium channels regulate neurotransmitter release from mouse motor nerve terminals. Eur. J. Neurosci. 20, 3313-3321.
Chandy, K.G. y Gutman, G.A. (1993) Nomenclature for mammalian potassium channel
genes. Trends Pharmacol. Sci. 14, 434. Chan, K.W., Sui, J.L., Vivaudou, M. y Logothetis, D.E. (1997) Specific regions of
heteromeric subunits involved in enhancement of G protein-gated K+ channel activity. J. Biol. Chem. 272, 6548-6555.
Chang, S.Y., Zagha, E., Kwon, E.S., Ozaita, A., Bobik, M., Martone, M.E., Ellisman,
M.H., Heintz, N. y Rudy, B. (2007) Distribution of Kv3.3 potassium channel subunits in distinct neuronal populations of mouse brain. J. Comp. Neurol. 502, 953–972.
Christie, M.J., North, R.A., Osborne, P.B., Douglass, J. y Adelman, J.P. (1990)
Heteropolymeric potassium channels expressed in Xenopus oocytes from cloned subunits. Neuron 4, 405-411.
Covarrubias, M., Wei, A.A. y Salkoff, L. (1991) Shaker, Shal, Shab, and Shaw express
independent K+ current systems. Neuron 7, 763-773.
Coetzee, W., Amarillo, Y., Chiu, J., Chow, A., Lau, D., McCormack, T., Moreno, H., Nadal, M.S., Ozaita, A., Pountney, D., Saganich, M., Vega-Saénz de Miera, E. y Rudy, B. (1999) Molecular diversity of K+ channels. Ann. NY Acad. Sci. 868, 233-285.
Critz, S.D., Wible, B.A., Lopez, H.S. y Brown, A.M. (1993) Stable expression and
regulation of a rat brain K+ channel. J. Neurochem. 60, 1175-1178.
De Zeeuw, C.I., Simpson, J.I., Hoogenraad, C.C., Galjart, N., Koekoek, S.K.E. y Ruigrok, T.H.J. (1998) Microcircuitry and function of the inferior olive. Trends Neurosci. 21, 391-400.

99
Tesis Doctoral Aranzazu Urresola Olabarrieta
Deuchars, S.A., Brooke, R.E., Frater, B. y Deuchars, J. (2001) Properties of
interneurons in the intermediolateral cell column of the rat spinal cord: Role of the potassium channel subunit Kv3.1. Neuroscience 106, 433-446.
Devaux, J., Alcaraz, G., Grinspan, J., Bennett, V., Joho, R., Crest, M. y Scherer, S.S.
(2003) Kv3.1b is a novel component of CNS nodes. J. Neurosci. 23, 4509-4518.
Drewe, J.A., Verma, S., Frech, G. y Joho, R.H. (1992) Distinct spatial and temporal expression patterns of K+ channel mRNAs from different subfamilies. J. Neurosci. 12, 538-548.
Du, J., Zhang, L., Weiser, M., Rudy, B. y McBain, C.J. (1996) Developmental
expression and functional characterization of the potassium-channel subunit Kv3.1b in parvalbumin-containing interneurons of the rat hippocampus. J. Neurosci. 16, 506-518.
Elezgarai, I., Díez, J., Puente, N., Azkue, J.J., Benítez, R., Bilbao, A., Knöpfel, T.,
Doñate-Oliver, F. y Grandes, P. (2003) Subcellular localization of the voltage-dependent potassium channel Kv3.1b in postnatal and adult rat medial nucleus of the trapezoid body. Neuroscience 118, 889-898.
Erisir, A., Lau, D., Rudy, B. y Leonard, C.S. (1999) Function of specific K(+) channels in sustained high-frequency firing of fast-spiking neocortical interneurons. J. Neurophysiol. 82, 2476-2489.
Espinosa, F., Marks, G., Heintz, N. y Joho, R.H. (2004) Increased motor drive and
sleep loss in mice lacking Kv3-type potassium channels. Genes Brain Behav. 3, 90-100.
Espinosa, F., McMahon, A., Chan, E., Wang, S., Ho, Ch.Sh., Heintz, N. y Joho, R.H.
(2001) Alcohol hypersensitivity, increased locomotion, and spontaneous myoclonus in
mice lacking the potassium channels Kv3.1 and Kv3.3. J. Neurosci. 21: 6657-6665.
Fukuda, M., Yamamoto, T., y Llinás, R., (2001) The isochronic band hypothesis and
climbing fibre regulation of motricity: an experimental study. Eur. J. Neurosci. 13, 315-
326.
Gan, L. y Kaczmarek, L.K. (1998) When, where, and how much? Expression of the Kv3.1 potassium channel in high-frequency firing neurons. J. Neurobiol. 37, 69-79.
Gan, L., Perney, T.M. y Kaczmarek, L.K. (1996) Cloning and characterization of the
promoter for a potassium channel expressed in high-frequency firing neurons. J. Biol. Chem. 271, 5859-5865.
Gittis, A.H. y du Lac, S. (2007). Firing properties of GABAergic versus non-GABAergic
vestibular nucleus neurons conferred by a differential balance of potassium currents. J. Neurophysiol. 97, 3986-3996.
Goldman-Wohl, D.S., Chan, E., Baird, D. y Heintz, N. (1994) Kv3.3b: a novel Shaw
type potassium channel expressed in terminally differentiated cerebellar Purkinje cells and deep cerebellar nuclei. J. Neurosci. 14, 511–522.
Grandes, P., Ortega, F. y Streit, P. (1994) Glutamate-immunoreactive climbing fibres
in the cerebellar cortex of the rat. Histochemistry 101, 427-437. Grigg, J.J., Brew, H.M. y Tempel, B.L. (2000) Differential expression of voltage-gated
potassium channel genes in auditory nuclei of the mouse brainstem. Hear Res. 140, 77-90.
Grissmer, S., Nguyen, A.N., Aiyar, J., Hanson, D.C., Mather, R.J., Gutman, G.A.,
Karmilowicz, M.J., Auperin, D.D. y Chandy, K.G. (1994) Pharmacological

100
Tesis Doctoral Aranzazu Urresola Olabarrieta
characterization of five cloned voltage-gated K+ channels, types Kv1.1, 1.2, 1.3, 1.5, and 3.1, stably expressed in mammalian cell lines. Mol. Pharmacol. 45, 1227-1234.
Gutman, G.A,, Chandy, K.G., Grissmer, S., Lazdunski, M., McKinnon, D., Pardo, L.A.,
Robertson, G.A., Rudy, B., Sanguinetti, M.C., Stühmer, W. y Wang, X. (2005) International Union of Pharmacology. LIII. Nomenclature and molecular relationships of voltage-gated potassium channels. Pharmacol. Rev. 57, 473-508.
Härtig, W., Singer, A., Grosche, J., Brauer, K., Ottersen, O.P. y Brückner (2001) Perineuronal nets in the rat medial nucleus of the trapezoid body surround neurons immunoreactive for various amino acids, calcium-binding proteins and the potassium channel subunit Kv3.1b. Brain Res. 899, 123-133.
Hendricks, R., Morest, D.K. y Kaczmarek, L.K. (1999) Shaw-like potassium currents in
the auditroy rhombencephalon throughout embryogenesis. J. Neurosci. Res. 15, 791-804. Hernández-Pineda, R., Chow, A., Amarillo, Y., Moreno, H., Saganich, M., Vega-Sáenz
de Miera, E.C., Hernández-Cruz, A. y Rudy, B. (1999) Kv3.1-Kv3.2 channels underlie a high-voltage-activating component of the delayed rectifier K+ current in projecting neurons from the globus pallidus. J. Neurophysiol. 82, 1512-1528.
Hille, B., Armstrong, C.M. y MacKinnon, R. (1999) Ion channels: from idea to reality. Nat. Med. 5, 1105-1109.
Ho, C.S., Grange, R.W. y Joho, R.H. (1997) Pleiotropic effects of a disrupted K+ channel
gene: reduced body weight, impaired motor skill and muscle contraction, but no seizures. Proc. Natl. Acad. Sci. USA 94, 1533-1538.
Hsu, S.M. Raine, L. y Fanger, H. (1981) Use of avidin-biotin-peroxidase complex (ABC)
in immunoperoxidase techniques: a comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cytochem. 29, 577.
Hurlock, E.C., Bose, M., Pierce, G. y Joho, R.H. (2009) Rescue of motor coordination by
Purkinje cell-targeted restoration of Kv3.3 channels in Kcnc3-null mice requires Kcnc1. J. Neurosci. 29, 15735-15744.
Hurlock, E.C., McMahon, A. y Joho, R.H. (2008) Purkinje-cell-restricted restoration of
Kv3.3 function restores complex spikes and rescues motor coordination in Kcnc3 mutants. J. Neurosci. 28, 4640-4648.
Isacoff, E.Y., Jan, Y.N. y Jan, L.Y. (1990) Evidence for the formation of
heteromultimeric potassium channels in Xenopus oocytes. Nature 345, 530-534.
Ito, M. (2008) Control of mental activities by internal models in the cerebellum. Nat.
Rev. Neurosci. 9, 304-313. Jan, L.Y. y Jan, Y.N. (1997) Cloned potassium channels from eukaryotes and
prokaryotes. Annu. Rev. Neurosci. 20, 91-123. Joho, R.H., Ho, C.S. y Marks, G.A. (1999) Increased gamma- and decreased delta-
oscillations in a mouse deficient for a potassium channel expressed in fast-spiking interneurons. J. Neurophysiol. 82, 1855-1864.
Joho, R.H., Street, C., Matsushita, S. y Knöpfel, T. (2006) Behavioral motor
dysfunction in Kv3-type potassium channel-deficient mice. Genes Brain Behav. 5, 472–482.
Lenz, S., Perney, T.M., Qin, Y., Robbins, E. y Chesselet, M.F. (1994) GABA-ergic
interneurons of the striatum express the Shaw-like potassium channel Kv3.1. Synapse 18, 55-66.

101
Tesis Doctoral Aranzazu Urresola Olabarrieta
Li, W., Kaczmarek, L.K. y Perney, T.M. (2001) Localization of two high-threshold
potassium channel subunits in the rat central auditory system. J. Comp. Neurol. 437, 196-218.
Lien, C.C. y Jonas, P. (2003) Kv3 potassium conductance is necessary and kinetically
optimized for high-frequency action potential generation in hippocampal interneurons. J. Neurosci. 23, 2058–2068.
Llinás, R.R. (1988) The intrinsic electrophysiological properties of mammalian neurons: Insights into central nervous system function. Science 242, 1654-1664.
Llinás, R. y Sugimori, M. (1980). Electrophysiological properties of in vitro Purkinje
cell dendrites in mammalian cerebellar slices. J. Physiol. 305, 197-213.
Luneau, C.J., Williams, J.B., Marshall, J., Levitan, E.S., Oliva, C., Smith, J.S., Antanavage, J., Folander, K., Stein, R.B., Swanson, R., Kaczmarek, L.K. y Buhrow, S.A. (1991) Alternative splicing contributes to K+ channel diversity in the mammalian central nervous system. Proc. Natl. Acad. Sci. USA 88, 3932-3936.
Macica, C.M. y Kaczmarek, L.K. (2001) Casein kinase 2 determines the voltage
dependence of the Kv3.1 channel in auditory neurons and transfected cells. J. Neurosci. 21, 1160-1168.
Macica, C.M., von Hehn, C.A., Wang, L.Y., Ho, C.S., Yokoyama, S., Joho, R.H. y
Kaczmarek, L.K. (2003) Modulation of the kv3.1b potassium channel isoform adjusts the fidelity of the firing pattern of auditory neurons. J Neurosci. 23, 1133-1141.
Martina, M., Schultz, J.H., Ehmke, H., Monyer, H. y Jonas, P. (1998) Functional and
molecular differences between voltage-gated channels of fast-spiking interneurons and pyramidal neurons of rat hippocampus. J. Neurosci. 18, 8111-8125.
Martina, M., Yao, G.L. y Bean, B.P. (2003) Properties and functional role of voltage-dependent potassium channels in dendrites of rat cerebellar Purkinje neurons. J. Neurosci. 23, 5698–5707.
Massengill, J.L., Smith, M.A., Son, D.I. y O'Dowd, D.K. (1997) Differential expression
of K4-AP currents and Kv3.1 potassium channel transcripts in cortical neurons that
develop distinct firing phenotypes. J. Neurosci. 17, 3136-3147.
Matsukawa, H., Wolf, A.M., Matsushita, Sh., Joho, R.H. y Knöpfel, T. (2003) Motor
dysfunction and altered synaptic transmission at the parallel fiber-Purkinje cell synapse
in mice lacking potassium channels Kv3.1 and Kv3.3. J. Neurosci. 23, 7677-7684.
McKay, B.E. y Turner, R.W. (2004) Kv3 K+ channels enable burst output in rat
cerebellar Purkinje cells. Eur. J. Neurosci. 20, 729-739.
McCormack, T., Vega-Saenz de Miera, E.C. y Rudy, B. (1990) Molecular cloning of a
member of a third class of Shaker-family K+ channel genes in mammals. Proc. Natl.
Acad. Sci. USA 87, 5227-5231.
McDonald, A.J. y Mascagni, F. (2006) Differential expression of Kv3.1b and Kv3.2
potassium channel subunits in interneurons of the basolateral amygdala. Neuroscience
138, 537-547.

102
Tesis Doctoral Aranzazu Urresola Olabarrieta
McMahon, A., Fowler, S.C., Perney, T.M., Akemann, W., Knöpfel, T. y Joho, R.H.
(2004) Allele-dependent changes of olivocerebellar circuit properties in the absence of
the voltage-gated potassium channels Kv3.1 and Kv3.3. Eur. J. Neurosci. 12, 3317-3327.
Metzger, F., Repunte-Canonigo, V., Matsushita, S., Akemann, W., Díez-García, J., Ho,
Ch.S., Iwasato, T., Grandes, P., Itohara, S., Joho, R.H. y Knöpfel, T. (2002) Transgenic
mice expressing a pH and Cl- sensing yellow-fluorescent protein under the control of a
potassium channel promoter. Eur. J. Neurosci. 15, 40-50.
Milner, T.E., Cadoret, G., Lessard, L. y Smith, A.M. (1995) EMG analysis of
harmalineinduced tremor in normal and three strains of mutant mice with Purkinje cell
degeneration and the role of the inferior olive. J Neurophysiol. 73, 2568-2577.
Ozaita, A., Martone, M.E., Ellisman, M.H. y Rudy, B. (2002) Differential subcellular
localization of the two alternatively spliced isoforms of the Kv3.1 potassium channel
subunit in brain. J. Neurophysiol. 88, 394-408.
Palay, S.L. y Chan-Palay, V. (1974) Cerebellar Cortex. Springer, Berlin Heidelberg
New York.
Parameshwaran, S., Carr, C.E. y Perney, T.M. (2001) Expression of the Kv3.1 potassium channel in the avian auditory brainstem. J. Neurosci. 21, 485-494.
Perney, T.M. y Kaczmarek, L.K. (1991) The molecular biology of K+ channels. Curr.
Opin. Cell Biol. 3, 663-670. Perney, T.M. y Kaczmarek, L.K. (1997) Localization of a high threshold potassium
channel in the rat cochlear nucleus. J. Comp. Neurol. 386, 178-202. Perney, T., Marshall, J., Martin, K.A., Hockfield, S. y Kaczmarek, L.K. (1992)
Expression of the mRNA for the Kv3.1 potassium channel gene in the adult and developing rat brain. J. Neurophysiol. 68, 756-766.
Pongs, O. (1992) Molecular biology of voltage-dependent potassium channels. Physiol.
Rev. 72, S69-S88. Porcello, D.M., Ho, C.S., Joho, R.H. y Huguenard, J.R. (2002) Resilient RTN fast
spiking in Kv3.1 null mice suggests redundancy in the action potential repolarization mechanism. J. Neurophysiol. 87, 1303-1310.
Prinz, A.A., Bucher, D. y Marder, E. (2004) Similar network activity from disparate
circuit parameters. Nat. Neurosci. 7, 1345-1352.
Puente, N., Mendizabal-Zubiaga, J., Elezgarai, I., Reguero, L., Buceta, I. y Grandes, P. (2010) Precise localization of the voltage-gated potassium channel subunits Kv3.1b and Kv3.3 revealed in the molecular layer of the rat cerebellar cortex by a pre-embedding immunogold method. Histochem. Cell Biol. 134, 403-409.
Raman, I.M. y Bean, B.P. (1999) Properties of sodium currents and action potential
firing in isolated cerebellar Purkinje neurons. Ann. N.Y. Acad. Sci. USA 868, 93-96.
Rettig, J., Heinemann, S.H., Wunder, F., Lorra, C., Parcej, D.N., Dolly, J.O. y Pongs, O. (1994) Inactivation properties of voltage-gated K+ channels altered by presence of beta-subunit. Nature 369, 289-294.
Rudy, B., Chow, A., Lau, D., Amarillo, Y., Ozaita, A., Saganich, M., Moreno, H., Nadal, M. S., Hernández-Pineda, R., Hernández-Cruz, A., Erisir, A., Leonard, C. y Vega-Saenz

103
Tesis Doctoral Aranzazu Urresola Olabarrieta
de Miera, E. (1999) Contributions of Kv3 channels to neuronal excitability. In Molecular and Functional Diversity of Ion Channels and Receptors, Annals of the New York Academy of Sciences 868, 304-343.
Rudy, B., Kentros, C., Weiser, M., Fruhling, D., Serodio, P., Vega-Sáenz de Miera, E.,
Ellisman, M.H., Pollock, J.A. y Baker, H. (1992) Region-specific expression of a K+ channel gene in brain. Proc. Natl. Acad. Sci. USA 89, 4603-4607.
Rudy B. y McBain C.J. (2001) Kv3 channels: voltage-gated K+ channels designed for
high-frequency repetitive firing. Trends Neurosci. 24, 517–526. Ruppersberg, J.P., Schröter, K.H., Sakmann, B., Stocker, M., Sewing, S. y Pongs, O.
(1990) Heteromultimeric channels formed by rat brain potassium-channel proteins. Nature 345, 535-537.
Sabatini, B.L. y Regehr, W.G. (1997) Control of neurotransmitter release by
presynaptic waveform at the granule cell to Purkinje cell synapse. J. Neurosci. 17, 3425–3435.
Salkoff, L., Baker, K., Butler, A., Covarrubias, M., Pak, M.D. y Wei, A. (1992) An
essential 'set' of K+ channels conserved in flies, mice and humans. Trends Neurosci. 15, 161-166.
Sánchez, J.A., Ho, C.S., Vaughan, D.M., Garcia, M.C., Grange, R.W. y Joho, R.H. (2000) Muscle and motor-skill dysfunction in a K+ channel-deficient mouse are not due to altered muscle excitability or fiber type but depend on the genetic background. Pflügers Arch. 440, 34-41.
Schorge, S., van de Leemput, J., Singleton, A., Houlden, H. y Hardy, J. (2010) Human
ataxias: a genetic dissection of inositol triphosphate receptor (ITPR1)-dependent signalling. Trends Neurosci. 33, 211-219.
Schröter, K.H., Ruppersberg, J.P., Wunder, F., Rettig, J., Stocker, M. y Pongs, O. (1991) Cloning and functional expression of a TEA-sensitive A-type potassium channel from rat brain. FEBS Lett. 278, 211-216.
Schulz, D.J., Baines, R.A., Hempel, C.M., Li, L., Liss, B. y Misonou, H. (2006) Cellular
excitability and the regulation of functional neuronal identity: from gene expression to neuromodulation. J. Neurosci. 26, 10362-10367.
Scott, V.E., Rettig, J., Parcej, D.N., Keen, J.N., Findlay, J.B., Pongs, O. y Dolly, J.O.
(1994) Primary structure of a beta subunit of alpha-dendrotoxin-sensitive K+ channels from bovine brain. Proc. Natl. Acad. Sci. USA. 91, 1637-1641.
Sekirnjak, C., Martone, M.E., Weiser, M., Deerinck, T., Bueno, E., Rudy, B. y Ellisman, M. (1997) Subcellular localization of K+ channel subunit Kv3.1b in selected rat CNS neurons. Brain Res. 766, 173-187.
Sheng, M., Liao, Y.J., Jan, Y.N. y Jan, L.Y. (1993) Presynaptic A-current based on
heteromultimeric K+ channels detected in vivo. Nature 365, 72-75. Shibata, R., Wakazono, Y., Nakahira, K., Trimmer, J.S. y Ikenaka, K. (1999)
Expression of Kv3.1 and Kv4.2 genes in developing cerebellar granule cells. Dev. Neurosci. 21, 87-93.
Song, P., Yang, Y., Barnes-Davies, M.,Bhattacharjee, A., Hamann, M., Forsythe, I.D.,
Oliver, D.L. y Kaczmarek, L.K. (2005) Acoustic environment determines phosphorylation state of the Kv3.1 potassium channel in auditory neurons. Nat. Neurosci. 8, 1335–1342.

104
Tesis Doctoral Aranzazu Urresola Olabarrieta
Southan, A.P. y Robertson, B. (2000) Electrophysiological characterization of voltage-
gated K(+) currents in cerebellar basket and Purkinje cells: Kv1 and Kv3 channel subfamilies are present in basket cell nerve terminals. J. Neurosci. 20, 114-122.
Uusisaari, M., Obata, K. y Knöpfel, T. (2007) Morphological and electrophysiological
properties of GABAergic and non-GABAergic cells in the deep cerebellar nuclei. J.
Neurophysiol. 97, 901-911.
Vega-Sáenz de Miera, E., Weiser, M., Kentros, C., Lau, D., Moreno, H., Serodio, P. y Rudy, B. (1994) Shaw-related K+ channels in mammals. En: Peraccia C. (ed.) Handbook of Membrane Channels. Academic, Orlando, págs.: 41-78.
Vincent, A., Lautermilch, N.J. y Spitzer, N.C. (2000) Antisense suppression of
potassium channel expression demonstrates its role in maturation of the action potential. J. Neurosci. 20, 6087-6094.
Vullhorst, D., Jockusch, H. y Bartsch, J.W. (2001) The genomic basis of K(V)3.4
potassium channel mRNA diversity in mice. Gene 264, 29-35. Wang, L.Y., Gan, L., Forsythe, I.D. y Kaczmarek, L.K. (1998a) Contribution of the
Kv3.1 potassium channel to high-frequency firing in mouse auditory neurons. J. Physiol. 509, 183-194.
Wang, L.Y., Gan, L., Perney, T.M., Schwartz, I. y Kaczmarek, L.K. (1998b) Activation
of Kv3.1 channels in neuronal spine-like structures may induce local potassium ion depletion. Proc. Natl. Acad, Sci. USA 95, 1882-1887.
Waters, M.F., Minassian, N.A., Stevanin, G., Figueroa, K.P., Bannister, J.P., Nolte, D., Mock, A.F., Evidente, V.G., Fee, D.B., Müller, U., Dürr, A., Brice, A., Papazian, D.M. y Pulst, S.M. (2006) Mutations in voltage-gated potassium channel KCNC3 cause degenerative and developmental central nervous system phenotypes. Nat. Genet. 38, 447-451.
Waters, M.F. y Pulst, S.M. (2008) Sca13. Cerebellum 7, 165-169.
Weiser, M., Bueno, E., Sekirnjak, C., Martone, M.E., Baker, H., Hillman, D., Chen, S.,
Thornhill, W. y Rudy, B. (1995) The potassium channel subunit Kv3.1b is localized to somatic and axonal membranes of specific populations of CNS neurons. J. Neurosci. 15, 4298-4314.
Weiser, M., Vega-Sáenz de Miera, E., Kentros, C., Moreno, H., Franzen, L., Hillman,
D., Baker, H. y Rudy, B. (1994) Differential expression of Shaw-related K+ channels in the rat central nervous system. J. Neurosci. 14, 949-972.
Welsh, J.P. Lang, E.J., Sugihara, I. y Llinás, R. (1995) Dynamic organization of motor
control within the olivocerebellar system. Nature 374, 453-457. Wigmore, M.A. y Lacey, M.G. (2000) A Kv3-like persistent, outwardly rectifying, Cs+-
permeable, K+ current in rat subthalamic nucleus neurones. J. Physiol. 527, 493-506. Xu, M., Cao, R., Xiao, R., Zhu, M.X. y Gu, C. (2007) The axon-dendrite targeting of Kv3
(Shaw) channels is determined by a targeting motif that associates with the T1 domain and ankyrin G. J. Neurosci. 27, 14158-14170.
Zagha E., Lang E.J. y Rudy B. (2008) Kv 3.3 channels at the Purkinje cells soma are necessary for generation of the classical complex spike waveform. J. Neurosci. 28, 1291-1300.
Zucker R.S. y Regehr W.G. (2002) Short-term synaptic plasticity. Annu. Rev. Physio.
64, 355–405.