sodium bicarbonate effects on the anaerobic digestion of refuse
TRANSCRIPT

J. Chem. Tech. Biotechnol. 1989, 45, 279-289
Sodium Bicarbonate Effects on the Anaerobic Digestion of Refuse
George B. Kasali,* Eric Seniorf & Irene A. Watson-Craik
Department of Bioscience and Biotechnology, University of Strathclyde, 204 George Street, Glasgow G1 lXW, UK
(Received 9 May 1988; revised version received 24 October 1988; accepted 25 October 1988)
ABSTRACT
Laboratory column and bottle cultures were used to examine the efficacy of NaHCO, additions ( I %, 2.5 % and 5 % w/v) to promote the solid-state refuse fermentation. Buffering capacity increase, by addition of 2.5 % NaHCO,, equivalent to 84 mg NaHCO, g dry weight refuse-', promoted both acidogenesis and methanogenesis, with the latter accelerating by a factor of six, compared with glass-distilled water controls. Increased addition, with 5 "/, NaHCO,, however, mediated partial suppression of methanogenesis and acidoyenesis with cation toxicity the most likely cause. Promotion of the refuse fermentation by NaHCO, addition, therefore, necessitates careful control of the concentration applied.
Key words: refuse, acidogenesis, methanogenesis, sodium bicarbonate.
1 INTRODUCTION
Effective exploitation of a refuse mass as a source of methane necessitates establishment and maintenance of a stable solid-state methanogenic fermentation. To facilitate this, manipulation of a specific site variable@), such as refuse composition and pretreatment,'-, refuse density and moisture ~ o n t e n t , ~ - ~ temperature,' or pH and alkalinity,2*3,'0 may be considered, possibly in conjunction with microbial seeding or nutrient addition, although constraints may be imposed by the increasingly complex site licence conditions.
* Present address: National Council for Scientific Research, PO Box CH-158. Chelstone, Lusaka, Zambia.
To whom all correspondence should be addressed.
279 J . Chem. Tech. Biotechnol. 0268-2575/89/$03.50 0 1989 Society of Chemical Industry. Printed in Great Britain

280 G. 8. Kasali, E . Senior, I . A. Warson-Craik
Following refuse placement, the methanogenic bacteria are soon outnumbered by the acidogenic species due to their sensitivity to oxygen and their lower specific growth rates. The net result is an accumulation of carboxylic acids, and thus a lowering of the pH, to the detriment of the methanogens. A general deceleration of the fermentation is then apparent, as observed by Mao and Pohland,' in sewage- sludge-seeded lysimeter studies, and by Leckie et ~ 1 . ~ and Harmsen" in situ.
To establish a stable refuse methanogenic fermentation it has been suggested',' that buffering agents such as lime, calcium carbonate or sodium bicarbonate should be added. Since acidogenic phase landfill leachate has been shown to have a pH of 5.7 and a bicarbonate content of 17 mg dm-3 in contrast with methanogenic-phase leachate of pH 7.0 and 12 000 mg dm-3 bicarbonate," this would appear to be a valid approach. Also, sodium bicarbonate additions have been used to maintain favourable pH values in fermenting refuse samples and, as a consequence, increase gas production.l4*l5
Together with pH maintenance, the potentially expensive' alkali additions required to establish the acetic acid:alkalinity ratio < 0.8 necessary to initiate methanogenesis," may effect chemical composition changes.I6 Thus promotion of refuse methanogenesis by calcium carbonate additionsI3 may be due to a combination of pH elevation and precipitation of potentially toxic heavy metals.' '
Despite such promotions, problems remain with respect to the mechanism of introducing strong alkalis to a digestion system and the possible toxic effects which may result. Also, large pH fluctuations, even in the presence of alkali additions, may be apparent due to acidogenesis and acidotrophy. This study was, therefore, initiated to make a preliminary examination of the effects of one selected alkali, sodium bicarbonate, on the interrelated processes of acidogenesis and methanogenesis in fermenting refuse.
2 EXPERIMENTAL MATERIALS AND METHODS
2.1 Refuse
Following emplacement, for a period in excess of 1 month, refuse was sampled from Kilgarth Landfill, Glasgow (UK) from a depth 2 2 m. The refuse was handsorted to remove stone, glass, metallic and ceramic components and then homogenised for 10 s with a kitchen blender (Krups).
2.2 Closed cultures
2.2.1 Refuse columns Samples (350 g) of homogenised refuse, with a moisture content of 55 % (w/w), were packed into each of four glass columns (940 cm3) which were fitted with calibrated gas collection cylinders" which contained a solution of sodium chloride (20% w/v) acidified with citric acid (0.5 % w/v). The columns were sealed at the base with Suba- seals (Gallenkamp) prior to individual saturation with 500 cm3 of oxygen-free nitrogen (OFN) (British Oxygen)-deoxygenated 1 %, 2.5 % or 5 (w/v) sodium bicarbonate or glass-distilled water (control). After closing with rubber bungs, the

Refuse digestion: sodium bicarbonate promotion 281
columns were maintained at 4°C overnight. Subsequently, the Suba-seals were removed and all excess liquids drained to poise the refuse moisture contents at field capacity (77 % w/w). The columns were then resealed prior to incubation at 30°C for 92 days, during which time gas samples were taken at regular intervals for methane assay.
2.2.2 Bottle cultures Bottles (164cm3) were batched with 10-g samples of the same refuse prior to individual supplementation with OFN-deoxygenated 1 %, 2.5 % or 5 % (w/v) sodium bicarbonate or glass-distilled water (control) to moisture contents of refuse field capacity. The bottles were closed with Suba-seals and incubated at 30°C for 48 days. The generated gases were displaced with OFN at regular intervals and triplicate samples for each treatment were removed at discrete time intervals for volatile fatty acid assay.
2.3 Analytical methods
2.3. I Methane Volumes of evolved methane were quantified using gas chromatography, as described previously.'8
2.3.2 Volatile fatty acids Volatile fatty acids were extracted and assayed by gas chromatography, as described previously."
2.3.3 p H A slurry of one part by weight refuse to two parts by volume glassdistilled water was prepared and allowed to stand at ambient temperature for 10 min after which the supernatant pH was determined by use of a Corning 7 pH meter.
2.3.4 Rejuse dry weight Samples of refuse were placed in dried ceramic crucibles of known weight before reweighing. The samples were then oven-dried (105°C) to constant weight.
3 RESULTS AND DISCUSSION
Two experimental systems were used in this study. Refuse columns were chosen to examine gas evolution to minimise variability by increasing the sample size. Triplicate bottle cultures were selected for volatile fatty acid metabolism examination since the method of analysis necessitated destructive sampling. Displacement of evolved gas was an essential feature of this method of batch cultivation since this ensured that possible gas overpressure-mediated fermentation balance changes" were obviated.
After deoxygenation, the pH values of 1 %, 2.5 % and 5 % sodium bicarbonate solutions and glass-distilled water were 8.8, 8.5, 8.3 and 7-0, respectively. In water

282 G . B . Kasali, E . Senior, I . A. Watson-Craik
sodium bicarbonate ionises as follows:
NaHCO,+Na++HCO; (1)
(2)
HC0i-+C032- + H f (3)
HCO; + H 2 0 --* H2C03 + -OH
Following supplementation, the gross pH values of the equivalent refuse samples were 7.2, 7.3, 7.7 and 7.1, thus indicating possible consumption of bicarbonate alkalinity rather than interaction of refuse carbonates with carbonic acid in bicarbonate formation.
TABLE 1 Volumes of Methane and Total Gas Evolved from Refuse Columns, in the Presence of NaHCO, or Glass-distilled Water Addition, During 92 Days
Incubation at 30°C
Addition Evolved gas volume (dm3 k g dry weight refuse-')
Methane Total
NaHCO, (1 w/v) NaHCO, (2.5 %) NaHCO, ( 5 % ) Glassdistilled water
(control)
55.7 63.9 16.2
17.0
81.9 103.9 48.8
32.3
During the 92day incubation period the presence of sodium bicarbonate effected increases in the total volumes ofgas evolved (Table 1). Comparable increases in the volumes of methane evolved were, however, only recorded with the 1 % and 2.5 % sodium bicarbonate-treated samples since 5 % sodium bicarbonate exerted different effects on the acidogens and methanogens although some of the gas may have been generated chemically :
H + +NaHCO,+Na++H,O+CO, (4)
in the presence of active acidogenesis. Together with total volume increases, sodium bicarbonate additions also effected
rate increases during the first four weeks of incubation (Fig. I). Subsequently, decelerations were evident with the 5 % (w/v)-supplemented column and were accompanied by similar reductions in the methane evolution rates (Fig. 2). The calculated geometric means of the total gas evolution rates were 0-99,1.25,0*67 and 0.34dm3 kg dry weight refuse-' day-' for the 1 %, 2 5 % and 5 % sodium bicarbonate and glassdistilled water-treated samples, respectively. Sodium bicarbonate (2.5 %)-mediated gas evolution rate increases were reported by DeWalle et a1." although, unlike our study, methanogenesis was severely stressed.
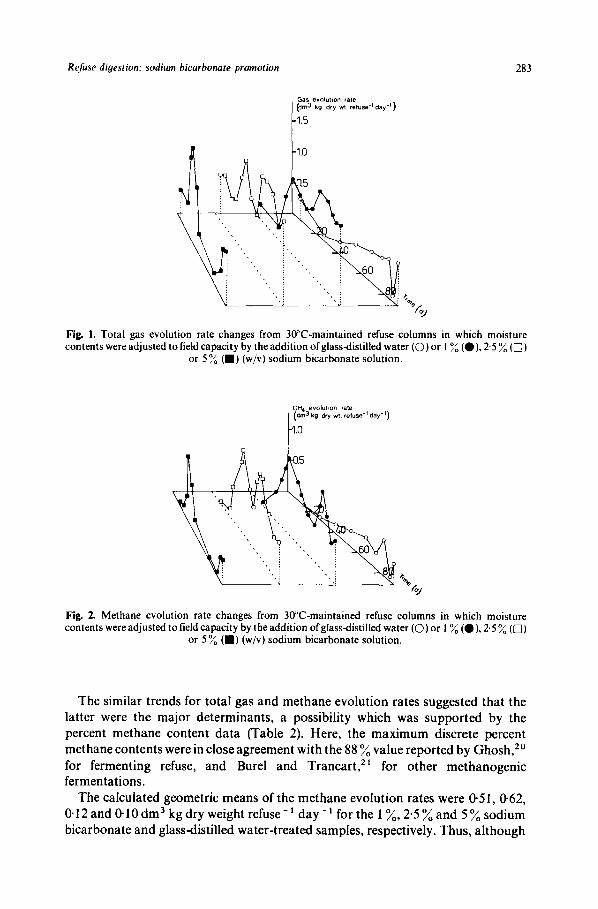
Refuse diyestion: sodium bicarbonate promotion 283
Gas evolulton rate I (dm3 kg dry w1 retuse.lday.l)
Fig. 1. Total gas evolution rate changes from 30°C-maintained refuse columns in which moisture contents were adjusted to field capacity by the addition of glassdistilled water (0) or 1 % (a), 2.5 % (0)
or 5 % (W) (w/v) sodium bicarbonate solution.
CH4 evolution rate kp dry wt retuse.'day-')
Fig. 2. Methane evolution rate changes from 30°C-maintained refuse columns in which moisture contents wereadjusted to fieldcapacity by theaddition ofglassdistilled water (0) or 1 %(a), 2 5 % (0)
or 5 % (W) (w/v) sodium bicarbonate solution.
The similar trends for total gas and methane evolution rates suggested that the latter were the major determinants, a possibility which was supported by the percent methane content data (Table 2). Here, the maximum discrete percent methane contents were in close agreement with the 88 %value reported by Ghosh," for fermenting refuse, and Burel and Trancart," for other methanogenic fermentations.
The calculated geometric means of the methane evolution rates were 0.51, 0.62, 0.12 and 0.10dm3 kgdry weight refuse-' day-' for the 1 %, 2*5%and 5 % sodium bicarbonate and glassdistilled water-treated samples, respectively. Thus, although

284 G. B . Kasali, E . Senior, I. A . Watson-Craik
TABLE 2 Percent Range and Mean Methane Contents of Gases Evolved from Refuse Columns, in the Presence of NaHCO, or Glass-
distilled Water Addition, During 92 Days Incubation at 30°C
Amendment Percent methane content
Range Mean
NaHCO, (1 "/, w/v) 1'3-90.5 70.3 NaHCO, (2.5 %) 0'3-89'8 71.4 NaHCO, (5%) 0'1-68.5 42.7 Glassdistilled water
(control) 0.4-85.5 53.5
1 % and 2.5 % sodium bicarbonate additions effected methane evolution rate increases, the relationship was not linear which suggested that the principal role of the alkali was maintenance of a favourable pH regime for methanogenesis.
An explanation for the 5 % sodium bicarbonate-mediated inhibitory effects during the later stages of the fermentation is not readily apparent. It may be speculated, however, that refuse acidogenesis could have promoted sodium ion dissociation-eqn (4). Unlike carbon dioxide, which may have either entered into reactions with other chemical species or evolved from the refuse, the sodium ions must have remained. It is possible, therefore, that the added Na' (14.5 g dm-3) exerted an inhibitory effect sinceconcentrations of 14 g dm-3 have been reported to effect a 50 % reduction in methanogenesis.22 Since carboxylic acid acidification of refuse increases the ionic strength, it is possible that ionisation of the refuse sodium salts increased as the acidogenic phase of catabolism progressed. In this eventuality, the cation toxicity exerted by the free sodium ions would be regulated by the carboxylic acid concentration. In our study, inhibition of methanogenesis in the refuse columns, after 5 weeks incubation (Fig. 2) was coincident with an accumulation of volatile fatty acids (acetate, propionate, butyrate, isobutyrate, isovalerate, valerate and caproate) in the equivalent bottle cultures. Caution must, however, be exercised in comparing the results of two different experimental systems.
Together with the methane volume and evolution rate increases, 1 % and 2.5 % sodium bicarbonate supplementation mediated fermentation balance changes (Table 2). Possible explanations for these changes include carbon dioxide retention due to increased gross pH values and increased carbon dioxide methanogenesis.
In actual landfills, methanogenesis has been reported to be the major process controlling acidotrophy.'. ' Thus, determination of the effects of sodium bicarbonate additions on acidogenesis could best be made during the early stage of the refuse fermentation when methanogenesis is minimal. During this period, acidogenesis increased with all the treatments although the promotion recorded with the 5 % sodium bicarbonate-supplemented samples was lower than the control (Fig. 3).

Rejuse digestion: sodium hicarbonute promotion
Total volatile fatty acld COK"
~
Lo (nu g dry wt refuse.') ! 285
Fig. 3. Changes in concentrations of 'total' volatile fatty acids (acetate, propionate, butyrdte, isobutyrate, isovalerate, valerate, caproate) in refuse samples during bottle culture cultivation at 30°C. Moisture contents wereadjusted to fieldcapacity by theaddition ofglassdistilled water ( 0 ) o r I (O), 2.5:; (m),
or 5 % (W) (w/v) sodium bicarbonate solution.
Although the relationship between acidogenesis and methanogenesis is often indirect, it may be speculated that in refuse a direct relationship exists between acetogenesis and methanogenesis since acetate is the dominant methane precursor." In our study, during the first 2 weeks of incubation, acetogenesis was promoted by 1 % and 2.5 "/, sodium bicarbonate but reduced by the 5 % treatment relative to the control (Fig. 4). Subsequently, with the exception of the 5 % treatment, reductions in discrete acetate concentrations (balances between acetogenesis and acetotrophy) were coincident with increased rates of methanogenesis in the refuse columns.
The time course profiles recorded with acetate were not repeated with propionate, butyrate, isobutyrate, isovalerate, valerate or caproate (Fig. 4) since, in general, the highest concentrations recorded were coincident with the highest rates of methanogenesis. Since propionate degradation in anoxic environments has been shown23 to be inhibited by high partial pressures of hydrogen it may be speculated that this mechanism was operative in the highly methanogenic samples. Similarly, since butyric acid bacteria generate hydrogen then butyrate production should have been influenced by the partial pressures of hydrogen in the samples.
After 92 days incubation the volatile fatty acid contents of the refuse columns were also determined (Table 3). Perhaps surprisingly, propionate accumulation characterised the 5 % and, more particularly, the 2.5 % sodium bicarbonate supplemented samples. Although methanogenic suppression could have accounted for the 5 :< treatment results it seems unlikely to have been the cause in the 2.5 % sample. Conversely, even though the existence of refuse microenvironments having high partial pressures of hydrogen could not be discounted, it is possible that the elevated propionate concentrations were due to increased dissolution of carbon dioxide, as a result of increased alkalinity which, in turn, could have inhibited propionate d e g r a d a t i ~ n . ~ ~
Following the protracted incubation period the final pH values of the 1 %, 2.5 %

286
Volatile fatty
(mp g dry WI refuse-() 18 acid cow3
G . B . Kasali, E . Senior, I . A . Watson-Craik
Fig. 4. Changes in concentrations of acetate (a), propionate (b), butyrate (c), isobutyrate (d), isovalerate (e). valerate (f) and caproate (g) in refuse samples during bottle culture cultivation at 30°C. Moisture contents were adjusted to field capacity by the addition ofglassdistilled water (0) or 1 a’, (a), 2.5 ”4 (0)
or 5 ”(, (m) (w/v) sodium bicarbonate solution.
and 5 % sodium bicarbonate and glassdistilled water-treated refuse were 8.0, 8.3, 7.9 and 7-2, respectively, which suggested that all the samples had generated additional alkalinity during the course of the fermentation.
4 CONCLUSION
Single additions of sodium bicarbonate solutions greatly improved methane production rates and concentrations in the solid-state refuse fermentation provided

TA
BL
E 3
T
erm
inal
Vol
atile
Fat
ty A
cid C
once
ntra
tions
Fol
low
ing I
ncub
atio
n at
30°
C fo
r 92
Day
s in
the
Pres
ence
ofN
aHC
O, o
r G
lass
-dis
tille
d W
ater
Add
ition
Add
ition
V
olat
ile
fatt
y ac
id (mg g
dry
weig
ht r
efis
e-')
Ace
tate
Pr
opio
nate
Is
oBut
yrat
e B
utyr
ate
IsoV
aler
ate
Val
erat
e C
apro
ate
~
NaH
CO
, (1
% w
/v)
NaH
CO
, (2
.5%
) N
aHC
O,
(5%
) G
lass
dist
illed
w
ater
(co
ntro
l)
~ ~
~ ~
~ ~
~ ~
7.89
3.
54
1.61
2.3
1
1.45
1 44
1.33
8.
49
1446
2.
13
2.24
2.
47
1.24
0.
67
23.4
0 12
.14
3.1
1 11
.88
2.77
2.
89
2:21
5.97
3.
21
1.43
2.
12
1.31
0.
86
0.81

288 G . B . Kasali, E . Senior, I . A . Watson-Craik
that the alkali concentrations maintained an acetic acid:alkalinity ratio < 08" and were below the cation toxicity threshold. However, although 2.5 % sodium bicarbonate was shown to promote the fermentation it is possible that the relatively high cost of the alkali might not be recovered in the value of the product. In this eventuality lower concentrations, even < 1 %, possibly in conjunction with the optimisation of a different site variable(s) such as refuse density and moisture content, pH or temperature might prove to be more commercially viable.
ACKNOWLEDGEMENTS
This work was supported by a studentship to G.B.K. from The National Council for Scientific Research, Zambia. Additional funding, which is gratefully acknowledged, was provided by Cleanaway Ltd.
REFERENCES
1. Rees, J. F., The fate of carbon compounds in the landfill disposal of organic matter. J . Chem. Tech. Biotechnol., 30 (1980) 161-75.
2. Buivid, M. G., Wise, D. L., Blanchet, M. J., Remedios, E. C., Jenkins, B. M., Boyd, W. F. & Pacey, J. G., Fuel gas enhancement by controlled landfilling of municipal solid waste. Resour. Conseru., 6 (1981) 3-20.
3. Pauss, A., Nyns, E.-J. & Naveau, H., Production of methane by anaerobic digestion of domestic refuse. In Anaerobic Digestion and Carbohydrate Hydrolysis of Waste, ed. G. L. Ferrero, M. P. Ferranti & H. Naveau. Elsevier Applied Science Publishers, London,
4. Leckie, 0. J., Pacey, J . G. & Halvadakis, C., Landfill management with moisture control. J . Enuiron. Eng. Diu. (ASCE), 105 (1979) 337-55.
5. DeWalle, F. B. & Chian, E. S. K., Energy recovery from landfilled solid waste. Biotechnol. Bioeng. Symp., 9 (1979) 317-28.
6. Klink, R. E. & Ham, R. K. , Effects of moisture movement on methane production in solid waste landfill samples. Resour. Recou. Conserv., 8 (1982) 2941.
7. Pfeffer, J . T., Temperature effects on anaerobic fermentation of domestic refuse. Biotechnol. Bioeng., 16 (1974) 771-87.
8. Rees, J . F., Optimisation of methane production and refuse decomposition in landfill by temperature control. J . Chem. Tech. Biotechnol., 30 (1980) 458-65.
9. Hartz, K. E., Klink, R. E. & Ham, R. K., Temperatureeffects: Methane generation from landfill samples. J . Enuiron. Eng. Diu. (ASCE), 108 (1982) 629-38.
10. Ehrig, H.-J., Quality and quantity of sanitary landfill leachate. Waste Manage. Res., 1
11. Mao, M. C. & Pohland, F. G., Continuing investigations on landfill stabilisation with leachate recirculation, neutralisation and sludge seeding. Special Research Report, Department of Civil Engineering, Georgia Institute of Technology, Atlanta, 1973.
12. Harmsen, J., Identification of organic compounds in leachate from a waste tip. Water Res., 17 (1983) 699-705.
13. Augenstein, D. C., Wise, D. L., Wentworth, R. L. & Cooney, C. L., Fuel gas recovery
1984, pp. 209-22.
(1983) 53-68.
from controlled landfilling of municipal wastes. Resour. Recou. Conseiu., 2 (1976) 103-17.
14. Klein, S. A., Anaerobic digestion of solid wastes. Compost Sci., 13(1) (1972) 6-11. 15. DeWalle, F. B., Chian, E. S. K. & Hammerberg, E., Gas production from solid waste in
landfills. J . Enuiron. Eng. Diu. (ASCE), 3 (1978) 415-31.

Refuse digestion: sodium bicarbonate promotion 289
16. Gossett, J . M., Stuckey, D. C., Owen, W. F. & McCarty, P. L., Heat treatment and anaerobic digestion of refuse. J . Enuiron. Eng. Diu. (ASCE), 108 (1982) 437-54.
17. Mosey, F. E., Swanwick, D. J . & Hughes, A. D., Factors affecting the availability of heavy metals to inhibit anaerobic digestion. Water Pollut. Control, 72 (1971) 668-78.
18. Kasali, G. B. & Senior, E., Temperature and moisture effects on the anaerobic digestion of refuse. J . Chem. Tech. Biotechnol., 44(1) (1988) 3 1 4 1 .
19. Hansson, G., End product inhibition in methane fermentations. Process Biochem., 17(6)
20. Ghosh, S., Solid-phase digestion of low moisture feeds. Biotechnol. Bioeng. Symp., 14
21. Burel, C. & Trancart, J . L., Mise au point d’un pilote de digestion anaerobie: conception et realisation. Trib. du Cebed., 38 (1985) 2 7 4 3 .
22. deBaere, L. A,, de Vocht, M., van Assche, P. & Verstraete, W., Influence of high NaCl and NH,CI salt levels in methanogenic associations. Water Res., 18 (1984) 543-8.
23. Boone, D. R., Mixed culture fermenter for simulating methanogenic digesters. A p p l . Enuiron. Microbiol., 48 (1984) 122-6.
24. Hansson, G. & Molin, N., End-product inhibition in methane fermentations: effects of carbon dioxide and methane on methanogenic bacteria utilising acetate. Eur. J . A p p l . Microbiol. Biotechnol., 13 (1981) 236-41.
(1982) 45-9.
(1984) 365-82.