serum catecholase: properties of the enzyme and
TRANSCRIPT

SERUM CATECHOLASE: PROPERTIES OF THE ENZYME AND NATURE OF AN ENDOGENOUS INHIBITOR*
BY BERNARD SHACTER
(From the Laboratory of Experimental Oncology, National Cancer Institute, National Institutes of Health, and the University of California
Medical School, San Francisco)
(Received for publication, August 29, 1949)
The occurrence of phenolases in plants has been known for many years (2). It has been only comparatively recently that such enzymes have been described in mammalian sources (3-5), including the histochemical demonstration of dopa oxidase activity in the leucocytes of blood (6). During an investigation of the relationship between enzymes and steroid hormones, an enzyme system was found in human blood serum which catalyzes the oxidation of catechol. Serum also contains an inhibitor, of variable concentration, which retards the oxidation of catechol during the early phases of the reaction. This report describes the properties of catecholase of human blood serum and the nature of the inhibitor. The activity of the enzyme and inhibitor in sera from various sources is de- scribed elsewhere (7).
EXPERIMENTAL
Methods
Fasting blood was drawn from the antecubital vein, and serum was separated after normal clotting had occurred. Catecholase activity of the serum was determined manometrically by measurement of oxygen uptake in the Warburg apparatus. Since the rate of catechol oxidation is markedly affected by even small changes in pH, the serum was first “neutralized” (8, 9) by removal of excess bicarbonate with hydrochloric acid. This was effected by adding serum and acid to the main chamber of Warburg vessels and alkali to the central well. The flasks, attached to lightly greased manometers, were then equilibrated in the Warburg bath at 37” for 1 hour. Most of the experiments were carried out at pH 7.4 with 1 ml. of serum. Under these conditions, generally 0.025 m.eq. of acid were re- quired for bicarbonate neutralization.
Following the neutralization equilibration, the flasks were removed from the bath, and the following solutions were added to the main chamber: 1 ml. of 0.5 M phosphate buffer; other materials studied for their effect on
* Presented in part at a meeting of the Federation of American Societies for Ex- perimental Biology at Detroit, April, 1949 (1).
697
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
698 SERUM CATECHOLASE
the reaction, adjusted to the pH of the buffer; and water to bring the volume to 2.5 ml. Controls contained water in place of serum. The sub- strate was dissolved in 0.2 M phosphate buffer, the solution was adjusted to the pH of the contents of the main compartment, and 0.5 ml. was added to the side arm. Substrate solutions were prepared immediately before use. The flasks with attached manometers were equilibrated in the bath for 10 minutes. The stop-cocks were then closed, the zero reading was taken, and the side arm contents were tipped in. Readings were taken every 10 minutes for several hours. The flasks were then removed from
MINUTES MINUTES
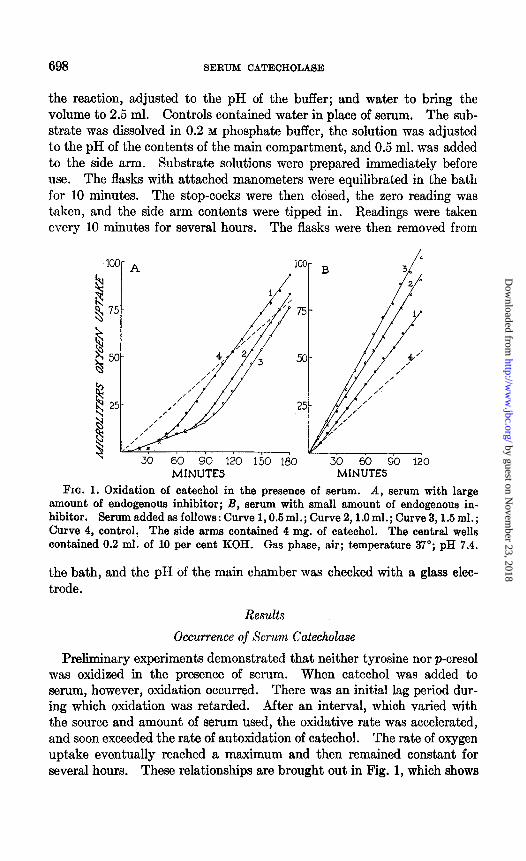
FIG. 1. Oxidation of catechol in the presence of serum. A, serum with large amount of endogenous inhibitor; B, serum with small amount of endogenous in- hibitor. Serumaddedasfollows:Curve1,0.5ml.;Curve2,1.0ml.;Curve3,1,5ml.; Curve 4, controll The side arms contained 4 mg. of catechol. The central wells contained 0.2 ml. of 10 per cent KOH. Gas phase, air; temperature 37”; pH 7.4.
the bath, and the pH of the main chamber was checked with a glass elec- trode.
Results
Occurrence of Serum Catecholuse
Preliminary experiments demonstrated that neither tyrosine nor p-cresol was oxidized in the presence of serum. When catechol was added to serum, however, oxidation occurred. There was an initial lag period dur- ing which oxidation was retarded. After an interval, which varied with the source and amount of serum used, the oxidative rate was accelerated, and soon exceeded the rate of autoxidation of catechol. The rate of oxygen uptake eventually reached a maximum and then remained constant for several hours. These relationships are brought out in Fig. 1, which shows
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
B. SHACTER 699
typical experiments on blood sera containing large and small amounts of inhibitor.
The factor in serum which catalyzes the oxidation of catechol is probably enzymatic in nature since, as described below, it satisfies several criteria for enzymatic activity, such as inactivation by heat and specificity. Of several mono- and diphenols tried as substrates, only catechol oxidation was accelerated in the presence of serum. The factor will therefore be referred to as serum catecholase. A subsequent section of this investiga-
30 60 90 120 150 180 2 4 6 8 MINUTES MG. CATECHOL
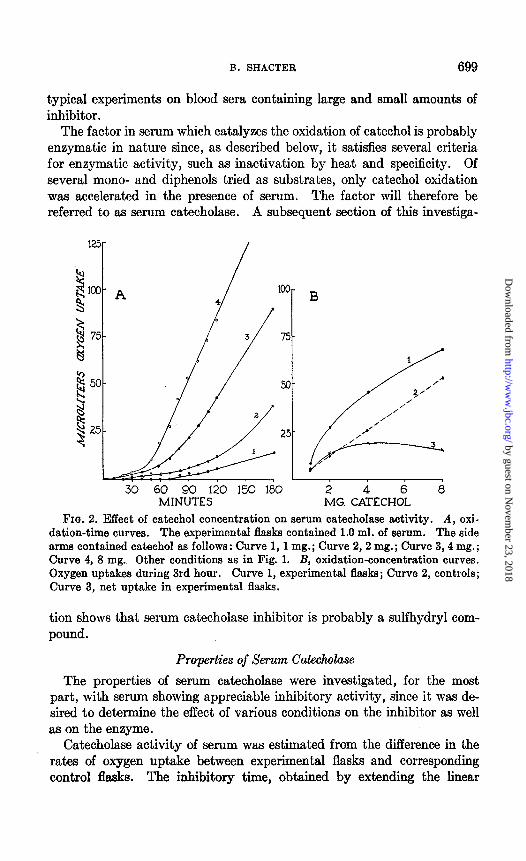
FIG. 2. Effect of catechol concentration on serum catecholase activity. A, oxi- dation-time curves. The experimental flasks contained 1.0 ml. of serum. The side arms contained catechol as follows: Curve 1,1 mg.; Curve 2,2 mg.; Curve 3,4 mg.; Curve 4, 8 mg. Other conditions as in Fig. 1. B, oxidation-concentration curves. Oxygen uptakes during 3rd hour. Curve 1, experimental flasks; Curve 2, controls; Curve 3, net uptake in experimental flasks.
tion shows that serum catecholase inhibitor is probably a sulfhydryl com- pound.
Properties of Serum Catecholase
The properties of serum catecholase were investigated, for the most part, with serum showing appreciable inhibitory activity, since it was de- sired to determine the effect of various conditions on the inhibitor as well as on the enzyme.
Catecholase activity of serum was estimated from the difference in the rates of oxygen uptake between experimental flasks and corresponding control flasks. The inhibitory time, obtained by extending the linear
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
700 SERUM CATECHOLASE
portion of the oxygen uptake curve to the time axis, was used as a measure of inhibitor activity.
Effect of Substrate Concentration-As the concentration of catechol added to serum was increased from 1 to 8 mg. per flask (Fig. 2), the inhibitory time decreased, while the rate of oxidation following the end of the lag period was augmented. Autoxidation of catechol proceeded in a linear fashion with time for all concentrations used, and the rate of oxygen up- take was proportional to the catechol concentration. Catecholase activity
30 60 90 120 150 180 30 60 90 120 150 180 MINUTES MINUTES
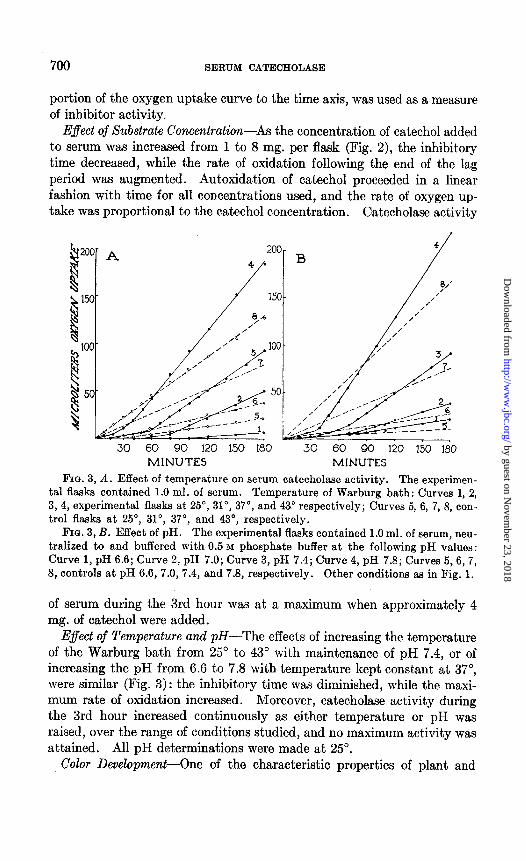
FIG. 3, A. Effect of temperature on serum catecholase activity. The experimen- tal flasks contained 1.0 ml. of serum. Temperature of Warburg bath: Curves 1, 2, 3,4, experimental flasks at 29, 31”, 37”, and 43” respectively; Curves 5, 6, 7, 8, con- trol flasks at 25”, 31”, 37”, and 43”, respectively.
FIG. 3, B. Effect of pH. The experimental flasks contained 1.0 ml. of serum, neu- tralized to and buffered with 0.5 M phosphate buffer at the following pH values: Curve 1, pH 6.6; Curve 2, pH 7.0; Curve 3, pH 7.4; Curve 4, pH 7.8; Curves 5,6,7, 8, controls at pH 6.6, 7.0, 7.4, and 7.8, respectively. Other conditions aa in Fig. 1.
of serum during the 3rd hour was at a maximum when approximately 4 mg. of catechol were added.
E$ect of Temperature and pH-The effects of increasing the temperature of the Warburg bath from 25’ to 43” with maintenance of pH 7.4, or of increasing the pH from 6.6 to 7.8 with temperature kept constant at 37”, were similar (Fig. 3): the inhibitory time was diminished, while the maxi- mum rate of oxidation increased. Moreover, catecholase activity during the 3rd hour increased continuously as either temperature or pH was raised, over the range of conditions studied, and no maximum activity was attained. All pH determinations were made at 25”.
Color Development-One of the characteristic properties of plant and
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
B. SHACTER 701
animal phenoloxidases is the transformation of the substrate to a colored. oxidation product (10). Serum catecholase also showed this property. No color developed during the lag period following addition of catechol to serum; however, as soon as the rate of oxygen uptake became accelerated, a pink color appeared which turned brown in a short time. The depth of color developed in 3 hours was more intense as the serum concentration, substrate concentration, temperature, or pH was raised, over the whole range of conditions observed. No appreciable color developed in control flasks.
MINUTE.5 MINUTES FIG. 4. Effect of heat on serum catecholase activity. A, the experimental flasks
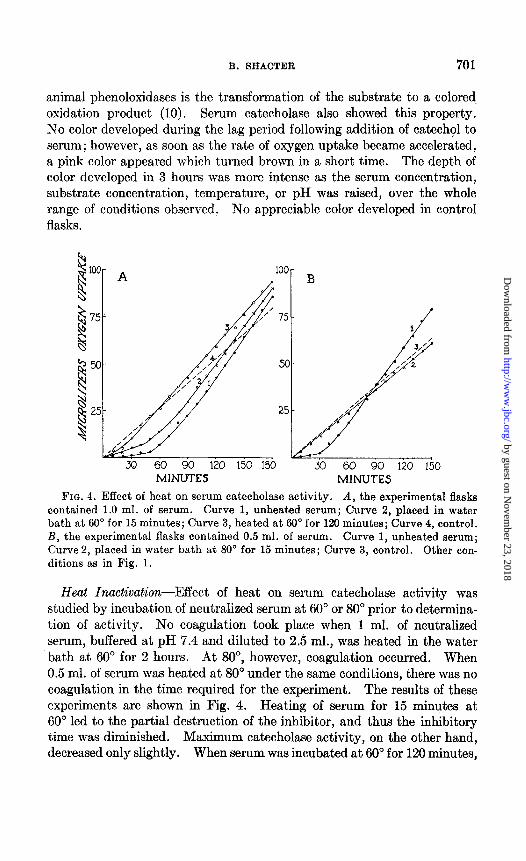
contained 1.0 ml. of serum. Curve 1, unheated serum; Curve 2, placed in water bath at 60” for 15 minutes; Curve 3, heated at 60” for 120 minutes; Curve 4, control. B, the experimental flasks contained 0.5 ml. of serum. Curve 1, unheated serum; Curve 2, placed in water bath at 80” for 15 minutes; Curve 3, control. Other con- ditions as in Fig. 1.
Heat Inactivation-Effect of heat on serum catecholase activity was studied by incubation of neutralized serum at 60” or 80” prior to determina- tion of activity. No coagulation took place when 1 ml. of neutralized serum, buffered at pH 7.4 and diluted to 2.5 ml., was heated in the water bath at 60” for 2 hours. At SO”, however, coagulation occurred. When 0.5 ml. of serum was heated at 80” under the same conditions, there was no coagulation in the time required for the experiment. The results of these experiments are shown in Fig. 4. Heating of serum for 15 minutes at 60” led to the partial destruction of the inhibitor, and thus the inhibitory time was diminished. Maximum catecholase activity, on the other hand, decreased only slightly. When serum was incubated at 60” for 120 minutes,
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
702 SERUM CATECHOLASE
the lag period was only transitory, while enzymatic activity, although diminished, was still present. Incubation of serum for 30 and 60 minutes at 60” produced intermediate effects on the inhibitor and on catecholase activity.
Inhibitor and enzyme were both completely inactivated when serum was maintained at a temperature of 80” for 15 minutes. Even under these conditions, however, when no enzymatic catalysis of catechol oxidation was
25
30 60 90 120 150 180 30 60 90 120 150 180 MINUTES MINUTES
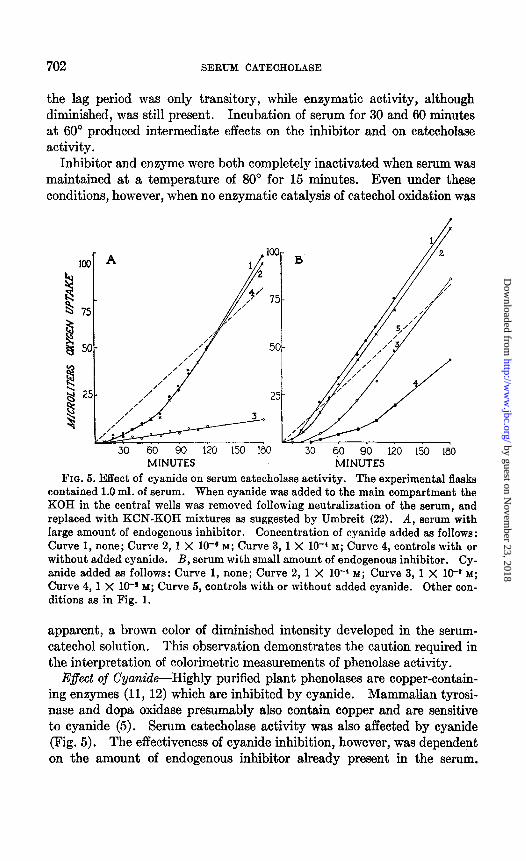
FIG. 5. Effect of cyanide on serum catecholase activity. The experimental flasks contained 1.0 ml. of serum. When cyanide w&s added to the main compartment the KOH in the central wells was removed following neutralization of the serum, and replaced with KCN-KOH mixtures as suggested by Umbreit (22). A, serum with large amount of endogenous inhibitor. Concentration of cyanide added as follows : Curve 1, none; Curve 2, 1 X 10-6 M; Curve 3,1 X lO+ M; Curve 4, controls with or without added cyanide. B, serum with small amount of endogenous inhibitor. Cy- anide added as follows: Curve 1, none; Curve 2, 1 X 10-’ M; Curve 3, 1 X 10-a M;
Curve 4, 1 X 10-* M; Curve 5, controls with or without added cyanide. Other con- ditions as in Fig. 1.
apparent, a brown color of diminished intensity developed in the serum- catechol solution. This observation demonstrates the caution required in the interpretation of calorimetric measurements of phenolase activity.
E$ect of Cyanide-Highly purified plant phenolases are copper-contain- ing enzymes (11, 12) which are inhibited by cyanide. Mammalian tyrosi- nase and dopa oxidase presumably also contain copper and are sensitive to cyanide (5). Serum catecholase activity was also affected by cyanide (Fig. 5). The effectiveness of cyanide inhibition, however, was dependent on the amount of endogenous inhibitor already present in the serum.
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
B. SHACTER 703
With serum which normally had a long lag period, a concentration of 1 X lo-* M cyanide extended the inhibitory time for the duration of the experiment. The same concentration of cyanide, on the other hand, in- creased the inhibitory time only slightly with serum which had a small amount of endogenous inhibitor, while maximum catecholase activity was unaffected. With a concentration of 1 X 10-a M cyanide, the inhibitory time was increased further, although ultimate enzymatic activity remained unchanged. When the cyanide concentration was increased to 0.01 M,
30 60 90 120 150 180 MINUTES
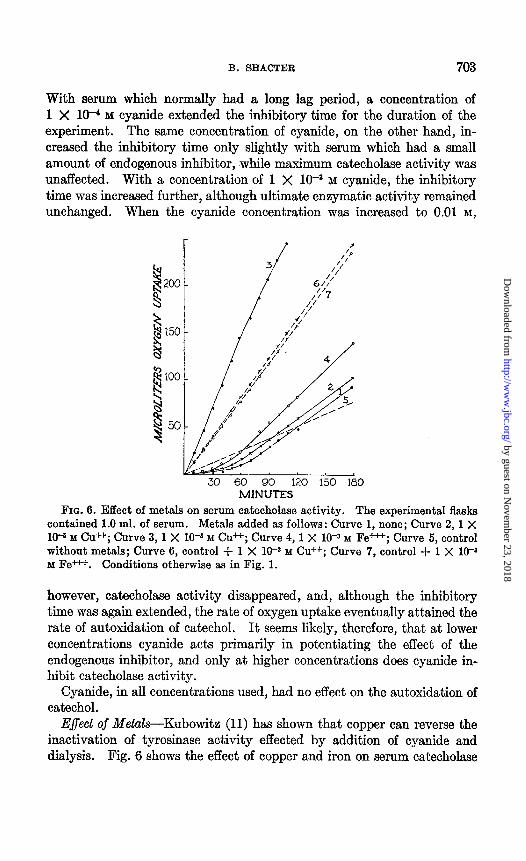
FIG. 6. Effect of metals on serum catecholase activity. The experimental flasks contained 1.0 ml. of serum. Metals added as follows: Curve 1, none; Curve 2, 1 X lo+ M cd+; Curve 3, 1 X 10T3 M cu *; Curve 4,1 X 10-a M Fe+++; Curve 5, control without metals; Curve 6, control + 1 X W3 M Cu ++; Curve 7, control + 1 X 10-* M Fe+++. Conditions otherwise as in Fig. 1.
however, catecholase activity disappeared, and, although the inhibitory time was again extended, the rate of oxygen uptake eventually attained the rate of autoxidation of catechol. It seems likely, therefore, that at lower concentrations cyanide acts primarily in potentiating the effect of the endogenous inhibitor, and only at higher concentrations does -cyanide in- hibit catecholase activity.
Cyanide, in all concentrations used, had no effect on the autoxidation of catechol.
E$ect of Metals-Kubowitz (11) has shown that copper can reverse the inactivation of tyrosinase activity effected by addition of cyanide and dialysis. Fig. 6 shows the effect of copper and iron on serum catecholase
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
704 SERUM CATECHOLASE
activity. Copper at 1 X 10V3 M concentration markedly accelerated the enzymatic and non-enzymatic oxidation of catechol; moreover, the in- hibitory time with serum was completely abolished. When the copper concentration was decreased to 1 X 10e5 M, maximum catecholase activity remained practically unchanged, while the inhibitory time was somewhat shortened.
The effect of iron on serum catecholase activity was in sharp contrast to that of copper. Although iron at 1 X lop3 M concentration accelerated catechol oxidation in the iron-catechol system as much as an equimolar
2OO- A
s EO- B
h E IOO- 3
F k 8 a 2
30 60 90 120 150 180 30 60 90 120 150 180 MINUTES MINUTES
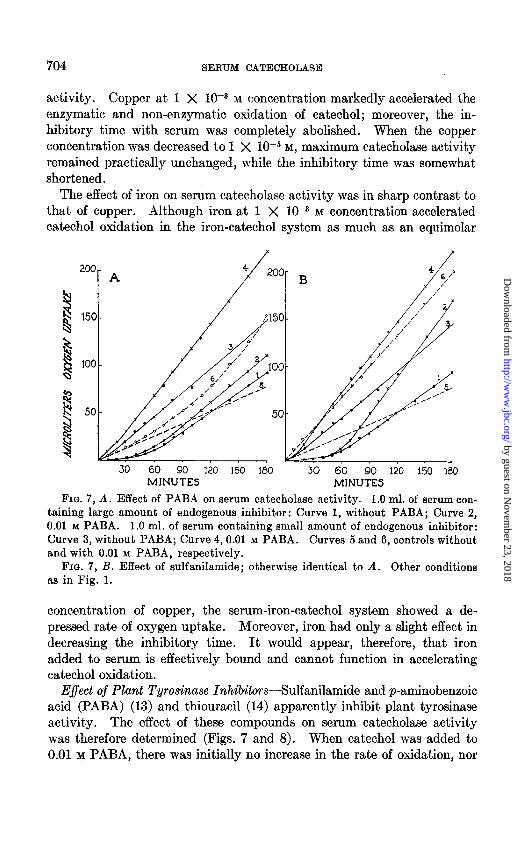
FIG. 7, A. Effect of PABA on serum catecholase activity. 1.0 ml. of serum con- taining large amount of endogenous inhibitor: Curve 1, without PABA; Curve 2, 0.01 M PABA. 1.0 ml. of serum containing small amount of endogenous inhibitor: Curve 3, without PABA; Curve 4,O.Ol M PABA. Curves 5 and 6, controls without and with 0.01 M PABA, respectively.
FIG. 7, B. Effect of sulfanilamide; otherwise identical to A. Other conditions as in Fig. 1.
concentration of copper, the serum-iron-catechol system showed a de- pressed rate of oxygen uptake. Moreover, iron had only a slight effect in decreasing the inhibitory time. It would appear, therefore, that iron added to serum is effectively bound and cannot function in accelerating catechol oxidation.
E$ect of Plant Tyrosinase InhibitorsSulfanilamide and p-aminobenzoic acid (PABA) (13) and thiouracil (14) apparently inhibit plant tyrosinase activity. The effect of these compounds on serum catecholase activity was therefore determined (Figs. 7 and 8). When catechol was added to 0.01 M PABA, there was initially no increase in the rate of oxidation, nor
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
B. SHACTER 705
was there, any immediate color development. After approximately 30 minutes, however, the rate of oxygen uptake became accelerated and a reddish color appeared. At the end of the 3rd hour the effect of PABA on catechol oxidation was very marked and a strong brown color had de- veloped. The same concentration of PABA, added to serum containing appreciable endogenous inhibitor, had no effect on the inhibitory time, while the accelerative phase of catechol oxidation was retarded. When added to serum which had a short lag period, PABA inhibited the ap- parent enzymatic activity. Thus catecholase activity during the 3rd
MINUTES
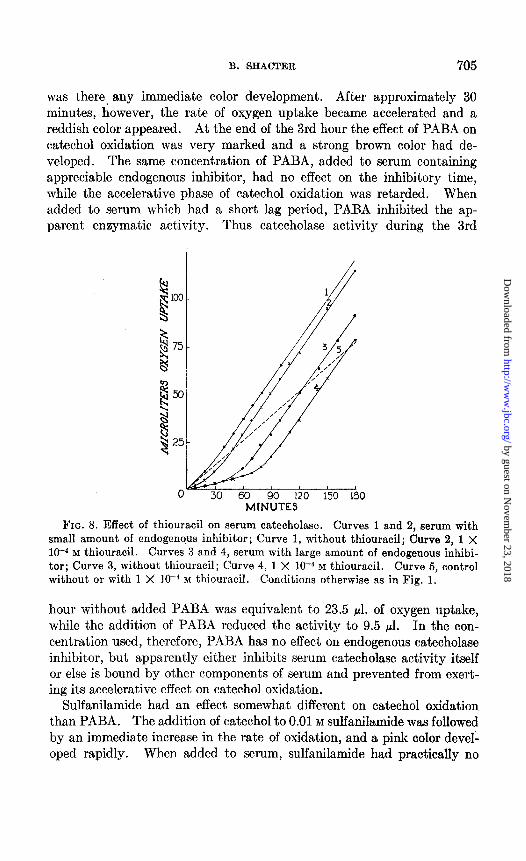
FIG. 8. Effect of thiouracil on serum catecholase. Curves 1 and 2, serum with small amount of endogenous inhibitor; Curve 1, without thiouracil; Curve 2, 1 X 10-4 M thiouracil. Curves 3 and 4, serum with large amount of endogenous inhibi- tor; Curve 3, without thiouracil; Curve 4, 1 X 10V4 M thiouracil. Curve 5, control without or with 1 X lo-* M thiouracil. Conditions otherwise as in Fig. 1.
hour without added PABA was equivalent to 23.5 ~1. of oxygen uptake, while the addition of PABA reduced the activity to 9.5 pl. In the con- centration used, therefore, PABA has no effect on endogenous catecholase inhibitor, but apparently either inhibits serum catecholase activity itself or else is bound by other components of serum and prevented from exert- ing its accelerative effect on catechol oxidation.
Sulfanilamide had an effect somewhat different on catechol oxidation than PABA. The addition of catechol to 0.01 M sulfanilamide was followed by an immediate increase in the rate of oxidation, and a pink color devel- oped rapidly. When added to serum, sulfanilamide had practically no
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
706 SERUM CATECHOLASE
effect on the inhibitory time, while apparent maximum catecholase activity was decreased.
Fig. 8 shows that the only effect of 1 X HP4 M thiouracil was to increase the inhibitory time. There waa no inhibition of maximum enzymatic activity. Thiouracil at a concentration of 1 X 10m4 M had no effect on the autoxidation of catechol. When the thiouracil concentration was in- creased to 1 X 10m3 M, however, both the enzymatic oxidation and the autoxidation of catechol were inhibited.
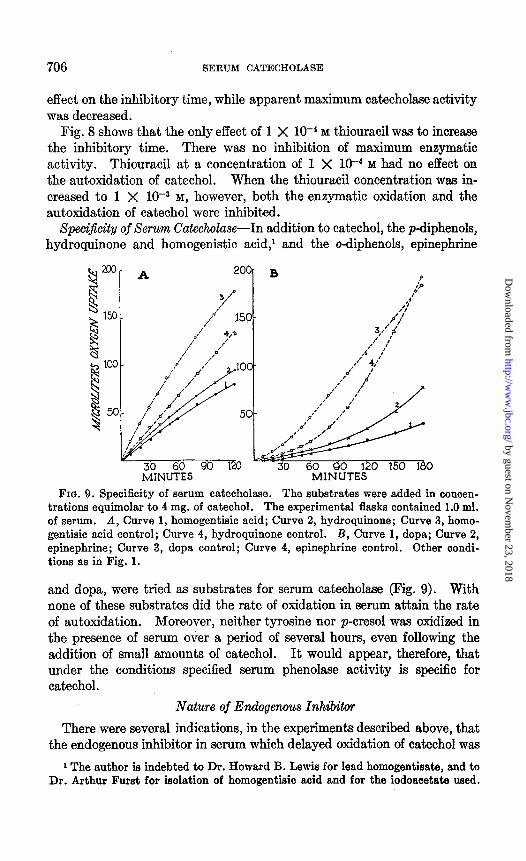
SpeciJicity of Serum Cakcholase-In addition to catechol, the p-diphenols, hydroquinone and homogenistic acid,’ and the o-diphenols, epinephrine
MINUTES FIQ. 9. Specificity of serum catecholase. The substrates were added in concen-
trations equimolar to 4 mg. of catechol. The experimental flasks contained 1.0 ml. of serum. A, Curve 1, homogentisic acid; Curve 2, hydroquinone; Curve 3, homo- gentisic acid control; Curve 4, hydroquinone control. B, Curve 1, dopa; Curve 2, epinephrine; Curve 3, dopa control; Curve 4, epinephrine control. Other condi- tions as in Fig. 1.
and dopa, were tried as substrates for Serum catecholaae (Fig. 9). With none of these substrates did the rate of oxidation in serum attain the rate of autoxidation. Moreover, neither tyroaine nor p-cresol was oxidized in the presence of serum over a period of several hours, even following the addition of small amounts of catechol. It would appear, therefore, that under the conditions specified serum phenolaae activity is specific for catechol.
Nature of Endogenous Inhibitor
There were several indications, in the experiments described above, that the endogenous inhibitor in serum which delayed oxidation of catechol was
1 The author is indebted to Dr. Howard B. Lewis for lead homogentisate, and to Dr. Arthur Fur& for isolation of homogentisic acid and for the iodoacetate wed.
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from
B. SHACTER 707
a sulfhydryl compound. Copper, for example, catalyzes the oxidation of reduced glutathione, while iron is ineffective in this regard (15, 16). More- over, cyanide inhibits almost completely the oxidation of sulfhydryl groups by various oxidizing agents (17). Figge (18) has shown that glutathione inhibits melanin formation in tyrosine-tyrosinase mixtures; and more re- cently it was found (19, 20) that human skin contains a dialyzable sulfhy- dry1 compound which inhibited the autoxidation of dopa and the catalytic oxidation of tyrosine by plant tyrosinase. The inhibition could be elimi- nated by the addition of copper or of the sulfhydryl reagents, iodoaceta- mide and p-chloromercuribenzoic acid. The effect of several sulfhydryl reagents on the endogenous inhibitor in serum was therefore determined.
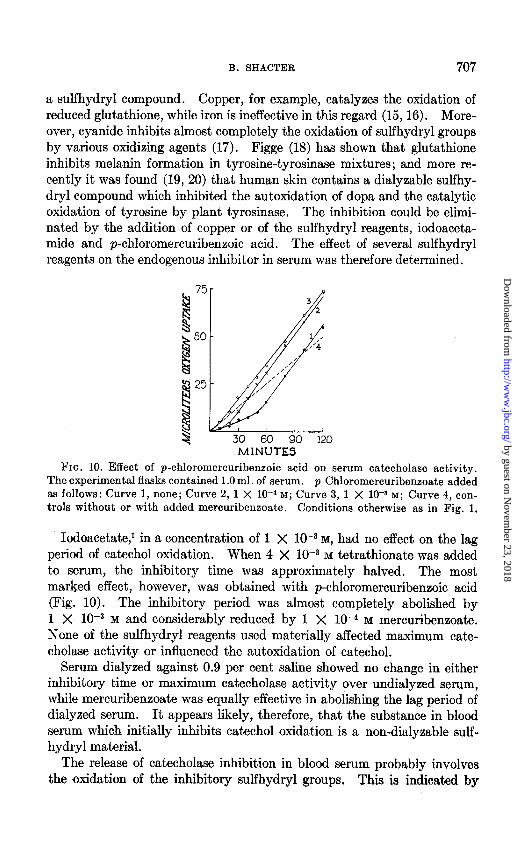
30 60 90 120 MINUTES
FIG. 10. Effect of p-chloromercuribeneoic acid on serum catecholase activity. The experimental flasks contained 1 .O ml. of serum. p-Chloromercuribenzoate added as follows: Curve 1, none; Curve 2, 1 X lc4 M; Curve 3, 1 X 10-s M; Curve 4, con- trols without or with added mercuribenzoate. Conditions otherwise as in Fig. 1.
Iodoacetate,’ in a concentration of 1 X lOWa M, had no effect on the lag period of catechol oxidation. When 4 X low3 M tetrathionate was added to serum, the inhibitory time was approximately halved. The most marked effect, however, was obtained with p-chloromercuribenzoic acid (Fig. 10). The inhibitory period was almost completely abolished by 1 X lo-+ M and considerably reduced by 1 X 1O-4 M mercuribenzoate. None of the sulfhydryl reagents used materially affected maximum cate- cholase activity or influenced the autoxidation of catechol.
Serum dialyzed against 0.9 per cent saline showed no change in either inhibitory time or maximum catecholase activity over undialyzed serum, while mercuribenzoate was equally effective in abolishing the lag period of dialyzed serum. It appears likely, therefore, that the substance in blood serum which initially inhibits catechol oxidation is a non-dialyzable sulf- hydryl material.
The release of catecholase inhibition in blood serum probably involves the ,oxidation of the inhibitory sulfhydryl groups. This is indicated by
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from

708 SERUM CATECHOLASE
the accelerative effect of copper on the release of inhibition and on the prolongation of inhibition with cyanide. Catechol is apparently necessary for the sulfhydryl oxidation, since neither keeping the serum refrigerated for several days before determination of catecholase activity nor increasing the time of the neutralization equilibration materially decreases the inhibi- tory time. The mechanism of oxidation possibly involves the initial oxi- dation of catechol to o-benzoquinone, which then oxidizes the sulfhydryl groups and is in turn reduced to catechol again, until all of the sulfhydryl groups are oxidized. Miller and Dawson (21) have utilized a similar re- action in their chronometric method for measurement of catecholase activ- ity, using ascorbic acid as the reducing agent.
SUMMARY
1. Human blood serum contains an enzyme which accelerates the oxida- tion of catechol, but not tyrosine, p-cresol, hydroquinone, homogentisic acid, epinephrine, or dopa; t,he system is referred to as serum catecholase.
2. Serum also contains an inhibitor, of variable concentration, which initially retards the oxidation of cathechol. The inhibitor has the charac- teristics of a non-dialyzable, sulfhydryl material.
3. Serum catecholase activity is accelerated and the inhibitory time decreased by increases in temperature from 25” to 43” and by increases in pH from 6.6 to 7.8.
4. Heat of 80” inactivates both the enzyme and the inhibitor. 5. Cyanide inhibits catecholase activity and prolongs the effect of the
endogenous inhibitor. 6. Copper abolishes the lag period in the oxidation of catechol by serum
and enhances catecholase activity. 7. Sulfanilamide and p-aminobenzoic acid have no effect on the en-
dogenous inhibitor, but themselves inhibit apparent catecholase activity. Thiouracil prolongs the inhibitory time but has no effect on maximum en- zymatic activity.
BIBLIOGRAPHY
1. Shatter, B., Federation Proc., 8, 144 (1949). 2. Bourquelot, E., and Bertrand, G., Compt. rend. Sot. biol., 47, 582 (1895). 3. Bloch, B., Am. J. Med. SC., 1’77, 609 (1929). 4. Cadden, J. F., and Dill, L. V., J. Biol. Chem., 143, 105 (1942). 5. Hogeboom, G. H., and Adams, M. H., J. Biol. Chem., 146, 273 (1942). 6. Peck, S. M., and Sobotka, H., J. Invest. Dermat., 4, 325 (1941). 7. Shatter, B., and Shimkin, M. B., J. Nat. Cancer Inst., 10,637 (1949). 8. MacLeod, J. M., and Rhoads, C. P., Proc. Sot. Exp. Biol. and Med., 41,268 (1939). 9. Friend, D. G., and Hastings, A. B., Proc. Sot. Exp. Biol. and Med., 46,137 (1940).
10. Nelson, J. M., and Dawson, C. R., Advances in EnzymoZ., 4, 99 (1944).
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from

B. SHACTER 709
11. Kubowitz, F., Rio&em. Z., 299, 32 (1938). 12. Keilin, D., and Mann, T., Proc. Roy. Sac. London, Series B, 126, 187 (1938). 13. Martin, G. J., Wisansky, W. A., and Ansbacher, S., Proc. Sot. Exp. Biol. and
Med., 47, 26 (1941). 14. Paschkis, K. E., Cantarow, A., Hart, W. M., and Rakoff, A. E., Proc. Sot. Exp.
Biol. and Med., 67, 37 (1944). 15. Voegtlin, C., Johnson, J. M., and Rosenthal, S. M., Pub. Health Rep., U. S. P.
H. S., 46, 2234 (1931). 16. Bernheim, F., and Bernheim, M. L. C., Cold Spring Harbor Symposia Quant.
Biol., 7, 174 (1939). 17. Anson, M. L., J. Gen. Physiol., 24, 399 (1940-41). 18. Figge, F. H. J., Proc. Sot. Exp. Biol. and Med., 46, 269 (1941). 19. Rothman, S., Krysa, H. F., and Smiljanic, A. M., Proc. Sot. Exp. Biol. and Med.,
62, 208 (1946). 29. Flesch, P., Proc. Sot. Exp. Biol. and Med., 70, 136 (1949). 21. Miller, W. H., and Dawson, C. R., J. Am. Chem. Sot., 63, 3375 (1941). 22. Umbreit, W. W., in Umbreit, W. W., Burris, R. H., and Stauffer, J. F., Mano-
metric techniques and related methods for the study of tissue metabolism, Minneapolis, 45 (1945).
by guest on Novem
ber 23, 2018http://w
ww
.jbc.org/D
ownloaded from

Bernard ShacterENDOGENOUS INHIBITOR
OF THE ENZYME AND NATURE OF AN SERUM CATECHOLASE: PROPERTIES
1950, 184:697-710.J. Biol. Chem.
http://www.jbc.org/content/184/2/697.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/184/2/697.citation.full.haccessed free atThis article cites 0 references, 0 of which can be by guest on N
ovember 23, 2018
http://ww
w.jbc.org/
Dow
nloaded from