retikulum endoplasma.pdf
TRANSCRIPT

RETIKULUM ENDOPLASMA
Clara Khoirunnisa
Dian Putri Permatasari
Shyfa F. Ruyani
Yussa Annisa Syabilla

PENDAHULUAN
Retikulum endoplasma (RE) merupakan organel selyang tidak statis dan dapatdianggap sebagai salah satukomponen dari suatu sistem dinamik yang lebih besaryaitusistem membran di dalam sel yang terdiri atas berbagaiorganel. Jaringan sistemmembran ini disebut jaringancytocavitary yang di dalamnya termasuk mitokondria,lisosom, badan golgi, badan mikro, dan membran inti.
Bersama-sama mereka membentuk sebuah jaringandidalam sel, memisahkannya menjadi dua kompartemenyaitu kompartemen sitoplasmik dan kompartemenintracavitary. Jadi, membran yang membangun sistem itudisatu sisi permukaan menghadap sitoplasma ekstra organel(sitosol) dan permukaan lain menghadap lumen dari sistemmembran.

• Umumnya semua organel dari jaringan cytocavitarysecara terus menerus melakukan komunikasi danpertukaran diantara pasangan tertentu atau antartetangga, sehingga terlihat melakukan sebuahinteraksi baik secara langsung maupun tidaklangsung.
• Struktur dan fungsi dari retikulum endoplasmabergantung dan tergantung pada organel darijaringan cavitary.
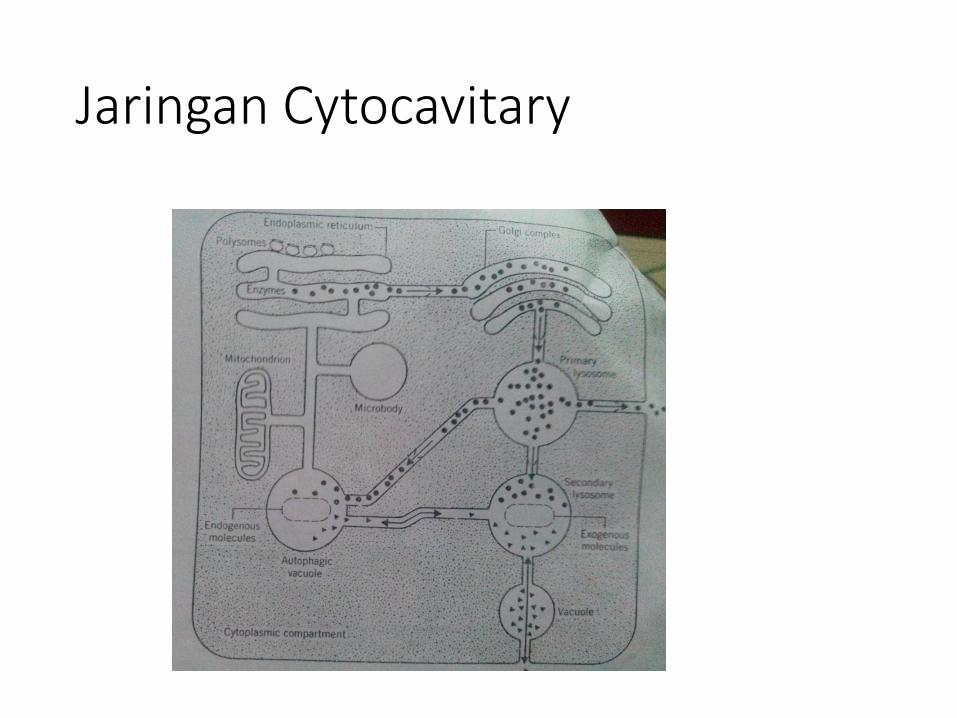
Jaringan Cytocavitary

6.1 SIFAT UMUM DARI RETIKULUM ENDOPLASMA• Tanpa adanya sistem membran yang dinamis,
bagian interior dari sel eukariotik akan sulitdijangkau.
• Substrat-substrat yang tidak mampu didifusikandengan cepat sesuai yang dibutuhkan dan produk-produk buangan akan terakumulasi dengan sia-siamenjadi racun. Hal tersebut bukan merupakanmasalah yg besar bagi sel prokariotik karenapendeknya jarak metabolisme yang terjadi.
• Tetapi jarak metabolisme pada sel eukariotik yangmenjadi masalah besar.

• Jaringan cavitary khususnya retikulum endoplasma,mencegah stagnasi pembentukan sistem sirkulasiinterseluler dan secara luas penyebaran enzim padaaktivitas katabolisme dan anabolisme.
• Substrat-substrat vital bisa menjangkau interior darisel dengan cepat oleh adanya fusi membran danpergerakan, dan material-material yang disintesisdan dirakit di dalam sel dapat dengan cepat ditransportasikan ke permukaan.

RETIKULUM ENDOPLASMA SECARA MIKROSKOPI• Secara mikroskopi, pada sel hati, terdapat dua
bagian besar dibentuk dari membran-membranmembentuk retikulum endoplasma.
• Membran-membran yang memiliki rupa yang kasar,karena diselubungi partikel ribosom, disebutRetikulum Endoplasma Kasar (RER).
• Membran-membran yang tidak memiliki partikelribosom dikenal sebagai Retikulum EndoplasmaHalus (SER).

RETIKULUM ENDOPLASMA KASAR DAN HALUS• RER adalah tempat dari sintesis protein di dalam sel,
khususnya, sintesis dari protein sekresi dan protein dari retikulum endoplasma itu sendiri.
• SER memperantarai berbagai reaksi sintesis lainnya baik seperti modifikasi kimia dari material-material molekular.
• Kehadiran dari keduanya RER dan SER pada sel hatiberhubungan dengan peran hati dalam metabolisme.
• Secara virtual semua reaksi dari metabolismeintermediary mengambil tempat di liver, dan beberapareaksi terbagi diantara dua tipe dari membran.

• Rasio REK dan REH pada setiap sel berbeda-beda tergantung pada fisiologis dari sel yang bersangkutan.
• Sel-sel yang lebih sering dilibatkan dalam sintesis protein sekresi , seperti pancreatic acinar atau sel plasma, memiliki lebih banyak RER. Tugas utama RER adalah untuk merakit dan mengekspor protein.
• Sel-sel yang terkonsentrasi pada produksi steroid,seperti pembuatan jaringan dari adernal bagian kortex,mengandung retikulum endoplasma yang didominasibagian halus. Sintesis kolesterol juga terjadi di REhalus tersebut seperti halnya beberapa reaksi darimodifikasi steroid membentuk progesteron dandeoxycorticosterone.

LAMELLAR, VESIKULAR, DAN TUBULAR
• Selain adanya RER dan SER , retikulum endoplasma biasanya juga hadir dalam tiga bentuk fisik yang berbeda di dalam sel.
Lamellar
• RER paling sering terlihat dalam bentuk lamellar. Bentuk ini tidak semata-semata timbunan dari lembaran-lembaran membran, tapi juga kumpulan dari kantung membran yang memipih.
• Salah satu sifat dari bentuk lamellar adalah adanya ribosom yang tidak merata (asimetri) pada membran ini yang mana terdapat sebuah ruang yang disebut Sisterna.

Vesikular
• Bentuk kantung (vesikular), kebanyakan terdapat dalam retikulum endoplasma halus.
• Penelitian secara in vivo menunjukan membran memiliki kecendrungan alami membentuk vesikel-vesikel. Secara in vitro menunjukan wujud membran tersebut sebagai sebuah vesikel di dalam sel.
Tubullar
• Bentuk tubular (pembuluh). Bentuk ini terutama dimiliki oleh retikulum endoplasma halus, menunjukkan sifat yang dinamik dari RE dan mempunyai hubungan erat dengan gerakan membran, pemisahan dan fusi dalam sistem membran (jaringan cytocavitary)

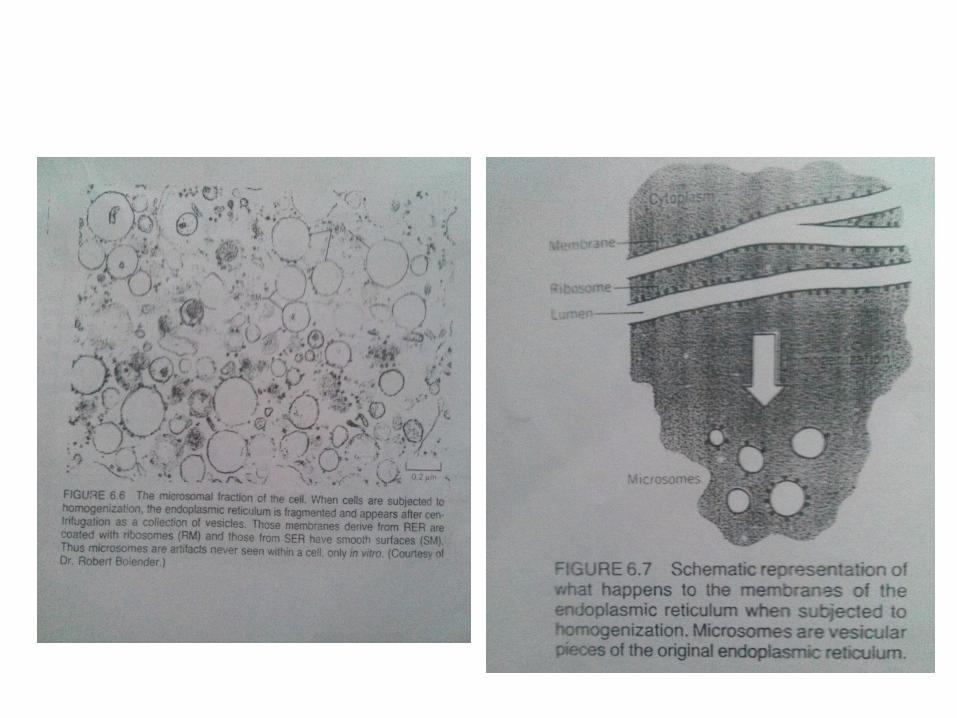
ISOLASI : FRAKSI MICROSOMAL
• Membran dari retikulum endoplasma dapatdiisolasikan dengan teknik homogenized jaringan kedalam sentrifugasi yang berbeda. Membran tersebutberputar ke bawah seperti sebuah peluru setelahmitokondria dihilangkan.
• Mikroskop elektron menunjukan bahwa membran-membran tersebut membentuk vesikel-vesikeltersebut dari salah satu antara bagian yang halusmaupun kasar dari membran. ( Gambar 6.6)Membran yang berupa vesikel tersebut adalahmiscrosomes.

• Retikulum endoplasma terdisrupsi dari bentuk asalnya sebagai sebuah kumpulan dari vesikel membran.
• Vesikel-vesikel yang dihasilkan merupakan turunan dari RER ditunjukan dengan adanya ribosom yang melekat pada vesikel dibagian permukaan luar.
• Dapat disimpulkan semua vesikel miscrosomal memiliki orientasi luar dan dalam yang sama seperti yang ada pada permukaan luar permukaan sitoplasma didalam sel yang utuh. (Gambar 6.7)
• Dapat dijelaskan bahwa microsomes bukan benar-benar sebagai organel dari sel , pada kenyataanya, merupakan hasil isolasi.
• Istilah microsomes digunakan ketika mengacu pada retikulum endoplasma yang terisolasi dan istilah retikulum endoplasma untuk struktur membran sebagai mana yang ada pada sel yang utuh.
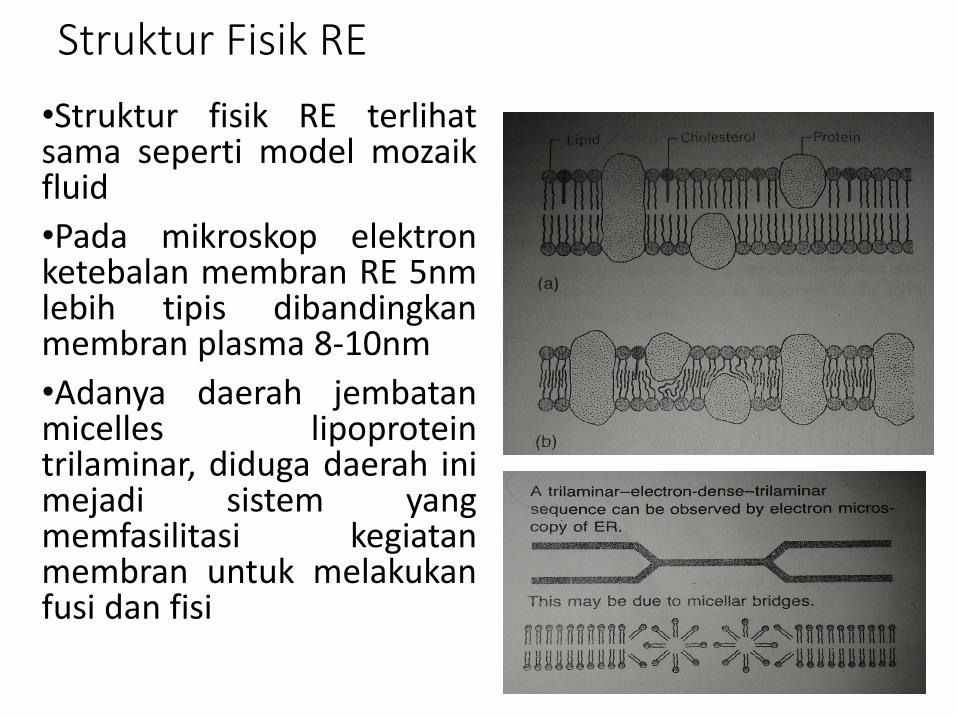
Struktur Fisik RE
•Struktur fisik RE terlihatsama seperti model mozaikfluid
•Pada mikroskop elektronketebalan membran RE 5nmlebih tipis dibandingkanmembran plasma 8-10nm
•Adanya daerah jembatanmicelles lipoproteintrilaminar, diduga daerah inimejadi sistem yangmemfasilitasi kegiatanmembran untuk melakukanfusi dan fisi

Struktur KimiaPada hati tikus, membran dari microsomenya
memiliki berat yang 60%-70% mengandung proteindan 30%-40% sisanya mengadung phospholipid.Setidaknya terdapat 33 polypeptides yang memilikistruktur fisika dan kimia yang berbeda sertapembagian dari microsomal phospholipid sebagaiberikut :
•phosphatidylcholine (55%)
•phosphatidylethanolamine (20-25%)
•phosphatidylserine (5-10%)
•phosphatidylinositol (5-10%)
•sphingomyelin (4-7%)

Unsur-unsur Pokok Enzim

Glukosa-6-fosfatase umumnya dibuat sebagai penandaenzim untuk RE membran. Kita akan membahas perannya ketikakita berurusan dengan metabolisme karbohidrat dalam RE
.
Enzim yang melimpah di RE adalah sitokrom P-450. Hal initerjadi pada tingkat setinggi 10% protein microsomal. Sitokrom inimerupakan salah satu anggota RE dalam pengangkutan elektronjaringan yang meliputi NADPH-sitokrom p-450 reduktase danphosphatidylcholine. Kompleks ini terlibat dalam reaksi hidroksilasiyang kita membahas secara lebih rinci dalam Bagian 6.4. enzimmembran penting lain yang berpartisipasi dalam aspek-aspektertentu dari pengangkutan elektron ini adalah sitokrom b5. Inijuga memiliki sifat reduktase, enzim yang mengandung NADH.
Ada banyak enzim RE yang kita tidak bisa bahas dalamlingkup teks ini. Diantaranya memiliki fungsi biosynthetic, sepertisintesis eter yang terkait dengan glycerolipids, sekelompokkompleks membran lipid. Serta lainnya yg memiliki peran katabolikserta mengenai kerusakan membran lipid

Topography of Enzyme in Lateral Plane
Vesikel-vesikel microsomal heterogen tergantungdengan kerapatan, ukuran, dan muatan permukaan, yangmemungkinkan bagian-bagian kecil menjadi kelompoksesuai dengan sifat-sifatnya. Semakin kecil vesikula,semakin besar tingkat heterogenitas. Vesikula yang lebihkecil akan memiliki ruang yg sedikit untuk enzim, secarastatistik diduga akan ditemukannya satu vesikel yangtidak sempurna pada beberapa komponen enzim jikaenzim yang didistribusikan secara acak dalam membran.Di sisi lain, tingkat kehidupan ditandai dengan asosiasilateral kelompok-kelompok enzim tertentu

data-data sejauh ini menunjukkan kuantitatif tetapiperbedaan-perbedaan qualitatitve tidak ada dalam susunan enzimdari vesikel. Jadi, misalnya, Semua vesikel mengandung glukosa-6-fosfatase, sitokrom b5 dan NADH-sitokrom p-450 reduktase.Meskipun semua enzim sama dalam setiap vesikel, vesikel ygberbeda mungkin berisi enzim dalam rasio yang berbeda.
Heterogenitas mencerminkan mobilitasnya dibatasi olehprotein dalam lapisan lipid bilayer atau mungkin mengungkapkantingkat struktural interaksi penting bagi kegiatan enzimatik. Selainitu, heterogenitas mungkin karena variasi dalam sifat ER dalam seltunggal atau dalam sel yang berbeda dalam satu jaringan atau organ,daripada heterogenitas dalam kurun waktu terus-menerus darimembran ER.

Topography of Enzyme in the Transverse Plane
Tabel 6.1 termasuk lokalisasi topografi berbagai enzim ER.Hasil lebih jelas daripada studi lateral distribusi. Semua enzim yangterdaftar, kecuali sitokrom p-450. Sitokrom p-450 terlihat sepertiprotein transmembran. Posisi enzim ini dalam membran yang tidakdiragukan lagi berhubungan dengan fungsi katalitik.
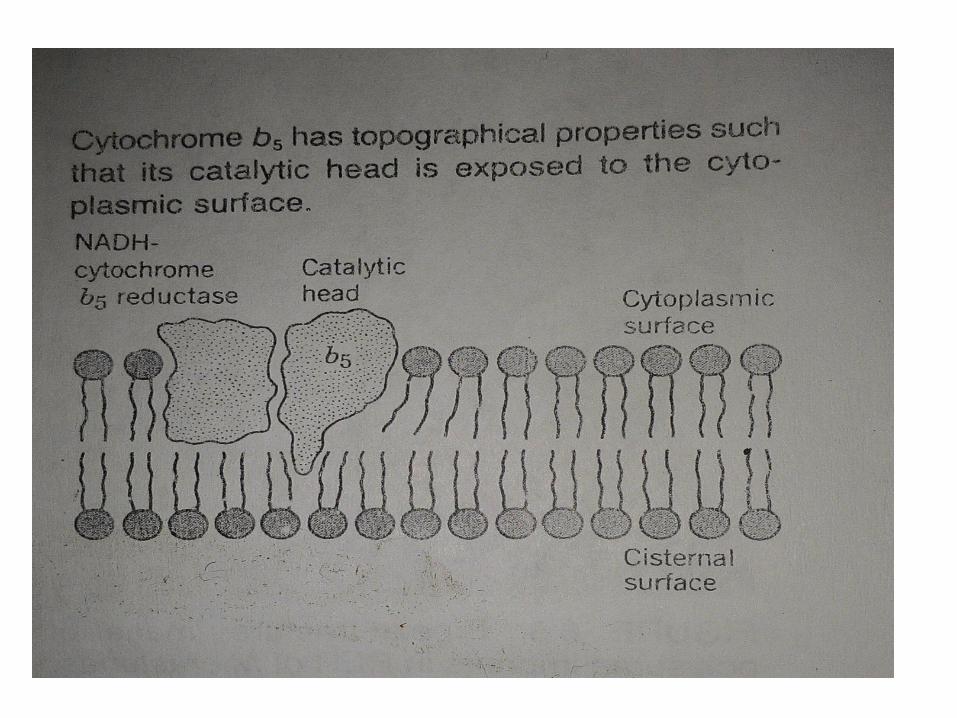
Studi rinci pada sitokrom b5 telah menyediakan beberapawawasan yang luar biasa terhadap sifat topografi enzim. Sitokromb5 adalah protein membran integral (berat molekul 11.000 daltons)yang condong ke arah permukaan sitoplasma. Hasil dari studidalam sejumlah laboratorium telah mengarah pada kesimpulanbahwa enzim memiliki hydrophilichead yang mengandung sisi aktifcatalytically dan hidrofobik ekor yang tidak aktif catalytically. Ekorhidrofobik larut dalam lemak bilayer membran. Kepala catalyticbebas berikatan dengan daerah permukaan hidrofil membrandimana berinteraksi dengan molekul substrat yang sesuai.

Bukti menunjukan bahwa NADH-sitokrom b5 reduktase, enzim inimenyuplai elektron ke sitokrom b5. Kedua enzim mungkinberinteraksi secara fisik dalam melaksanakan fungsi katalitikmereka.
Meskipun jenis interaksi protein integral membran dengankedua bagian hidrofobik dan hidrofil membran mungkin umum,nampaknya tidak melanggar aturan. Protein membran tertentuyang lain menampilkan sifat yg kurang amphipathic, dan mungkinlebih, atau benar-benar, tenggelam di wilayah hidrofobikmembran.

HUBUNGAN ANTARA RE KASAR DAN HALUS
Seperti telah ditunjukkan, terdapat perbedaanfungsional antara RE halus dan kasar. SER terlibat dalamsintesis dan metabolisme molekul kecil tertentu danmekanisme cellular detocification, sedangkan RER terlibatdalam sintesis dan transportasi protein.

RE Kasar Memberikan Perkembagan Pada RE Halus
Dua bentuk yang berbeda dari segi morfologi yg berdekatandalam sel. Salah satu bentuk SER, mungkin berasal dari yang lain.Point pada kasus ini, Phenobarbital(sejenis obat yg memiliki efekpenenang) merangsang proliferasi membran RE. Hal ini terjadi dlmdua tahap proses. Pertama, RER proliferasi dan kemudian SERberproliferasi. Fungsi dari SER yg terkait dengan Phenobarbital yaitureaksi detoksifikasi, enzim detoksifikasi(yg telah dibentuk)bergabung ke dalam membran dengan ribosom terikat. Makamunculnya RER pertama, diikuti dengan peningkatan SER.

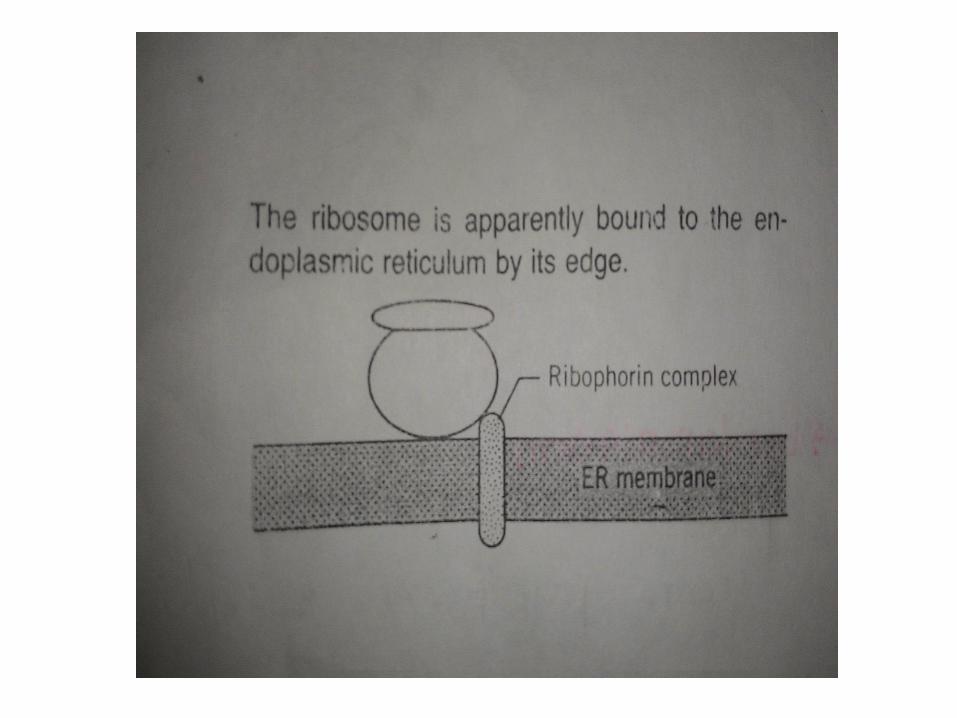
Dengan demikian, RER dan SER memiliki fungsionalberkaitan tetapi tidak sama. Sejumlah penelitian telahmenunjukkan bahwa microsomes kasar dan halus, dapatdipisahkan oleh tekhnik kepadatan gradien. Namun, banyakpeneliti telah melaporkan perbedaan antara microsomeskasar dan microsome halus, ada bukti yg menunjukkansetidaknya terdapat dua protein membran yang merupakankarakteristik dari microsomes kasar dan memiliki fungsisebagai tempat melekatnya ribosom. Protein ini disebutribophorin, memiliki berat molekul 63.000 dan 65.000daltons. Diduga bahwa fungsi lain dari ribophorin yaitudapat membentuk jaringan perancah seluruh membraninterkoneksi sisi pelekatan ribosom. Jaringan filamen inimemberi RER bentuk lamellar yang khas, yang cenderunglebih kaku dan stabil daripada bentuk vesikuler dan tubularpada SER

Ribosome Binding Sites on ERSintesis protein dibawa keluar kedua ribosom bebas dan
terikat menuju RE. Ribosom bebas menghasilkan protein larut disitosol, sedangkan ribosom terikat menerjemahkan sekresi proteindan protein membran integral. Dalam sel hewan ribosom berbarisdi messenger RNA yang membentuk polysomes, yang mungkin jugabebas atau terikat di ER ketika memproduksi protein.
Ketika ribosom melekat di ER, mereka menempel padasubunit besar. Umumnya diterima ribophorins I dan II, dua proteinintegral transmembran, adalah protein yang mengikat ribosom dandengan demikian dpt menempel pada membran.
Teknik membekukan-fraktur 11-nm partikel telah diamatipada ER membran yang tampaknya setara struktur ribosommengikat situs. Diperkirakan bahwa setiap partikel tersusun atassatu molekul ribophorins I dan II ditambah dengan minculnya rantaipolypeptida.

Gambar 6.9 mocrograph elektron terdapat bekas polysomepada membran RER. Pada umumnya, satu partikel 11 nm untuksetiap ribosom jejak di polysome dan partikel ini terletak diperipherally dekat jejak. Oleh karena itu akan muncul seolah-olahsubunit besar ribosom berlabuh di partikel integral membran tidaksecara langsung di pusat subunit tetapi lebih paada ke satu sisi.
Hubungan antara ribophorins dan partikel-partikel yangdilihat dengan mikroskop elektron tidak pasti, namun diyakinibahwa mereka sama. Dengan demikian kita dapat melihat proteinkompleks yang bertugas untuk mengikat ribosom di RER.
SER dan RER dalam segi fungsi dan struktural saling terkait,namun berbeda dalam sifat dasar mereka

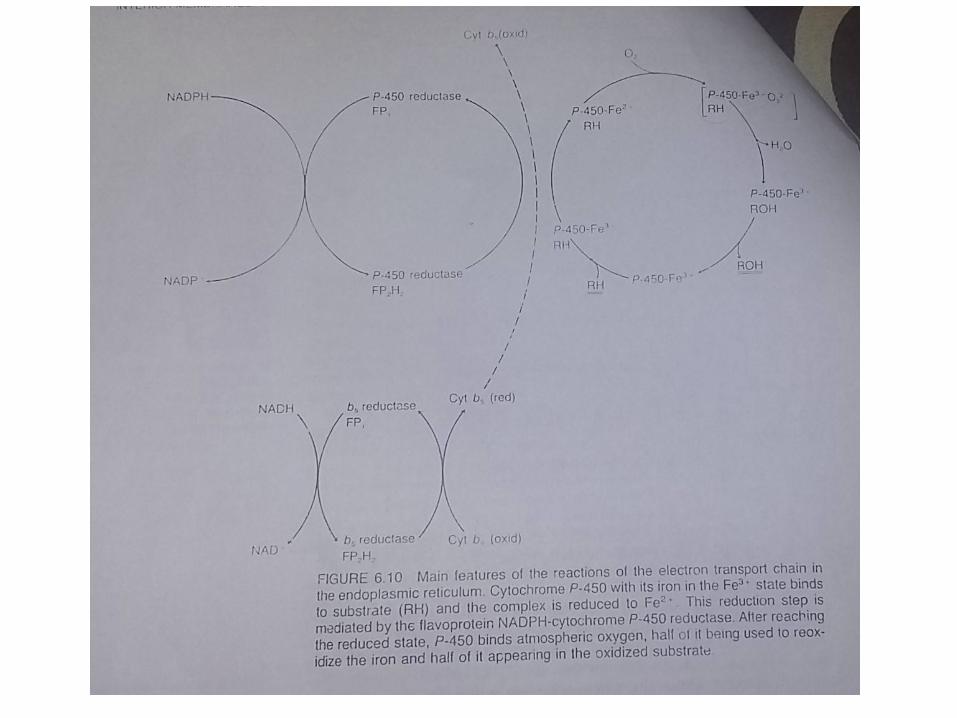
The Hydroxylation System of The Endoplasmic Reticulum

The Mixed-Function Oxidase System • Pada proses ini terjadi penambahan elektron
• Reaksi diatas disebut reaksi mixed-function oxidase atau monooxygenases yang akan dikatalisasi oleh enzim
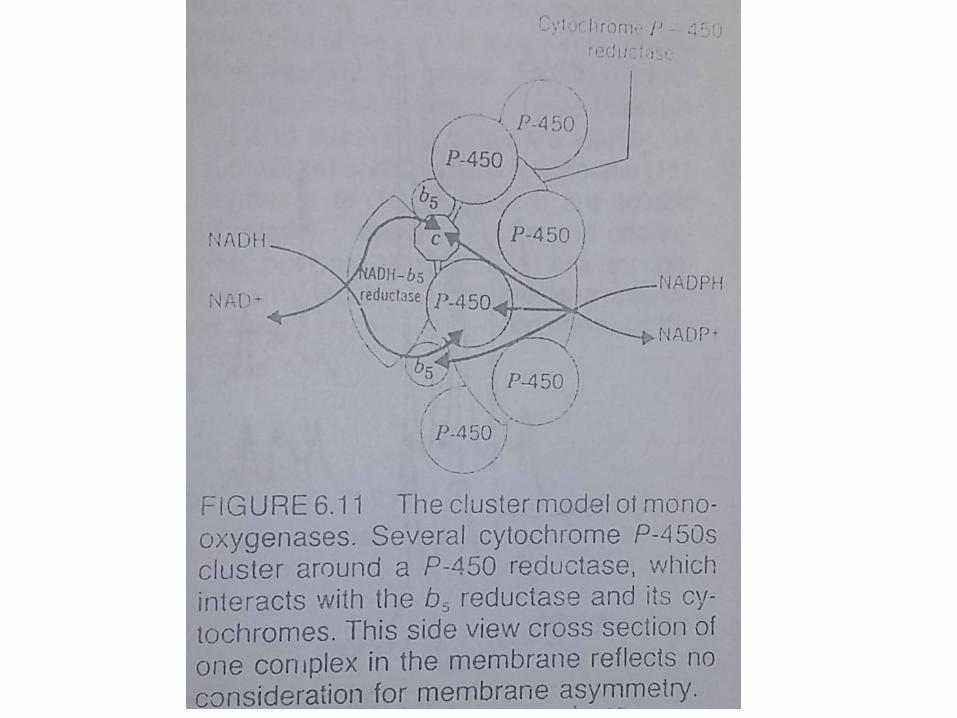

Induction of Enzym of Endoplasmic Reticulum• Phenobarbital akan berperan sebagai induktor
untuk meningkatkan kerja enzim.
• Pertama aktivitas enzim terdapat di RER
• Setelah 6 jam pemberian obat, aktivitas enzim meningkat di SER
• Hal ini menunjukkan bahwa obat menginduksi pembentukan enzim baru yang di translasi di RER kemudian enzim tersebut akan berpindah ke SER

• Selain meningkatkan kerja enzim induksi obat jugamenstimulasi proses biosintesis seperti sintesiskolesterol dan fosfolipid
• Sintesis fosfolipid berlangsung lebih awaldibandingkan dengan sintesis protein
• Komponen monooxygenase sistem akan ditambahkanpada cytochrome P-450 dan NADPH-cytochrome P-450 tertentu

• Membran akan terus berpoliferasi dan kemudian terlepas membentuk formasi autofagosom
• Membran RE hasil poliferasi tidak membawa semua enzim hasil induksi, karena tidak semua enzim bertambah ketika terjadi proses induksi

Properties of Cytochrome P-450
• Cytochrome P-450 merupakan protein integral transmembran yang juga merupakan enzim utama dalam RE yang ada di sel hati.
• Komposisi Cytochrome sebesar 10% dari total protein non-induksi
• Jika diinduksi akan bertambah menjadi 25-30 % dari total protein

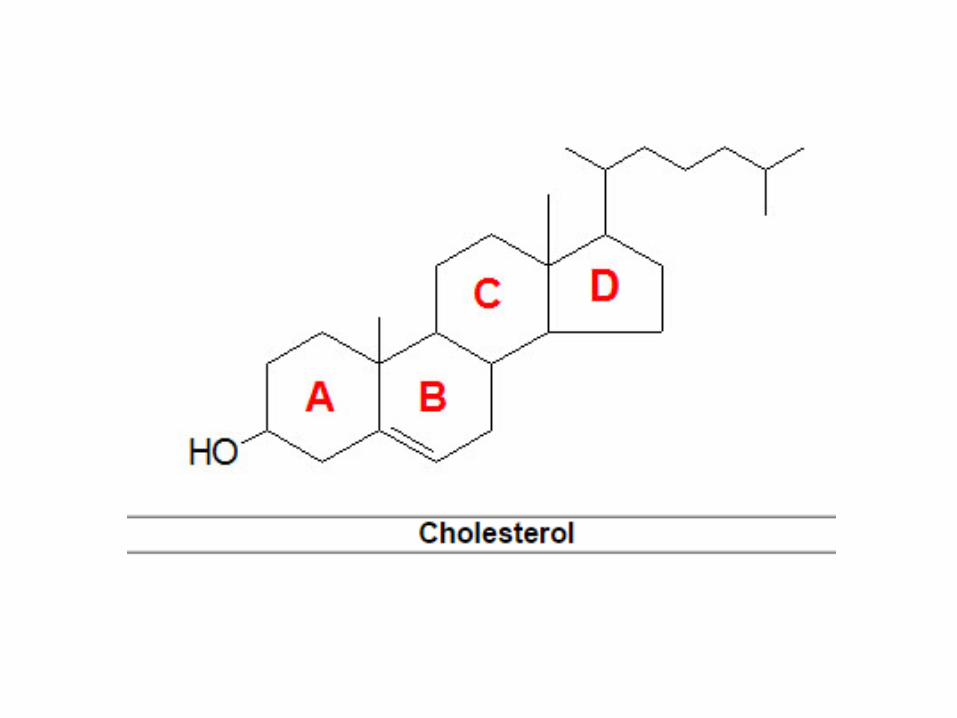
Biosintesis Kolesterol
• Kolesterol merupakan molekul penting yangmemiliki peran pada struktur membran sebagaiprekusor sintesis hormon steroid, asam empedudan vitamin d
• Biosintesis kolesterol yang berlangsung di dalam REmelibatkan sejumlah enzim.
• Enzim-enzim tersebut ada dalam bentuk enzimterlarut, dan yang lain dalam bentuk enzim yangterikat pada membran.

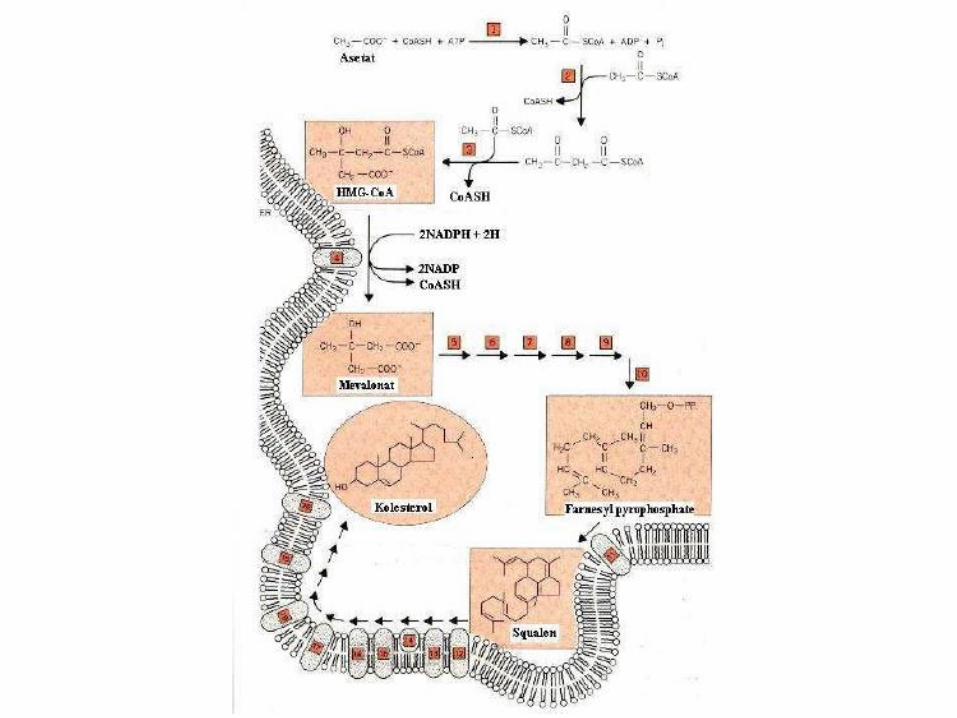
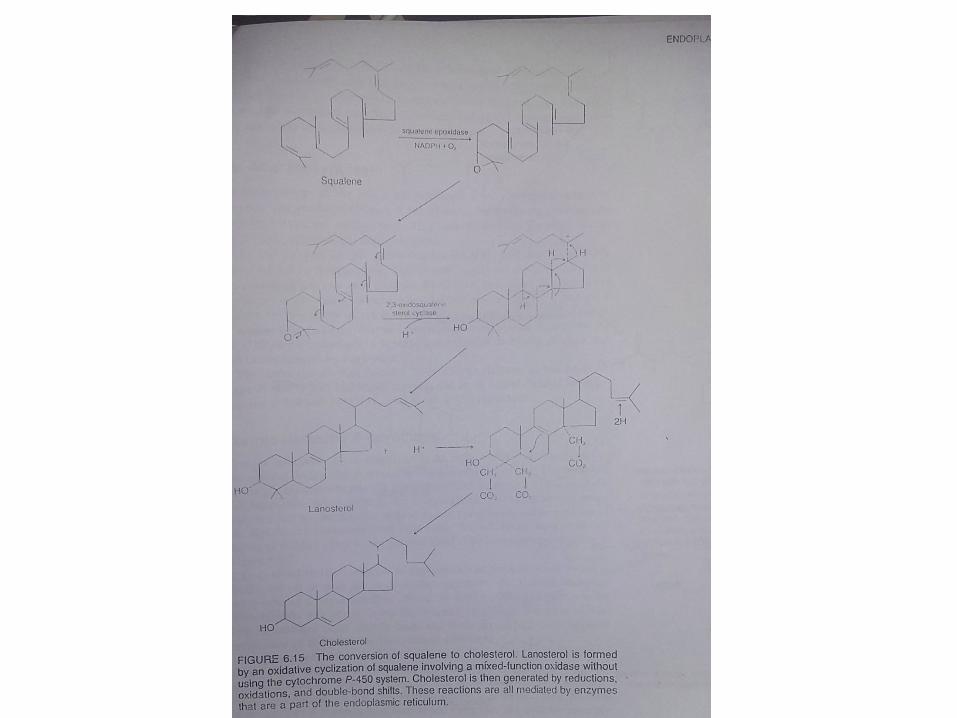
Tahapan Biosintesis Kolesterol
• kolesterol disintesis dari asetat dan secara bertahapmenghasilkan β-hidroksi-β-metilglutaril coenzim A(HMG-CoA).
• HMG-CoA selanjutnya diubah menjadi mevalonatdengan bantuan enzim HMG-CoA reduktase yangmerupakan enzim terikat membran
• Mevalonat selanjutnya diubah secara bertahapmenjadi Fernesyl pirofosfat

• Semua enzim yang berperan dalam perubahanmevalonat menjadi Farnesyl pirofosfat merupakanenzim-enzim terlarut.
• Farnesyl pirofosfat selanjutnya diubah menjadisqualen dengan bantuan squalen sintetase.
• Squalen sintetase selanjutnya diubah secara bertahapmenjadi kolesterol.

Biosintesis Asam Empedu
• Biosintesis asam empedu menggambarkan pola enzim dan produk yang sangat kompleks (Gambar 6.16).
• Enzim kolesterol 7α-hidroksilasi mengkatalisis reaksi pertama pada biosintesis asam empedu
• Reaksi hidroksilasi yang mengubah 7α-hidroksilasi pada kolesterol menjadi asam empedu membutuhkan NADPH dan molekul oksigen, dan tergantung pada sitokrom P-450 dan NADPH-sitokrom P-450 reduktase.

Biosintesis Asam Empedu

Biosintesis Hormon Steroid
• Hormon steroid yang disintesis dalam sel yang menyusunjaringan korteks adrenal, ovarium, testis, dan plasenta.
• Hormon steroid diproduksi secara terkoordinasi olehenzim yang sebagian terletak di mitokondria dan sebagiandi retikulum endoplasma.
• Karena itu, kunci metabolit harus bolak-balik melintasimembran mitokondria untuk menyelesaikan jalurbiosintesis.
• Pregnenolon adalah prekursor utama untuk semuahormon steroid.
• Pregnenolon berasal dari kolesterol oleh pembelahanoksidatif dari rantai sisi dalam mitokondria.

Biosintesis Hormon Steroid
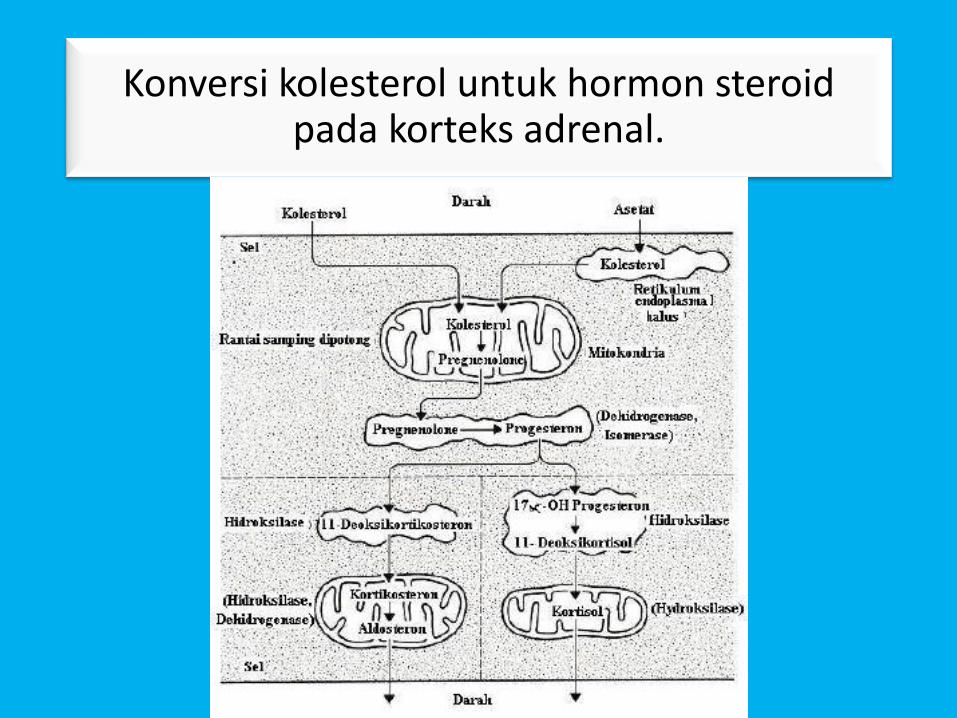
Konversi kolesterol untuk hormon steroid pada korteks adrenal.

• Reaksi hidroksilasi akhir yang membubuhkankelompok hidroksil pada posisi 11β dari dua produkini dikatalisasi oleh campuran-fungsi oksidase yangberoperasi di dalam mitokondria.
• Ini tergantung pada sitokrom P-450 rantai transportelektron yang sama dalam banyak hal tetapi tidakidentik dengan P-450 di retikulum endoplasma.
• Reaksi mitokondria dengan demikian diperlukanuntuk menyelesaikan pembentukan tiga hormonsteroid adrenal utama: kortikosteron, aldosteron,dan kortisol.

METABOLISME KARBOHIDRAT
• Retikulum endoplasma berperan dalam metabolisme karbohidrat, tetapi bukti keterlibatan langsung tidak jelas dan konklusif.
• Sistem di mana retikulum endoplasma mungkin memiliki peran penting, yaitu :
1. Homeostasis Glukosa darah
2. Sintesis glikoprotein

Homeostasis Glukosa Darah
• Glukosa-6-fosfatase adalah enzim yang terikat eratdengan membran retikulum endoplasma.
• Hal ini biasanya dianggap sebagai gluconeogenicphosphohydrolase yang mengkatalisis pelepasanglukosa bebas dari bentuk terfosforilasi dalam hati,sehingga dapat mempertahankan tingkat homeostatisglukosa dalam darah untuk pemeliharaan sel-sel darahmerah dan jaringan saraf.
• ini mungkin berfungsi dalam pemecahan glikogen(glikogenolisis).

• Pada mamalia, sebelum kelahirannya belummemiliki enzim, karena embrio menerimapasokan glukosa melalui plasenta dari sirkulasimaternal.
• Segera setelah lahir ada peningkatan yangsignifikan pada aktivitas glukosa-6-fosfatase.
• Dalam kehidupan hewan yang dimulai dengantelur, enzim sudah ada dalam telur, karena tidakada pasokan eksogen glukosa.

• Reaksi Glukosa-6-fosfatase

• Lokasi glukosa-6-fosfatase tidak terbatas padahati, meskipun aktivitasnya dalam bentuksintetis dan hidrolitik telah diamati dalam hatilebih dari 20 spesies mamalia, burung, amfibi,dan invertebrata.
• Tetapi juga telah sering diamati pada jaringanginjal dan telah dipantau dalam sel epitel ususkecil, dan dalam jumlah yang lebih rendahtetapi signifikan dalam organ seperti otak,pankreas, kelenjar adrenal, dan testis.

Sintesis Glikoprotein
• Sebagian besar protein yang ditakdirkan untuktransportasi ke luar sel adalah glikoprotein. Iniadalah protein kompleks yang mempunyai satuatau lebih unit karbohidrat yang melekat.
• Peran retikulum endoplasma menyerap proteinsekretori ke sisterna dan mulai modifikasi awalberbagai jenis protein sekretori melaluipenambahan residu karbohidrat pada ujungproksimal dari polipeptida dan proses selanjutnyaberlangsung di dalam badan golgi sepertidirangkum pada gambar 6.20.
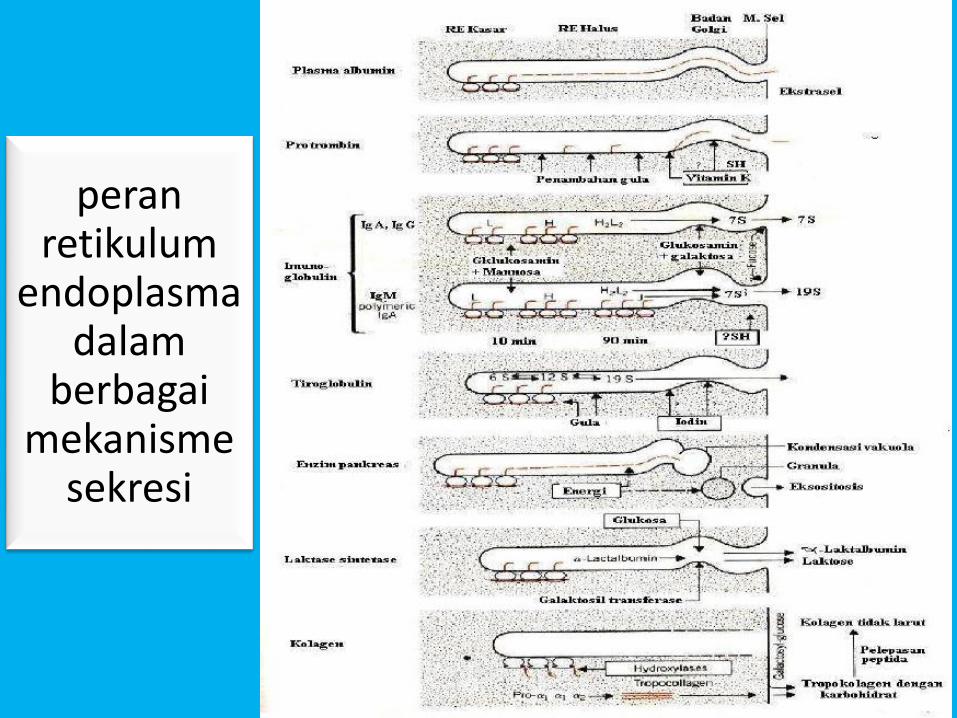
peran retikulum
endoplasma dalam
berbagai mekanisme
sekresi