respiration and oxidation...
TRANSCRIPT

Respiration and Glucose Oxidation in Human
and Guinea Pig Leukocytes: Comparative Studies
ROBERTL. BAEHNER,NEALGiLMAN, and MANFBEDL. KARNOvSKY
From the Departments of Pediatrics and Biological Chemistry, HarvardMedical School, and the Division of Research Hematology, Children'sHospital Medical Center, Boston, Massachusetts 02115
A B S T R A C T A comparison has been made of themetabolic shifts in human and guinea pig leukocyteswhen they phagocytize. Respiration of guinea pig poly-morphonuclear leukocytes (PMN) and the incrementduring phagocytosis were each about 2k-fold that ofhuman PMN. This was also true of the direct oxidationof glucose-6-P (hexose monophosphate shunt). Enzymespotentially responsible for these phenomena have beencompared in each species. Cyanide-insensitive NADHoxidase and NADPHoxidase were measured and onlythe former exhibited adequate activity to account for therespiratory stimulus durintg phagocytosis. The hydro-gen peroxide formed by this enzyme stimulates the hexosemonophosphate shunt by oxidizing glutathione whichupon reduction by an NADPH-linked glutathione re-ductase provides NADPto drive the hexose monophos-phate shunt. Other linkages between respiratory stimu-lation and that of the hexose monophosphate shunt alsopertain in the guinea pig.
INTRODUCTION
Metabolic changes concomitant with phagocytosis inleukocytes have been the subject of detailed investiga-tion for the past decade (1-6). The initial observationwas made by Baldridge and Gerard (7) that canine leu-kocytes display a burst of "extra respiration" duringphagocytosis. This respiratory increment was thoughtto be intrinsic to the phagocytic act, i.e., the ingestionprocess per se, until it was shown by Sbarra and Karn-ovsky (1) that guinea pig leukocytes ingested polysty-rene spherules to the same extent in a pure nitrogenatmosphere as they did in an aerobic milieu. The latterauthors concluded from the above and from evidence ob-
Dr. Baehner is an Established Investigator of the Ameri-can Heart Association.
Received for publication 5 September 1969 and in revisedform 19 November 1969.
tained with metabolic inhibitors that the process isdriven by metabolic energy derived from glycolysis.The respiratory increase and marked stimulation ofdirect oxidation of glucose-6-phosphate (hexose mono-phosphate shunt, HMP) are concomitants of the proc-ess, whatever their function.
The enzymatic basis of the respiratory burst duringphagocytosis has been the subject of some controversy.Some workers believe that it is due to the oxidation ofNADHby a flavoprotein oxidase (8). Others are of theopinion that an oxidase for NADPHis cardinal (9).In the intact cell, the respiratory burst is accompaniedby Ha02 production (3); both enzyme systems sug-gested above do produce this substance when theirsubstrates are oxidized (8-10).
Recently interest in the metabolism of the humanperipheral blood leukocyte has been kindled afresh bystudies on leukocytes from children with chronic granu-lomatous disease (CGD) (11, 12). In that condition,ingestion of bacteria (or other particles) by leukocyteshas been reported to be normal, but the respiratory in-crement and the stimulation of direct oxidation of glu-cose-6-phosphate is depressed.
The purposes of our paper are as follows: (a) Tocompare human and guinea pig polymorphonuclear leu-kocytes (PMN) with respect to their respiratory re-sponses, and increases in HMPactivity during phago-cytosis. In assessing the metabolic aspects of the humanPMN, the behavior of guinea pig PMNhas previouslybeen accepted as a yardstick and comparative data ob-tained under similar conditions would be helpful; (b)To investigate the quantitative aspects of the two en-zymatic activities mentioned above (i.e. NADHoxi-dase and NADPHoxidase). Such information shouldpermit one to admit the possibility of involvement ofan enzyme in the respiratory burst during phagocytosisor suggest its inadmissibility, on the basis of quantita-tive adequacy; (c) To establish, particularly for human
692 The Journal of Clinical Investigation Volume 49 1970

PMN, a linkage between the respiratory increment ofphagocytosis and the stimulation of the HMP. Since theHMPis regulated by the availability of NADP, adequateoxidation of NADPHper se would provide a directlink to the increased oxidation of glucose-6-phosphate.If such oxidation of NADPHwere inadequate, and thatof NADHadequate, it becomes necessary to establishthe connection between NADH oxidation and theprovision of NADPfor the increased HMPactivity.
METHODS
Preparation of leukocyte suspensionsHuman peripheral blood was collected in heparinized'
plastic syringes from patients with granulocytosis due to in-fection. The syringes were inverted and the red cells sedi-mented at 37'C for 30-60 min. The supernatant plasmacontaining the PMN was decanted into 40-ml siliconizedglass centrifuge tubes. For studies of leukocyte homogenates,the red cells were lysed using three volumes of 0.87%NH4Cl to one volume of white cell suspension in plasma.This procedure, if rapidly performed (5 min), did not al-ter the activity of the leukocytic enzymes assayed in thisstudy. The plasma was centrifuged at 250 g for 5 min andleukocyte button was washed twice with Krebs-Ringer phos-phate solution (KRP), pH 7.4. The yield was between 0.5ml and 1.0 ml of packed leukocytes per 50 ml of wholeblood, and red cell contamination was 1: 1 if they were notremoved with NH4Cl. The percentage of mature granu-locytes varied between 80 and 90% of the leukocytes.Donors with a shift to the left to the metamyelocyte stagewere not used for this study.
Guinea pig peritoneal leukocytes were prepared by themethod of Stiihelin, Karnovsky, Farnham, and Suter (13)using a 12%o suspension of sterile sodium caseinate. 15 ml ofcaseinate were injected into the peritoneal cavity and theleukocytes were isolated 15-18 hr later. The usual yield was1 ml of packed leukocytes per animal. The percentage ofmature neutrophilic granulocytes was greater than 90%of the leukocytes.
Studies of respiration, glucose oxidation, andformate oxidation by whole cellsLeukocytes were suspended in KRP with 5-10 mmglu-
cose to a concentration of 10% (v/v). Respirometry wasperformed in micro Warburg flasks 2 with air as the gasphase and a total fluid volume of 1.1 ml at 37'C, usingstandard manometric techniques. Incubations were carriedout for 45 min after the addition of latex particles (0.81 ,diameter,' 0.2 ml 1: 3 v/v dilution in KRP) tipped from theside arm. Total cell protein per- flask varied between 2.9mg and 5.5 mg which represented a cell population of be-tween 2.8 X 107 and 6.0 X 107. '4CO2 evolved from the oxi-dation of glucose-l-'C and formate-14C ' was trapped incenter wells containing 0.1 ml 5 N NaOH; where res-piratory measurements were not required 10-ml Erlenmeyerflasks equipped with center wells and vaccine ports were
1Liquaemin Sodium "10," Organon Inc., West Orange,N. J.
'Arthur H. Thomas Co., Philadelphia, Pa.S Bacto-Latex, Difco Laboratories, Detroit, Mich.' NewEngland Nuclear Corporation, Boston, Mass.
used.' The contents of the wells were transferred to vialscontaining 15 ml phosphor solution and the radioactivity ofsuitable standards and samples was determined in anAnsitron liquid scintillation counter. The specific activityof the substrates was determined so that radioactivity mea-surements could be converted to nmoles mg-1 protein hr-1.
Preparation of leukocyte homogenates and sub-cellular fractionsSuspensions of intact leukocytes were centrifuged 10 min
at 250 9 at 4'C, and the pellet of cells was then resuspendedand washed once either with cold 0.34 M sucrose bufferedwith NaHCO3 to pH 7.4 or with cold 0.154 M KCl con-taining 3.2 X 10' M KHCO8 (alkaline isotonic KC1).The washed cells were again centrifuged 10 min at 250 gat 4'C. The cell button was then suspended at 15% con-centration (v/v) with the appropriate homogenizing mediumand a 5 min homogenization was performed utilizing aTeflon pestle' in a 50 ml glass tube packed in ice. This pro-cedure disrupted 95-98% of the cells (microscopic examina-tion). The homogenate was then centrifuged 10 min at 250 9at 4°C and the supernatant (Si) was isolated. The buttonwas resuspended in the appropriate homogenizing mediumand a second homogenization procedure was performed inidentical fashion. The supernatant fluid obtained (S2) wascombined with Si and again centrifuged for 10 min at 250 9at 4°C. The buttons contained nuclei and disrupted mem-branes and a few unbroken cells; this fraction was desig-nated the "debris fraction." The supernatant fluid wascentrifuged at 40,000 g' for 15 min at 4°C. The clear super-natant fluid obtained after this centrifugation was desig-nated "supernatant fraction" and the button, which con-tained large and small granules, was resuspended in sucroseor KCl (concentrations as above) and was designated the"granule fraction." The protein content of these fractions andthe whole homogenate was determined by the method ofLowry, Rosebrough, Farr, and Randall (14).
Enzymatic assays of subcellular fractionsNADHoxidase was assayed using cuvettes with 1 mm
light path in a Perkin-Elmer spectrophotometer model 202equipped with a scale expansion continuous recorder7 andthermostat.' The NADHconcentration was 2.5 X 10' M in0.1 M potassium phosphate buffer, pH 7.0, in the presence of1 mMKCN. The decrease in absorbance at 340 mi -was re-corded for 15 min and converted to nanomoles NADHoxi-dized mg1 protein min1. For comparative studies withNADPHoxidase, 1 cm light path cuvettes were used atNADPHand NADH concentrations of 1.6 X 10' mole/liter. NADPHoxidase activity was also assayed using theClark membrane electrode' attached to a polarograph formeasuring PoN and an expanded scale recorder set torecord a change in PoN of 50 mmHg over the scale. Theelectrode well contained a fluid volume of 1.8 ml and wassurrounded by a constant temperature water bath set at37'C. NADPHoxidase activity was measured by the rateof decrease in Po, after addition of the reduced pyridinenucleotide at a final NADPHconcentration of 7.0 X 10-'mole/liter.
'Kontes Glass Co., Vineland, N. J.eInternational Model B-20. International Equipment Co.,
Needham Heights, Mass.'Leeds and Northrup, Philadelphia, Pa.'Haake, F. J., West Germany.' Instrumentation Laboratory Inc., Boston, Mass.
Comparative Studies of Metabolism in Leukocytes 693

Lactate dehydrogenase was assayed by recording the rateof decrease in absorbance of either NADHor NADPHat340 myA as described by Evans and Karnovsky (15). Thereaction mixture had a final volume of 3.0 ml and a finalconcentration of potassium phosphate buffer 0.03 mole/liter,pH 5.6 or 7.4; NADHor NADPHwere 3.0 X 10' mole/liter; pyruvate was 3 X 10' mole/liter; and supernatantfraction had 0.10 mg protein per assay.
Glutathione reductase was measured by the method ofRacker (16), recording the rate of NADPHdisappearanceat 340 m/A after the addition of oxidized glutathione andsuitable enzyme fractions to the reaction mixture. The reac-tion mixture contained a final volume of 1.0 ml in 1 cmlight path quartz cuvettes with a 1.5 ml capacity, and afinal concentration of 0.2 M potassium phosphate buffer,pH 7.6, 0.01% bovine serum albumin, NADPH1.0 X 10'mole/liter, oxidized glutathione 6.5 X 10' mole/liter, andsupernatant enzyme fractions containing between 0.10 mgand 0.50 mg protein.
NADPHdehydroascorbate reductase was assayed by re-cording the rate of disappearance of NADPHat 340 m~iat 370C. Dehydroascorbate was prepared by the method ofDamron, Monier, and Roe (17), by the addition of onedrop of concentrated bromine to 6.0 X 10' M ascorbic acidand subsequent aeration for 30 min. This sample was thenbuffered with Na2CO8 to pH 7.0. The reaction mixture con-tained a final volume of 1.0 ml and a final concentration of0.25 M sodium phosphate buffer, pH 7.0, NADPH 1.8 X10' mole/liter, dehydroascorbate 2.0 X 10' mole/liter, andleukocyte homogenate fractions containing 1.0 mg protein.
Measurement of potential hydrogen donors inthe human leukocytes
Reduced glutathione was determined using a micro-adapta-tion of the method of Beutler, Duron, and Kelly (18) basedupon the development of relatively stable yellow color when5,5'-dithiobis (2-nitrobenzoic acid) is added to sulfhydryl
compounds.Ascorbic acid was determined using the method of Lowry,
Lopez, and Bessey (19) who adapted the method of Roe andKuether (20), which is based upon the colorimetric deter-mination of a solution of copper sulfate-thiourea-dinitro-phenylhydrazine.
RESULTSComparison of respiratory responses and oxidation of
glucose-6-P. The respiratory response of human periph-eral blood leukocytes during phagocytosis was comparedto that of guinea pig peritoneal leukocytes (Table I).Human leukocytes have a lower oxygen consumption atrest and during phagocytosis but the ratio of activitiesis similar to that in guinea pig cells. It should be em-phasized that the results pertain to both species eitheron the basis of cell number, or on the basis of cell pro-tein, since 1.0 X 1OT leukocytes from human peripheralblood or guinea pig peritoneal cavity yield approxi-mately 1.5 mg protein.
Glucose-6-phosphate oxidation via the hexose mono-phosphate shunt was assessed by measurement of '4CO2evolution from glucose-1-"4C. The results, expressed asnanomoles glucose oxidized mg' protein hr-1 were alsocompared for human and guinea pig leukocytes, at restand after phagocytosis (Table I). The resting valuesare quite similar but the increment during phagocytosiswas much greater for guinea pig leukocytes. In orderto evaluate more accurately the amount of glucose thatcycles through the shunt, 14CO- evolution was measuredusing glucose -1-2C and glucose-2-2C in human leuko-
TABLE IRespiratory and Oxidative Activities of Resting and Phagocytizing Leukocytes*
Guinea pig (4) Human (4)
Resting Phagocytizing Resting Phagocytizing
Oxygen consumption .... ,uliters' 2.5 6.2 1.1 2.5(2.0-3.2) (5.0-7.5) (1.0-1.4) (2.0-2.7)
nmoles 112 277 49 112Ad 165 63
Glucose oxidationt ...... nmoles 8 56 6 26(5-12) (48-62) (4.9-7.0) (18-33.6)
A\ 48 20
NADPHproduction§ .... nmole5 19 134 14 . 62A 115 48
Formate oxidation!. timbles 2.9 7.8 0.6 2,8.(0.8-6.9) (2.6-17.0) (0.2-1.1) (1 .1-5.9)
A 4.9 2.2
* All data reported mg-' protein hr-'. Ranges are given in parentheses.t Glucose-1-'4C - "4CO2.§ Corrected for the oxidation of carbon-2 of glucose (see text).11; Formate-"C "CO2'
694 R. L. Baehuer, N. Gilman, and M. L. Karnovsky
cytes at rest and during exposure to methylene blue(2.0 X 10' mole/liter). In two such experiments, it wasfound that the amount of glucose-2-`4C that yielded 14CO2in both resting leukocytes and after treatment of methyl-ene blue was 20% of the value for glucose-1-"4C. Be-cause the oxidation of 14C-2 of glucose to 14CO2 is rela-tively small, the data for human PMNwere extrapolatedto guinea pig cells. Since 2 moles of NADPHare pro-duced per mole of glucose oxidized to C02 by thispathway the extra oxidation of glucose during phago-cytosis represents 115 nmoles NADPHmg'1 protein hr""(for the guinea pig leukocyte) and 48 nmoles NADPHmg-' protein hr"" (for the human leukocyte).
Comparison of formate oxidation. The oxidation offormate-1'C has been shown by Iyer, Islam, and Quastel(3) to be an indication of intracellular hydrogen peroxideproduction. The rate of formate oxidation in wholeguinea pig leukocytes at rest was fivefold greater thanin human leukocytes, and during phagocytosis threefoldgreater than the value for human leukocytes (Table I).It should be emphasized that the oxidation of formateamounts to only a small fraction (3%) of the oxygenconsumption.
NADPHoxidase in human and guinea pig leukocytes.NADPHoxidase insensitive to 1 mmKCNwas verifiedin the granule fraction of guinea pig leukocyte homoge-nates as well as human leukocyte homogenates (TableII). Since the granule protein represents 25% of thetotal cell protein (both in guinea pig [21] and humancells) the rate of NADPH oxidation catalyzed byNADPHoxidase would be about 23 nmoles mg'" pro-tein hr"1 for the intact guinea pig leukocyte and 5 nmolesmg-' protein hr"" for the intact human leukocyte. Thisis inadequate activity to cover the reoxidation ofNADPHproduced during phagocytosis (see Table I),i.e., 115 nmoles mg-1 protein hr"" for guinea pig and 48nmoles mg"" protein hr"" for human leukocytes.
TABLE I INIADPH Oxidase of Leukocyte Granules*
Oxygen uptake
1si nmoles
Guinea pig (3) 2.0 90(1.05-2.60)
Human (3) 0.4 18(0.34-0.60)
* mg-' granule protein hr-1. Range and number of observa-tions in parentheses. Cyanide was present at 1 mmole/liter;temperature, 370C. It should be noted that granule proteinconstitutes 25%of total cell protein. Consequently, the valuesfor whole cell protein are about one-quarter of those reportedabove.
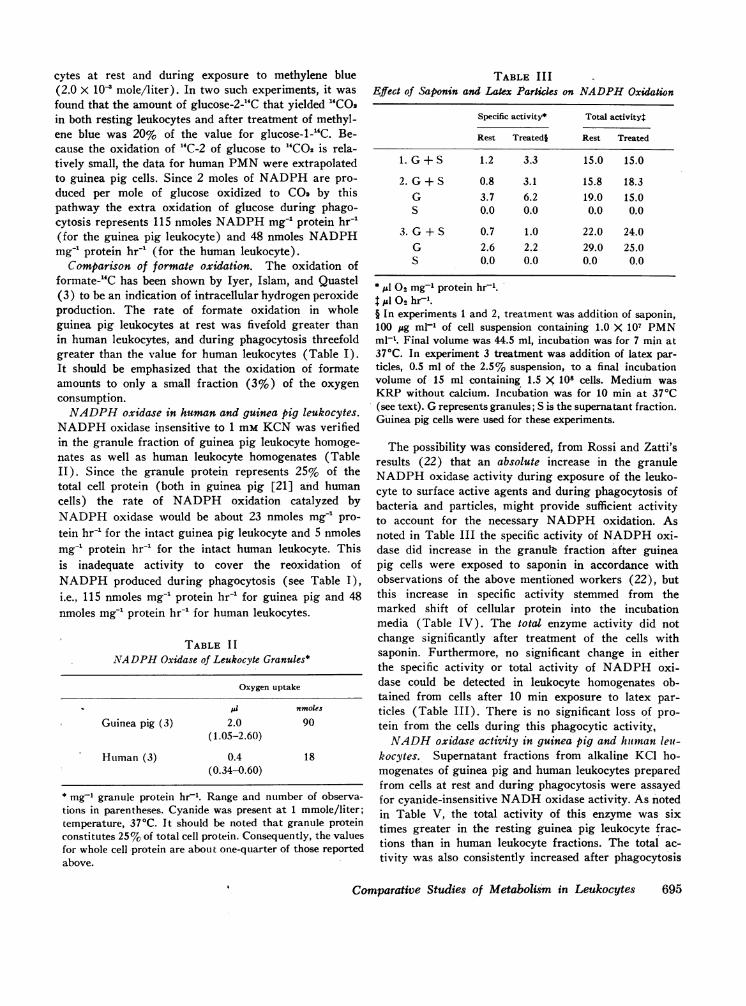
TABLE IIIEffect of Saponin and Latex Particles on NADPHOxidation
Specific activity* Total activity:
Rest Treatedi Rest Treated
1. G+S 1.2 3.3 15.0 15.0
2. G+ S 0.8 3.1 15.8 18.3G 3.7 6.2 19.0 15.0S 0.0 0.0 0.0 0.0
3. G + S 0.7 1.0 22.0 24.0G 2.6 2.2 29.0 25.0S 0.0 0.0 0.0 0.0
*jul O mg- protein hrf1.t JAI 02 hr-".§ In experiments 1 and 2, treatment was addition of saponin,100 ,sg ml"1 of cell suspension containing 1.0 X 107 PMNml-'. Final volume was 44.5 ml, incubation was for 7 min at370C. In experiment 3 treatment was addition of latex par-ticles, 0.5 ml of the 2.5% suspension, to a final incubationvolume of 15 ml containing 1.5 X 108 cells. Medium wasKRP without calcium. Incubation was for 10 min at 370C(see text). Grepresents granules; S is the supernatant fraction.Guinea pig cells were used for these experiments.
The possibility was considered, from Rossi and Zatti'sresults (22) that an absolute increase in the granuleNADPHoxidase activity during exposure of the leuko-cyte to surface active agents and during phagocytosis ofbacteria and particles, might provide sufficient activityto account for the necessary NADPHoxidation. Asnoted in Table III the specific activity of NADPHoxi-dase did increase in the granule fraction after guineapig cells were exposed to saponin in accordance withobservations of the above mentioned workers (22), butthis increase in specific activity stemmed from themarked shift of cellular protein into the incubationmedia (Table IV). The total enzyme activity did notchange significantly after treatment of the cells withsaponin. Furthermore, no significant change in eitherthe specific activity or total activity of NADPHoxi-dase could be detected in leukocyte homogenates ob-tained from cells after 10 min exposure to latex par-ticles (Table III). There is no significant loss of pro-tein from the cells during this phagocytic activity,
NADHoxidase activity in guinea pig and human leu-kocytes. Supernatant fractions from alkaline KCl ho-mogenates of guinea pig and human leukocytes preparedfrom cells at rest and during phagocytosis were assayedfor cyanide-insensitive NADHoxidase activity. As notedin Table V, the total activity of this enzyme was sixtimes greater in the resting guinea pig leukocyte frac-tions than in human leukocyte fractions. The total ac-tivity was also consistently increased after phagocytosis
Comparative Studies of Metabolism in Leukocytes 695

in both species. The total enzyme activity was clearlypresent in adequate quantities to mediate the respiratoryresponse in phagocytizing guinea pig cells, but ap-parently less definitely so in human cells. However, ifthe optimal pH is taken into account (Table V) theenzyme in both species would be sufficient to cover therespiratory needs.
If alkaline KCl solution was used as the homogeniza-tion medium, NADHoxidase was completely solubilized(Fig. 1). However, if 0.34 M sucrose was used for ho-mogenization, half of the NADHoxidase was shifted tothe granule fraction. It could be washed off such gran-ules with isotonic alkaline KCl. This was true for guineapig cells but was less evident for human cells.
Cagan and Karnovsky (8) reported that NADHoxi-dase of guinea pig leukocytes has a Km of 1.0 X 103mole/liter for NADHand a pH optimum of 4.5. NADHoxidase' from human leukocytes has a Km of 4 X 10-4mole/liter and a pH optimum similar to that of theguinea pig enzyme. Under the conditions of these mea-surements at pH 6, the guinea pig enzyme was foundin this work to have a Km of 2 X 10' mole/liter.
Linkage of respiration to the hexose monophosphatedirect oxidative pathway. Since it appears from theabove, and from other data (23), that NADHoxidaseis the enzyme responsible for the increased respirationduring phagocytosis, the mode of regeneration of NADPon which the shunt depends, has been considered.
Evans and Karnovsky (15) have noted a NADPH-linked LDH in guinea pig leukocytes which was opera-tive at low pH. Through the agency of this enzyme py-ruvate (available in glycolysis because of the action ofthe NADHoxidase and consequent deficit of NADH)could regenerate NADP. No such activity was found in
TABLE IVRelease of Protein from PMNduring Saponin Treatment
or Phagocytosis*
Granulesand
Medium Debris supernatant
Experiment IWithout saponin 27.9 24.5 47.6With saponin 77.6 10.6 11.8
Experiment IIWithout latex 4.7 54.0 41.3With latex 5.1 31.7 63.2
* The protein present in the external medium and in cellfractions was measured after 7 min incubation with or withoutsaponin; or after adding latex (see text and footnote of TableIII for details).
TABLE V
NADHOxidase Activity*
Guinea pig Human(4) (4)
Respiratory incrementi 165 63NADHoxidase pH 7.0 (resting) 144 24
(62-194) (17-31)pH 7.0 (phagocytizing) 219 39
(115-390) (22-57)pH 5.0§ ca. 400 ca. 80
* Nanomoles mg-' protein hr-1.¢ From Table I.§ Based upon pH optimum curves which are similar for enzyme from bothspecies.
the alkaline KCl supernatant fractions from human leu-kocytes.
Since leukocytes contain very large amounts of as-corbate (24), dehydroascorbate-NADPH reductase ac-tivity was considered, and was found in the granulefractions of human leukocyte homogenates. No activitywas found in the supernatant fractions. The nonenzy-matic oxidation of NADPHalso occurred very rapidlyin the presence of dehydroascorbate. The addition of 1mg of granule fraction protein produced an enhance-ment of NADPHoxidation but this rate was on its ownonly about half that necessary to support the reoxida-tion of NADPHin the hexose monophosphate shuntduring phagocytosis. No ascorbate oxidase activity wasdetected in the homogenates of leukocytes with the oxy-gen electrode and ascorbic acid (5.6 mmoles/liter).This couple is, therefore, not regarded as a good candi-date for the activity sought.
In red cells NADPH-linked glutathione reductase,coupled to glutathione peroxidase (25), has been con-sidered as an important means of oxidizing NADPHand dealing with H202. This latter is formed by NADHoxidase (8) (Tables I and V). This system has beenstudied extensively the guinea pig PMN.'0 Although theglutathione reductase was present in those cells invery large amounts (many times the activity needed tosatisfy the requirements of the HMP) the peroxidasedid not appear to be present. Addition of GSSGto su-pernatant fractions of guinea pig PMNraised the con-version of glucose carbon-1 to C02 threefold. Further,the oxidation of reduced glutathione was quite rapidin the presence of Hs20, and the addition of peroxide ora peroxide-generating system in the presence of GSHto a homogenate supernatant fraction greatly stimulatedconversion of glucose carbon-1 to C02 (27).'1 The sys-tem was explored in the human PMN.
A potent glutathione reductase was identified in thesupernatant fractions of human leukocytes from fournormal subjects, which could adequately account for the
10Noseworthy, J., and M. L. Karnovsky. In preparation.
696 R. L. Baehner, NV. Gilman, and M. L. Karnovsky

SUCROSE(0.34M)
S G S GFIGURE 1 Distribution of NADHoxidase activity in homogenates of guineapig leukocytes. (Data expressed as percentage of total activity, nanomoles 02hr'.) S indicates soluble fraction, G indicates granule fraction, AIK is "alka-line isotonic KCI."
required reoxidation of NADPHduring phagocytosis,i.e., 898 nmoles NADPHmg' protein hr' (range 564-1284). The average glutathione content of human leu-kocytes from five individuals was 219 nanomoles mg'cell protein (range 151-297). The completion of theglutathione reductase cycle would be dependent upon
an intracellular means of oxidation of the GSHto GSSG,presumably through the agency of H202.
The enzymatic and nonenzymatic rates of GSHoxi-dation were measured with and without the addition ofH302 in Krebs-Ringer phosphate buffer at 370C usingsubstrate concentrations in the range of quantitative de-terminations of GSH and H202 present per milligramcell protein. The estimation of intracellular hydrogenperoxide concentration was derived from the value forthe increment of formate-14C oxidation obtained fromTable I (2.2 nmoles H202 mg-1 protein hr') and fromthe value that 1 ml of packed leukocytes contains ap-
proximately 50 mg protein, i.e., 110 nmoles hydrogenperoxide ml1 packed leukocytes or 110 AM H202. Anamount of peroxide was chosen that was double thisminimum value, which represents (see above) 3% ofthe H202 that might have been derived from the oxygen
uptake during phagocytosis. The rate of nonenzymaticoxidation of GSHover a 15 min period was 10 timesfaster in the presence of 220 AM H202 at pH 5.6 (200nmoles hr') and was 16 times faster at the same H20s
concentration at pH 7.4 (320 nmoles hr') than in theabsence of H202 (20 nmoles hr'). The addition of 0.05mg of leukocyte homogenate protein (granule and super-
natant fractions) in the presence of 1 mmKCN toblock catalase did not accelerate the oxidation of GSH,with 100 AM H202. Therefore, although a glutathioneperoxidase could not be identified, a rapid oxidation of
GSHoccurred, at H202 concentrations lower than thosethat might be present in phagocytizing human leukocytes,which was sufficient to cover the requirements of theHMP.
DIS CUSSION
Guinea pig leukocytes have an oxygen consumption(QO2) that is 2.5 times greater than that of human leu-kocytes either at rest or during phagocytosis. The mean
oxygen uptake of guinea pig leukocytes during phago-cytosis is approximataely 100 nmoles mg' protein hr1greater than that of human leukocytes. This impliesthat the enzyme activity(s) responsible for the cyanide-insensitive respiratory burst that occurs during ingestionmust be greater in the guinea pig than the human leu-kocyte. The differences we noted between the two celltypes mentioned could be due to variation betweenspecies or to the origin of the cells, i.e., peritoneal cavityof guinea pigs and peripheral venous blood of humans.However, it is clear from this and other studies (2, 26,27) that cells of different species and obtained by dif-ferent means from the animal do exhibit grossly similarmetabolic patterns.
There have been reports implicating cyanide insen-sitive oxidases for both reduced di- and reduced tri-phosphopyridine nucleotides as being responsible for thecyanide-insensitive respiratory burst of phagocytosis(4, 9, 10, 22). The cyanide-insensitive NADPHoxidaseactivity is only 10-20% of that needed to account forthe respiratory burst (See Table II) and is, therefore,of questionable importance. However, Zatti and Rossi(9) observed a fivefold increase in NADPHoxidaseactivity in the granule fraction of guinea pig leukocytesafter the cells were treated with saponin or after phago-
Comparative Studies of Metabolism in Leukocytes
100
50
A I K
697

cytosis of heat-killed bacteria. If there were such astimulation of enzyme activity after exposure of the cellsto surface active agents or after phagocytosis, NADPHoxidase could, indeed, be invoked to explain the respira-tory burst. The above authors expressed their resultsas specific activities, i.e., activity per unit of protein.We found in experiments similar to theirs that the in-crease in NADPHoxidase specific activity after ex-posure of the cells to saponin was due to loss of cellprotein into the incubation medium in excess of any lossof NADPHoxidase itself. The total activity of NADPHoxidase of granules isolated from the treated cells didnot change (see Table III). Further, when inert par-ticles (latex spherules) were used instead of bacteriain phagocytosis experiments similar to those of Zattiand Rossi, no increase in granule NADPHoxidase spe-cific activity was noted, even though ingestion of thesespherules produces a normal respiratory burst. It ispossible that in the experiments of Zatti and Rossi withthe phagocytizing cells, some effects not related tophagocytosis per se were introduced, i.e., effects due tostill operative enzymes of the dead bacteria they used ornonspecific effects such as those due to surfactants, en-dotoxins, or other components of the organisms (28-30). These could have produced a situation similar tothat described above for saponin. It thus appears to usthat the respiratory response of phagocytizing PMNcannot be ascribed to the action of an NADPHoxidasebecause there is insufficient enzymatic activity presentat rest or during phagocytosis.
As reported above, there is sufficient NADHoxidaseto match the respiratory burst during phagocytosis es-pecially if the pH within phagocytizing cells is con-sidered. Further, the amount of this enzyme present inguinea pig cells is far greater than in human PMN inline with the oxygen uptake of the intact cells.
The precise location of NADHoxidase within livingleukocytes has, however, not been established. Priorstudies of this enzyme (8) were performed on cellshomogenized in isotonic potassium chloride solution andall of the enzyme activity was soluble. If 0.34 M sucrosewas used as the homogenization medium, about one-halfof the enzyme could be recovered in the granule fraction.The activity can be removed from these granules by abrief exposure to isotonic potassium chloride solution.In the case of the enzyme from human PMN, the great-est proportion (90%) was immediately solubilized byhomogenization in sucrose. The site of action and modeof activation of the NADHoxidase in phagocytizingcells are not known. The binding to granules under someconditions and release under others might offer cluesto the situation in the whole cell at least in the caseof the guinea pig. For example, one might think thatthe enzyme might be granule-associated in the resting
cell. Perhaps the fusion of the granules with the phago-cytic vacuole releases it to the immediately adjacentcytoplasm. The substrate (NADH) would also be ex-pected to be in the cytoplasm. There would then be azone of high activity (NADH oxidase) in the imme-diate region of the phagocytic vacuole producing H202.This H202 would be immediately adjacent to the phago-cytic vacuole, and could diffuse into the latter where itcould interact with myeloperoxidase (released fromgranules into the vacuole) and perhaps a halide to pro-duce the bactericidal activity defined by Klebanoff (31,32) and by McRipley and Sbarra (33). Further studyis certainly needed at the ultrastructural level to un-ravel these events.
In addition to the question of whether sufficient en-zyme is available to meet the cellular requirements, andthat concerning location, we must consider the linkage ofthe respiratory burst to the increase in the HMP. TheHMPhas been shown to produce about two-thirds of thereduced pyridine nucleotide equivalent to the increase inoxygen consumption during phagocytosis (Table I). Thisestimate is based upon the conversion of glucose-1-'4Cand glucose-2-14C to 14CO2 and calculated as two molesof NADPHproduced for each mole of glucose cycledthrough the HMP. This value was calculated from thespecific activity of the exogenous glucose. The specificactivity of intracellular glucose-6-P might well belower, due for instance to the contribution to that poolfrom glycogen. If this were so, an even better stoichio-metric match of the increase in HMPto the increase inoxygen uptake would pertain. We believe, therefore,that the value of "two-thirds" mentioned above is con-servative.
In explaining the linkage between NADHoxidationand the HMP, whose activity depends upon NADP(34), several possibilities exist. Evans and Karnovskyfound a lactate dehydrogenase (LDH) in the guinea pigleukocyte that utilized NADPHat acid pH (15); thatenzyme could link respiration to the HMP by sub-stituting NADPH (which would be converted toNADP) for the NADH normally involved in lac-tate production but which is here oxidized directly byoxygen. In human leukocytes NADPH-linked lactatedehydrogenase does not appear to be present, and ad-ditional explanations for the linkage must be found inthose cells. Evans and Kdplan (35) have described aNADH-NADP+transhydrogenase in human leukocyteswhich could qualify as a link enzyme, but Evans andKarnovsky (15) showed that classical transhydrogenaseis minimal in guinea pig cells and in both species mito-chondria are sparse. A third possible link pathway couldbe via the oxidation and reduction of glutathione.
An important mechanism for the oxidation of reducedglutathione involves hydrogen peroxide. The latter is
698 R. L. Baehner, N. Gilman, and M. L. Karnovsky

produced during phagocytosis. Iyer, Islam, and Quastel(3) were the first to note that this occurred when theyobserved that formate-14C was converted to 14CO2 byingesting PMN. These authors correctly inferred thatthe reaction involved H202 and catalase. However, nodata have been available to indicate what proportion ofthe H202 that results from respiratory activity mightbe involved in this catalatic oxidation of formate. Ifthe entire respiratory burst is via NADHoxidase, H202production would be equal on a molar basis to the oxygenconsumed (8). As noted in Table I, the oxidation offormate by H202 would represent only 3% of the H202formed.
Mills has isolated and purified a glutathione peroxi-dase in the human red cell (25). This enzyme could bea link in the chain of events here, but to date it has notbeen found in the guinea pig or human leukocyte thoughit has been reported to be present in rather low activityin the leukocyte of the rat (27). Even if this enzyme isabsent, the rate of nonenzymatic oxidation of reducedglutathione by H202 should be considered. In the pres-ence of such minimal concentrations of peroxide as arerepresented by the increased conversion of formate toCO (Table I), glutathione is oxidized sufficiently rapidlyto account for the stimulation of the HMP. The involve-ment of glutathione in the latter phenomena involves theNADPH-linked reduction of oxidized glutathione in thepresence of glutathione reductase.
Noseworthy and Karnovsky10 and Reed (27) haveidentified glutathione reductase in the guinea pig andrat leukocyte, respectively. The data in this report sub-stantiate the presence of this enzyme in human leuko-cytes in more than adequate amounts to provide NADPfrom NADPH during phagocytosis. Further, Nose-worthy and Karnovsky10 have stimulated the HMPbyaddition of oxidized glutathione itself to a homogenateof guinea pig leukocytes or by addition of H202 to anorganelle-free supernatant of these cells containing re-duced glutathione. In this species, in contrast to thehuman, regeneration of NADPfrom NADPHmay thusoccur by at least two pathways, viz. the glutathione"cycle," and NADPH-linked lactate dehydrogenase.
In cells from children with chronic granulomatousdisease (CGD), a deficiency of NADHoxidase (andconsequently peroxide production) has been reported(11, 23). The HMP in such cells is also abnormallylow during phagocytosis. In accordance with the ideasexpressed above are the findings of Baehner, Karnovsky,and Nathan (36) who have stimulated the HMP ofCGDleukocytes by allowing them to phagocytize latexparticles coated with glucose oxidase, an H202 gener-ating system. Under such conditions, the HMPwas ele-vated in these cells.
The data that we have presented indicate that anNADH oxidase is probably responsible for the re-spiratory burst during phagocytosis. Further, the prod-uct of this pathway, H202 appears to be important instimulation of the HMPvia the mediation of glutathioneas reported by Reed (27).
The quantitative assessment of the multiple fates ofH202, i.e. the reaction with glutathione, catalase cata-lyzed reactions, or the very important myeloperoxidase-linked reactions that have bactericidal implications (31-33), remains to be decided, as well as the potential roleof such mediators of oxidation-reduction reactions asascorbate.
ACKNOWLEDGMENTSThis research was supported by U. S. Public Health ServiceGrants 1-R01 A108173 and AI-03260.
REFERENCES1. Sbarra, A. J., and M. L. Karnovsky. 1959. The bio-
chemical basis of phagocytosis. I. Metabolic changes dur-ing the ingestion of particles by polymorphonuclearleukocytes. J. Biol. Chem. 234: 1355.
2. Cohn, Z. A., and S. I. Morse. 1960. Functional and meta-bolic properties of polymorphonuclear leucocytes. I. Ob-servation on the requirements and consequences of par-ticle ingestion. J. Exp. Med. 111: 667.
3. Iyer, G. Y. N., D. F. M. Islam, and J. H. Quastel. 1961.Biochemical aspects of phagocytosis. Nature (London).192: 535.
4. Karnovsky, M. L. 1962. The metabolic basis of phago-cytosis. Physiol. Rev. 42: 145.
5. Ohta, H. 1964. Metabolic changes of neutrophile leuko-cytes during phagocytosis. Acta Haematol. Jap. 29: 554.
6. Rossi, F., and M. Zatti. 1966. Effect of phagocytosis onthe carbohydrate metabolism of polymorphonuclear leu-cocytes. Biochim. Biophys. Acta. 121: 110.
7. Baldridge, C. W., and R. W. Gerard. 1933. The extrarespiration of phagocytosis. Amer. J. Physiol. 103: 235.
8. Cagan, R. H., and M. L. Karnovsky. 1964. Enzymaticbasis of the respiratory stimulation during phagocytosis.Nature (London). 204: 255.
9. Zatti, M., and F. Rossi. 1966. Mechanism of the respira-tory stimulation in phagocytosing leukocytes. The KCN-insensitive oxidation of NADPH2. Experientia. 22: 758.
10. Roberts, J., and J. H. Quastel. 1964. Oxidation of re-duced triphosphopyridine nucleotide by guinea pig poly-morphonuclear leukocytes. Nature (London). 202: 85.
11. Holmes, B., A. R. Page, and R. A. Good. 1967. Studiesof the metabolic activity of leukocytes from patients witha genetic abnormality of phagocytic function. J. Clin. In-vest. 46: 1422.
12. Baehner, R. L., and D. G. Nathan. 1967. Leukocyte oxi-dase: defective activity in chronic granulomatous disease.Science (Washington). 155: 835.
13. Stahelin, H. M., M. L. Karnovsky, A. E. Farnham, andE. Suter. 1957. Studies on the interaction between phago-cytes and tubercle bacilli. III. Some metabolic effects inguinea pigs associated with infection with tuberclebacilli. J. Exp. Med. 105: 265.
14. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurements with the Folinphenol reagent. J. Biol. Chem. 193: 265.
Comparative Studies of Metabolism in Leukocytes 699

15. Evans, W. H., and M. L. Karnovsky. 1962. The bio-chemical basis of phagocytosis. IV. Some aspects of car-bohydrate metabolism during phagocytosis. Biochemistry.1:159.
16. Racker, E. R. 1955. Glutathione reductase from bakers'yeast and beef liver. J. Biol. Chem. 217: 855.
17. Damron, C. M., M. M. Monier, and J. H. Roe. 1952.Metabolism of L-ascorbic acid, dehydro-L-ascorbic acid,and diketo-L-gluconic acid in the guinea pig. J. Biol.Chem. 195: 599.
18. Beutler, E., 0. Duron, and B. M. Kelly. 1963. Improvedmethod for the determination of blood glutathione. J.Lab. Clin. Med. 61: 882.
19. Lowry, 0. H., J. A. Lopez, and 0. A. Bessey. 1945. Thedetermination of ascorbic acid in small amounts of bloodserum. J. Biol. Chem. 160: 609.
20. Roe, J. H., and C. A. Kuether. 1943. The determinationof ascorbic acid in whole blood and urine through the2,4-dinitrophenyl hydrazine derivative of dehydroascorbicacid. J. Biol. Chem. 147: 399.
21. Oren, R., A. E. Farnham, K. Saito, E. Milofsky, andM. L. Karnovsky. 1963. Metabolic patterns in three typesof phagocytizing cells. J. Cell. Biol. 17: 487.
22. Rossi, R., and M. Zatti. 1968. Mechanism of the respira-tory stimulation in saponin-treated leucocytes. The KCN-insensitive oxidation of NADPH. Biochirn. Biophys.Acta. 153: 296.
23. Baehner, R. L., and M. L. Karnovsky. 1968. Deficiencyof reduced nicotinamide-adenine dinucleotide oxidase inchronic granulomatous disease. Science (Washington).162: 1277.
24. Crandon, J. H., B. Landau, S. Mikal, J. Balmanno, M.Jefferson, and M. Mahoney. 1958. Ascorbic acid economyin surgical patients as indicated by blood ascorbic acidlevels. N. Engl. J. Med. 258: 105.
25. Mills, G. C. 1959. The purification and properties ofglutathione peroxidase of erythrocytes. J. Biol. Chem.234: 502.
26. Becker, H. J., G. Munder, and H. Fischer. 1958. 'Oberden leukocytocytenstoff wechsel bei der phagocytose.Z. Physiol. Chem. Hoppe-Seylers. 313: 266.
27. Reed, P. W. 1969. Glutathione and the hexose mono-phosphate shunt in phagocytizing and hydrogen peroxide-treated rat leukocytes. J. Biol. Chem. 244: 2459.
28. Strauss, B. S., and C. A. Stetson, Jr. 1960. Studies onthe effect of certain macromolecular substances on therespiratory activity of the leucocytes of peripheral blood.J. Exp. Med. 112: 653.
29. Graham, R. C., Jr., M. J. Karnovsky, A. W. Shafer,E. A. Glass, and M. L. Karnovsky. 1967. Metabolicand morphological observations on the effect of surface-active agents on leukocytes. J. Cell. Biol. 32: 629.
30. Cohn, Z. A., and S. I. Morse. 1960. Functional andmetabolic properties of polymorphonuclear leucocytes.II. The influence of a lipopolysaccharide endotoxin.J. Exp. Med. 111: 689.
31. Klebanoff, S. J. 1968. Myeloperoxidase-halide-hydrogenperoxide antibacterial system. J. Bacteriol. 95: 2131.
32. Klebanoff, S. J., and L. R. White. 1969. Iodination defectin the leukocytes of a patient with chronic granulomatousdisease of childhood. N. Engl. J. Med. 280: 460.
33. McRipley, R. J., and A. J. Sbarra. 1967. Role of thephagocyte in host-parasite interactions. XII. Hydrogenperoxide-myeloperoxidase bactericidal system in thephagocyte. J. Bacteriol. 94: 1425.
34. Beck, W. S. 1958. Occurrence and control of the phos-phogluconate oxidation pathway in normal and leukemicleukocytes. J. Biol. Chem. 232: 271.
35. Evans, A. E. and N. 0. Kaplan. 1966. Pyridine nucleo-tide transhydrogenase in normal human and leukemic leu-kocytes. J. Clin. Invest. 45: 1268.
36. Baehner, R. L., M. L. Karnovsky, and D. G. Nathan.1969. Correction of oxidative deficiencies of leukocytesin chronic granulomatous disease. J. Clin. Invest. 48: Sa.(Abstr.)
700 R. L. Baehner, N. Gilman, and M. L. Karnovsky