patterns of genetic, phenotypic, and acoustic variation
TRANSCRIPT

Ecology and Evolution. 2017;7:2169–2180. | 2169www.ecolevol.org
Received:17August2016 | Revised:30December2016 | Accepted:5January2017DOI:10.1002/ece3.2782
O R I G I N A L R E S E A R C H
Patterns of genetic, phenotypic, and acoustic variation across a chiffchaff (Phylloscopus collybita abietinus/tristis) hybrid zone
Daria Shipilina1 | Maksym Serbyn2 | Vladimir Ivanitskii1 | Irina Marova1 | Niclas Backström3
ThisisanopenaccessarticleunderthetermsoftheCreativeCommonsAttributionLicense,whichpermitsuse,distributionandreproductioninanymedium,providedtheoriginalworkisproperlycited.© 2017 The Authors. Ecology and EvolutionpublishedbyJohnWiley&SonsLtd.
1DepartmentofVertebrateZoology, LomonosovMoscowStateUniversity,Moscow,Russia2DepartmentofPhysics,UniversityofCalifornia,Berkeley,CA,USA3DepartmentofEvolutionaryBiology,EvolutionaryBiologyCentre(EBC),UppsalaUniversity,Uppsala,Sweden
CorrespondenceNiclasBackström,DepartmentofEvolutionaryBiology,EvolutionaryBiologyCentre(EBC),UppsalaUniversity,Uppsala,Sweden.Email:[email protected]
Funding informationRoyalSwedishAcademyofScience(KVA),Grant/AwardNumber:FOA13H-088;Vetenskapsrådet,Grant/AwardNumber:2013-4508;KungligaFysiografiskaSällskapetiLund;HelgeAx:sonJohnsonsFoundation;LarsHiertaMemorialFoundation;RussianFoundationofBasicResearch(RFBR),Grant/AwardNumber:14-04-01259and15-29-02771;RussianScienceFoundation(RSF),Grant/AwardNumber:14-50-00029;GordonandBettyMooreFoundation’sEPiQSInitiativeGrant,Grant/AwardNumber:GBMF4307.
AbstractCharacterizingpatternsofevolutionofgeneticandphenotypicdivergencebetweenincipientspecies isessential tounderstandhowevolutionof reproductive isolationproceeds.Hybridzonesareexcellentforstudyingsuchprocesses,astheyprovideop-portunitiestoassesstraitvariationinindividualswithmixedgeneticbackgroundandtoquantifygeneflowacrossdifferentgenomicregions.Here,wecombineplumage,song,mtDNAandwhole-genomesequencedataandanalyzevariationacrossasym-patriczonebetweentheEuropeanandtheSiberianchiffchaff(Phylloscopus collybita abietinus/tristis)tostudyhowgeneexchangebetweenthelineagesaffectstraitvaria-tion.Ourresultsshowthatchiffchaffwithinthesympatricregionshowmoreextensivetraitvariation thanallopatricbirds,witha largeproportionof individualsexhibitingintermediatephenotypiccharacters.Thegenomicdifferentiationbetweenthesubspe-ciesislowerinsympatrythaninallopatryandsympatricbirdshaveamixofgeneticancestryindicatingextensiveongoingandpastgeneflow.Patternsofphenotypicandgeneticvariationalsovarybetweenregionswithinthehybridzone,potentiallyreflect-ing differences in population densities, age of secondary contact, or differences inmaterecognitionormatepreference.Thegenomicdatasupportthepresenceoftwodistinctgeneticcladescorrespondingtoallopatricabietinusandtristisandthatgeneticadmixture is the forceunderlying traitvariation in thesympatric region—theprevi-ouslydescribedsubspecies(“fulvescens”)fromtheregionisthereforenotlikelyadis-tinct taxon. In addition, we conclude that subspecies identification based onappearance isuncertainasan individualwithanapparentlydistinctphenotypecanhaveaconsiderableproportionofthegenomecomposedofmixedalleles,orevenamajorpartofthegenomeintrogressedfromtheothersubspecies.Ourresultsprovideinsightsintothedynamicsofadmixtureacrosssubspeciesboundariesandhaveimpli-cationsforunderstandingspeciationprocessesandforthe identificationofspecificchiffchaffindividualsbasedonphenotypiccharacters.
K E Y W O R D S
Chiffchaff,geneticdifferentiation,hybridzone,hybridization,introgression,speciation

2170 | SHIPILINA et AL.
1 | INTRODUCTION
Populationdivergenceandspeciationaretheultimatesourcesofbio-diversity,andcharacterizationofpatternsandmechanismsduringthespeciationprocessisessentialtounderstandhowbiodiversityisgen-eratedandmaintained(Schluter,2009).Speciationcanbedrivenbydifferentfactors;aratherdistinctpartitionispresentbetween(1)spe-ciationasaresultofstochasticaccumulationofmutationsthatcon-tribute to reproductive isolation between geographically separatedtaxaand(2)speciationasaresultofdivergentnaturalselection,thatisthatlocaladaptationbyitselfdrivesthespeciationprocess(Bierne,Gagnaire,&David,2013;Coyne&Orr,2004;Nosil&Feder,2012;Nosil, Harmon, & Seehausen, 2009; Price, 2008; Schluter, 2009).Distinguishingbetween these scenariosmakes itpossible tounder-stand themechanisms underlying variation in observed patterns ofbiodiversityacross regions.Hybrid zones,geographic regionswherethereisanoverlapbetweendistributionrangesandgeneflowoccursbetweentaxonomicunits,canserveas“naturallaboratories”allowingforcloseobservationofmicroevolutionaryprocessesthatmayleadtopopulationdifferentiationand,ultimately,speciation(Abbott,Barton,&Good,2016;Barton&Hewitt,1989;Payseur&Rieseberg,2016).InthePalearcticandNearcticregions,suchhybridzonesarenaturallymostoftensecondarycontactzones,theresultsofdistributionrangeexpansionsfollowingthelatestglaciationperiod(Hewitt,2004,2011).Bystudyingthesecondarycontacthybridzonesclosely,wemayinves-tigateifthereareanybarrierstogeneflowbetweenthetaxainvolvedand combine that with information about potential forces behinddivergence,that is iftaxahavedivergedbecauseofrandomacquisi-tionof incompatibility alleles (observed trait differences not relatedtoecology)orifadaptationhasplayedamajorroleinthedivergenceprocess (potential trait differences clearly associated with ecology)(Toews&Irwin,2008).Inaddition,studyingsecondarycontactzonescangive informationabout thedesignationof lineageswithuncleartaxonomicstatusbyallowingforquantificationofgeneticdifferentia-tionandpotentialintrogressionbetweeninvolvedtaxa(Abbottetal.,2016;Delmoreetal.,2015;Payseur&Rieseberg,2016).
TheOldWorldleafwarblergroup(genusPhylloscopus)containsalargesetofspeciesthatexpressconsiderableresemblanceinplumagecolorationandmorphology(delHoyo,Elliot,&Christie,2006).Thelargenumberofspecieswithinthegenusincombinationwithasubstantialvariationinthedegreeofphenotypicdifferentiationbetweenspeciespairshashelpedprovideunderstandingofwhat typesofprematingreproductive isolationbarriers act at different stages of the specia-tionsuccession(Bensch,Grahn,Müller,Gay,&Åkesson,2009;Price,2010).For instance,pioneeringworkfocusingongenus levelphylo-geneticanalyses(Bensch,Irwin,Irwin,Kvist,&Åkesson,2006;Helbigetal.,1996),targetedanalysisofthegeneticunderpinningsofmigra-torybehavior (Bensch,Åkesson,& Irwin,2002;Benschetal.,2009;Lundberg,Åkesson,&Bensch,2011;Lundbergetal.,2013),theroleofsex-chromosomesindrivingdivergence(Helbig,Salomon,Bensch,&Seibold,2001)andgenomicdifferentiationbetweenpopulationsofa“ringspecies”(Alcaide,Scordato,Price,&Irwin,2014;Irwin,Alcaide,
Delmore, Irwin, & Owens, 2016) has increased the understandingof the forces underlying reproductive isolation between some taxawithinthegenus,butwearestillfarfromadetailedunderstandingofthemechanismsunderlyingthedivergenceprocesses.
The chiffchaff superspecies complex contains four distinct spe-ciesandasetofsubspecieswidelydistributedwithin thePalearcticregion (Figure S1). The Canarian chiffchaff (P. canariensis), includ-ingsubspeciescanariensisand,asofnowextinct,exsul, inhabits theCanaryIslands.ThedistributionrangeoftheIberianchiffchaff(P. iberi-cus)spanstheIberianPeninsulaandthePyreneesandsouthwesternFrance.Themountainchiffchaff (P. sindianus)breeds inhigh-altitudehabitatsintheCaucasusMountains(P. s. lorenzii)andinpartsofeast-ern–centralAsia (P. s. sindianus). The common chiffchaff (P. collybita)has traditionally been separated into six different subspecies: colly-bita(distributedthroughoutsouthernandcentralEurope),brevirostris (northern Asia Minor), caucasicus (lowlands in Caucasus),menzbieri (KopetDaghandsouthwesternTranscaspia),abietinus(north-easternEurope to the Ural Mountains), and tristis (Ural Mountains to eastSiberia;Stepanyan,1990;Svensson,1992).However,thetaxonomicstatusoftristishasbeenatopicofsomedebateoverwhetherornotitshouldbedesignatedfullspeciesstatus(delHoyoetal.,2006).Inthisstudy,wewill refertobothabietinusandtristisassubspeciesofthecommonchiffchaff(P. collybita).
In some areas of the superspecies distribution range, chiffchaffspecies and/or subspeciesoccur in sympatry.The largest such zonewasidentifiedalreadyinthelatenineteenthcentury(Sushkin,1897).Thisregionislikelyasecondarycontactzoneformedduringtherecol-onizationfromglacialrefugiafollowingtheretractionoftheicecoverafterthelatestglaciationperiod,butithasstillnotbeendescribedinmuchdetail.ThezonemoreorlesscoincideswiththeUralMountainsall theway from the southern edge to theWhite Sea (Komarova&Shipilina, 2010; Marova, Fedorov, Shipilina, & Alekseev, 2009) andincludes two subspecies of the common chiffchaff: the “Siberian”chiffchaff(P. c. tristis)andthe“European”commonchiffchaff(P. c. abi-etinus)–twosubspecieswithdifferencesinhabitatpreference,bodysize,plumagecharacteristics,songtype,andmigrationroutes/winter-ingquarters (Dean&Svensson,2005;Helbigetal.,1996;Marova&Alekseev,2008;Ticehurst,1938).Theinterindividualvariationinplum-agecoloration,bodysize,andsongtypeismorepronouncedwithinthesympatricregionthanwithinallopatricpopulationsofeitherabietinus or tristis.Inparticular,someindividualsperformingintermediatesong,so-calledmixedsingers(Lindholm,2008;Marova&Leonovich,1993)andintermediateplumagephenotypeshavepreviouslybeenidentifiedinthesympatriczone(Marovaetal.,2009).Thisparticularregionhasalsobeenthoughttobe inhabitedbyathirdsubspecies,“P. c. fulves-cens”, expressing intermediate characters (plumage phenotype andsongtype)tothoseobservedinabietinusandtristis(Dean&Svensson,2005; Severtsov, 1873; Stepanyan, 1990).However, a set of recentstudies are not consistentwith this hypothesis: aweak associationbetweensongtype,plumagecolor,andmitochondrialhaplotypehasbeenfoundwithinthesympatricregionbutaconsiderableproportionofbirdsdeviatedfromthepattern(Marova&Alekseev,2008;Marova,Shipilina,Fedorov,& Ivanitskii,2013;Marovaetal.,2009).This is in

| 2171SHIPILINA et AL.
agreement with recurrent intercrossing between subspecies in theregion.Infurthersupportofthat,asmall-scalegeneticanalysisusingrestriction enzymedigestionof a singlemitochondrial gene showedhaplotype differentiation between abietinus and tristis (Helbig etal.,1996;Marova etal., 2009), but birdswith distinct abietinus pheno-typesometimescarriedthetristismitochondrialhaplotypeorhadthetypicaltristissongdialectandviceversa(Marovaetal.,2009,2013).However, information from a single, maternally inherited geneticmarkerdoesnotnecessarilyprovideconclusiveevidence forgeneticadmixtureandruleoutthepresenceofathirdsubspecies.Tomakeamoredetailedinvestigationofthegenomicconsequencesofadmixturebetweenabietinusandtristisinregionsofsympatryandtoultimatelytestthevalidityofthepreviouslydescribed“fulvencens”subspecies,wecombinepreviouslyavailabledataonmorphology,acoustics,andmito-chondrialDNA(Marovaetal.,2009,2013)withnovelwhole-genomesequence data.We hypothesize that reproductive barriers betweenabietinus and tristis are incomplete and that genetic admixture hasgivenrisetotheintermediatephenotypesfrequentlyobservedwithinthesympatriczone. Inaddition,totest if relativepopulationdensityandtimesincerecolonizationmayaffectpatternsofgeneticvariationinintermixedpopulations,weanalyzesamplesfromtwodistinctgeo-graphiclocationsthatdifferinrelativedensitiesofabietinusandtristis andindistancetopotentialglacialrefugia:thesouthernmost(southernUralmountains,predominantlymixed forest,closer toglacial refuge)and northernmost (Arkhangelsk region, boreal forest, further awayfromglacialrefuge)partsofthesympatriczone.
2 | METHODS
2.1 | Capture of specimens and DNA extractions
Activechiffchaffmaleswerecapturedbyplaybackandmistnettingduringbreedingseasonsof2007–2009(seeFigureS1forsamplinglocationsandsamplesizes).Thedatawerecollected inthefollow-ingway: first,male songwas recorded for at least threeminutes.Then, individualswerecaught inamistnetbysoundtrapping.Wequantifiedplumagecoloration(morphotype)andtookabloodsam-ple.Thekeyfor themorphological identificationof thesubspeciesisyellow(lipochrome)colorationintensityontheventralsideofthebody: throat, breast, and belly (Svensson, 1992; Ticehurst, 1938).Individualswereclassifiedasabitinusiftheyhadbrightyellowstripesonthebreastandyellowandgreentonesinoverallcolorationandastristisiftheyshowedabsenceofanyyellowhuesincolorationexceptontheunderwing(Svensson,1992;Ticehurst,1938).Birdsclassifiedasintermediatehadyellowishfeatherstovaryingextents,butalwaysclearlylessthanabietinusormorethantristis(Marovaetal.,2009).Intotal,245individualswereincludedinthisstudy;however,duetospecificsofthesamplingprocess,wewerenotabletocharacterizeallofthemforallsetsofparameters:morphotype,acoustics,mtDNAhaplotype,andnuclearSNPs.Of the245sampled individuals,228birdswerecharacterizedasabietinus, tristis, or intermediatebasedon morphological characters (Marova & Alekseev, 2008; Marovaetal.,2009).
For 138 birds,we could record the song andmake high-qualitysonogramsforquantificationofacousticpatterns(Marova&Alekseev,2008;Marovaetal.,2009).Songsoftristisandabietinuscanbeeasilydistinguishedbyearorbysonogramanalysisduetosignificantdiffer-encesintempoandstructure(Ticehurst,1938;Marovaetal.,2009).Acleardistinctionbetweensongtypesisthepresenceofascendingnotesinthetypicaltristissongandtheabsenceofsuchnotesinthesongofabietinus,theconsiderablyshorterintervalbetweennotesintristis(4.7–7.2notespersecond)thaninabietinus(2.8–3.3notespersecond), and the almost nonoverlapping frequency ranges of notes(abietinus: 3.7–4.6kHz; tristis: 2.9–3.7kHz). To designate recordedbirdstoaspecificsongtype,wecalculatedthepercentageofascend-ingelements inthreerandomlyselectedthree-second long intervalsofsongsofeachrecordedmale(visualanalysisofspectrograms)andmeasuredthefrequencyrangeandnoterateusingSyrinx2.3(J.Burt,Seattle,WA,USA).
Bloodsampleswerecollectedfromspecimensbycarefulpiercingof thebrachialveinusing standard syringes. In total, blood samplesfrom197of the245chiffchaffswere collectedand storedonfilterpaper.Samplingsitesweredistributedbothwithintheallopatricrangesofabietinus (n=16)andtristis (n=37)andwithinboththenorthern(intotal67samples;basedonplumagecharacters,thesewere24tristis,18 abietinus,and21withintermediateplumagephenotype,for4birdsmorphologywasnotdetermined)andthesouthern(intotal125sam-ples;41tristis,44abietinus,and27withintermediatephenotype,for13birdsphenotypewasnotdetermined)partofthesympatriczone(FigureS1).All samples fromthenorthernsympatriczonearenoveltothisstudywhilethemajorityofthedatafromallopatricregionsandthesouthernsympatriczonehavebeenusedintwopreviousstudiesofplumage,song,andmtDNAvariation(Marovaetal.,2009,2013).Intotal,therewere105individualsthatwereanalyzedusingallthreecharacteristics:morphotype,songtype,andmtDNAhaplotype.DNAfromwhole blood sampleswas extracted using a standard phenol–chloroformprotocol(Sambrook,Fritsch,&Maniatis,1989).Thequan-tityandqualityoftheDNApreparationswasassessedwithagarosegel (1%) electrophoresis andmeasurementwith theQubit (ThermoFisherScientificInc.)andtheNanoDrop(ThermoFisherScientificInc.)instruments.
2.2 | mtDNA analyses
Onehundredandninety-twobirdscouldbescoredformtDNAhap-lotype at a region in the Cytochrome B (CytB) locus by restrictionanalysis.Weanalyzed the same389-bp-long region thathasprevi-ouslybeenpublished for thechiffchaff (GenBankaccessionentries:Z73479.1,Z73482.1).WithinthisregionofCytB,fiveSNPshavepre-viouslybeenfoundtobediagnosticbetweenallopatricEuropeanandSiberian chiffchaff (Helbig etal., 1996). For restriction analysis, wechosetwoofthosediagnosticSNPswhichallowedustodistinguishbetweendistinctallopatrichaplotypesandathirdSNPthatmade itpossibletoidentifythepresenceofapreviouslyidentifiedhaplotypethatalsohasbeenfoundtobeuniquetotristisandthatdifferstothemostcommontristishaplotypeatasinglenucleotideposition(Marova

2172 | SHIPILINA et AL.
etal.,2009).Restrictionanalysiswasperformedusingtheendonucle-aseHinfI(GANTC).Foreachsample,weused0.3μlofHinf1mixedwithBufferR(1μl)andbi-distilledwater(4μl),and5μlofextractedDNAwasaddedtothemix.Therestrictionreagentmixwasheldat37°Cfor12hr,andfragmenttypeswerescoredbyregularpolyacryla-midegelelectrophoresis.AllscoredmtDNAvariantshavebeensub-mittedtodatadryad.organdareavailableunderdoi:10.5061/dryad.g74v0.
2.3 | Whole- genome sequencing and SNP detection
Ofthe223sampledspecimensthatwerescoredforplumagecolor,we selected 10 individuals with the highest DNA yield from eachsubspeciesandeacharea (i.e.,10allopatric tristis, 10allopatricabi-etinus, 10 sympatric tristis, and 10 sympatric abietinus) and pre-pared standard insert size (380bp) libraries for 100bp paired-endsequencing on the Illumina HiSeq2000 platform. Library prepara-tion and sequencing was performed by the SNP&SEQ TechnologyPlatform inUppsala. The sequencingyieldwas in total 147Gbandvariedbetween2.7and5.1Gbforeachsample.Therangefor indi-vidualsampleswas1.6–4.3GbafterfilteringoutbaseswithQ-scores<30usingConDeTri(Smeds&Künstner,2011),intotal113Gb.Thequality-filterednuclearsequencereadsweremappedtotheFicedula albicollis genome sequence (Ellegren etal., 2012; Kawakami etal.,2014)usingtheBurrows–Wheeleralgorithmas implemented inthesoftwareBWA(version0.5.9,Li&Durbin,2010).SAMtools(Lietal.,2009)wasusedtoconvertfilestoBAMformatandtovisuallyinspectsomeofthealignments.Wesubsequentlymergedallmappedreadsfrom individual samples into a single BAM file using Picard Tools(http://broadinstitute.github.io/picard/). We applied the methodsin the packageGATK (McKenna etal., 2010) for nucleotide qualityscorerecalibration,localinsertion/deletionrealignmentandremovalofduplicates,andcalledSNPsacrossallsamplesusingfilteringparam-etersandSNPqualityrecalibrationrecommendedintheGATKBestPractices(DePristoetal.,2011;VanderAuweraetal.,2013),withoutatrainingsetasnogenomicSNPdatawerepreviouslyavailableforthesesubspecies.Specifically,thiswasperformedintwosteps:first,the RealignerTargetCreator and IndelRealigner packageswere usedforlocalrealignmenttopurgereadsthatmighthavebeenerroneouslymapped, and then, theUnifiedGenotyper packagewas used to callhigh-qualitySNPs(criteria:Q-score>30,minimumcoverage5readsperSNPper individual)andsave these invcf formatfiles.TheSNPcallingprocedureresultedin4.9millionhigh-qualitySNPsthatcouldbeusedfordownstreamanalysis(FigureS2).
2.4 | Analyses of genomic SNP variation
Processingofvcffileswasperformed in thepackagePyVCF in thePythonenvironment (https://github.com/jamescasbon/PyVCF).Twofurtherfilteringstepswereperformedtogeneratethefinaldatasetsusedforanalysis.WeinitiallyappliedstringentcriteriaandselectedasetofSNPs,inwhichwehadhigh-qualitySNPscoreinformationfromat least 9 of the 10 individuals from each population, respectively.
After this filtering step, only 18,014 SNPs were retained—this isfromhereonreferredtoasthe“high-stringencydataset”.Thishigh-stringencydatasetwassubsequentlyusedtoquantifyandvisualizepatternsofdifferentiationbetweenpopulationsbutdidnotcontainmorethan50fixeddifferences(50fixeddifferences/18,014SNPsintotal=0.3%)betweenabietinus and tristis. Toobtain a larger setofpolymorphismsforassessmentofratesoffixed,private,andsharedpolymorphismsbetweenandwithinabietinusandtristis,respectively,andforanalysisofgenomiccompositionofbirdssampledinthesym-patricregion,weselectedsitesthatwerepresentinatleastsixdiffer-ent individuals inbothallopatricabietinusandallopatrictristis.Afterthatfilteringstep,1,233,236high-qualitySNPswereretained—fromhereonreferredtothe“low-stringencydataset”.Itshouldbenotedthat the relative frequency of fixed differences did not change forthe lowerstringencydataset (3,555fixedSNPs/1,233,236SNPs intotal=0.3%). All genotypes have been submitted to datadryad.organdareavailableunderdoi:10.5061/dryad.g74v0.
We used in-house developed Python scripts (available uponrequest) to calculate the number of fixed substitutions betweenallopatricpopulations,aswellasprivateandsharedpolymorphismswithin populations using the larger set of SNPs (1,233,236 SNPs).Fixation indices (FST) were calculated for both allopatric and sym-patric population comparisons (allopatric abietinus versus allopat-ric tristis and sympatricabietinus versus sympatric tristis) using thehigh-stringencydataset(18,014SNPs)andthepackagePopGenome(http://popgenome.weebly.com) in the R environment (Pfeifer,Wittelsbuerger, Ramos-Onsins, & Lercher, 2014). Error estimateswere generated by jackknife resampling over sets of 50 SNPs. Inaddition to overall FST-estimates, the degree of genetic differenti-ationbetween all 40 samples thatwerewhole-genome sequencedgrouped as (1) allopatricabietinus, (2) allopatric tristis, (3) southernsympatricsamples (sample id:sS1-S10),and (4)northernsympatricsamples (sample id:sN1-N10),respectively)wasvisualizedbyusingaprincipalcomponentanalysisofgeneticvariationas implementedinthepackagesmartPCA(EigenSoft)forR(Patterson,Price,&Reich,2006).Preparationofdatafilesforthatanalysiswascarriedoutusingthe program PyVCF (https://github.com/jamescasbon/PyVCF), andgraphswereconstructedusingtheggplot2 library inR. Inaddition,weappliedapopulationassignmentanalysisbasedonallelefrequen-ciesacrossthehigh-stringencySNPsasimplementedintheprogramSTRUCTURE 2.3.4 (Pritchard, Stephens, & Donnelly, 2000) usingthe admixturemodel.We ran Structure for 50,000 iterations afteraburn-inof20,000iterations,fivetimesforeachofsevenKvalues(K = 1 to K=7populations)andevaluatedtheoptimalKasdescribedinEvannoetal.(2005).AllfiveindependentrunsforeachK resulted inconsistentresults.AsummaryofalldatatreatmentsandanalyticalstepsforeachdatasetispresentedinFigureS2.
3 | RESULTS
Within the allopatric regions, all birds were scored as typical abi-etinus or tristis, respectively,both regardingplumagecharacteristics
| 2173SHIPILINA et AL.
andsongtype.Inthesympatricregions,thevariationinplumageandsong typewasgreaterwithseveral individuals scoredas intermedi-ate/mixedphenotypes(summarizedinTable1).Specifically,wefoundthat33%ofallsampledindividualshadintermediateplumagecharac-ters(onlypaleyellowhueandnegligibleamountofyellowfeathersonventralside)inthenorthernsympatricregion,andthecorrespondingvalueforthesouthernsympatricregionwas24%.Forthedistributionofsongtypes,therewasagainaconsiderableproportionofindividu-alsinthesympatricregionsperformingmixedsong—thatisamixtureoftypicaltristisandabietinusnotesandintermediatefrequencyrangeandsongspeed(notespertimeunit)—63%inthenorthernand40%inthesouthernsympatricregion.
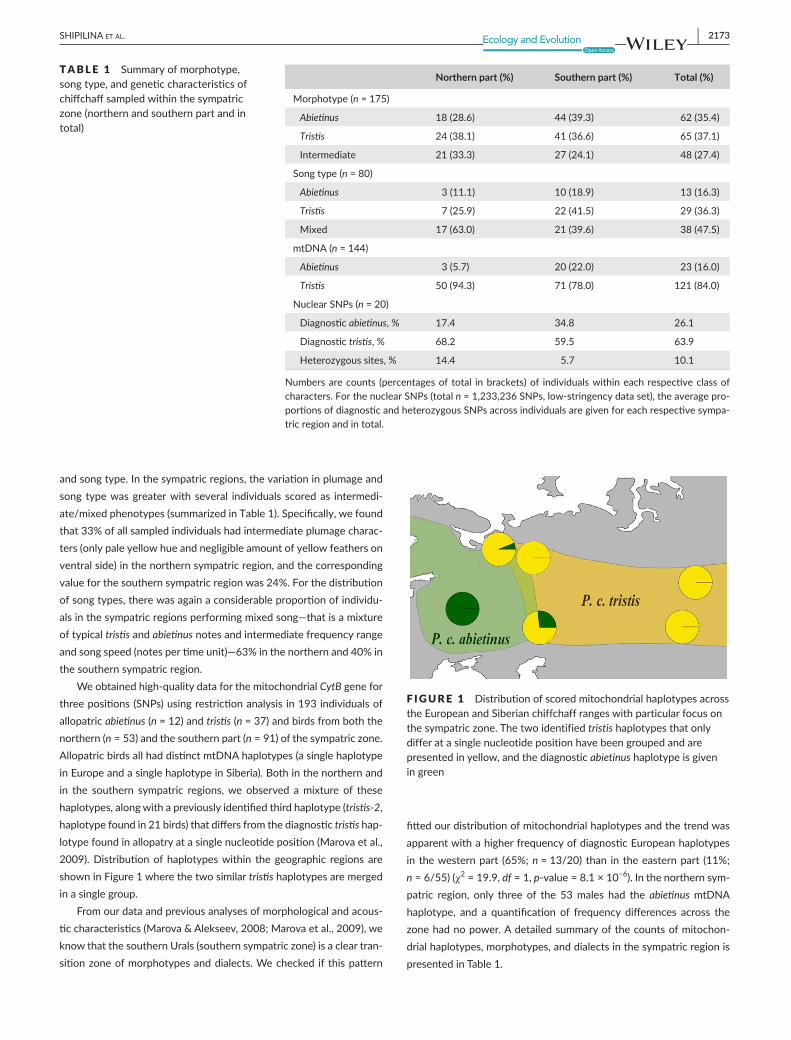
Weobtainedhigh-qualitydataforthemitochondrialCytBgeneforthreepositions(SNPs)usingrestrictionanalysisin193individualsofallopatricabietinus (n=12)andtristis (n=37)andbirdsfromboththenorthern(n=53)andthesouthernpart(n=91)ofthesympatriczone.AllopatricbirdsallhaddistinctmtDNAhaplotypes(asinglehaplotypeinEuropeandasinglehaplotypeinSiberia).Bothinthenorthernandin the southern sympatric regions,we observed amixture of thesehaplotypes,alongwithapreviouslyidentifiedthirdhaplotype(tristis-2,haplotypefoundin21birds)thatdiffersfromthediagnostictristishap-lotypefoundinallopatryatasinglenucleotideposition(Marovaetal.,2009).Distributionofhaplotypeswithin thegeographic regionsareshowninFigure1wherethetwosimilartristishaplotypesaremergedinasinglegroup.
Fromourdataandpreviousanalysesofmorphologicalandacous-ticcharacteristics(Marova&Alekseev,2008;Marovaetal.,2009),weknowthatthesouthernUrals(southernsympatriczone)isacleartran-sitionzoneofmorphotypesanddialects.Wechecked ifthispattern
fittedourdistributionofmitochondrialhaplotypesandthetrendwasapparentwithahigherfrequencyofdiagnosticEuropeanhaplotypesinthewesternpart (65%;n=13/20)than intheeasternpart (11%;n=6/55)(χ2=19.9,df=1,p-value=8.1×10−6).Inthenorthernsym-patric region, only three of the 53males had the abietinusmtDNAhaplotype, and a quantification of frequency differences across thezonehadnopower.Adetailedsummaryofthecountsofmitochon-drialhaplotypes,morphotypes,anddialectsinthesympatricregionispresentedinTable1.
Northern part (%) Southern part (%) Total (%)
Morphotype(n=175)
Abietinus 18(28.6) 44(39.3) 62(35.4)
Tristis 24(38.1) 41(36.6) 65(37.1)
Intermediate 21(33.3) 27(24.1) 48(27.4)
Songtype(n=80)
Abietinus 3(11.1) 10(18.9) 13(16.3)
Tristis 7(25.9) 22(41.5) 29(36.3)
Mixed 17(63.0) 21(39.6) 38(47.5)
mtDNA(n=144)
Abietinus 3(5.7) 20(22.0) 23(16.0)
Tristis 50(94.3) 71(78.0) 121(84.0)
NuclearSNPs(n=20)
Diagnosticabietinus,% 17.4 34.8 26.1
Diagnostictristis,% 68.2 59.5 63.9
Heterozygoussites,% 14.4 5.7 10.1
Numbersarecounts (percentagesoftotal inbrackets)of individualswithineachrespectiveclassofcharacters.ForthenuclearSNPs(totaln=1,233,236SNPs,low-stringencydataset),theaveragepro-portionsofdiagnosticandheterozygousSNPsacrossindividualsaregivenforeachrespectivesympa-tricregionandintotal.
TABLE 1 Summaryofmorphotype,songtype,andgeneticcharacteristicsofchiffchaffsampledwithinthesympatriczone(northernandsouthernpartandintotal)
F IGURE 1 DistributionofscoredmitochondrialhaplotypesacrosstheEuropeanandSiberianchiffchaffrangeswithparticularfocusonthesympatriczone.Thetwoidentifiedtristishaplotypesthatonlydifferatasinglenucleotidepositionhavebeengroupedandarepresentedinyellow,andthediagnosticabietinushaplotypeisgiveningreen
2174 | SHIPILINA et AL.
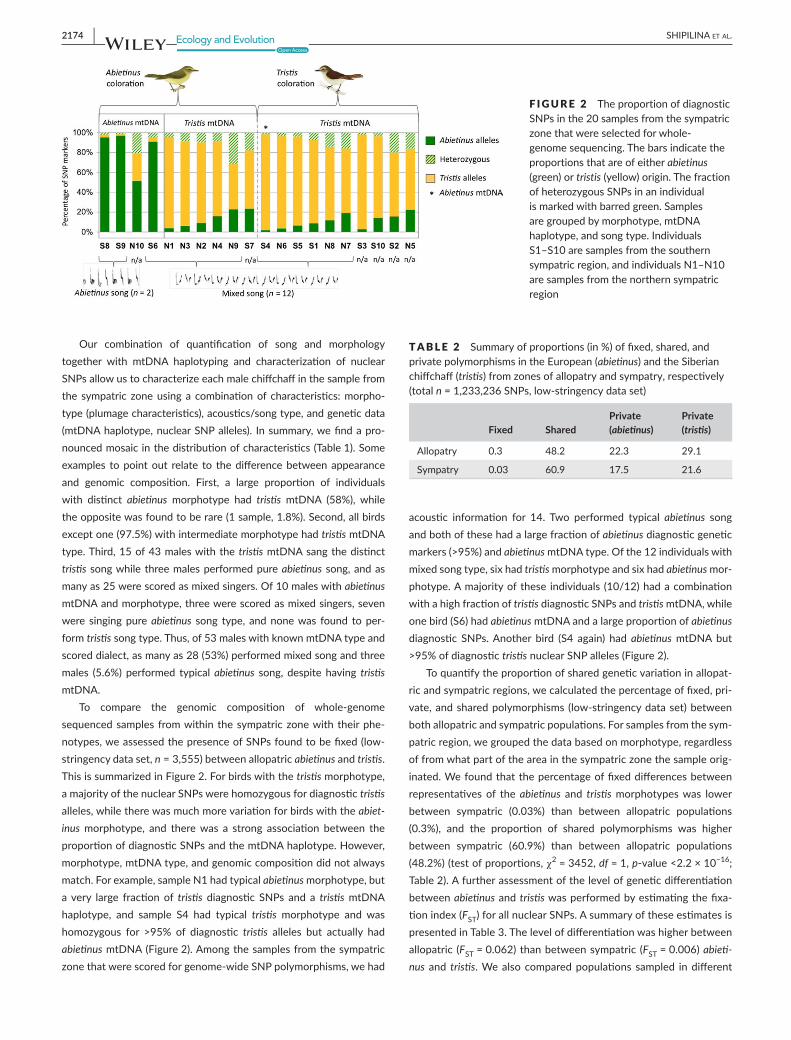
Our combination of quantification of song and morphologytogether with mtDNA haplotyping and characterization of nuclearSNPsallowustocharacterizeeachmalechiffchaffinthesamplefromthe sympatric zoneusinga combinationof characteristics:morpho-type(plumagecharacteristics),acoustics/songtype,andgeneticdata(mtDNAhaplotype,nuclearSNPalleles). Insummary,wefindapro-nouncedmosaicinthedistributionofcharacteristics(Table1).Someexamples topointout relate to thedifferencebetweenappearanceand genomic composition. First, a large proportion of individualswith distinct abietinus morphotype had tristis mtDNA (58%), whiletheoppositewasfoundtoberare(1sample,1.8%).Second,allbirdsexceptone(97.5%)withintermediatemorphotypehadtristismtDNAtype.Third,15of43maleswiththetristismtDNAsangthedistincttristissongwhilethreemalesperformedpureabietinussong,andasmanyas25werescoredasmixedsingers.Of10maleswithabietinus mtDNAandmorphotype,threewerescoredasmixedsingers,sevenwere singingpureabietinus song type, andnonewas found toper-formtristissongtype.Thus,of53maleswithknownmtDNAtypeandscoreddialect,asmanyas28(53%)performedmixedsongandthreemales (5.6%) performed typicalabietinus song, despite having tristis mtDNA.
To compare the genomic composition of whole-genomesequencedsamples fromwithinthesympatriczonewiththeirphe-notypes,weassessedthepresenceofSNPsfoundtobefixed(low-stringencydataset,n=3,555)betweenallopatricabietinusandtristis. ThisissummarizedinFigure2.Forbirdswiththetristismorphotype,amajorityofthenuclearSNPswerehomozygousfordiagnostictristis alleles,whiletherewasmuchmorevariationforbirdswiththeabiet-inusmorphotype, and therewas a strong association between theproportionofdiagnosticSNPsandthemtDNAhaplotype.However,morphotype,mtDNAtype,andgenomiccompositiondidnotalwaysmatch.Forexample,sampleN1hadtypicalabietinusmorphotype,butavery large fractionof tristis diagnostic SNPs and a tristismtDNAhaplotype, and sample S4 had typical tristis morphotype and washomozygous for >95% of diagnostic tristis alleles but actually hadabietinusmtDNA(Figure2).Amongthesamplesfromthesympatriczonethatwerescoredforgenome-wideSNPpolymorphisms,wehad
acoustic information for 14. Two performed typical abietinus songandbothofthesehadalargefractionofabietinusdiagnosticgeneticmarkers(>95%)andabietinusmtDNAtype.Ofthe12individualswithmixedsongtype,sixhadtristismorphotypeandsixhadabietinusmor-photype.Amajorityofthese individuals (10/12)hadacombinationwithahighfractionoftristisdiagnosticSNPsandtristismtDNA,whileonebird(S6)hadabietinusmtDNAandalargeproportionofabietinus diagnostic SNPs.Another bird (S4 again) hadabietinusmtDNAbut>95%ofdiagnostictristisnuclearSNPalleles(Figure2).
Toquantifytheproportionofsharedgeneticvariationinallopat-ricandsympatricregions,wecalculatedthepercentageoffixed,pri-vate,andsharedpolymorphisms (low-stringencydataset)betweenbothallopatricandsympatricpopulations.Forsamplesfromthesym-patricregion,wegroupedthedatabasedonmorphotype,regardlessoffromwhatpartoftheareainthesympatriczonethesampleorig-inated.Wefoundthatthepercentageoffixeddifferencesbetweenrepresentativesof theabietinus and tristismorphotypeswas lowerbetween sympatric (0.03%) than between allopatric populations(0.3%), and the proportion of shared polymorphisms was higherbetween sympatric (60.9%) than between allopatric populations(48.2%)(testofproportions,χ2=3452,df=1,p-value<2.2×10−16; Table2).Afurtherassessmentofthelevelofgeneticdifferentiationbetweenabietinusandtristiswasperformedbyestimatingthefixa-tionindex(FST)forallnuclearSNPs.AsummaryoftheseestimatesispresentedinTable3.Thelevelofdifferentiationwashigherbetweenallopatric (FST=0.062)thanbetweensympatric (FST=0.006)abieti-nus and tristis.Wealsocomparedpopulationssampled indifferent
F IGURE 2 TheproportionofdiagnosticSNPsinthe20samplesfromthesympatriczonethatwereselectedforwhole-genomesequencing.Thebarsindicatetheproportionsthatareofeitherabietinus (green)ortristis(yellow)origin.ThefractionofheterozygousSNPsinanindividualismarkedwithbarredgreen.Samplesaregroupedbymorphotype,mtDNAhaplotype,andsongtype.IndividualsS1–S10aresamplesfromthesouthernsympatricregion,andindividualsN1–N10aresamplesfromthenorthernsympatricregion
TABLE 2 Summaryofproportions(in%)offixed,shared,andprivatepolymorphismsintheEuropean(abietinus)andtheSiberianchiffchaff(tristis)fromzonesofallopatryandsympatry,respectively(totaln=1,233,236SNPs,low-stringencydataset)
Fixed SharedPrivate (abietinus)
Private (tristis)
Allopatry 0.3 48.2 22.3 29.1
Sympatry 0.03 60.9 17.5 21.6
| 2175SHIPILINA et AL.
partswithinthesympatriczone.Weobservedaslightlylowerlevelof genetic differentiation between sympatric abietinus and tristis populationswithinthesouthernregion(FST=0.013)thanwithinthenorthernregion(FST=0.016).
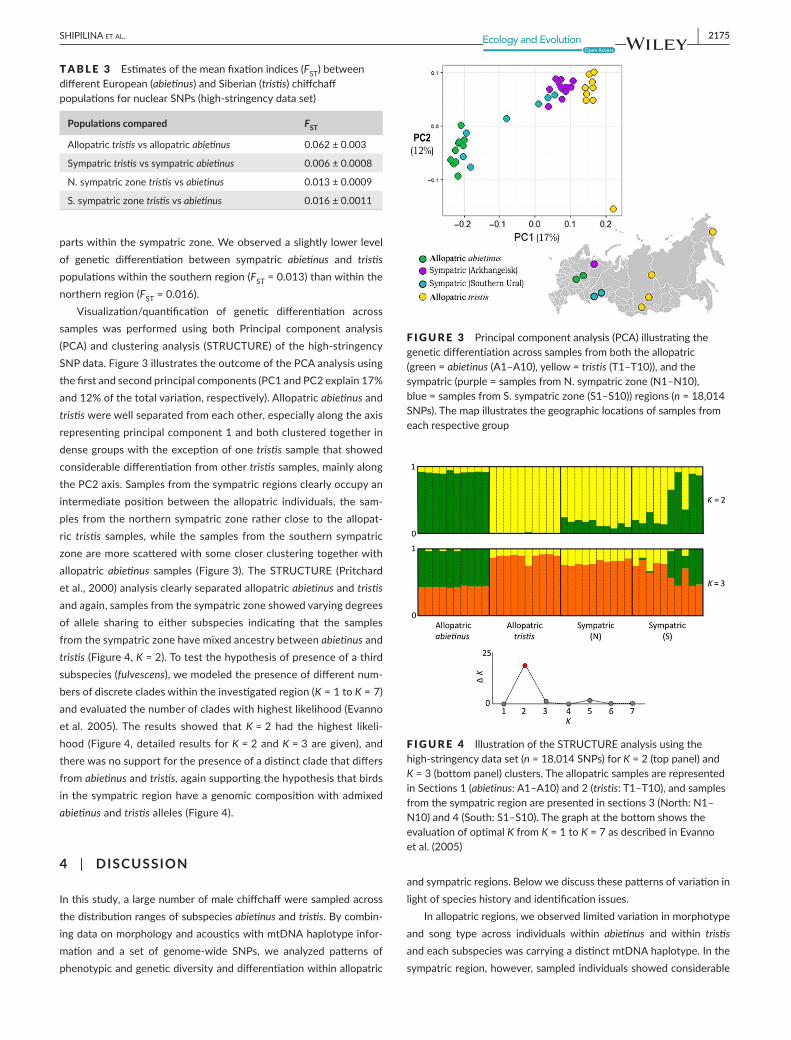
Visualization/quantification of genetic differentiation acrosssamples was performed using both Principal component analysis(PCA)andclusteringanalysis (STRUCTURE)ofthehigh-stringencySNPdata.Figure3illustratestheoutcomeofthePCAanalysisusingthefirstandsecondprincipalcomponents(PC1andPC2explain17%and12%ofthetotalvariation,respectively).Allopatricabietinusandtristiswerewellseparatedfromeachother,especiallyalongtheaxisrepresentingprincipalcomponent1andbothclusteredtogetherindensegroupswiththeexceptionofonetristissamplethatshowedconsiderabledifferentiationfromothertristissamples,mainlyalongthePC2axis.Samplesfromthesympatricregionsclearlyoccupyanintermediatepositionbetween theallopatric individuals, the sam-plesfromthenorthernsympatriczoneratherclosetotheallopat-ric tristis samples,while the samples from the southern sympatriczonearemorescatteredwithsomecloserclusteringtogetherwithallopatricabietinus samples (Figure3).TheSTRUCTURE (Pritchardetal.,2000)analysisclearlyseparatedallopatricabietinusandtristis andagain,samplesfromthesympatriczoneshowedvaryingdegreesof allele sharing to either subspecies indicating that the samplesfromthesympatriczonehavemixedancestrybetweenabietinusandtristis(Figure4,K=2).Totestthehypothesisofpresenceofathirdsubspecies(fulvescens),wemodeledthepresenceofdifferentnum-bersofdiscretecladeswithintheinvestigatedregion(K = 1 to K =7)andevaluatedthenumberofcladeswithhighestlikelihood(Evannoetal. 2005).The results showed thatK=2 had the highest likeli-hood(Figure4,detailedresultsforK=2andK=3aregiven),andtherewasnosupportforthepresenceofadistinctcladethatdiffersfromabietinusandtristis,againsupportingthehypothesisthatbirdsinthesympatricregionhaveagenomiccompositionwithadmixedabietinusandtristisalleles(Figure4).
4 | DISCUSSION
Inthisstudy,alargenumberofmalechiffchaffweresampledacrossthedistributionrangesofsubspeciesabietinusandtristis.Bycombin-ingdataonmorphologyandacousticswithmtDNAhaplotypeinfor-mation and a set of genome-wide SNPs, we analyzed patterns ofphenotypicandgeneticdiversityanddifferentiationwithinallopatric
andsympatricregions.Belowwediscussthesepatternsofvariationinlightofspecieshistoryandidentificationissues.
Inallopatricregions,weobservedlimitedvariationinmorphotypeand song type across individuals within abietinus and within tristis andeachsubspecieswascarryingadistinctmtDNAhaplotype.Inthesympatricregion,however,sampledindividualsshowedconsiderable
TABLE 3 Estimatesofthemeanfixationindices(FST)betweendifferentEuropean(abietinus)andSiberian(tristis)chiffchaffpopulationsfornuclearSNPs(high-stringencydataset)
Populations compared FST
Allopatrictristisvsallopatricabietinus 0.062±0.003
Sympatrictristisvssympatricabietinus 0.006 ± 0.0008
N.sympatriczonetristis vs abietinus 0.013±0.0009
S.sympatriczonetristis vs abietinus 0.016 ± 0.0011
F IGURE 3 Principalcomponentanalysis(PCA)illustratingthegeneticdifferentiationacrosssamplesfromboththeallopatric(green=abietinus(A1–A10),yellow=tristis(T1–T10)),andthesympatric(purple=samplesfromN.sympatriczone(N1–N10),blue=samplesfromS.sympatriczone(S1–S10))regions(n=18,014SNPs).Themapillustratesthegeographiclocationsofsamplesfromeachrespectivegroup
F IGURE 4 IllustrationoftheSTRUCTUREanalysisusingthehigh-stringencydataset(n=18,014SNPs)forK=2(toppanel)andK=3(bottompanel)clusters.TheallopatricsamplesarerepresentedinSections1(abietinus:A1–A10)and2(tristis:T1–T10),andsamplesfromthesympatricregionarepresentedinsections3(North:N1–N10)and4(South:S1–S10).ThegraphatthebottomshowstheevaluationofoptimalKfromK = 1 to K=7asdescribedinEvannoetal.(2005)

2176 | SHIPILINA et AL.
variation in both traits andwe found amixture of the two distinctmtDNAhaplotypes(togetherwith21caseswherebirdscarriedathirdmtDNAhaplotypethatpreviouslyhasbeenshowntodifferfromthetypical tristis haplotype at a single position).This supports previousstudiesindicatingpresenceofmtDNAintrogressionbetweenthetwosubspecies(Marovaetal.,2009,2013)butdoesnotprovideanyinfor-mationaboutpotentialnucleargenomicgeneexchange.Toinvestigatethepatternsofadmixture inmoredetail,weanalyzedgenome-widegenetic variation using SNP data.We conducted a set of differenttypesofanalysesusingthenuclearSNPsthatallsupportconsiderablegenetic admixture: principal component analysis, assignment tests,assessmentofgeneticdifferentiation,andanalysisofthecontributionofsubspeciesspecific“diagnostic”geneticmarkerstosampleswithinthe zone of sympatry. The principal component analysis showed aclear separationof the allopatric populations ofabietinus and tristis intotwodistinctclustersandanintermediatepositionformostoftheinvestigated samples fromwithin the sympatric zone.This indicatesthat gene flow between the two subspecies has resulted in higherdegreeofallelesharingbetweenindividualsinsympatry.Supportforthis also comes from our clustering analysiswhich has the highestlikelihoodfortwodistinctpopulations(allopatricabietinusandtristis,respectively),whilesamplesfromthesympatriczonehaveageneticset-upbuiltupbyintermixedcontributionsfromthetwosubspecies.Whenassessingthefractionsofsharedandfixedpolymorphismsandestimatingtheglobalgeneticdifferentiation(FST),weobservedasub-stantially lower leveloffixeddifferences (highergeneticdifferentia-tion)andhigherfractionofsharedpolymorphisms(lowergeneticdif-ferentiation)betweensubspeciesinsympatrythaninallopatry,againsupportingascenariowithadmixtureandallelesharingintheregionwheresubspeciesrangesoverlap,althoughthisshouldbetakenascir-cumstantialratherthanhardevidence,asthegroupingofindividualsinthesympatricregionwasbasedonmorphotypealone.
ItshouldbenotedthatthePCAanalysisrevealedthatonetristis sample showedcleardifferentiation to all other tristis samples.ThissampleoriginatesfromtheChukotkaregioninfareasternSiberiaandishenceconsiderablygeographically separated fromallother tristis. Inaddition,thetristis-2mtDNAhaplotypewasnotfoundinallopat-ric tristis.Theseobservations indicatethatthere isgeneticstructurealsowithinthetristissubspecies,butextendedsamplingisobviouslynecessarytoquantifythedegreeofpotentialdifferentiationbetweentristispopulationsandhowthatrelatestoobserved“intrasubspecific”variationinphenotypictraitssuchassongandplumagecolor(Marovaetal.,2009,2013).Wealsoobservedasomewhathigherfractionofprivate polymorphisms in tristis (29% of all SNPs) than in abietinus (22%ofallSNPs).Thatindicatesaslightlylargereffectivepopulationsizeintrististhaninabietinusandisinagreementwiththelargerdis-tributionrangebutcouldalsobeaconsequenceofthatcurrenttristis populationsexpandedfrommultipleglacialrefugia(Nazarenko,1982,1985;Vorontsov,1949).
Withinthesympatricregion,wefoundstrongassociationbetweengeneticset-upandappearanceinsomeindividuals,twobirdswithabi-etinusmorphotypeandsongalsohadtheabietinusmtDNAandcloseto 100% abietinus diagnostic nuclear SNPs, and 9 birdswith tristis
morphotypehadtristismtDNAandamajorfractionoftristisdiagnosticnuclearSNPs.(Itshouldbenotedthatwealsoobserveasmallfractionof heterozygous sites and sites diagnostic for the other subspecies(i.e.,abietinus alleles in a tristis sample) in these particular individu-als. Itcouldeither indicate that i) thesesamplesare representativesformulti-generation recurrent backcrossings into oneof the paren-tal subspecies, and/or ii) thatour selectionofdiagnosticSNPscon-tainsaminorproportionofmarkersthatinfactaresharedorprivate).However,sixbirdsthatwerescoredastristisbasedonplumagechar-actersandmtDNAperformed themixedsong type,and therewerealsoseveralexampleswheretheappearanceandsongdidnotmatchthegeneticset-up.Themostevidentcasesincludefivebirdswithtyp-icalabietinusmorphotypethatactuallyhadthetristismtDNA,andamajorityofdiagnostictristisnuclearSNPsthatalsoperformedmixedsongandonebird,alsoamixedsinger,thathadthetypicaltristismor-photypeandalmost100%tristisdiagnosticnuclearSNPsbutabietinus mtDNA.ThisisinlinewithpreviousanalysisofcorrelationsbetweenmtDNAandphenotypeinchiffchaffsampledacrossringingstationsinNetherlandswherealargeproportionofbirdsidentifiedasabietinusinhandactuallyhadatypicaltristismtDNAtype(DeKnijff,vanderSpek,&Fischer,2012)and indicates that speciesdeterminationbasedonmorphologycanbeuncertain.Hence,manybirdsthatappearasobvi-ous,distinctabietinus or tristiscanharborageneticset-upthatisamixbetweenspeciesorevenalmostcompletelyfixedforforeignalleles.Weconcludefromthecombinedanalysesofphenotypes,mtDNA,andgenome-wideSNPsinbothallopatryandsympatrythatextensivehis-toricalandongoinggeneflowbetweenthesubspecieshaveresultedinatransitionzoneofmorphotypesandsongtypes.
Onarelatednote,itmightbeofinteresttouseourresultstospec-ulateinmoredetailaroundthetaxonomicstatusofchiffchaffpopula-tions,inhabitingthe1,500kmlongregionfromtheArkhangelskregionin the north to the southernUralMountains in the south and alsotheregioncoveringpartsofthewestSiberianplain.Basedonspear-heading naturalist work during the nineteenth century (Severtsov,1873),ithasbeensuggestedthattheregionharborsadistinctchiff-chaff taxon, “fulvescens”, described as expressing intermediate char-acters (morphotype,song)comparedtoabietinusandtristis (Dean&Svensson,2005).LaterobservationssuggestedthatchiffchaffsampledinthewestSiberianplainarea(betweentheUralmountainsandtheYeniseiriver)sometimesshowedamorphotypethatdeviatedslightlyfromtristissamplesfromeastoftheYeniseiriver,inessenceshowingpartlymoreyellowishplumage (Dean&Svensson,2005).AdditionalstudiesinthesouthernUralsconcludedthatbirdsinthisregionshow-ingintermediatemorphotypeandmixedsongprobablywerehybridsand/orbackcrossesbetweenabietinusandtristis(Marovaetal.,2009,2013;Ticehurst, 1938), initially these formswere named “riphaeus” (Ticehurst,1938).Asmentionedabove,weobservedacleartransitionzonebetweendistinctabietinusandtristisacrossthisregion,thediag-nosticmtDNAhaplotypeswere intermixedintheareaandgenome-widenuclearSNPvariationdidnot indicateanyadditional structur-ingbesidesthatallopatricabietinusandtristisaregeneticallydistinct,butexchangesubstantialportionsofthegenomewheredistributionranges overlap. This leads to the conclusion that the “fulvescens”

| 2177SHIPILINA et AL.
(“riphaeus”) form represents individualswith amixed genetic set-upfrombothtristisandabietinusratherthanadistincttaxonandthatthemostplausibleexplanationfortheoccurrenceof“fulvescens”typebirdseastoftheimmediatecontactzoneisthattheserepresentindividualswithintrogressedabietinusallelesonatristisgenomicbackground(seealsodiscussionondirectionalgeneflowbelow).This scenariocouldeasily be envisioned if hybrids and backcrosses preferentially inter-breedwith“pure”tristisgeneratingamoredilutedtransitionzoneeastoftheimmediatecontactzone.
Theevolutionofreproductiveisolationisgenerallyalong-drawn-out process, initiated by divergence in traits related to prezygoticbarriersandendingwithpostzygoticgeneticincompatibilities(Coyne&Orr,2004;Price,2008).Thetwosubspeciesabietinusandtristisdif-ferinplumagecolor,vocalizations,size,andhabitatpreference,andtheyhavedistinctbreedingandwinteringgroundsexceptforazonearoundtheUralmountainswheretheyco-occurduringbreedingsea-son (delHoyoetal., 2006;Komarova&Shipilina, 2010;Marova&Alekseev,2008;Marovaetal.,2009,2013;Svensson,1992).Giventhe specific characteristics of each subspecies and the relativelydeep divergence (1.5–2%mtDNA divergence; Helbig etal., 1996),onecanassumethatbothlocaladaptationandstochasticprocessesduringallopatricseparationhaveplayedaroleindrivingphenotypicdivergenceandthatindividualswithintermediatecharactersresult-ing from hybridization and backcrossingmay have reduced fitnessas a consequence of lower attractiveness to mates, intermediatemigration patterns, and nonoptimal utilization of specific habitats(Rundle&Nosil,2005).Insupportofthat,allopatrictristisandabiet-inusshownegligiblereactionstotheothersubspecies’vocalizations(Marovaetal.,2009;Marovaetal.2017;Martens&Meinche,1989),indicating that song recognition could be a strong prezygotic bar-rier.However,withinsympatricregions,chiffchaffofbothsubspeciesrespondmore actively to alien song types—theunderlyingmecha-nismbehind that difference is not known (Marova etal., 2009). Inaddition, both abietinus and tristis females have been observedpairingupwithmaleswith intermediateplumagecharacters in thefield(unpublishedobservations)indicatingthatpotentialprezygoticbarriers related tobothvocalizationsandmorphologyarestillper-meable. Our analysis of genomic composition of birds within thesympatric zone showed that several individuals express diagnosticplumagecharactersandperformdiagnosticsongdespiteharboringtheforeignmtDNAtypeand/oraconsiderableproportionofforeignnuclearalleleswhichindicatesthatthegeneticelementsunderlyingmorphologicalandvocalizationdifferencesoccurinarestrictedpor-tionofthegenome.Itistooearlytospeculateiftheseregionsalsocontain genes of importance for reproductive isolation. However,investigatingthegenome-widepatternofdifferentiationforamuchlargersetofnuclearmarkersand investigatingthegenomicadmix-ture proportions in birdswith intermediate phenotypeswill give achance to identify regions that show elevated differentiation andassess ifthosehaveappearedasaresultofrestrictedgeneflowinparticulargenomicregionsorasaconsequenceofadaptationtodif-ferenthabitatsduringseparation(Harrison&Larson,2016;Wolf&Ellegren,2016).
Itislikelythatthecurrentregionofsympatryisasecondarycon-tact zone formedafter recolonization fromglacial refugia, and suchzonesmight have formed recurrently duringwarmer periods duringthe Pleistocene glaciation cycles allowing for episodic interspe-cificgeneflow.Wefoundthatintrogressionseemtobebiasedwitha higher proportionof tristismtDNAhaplotypes and nuclear alleleswithin the sympatric region meaning that backcrossing predomi-nantlyoccursintothetristislineage.Thisdirectionalintrogressionmayexplainthepresenceofbirdswithsomewhatyellowishplumageinthewest Siberianplain (seediscussion aboveon “fulvescens”), as thesecouldcarryafractionofabietinusallelesthathavespreadthroughthetristispopulationeastwardsbyrecurrentbackcrossings.Asoursam-plingwasrestrictedtoonly includemales,weareunfortunatelynotabletocomparetothegeographicdistributionofmtDNAhaplotypesandcorrelationbetweenmtDNAandgenomiccompositioninfemales.As many avian taxa have female-biased dispersal (Mabry, Shelley,Davis,Blumstein,&VanVuren,2013)andacomparativelyhighrateofmtDNAintrogressionacrossdivergent lineages(reviewedinRheindt& Edwards, 2011), it is plausible that additional analyses includingfemale samples fromacross thedistribution rangesofabietinus andtristiswouldprovidemorequantitative informationonthepresenceandpotentialbiasinintrogressionpatternsacrossthetwosubspecies.
The introgressionbias into tristis appeared tobestronger in thenorthernsympatricregion.ItisknownfrompreviousanalysesthatthesouthernUralsprobablyservedasarefugiumwithforestvegetationthatpreglacialfaunaofEuropeinhabitedduringglaciation(Vorontsov,1949).ItispossiblethattheancestorsofcurrentEuropeanchiffchaffhave been present in this region also during glacial maxima. Afterthe retreatof the ice cover, Siberian chiffchaff thenprobably recol-onizedSiberiafromthesoutheastfromone(orboth)oftheSiberianAltai/Sayanrefugia (Nazarenko,1982,1985)coming incontactwithEuropeanchiffchaffinthesouthernUralsrelativelyearlyaftertheicesheetretracted.TheArkhangelskregion,incontrast,wasfullycoveredby glaciation, and both European and Siberian chiffchaff probablycolonized thatareamuch later indicating that thenorthernsympat-ricregionisyoungerthanthesouthernzone.Ifhybridizationreducesfitness, asmight be expected given the considerable differences inplumage, song, andmigration routes, a longer history of secondarycontactmayhaveledtoincreasedreinforcementofprezygoticbarriersinthesouthernsympatricregion,decreasingtherateofbackcrossingintotristis,butitisnottheonlyexplanationforthispattern.Anotheraspect,albeitnotmutuallyexclusive,relatestotheabundanceofeachsubspeciesindifferentregionsofthesympatriczone.Inthenorthernzone,thepredominanthabitattypeisboreal(coniferous)forestwhilethehabitat ismore fragmented in the southern zonewith apatchydistributionofmixedandborealforest.Abietinusisbreedinginhigherdensitiesinmixedforestwhiletristisismorecommoninboreal(conif-erous)forestandtherelativedensityoftristisishigherinthenorthernzone(Komarova&Shipilina,2010).Thislikelyallowsformoreoppor-tunities for backcrossing into tristis in the northern zone. Anotherquestionrelatedtothebiasedintrogressioniswhetherabietinusandtristis differ in the ability to distinguish pure “consubspecifics” fromhybrids/backcrosses.Theobservedpatternwould thensuggest that

2178 | SHIPILINA et AL.
tristis aremoreprone tomatewithhybridsbutmoredataonmatepreferencewillbeneededbeforethiscanbeelucidated.
In many avian study systems, there is clear evidence for male-biasedgeneflowasaresultofreducedfitnessinfemalehybridsduetoexposureofrecessiveincompatibilityallelesontheZ-chromosome(Haldane’s rule, Haldane, 1922; Price, 2008). This has also beenreportedinanothercontactzoneinvolvingadifferentpairofchiffchaffspecies(Bensch,Helbig,Salomon,&Seibold,2002).Ourobservationthatonebirdwith tristismorphotypeand>95% tristisnuclearSNPsactuallyharboredabietinusmtDNAindicatesthatfemalehybridsresult-ingfromacrossbetweenamaletristisandafemaleabietinusarefer-tileandcaninterbreedwithtristisallowingforabietinusmitochondrialintrogression into tristis.However, there isno individual inourdatasetthatshowstheoppositepattern(predominantlyabietinusnuclearallelesandtristismtDNA),allindividualswithtristismtDNAalsohavethe largest proportion of diagnostic tristis nuclear SNPs.This couldpotentiallybearesultoflowfitnessofhybridfemalesfromtherecip-rocalcross(maleabietinus,femaletristis),orthatthesehybridfemalesdonotmatewithabietinusmales,butthesamplesizeislimited,andextendedsamplingandmtDNAandnucleargenomeanalyseswouldbeneededbeforethiscanbeverified.
5 | CONCLUSION
Inthisstudy,wecombinegeneticdatawithmorphologyandvocaliza-tionsacrossahybridzonebetweenEuropean(abietinus)andSiberian(tristis)chiffchaff.Ourresultsindicatethattheoverallgeneticdiffer-entiationbetweensubspeciesislowdespiteconsiderablephenotypicdivergence.Alargeproportionofbirdsinthehybridzoneexhibitinter-mediatephenotypiccharactersandamixofgeneticancestry,indicat-ing extensive ongoing and past gene flow. Patterns of phenotypicand genetic variation vary between the northern and the southernpartofthehybridzone,probablyreflectingdifferencesinpopulationdensitiesand/orrelativeageofsecondarycontact.Thedataindicatethatbirdsshowingintermediatecharactersverylikelyrepresentindi-viduals resulting from recurrent backcrossings and introgression of(predominantly)abietinusallelesintoatristisgenomicbackground,andthepreviouslydescribedsubspecies“fulvescens”isthereforenottobeconsideredadistincttaxon.Thedataalsopointtothedifficultiesinidentificationofspecific individualsbasedonphenotypiccharactersalone.
ACKNOWLEDGMENTS
N.B. is supported by a Junior Research Grant from the SwedishResearchCouncil(VR2013-4508).Weacknowledgeadditionalfund-ingforDNAquantificationandqualitycontrol,librarypreparationandsequencing from Kungliga Fysiografiska Sällskapet i Lund (Nilsson-Ehle Donations), Helge Ax:son Johnsons Foundation and the LarsHierta Memorial Foundation. The SNP&SEQ Technology PlatformandUppsalaGenomeCenterperformedthelibrarypreparationsandthe sequencing supported by Science for Life Laboratory (SciLife,
Uppsala); anational infrastructure fundedby theSwedishResearchCouncil(VR-RFI)andtheKnutandAliceWallenbergFoundation.ThecomputationswereperformedonresourcesprovidedbySNICthroughUppsala Multidisciplinary Center for Advanced ComputationalScience(UPPMAX;Lampa,Dahlö,Olason,Hagberg,&Spjuth,2013)underProject#b2013146.SamplecollectionandprimaryprocessingweresupportedbytheRussianFoundationofBasicResearch(RFBR)(14-04-01259, 15-29-02771) and the Russian Science Foundation(RSF)(14-50-00029).M.S.issupportedbyGordonandBettyMooreFoundation’sEPiQSInitiativeGrant(GBMF4307).
CONFLICT OF INTERESTS
Theauthorsdeclarenofinancialornonfinancialconflictofinterests.
REFERENCES
Abbott,R.J.,Barton,N.H.,&Good,J.M.(2016).Genomicsofhybridizationanditsevolutionaryconsequences.Molecular Ecology,25,2325–2332.
Alcaide,M., Scordato, E. S., Price,T.D., & Irwin,D. E. (2014). Genomicdivergenceinaringspeciescomplex.Nature,511,83–85.
Barton,N.H.,&Hewitt,G.M. (1989).Adaptation,speciationandhybridzones.Nature,341,497–503.
Bensch,S.,Åkesson,S.,&Irwin,D.E.(2002).TheuseofAFLPtofindaninformativeSNP:Geneticdifferencesacrossamigratorydivideinwil-lowwarblers.Molecular Ecology,11,2359–2366.
Bensch,S.,Grahn,M.,Müller,N.,Gay,L.,&Åkesson,S.(2009).Genetic,mor-phological,andfeatherisotopevariationofmigratorywillowwarblersshowgradualdivergenceinaring.Molecular Ecology,18,3087–3096.
Bensch,S.,Helbig,A.J.,Salomon,M.,&Seibold,I.(2002).Amplifiedfrag-ment length polymorphism analysis identifies hybrids between twosubspeciesofwarblers.Molecular Ecology,11,473–481.
Bensch,S.,Irwin,D.E.,Irwin,J.H.,Kvist,L.,&Åkesson,S.(2006).Conflictingpatterns ofmitochondrial and nuclearDNA diversity inPhylloscopus warblers.Molecular Ecology,15,161–171.
Bierne,N.,Gagnaire,P.-A.,&David,P.(2013).Thegeographyofintrogres-sion inapatchyenvironmentandthe thorn in thesideofecologicalspeciation.Current Zoology,59,72–86.
Coyne, J. A., & Orr, H. A. (2004). Speciation. Sunderland, MA: SinauerAssociates.
DeKnijff,P.,vanderSpek,V.,&Fischer,J.(2012).GeneticidentityofgreychiffchaffstrappedintheNetherlandsinautumnsof2009–11.Dutch Birding,34,386–392.
DePristo,M.A.,Banks,E.,Poplin,R.,Garimella,K.V.,Maguire,J.R.,Hartl,C.,…Daly,M.J.(2011).Aframeworkforvariationdiscoveryandgeno-typingusingnext-generationDNAsequencingdata.Nature Genetics,43,491–498.
Dean,A.R.,&Svensson,L.(2005).Siberianchiffchaffrevisited.British Birds,98,396–410.
Delmore,K.E.,Hubner,S.,Kane,N.C.,Schuster,R.,Andrew,R.L.,Camara,F.,···Irwin,D.E.(2015).Genomicanalysisofamigratorydividerevealscandidategenesformigrationandimplicatesselectivesweepsingen-eratingislandsofdifferentiation.Molecular Ecology,24,1873–1888.
Ellegren,H.,Smeds,L.,Burri,R.,Olason,P.,Backström,N.,Kawakami,T.,···Wolf, J. B.W. (2012). The genomics of species differentiation inFicedulaflycatchers.Nature,491,756–760.
Evanno, G., Regnaut, S. and Goudet, J. 2005. Detecting the number ofclustersofindividualsusingthesoftwarestructure:asimulationstudy.Molecular Ecology,14: 2611–2620.
Haldane,J.B.S.(1922).Sexratioandunisexualsterilityinhybridanimals.Journal of Genetics,12,279–281.

| 2179SHIPILINA et AL.
Harrison,R.G.,&Larson,E.L.(2016).Heterogeneousgenomedivergence,differentialintrogression,andtheoriginandstructureofhybridzones.Molecular Ecology,25,2454–2466.
Helbig, A. J., Martens, J., Seibold, I., Henning, F., Schottler, B., &Wink,M. (1996). Phylogeny and species limits in the Palaearctic chiffchaffPhylloscopus collybita complex: Mitochondrial genetic differentiationandbioacousticevidence.Ibis,138,650–666.
Helbig,A. J., Salomon,M., Bensch, S., & Seibold, I. (2001).Male-biasedgene flow across an avian hybrid zone: Evidence from mitochon-drial and microsatellite DNA. Journal of Evolutionary Biology, 14, 277–287.
Hewitt,G.M.(2004).Thestructureofbiodiversity-insightsfrommolecularphylogeography.Frontiers in Zoology,1,4.
Hewitt, G. M. (2011). Quaternary phylogeography: The roots of hybridzones.Genetica,139,1–22.
delHoyo, J., Elliot,A.,&Christie,D.A. (2006).Handbook of the birds of the world. Old world flycatchers to old world warblers. Barcelona: LynxEdicions.
Irwin,D.E.,Alcaide,M.,Delmore,K.E.,Irwin,J.H.,&Owens,G.L.(2016).Recurrentselectionexplainsparallelevolutionofgenomic regionsofhighrelativebutlowabsolutedifferentiationinaringspecies.Molecular Ecology,25,4488–4507.
Kawakami,T.,Smeds,L.,Backström,N.,Husby,A.,Qvarnström,A.,Mugal,C.,···Ellegren,H.(2014).Ahigh-densitylinkagemapenablesasecond-generationcollaredflycatchergenomeassemblyandrevealsthepat-ternsof avian recombination ratevariationand chromosomal evolu-tion.Molecular Ecology.,23,4035–4058.
Komarova, A. F., & Shipilina, D. A. (2010). Hybrid population of eastern European and Siberian forms of chiffchaff (Phylloscopus collybita abieti-nus, Ph. (c.) tristis) in the Arkhangelsk area.XVIIInternationalconferenceofstudents,post-graduatestudentsandyoungscientists;Lomonosov:MAKSPress,211–212.
Lampa,S.,Dahlö,M.,Olason,P.I.,Hagberg,J.,&Spjuth,O.(2013).Lessonslearned from implementing a national infrastructure in Sweden forstorageandanalysisofnext-generationsequencingdata.Gigascience,25,9.
Li, H., & Durbin, R. (2010). Fast and accurate long-read alignmentwithBurrows-Wheelertransformation.Bioinformatics,26,589–595.
Li,H.,Handsaker,B.,Wysoker,A.,Fennell,T.,Ruan,J.,Homer,N.,···Durbin,R. (2009). The Sequence Alignment/Map format and SAMtools.Bioinformatics,25,2078–2079.
Lindholm,A. (2008).MixedsongsofchiffchaffsinnorthernRussia.Alula,3,108–115.
Lundberg,M.,Åkesson,S.,&Bensch,S.(2011).Characterizationofadiver-gent chromosome region in thewillowwarblerPhylloscopus trochilus using avian genomic resources. Journal of Evolutionary Biology, 24,1241–1253.
Lundberg,M.,Boss,J.,Canbäck,B.,Liedvogel,M.,Larson,K.W.,Grahn,M.,···Wright,A.(2013).Characterisationofatranscriptometofindsequence differences between two differentiallymigrating subspe-ciesofthewillowwarblerPhylloscopus trochilus. BMC Genomics,14,e330.
Mabry,K.E.,Shelley,L.E.,Davis,K.E.,Blumstein,D.T.,&VanVuren,D.H.(2013).Socialmatingsystemandsex-biaseddispersalinmammalsandbirds:Aphylogeneticanalysis.PLoS One,8,e57980.
Marova, I., Shipilina, D., Fyodorov, V., Alekseev, V., Ivanitskii, V. (2017).Mixed singing and hybridization between two chiffchaff taxa(Phylloscopuscollybitaabietinus -Ph.c. tristis) insecondarycontactzones.[SubmittedManuscriptInPress]
Marova,I.M.,&Alekseev,V.N.(2008).Populationstructureanddistribu-tionofvocaldialectsof chiffchaff (Phylloscopus collybita) in southernUralmountains.Proceedings of the South Ural State Natural Reserve,1,306–318.
Marova, I. M., Fedorov,V.V., Shipilina, D.A., &Alekseev,V. N. (2009).Genetic andvocal differentiation in hybrid zones of passerine birds:
SiberianandEuropeanchiffchaffs(Phylloscopus [collybita] tristisandPh. [c.] abietinus) in the southernUrals.Doklady Biological Sciences,427,384–386.
Marova, I.M.,&Leonovich,V.V. (1993).HybridizationbetweenSiberian(Phylloscopus collybita tristis) and east European (Ph. collybita abieti-nus)chiffchaffsintheareaofsympatry.Sbornik Trudov Zoologicheskogo Muzeya, Moskovskogo Gosudarstvennogo Universiteta,30,147–163.
Marova, I. M., Shipilina, D. A., Fedorov, V. V., & Ivanitskii, V. V. (2013).Siberian and East European chiffchaffs: Geographical distribution,morphological features, vocalization, phenomenon of mixed singing,andevidencesofhybridizationinsympatryzone.InN.R.Garcia&J.L.CJ (Eds.),El mosquitero ibérico (pp.119–139). Leon:Grupo IbericodeAnillamiento.
Martens,J.,&Meinche,C.(1989).Dersibirischezilpzalp(Phylloscopus col-lybita tristis):Gesangundreactioneinermitteleuropäischenpopulationimfreilandversuch.Journal of Ornithology,130,455–473.
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K.,Kernytsky,A.,Garimella,K.,Altshuler,D.,Gabriel,S.,Daly,M.,etal.(2010). The genome analysis toolkit: AMapReduce framework foranalyzingnext-generationDNAsequencingdata.Genome Research,20,1297–1303.
Nazarenko, A. A. (1982). On faunistic cycles (extinction-expansion-extinction) of a woodland ornithofauna of the eastern Paleoarctic.Journal of Fundamental Biology,43,823–835.
Nazarenko,A.A.(1985).Selectedhistoricalandbiogeographicalproblems,relatedtoHimalayas(addressedforwoodlandornithofauna).Journal of Fundamental Biology,46,41–54.
Nosil, P., & Feder, J. L. (2012). Genomic divergence during speciation:Causes and consequences. Philosophical Transactions of the Royal Society B: Biological Sciences,367,332–342.
Nosil, P., Harmon, L. J., & Seehausen, O. (2009). Ecological explana-tions for (incomplete) speciation. Trends in Ecology & Evolution, 24,145–156.
Patterson, N., Price,A. L., & Reich, D. (2006). Population structure andeigenanalysis.PLoS Genetics,2,e190.
Payseur,B.A.,&Rieseberg,L.H.(2016).Agenomicperspectiveonhybrid-izationandspeciation.Molecular Ecology,25,2337–2360.
Pfeifer,B.,Wittelsbuerger,U.,Ramos-Onsins,S.E.,&Lercher,M.J.(2014).PopGenome: An efficient Swiss army knife for population genomicanalysesinR.Molecular Biology and Evolution,31,1929–1936.
Price,T.D. (2008).Speciation in birds.GreenwoodVillage,CO:Roberts&CompanyPublishers.
Price,T.D.(2010).Therolesoftimeandecologyinthecontinentalradi-ation of the Old World leaf warblers (Phylloscopus and Seicercus).Philosophical Transactions of the Royal Society B: Biological Sciences,365,1749–1762.
Pritchard, J. K., Stephens,M., &Donnelly, P. (2000). Inference of pop-ulation structure using multilocus genotype data. Genetics, 155,945–959.
Rheindt, F. E., & Edwards, S. V. (2011). Genetic introgression: An inte-gral but neglected component of speciation in birds. The Auk, 128, 620–632.
Rundle,H.D.,&Nosil,P. (2005).Ecologicalspeciation.Ecology Letters,8,336–352.
Sambrook,J.,Fritsch,E.F.,&Maniatis,T.(1989).Molecular cloning: A labo-ratory manual.ColdSpringHarbor,NY:ColdSpringHarborLaboratoryPress.
Schluter,D.(2009).Evidenceforecologicalspeciationanditsalternative.Science,323,737–741.
Severtsov,N.A.(1873).Vertical and horizontal distribution of Turkestan ani-mals,Vol.2.MoscowRussianFederation:Proceedingsof society fornaturalhistory,anthropologyandethnography.
Smeds, L., & Künstner,A. (2011). ConDeTri - a content dependent readtrimmerforIlluminadata.PLoS One,6,e26314.
Stepanyan,L.S.(1990).List of ornithofauna of USSR.Moscow:Nauka.

2180 | SHIPILINA et AL.
Sushkin,P.P.(1897).BirdsoftheUfaprovince.Proceedings of the knowledge about the flora and the fauna of the Russian Empire[InRussian],4,1–331.
Svensson, L. (1992). Identification guide to European passerines. Thetford,UK:BritishTrustforOrnithology.
Ticehurst,C.(1938).A systematic review of the genus Phylloscopus.London:OxfordUniversityPress.
Toews,D.P.,& Irwin,D.E. (2008).Crypticspeciation inaHolarcticpas-serinerevealedbygeneticandbioacousticanalyses.Molecular Ecology,17,2691–2705.
Van derAuwera, G.A., Carneiro,M.O., Hartl, C., Poplin, R., DelAngel,G.,Levy-Moonshine,A.,Jordan,T.,Shakir,K.,Roazen,D.,Thibault,J.,et al. (2013). From FastQ data to high confidence variant calls: Thegenome analysis toolkit best practices pipeline. Current Protocols in Bioinformatics,11,11–33.
Vorontsov, E. M. (1949). Birds of Kama Urals (Molotov oblast), p. 431,Gorkiy:GorkovskyGosUniversitet.
Wolf,J.B.W.,&Ellegren,H.(2016).Makingsenseofgenomicislandsofdifferentiationinlightofspeciation.Nature Reviews Genetics(AdvanceOnlinePublication),18,87–100.
SUPPORTING INFORMATION
AdditionalSupportingInformationmaybefoundonlineinthesupport-inginformationtabforthisarticle.
Figure S1ApproximatedistributionrangesofP. c abietinus(green)andP. c. tristis(yellow) Figure S2AnillustrationofthedifferentstepsinthegenerationandanalysisofnuclearDNA
How to cite this article:ShipilinaD,SerbynM,IvanitskiiV,MarovaI,BackströmN.Patternsofgenetic,phenotypic,andacousticvariationacrossachiffchaff(Phylloscopus collybita abietinus/tristis)hybridzone.Ecol Evol. 2017;7:2169–2180. https://doi.org/10.1002/ece3.2782