original article volasertib suppresses the growth of … article volasertib suppresses the growth of...
TRANSCRIPT

Am J Cancer Res 2016;6(11):2476-2488www.ajcr.us /ISSN:2156-6976/ajcr0037182
Original ArticleVolasertib suppresses the growth of human hepatocellular carcinoma in vitro and in vivo
Di-Wei Zheng1*, You-Qiu Xue1,2*, Yong Li3*, Jin-Ming Di4, Jian-Ge Qiu1, Wen-Ji Zhang1, Qi-Wei Jiang1, Yang Yang1, Yao Chen1, Meng-Ning Wei1, Jia-Rong Huang1, Kun Wang1, Xing Wei1, Zhi Shi1
1Department of Cell Biology & Institute of Biomedicine, College of Life Science and Technology, Jinan University, National Engineering Research Center of Genetic Medicine, Guangdong Provincial Key Laboratory of Bioen-gineering Medicine, Guangzhou 510632, Guangdong, China; 2Center for Clinic Immunology, Third Hospital at Sun Yat-sen University, Guangzhou 510630, China; 3Department of Gastrointertinal Surgery & General Surgery, Guangdong General Hospital, Guangdong Academy of Medical Sciences, Guangzhou, Guangdong 510080, China; 4Department of Urology, The 3rd Affiliated Hospital of Sun Yat-sen University, Guangzhou, Guangdong 510630, China. *Equal contributors.
Received August 3, 2016; Accepted August 9, 2016; Epub November 1, 2016; Published November 15, 2016
Abstract: Hepatocellular carcinoma (HCC) is the sixth most frequent malignant tumor with poor prognosis, and its clinical therapeutic outcome is poor. Volasertib, a potent small molecular inhibitor of polo-like kinase 1 (PLK1), is currently tested for treatment of multiple cancers in the clinical trials. However, the antitumor effect of volasertib on HCC is still unknown. In this study, our data show that volasertib is able to induce cell growth inhibition, cell cycle arrest at G2/M phase and apoptosis with the spindle abnormalities in human HCC cells. Furthermore, volasertib also increases the intracellular reactive oxidative species (ROS) levels, and pretreated with ROS scavenger N-acety-L-cysteine partly reverses volasertib-induced apoptosis. Moreover, volasertib markedly inhibits the subcutaneous xenograft growth of HCC in nude mice. Overall, our study provides new therapeutic potential of volasertib on hepa-tocellular carcinoma.
Keywords: Hepatocellular carcinoma, volasertib, ROS, apoptosis
Introduction
Hepatocellular carcinoma (HCC) is the sixth most frequent malignant tumor and the third leading cause of cancer-related mortality in the world [1]. Most cases of HCC are as a result of viral infection (hepatitis B or C) and metabolic toxins (alcohol or aflatoxin), and the patients with HCC are usually diagnosed when tumor is in an advanced stage [2]. Although treatment of HCC includes the combination of surgery, chemotherapy or radiotherapy, the 5-year sur-vival rates of patients with HCC is poor [3]. Consequently, it is necessary to develop new therapeutic strategies against HCC.
The polo-like kinases (PLKs) are a family of con-served serine/threonine kinases and play the critical role in the regulation of multiple stages of cell cycle, including mitotic entry/exit, centro-some maturation, and maintenance of the bipolar spindle [4]. It has been reported that PLK1 is overexpressed in up to 80% of cancers
and associated with poorer prognosis [5-7]. In human HCC, PLK1 expression is significantly upregulated and correlated with venous inva-sion, tumor nodules, Edmondson grade and survival rates [8, 9]. Knockdown of PLK1 signifi-cantly inhibits the growth of HCC cells by induc-ing cell cycle arrest at G2/M phase and apopto-sis [10, 11]. Additionally, the HBV X protein can enhance the expression of PLK1, and inhibition of PLK1 suppresses the HBV X protein-mediat-ed oncogenic transformation in HCC [12]. Therefore, PLK1 is a promising therapeutic tar-get of HCC. Volasertib (BI 6727) is a potent small molecule inhibitor of PLK1 (IC50=0.87 nM) by competing with ATP for binding to the ATP binding pocket, and 6- and 65-fold greater activity against two closely related kinases PLK2 (IC50=5 nM) and PLK3 (IC50=56 nM) [13]. In the preclinical studies, volasertib shows broad antitumor activity in several cancer mod-els including hematological malignancies, mel-anomas, cervical cancer and sarcomas [13-17].

Volasertib suppresses HCC growth
2477 Am J Cancer Res 2016;6(11):2476-2488
However, the antitumor effect of volasertib on HCC is still unknown. In this study, we demon-strated that volasertib could potently suppress the growth of human HCC in vitro and in vivo.
Material and methods
Cell culture and reagents
Human hepatoma cancer cell lines BEL7402, HepG2, SMMC7721 and SK-Hep-1 were cul-tured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/ml) and strepto-mycin (100 ng/ml) in a humidified incubator at 37°C with 5% CO2. Volasertib was purchased from ApexBio Technology. N-acetly-L-cysteine (NAC) and dihydroethidium (DHE) were pur-chased from Sigma-Aldrich. Anti-PARP (9542), anti-β-tubulin (A11031) and Anti-GAPDH (KM- 9002) antibodies were from Cell Signaling Technologies, Ruiying Biological and Sungene Biotech, respectively.
Cell viability assays
Cells were firstly seeded into a 96-well plate at a density of 5~10×103 cells per well, and incu-bated with drugs in three parallel wells for 72 h. Then 3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyl-tetrazolium bromide (MTT) was added to each well at a final concentration of 0.5 mg/ml. After incubation for 4 h, formazan crystals were dis-solved in 100 μl of DMSO, and absorbance at 570 nm was measured by plate reader. The concentrations required to inhibit growth by 50% (IC50) were calculated from survival curves using the Bliss method [18, 19]. For drug com-binational experiments, cells were treated with the indicated concentrations of volasertib com-bined with cisplatin for 72 h. The data were analyzed by CompuSyn software with the results showed as combination index (CI) val-ues according to the median-effect principle, where CI <1, =1, and >1 indicate synergism, additive effect and antagonism, respectively [20, 21].
Cell cycle assays
Cells were harvested and washed twice with cold phosphate-buffered saline (PBS), then fixed with ice-cold 70% ethanol for 30 min at 4°C. After centrifugation at 200×g for 10 min, cells were washed twice with PBS and resus-pended with 0.5 ml PBS containing PI (50 μg/
ml), 0.1% Triton X-100, 0.1% sodium citrate, and DNase-free RNase (100 μg/ml), and detected by FCM after 15 min incubation at room temperature in the dark. Fluorescence was measured at an excitation wavelength of 480 nm through a FL-2 filter (585 nm). Data were analyzed using ModFit LT 3.0 software (Becton Dickinson) [22, 23].
Apoptosis assays
Cell apoptosis was evaluated with flow cytome-try (FCM) assay. Briefly, cells were harvested and washed twice with PBS, stained with Annexin V-FITC and propidium iodide (PI) in the binding buffer, and detected by FACS Calibur FCM (BD, CA, USA) after 15 min incubation at room temperature in the dark. Fluorescence was measured at an excitation wave length of 480 nm through FL-1 (530 nm) and FL-2 filters (585 nm). The early apoptotic cells (Annexin V positive only) and late apoptotic cells (Annexin V and PI positive) were quantified [24, 25].
Reactive oxygen species (ROS) assay
Cells were incubated with 10 μM of DHE for 30 min at 37°C, and observed under fluorescence microscope (Olympus, Japan) immediately after washing twice with PBS. Five fields were taken randomly for each well [26, 27].
Western blot analysis
Cells were harvested and washed twice with cold PBS, then resuspended and lysed in RIPA buffer (1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 10 ng/ml PMSF, 0.03% aprotinin, 1 μM sodium orthovanadate) at 4°C for 30 min. Lysates were centrifuged for 10 min at 14,000×g and supernatants were stored at -80°C as whole cell extracts. Total protein con-centrations were determined with Bradford assay. Proteins were separated on 12% SDS-PAGE gels and transferred to polyvinylidene difluoride membranes. Membranes were blocked with 5% BSA and incubated with the indicated primary antibodies. Corresponding horseradish peroxidase (HRP)-conjugated sec-ondary antibodies were used against each pri-mary antibody. Proteins were detected using the chemiluminescent detection reagents and films [28, 29].
Immunofluorescence assay
BEL 7402 and Hep G2 cells grown on glass cov-erslips were washed thrice with PBS and fixed
Volasertib suppresses HCC growth
2478 Am J Cancer Res 2016;6(11):2476-2488
in 4% paraformaldehyde. The fixed cells were washed thrice with PBS containing 0.05% Tw- een 20, and then treated with PBS containing 0.2% Triton X-100 for 10 min at room tempera-ture followed by washing, blocking and incuba-tion with anti-β-tubulin for 1 hr at 37°C. The coverslips were washed, treated with Hoechst 33342 (5 μg/ml in PBS) for 5 min, washed again. The stained cells were analyzed by using the inverted Olympus microscope IX 70 FluoView system [30, 31].
Nude mouse assays
Balb/c nude mice were obtained from the Ba- lb/c nude mice were obtained from the Guang- dong Medical Laboratory Animal Center and maintained with sterilized food and water. Six female nude mice with 5 weeks old and 20 g weight were used for each group. Each mouse was injected subcutaneously with BEL7402 and HepG2 cells (2×106 in 100 μl of medium) under the shoulder. When the subcutaneous tumors were approximately 0.3×0.3 cm2 (two perpendicular diameters) in size, mice were randomized into four groups, and were injected intraperitoneally with vehicle alone (0.5% car-boxymethyl cellulose) and volasertib alone (15 mg/kg) every two days, respectively. The body weights of mice and the two perpendicular
diameters (A and B) of tumors were recorded. The tumor volume (V) was calculated according to the formula:
v 6 2A B 3= +r ` j
The mice were anaesthetized after experiment, and tumor tissues were excised from the mice and weighted. The rate of inhibition (IR) was cal-culated according to the formula [32, 33]:
IR 1Mean tumor weight of control group
Mean tumor weight of experimental group100%= - #
Statistical analysis
All results are expressed as mean ± standard deviation (SD). Statistical analysis of the differ-ences between two groups is performed with Student’s t-test. Values of P<0.05 were consid-ered as significant differences.
Results
Volasertib inhibits the growth of HCC cells in vitro
To investigate the effect of volasertib on the growth of HCC, cell survival was detected by
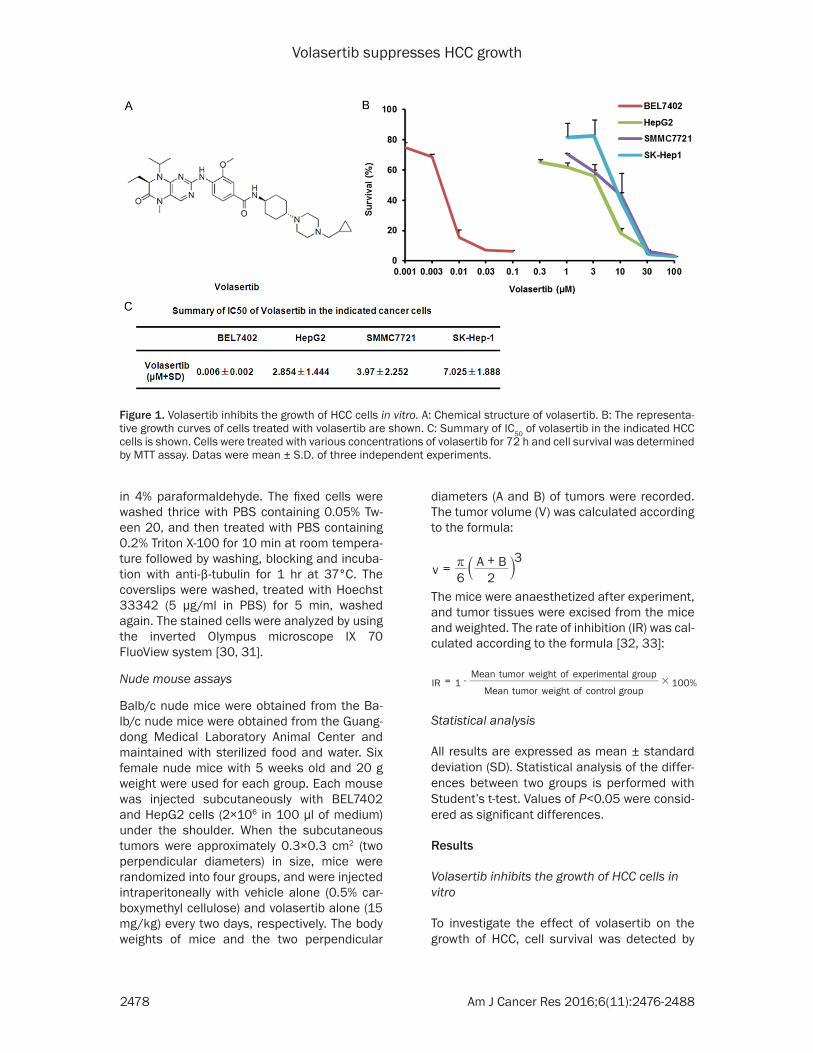
Figure 1. Volasertib inhibits the growth of HCC cells in vitro. A: Chemical structure of volasertib. B: The representa-tive growth curves of cells treated with volasertib are shown. C: Summary of IC50 of volasertib in the indicated HCC cells is shown. Cells were treated with various concentrations of volasertib for 72 h and cell survival was determined by MTT assay. Datas were mean ± S.D. of three independent experiments.
Volasertib suppresses HCC growth
2479 Am J Cancer Res 2016;6(11):2476-2488
MTT assay. The four HCC cell lines BEL7402, HepG2, SMMC7721 and SK-Hep-1 were treat-ed with various concentration of volasertib for 72 hr. As shown in Figure 1, the survivals of four cell lines were decreased in a dose-depen-dent manner after volasertib treatment. BEL7402 cells were most sensitive to volasert-ib with the IC50 values of 0.006 μM. The IC50 values of volasertib in other three HCC cell lines HepG2, SMMC7721 and SK-Hep1 were 2.854 μM, 3.970 μM and 7.025 μM, respectively.
Volasertib induces cell cycle arrest at G2/M phase in HCC cells
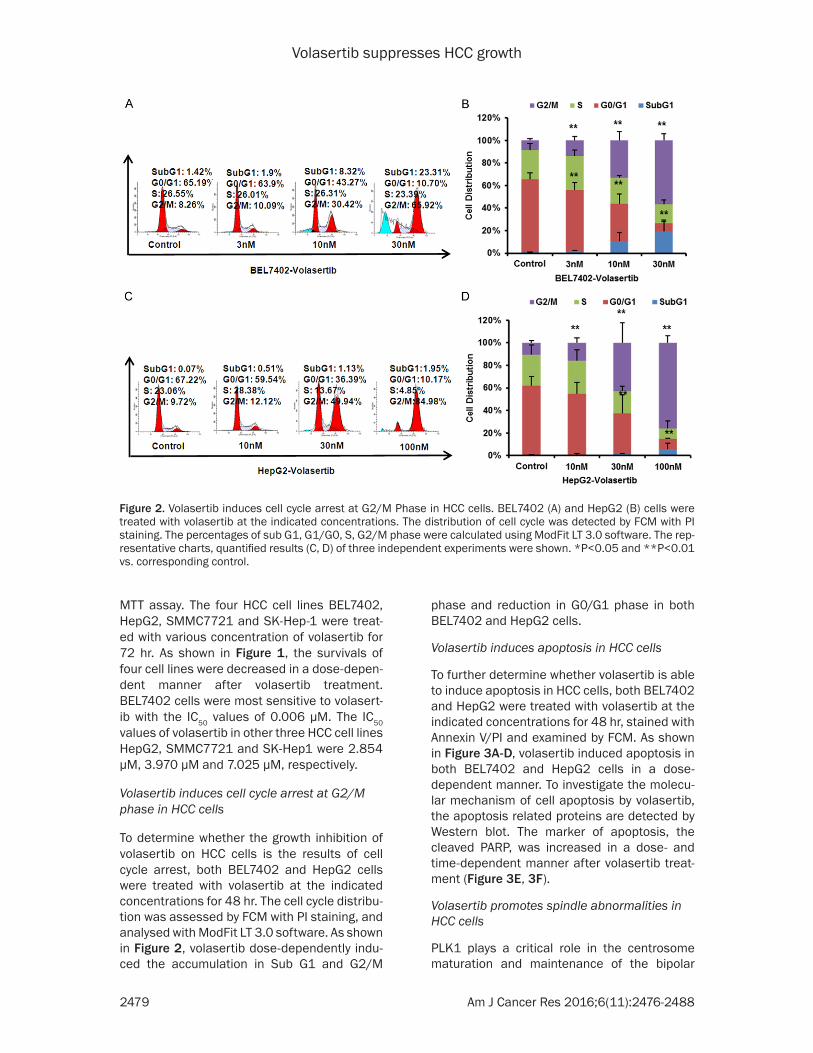
To determine whether the growth inhibition of volasertib on HCC cells is the results of cell cycle arrest, both BEL7402 and HepG2 cells were treated with volasertib at the indicated concentrations for 48 hr. The cell cycle distribu-tion was assessed by FCM with PI staining, and analysed with ModFit LT 3.0 software. As shown in Figure 2, volasertib dose-dependently indu- ced the accumulation in Sub G1 and G2/M
phase and reduction in G0/G1 phase in both BEL7402 and HepG2 cells.
Volasertib induces apoptosis in HCC cells
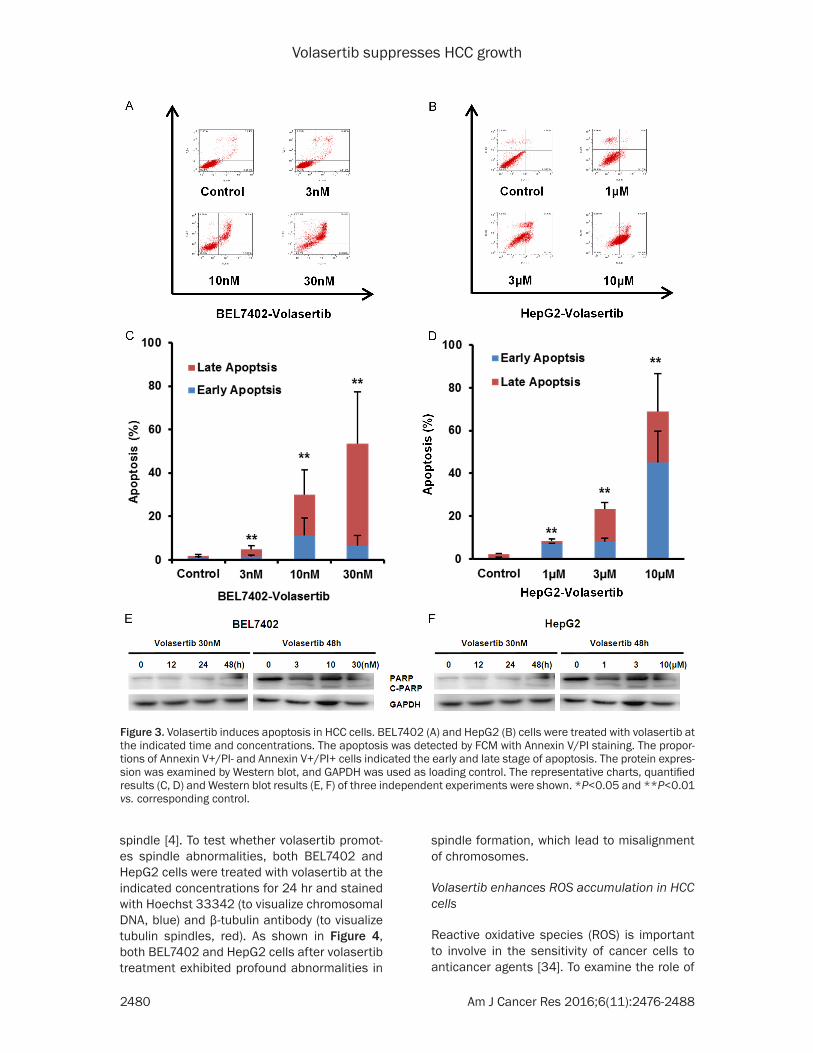
To further determine whether volasertib is able to induce apoptosis in HCC cells, both BEL7402 and HepG2 were treated with volasertib at the indicated concentrations for 48 hr, stained with Annexin V/PI and examined by FCM. As shown in Figure 3A-D, volasertib induced apoptosis in both BEL7402 and HepG2 cells in a dose-dependent manner. To investigate the molecu-lar mechanism of cell apoptosis by volasertib, the apoptosis related proteins are detected by Western blot. The marker of apoptosis, the cleaved PARP, was increased in a dose- and time-dependent manner after volasertib treat-ment (Figure 3E, 3F).
Volasertib promotes spindle abnormalities in HCC cells
PLK1 plays a critical role in the centrosome maturation and maintenance of the bipolar
Figure 2. Volasertib induces cell cycle arrest at G2/M Phase in HCC cells. BEL7402 (A) and HepG2 (B) cells were treated with volasertib at the indicated concentrations. The distribution of cell cycle was detected by FCM with PI staining. The percentages of sub G1, G1/G0, S, G2/M phase were calculated using ModFit LT 3.0 software. The rep-resentative charts, quantified results (C, D) of three independent experiments were shown. *P<0.05 and **P<0.01 vs. corresponding control.
Volasertib suppresses HCC growth
2480 Am J Cancer Res 2016;6(11):2476-2488
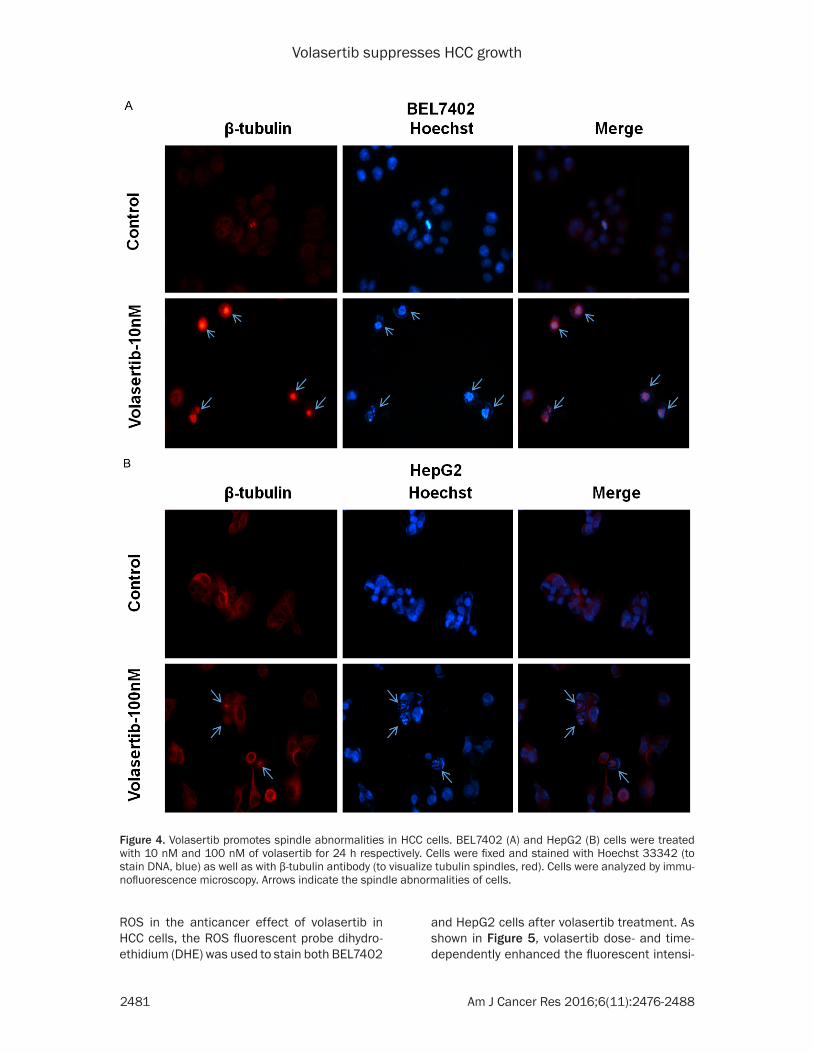
spindle [4]. To test whether volasertib promot- es spindle abnormalities, both BEL7402 and HepG2 cells were treated with volasertib at the indicated concentrations for 24 hr and stained with Hoechst 33342 (to visualize chromosomal DNA, blue) and β-tubulin antibody (to visualize tubulin spindles, red). As shown in Figure 4, both BEL7402 and HepG2 cells after volasertib treatment exhibited profound abnormalities in
spindle formation, which lead to misalignment of chromosomes.
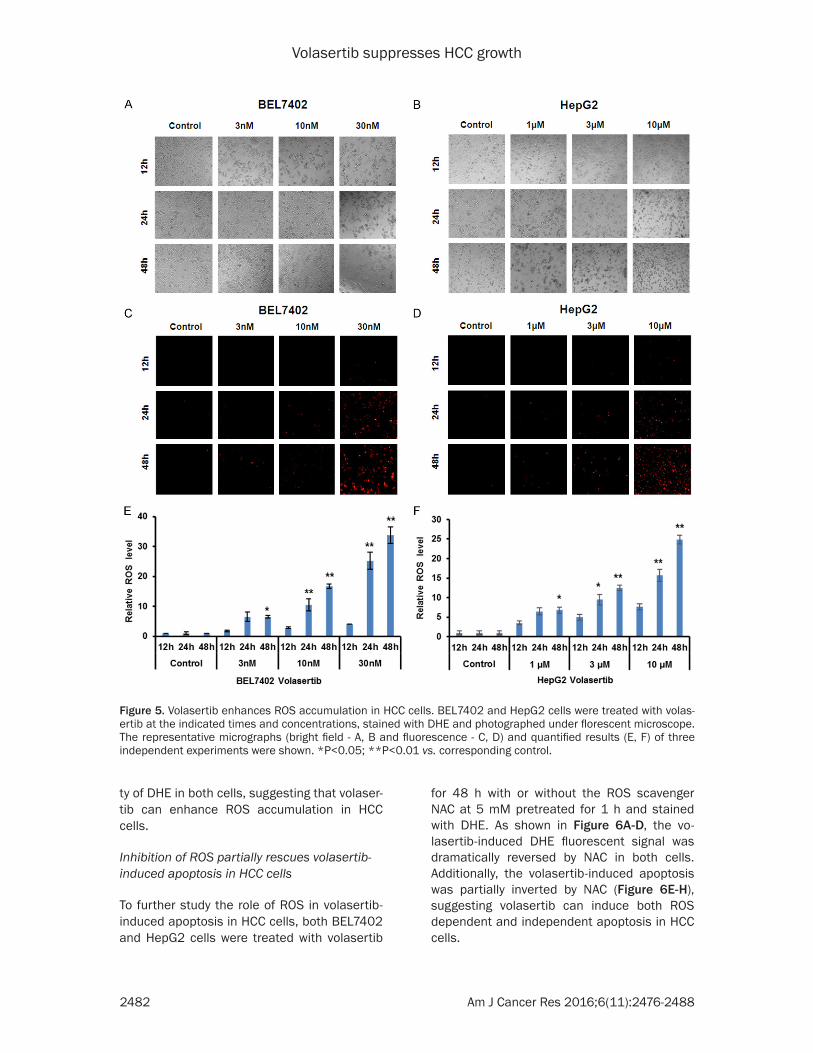
Volasertib enhances ROS accumulation in HCC cells
Reactive oxidative species (ROS) is important to involve in the sensitivity of cancer cells to anticancer agents [34]. To examine the role of
Figure 3. Volasertib induces apoptosis in HCC cells. BEL7402 (A) and HepG2 (B) cells were treated with volasertib at the indicated time and concentrations. The apoptosis was detected by FCM with Annexin V/PI staining. The propor-tions of Annexin V+/PI- and Annexin V+/PI+ cells indicated the early and late stage of apoptosis. The protein expres-sion was examined by Western blot, and GAPDH was used as loading control. The representative charts, quantified results (C, D) and Western blot results (E, F) of three independent experiments were shown. *P<0.05 and **P<0.01 vs. corresponding control.
Volasertib suppresses HCC growth
2481 Am J Cancer Res 2016;6(11):2476-2488
ROS in the anticancer effect of volasertib in HCC cells, the ROS fluorescent probe dihydro-ethidium (DHE) was used to stain both BEL7402
and HepG2 cells after volasertib treatment. As shown in Figure 5, volasertib dose- and time-dependently enhanced the fluorescent intensi-
Figure 4. Volasertib promotes spindle abnormalities in HCC cells. BEL7402 (A) and HepG2 (B) cells were treated with 10 nM and 100 nM of volasertib for 24 h respectively. Cells were fixed and stained with Hoechst 33342 (to stain DNA, blue) as well as with β-tubulin antibody (to visualize tubulin spindles, red). Cells were analyzed by immu-nofluorescence microscopy. Arrows indicate the spindle abnormalities of cells.
Volasertib suppresses HCC growth
2482 Am J Cancer Res 2016;6(11):2476-2488
ty of DHE in both cells, suggesting that volaser-tib can enhance ROS accumulation in HCC cells.
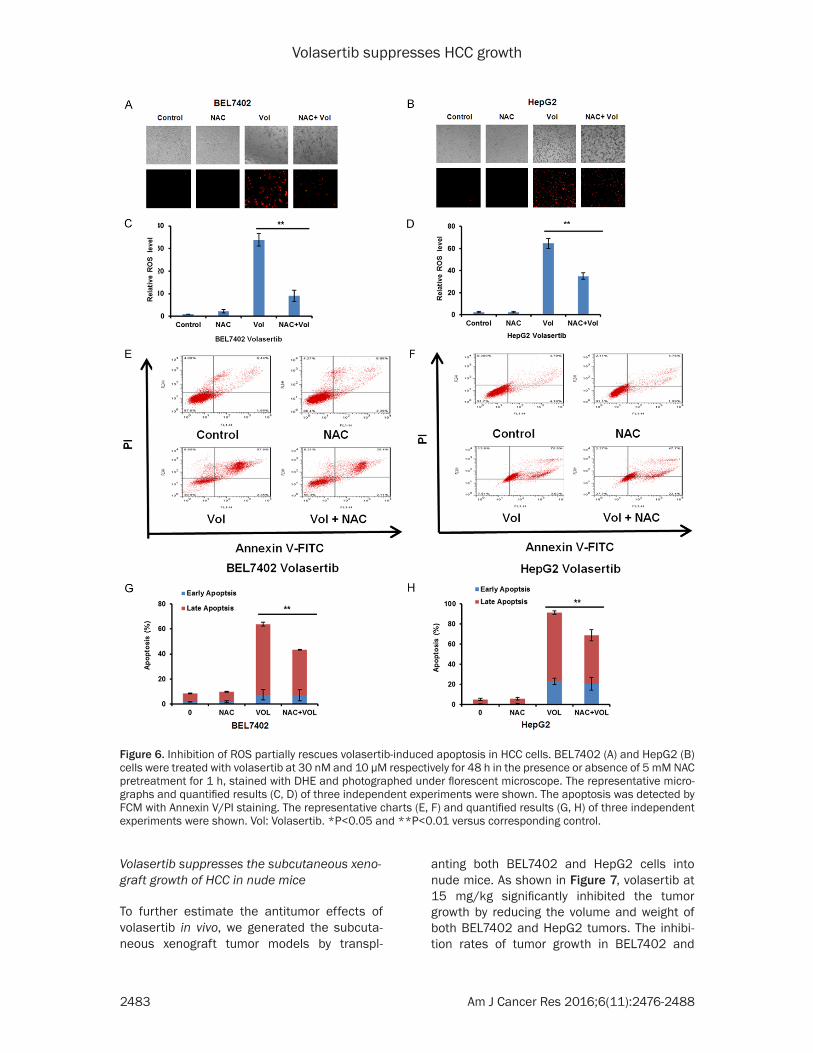
Inhibition of ROS partially rescues volasertib-induced apoptosis in HCC cells
To further study the role of ROS in volasertib-induced apoptosis in HCC cells, both BEL7402 and HepG2 cells were treated with volasertib
for 48 h with or without the ROS scavenger NAC at 5 mM pretreated for 1 h and stained with DHE. As shown in Figure 6A-D, the vo- lasertib-induced DHE fluorescent signal was dramatically reversed by NAC in both cells. Additionally, the volasertib-induced apoptosis was partially inverted by NAC (Figure 6E-H), suggesting volasertib can induce both ROS dependent and independent apoptosis in HCC cells.
Figure 5. Volasertib enhances ROS accumulation in HCC cells. BEL7402 and HepG2 cells were treated with volas-ertib at the indicated times and concentrations, stained with DHE and photographed under florescent microscope. The representative micrographs (bright field - A, B and fluorescence - C, D) and quantified results (E, F) of three independent experiments were shown. *P<0.05; **P<0.01 vs. corresponding control.
Volasertib suppresses HCC growth
2483 Am J Cancer Res 2016;6(11):2476-2488
Figure 6. Inhibition of ROS partially rescues volasertib-induced apoptosis in HCC cells. BEL7402 (A) and HepG2 (B) cells were treated with volasertib at 30 nM and 10 μM respectively for 48 h in the presence or absence of 5 mM NAC pretreatment for 1 h, stained with DHE and photographed under florescent microscope. The representative micro-graphs and quantified results (C, D) of three independent experiments were shown. The apoptosis was detected by FCM with Annexin V/PI staining. The representative charts (E, F) and quantified results (G, H) of three independent experiments were shown. Vol: Volasertib. *P<0.05 and **P<0.01 versus corresponding control.
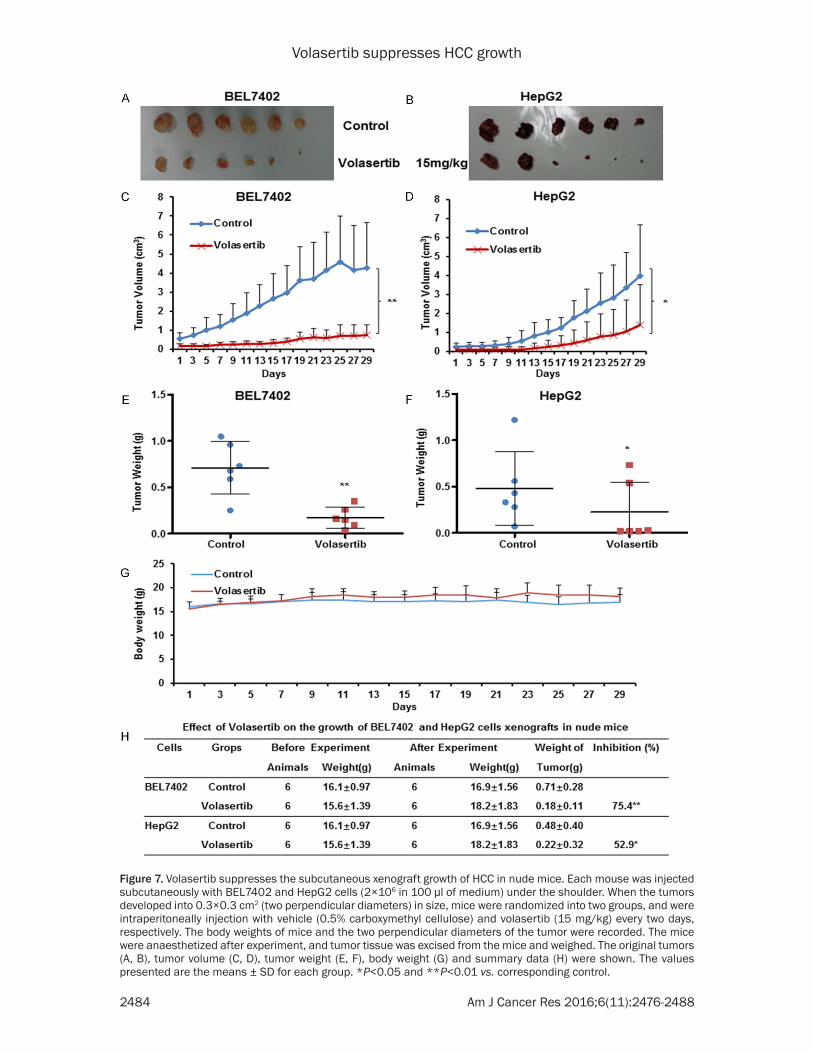
Volasertib suppresses the subcutaneous xeno-graft growth of HCC in nude mice
To further estimate the antitumor effects of volasertib in vivo, we generated the subcuta- neous xenograft tumor models by transpl-
anting both BEL7402 and HepG2 cells into nude mice. As shown in Figure 7, volasertib at 15 mg/kg significantly inhibited the tumor growth by reducing the volume and weight of both BEL7402 and HepG2 tumors. The inhibi-tion rates of tumor growth in BEL7402 and
Volasertib suppresses HCC growth
2484 Am J Cancer Res 2016;6(11):2476-2488
Figure 7. Volasertib suppresses the subcutaneous xenograft growth of HCC in nude mice. Each mouse was injected subcutaneously with BEL7402 and HepG2 cells (2×106 in 100 μl of medium) under the shoulder. When the tumors developed into 0.3×0.3 cm2 (two perpendicular diameters) in size, mice were randomized into two groups, and were intraperitoneally injection with vehicle (0.5% carboxymethyl cellulose) and volasertib (15 mg/kg) every two days, respectively. The body weights of mice and the two perpendicular diameters of the tumor were recorded. The mice were anaesthetized after experiment, and tumor tissue was excised from the mice and weighed. The original tumors (A, B), tumor volume (C, D), tumor weight (E, F), body weight (G) and summary data (H) were shown. The values presented are the means ± SD for each group. *P<0.05 and **P<0.01 vs. corresponding control.

Volasertib suppresses HCC growth
2485 Am J Cancer Res 2016;6(11):2476-2488
HepG2 was 75.4% and 52.9%, respectively. Ad- ditionally, the weight of nude mice did not exhibit the statistical difference during experi-ments, which suggests volasertib at the indi-cated dose did not cause toxicities in mice (Figure 7G).
Discussion
In this study, we show that volasertib can potently induce cell growth inhibition, cell cycle arrest at G2/M phase and apoptosis with abnormal spindle formation in human HCC cells in vitro and in vivo, which are consistent with previous reports that have demonstrated that volasertib is able to induce mitotic arrest and apoptosis by targeting PLK1 in several other cancer models [13, 14]. Furthermore, volasert-ib also increases the intracellular ROS levels, and pretreated with ROS scavenger NAC par-tially reverses volasertib-induced apoptosis. It has been reported that another PLK1 inhibitor BI 2536 alone or in combination with histone deacetylase (HDAC) inhibitor vorinostat could also enhance the intracellular ROS production to trigger cell death either in imatinib mesylate-sensitive or -resistant BCR/ABL+ leukemia cells, and the ROS scavenger Mn (III) tetrakis (4-ben-zoic acid)porphyrin chloride (MnTBAP) blocks ROS generation as well as lethality in BI 2536/vorinostat-treated cells [35]. Similarly, BI 2536 alone or combined with microtubule-destabiliz-ing drug vincristine dramatically promotes the production of ROS to induce cell apoptosis in rhabdomyosarcoma cells, and the antioxidants MnTBAP, NAC as well as α-tocopherol signifi-cantly rescue from BI 2536/vincristine-induced apoptosis [36]. Therefore, ROS plays an impor-tant role in cancer treatment with PLK1 inhibi-tors including volasertib, which usually triggers cancer cell death by increasing the intracellular ROS.
Currently, the clinical data indicate that volas-ertib has the promising clinical efficacy in a wide range of malignancies. The phase I clinical trials of volasertib show a favorable pharmaco-kinetic profile and encouraging preliminary anti-tumour activity with manageable toxicities such as anaemia, neutropenia and thrombocytope-nia [37-40]. But volasertib has insufficient anti-tumor activity as a monotherapy in the phase II trial of locally advanced or metastatic urothe-lial cancer [41]. Accordingly, multiple groups
have investigated the combined anticancer effects of volasertib and other chemotherapeu-tic drugs in the preclinical and clinical studies. Combination of volasertib with microtubule-interfering drugs including vincristine, vinblas-tine, vinorelbine or eribulin shows the synergis-tic induction of apoptosis in Ewing sarcoma and other pediatric malignant cells [42, 43]. Vol- asertib alone or in combination with azaciti-dine, cytarabine, decitabine and quizartinib is highly potent in multiple preclinical models of acute myeloid leukemia (AML) [44]. Moreover, volasertib powerfully inhibits the growth of bladder and cervical cancer cells, and potenti-ates the activity of cisplatin [17, 45]. Inspiringly, a recent phase I study has demonstrated that volasertib plus cisplatin or carboplatin has ma- nageable safety and antitumor activity without pharmacokinetics interference [46]. Another phase I study shows that volasertib combined with nintedanib also has manageable safety profile and antitumor effect without clearly pharmacokinetic disturbance [47]. But volaser-tib in combination with afatinib exhibits limited antitumor activity in a phase I study of 57 patients with advanced solid tumors [48]. In a phase II trial including 87 patients with AML, co-treatment with volasertib and cytarabine significantly prolongs median overall survival and median event-free survival but increases frequency of side effects compared with cyta-rabine alone [49]. But the results of another phase II trial in patients with advanced non-small-cell lung cancer exhibits that volasertib in combination with pemetrexed does not improve efficacy without increasing adverse events and pharmacokinetics interference compared with pemetrexed alone [50]. However, the combina-tion of volasertib with other anticancer agents for the treatment of HCC still needs to be test-ed in the future.
In summary, our study demonstrated that vola-sertib could potently induce cell growth inhibi-tion, cell cycle arrest and apoptosis in HCC cells in vitro and in vivo. The beneficial therapy of volasertib appears to be potential treatment strategy in patients with HCC.
Acknowledgements
This work was supported by funds from the Chinese National Natural Science Foundation No. 31271444 and No. 81201726 (Z. S.), and

Volasertib suppresses HCC growth
2486 Am J Cancer Res 2016;6(11):2476-2488
No. 31171304 (X. W.), the Guangdong Natural Science Funds for Distinguished Young Scholar No. 2014A030306001 (Z. S), the Guangdong Special Support Program for Young Talent No. 2015TQ01R350 (Z. S.) and the Science and Technology Program of Guangdong No. 2016A- 050502027 (Z. S.) and No. 2013B090500109 (X. W.), the Science and Technology Program of Guangzhou No. 201510010123 and No. 201300000041 (X. W.), and the Foundation for Research Cultivation and Innovation of Jinan University No. 21616119 (Z. S.).
Disclosure of conflict of interest
None.
Address correspondence to: Drs. Zhi Shi and Xing Wei, Department of Cell Biology & Institute of Bio- medicine, College of Life Science and Technology, Jinan University, National Engineering Research Center of Genetic Medicine, Guangdong Provincial Key Laboratory of Bioengineering Medicine, Room 708, The 2nd Engineer and Scientific Building, 601 Huangpu West Road, Guangzhou 510632, China. Tel: +86-20-852-245-25; Fax: +86-20-852-259-77; E-mail: [email protected] (ZS); [email protected] (XW)
References
[1] Siegel RL, Miller KD and Jemal A. Cancer sta-tistics, 2016. CA Cancer J Clin 2016; 66: 7-30.
[2] Bruix J, Sherman M; Practice Guidelines Com-mittee, American Association for the Study of Liver Diseases. Management of hepatocellular carcinoma. Hepatology 2005; 42: 1208-1236.
[3] Smith AD, Dunk AA, Tuttle-Newhall JE and Trot-ter JF. Hepatocellular carcinoma. Lancet 2004; 363: 898-899.
[4] Zitouni S, Nabais C, Jana SC, Guerrero A and Bettencourt-Dias M. Polo-like kinases: struc-tural variations lead to multiple functions. Nat Rev Mol Cell Biol 2014; 15: 433-452.
[5] Gjertsen BT and Schoffski P. Discovery and de-velopment of the Polo-like kinase inhibitor volasertib in cancer therapy. Leukemia 2015; 29: 11-19.
[6] Degenhardt Y and Lampkin T. Targeting Polo-like kinase in cancer therapy. Clin Cancer Res 2010; 16: 384-389.
[7] Strebhardt K and Ullrich A. Targeting polo-like kinase 1 for cancer therapy. Nat Rev Cancer 2006; 6: 321-330.
[8] He ZL, Zheng H, Lin H, Miao XY and Zhong DW. Overexpression of polo-like kinase1 predicts a
poor prognosis in hepatocellular carcinoma patients. World J Gastroenterol 2009; 15: 4177-4182.
[9] Sun W, Su Q, Cao X, Shang B, Chen A, Yin H and Liu B. High expression of polo-like kinase 1 is associated with early development of he-patocellular carcinoma. Int J Genomics 2014; 2014: 312130.
[10] Mok WC, Wasser S, Tan T and Lim SG. Polo-like kinase 1, a new therapeutic target in hepato-cellular carcinoma. World J Gastroenterol 2012; 18: 3527-3536.
[11] He Z, Wu J, Dang H, Lin H, Zheng H and Zhong D. Polo-like kinase 1 contributes to the tumori-genicity of BEL-7402 hepatoma cells via regu-lation of Survivin expression. Cancer Lett 2011; 303: 92-98.
[12] Studach LL, Rakotomalala L, Wang WH, Hull-inger RL, Cairo S, Buendia MA and Andrisani OM. Polo-like kinase 1 inhibition suppresses hepatitis B virus X protein-induced transforma-tion in an in vitro model of liver cancer progres-sion. Hepatology 2009; 50: 414-423.
[13] Rudolph D, Steegmaier M, Hoffmann M, Grau-ert M, Baum A, Quant J, Haslinger C, Garin-Chesa P and Adolf GR. BI 6727, a Polo-like ki-nase inhibitor with improved pharmacokinetic profile and broad antitumor activity. Clin Can-cer Res 2009; 15: 3094-3102.
[14] Gorlick R, Kolb EA, Keir ST, Maris JM, Reynolds CP, Kang MH, Carol H, Lock R, Billups CA, Kur-masheva RT, Houghton PJ and Smith MA. Initial testing (stage 1) of the Polo-like kinase inhibi-tor volasertib (BI 6727), by the Pediatric Pre-clinical Testing Program. Pediatr Blood Cancer 2014; 61: 158-164.
[15] Liu X. Targeting Polo-Like Kinases: A Promising Therapeutic Approach for Cancer Treatment. Transl Oncol 2015; 8: 185-195.
[16] Brandwein JM. Targeting polo-like kinase 1 in acute myeloid leukemia. Ther Adv Hematol 2015; 6: 80-87.
[17] Xie FF, Pan SS, Ou RY, Zheng ZZ, Huang XX, Jian MT, Qiu JG, Zhang WJ, Jiang QW, Yang Y, Li WF, Shi Z and Yan XJ. Volasertib suppresses tumor growth and potentiates the activity of cisplatin in cervical cancer. Am J Cancer Res 2015; 5: 3548-3559.
[18] W CS. Tables for convenient calculation of me-dian-effective dose (LD50 or ED50) and in-structions in their use. Biometrics 1952; 8: 249-263.
[19] Shi Z, Tiwari AK, Shukla S, Robey RW, Singh S, Kim IW, Bates SE, Peng X, Abraham I, Ambud-kar SV, Talele TT, Fu LW and Chen ZS. Sildenafil reverses ABCB1- and ABCG2-mediated chemo-therapeutic drug resistance. Cancer Res 2011; 71: 3029-3041.

Volasertib suppresses HCC growth
2487 Am J Cancer Res 2016;6(11):2476-2488
[20] Chou TC, Talalay P. Quantitative analysis of dose-effect relationships: the combined ef-fects of multiple drugs or enzyme inhibitors. Adv Enzyme Regul 1984; 22: 27-55.
[21] Jiang QW, Cheng KJ, Mei XL, Qiu JG, Zhang WJ, Xue YQ, Qin WM, Yang Y, Zheng DW, Chen Y, Wei MN, Zhang X, Lv M, Chen MW, Wei X and Shi Z. Synergistic anticancer effects of trip-tolide and celastrol, two main compounds from thunder god vine. Oncotarget 2015; 6: 32790-32804.
[22] Lv M, Qiu JG, Zhang WJ, Jiang QW, Qin WM, Yang Y, Zheng DW, Chen Y, Huang JR, Wang K, Wei MN, Cheng KJ and Shi Z. Wallichinine re-verses ABCB1-mediated cancer multidrug re-sistance. Am J Transl Res 2016; 8: 2969-80.
[23] Chen X, Gong L, Ou R, Zheng Z, Chen J, Xie F, Huang X, Qiu J, Zhang W, Jiang Q, Yang Y, Zhu H, Shi Z and Yan X. Sequential combination therapy of ovarian cancer with cisplatin and gamma-secretase inhibitor MK-0752. Gynecol Oncol 2016; 140: 537-544.
[24] Shi Z, Li Z, Li ZJ, Cheng K, Du Y, Fu H and Khuri FR. Cables1 controls p21/Cip1 protein stability by antagonizing proteasome subunit alpha type 3. Oncogene 2015; 34: 2538-2545.
[25] Shi Z, Liang YJ, Chen ZS, Wang XW, Wang XH, Ding Y, Chen LM, Yang XP and Fu LW. Reversal of MDR1/P-glycoprotein-mediated multidrug resistance by vector-based RNA interference in vitro and in vivo. Cancer Biol Ther 2006; 5: 39-47.
[26] Gong LH, Chen XX, Wang H, Jiang QW, Pan SS, Qiu JG, Mei XL, Xue YQ, Qin WM, Zheng FY, Shi Z and Yan XJ. Piperlongumine induces apopto-sis and synergizes with cisplatin or paclitaxel in human ovarian cancer cells. Oxid Med Cell Longev 2014; 2014: 906804.
[27] Chen XX, Xie FF, Zhu XJ, Lin F, Pan SS, Gong LH, Qiu JG, Zhang WJ, Jiang QW, Mei XL, Xue YQ, Qin WM, Shi Z and Yan XJ. Cyclin-dependent kinase inhibitor dinaciclib potently synergizes with cisplatin in preclinical models of ovarian cancer. Oncotarget 2015; 6: 14926-14939.
[28] Luo Y, Jiang QW, Wu JY, Qiu JG, Zhang WJ, Mei XL, Shi Z and Di JM. Regulation of migration and invasion by Toll-like receptor-9 signaling network in prostate cancer. Oncotarget 2015; 6: 22564-22574.
[29] Zhang J, Lan SJ, Liu QR, Liu JM and Chen XQ. Neuroglobin, a novel intracellular hexa-coordi-nated globin, functions as a tumor suppressor in hepatocellular carcinoma via Raf/MAPK/Erk. Mol Pharmacol 2013; 83: 1109-1119.
[30] Shi Z, Park HR, Du Y, Li Z, Cheng K, Sun SY, Fu H and Khuri FR. Cables1 complex couples sur-vival signaling to the cell death machinery. Cancer Res 2015; 75: 147-158.
[31] Shi Z, Tiwari AK, Shukla S, Robey RW, Kim IW, Parmar S, Bates SE, Si QS, Goldblatt CS, Abra-
ham I, Fu LW, Ambudkar SV and Chen ZS. In-hibiting the function of ABCB1 and ABCG2 by the EGFR tyrosine kinase inhibitor AG1478. Biochem Pharmacol 2009; 77: 781-793.
[32] Mei XL, Yang Y, Zhang YJ, Li Y, Zhao JM, Qiu JG, Zhang WJ, Jiang QW, Xue YQ, Zheng DW, Chen Y, Qin WM, Wei MN and Shi Z. Sildenafil inhib-its the growth of human colorectal cancer in vitro and in vivo. Am J Cancer Res 2015; 5: 3311-3324.
[33] Qiu JG, Zhang YJ, Li Y, Zhao JM, Zhang WJ, Ji-ang QW, Mei XL, Xue YQ, Qin WM, Yang Y, Zheng DW, Chen Y, Wei MN and Shi Z. Tra-metinib modulates cancer multidrug resis-tance by targeting ABCB1 transporter. Onco-target 2015; 6: 15494-15509.
[34] Xue YQ, Di JM, Luo Y, Cheng KJ, Wei X and Shi Z. Resveratrol oligomers for the prevention and treatment of cancers. Oxid Med Cell Longev 2014; 2014: 765832.
[35] Dasmahapatra G, Patel H, Nguyen T, Attkisson E and Grant S. PLK1 inhibitors synergistically potentiate HDAC inhibitor lethality in imatinib mesylate-sensitive or -resistant BCR/ABL+ leu-kemia cells in vitro and in vivo. Clin Cancer Res 2013; 19: 404-414.
[36] Hugle M, Belz K and Fulda S. Identification of synthetic lethality of PLK1 inhibition and mi-crotubule-destabilizing drugs. Cell Death Differ 2015; 22: 1946-1956.
[37] Schoffski P, Awada A, Dumez H, Gil T, Bar-tholomeus S, Wolter P, Taton M, Fritsch H, Glomb P and Munzert G. A phase I, dose-esca-lation study of the novel Polo-like kinase inhibi-tor volasertib (BI 6727) in patients with ad-vanced solid tumours. Eur J Cancer 2012; 48: 179-186.
[38] Lin CC, Su WC, Yen CJ, Hsu CH, Su WP, Yeh KH, Lu YS, Cheng AL, Huang DC, Fritsch H, Voss F, Taube T and Yang JC. A phase I study of two dosing schedules of volasertib (BI 6727), an intravenous polo-like kinase inhibitor, in pa-tients with advanced solid malignancies. Br J Cancer 2014; 110: 2434-2440.
[39] Nokihara H, Yamada Y, Fujiwara Y, Yamamoto N, Wakui H, Nakamichi S, Kitazono S, Inoue K, Harada A, Taube T, Takeuchi Y and Tamura T. Phase I trial of volasertib, a Polo-like kinase in-hibitor, in Japanese patients with advanced solid tumors. Invest New Drugs 2016; 34: 66-74.
[40] Kobayashi Y, Yamauchi T, Kiyoi H, Sakura T, Hata T, Ando K, Watabe A, Harada A, Taube T, Miyazaki Y and Naoe T. Phase I trial of volaser-tib, a Polo-like kinase inhibitor, in Japanese patients with acute myeloid leukemia. Cancer Sci 2015; 106: 1590-1595.
[41] Stadler WM, Vaughn DJ, Sonpavde G, Vogel-zang NJ, Tagawa ST, Petrylak DP, Rosen P, Lin

Volasertib suppresses HCC growth
2488 Am J Cancer Res 2016;6(11):2476-2488
CC, Mahoney J, Modi S, Lee P, Ernstoff MS, Su WC, Spira A, Pilz K, Vinisko R, Schloss C, Frit-sch H, Zhao C and Carducci MA. An open-label, single-arm, phase 2 trial of the Polo-like kinase inhibitor volasertib (BI 6727) in patients with locally advanced or metastatic urothelial can-cer. Cancer 2014; 120: 976-982.
[42] Weiss LM, Hugle M, Romero S and Fulda S. Synergistic induction of apoptosis by a polo-like kinase 1 inhibitor and microtubule-inter-fering drugs in Ewing sarcoma cells. Int J Can-cer 2016; 138: 497-506.
[43] Abbou S, Lanvers-Kaminsky C, Daudigeos-Du-bus E, LE Dret L, Laplace-Builhe C, Molenaar J, Vassal G, Geoerger B; within the ITCC Biology and Preclinical Evaluation Committee. Polo-like Kinase Inhibitor Volasertib Exhibits Antitu-mor Activity and Synergy with Vincristine in Pediatric Malignancies. Anticancer Res 2016; 36: 599-609.
[44] Rudolph D, Impagnatiello MA, Blaukopf C, Sommer C, Gerlich DW, Roth M, Tontsch-Grunt U, Wernitznig A, Savarese F, Hofmann MH, Al-brecht C, Geiselmann L, Reschke M, Garin-Chesa P, Zuber J, Moll J, Adolf GR and Kraut N. Efficacy and mechanism of action of volasert-ib, a potent and selective inhibitor of Polo-like kinases, in preclinical models of acute myeloid leukemia. J Pharmacol Exp Ther 2015; 352: 579-589.
[45] Brassesco MS, Pezuk JA, Morales AG, de Oliveira JC, Roberto GM, da Silva GN, Francisco de Oliveira H, Scrideli CA and Tone LG. In vitro targeting of Polo-like kinase 1 in bladder carci-noma: comparative effects of four potent in-hibitors. Cancer Biol Ther 2013; 14: 648-657.
[46] Awada A, Dumez H, Aftimos PG, Costermans J, Bartholomeus S, Forceville K, Berghmans T, Meeus MA, Cescutti J, Munzert G, Pilz K, Liu D and Schoffski P. Phase I trial of volasertib, a Polo-like kinase inhibitor, plus platinum agents in solid tumors: safety, pharmacokinetics and activity. Invest New Drugs 2015; 33: 611-620.
[47] de Braud F, Cascinu S, Spitaleri G, Pilz K, Clem-enti L, Liu D, Sikken P and De Pas T. A phase I, dose-escalation study of volasertib combined with nintedanib in advanced solid tumors. Ann Oncol 2015; 26: 2341-6.
[48] Machiels JP, Peeters M, Herremans C, Sur-mont V, Specenier P, De Smet M, Pilz K, Strel-kowa N, Liu D and Rottey S. A phase I study of volasertib combined with afatinib, in advanced solid tumors. Cancer Chemother Pharmacol 2015; 76: 843-851.
[49] Dohner H, Lubbert M, Fiedler W, Fouillard L, Haaland A, Brandwein JM, Lepretre S, Reman O, Turlure P, Ottmann OG, Muller-Tidow C, Kramer A, Raffoux E, Dohner K, Schlenk RF, Voss F, Taube T, Fritsch H and Maertens J. Ran-domized, phase 2 trial of low-dose cytarabine with or without volasertib in AML patients not suitable for induction therapy. Blood 2014; 124: 1426-1433.
[50] Ellis PM, Leighl NB, Hirsh V, Reaume MN, Blais N, Wierzbicki R, Sadrolhefazi B, Gu Y, Liu D, Pilz K and Chu Q. A Randomized, Open-Label Phase II Trial of Volasertib as Monotherapy and in Combination With Standard-Dose Peme-trexed Compared With Pemetrexed Monother-apy in Second-Line Treatment for Non-Small-Cell Lung Cancer. Clin Lung Cancer 2015; 16: 457-465.