interleukin 10 suppresses tumor growth and …. 2, 1969-1979, december 1996 clinical cancer research...
TRANSCRIPT

Vol. 2, 1969-1979, December 1996 Clinical Cancer Research 1969
Interleukin 10 Suppresses Tumor Growth and Metastasis of Human
Melanoma Cells: Potential Inhibition of Angiogenesis1
Suyun Huang, Keping Xie, Corazon D. Bucana,
Stephen E. Ullrich, and Menashe Bar-Eli2
Departments of Cell Biology [S. H., K. X., C. D. B., M. B-E.l and
Immunology [S. E. UI, The University of Texas M. D. Anderson
Cancer Center, Houston. Texas 77030
ABSTRACT
Interleukin 10 (IL-lO) inhibits the production of a wide
range of cytokines in various cell types. The purpose of this
study was to determine whether the expression of the IL-JO
gene can influence tumor growth and metastatic properties
of human melanoma cells. The human melanoma cell line,
A375P, which does not produce endogenous IL-b, was
transfected with a hygromycin expression vector (control) or
a vector containing full-length murine IL-lO cDNA. A375P
parental cells, A375P-Hygro, and A375P-IL-l0-positive cells
were injected s.c. and i.v. into nude mice. A375P-IL-10 cells
produced significantly slower growing s.c. tumors and fewer
lung metastases than control cells. The tumorigenicity of the
human melanoma A375SM and the murine melanoma B16-
BL6 cells was also significantly inhibited when they were
admixed with A375P-IL-10 but not with A375P-Hygro be-
fore s.c. injection into nude mice. The suppression of tumor
growth and metastasis was directly correlated with a de-
crease in neovascularity determined by immunostaining
with anti-factor VIII. Because tumor-associated macro-
phages are the major source of angiogenic molecules in
melanoma, we used reverse transcription-PCR to demon-
strate that IL-lO down-regulates the production of vascular
endothelial growth factor, the most potent angiogenic factor
in activated macrophages. Other factors involved in angio-
genesis such as IL-1f�, tumor necrosis factor-a, IL-6, and the
proteinase matrix metalloproteinase-9 were also inhibited in
activated macrophages by supernatants from A375P-IL-10
cells Collectively, these data suggest that the production of
IL-lO by tumor cells inhibits macrophages-derived angio-
genic factors, and hence, tumor growth and metastasis.
INTRODUCTION
IL3- I 0 was first described as a product secreted by munne
Th2 clones of helper T cells that suppressed cytokine production
by Thi clones (1); therefore, IL-lO was designated as cytokine
synthesis inhibition factor (2-4). Subsequently, studies have
shown that IL- [0 inhibits the production of a wide range of
cytokines by various cell types involved in the immune re-
sponse. For example, IL-b inhibits the production of IL-2,
IL-3, lymphotoxin, IFN-�1’, and granulocyte-macrophage cob-
ny-stimulating factor in Thl cells (1, 5); IL-4 and IL-S in Th2
cells (6); and IFN-�y and TNF-a in NK cells (7, 8). A key
determinant of the role of IL-b in the regulation of immune
responses may lie in its deactivating effects on macrophages.
Indeed, in macrophages/monocytes, IL- 10 inhibits the produe-
(ion of IL-l, IL-6, IL-8, TNF-a, and MMP-9 (3, 9-12), down-
regulates the expression of the MHC class II (I 3), and sup-
presses the production of reactive nitrogen oxides (14).
Because macrophages are a major component of the lym-
phoreticular infiltrate of melanoma and because many of the
cytokines inhibited by IL-lO (IL-l, IL-6, IL-8, and TNF-a) and
the proteinase MMP-9 are important in angiogenesis, we have
initiated studies to examine the role of IL-lO in the growth and
metastasis of human melanoma tumors. Angiogenesis is a crit-
ical component of the metastatic pathway; malignant tumors do
not grow beyond 2-3 mm3 in size unless they stimulate the
formation of new blood vessels (15). The development of new
vaseulature also provides an essential exit route for metastasiz-
ing tumor cells from the primary tumor to the bloodstream to
colonize in distant organs (IS, 16). A recent report by Richter et
a!. (17) showed that transduction of Chinese hamster ovary
(CHO) cells to express the murine IL-JO gene resulted in tumor
suppression in nude mice (17).
In the present report, we show that expression of IL-b in
human melanoma cells suppressed their tumorigenicity and met-
astatic potential in nude mice. We provide evidence that IL-lO
exerts its antitumor and antimetastatic activity by inhibiting
angiogenesis in vivo. The in vivo decrease in vaseularization
found in IL-lO-secreting tumors is most likely due to the ability
of IL-lO to down-regulate the synthesis of VEGF and other
angiogenie cytokines produced by macrophages. Thus. IL-lO
may be an effective antitumor and antimetastatic agent.
Received 5/22/96: revised 9/1 3/96: accepted 9/23/96.
I This work was supported in part by NIH Grant CA 41525, the Wendy
Will Case Cancer Fund, Inc. (to M. B. E.), and NIH Grant AR 40824 (to
S. E. U.).2 To whom requests for reprints should be addressed, at Department of Cell
Biology. Box 173. The University ofTexas M. D. Anderson Cancer Center.
15 15 Holcombe Boulevard. Houston, TX 77030. Phone: (7 13) 794-4004:
Fax: (7 13) 792-8747: E-mail: AN [email protected].
3 The abbreviations used are: IL. interleukin: rMuIL. recombinant murine
IL: TNF, tumor necrosis factor; NK, natural killer: MMP, matrix metalbo-
proteinase: VEGF. vascular endothelial growth factor: FBS, fetal bovine
serum; iNOS, inducible nitric oxide synthase: bFGF, basic fibroblast
growth factor: MU, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide: LPS. lipopolysaceharide; RT-PCR. reverse transcription-PCR:
G3PDH, glyceraldehyde-3-phosphate dehydrogenase.
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

1970 Antiangiogenic Effect of Interleukin 10
MATERIALS AND METHODS
Animals. Male athymic BALB/c nude mice were pur-
chased from the Animal Production Area of the National Cancer
Institute, Frederick Cancer Research Facility (Frederick, MD).
The mice were housed in laminar flow cabinets under specific
pathogen-free conditions and used at 8 weeks of age. Animals
were maintained in facilities approved by the American Asso-
ciation for Accreditation of Laboratory Animal Care and in
accordance with current regulations and standards of the United
States Department of Agriculture, Department of Health and
Human Services, and National Institutes of Health.
Cell Lines and Culture Conditions. The A37SP human
melanoma cell line was originally established in culture from a
lymph node metastasis of a 54-year-old melanoma patient. The
highly metastatic derivative A37SSM was established from
pooled lung metastases produced by the A37SP cells injected
iv. into nude mice (18). The murine Bl6-BL6 cell line was
established in culture after injecting B16 cells into mouse blad-
der in vitro (19). B16-BL6 cells are highly tumorigenic and
highly invasive in vivo (19). The cells were maintained in
culture as adherent monolayers in Eagle’s MEM supplemented
with 10% FBS, sodium pyruvate, nonessential amino acids,
L-glutamine, 2-fold vitamin solution, and penicillin-streptomy-
cm (Flow Laboratories, Rockville, MD). All cultures were free
of Mycoplasma and pathogenic murine viruses (assayed by
Microbiological Associates, Bethesda, MD). Cultures were
maintained for no longer than 4 weeks after recovery from
frozen stocks.
Expression Vectors and Cell Transfections. A full-
length murine IL-lO eDNA was cloned into the PvuII-XhoI site
of the pCEP4 expression vector (20). The expression of IL-lO
eDNA was under the control of the cytomegalovirus promoter.
A37SP cells were transfected by using the Lipofectamine rea-
gent (Life Technologies, Inc., Gaithersburg, MD) and 1-2 p.g of
DNA. Transfections were carried out as we described previously
(21-23). Three days after transfection, hygromycin (Boehringer
Mannheim Corp., Indianapolis, IN) was added at a final con-
centration of ISO p.g/ml, and cultures were maintained in this
medium. Fourteen days later, hygromycin-resistant clones were
isolated by trypsinization and established in culture.
ELISA for Murine IL-lO. The production of IL-lO by
the transfected cells was measured by ELISA with a kit pur-
chased from Endogen (Cambridge, MA) and performed accord-
ing to the manufacturer’s instructions as described previously
(24). A standard curve using authentic rMuIL-lO (Genzyme,
Cambridge, MA) with specific activity of S X l0� units/mg was
used for quantitation. Results are expressed as nanograms of
IL-l0/l06 cells/24 h. IL-b could not be detected in conditioned
medium from parental or control Hygro-transfected cells.
Northern Blot Analysis. Poly(A)� mRNA was ex-
tracted from macrophages or tumor cells growing in culture
using the FastTrack mRNA isolation kit (Invitrogen Co., San
Diego, CA). mRNA was electrophoresed on a 1% denaturing
formaldehyde/agarose gel, electrotransferred at 0.6 amp to
GeneScreen nylon membrane (DuPont/NEN, Boston, MA), and
UV cross-linked with 120,000 1iJ/cm2 using a UV Stratalinker
1800 (Stratagene, La Jolla, CA). Hybridizations were performed
as described previously (25-27). The eDNA probes used in
these analyses were a 1 .3-kb PstI eDNA fragment correspond-
ing to rat glyeeraldehyde 3-phosphate dehydrogenase (28), a
1.0-kb MMP-9 eDNA probe (12), a 0.6-kb BatnHI-KpnI eDNA
fragment of the murine IL-JO gene, (20), a 4.0-kb XbaI-HindIII
fragment of iNOS (29), and a 1 .4-kb EcoRI eDNA fragment of
bovine bFGF (30). Each eDNA probe was purified by agarose
gel electrophoresis and recovered by using GeneClean (BIO
101, Inc., La Jolla, CA). Specific oligonueleotide probes for
IL-I, IL-6, and TNF-a were purchased from R & D Systems
(Minneapolis, MN). All probes were radiolabeled by using the
random primer technique with [a-32P]deoxyribonueleotide
triphosphates.
In Vitro Growth Rates. Tumor cells were suspended in
MEM containing 10% FBS and plated at a concentration of 2 X
l0� cells/well in six-well tissue culture plates (Costar, Cam-
bridge, MA). Three wells of cells were trypsinized, cells were
counted at 12, 24, 48, and 72 h, and the doubling time was
calculated. In vitro growth rates were also measured by the
[3H]thymidine uptake and the MIT tests. In the [3Hjthymidine
uptake test, S X i0� cells/well were incubated in 96-well plates
with 200 p.1 of MEM containing 10% FBS for 72 h. Five-tenths
�.tCi of [3H]thymidine in 20 pA of MEM was added to each well.
After 18 h incubation, cells were harvested onto glass fiber
sheets. After drying, filters were placed in plastic bags, and 10
ml of scintillation fluid were added to each bag and sealed by
heat. Incorporated radioactivity was measured using a �3 counter
(1205 Betaplate; Wallac Oy, Finland) and reported as epm. Cell
viability was also determined by the MTT (Sigma Chemical
Co., St. Louis, MO) assay. MiT was added at a concentration of
0.1 mg/mI for 2 h at 37#{176}C,and absorbanee was determined at
450 nm.
Tumor Cell Injections. To prepare tumor cells for moe-
ulation, cells in exponential growth phase were harvested by
brief exposure to 0.25% trypsin-0.02% EDTA solution (w/v).
The flask was sharply tapped to dislodge the cells, and supple-
mented medium was added. The cell suspension was pipetted to
produce a single-cell suspension. The cells were washed and
resuspended in Ca2�- and Mg2�-free HBSS to the desired cell
concentration. Cell viability was determined by trypan blue
exclusion, and only single-cell suspensions of >90% viability
were used. s.c. tumors were produced by injecting 1 X 106
tumor cells (A37SP, A375P-Hygro, and A37SP-IL- 10) in 0.2 ml
HBSS over the right scapular region. The growth of s.c. tumors
was monitored by weekly examination of the mice and meas-
urement of tumors with calipers (31).
The tumorigenicity of A37SSM cells was determined by
s.c. injection of 1 X 106 A37SSM alone, A37SP-Hygro alone,
A37SP-IL-l0 alone, and I X 106 A37SSM cells admixed with
1 X 106 A37SP-Hygro cells or 1 X 106 A37SSM cells admixed
with A37SP-IL-l0 cells (ratio of I : I , all in 0.2 ml of HBSS) into
BALB/e nude mice. The mice were examined weekly, and the
tumors were measured over a period of S weeks. The tumori-
genicity of the murine B16-BL6 melanoma cells was deter-
mined by s.c. injection of 0.2 X 106 Bl6-BL6 cells alone, 1 X
106 A37SP-Hygro alone, I X 106 A37SP-IL-lO alone, and
0.2 X 106 Bl6-BL6 admixed with 1 X 106 A37SP-Hygro cells
or0.2 X 106 Bl6-BL6 cells admixed with I X 106 A37SP-IL-l0
cells (ratio of 1:5, all in 0.2 ml HBSS). Since B16-BL6 is a
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Clinical Cancer Research 1971
fast-growing tumor in nude mice, the mice were examined every
3 days, and the tumors were measured over a period of 16 days.
Experimental Lung Metastasis. The production of ex-
perimental lung metastases was evaluated by injection of 1 X
106 A37SP cells, A37SP-Hygro cells, and A37SP-IL-l0 cells
into the tail vein of nude mice, all in 0.2 ml of HBSS. The mice
were killed when moribund or 6 weeks after injection. The lungs
were removed, rinsed in HBSS, and fixed in Bouin’s solution.
The number of tumor colonies was determined under a dissect-
ing microscope, as we described previously (21-23).
Immunohistochemistry. Cryostat sections of tissues
were fixed with 4% paraformaldehyde in PBS (pH 7.5) for 10
mm at room temperature, washed twice with PBS, and treated
with 1% Triton X-lOO for S mm. The sections were washed
three times, and endogenous peroxidase was blocked with 3%
hydrogen peroxide in methanol for I 2 mm. The samples were
then washed three times with PBS and incubated with a protein-
blocking solution consisting of PBS containing 1% normal goat
serum and 10% horse serum for 20 mm at room temperature.
Excess blocking reagent was drained off, and the samples were
incubated with the appropriate dilution of horseradish peroxi-
dase-conjugated rabbit anti-factor VIII-related antigen (Dako
Corp., Carpinteria, CA) for 15-18 h at 4#{176}C.The samples were
rinsed four times with PBS, rinsed briefly with distilled water,
and incubated with 3,3’-diaminobenzidine (Research Genetics,
Huntsville, AL) for 20 mm at room temperature. The sections
were then washed three times with distilled water, counter-
stained with aqueous hematoxylin, washed, mounted with Per-
mount, and examined in a bright-field microscope. The positive
reaction was indicated by a reddish-brown precipitate. For each
tumor injected, at least 10 microscopic fields were examined
using a X40 objective.
Frozen sections were cut and stained for infiltrating macro-
phages with F4/80 rat monoclonal antibody, which binds to a Mr
160,000 plasma membrane glycoprotein present on mouse
mononuclear phagocytes (32). The hybridoma cells producing
the antibody were the gift of Dr. David Hume (University of
Queensland, Queensland, Australia). The sections were then
incubated with peroxidase-conjugated mouse antirat immuno-
globulin antibody (Boehringer Mannheim Corp.). Immunohis-
tochemical staining was evaluated in at least I 0 random micro-
scopic fields (Nikon Optiphot microscope), as we described
previously (22). Sections obtained from the same portion of a
given group of tumors exhibited a uniform pattern of macro-
phage infiltration; thus, we are confident that the analysis done
in a given section is representative of the tumor being tested.
The images were projected and recorded by digitizing the image
using Sony 3 CCD color video camera and an Optimas Image
Analysis software (Optimas Corp., Bothell, WA) and a digital
printer (Sony Corp., Tokyo, Japan).
Isolation and Treatment of Macrophages. Peritoneal
exudate macrophages were collected by peritoneal lavage from
mice given an i.p. injection of 2.0 ml of thioglycollate broth
(Baltimore Biological Laboratory, Coekeysville, MD) 4 days
before the harvest. The peritoneal exudate macrophages were
washed in Ca2�- and Mg2�-free HBSS and resuspended in
serum-free medium. Cells (4 X l0�) were plated into each tissue
culture dish (150 X 25 mm: Corning Inc., Corning, NY). After
a 3-h incubation, the nonadherent cells were removed by wash-
ing with medium. More than 98% of the adherent cell popula-
tions were macrophages according to morphological, immuno-
cytochemical, and phagocytie criteria. These cultures were then
treated with LPS (200 ng/ml) alone or LPS plus supernatants
from A37SP melanoma cells transfected with the control hvgro
gene or the IL-JO gene for 24 h. The macrophages were then
harvested, and mRNA was isolated for Northern blot analysis.
Semiquantitative RT-PCR Assay for the Expression of
VEGF in IL-lO-treated Macrophages. The effect of
rMuIL-lO on the expression of VEGF in macrophages was
measured by multiple RT-PCR for VEGF and the mouse
G3PDH gene (control amplimer set). Poly(A)� mRNA was
extracted from untreated and IL-lO-treated (25, 50, and 100
ng/ml per 72 h) macrophages as described above. eDNA was
synthesized from the isolated mRNA by reverse transcription in
20-pA reactions using the Promega Reverse Transcription Sys-
tem (Promega Corp. , Madison, WI) as recommended by the
manufacturer. PCR was run for 30 cycles at 93#{176}C(30 s), 57#{176}C
(I mm), and 72#{176}C(1 mm) with 3 �i.l of the eDNA 2.5 units
Taq Polymerase (Promega Corp.) and VEGF primers 5’ to 3’
of sense CACATAGGAGAGATGAGCTTC and antisense
CCGCCTCGGCTTGTCACAT (33). The primers correspond to
exons 4 and 8 of the VEGF gene. After one round of PCR, two
major bands of 230 and 300 bp were identified corresponding to
VEGF165 and VEGF189, respectively (34). We coamplified in
the same tube a fragment of 983 bp corresponding to the mouse
G3PDH gene (Clontech, Palo Alto, CA) for positive control and
to monitor genomic DNA contamination. Since the G3PDH
fragment spans exon-intron boundaries, the generated 983-bp
fragment could be obtained only from a reverse-transcribed
eDNA. The two primers used to amplify the G3PDH fragment
are: sense S ‘-TGAAGGTCGGTGTGAACGGATTTGGC-3’
and antisense 5’-CATGTAGGCCATGAGGTCCACCAC-3’.
When the primers for VEGF and G3PDH were added at the
beginning of the reaction, amplification was linear for both
genes over 30 PCR cycles. In addition, to further ensure that our
eDNA were not contaminated with genomic DNA, each sample
was amplified with a set of primers corresponding to the f3-actin
gene. These primers will generate a 62 1-bp fragment from
eDNA and a 73l-bp fragment from genomic DNA to include the
120-bp of intron S. The two �3-actin primers are: sense
5’ACACTGTGCCCATCTACGAGG-3’ and antisense 5’-
AGGGGCCGGACTCGTCATACT-3’. The human colon carci-
noma cell line SW620 served as a positive control for the
expression of VEGF.
Statistical Analysis. The in vitro data were analyzed for
significance by the Student’s t test (two-tailed), and the in viva
data were analyzed by the Mann-Whitney test.
RESULTS
Expression of IL-lO in A375P Human Melanoma Cells.
Human A375P melanoma cells were selected for these studies
because preliminary experiments had shown that these cells did
not express or secrete endogenous IL-lO. Following gene trans-
fection with the eDNA encoding murine IL-lO, seven hygro-
resistant clones transfected with the pCEP4-IL-l0 expression
vector and one control clone transfeeted with the naked expres-
sion vector alone (without IL-b eDNA) were analyzed for
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

a
IL-lO
0)0)ECu
EC.)
0)N
(1)
0
E
I-
0 1 2 3 4 5 6 7 8
1972 Antiangiogenic Effect of Interleukin 10
4 .s
-I-..vyw vwwww
123456789
IL-lO
GAPDH
Fig. 1 Northern blot analysis for the expression of murine IL-b inparental A375P cells (Lane 1), in Hygro-transfected cells (Lane 2), and
in seven pCEP4-IL-l0-transfected clones (Lanes 3-9). The same blot
was hybridized with a G3PDH (GAPDI-I) probe to verify that intact
RNA in similar amounts was loaded in each lane.
Time after Tumor Injection (week)
Fig. 2 The tumorigenicity of A375P parental cells (0), two Hygro-transfected clones Hl and H2 (LI)’ and five A375P-IL-l0 clones 1-1
through I-S (#{149})7 weeks after s.c. injection into nude mice. Tumor cells(1 X 106) in 0.2 ml of HBSS were injected over the right scapular
region. The growth of s.c. tumors was monitored by weekly examination
and measurement of tumors with calipers.
IL-lO expression. Northern blot analysis using the IL-lO eDNA
as a probe detected appreciable levels of InRNA transcripts in
all seven clones (Fig. 1, Lanes 3-9) but not in wild-type A375P
cells or in cells transfected with expression vector alone (Fig. 1,
Lanes J and 2, respectively). Secretion of IL-lO by the trans-
fected clones was assayed by ELISA. The seven isolated clones
were found to secrete IL-lO in the range of 10-100 ng/ml per
106 cells every 24 h.
Tumorigenicity of IL-lO-transfected Cells. To deter-
mine the tumorigenicity of the IL-lO-transfected clones, we
injected 1 X 106 cells from five different clones, designated 1-1
through 1-5, s.c. into BALB/c nude mice (Fig. 2). Parental
A375P and two Hygro-transfected clones (Hl and H2) grew
progressively, but the growth of the IL- lO-transfected cells was
much slower as compared to that of parental and A375P-Hygro
tumors. Fig. 2 shows that parental and A375P-Hygro tumors
Vitro Vivo
�-�-�‘ �-�--�
1 2 3 4 5 6 7 8 9 10
GAPDH
Fig. 3 Expression of IL-b in tumors in viva 35 days after s.c. injec-
(ions. Two tumors that express IL-lO in vitro, I-2 and 1-3 (Lanes 3 and5), continued to express IL-b mRNA in viva 35 days after s.c. injec-
tions (Lanes 7 and 9, respectively). In contrast, two tumors produced bythe control A375P-Hygro cells (Lanes 6 and 8) and one tumor produced
by the parental cells (Lane 10) did not express IL-b mRNA in vivo.
Lanes 1-5, cells in tissue culture; Lanes 6-JO, tumors growing s.c. in
nude mice 35 days after injections. Lane 1. A375P-parental: Lane 2.
A375P-H1; Lane 3, A375P-I-2: Lane 4. A375P-H2; Lane 5, A375P-I-3;
Lane 6, A375PH1; Lane 7, A375P-I-2: Lane 8, A375P-H2; Lane 9,
A375P-I-3: Lane 10, A375P-parental.
grew in all mice that received injections (100% tumor uptake),
reaching 1-1 .5 cm in mean diameter within 6 weeks. In contrast,
four of the IL-b transfected cells formed small tumors (3-5 mm
in mean diameter) 7 weeks after injections. Growth of the four
A375P-IL-lO cells differed significantly from parental and
A375P-Hygro cells at all times (P < 0.01), while the difference
between the fifth IL-lO-expressing clone (clone 1-1) and A375P
and A375P-Hygro was statistically significant 6 and 7 weeks
after injection (P < 0.05). We observed a direct correlation
between the amount of IL-b secreted by each individual clone
in vitro and its growth rate in vivo. For example, clone 1-3,
which secretes 100 ng/ml per 106 cells every 24 h IL-lO (Fig. 2,
bottom #{149}line) grew slower than other clones tested, whereas
clone 1-1 (10 ng/mlIlO6 eells/24 h; Fig. 2, upper S line) grew
more rapidly than the other IL-lO-transfeeted cells but slower
than the parental or Hygro-transfected cells (Fig. 2). IL-lO was
detected only in the serum of animals injected with IL-b-
transfected clones (1000-8000 pg/mI) but not in animals in-
jected with parental or A375P-Hygro cells. Interestingly
enough, the one A375P-IL-lO clone (1-1) that grew more pro-
gressively than the other IL-lO-transfected clones exhibited the
lowest levels of IL-lO in the serum (1000 ± 50 pg/ml).
Tumors from two mice that received s.c. injections of
A375P-IL-bO cells were harvested 35 days after injection and
assayed for IL-JO gene expression by Northern blot analysis.
Fig. 3 demonstrates that clones 1-2 and 1-3 that expressed IL-lO
in culture before s.c. injections (Lanes 3 and 5) continued to
express the IL-b mRNA transcript in viva (Lanes 7 and 9). In
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

A375P-13
A375P-H I
A375P
0.0 1.0 2.0 3.0 4.0
3H-Thymidlne Incorporation (cpm)
A375P-13
A375P-H1
A375P
Cell Number (x104)
A375P-13
A375P-H1
I’
A375P
Clinical Cancer Research 1973
contrast, two tumors produced by the control A375P-Hygro
cells and one tumor produced by the parental cells did not
express IL-lO mRNA in viva (Fig. 3, Lanes 6, 8, and JO,
respectively).
To determine if the growth inhibition exhibited by IL-lO-
expressing tumors was due to a direct effect of IL-lO on tumor
cell proliferation, we compared the growth of A375P-Hygro and
A375P-IL-l0 clones to that of parental A375P cells in vitro.
Using three different assays to determine in vitro growth rates,
no significant differences in cell proliferation were found among
the three cell populations (Fig. 4).
Experimental Lung Metastasis. We next analyzed the
capacity of IL-lO transfected cells to produce experimental lung
metastases after their iv. injection into the lateral tail vein of
BALB/c nude mice. As shown in Table 1, A375P and A375P-
Hygro (four clones were analyzed, Hl-H4) produced lung tu-
mor colonies in all mice injected. In contrast. experimental
metastasis of the seven IL-lO-transfected clones analyzed (1-1
through 1-7) was totally inhibited (clones 1-2 and 1-3) or pro-
duced a few lung metastases in some of the mice receiving
injections (clones I-b, 1-4, 1-5, 1-6, and 1-7: P < 0.001).
Effect of IL-lO on Tumor-infiltrating Macrophages.
Tumors are often infiltrated by macrophages. Because of their
many functions and states of activation, it has been proposed
that macrophages may exert tumor growth-promoting and -sup-
pressive properties (35). One of the major roles of IL-lO in the
regulation of immune responses may be due to its deactivating
activity on macrophages. For example, IL-lO down-regulates
MHC class II expression and suppresses the production of
several proinflammatory cytokines, such as IL-la, IL-6, IL-8,
TNF-ct, granuloeyte-macrophage and granubocyte colony-slim-
ulating factors, and reactive nitrogen oxides (3, 9-14).
To obtain evidence for the interaction between IL-lO-
transfeeted cells and the host macrophages, histological exam-
ination of tumors injected s.c. was performed 45 days after
injection (see “Materials and Methods”). Specific in situ stain-
ing for macrophages with F4/80 antibody revealed that A375P
and A375P-Hygro cells had a different macrophage content
from A375P-IL-lO cells. Although the same number of macro-
phages was observed in the periphery of the tumors, a large
number of infiltrating macrophages was observed within the
tumor produced by A375P-Hygro cells, compared with only a
few within the tumor produced by A375P-IL-10-expressing
cells (Fig. 5, A and B, respectively). Staining of the tumor
produced by A375P cells (data not shown) was similar to that
produced by A375P-Hygro cells (Fig. SA). IL-lO secreted by the
tumor cells may prevent the migration of macrophages from the
periphery into the tumor tissue and inhibits the production of
angiogenic and other tumor growth-promoting factors produced
by the few infiltrating and the large number of surrounding
maerophages.
IL-lO Inhibits Angiogenesis in Vivo. Malignant tumors
do not grow beyond 2-3 mm3 and cannot metastasize unless
they stimulate the formation of new blood vessels and thus
provide a route for the increased in-flow of nutrients and oxygen
and out-flow of waste products. Tumor angiogenesis also pro-
vides an essential exit route for metastasizing tumor cells from
the primary tumor to the bloodstream (16, 36). From the many
cells and cell products within a tumor that serve as inducers or
0.0 0.5 1.0 1.5 2.0
OD(at450 nm)
Fig. 4 In vitro growth rate of A375P (parental). Hygro- (A375P-Hl),
and IL-lO-transfected cells A375P-13. In vitro growth rate was meas-
ured by three different methods (see “Materials and Methods”): no
significant differences in proliferation rate were found among the three
cell populations.
modulators of angiogenesis. macrophages have emerged as a
major component (37). Several studies have indicated recently
that IL- I 0 inhibits the secretion of IL- I , IL-6, IL-8, TNF-a, and
MMP-9 by monocytes/macrophages. These cytokines are
known to be promoters of angiogenesis (37). We, therefore,
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

1974 Antiangiogenie Effect of Interleukin 10
Table 1 Suppression of lung metastasis in nude mice of A375P
human melanoma cells transfected with murine IL-b
Experimental lung metastasis”
Cell lines Metastases [neidenee” Median� (range)
A375P 4, 6, 13, 17, 56 5/5 13 (4-56)
A375P-Hl 3, 6, 8, 11, 43 5/5 8 (3-43)
A375P-H2 3, 4, 23, 77, 83 5/5 23 (3-83)
A375P-H3 1, 9, 12, 23, 24 5/5 12 (1-24)
A375P-H4 16, 29, 42, 98, 182 5/5 42 (16-182)A375P-Il 0,0,0,0,3 1/5 0”(0-3)A375P-I2 0, 0, 0, 0, 0 0/5 0”A375P-13 0, 0, 0, 0, 0 0/5 0”A375P-I4 0, 0, 0, 2, 10 2/5 0” (0-10)
A375P-I5 0, 0, 0, 3, 1 2/5 0” (0-1)
A375P-I6 0, 0, 0, 3, 13 2/5 0” (0-13)
A375P-I7 0, 0, 0, 0, 1 1/5 0d (0-1)
(2 Cells (I X 106) in 0.2 ml ofHBSS were injected iv., and the mice
were killed 60 days later.b Number of positive mice per number of injected mice.C Median (range) number of lung tumor colonies counted with the
aid of a dissecting microscope.
d p < 0.001 compared with A375P or A375P-Hl-H4.
sought to test the hypothesis that IL-lO exerts its antitumor
effect by inhibiting angiogenesis.
To determine the extent of vascularization in the tumors
produced s.c. by A375P, A375P-Hygro, and A375P-IL-l0 cells,
we used immunohistochemical techniques to identify cells re-
acting with antibodies against factor VIII, which specifically
stain endothelial cells. Two sections of equal size tumors were
selected for immunostaining with anti-factor VIII antibody, one
on the periphery of the tumor (Fig. 5, Cl and Dl) and one in the
center (Fig. 5, C2 and D2). We found very few blood vessels in
the skin surrounding the tumors produced by A375P-IL-lO cells
(Fig. 5, Dl), whereas blood vessels were virtually absent within
the IL-lO-secreting tumors (Fig. 5D2). In contrast, large num-
bers of blood vessels were observed surrounding and within the
tumor tissue of A375P-Hygro cells (Fig. 5, Cl and C2, respec-
lively). The same extent of vascularization was observed in
tumors produced by the parental A375P cells.
IL-lO Down-Regulates VEGF Expression in Macro-
phages. VEGF and bFGF are the most potent factors involved
in angiogenesis. The A375P human melanoma cells used in our
study express basal levels of VEGF and bFGF. The enforced
expression of IL-b in these cells had no effect on the expres-
sion of VEGF and bFGF (data not shown). Collectively, these
data and the data presented in Fig. 4 suggest that IL-lO does not
act in an autocrine fashion but rather it may affect the host
response in a paracrine fashion. Presumably, the amount of
VEGF and bFGF secreted by the tumor cells themselves might
not be sufficient to induce/establish an angiogenic process
within the tumor. Since infiltrating macrophages and macro-
phages surrounding the tumor can be a rich source of these
molecules, in the next set of experiments we wanted to deter-
mine whether the decrease in vascularization that we observed
in the IL-lO-secreting tumors is due to the down-regulation of
VEGF and bFGF production in macrophages exposed to IL-lO.
Since the conventional Northern blot analysis was not sensitive
enough to detect VEGF mRNA transcripts in macrophages, we
used semiquantitative RT-PCR to determine the effect of
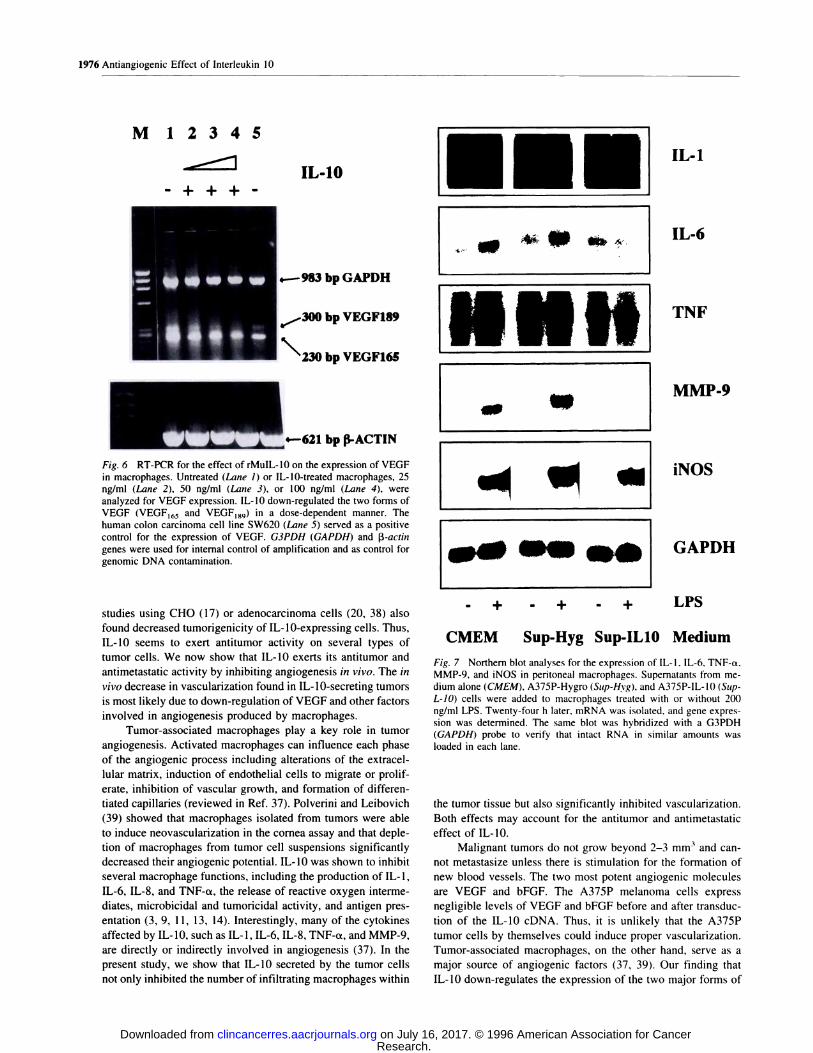
rMuIL-10 on the expression of VEGF in macrophages. Fig. 6
shows that IL-b down-regulates, in a dose-dependent manner,
the expression of the two major forms of VEGF, VEGF,65
(230-bp band on the gel) and VEGF,89 (300-bp band). Treat-
ment of the macrophages with 100 ng/ml rMuIL-lO abolished
the VEGF189 transcript (Fig. 6, Lane 4) and reduced signifi-
cantly the RNA expression of the VEGF165 form. bFGF mRNA
synthesis was hardly detectable in macrophages by RT-PCR
before and after treatment with IL-b, implying that VEGF is
the dominant angiogenic factor secreted by macrophages.
Supernatants from IL-lO-transfected Tumor Cells
Down-Regulate the Expression of IL-i, IL-6, TNF-a, and
MMP-9 by Macrophages. Recent studies have indicated that
rMuIL-lO added to macrophages in vitro inhibits the production
of IL- 1 , IL-6, TNF-a ( 1 1 ), and collagenase type IV MMP-9
(12). In the next set of experiments, we wanted to determine
whether the IL-lO secreted by the tumor cells studied here had
the same biological activity. To this end, supernatants collected
from parental A375P, A375P-Hygro, and A375P-IL-l0 (clone
1-3, secreting 100 ng/106 cells every 24 h) cells grown in culture
were added to peritoneal macrophages before and after stimu-
lation with LPS. The RNA expression of the IL-J, IL-6, TNF-a,
and MMP-9 genes was determined by Northern blot analysis.
Fig. 7 shows that IL-lO inhibited the transcription of these four
genes in LPS-treated macrophages, whereas the expression of
the iNOS gene was not affected (Fig. 7).
Tumor Growth Inhibition of Other Murine and Human
Melanoma Cells Admixed with A375P-IL-10 In the last set
of experiments, we wanted to determine the tumorigenicity of
other murine and human melanoma cells in nude mice when
co-injected with A375P-IL-bO cells. We chose to work with the
murine melanoma cell line Bl6-BL6 and with the human mel-
anoma cell line A375SM because both are highly tumorigenic in
nude mice and produce no IL-lO. A375SM cells alone or ad-
mixed with A375P-Hygro before injection (in a ratio of 1 : 1, see
“Materials and Methods”) grew progressively in nude mice. In
contrast, the growth of A375SM cells was significantly inhibited
(P < 0.01, at all points measured) when admixed with A375P-
IL-lO (clone 1-3) before injection (Fig. 8A). The murine Bl6-
BL6 cell line is a fast-growing tumor in nude mice (Fig. 8: note
the difference in the time scale between A and B). Injection of
Bl6-BL6 cells alone or admixed with A375P-Hygro grew pro-
gressively in nude mice, reaching 1-1.5 em in mean diameter
within 2 weeks. In contrast, the tumorigenicity of B16-BL6 cells
was inhibited significantly (P < 0.01) when admixed with
A375P-IL-bO cells (in a ratio of 1:5) in spite of its fast growth
rate (Fig. 8B). Thus, the inhibitory effect of IL-lO was not
limited to one melanoma cell line and was not restricted to
malignant cells of human origin only. Collectively, these data
indicate that the presence of IL-lO within a tumor inhibits
macrophage-derived angiogenic factors, and hence, tumor
growth and eventually metastasis.
DISCUSSION
In this study, we demonstrated that enforced expression of
IL-lO in human melanoma cells significantly suppressed their
tumorigenieity and metastatic potential in nude mice. Recent
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

. -:-.._�. #{149}�‘!�.
� .-.�. ,%
�#{149}L � �,,. . � �
�. ,.l’%� �.‘#{176}� � �,P
�.‘ . -‘. � #{182}
� � #{149}�“.� ���lh.::d’:.� *: � B
>
0
U-
,. �p�’� � , �. �- a � �
. � .��-“:
� ..
�- � .�: � � �� ‘::� ‘�-‘: ‘ ‘ #{149}:,� ‘�‘ � � C2
Clinical Cancer Research 1975
0
U-
A375P-Hygro A375P-IL-1 0
- . :,- � ��..d ‘ ‘��#{149}t .�‘ a
#{149}4� ‘ �. ./ %.,. s..’ ‘.. �
#{149}�# I’ .. _‘� _,.4. . � ph’
‘�!
_.,. 4’..� p
� �; Y
, �‘,‘:. � �
� , � :‘,:::-/�(� �.
� V �4#{231}�”� ‘��‘ � � �
‘1��%js .,� ‘�#{149};:‘ � � � ‘�‘ Dl
� #{149}a,4��#{149} � 4 � .- �.,,,,� #{149}..‘; � ‘ - a,. �‘, � -.,1Ii .�(, � �
� ;. ‘. #{149} .,.. ‘ .. ,.h � � � #{149}�f _*l,. #{149} 4, � #{149}% #{149}I�d �#{149} 4$ #{149}.4 #{149}� #{176}O’� 4’t .‘�‘
- � � �3 -.�.. #{149}.p.�4�-�* #{149}�.(‘� #{149} �, . ,, ‘.,� a, � �� � #{149}� , S� S #{149}
� � 4 _ _ �‘, .. � � 4� .p1� #{149}.‘p4 #{149}‘��4 � �‘ �.‘ 4*’
).�. �., ..� . �: . #{149}� #{149}..IaI,;D�� �I,.1., ‘‘- � I s -� #{149}�4� ., � I �. #{149} � � ‘F 4IPl�# .,� I #{149},.� � #{149}? : #{149}� � � . ‘�‘��#{149}
4% #{149}�#{149} .; I � , #{149}. #{149}A#Ei�. #{149}S � � �4 �‘ � f. S #{149} , a� � . � k #{149}#{149}�#{149}
� ‘ 1’ � ‘ -‘ , ‘‘ . *. �..* �-‘ . �54
. � ,, , . . ,�� ,t., a�,c.,. �
I. .�?+ffi�r� :.lb,,� #{149}Id:l.�:.T�..: ,�‘!�:,i�.qi’�
� O’v.�, I �‘ a � � ‘ � _�*��1#{149}� #{149}#{149}� � f�D2
Fig. 5 A and B, immunohistochemical staining for tumor-infiltrating macrophages in mice bearing tumors after injection of A375P-Hygro (A) or
A375P-IL-l0 (B) cells. Frozen sections were cut and stained with F4/80 rat monoclonal antibody. The pattern shown is representative of more than
10 random fields examined. Note that in A375P-Hygro tumor (A). macrophages had infiltrated the tumor tissue. whereas in A375P-IL-bO tumor (B).
they were absent within the tumor but still present around the tumor tissue. C and D, immunohistochemical staining with anti-factor VIII antibody
in tumor produced by A375P-Hygro (Cl and C2) and in tumor produced by A375P-IL-I0 (DI and D2). For each tumor, two sections were analyzed,one close to the periphery (CI and Dl) and one representing the center of the tumor (C2 and D2). Note neovascularization in A375P-Hygro tumors(CI and C2) but very few in A375P-IL-lO tumors (DI and D2).
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
IL-bIL-i
IL-6
I-� --k- --�-- -
GAPDH
1976 Antiangiogenic Effect of Interleukin 10
M 12345
bp GAPDH
bp VEGF189
bp VEGF16S
bp I�-ACTIN
Fig. 6 RT-PCR for the effect of rMuIL-10 on the expression of VEGF
in macrophages. Untreated (Lane 1) or IL-lO-treated macrophages, 25ng/ml (Lane 2), 50 ng/ml (Lane 3), or 100 ng/ml (Lane 4), wereanalyzed for VEGF expression. IL-b down-regulated the two forms of
VEGF (VEGF165 and VEGF189) in a dose-dependent manner. Thehuman colon carcinoma cell line SW620 (Lane 5) served as a positive
control for the expression of VEGF. G3PDH (GAPDH) and �3-actin
genes were used for internal control of amplification and as control for
genomie DNA contamination.
studies using CHO (17) or adenocarcinoma cells (20, 38) also
found decreased tumorigenicity of IL-lO-expressing cells. Thus,
IL- 10 seems to exert antitumor activity on several types of
tumor cells. We now show that IL-b exerts its antitumor and
antimetastatic activity by inhibiting angiogenesis in vivo. The in
vivo decrease in vascularization found in IL-lO-secreting tumors
is most likely due to down-regulation of VEGF and other factors
involved in angiogenesis produced by macrophages.
Tumor-associated macrophages play a key role in tumor
angiogenesis. Activated macrophages can influence each phase
of the angiogenic process including alterations of the extracel-
lular matrix, induction of endothelial cells to migrate or prolif-
erate, inhibition of vascular growth, and formation of differen-
tiated capillaries (reviewed in Ref. 37). Polverini and Leibovich
(39) showed that macrophages isolated from tumors were able
to induce neovascularization in the cornea assay and that deple-
tion of macrophages from tumor cell suspensions significantly
decreased their angiogenie potential. IL-b was shown to inhibit
several macrophage functions, including the production of IL- 1,
IL-6, IL-8, and TNF-a, the release of reactive oxygen interme-
diates, microbicidal and tumoricidal activity, and antigen pres-
entation (3, 9, 1 1, 13, 14). Interestingly, many of the cytokines
affected by IL- 10, such as IL- 1 , IL-6, IL-8, TNF-ct, and MMP-9,
are directly or indirectly involved in angiogenesis (37). In the
present study, we show that IL-lO secreted by the tumor cells
not only inhibited the number of infiltrating macrophages within
�- � 4... 4$ #{149}b.�
H
--�
a#{149}�IS�#{149}
TNF
MMP-9
INOS
- + - + - + LPS
CMEM Sup-Hyg Sup-IL1O Medium
Fig. 7 Northern blot analyses for the expression of IL-l, IL-6, TNF-a.
MMP-9, and iNOS in peritoneal macrophages. Supernatants from me-dium alone (CMEM). A375P-Hygro (Sup-Hvg), and A375P-IL-10 (Sup-
L-1O) cells were added to macrophages treated with or without 200ng/ml LPS. Twenty-four h later, mRNA was isolated, and gene expres-
sion was determined. The same blot was hybridized with a G3PDH(GAPDJ-I) probe to verify that intact RNA in similar amounts was
loaded in each lane.
the tumor tissue but also significantly inhibited vascularization.
Both effects may account for the antitumor and antimetastatie
effect of IL-b.
Malignant tumors do not grow beyond 2-3 mm3 and can-
not metastasize unless there is stimulation for the formation of
new blood vessels. The two most potent angiogenic molecules
are VEGF and bFGF. The A375P melanoma cells express
negligible levels of VEGF and bFGF before and after transdue-
tion of the IL-b eDNA. Thus, it is unlikely that the A375P
tumor cells by themselves could induce proper vascularization.
Tumor-associated macrophages, on the other hand, serve as a
major source of angiogenie factors (37, 39). Our finding that
IL-b down-regulates the expression of the two major forms of
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

weeks Days
Time after Tumor Injection
C)C,
ECu
EC.)
C,N
U)L.
0E
I-
Clinical Cancer Research 1977
Fig. 8 A. suppressed tumorigenicity of human melanoma A375SM cells when admixed with A375P-IL-l0 but not with A375P-Hygro cells. In
mixture experiments, I X 106 A375SM cells were admixed with I X 106 A375P-Hygro or I X 106 A375P-IL-l0 cells before s.c. injection into nude
mice. B, suppressed tumorigenicity of murine B16-BL6 melanoma cells when admixed with A375P-IL-l0 cells but not with A375P-Hygro cells. In
mixture experiments. 0.2 X l0�’ B16-BL6 cells were admixed with I X 10” A375P-Hygro or I X 106 A375P-IL-l0 cells before injection s.c. into
nude mice. *, significance as compared with equivalent points of cells admixed with A375P-Hygro cells.
VEGF, VEGFI#{212}Sand VEGF15�, in macrophages may provide an
explanation for the decreased vascularization that we observed
in IL-lO-seereting tumors in vito. The protein form of VEGF165
is secreted as more soluble cellular product, whereas VEGF189
is secreted as less soluble protein that becomes tightly bound to
the extracellular matrix (34). Together with the observations that
a large spectrum of other molecules involved in angiogenesis,
such as IL-I �, TNF, IL-6, and the proteinase MMP-9 were also
inhibited in activated macrophages by supernatants from
A375P-IL-lO cells, our data suggest that the antiangiogenie
effect of IL-lO is most likely due to its ability to down-regulate
angiogenic molecules secreted by macrophages. One possibility
for the antiangiogenic effect of IL-lO is the production of
angiostatin, an inhibitor of angiogenesis (40). The source of
angiostatin is not known. It was postulated that metalboelastase
secreted by macrophages is responsible for the production of
angiostatin. However, recent studies have shown that IL-b had
no effect on the secretion of metalboelastase by macrophages
(4 1 ). suggesting that IL- I 0 does not promote the production of
angiostatin. The attachment of A375P cells to extracellular
matrix proteins such as laminin and fibroneetin was unaffected
by IL-lO expression (data not shown), arguing against the pos-
sibility that changes in adhesion properties of the tumor cells
could account for the observed decrease in tumorigenicity and
metastasis.
The metastatic potential of tumor cells depends, among
other things, on proper vascularization of the tumor and its
ability to degrade type IV collagen. Of particular interest is the
ability of IL-b to inhibit IL-8 production by macrophages and
neutrophils. IL-8 is a macrophage-derived mediator of angio-
genesis (42). We have recently shown that IL-8 promotes met-
astatie potential in primary melanoma cells through the induc-
tion of the type IV collagenase MMP-2 (26). It is, therefore,
possible that the inhibition of MMP-9 ( I 2) and the up-regulation
of TIMP-l (43) by IL-b in macrophages is mediated via the
down-regulation of IL-8 production in these cells. It was re-
cently shown that IL-lO inhibits nuclear localization of the
nuclear factor kappa B (NF-kB), a transcription factor involved
in the expression of IL-8 and other inflammatory cytokine genes
(44). In this regard. it is possible that the dramatic decrease in
the ability of IL-lO-seereting cells to produce lung metastases
following iv. injections could be due to down-regulation of
MMP-9 ( 12) and up-regulation of TIMP-l (43) by IL-b. An-
other possible mechanism (not mutually exclusive with those
indicated above) is that IL-lO-transfeeted cells might became
sensitive to NK cells (20, 45). Indeed, recent studies have
suggested that the antimetastatie effect of IL- 10 in a murine
model of breast cancer might be dependent on the activity of NK
cells (45).
A possible molecular mechanism for the deactivation of
LPS-aetivated monocytes/macrophages by IL-lO has been pro-
vided recently by Geng et a!. (46). Activation of macrophages
by LPS is mediated through the activation ofthe protein tyrosine
kinase p56�’, and this is associated with tyrosine phosphoryl-
ation of the proto-oncogene Vav. LPS also increases Ras aeti-
vation in monocytes. IL-b has been shown to completely
inhibit the p56” tyrosine kinase activation and all other subse-
quent events in the p56” pathway. including Ras activation
(46).
Macrophages exert their tumoricidal activity through the
induction of iNOS and secretion of NO (47, 48). In our studies,
we were unable to demonstrate any changes in iNOS expression
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

1978 Antiangiogenic Effect of Interleukin 10
and NO production in macrophages after exposure to superna-
tants from IL-lO-transfected tumor cells (Fig. 7 and data not
shown). This makes it unlikely that the decrease in tumor
growth of IL-lO-transfected cells was due to an increase in
NO-driven cytotoxicity of macrophages against the tumor cells.
Recently, it has been shown that IL-l2 possesses an anti-
angiogenic activity that is mediated via IFN-�y secreted by
accessory cells (49, 50). In our nude mice model (lacking
T-cells), it is unlikely that IL-lO exerts its antiangiogenic activ-
ity through IFN-’y because: (a) IL-lO down-regulates IFN-’y
production by NK cells (5 1); (b) macrophages were negative for
IFN--y production before and after treatment with supernatants
from IL-lO-transfected cells; (c) B cells are an unlikely source
for IFN-�y; and (tO the inhibition of angiogenesis cannot be
attributed to IFN-y secreted by the tumor cells themselves due
to the IFN-y species-specific mode of action.
IL-lO secreted by the A375P-IL-l0 cells also inhibited
tumor growth of other IL- 10-negative murine and human mel-
anoma cells admixed with A375P-IL-lO cells before injection
(Fig. 8). These results indicate that: (a) the inhibitory effect of
IL-lO is not limited to one melanoma cell line; (b) IL-b can
affect bystander tumor cells; and (c) IL-lO need not be secreted
by the tumor cells themselves to exert its antitumor effects. To
further evaluate the antitumor and antimetastatic effect of IL-lO,
experiments are now under way in our laboratory to examine
tumor growth and metastasis of murine melanoma cells in
transgenic mice expressing IL- 10.
In conclusion, IL- 10 has now been shown to be a potent
inhibitor of tumor growth in three different tumor model sys-
tems (Refs. 17, 20, 37, and 45 and this study). In addition, we
now show, for the first time, that IL-lO is a potent antimetastatic
cytokine and that IL-lO may exert its antitumor and antimeta-
static activities through inhibition of angiogenesis. Recent stud-
ies have demonstrated that IL-lO given systemically to mice
(52) or humans (53, 54) had only few of the toxic effects
associated with the administration of many other cytokines.
Therefore, the efficacy of IL-b as an antimetastatic agent
should be further explored, both as an independent treatment
and in combination with other inhibitors of neovascularization.
ACKNOWLEDGMENTS
We thank Drs. Isaiah J. Fidler, Ronald Palacios, and Hector Mar-tinez-Vabdez for critical review; Drs. Paobo Dellabona and Ronald
Palacios for the murine IL-b expression vector; Dr. Carl Nathan for theiNOS probe; Donna Reynolds for the immunohistochemistry staining;
Craig McClain for the IL-b ELISA assays: Kenneth Dunner, Jr. for
photography; and Patherine Greenwood for expert preparation of themanuscript.
REFERENCES
I. Fiorentino, D. F., Bond, M. W., and Mosmann, T. R. Two types ofmouse helper T cell. IV. Th2 clones secrete a factor that inhibitscytokine production by Thl clones. J. Exp. Med., 170: 2081-2095,
1989.
2. Moore, K. W., Vieira, P., Fiorentino, D. F., Trounstine, M. L., Khan,
1. A., and Mosmann, T. R. Homology of cytokine synthesis inhibitoryfactor (IL-lO) to the Epstein-Barr virus gene BCRFJ. Science (Wash-ington DC), 248: 1230-1234, 1990.
3. Howard, M., and O’Garra, A. Biological properties of interleukin 10.Immunol. Today, J3: 198-200, 1992.
4. Zlotnik, A., and Moore, K. W. Interleukin 10. Cytokine. 3: 366-379.1991.
5. Fiorentino, D. F., Zbotnik, A., Vieira, P., Mosmann, T. R., Howard,
M., Moore, K. W., and O’Garra, A. IL-b acts on the antigen-presentingcell to inhibit cytokine production by Thl cells. J. Immunol., 146:
3444-3451, 1991.
6. Moore, K. W., O’Garra, A.. de Waal Malefyt. R.. Vieira, P., andMosmann. T. R. Interleukin 10. Annu. Rev. Immunol., 11: 165-190,1993.
7. Hsu, D-H., de Waal Malefyt, R., Fiorentino, D. F., Dang, M-N.,
Vieira, P., de Vries, J., Spits, H., Mosmann, T. R., and Moore, K. W.
Expression of IL-b activity by Epstein-Barr virus protein BCRF1.Science (Washington DC), 250: 830-832, 1990.
8. Hsu, D-H., Moore, K. W., and Spits, H. Differential effects ofinterleukin-4 and -10 on interleukin-2-induced interferon--y synthesis
and lymphokine-activated killer activity. mt. Immunol.. 4: 563-569,
1992.
9. de Waal Malefyt. R., Abrams, J., Bennett, B., Figdor, C. G.. and deVries, J. E. Interleukin 10 (IL-b) inhibits cytokine synthesis by humanmonocytes: an autoregulatory role of IL- 10 produced by monoeytes. J.Exp. Med., 174: 1209-1220, 1991.
10. Bogdan, C., Vodovotz, Y., and Nathan, C. Macrophage deactivationby interleukin-lO. J. Exp. Med., 174: 1549-1555, 1991.
I I. Fiorentino, D. F., Zlotnik, A., Mosmann, T. R., Howard, M., andO’Garra, A. IL-b inhibits cytokine production by activated macro-phages. J. Immunol., 147: 3815-3822, 1991.
12. Mertz, P. M., DeWitt, D. L., Stetler-Stevenson, W. G., and Wahl,L. M. Interleukin-lO suppression of monocyte prostaglandin H syn-
thase-2. J. Biol. Chem., 269: 21322-21329, 1994.
13. de Waal Malefyt, R., Haanen, J., Spits, H., Roncarobo, M-G., teVelde, A., Figdor, C., Johnson, K., Kastelein, R., Yssel, H., and deVries, J. E. Interleukin 10 (IL-lO) and viral IL-b strongly reduce
antigen-specific human T cell proliferation by diminishing the antigen-
presenting capacity of monocytes via downregulation of class II majorhistocompatibility complex expression. J. Exp. Med.. 174: 915-924.1991.
14. Gazzinelli, R. T., Oswald, I. P., James, S. L., and Sher, A. IL-b
inhibits parasite killing and nitrogen oxide production by IFN--y-aeti-
vated macrophages. J. Immunol., 148: 1792-1796. 1992.
15. Folkman, J., and Klagsbrun, M. Angiogenie factors. Science(Washington DC), 235: 442-447, 1987.
16. Rak, J. W., St. Croix, B. D., and Kerbel, R. S. Consequences ofangiogenesis for tumor progression. metastasis and cancer therapy.
Anticancer Drugs, 6: 3-18. 1995.
17. Richter, G., Kruger-Krasagakes, S., Hem, G., Hubs, C., Schmitt, E.,
Diamanstein, T., and Blankenstein, T. Interleukin 10 transfeeted into
Chinese hamster ovary cells prevents tumor growth and macrophage
infiltration. Cancer Res., 53: 4134-4137, 1993.
18. Li, L., Price, J. E., Fan, D., Zhang, R. D., Bucana, C. D., and Fidler,
I. J. Correlation of growth capacity of human tumor cells in hard agarose
with their in viva proliferation capacity at specific metastatie sites.J. NatI. Cancer Inst., 81: 1406-1412, 1989.
19. Hart, I. R. Selection and characterization of an invasive variant ofthe Bl6 melanoma. Am. J. Pathol., 97: 587-600, 1979.
20. Giovarelli, M.. Musiani, P., Modesti. A., Dellabona, P., Casorati,
G., Allione, A., Consalvo, M., Cavallo, F., diPierro, F., De Giovanni, C.,Musso, T., and Forni, G. Local release of IL-b by transfected mouse
mammary adenocareinoma cells does not suppress but enhances antitu-
mor reaction and elicits a strong cytotoxic lymphocyte and antibody-dependent immune memory. J. Immunol.. 55: 31 12-3123, 1995.
21. Radinsky, R.. Fidler. I. J.. Price, J. E., Esumi, N.. Tsan, R., Petty,C. M., Bucana, C. D., and Bar-Eli, M. Terminal differentiation and
apoptosis in experimental lung metastases of human osteogenic sarcoma
cells by wild type p53. Oneogene. 9: 1877-1883, 1994.
22. Huang, S., Singh, R. K., Xie. K., Gutman, M., Berry. K. K., Bucana,
C. D.. Fidler, I. J., and Bar-Eli, M. Expression of the JE/MCP-1 gene
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

Clinical Cancer Research 1979
suppresses metastatic potential in murine colon carcinoma cells. Cancer
Immunol. Immunother., 39: 231-238, 1994.
23. Huang. S.. Xie. K.. Singh. R. K.. Gutman. M.. and Bar-Eli, M.
Suppression of tumor growth and metastasis of murine renal adenocar-
cinoma by syngeneic fibroblasts genetically engineered to secrete the
JE/MCP-l cytokine. J. Interferon Cytokine Res., /5. 655-665, 1995.
24. Ullrich, S. E. Mechanism involved in the systemic suppression of
antigen-presenting cell function by UV irradiation: keratinocyte-derived
IL-b modulates antigen-presenting cell function of splenie adherent
cells. J. Immunol., 152: 3410-3416, 1994.
25. Luca, M., Hunt, B., Bucana, C. D., Johnson, J. P., Fidler, I. J., andBar-Eli, M. Direct correlation between MUCI8 expression and meta-static potential of human melanoma cells. Melanoma Res., 3: 35-41,
1993.
26. Singh, R. K., Gutman, M., Reich, R., and Bar-Eli, M. Ultraviolet B
irradiation promotes tumorigenie and metastatic properties in primary
cutaneous melanoma via induction of interleukin 8. Cancer Res.. 55:
3669-3674, 1995.
27. Luca, M., Xie, S., Gutman, M., Huang, S., and Bar-Eli, M. Abnor-
malities in the CDKN2 (P16�4/MTS-1) gene in human melanomacells: relevance to tumor growth and metastasis. Oneogene, 11: 1399-1402, 1995.
28. Fort, R., Marty. L., Piechaczyh, M., Sabrouty. S. E.. Dani. C..
Jeanteur, R., and Blanchard, J. M. Various rat adult tissues express only
major mRNA species from the glyceraldehyde-3-phosphate-dehydro-
genase multigenie family. Nucleic Acids Res.. 13: 1431-1442, 1985.
29. Xie, Q-W., Cho. H. J., Calaycay. J., Mumford, R. A., Swiderek,
K. M.. Lee, T. D.. Ding. A., Troso. T., and Nathan, C. Cloning and
characterization of inducible nitric oxide synthase from mouse macro-
phages. Science (Washington DC), 266: 225-228, 1992.
30. Rogelj. S., Weinberg, R. A., Fanning, P., and Klagsbrun, M. Basicfibroblast growth factor fused to a signal peptide transforms cells.
Nature (Lond.), 331: 173-175, 1988.
3 1 . Fidler. I. J. Orthotopic implantation of human colon carcinomas intonude mice provides a valuable model for the biology and therapy ofcancer. Cancer Metastasis Rev., 10: 229-243, 1991.
32. Austyn, J. M., and Gordon, S. F4/80, a monoebonal antibody di-
reeted specifically against the mouse macrophages. Eur. J. Immunol.,
11: 805-815, 1981.
33. Anthony. F. W., Wheeler, T.. Elcoek, C. L., Pickett, M., and
Thomas, E. J. Identification of a specific pattern of vascular endothelial
growth factor mRNA expression in human placenta and cultured pla-eental fibroblasts. Placenta, 15: 557-561 , I 994.
34. Houck. K. A.. Leung, D. W., Rowland, A. M., Winer, J., andFerrara, N. Dual regulation of vascular endothelial growth factor bio-availability by genetic and proteolytic mechanism. J. Biol. Chem., 267:
2603 1-26037, 1992.
35. Mantovani, A.. Botazzi, B., Colotta, F., Sozzani, S., and Ruco, L.
The origin and function of tumor-associated macrophages. Immunol.Today. 13: 265-270, 1992.
36. Folkman, J. The role of angiogenesis in tumor growth. Semin.
Cancer Biol., 3: 65-71. 1992.
37. Sunderkotter, C., Steinbrink, K., Goebeler, M., Bhardwaj. R., andSorg, C. Maerophages and angiogenesis. J. Leukocyte Biol., 55: 410-422, 1994.
38. Allione, A., Consalvo, M., Nanni, P., Lollini, P. L., Cavallo, F..
Giovarelli, M., Fomi, M., Forni, G. A., Colombo, M. P., Dellabona, P.,
Hock, H.. Blankenstein. T., Rosenthal, F. M., Gansbacher, G., Bosco,M. C., Musso, T., Guesella, L., and Guido. F. Immunizing and curativepotential of replicating and nonreplicating murine mammary adenocar-cinoma cells engineered with interleukin-l, IL4, 1L6, IL7, ILbO, tumor
necrosis factor a, granulocyte macrophage colony stimulating factor,
and -y-interferon gene or admixed with conventional adjuvants. CancerRes.. 54: 6022-6026, 1994.
39. Polverini, P. J., and Leibovich, S. J. Induction of neovaseularization
in viva and endothelial proliferation in vitro by tumor-associated macro-
phages. Lab. Invest., 51: 635-642, 1984.
40. O’Reilly, M. S., Holmgren, L., Shing, Y.. Chen, C., Rosenthal,
R. A., Moses, M., Lane, W. S., Cao, Y., Helene Sage, E.. and Folkman.
J. Angiostatin: a novel angiogenesis inhibitor that mediates the suppres-sion of metastases by a Lewis lung carcinoma. Cell, 79: 315-328. 1994.
41. Kumar, R., Dong, Z., and Fidler, I. J. Differential regulation of
metalloelastase activity in murine peritoneal macrophages by GM-CSF
and M-CSF. J. Immunol., in press, 1996.
42. Koch, A. E., Polverini, P. J., Kunkel. S. L., DiPietro. L. A., Elner.
V. M., Elner, S. G., and Stricter, R. M. Interleukin-8 as a macrophage-derived mediator of angiogenesis. Science (Washington DC), 258:
1798-1801. 1992.
43. Lacey, S., Nicod, L. P., Chicheportiche, R.. Welgus, H. G.. andDayer, J. M. IL- 10 inhibits metalboproteinase and stimulates TIMP- I
production in human mononuclear phagocytes. J. Clin. Invest.. 96:
2304-2310, 1995.
44. Wang, P., Wu, P., Siegel, M. I., Egan. R. W., and Billah, M. M.
Interleukin (lL)-b0 inhibits nuclear factor K B (NF K B) activation in
human monocytes-IL-bO and IL-4 suppress cytokine synthesis by dif-
ferent mechanisms. J. Biol. Chem., 270: 9558-9563, 1995.
45. Kundu, N., Beath, L. T., Jackson, M. J., and Fulton, A. M. Anti-metastatic and antitumor activities of interleukin- 10 in a murine model
of breast cancer. J. Natl. Cancer Inst., 88: 536-541, 1996.
46. Geng, Y., Gulbins, E., Altman, A., and Lotz. M. Monocyte deac-
tivation by interleukin 10 via inhibition of tyrosine kinase activity and
the Ras signaling pathway. Proc. Natl. Acad. Sci. USA, 91: 8602-8606,
1994.
47. Hibbs, J. B., Taintor, R. R., Jr., and Vavrin, Z. Macrophage cyto-
toxicity: role for L-arginme deaminase activity and amino nitrogen to
nitrate. Science (Washington DC), 235: 473-476, 1987.
48. Stuehr, D. J., and Nathan, C. F. Nitric oxide: a macrophage productresponsible for cytostasis and respiratory inhibition in tumor target cells.
J. Exp. Med., 169: 1543-1555, 1989.
49. Voest, E. E., Kenyan, B. M., O’Reilly, M. S.. Truitt, G., D’Amato,R. J., and Folkman, J. Inhibition of angiogenesis in vito by interleukin
12. J. NatI. Cancer Inst., 87: 581-586, 1995.
50. Kerbel, R. S., and Hawley, R. G. Interleukin 12: newest member of
the antiangiogenic club. J. NatI. Cancer Inst., 87: 557-559, 1995.
51 . D’Andrea, A., Aste-Amezaga, M.. Valiante, N. M., Ma, X.. Kubin,M.. and Trinehieri, G. Interleukin 10 (IL-lO) inhibits human lymphocyteinterferon -y-production by suppressing natural killer cell stimulatory
factorflL-l2 synthesis in accessory cells. J. Exp. Med.. 178: 104 1-1048.
1993.
52. Powrie, F., Menon, S., and Coffman, R. L. Interleukin 4 andinterleukin 10 synergize to inhibit cell mediated immunity in vito. Eur.J. Immunol., 23: 3043-3049, 1993.
53. Chernoff, A. E., Granowitz. E. V., Shapiro, L., Vannier, E..
Lonnemann, G., Angel, J. B.. Kennedy. J. S., Robson, A. R.. Wolff.
S. M.. and Dinarello, C. A. A randomized, controlled trial of IL-b in
humans: inhibition of inflammatory cytokine production and immune
response. J. Immunol., 154: 5492-5499, 1995.
54. Huhn, R. D., Radwanski, E., O’Connell, S. M., Sturgill, M. G..Clarke, L., Cody, R. P., Aifrine, M. B., and Cutler. D. L. Pharmacoki-netics and immunomodulatory properties of intravenously administered
recombinant human interleukin-lO in healthy volunteers. Blood, 87:
699-705, 1996.
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

1996;2:1969-1979. Clin Cancer Res S Huang, K Xie, C D Bucana, et al. human melanoma cells: potential inhibition of angiogenesis.Interleukin 10 suppresses tumor growth and metastasis of
Updated version
http://clincancerres.aacrjournals.org/content/2/12/1969
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on July 16, 2017. © 1996 American Association for Cancerclincancerres.aacrjournals.org Downloaded from